
Mother–offspring reunion in the South American sea lion Otaria flavescens at Isla de Lobos...
15
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Trimble, Micaela] On: 9 August 2010 Access details: Access Details: [subscription number 925481424] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Ethology Ecology & Evolution Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t916668712 Mother-offspring reunion in the South American sea lion Otaria flavescens at Isla de Lobos (Uruguay): use of spatial, acoustic and olfactory cues M. Trimble a ; S. J. Insley b a Cetáceos Uruguay, Sección Etología, Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay b Department of Biology, University of Victoria, Victoria, BC, Canada Online publication date: 09 August 2010 To cite this Article Trimble, M. and Insley, S. J.(2010) 'Mother-offspring reunion in the South American sea lion Otaria flavescens at Isla de Lobos (Uruguay): use of spatial, acoustic and olfactory cues', Ethology Ecology & Evolution, 22: 3, 233 — 246 To link to this Article: DOI: 10.1080/03949370.2010.502318 URL: http://dx.doi.org/10.1080/03949370.2010.502318 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Mother–offspring reunion in the South American sea lion Otaria flavescens at Isla de Lobos...

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Trimble, Micaela]On: 9 August 2010Access details: Access Details: [subscription number 925481424]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Ethology Ecology & EvolutionPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t916668712
Mother-offspring reunion in the South American sea lion Otaria flavescensat Isla de Lobos (Uruguay): use of spatial, acoustic and olfactory cuesM. Trimblea; S. J. Insleyb
a Cetáceos Uruguay, Sección Etología, Facultad de Ciencias, Universidad de la República, Montevideo,Uruguay b Department of Biology, University of Victoria, Victoria, BC, Canada
Online publication date: 09 August 2010
To cite this Article Trimble, M. and Insley, S. J.(2010) 'Mother-offspring reunion in the South American sea lion Otariaflavescens at Isla de Lobos (Uruguay): use of spatial, acoustic and olfactory cues', Ethology Ecology & Evolution, 22: 3,233 — 246To link to this Article: DOI: 10.1080/03949370.2010.502318URL: http://dx.doi.org/10.1080/03949370.2010.502318
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Ethology Ecology & Evolution 22: 233–246, 2010
ISSN 0394-9370 print/ISSN 1828-7131 online© 2010 Dipartimento di Biologia Evoluzionistica dell’Università, Firenze, ItaliaDOI: 10.1080/03949370.2010.502318http://www.informaworld.com
TEEE0394-93701828-7131Ethology Ecology & Evolution, Vol. 22, No. 3, Jul 2010: pp. 0–0Ethology Ecology & EvolutionMother–offspring reunion in the South American sea lion Otaria flavescens at Isla de Lobos (Uruguay): use of spatial, acoustic and olfactory cues
Mother–offspring reunion in the South American sea lionM. Trimble and S.J. InsleyM. TRIMBLE 1,3 and S.J. INSLEY 2
1 Cetáceos Uruguay, Sección Etología, Facultad de Ciencias, Universidad de la República,Iguá 4225, Montevideo 11400, Uruguay2 Department of Biology, University of Victoria, PO Box 3020, Station CSC, Victoria, BCV8W 3N5, Canada
Received 4 May 2009, accepted 12 February 2010
In the South American sea lion (Otaria flavescens), mothers and pups are regu-larly separated throughout lactation, demanding a well-developed individual recogni-tion system. The aim of this study was to investigate and quantify the searching andreunion behaviour between mothers and pups at Isla de Lobos, Uruguay. We recordeddetails of searches and reunions using focal animal sampling during two breeding sea-sons. Our results support the hypotheses that mother–pup dyads combine spatial,acoustic and olfactory cues for successful recognition and reunion behaviour and thatpups play an active role in the process. A total of 74% of the females returning from seavisited the last zone (4 × 4 m grid cell) where they had been with their pup prior to sep-arating. Pups spent close to 23% of their time at that location while their mothers wereaway foraging at sea. Female call rate was significantly higher in successful searches(i.e. those resulting in a reunion) compared to unsuccessful ones (Mann–Whitney testU = 72.5, P < 0.001). During reunions, 90% of the pups called prior to physical contactand search success varied significantly with their movement towards the mother(Fisher exact test P = 0.039). Lastly, mothers conducted olfactory investigations of theiroffspring during 97.6% of the reunions; they also conducted olfactory investigations of53.6% of the non-offspring that approached them. In sum, multiple sensory modalitiesappear to play important roles during O. flavescens mother–offspring reunion behavi-our, as has often been described but rarely quantified for other otariid species.
KEY WORDS: mother–offspring recognition, reunions, otariids, South American sealion, Otaria flavescens.
INTRODUCTION
In order to maximise their fitness, parents should avoid parental investment in unre-lated offspring (CLUTTON-BROCK 1991). To do so, parents must recognise their offspring or
3 Corresponding author: Micaela Trimble, The Natural Resources Institute, University of Manitoba,303-70 Dysart Road, Winnipeg, Manitoba R3T 2N2, Canada (E-mail: [email protected]).
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

234 M. Trimble and S.J. Insley
differentiate them from unrelated young during the period of parental care (HOLMES &SHERMAN 1983). In gregarious and precocial mammals, in which offspring are likely tointermingle with unrelated neonates, mothers tend to rapidly develop the ability toindividually recognise their offspring (CLUTTON-BROCK 1991). Gregarious and precocialdescribes most otariid pinnipeds (fur seals and sea lions). Their parent–offspring rec-ognition is known to be well developed although both descriptive and experimentalstudies have focused on the acoustic/auditory modality, despite the known importanceof visual and chemical information (reviewed in INSLEY et al. 2003).
Following parturition, female otariid pinnipeds alternate foraging trips at seawith nursing periods ashore (BOWEN 1991) and thus without the ability to find and dif-ferentiate their offspring, often in dense aggregations, their many reunions would beunsuccessful. One approach used to investigate the sensory modalities involved in thisprocess is to study the searching and reunion behaviour after mother–pup separations.Although searching and reunion behaviour has been anecdotally or qualitativelydescribed for several otariid species (e.g. BARTHOLOMEW 1959; VAZ-FERREIRA & ACHAVAL
1979; TRILLMICH 1981), only a few studies have quantified the behavioural details(GISINER & SCHUSTERMAN 1991; INSLEY 2001; DOBSON & JOUVENTIN 2003; PHILLIPS 2003).The results suggest that females often use multimodal sensory cues while searching fortheir pup: females appear to remember the last location where suckling took placeusing spatial cues; acoustic cues (i.e. individually distinctive calls) would act to locate,establish contact with, and attract the pup; and olfactory cues would be used for thefinal confirmation of pup identity. Visual cues appear to play an important role in thesequence of behaviours leading to reunion, but they seem more likely to be primingand localising devices rather than individual identifiers (INSLEY et al. 2003).
Initially, most authors agreed that otariid mothers took the more active role inrecognition and that pups reacted indiscriminately to any searching female (e.g.BARTHOLOMEW 1959; MCNAB & CRAWLEY 1975). However, further research in many ota-riid species indicated that the pups do play an important role in mother–offspringreunions (GISINER & SCHUSTERMAN 1991; INSLEY 2001; DOBSON & JOUVENTIN 2003; PHILLIPS
2003), even though their contribution might not be equal to that of their mothers(INSLEY 2001).
The South American sea lion (Otaria flavescens Shaw 1800) is a pinniped in thefamily Otariidae, distributed from the Atlantic coast near Rio de Janeiro (ca 23°S),Brazil, around the southern limit of South America, and up the Pacific coast, to Zorritos(ca 4°S), Peru (VAZ-FERREIRA 1981). The Uruguayan South American sea lion popula-tion has been estimated to be 12,000 individuals (PÁEZ 2006). It is a polygynous speciesthat breeds during the austral summer. Most births occur in January and after aperinatal period of about a week, mothers begin to alternate foraging trips at sea(ca 3 days) with nursing bouts ashore (ca 2 days). Lactation may last over a year, dur-ing which time mothers suckle only their own offspring (CAMPAGNA & LE BOEUF 1988).True allo-nursing and adoption of unrelated pups has not been observed; however,pups are known to sometimes steal milk from unaware females, especially duringtimes of food shortage (e.g. ENSO events), and often with severe consequences whendetected (SOTO et al. 2004). By experimentally separating mothers and pups andobserving their reunions at Isla de Lobos (Uruguay), VAZ-FERREIRA & ACHAVAL (1979)noted that acoustic and olfactory communication was important. FERNÁNDEZ-JURICIC etal. (1999) showed that both mother and pup calls of O. flavescens in Península Valdés(Argentina) were individually distinctive, and described a brief period of olfactory rec-ognition following vocal recognition. Both behaviours, calling and sniffing, occur athigh rates immediately after birth (FERNÁNDEZ-JURICIC et al. 1999; M. TRIMBLE pers.
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

Mother–offspring reunion in the South American sea lion 235
obs.), suggesting that this period might be important for memorising acoustic andolfactory cues.
The aim of this study was to provide quantitative descriptions of the searchingand reunion behaviour between mother and pup South American sea lions at Isla deLobos, Uruguay, during the first two months of a pup’s life. We tested the followingspecific hypotheses: (1) mothers use spatial cues when returning from sea by visitingthe last place where they were with their pup; (2) search success depends on the emis-sion of calls made by mothers and pups; (3) pup movement towards the mother isimportant for reunions, indicating that pups play an active role in maternal fidelity;and (4) at close range mothers always conduct olfactory confirmations of pup identity,indicating that chemical cues play a key role during identification.
MATERIALS AND METHODS
Study area and study period
Fieldwork was carried out at Isla de Lobos (35°01’S; 54°52’ W; Fig. 1(a),(b)), where thereare about 800 adult female and 350 adult male O. flavescens (E. PÁEZ pers. comm.). This is a 0.42km2 granite island is located 11 km from the city of Punta del Este in the Atlantic Ocean. Its long-est axis is ca 1230 m, oriented from south to north, and its maximum width is ca 690 m from westto east.
The study area was the zone known as “La Cachimba-El Mero” (Fig. 1(c)), where ca 80pups are born each summer (M. TRIMBLE pers. obs.), representing 20–25% of the total pupping atIsla de Lobos (E. PÁEZ pers. comm.). This zone, 75 m long and 25 m wide, is located at the
Fig. 1. — (a) South America map indicating the location of Isla de Lobos (Uruguay). (b) Aerial picture ofIsla de Lobos. The asterisk (*) indicates the study area (enlarged in c). (c) Zone “La Cachimba-El Mero”.
(a)
(b)
(c)
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

236 M. Trimble and S.J. Insley
north-north-western section of the island and consists of a flat rocky area with a slope of no morethan 10°. Near the northern edge of the colony there is a small beach of sand and gravel. TheSouth American fur seal, Arctocephalus australis, also breeds in this zone. Since fur seals aremuch more abundant, O. flavescens breeding groups are surrounded by them.
The study was conducted during the 2006 and 2007 breeding seasons (11–26 January, 8–21February and 8–21 March; 27 December–22 March respectively). Except when prevented byheavy rain, sampling was conducted every day between 07:00 and 19:00, totalling 1215 hours.
Individual identification
Mothers were paint-marked or identified by natural marks such as scars (especially in thesecond season) whereas pups were paint-marked or flipper-tagged in both foreflippers using All-flex No. 12 numbered plastic tags. Pups were also sexed during tagging.
During the first season we marked 11 mothers and 25 pups, including seven dyads (i.e.both members of the mother–pup pair marked; marked dyads were observed nursing prior to and/or after marking to confirm they were a mother–offspring pair). One additional mother was identi-fied by a scar. During the second season we marked 14 mothers and 26 pups, again includingseven marked dyads. Twenty-two additional mothers could be identified by natural marks, but nottheir pups. Considering both seasons together, there were 48 identified mothers (14 with markedpups) and 37 identified pups with non-identified mothers. The low number of mother–pup pairsmarked was due to the difficulty of marking the other member of the pair once the pup/motherhad been marked.
Data recording
We recorded a total of 120 searches conducted by 56 different mothers, which were eitheridentified females or mothers of marked pups. Due to the high number of identified pups withnon-identified mothers, searches initiated by unknown females were regularly recorded duringour daily observations. When such females were confirmed as mothers of a marked pup (i.e. nurs-ing occurred), the search was included in subsequent analyses. The only analysis for which thesereunions were not included was when determining the percentage of successful searches (thesesearches were all successful because, by definition, we were not able to determine whether anunmarked searching female was a marked pup’s mother until its search was successful). The like-lihood of having misidentified mother–pup pairs was considered very low; maternal behaviour inO. flavescens is only directed towards offspring (e.g. VAZ-FERREIRA & ACHAVAL 1979; VAZ-FERREIRA
1981; CAMPAGNA & LE BOEUF 1988). Despite a reasonable sample size of identifiable animals (i.e.marked or with natural marks) and substantial observer effort, many individuals were observedinfrequently for numerous reasons (e.g. movement out of the study area). In addition, the numberof marked dyads which could enable us to track the behaviour of both members of a pair was low.
Searches initiated by mothers were recorded by focal animal sampling (ALTMANN 1974),using an audio-tape recorder (Panasonic RQ-L11) for comments. The beginning of a search wasdefined as when a mother began vocalising and emitted at least two calls, either when returningfrom sea or during her time on land. Calls, throughout this study, are limited to only the affiliativevocalisations used in mother–pup communication (“PAC: pup attraction calls” emitted by mothersand “MAC: mother attraction calls” emitted by pups; FERNÁNDEZ-JURICIC et al. 1999; PHILLIPS
2003).The parameters recorded in each search were the duration (in minutes) from the first call
to the reunion or to the end of the search; mother call rate (number of calls/search duration);mother substrate sniffing (presence/absence); mother response to non-offspring (see below);number and success of non-offspring that attempted suckling (for allo-nursing to be classed assuccessful it had to last a minimum of 10 sec); and pup behaviour during the search (if it calledand/or moved towards the mother). Mother responses to non-offspring were classified as ignoring
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

Mother–offspring reunion in the South American sea lion 237
(mother behaviour did not change), sniffing and ignoring, sniffing and rejecting, rejecting with-out sniffing. Rejection behaviour was initially scored during 2006 as present or absent. During2007, rejection behaviour was broken down in more detail; defined as either threats (i.e. thefemale opened her mouth and/or emitted an aggressive vocalisation) or physical attacks (includ-ing physical contact, e.g. bites, head-darts, throwing; HARCOURT 1991; MILLER 1991; FERNÁNDEZ-JURICIC et al. 1999).
The importance of spatial cues was studied in 2007 using a grid with 4 × 4 m squares madeon an aerial photograph of the study area. Each day, scan samples were conducted every hourbetween 07:00 and 18:00 in order to record the location of each marked animal (ALTMANN 1974).This enabled us to determine whether mothers visited the last location where they had been withtheir pup before the separation (i.e. moved to the “last location”) and estimate how much timepups spent there during the periods of maternal absence. Given that females tend to stay in oneplace for much longer than 1 hr periods, the hourly scans should have been sufficient to recordthe majority of last location visits but would be conservative.
A search was considered to be successful when mother and pup reunited (i.e. establishedphysical contact and remained together after naso-nasal inspection) and/or nursing occurred(this was a criterion for searches with only one marked member of the dyad). In successfulsearches, reunions were described by recording which dyad member moved towards each other(mother, pup or both), the occurrence of pup calls prior to the reunion and the occurrence ofnaso-nasal inspection. This was possible for unmarked pups by recording the basic behaviour(i.e. movement towards or away, if they called) of every pup who was nearby the female until asearch was successful. During successful searches the post-reunion behaviour of the dyad wasrecorded for 10 min (only in the 2007 season). In each post-reunion minute we recorded theoccurrence of pup calls, number of mother calls, number of naso-nasal inspections, sucklingbehaviour, female substrate sniffing, and mother–non-offspring interactions.
Unsuccessful searches (i.e. when a mother did not find her pup) were divided into twogroups: (1) the female remained on land without calling for 10 min (PHILLIPS 2003); and (2) thefemale swam away from the study area. In both cases, the end of the search was the time at whichthe last call was emitted.
Statistical analyses
Unless stated, all data were used for each analysis. As noted and explained previously,mother–pup pairs with unmarked females were not included in the tally of search success. In thefew cases where more than one search was recorded per female, we analysed only the first inorder to avoid pseudo-replication. As the number of searches initiated during the stay on landwas low (n = 12), we focused most of our analyses (searching, reunion and post-reunion behav-iour) on the 54 searches of mothers returning from sea. No differences in searching behaviourwere found (or expected) between the 2006 and 2007 seasons (e.g. in search success, search dura-tion, mother call rate) and so these data were pooled.
The difference in searching behaviour was tested between from sea and on land searches(the two principal search contexts) in two ways: (1) a pooled comparison of means of from sea (n= 44; 10 females were excluded for which an on land search was also recorded) and on landsearches (n = 12) of different mothers; and (2) a paired comparison of 10 mothers with onesearch in each context.
To study the within-season variation in searching and reunion behaviour, we defined twoperiods of 32 days each: (1) Month 1, from 15 January to 15 February, which corresponds withthe first month of pup life; and (2) Month 2, from 16 February to 19 March. To determinewhether there was seasonal variation in the proportion of time (based on the number of scans)pups spent in the last location, analysis was restricted to pups with at least 10 observations perperiod.
Statistical analyses were conducted with Statistica 6.0 software. Non-parametric statisticaltests were used because the data violated the normality and variance homogeneity assumptions
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

238 M. Trimble and S.J. Insley
of parametric statistics (SOKAL & ROHLF 1981). All tests were two-tailed and significance in allcases was P < 0.05. Data are presented as median ± median absolute deviation (MAD; i.e. themedian of the set of differences between each data point and the median of the data).
RESULTS
Mothers and pups reunited in 58.3% (21/36) of the searches. However, afterunsuccessful searches, many females initiated new searches later the same day.Successful searches were significantly shorter in duration (Table 1), and this did notvary seasonally (Table 2).
Spatial cues
Seventy-four percent (14/19) of females returning from sea visited the last loca-tion where they had been with their pup before the separation, most of them (10/14)within the first 5 min of the search. Since the average number of squares encounteredduring searches was less than two (mean = 1.95, SD = 1.72, n = 19 searches), and givena total of 70 squares which are used by mother–pup pairs in the study area, the proba-bility of visiting any particular square by chance was just 2.86% (2/70). This holds trueif the probability of visiting all squares was equal (e.g. there was no preferential routeused by females). Although our dataset is insufficient to truly test this, our analysisindicated that no particular squares were visited more often than others. In contrast,visiting the last location did not appear to affect search success (Table 1). Further-more, although 43.8% (7/16) of females sniffed the substrate after arriving on land, this
Table 1.
Characteristics of successful (mother and pup reunited) and unsuccessful searches initiated byindividual O. flavescens mothers returning from sea (median ± MAD are shown for continuous variables,
* P < 0.05, ** P < 0.01).
VariableSuccessful searches
Unsuccessful searches
Test P
Duration (min) 6 ± 3 (n = 39) 13 ± 8 (n = 15)Mann–Whitney
test U = 138.50.003**
Mother call rate (calls/min)
4.25 ± 1.75 (n = 39) 1.65 ± 0.46 (n = 15)Mann–Whitney
test U = 72.5< 0.001**
Mother call rate; first 6 min only (calls/min)
4.67 ± 1.67 (n = 39) 2.35 ± 1.52 (n = 15)Mann–Whitney
test U = 1470.008**
No. mothers visited the last location
12/16 2/3 Fisher exact test 0.624
No. pups moved towards their mother
36/38 2/4 Fisher exact test 0.039*
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

Mother–offspring reunion in the South American sea lion 239
behaviour was not significantly associated with visiting the last location (5/12 whenthey visited it vs 2/4 when they did not, Fisher exact test P = 0.608). Substrate sniffingwas also frequently observed (12/25 or 48%) during the post-reunion 10 min period.
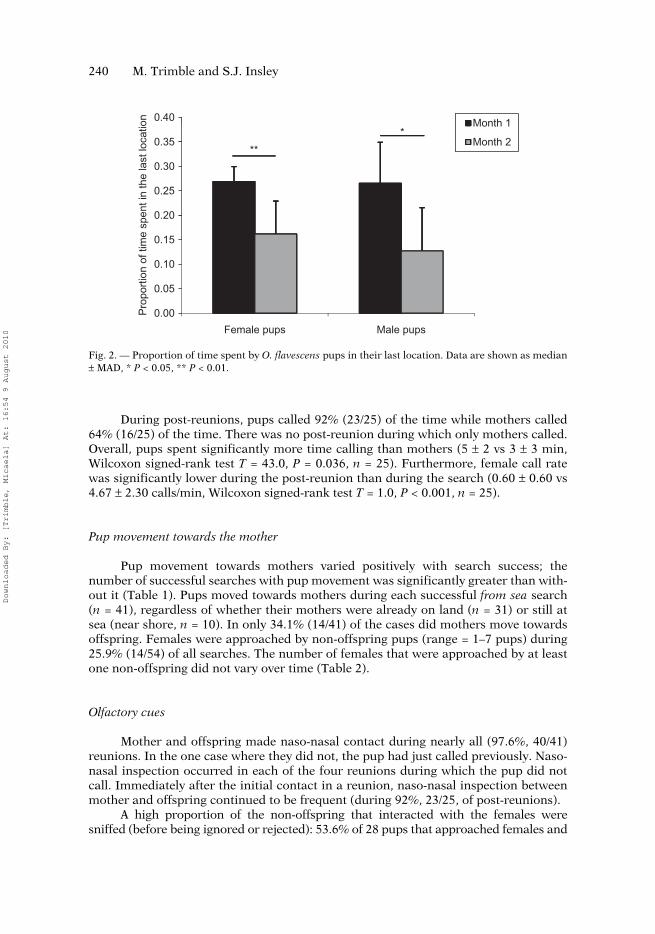
Time spent by pups in their last location varied between 11.4 and 48.8% whenconsidering the entire period (Months 1 and 2; 0.226 ± 0.051, n = 21). However, itdecreased significantly as pups matured (Table 2, Fig. 2). There was no sex differencein the time that pups spent in the last location (entire period: 0.230 ± 0.045 in females(1) vs 0.174 ± 0.032 in males (2i, Mann–Whitney test U = 40.0, P = 0.320, n1 = 12, n2 = 9;Month 1: U = 41.0, P = 0.356, n1 = 12, n2 = 9; Month 2: U = 30.0, P = 0.626, n1 = 10,n2 = 7; note that the same pups were observed in Months 1 and 2, thus Month 1 n +Month 2 n ≠ entire period n).
Mother and pup calls
Successful searches showed significantly higher mother call rates than unsuc-cessful ones (Table 1). Since call rate may decrease with time, and considering thatunsuccessful searches were longer than successful searches, we also compared mothercall rate over the first 6 min of calling in both categories (i.e. the median durationvalue of successful searches) and found out that differences were still significant(Table 1). There was no significant seasonal variation in mother call rate (Table 2).Pups called while moving in 90.2% (37/41) of all reunions. There was measurable tem-poral variation in pup reunion behaviour; some pups did not call before reunion in thefirst month while all called during the second month (Table 2).
Table 2.
Seasonal variation in searching and reunion behaviour between O. flavescens mothers and pups (median± MAD are shown for continuous variables, * P < 0.05, ** P < 0.01).
Variable Month 1 Month 2 Test P
Duration of successful searches (min)
6.5 ± 4.5 (n = 18) 5 ± 3 (n = 21)Mann–Whitney
test U = 1570.367
Mother call rate in successful searches (calls/min)
3.81 ± 1.69 (n = 18) 4.67 ± 1.33 (n = 21)Mann–Whitney
test U = 1610.430
Proportion of time pups spent in the last location (n = 17)
0.265 ± 0.059 0.151 ± 0.072Wilcoxon
signed-rank test T = 4.0
< 0.001**
No. mothers visited the last location
8/10 6/9Fisher exact
test0.444
No. pups moved without calling towards their mother
4/20 0/21Fisher exact
test0.048*
No. mothers approached by non-offspring
8/23 6/31 c254,1 0.201
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010
240 M. Trimble and S.J. Insley
During post-reunions, pups called 92% (23/25) of the time while mothers called64% (16/25) of the time. There was no post-reunion during which only mothers called.Overall, pups spent significantly more time calling than mothers (5 ± 2 vs 3 ± 3 min,Wilcoxon signed-rank test T = 43.0, P = 0.036, n = 25). Furthermore, female call ratewas significantly lower during the post-reunion than during the search (0.60 ± 0.60 vs4.67 ± 2.30 calls/min, Wilcoxon signed-rank test T = 1.0, P < 0.001, n = 25).
Pup movement towards the mother
Pup movement towards mothers varied positively with search success; thenumber of successful searches with pup movement was significantly greater than with-out it (Table 1). Pups moved towards mothers during each successful from sea search(n = 41), regardless of whether their mothers were already on land (n = 31) or still atsea (near shore, n = 10). In only 34.1% (14/41) of the cases did mothers move towardsoffspring. Females were approached by non-offspring pups (range = 1–7 pups) during25.9% (14/54) of all searches. The number of females that were approached by at leastone non-offspring did not vary over time (Table 2).
Olfactory cues
Mother and offspring made naso-nasal contact during nearly all (97.6%, 40/41)reunions. In the one case where they did not, the pup had just called previously. Naso-nasal inspection occurred in each of the four reunions during which the pup did notcall. Immediately after the initial contact in a reunion, naso-nasal inspection betweenmother and offspring continued to be frequent (during 92%, 23/25, of post-reunions).
A high proportion of the non-offspring that interacted with the females weresniffed (before being ignored or rejected): 53.6% of 28 pups that approached females and
Fig. 2. — Proportion of time spent by O. flavescens pups in their last location. Data are shown as median± MAD, * P < 0.05, ** P < 0.01.
0.15
0.00
0.05
0.10
0.20
0.25
0.30
0.35
0.40
Female pups
Pro
port
ion
of ti
me
spen
t in
the
last
loca
tion Month 1
Month 2**
*
Male pups
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

Mother–offspring reunion in the South American sea lion 241
91.7% of 24 pups approached by them. Eighteen females (33.3%) sniffed non-offspringduring the search.
Females responded differently to pups according to which approached which:60.7% of non-offspring pups that approached females were rejected (either threatenedor physically attacked) compared to 41.7% of pups that were approached by them(c2
47,2 = 7.64, P = 0.022; Fig. 3). Of 21 pups that were rejected during 40 searches, 76%were physically attacked while 24% were only threatened. Interestingly, only 36.4% ofthe approaching pups that were rejected retreated from the female after her first threator physical attack. The remaining pups were repeatedly attacked after failing toretreat. About 18% of approaching non-offspring attempted to suckle the females butall were physically attacked, resulting in zero allo-nursing events.
Search success rate was significantly higher when mothers had just returnedfrom sea (59.3%) compared to when they initiated the search on land (9.1%; Table 3).In addition, the call rate of mothers (when considering the entire search or just thefirst 6 min of calling) and the number of pups that moved towards the calling motherwere also significantly higher during from sea compared to on land searches (Table 3).
DISCUSSION
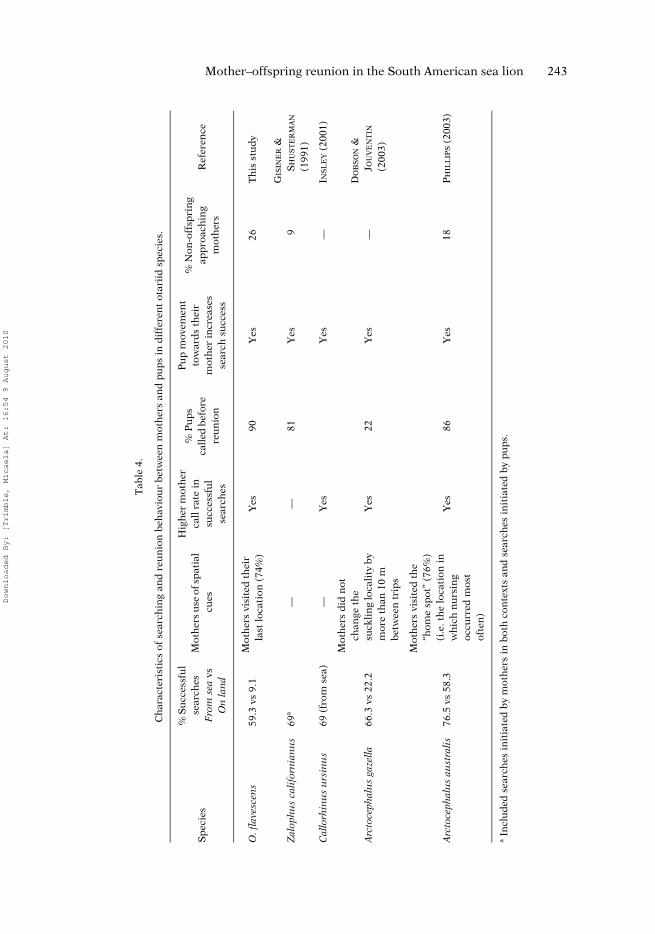
The first of our four working hypotheses was that mothers use spatial cues whenreturning from sea by visiting the last place where they were with their pup. Such use ofspatial cues by searching females has only been reported in two otariid species(A. australis and A. gazella; Table 4) and here O. flavescens females behaved similarly toA. australis females (PHILLIPS 2003). The high percentage of females that visited their lastlocation (74%), together with the lack of association between this behaviour and sub-strate sniffing (possible chemical cues), indicates the likely importance of spatial mem-ory and spatial cues during reunions. However, visiting the last location was not directlyrelated to search success, pointing to the importance of other factors such as pup behav-iour. That pups spent 23% of their time at their last location while their mothers were atsea is consistent with associating their mother’s return with this location. Thus bothmother and offspring appear to use spatial cues to increase the probability of reunion.
Our second hypothesis was that search success depends on the emission of callsmade by mothers and pups. The higher call rate of mothers found during successful
Fig. 3. — Proportion of O. flavescens female responses to non-offspring when pups approached them(n = 23 non-offspring) and when pups were approached by them (n = 24 non-offspring).
0.0
0.2
0.4
0.6
0.8
Sniffed and ignored
Pro
port
ion
of fe
mal
e re
spon
seto
non
-offs
prin
g
Pups approached females
Pups approached by females
Sniffed and rejected Rejected without sniffing
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

242 M. Trimble and S.J. Insley
searches and their lower call rate post-reunion supports this hypothesis. The highnumber of reunions in which pups called while moving (90%) would provide motherswith the opportunity to evaluate pup identity at a distance via acoustic cues. High callrates by mothers during successful searches have also been reported in other otariids(Table 4). In pups, O. flavescens called more frequently during reunions than the otherfour otariid species for which there are data (Table 4), implying the relative import-ance of acoustic cues. The variation among species may be a function of the differentacoustic environments. Conversely, the observation that some reunions were not pre-ceded by pup calls indicates that calling is not essential for reunions to be achieved.The same observation was made in A. gazella (DOBSON & JOUVENTIN 2003), A. australis(PHILLIPS 2003) and C. ursinus (S.J. INSLEY unpub.). Additionally, pup calls may alonebe sufficient for mothers to individually identify their offspring (FERNÁNDEZ-JURICIC et al.1999). This would agree with observations made in the present study; during one reun-ion the pup had called but naso-nasal investigation did not occur.
Our third hypothesis was that pup movement towards the mother is important forreunions, indicating that pups play an active role in maternal fidelity. As found in otherotariid species (Table 4), search success in O. flavescens varied significantly in accord-ance with pup movement towards the mother, showing that pups play an active role inreunions. In addition, pup responses to calling mothers suggest vocal discrimination.Selectivity in pup response is further supported by the significantly higher number offemales that were approached only by their offspring in comparison with the number offemales that were approached only by non-offspring. Our results also indicate that the
Table 3.
Searching and reunion behaviour between O. flavescens mothers and pups according to context (fromsea vs on land searches) (median ± MAD are shown for continuous variables, ** P < 0.01).
Unpaired comparison
Paired comparison Wilcoxon
signed-rank test (n = 10)
Variable From sea On land Test P T P
No. successful searches
16/27 1/11 Fisher exact test 0.005**
No. pups moved towards their mother
32/32 3/9 Fisher exact test <0.001**
No. mothers approached by non-offspring
9/44 1/12 Fisher exact test 0.309
Duration (min)6.5 ± 3.5
(n = 44)12 ± 9 (n = 12)
Mann–Whitney test U = 193.5
0.159 17.0 0.515
Mother call rate (calls/min)
3.60 ± 1.89 (n = 43)
1.39 ± 0.80 (n = 12)
Mann–Whitney test U = 111
0.003** 7.0 0.067
Mother call rate; first 6 min only (calls/min)
3.83 ± 1.67 (n = 39)
1.58 ± 1.08 (n = 15)
Mann–Whitney test U = 126
0.007**
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010
Mother–offspring reunion in the South American sea lion 243
Tab
le 4
.
Ch
arac
teri
stic
s of
sea
rch
ing
and
reu
nio
n b
ehav
iou
r b
etw
een
mot
her
s an
d p
up
s in
dif
fere
nt
otar
iid
sp
ecie
s.
Sp
ecie
s
% S
ucc
essf
ul
sear
ches
F
rom
sea
vs
On
lan
d
Mot
her
s u
se o
f sp
atia
l cu
es
Hig
her
mot
her
ca
ll r
ate
in
succ
essf
ul
sear
ches
% P
up
s ca
lled
bef
ore
reu
nio
n
Pu
p m
ove
men
t to
war
ds
thei
r m
oth
er i
ncr
ease
s se
arch
su
cces
s
% N
on-o
ffsp
rin
g ap
pro
ach
ing
mo
ther
sR
efer
ence
O. f
lave
scen
s59
.3 v
s 9.
1M
oth
ers
visi
ted
th
eir
last
loc
atio
n (
74%
)Y
es90
Yes
26T
his
stu
dy
Zal
oph
us
cali
forn
ian
us
69a
——
81Y
es9
GIS
INE
R &
S
HU
ST
ER
MA
N
(199
1)
Cal
lorh
inu
s u
rsin
us
69 (
fro
m s
ea)
—Y
esY
es—
INS
LE
Y (
2001
)
Arc
toce
phal
us
gaze
lla66
.3 v
s 22
.2
Mot
her
s d
id n
ot
chan
ge t
he
suck
lin
g lo
cali
ty b
y m
ore
th
an 1
0m
b
etw
een
tri
ps
Yes
22Y
es—
DO
BS
ON
&
JOU
VE
NT
IN
(200
3)
Arc
toce
phal
us
aust
rali
s76
.5 v
s 58
.3
Mo
ther
s vi
site
d t
he
“hom
e sp
ot”
(76%
) (i
.e. t
he
loca
tion
in
w
hic
h n
urs
ing
occu
rred
mos
t of
ten
)
Yes
86Y
es18
PH
ILL
IPS (
2003
)
a In
clu
ded
sea
rch
es i
nit
iate
d b
y m
oth
ers
in b
oth
con
text
s an
d s
earc
hes
in
itia
ted
by
pu
ps.
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

244 M. Trimble and S.J. Insley
pup’s active role changes over time, becoming more vocal with increased mobility. Wefound that pup site fidelity to their last location decreased as their size and mobilityincreased, the frequency of calling while moving was higher in the second month, andpups moved towards mothers regardless of age. In A. australis pups (5 to 35 days old),those that only moved during reunions tended to be younger, while those pups that onlycalled during reunions tended to be older (PHILLIPS 2003). Z. californianus pups werealso more actively involved in reunions as they grew older: the relative proportion ofreunions without pup movement declined from 31.8% when they were less than2 weeks old, to 8.9% when they were 4–6 weeks old (GISINER & SCHUSTERMAN 1991).
Our fourth hypothesis was that at close range mothers always conduct olfactory con-firmations of pup identity, indicating that chemical cues play a key role during identifica-tion. This hypothesis was supported by our observations of very high frequencies of naso-nasal inspection during reunions (97.6%), and that most approaching non-offspring weresniffed by searching females (53.6%). The four cases in which mothers sniffed andaccepted offspring that had not yet called suggest that olfactory cues alone are sufficientfor offspring recognition by mothers. Although either sensory modality (olfaction andacoustic/audition) may alone be sufficient for recognition, they were usually used together,underscoring the importance of offspring recognition and the high cost of maternal care.Olfactory cues appear to be important during mother–offspring recognition in most pin-niped species where recognition has been studied; naso-nasal inspection occurs duringmaternal reunions and females sniff approaching non-offspring (e.g. DOBSON & JOUVENTIN
2003; PHILLIPS 2003). Olfaction in pinnipeds, as with most mammals (LÉVY et al. 2004),appears to be well developed for use in air although its functional role in recognition andother aspects of communication is largely untested (INSLEY et al. 2003).
Finally, some of our findings are relevant to understanding the cost of recognitionby both mothers and pups. Search success in O. flavescens found during the present studywas lower than what has been found in other otariids (Table 4). These differences couldreflect the difficult circumstances faced by females at Isla de Lobos caused by both biotic(e.g. high animal densities, sympatric presence of A. australis) and abiotic (e.g. high ambientnoise, convoluted rocky substrate, high wind) factors. However, all pups and motherseventually reunited during our observations, indicating that mother–offspring recogni-tion is not deficient. As has been found in other otariids (e.g. INSLEY 2001; DOBSON &JOUVENTIN 2003), unsuccessful searches took significantly more time than successfulones, indicating an increased search cost to mothers when they do not find their pups.The risk to a pup of being injured by non-mothers can also represent a substantial cost.Non-offspring pups approached only 25.9% of the searching females that were returningfrom sea, probably due to the risk of being injured. We found this risk to be quite evident,with the majority of rejected pups receiving physical attacks from non-mothers. However,this was not consistent with the high number of pups that did not retreat after the firstthreat or physical attack from the female. Pup responses could be related to their level ofhunger, which in turn might be an important determinant of search success (CHARRIER etal. 2002). Risk-taking by pups appears to peak during times of food shortage (e.g. ENSOevents) and appears to be when pups are most likely to approach non-mothers (SOTO et al.2004). However, it is important to make the distinction between milk stealing (observedin this species; SOTO et al. 2004) and allo-nursing (not observed in this species); allo-nursing is thought to confer selective advantage via several possible means (e.g. experi-ence, kin selection, milk evacuation) in a number of species, notably primates, and canco-exist with well-developed parent–offspring behaviour (ROULIN 2002).
To sum, as previously suggested in other otariid species, our main results indic-ate that a combination of sensory modalities is involved in mother–offspring reunions
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

Mother–offspring reunion in the South American sea lion 245
in O. flavescens. Spatial cues are initially important when the mother returns from thesea. Next, acoustic cues are used by both dyad members, first at long range and then atshort range. Finally, the process is completed using olfactory cues to confirm the pup’sidentity. The relative importance of each sensory modality, as well as possible asym-metries in mother–offspring recognition, needs to be addressed experimentally. Suchexperiments should consider the effect of search context and pup age.
ACKNOWLEDGEMENTS
We are grateful to L. Olivera, N. Veiga, W. Melo and M. Casella for their help in marking theanimals and for giving us advice during our stay on the island. We would also like to thank G. Arévalo,M.J. José Benítez, E. Cawen, V. Franco Trecu, R. Frau, E. Görke, M. Ihara, C. Jaureguy, H. Katz,P. Mai, G. Martinez, S. Noya, E. Páez, M. Quevedo, M. Rivas and V. Samsa for their assistance in thefield. DINARA (National Direction of Aquatic Resources) provided us with permits (No. 264/2005 andNo. 705/2006) and logistical support. Marking procedures were also approved by the Honorary Com-mission of Animal Experimentation (CHEA, Uruguay, No. 01/02/2006). We thank E. Páez and A. Fer-reira for discussing details about data analysis. M. Trimble received funding from SZU (UruguayanZoological Society, Eduin Palerm Award) and CSIC (Sectorial Commission of Scientific Research,Uruguay) and equipment was provided by Idea Wild (USA). PEDECIBA (Programme for the Develop-ment of Basic Sciences, Uruguay) and ANII (National Agency of Research and Innovation, Uruguay)provided M. Trimble with a MSc scholarship. We thank I. Charrier for her useful comments on earlyversions of this manuscript. Two anonymous referees also made valuable suggestions.
REFERENCES
ALTMANN J. 1974. Observational study of behaviour: sampling methods. Behaviour 49: 227–265.BARTHOLOMEW G.A. 1959. Mother-young relations and the maturation of pup behaviour in the
Alaska fur seal. Animal Behaviour 7: 163–171.BOWEN W.D. 1991. Behavioural ecology of pinniped neonates, pp. 66–127. In: Renouf D., Ed. The
behaviour of pinnipeds. London: Chapman & Hall.CAMPAGNA C. & LE BOEUF B. 1988. Reproductive behaviour of southern sea lions. Behaviour 104:
233–261.CHARRIER I., MATHEVON N., HASSNAOUI M., CARRARO L. & JOUVENTIN P. 2002. The subantarctic fur
seal pup switches its begging behaviour during maternal absence. Canadian Journal ofZoology 80: 1250–1255.
CLUTTON-BROCK T.H. 1991. The evolution of parental care. Princeton: Princeton University Press.DOBSON F.S. & JOUVENTIN P. 2003. How mothers find their pups in a colony of Antarctic fur seals.
Behavioural Processes 61: 77–85.FERNÁNDEZ-JURICIC E., CAMPAGNA C., ENRIQUEZ V. & ORTIZ C.L. 1999. Vocal communication and
individual variation in breeding South American sea lions. Behaviour 136: 495–517.GISINER R. & SCHUSTERMAN R.J. 1991. California sea lion pups play an active role in reunions with
their mothers. Animal Behaviour 41: 364–366.HARCOURT R. 1991. Maternal aggression in the South American fur seal in Peru. Canadian Journal
of Zoology 70: 320–325.HOLMES W.G. & SHERMAN P.W. 1983. Kin recognition in animals. American Science 71: 46–55.INSLEY S.J. 2001. Mother-offspring vocal recognition in northern fur seals is mutual but assymet-
rical. Animal Behaviour 61: 129–137.INSLEY S.J., PHILLIPS A.V. & CHARRIER I. 2003. A review of social recognition in pinnipeds. Aquatic
Mammals 29: 181–201.LÉVY F., KELLER M. & POINDRON P. 2004. Olfactory regulation of maternal behavior in mammals.
Hormones and Behavior 46: 284–302.
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010

246 M. Trimble and S.J. Insley
MCNAB A.G. & CRAWLEY M.C. 1975. Mother and pup behaviour of the New Zealand fur seal, Arcto-cephalus forsteri (Lesson). Mauri Ora 3: 77–88.
MILLER E.H. 1991. Communication in pinnipeds, with special reference to non acoustic signaling,pp. 128–235. In: Renouf D., Ed. The behaviour of pinnipeds. London: Chapman & Hall.
PÁEZ E. 2006. Situación de la administración del recurso lobos y leones marinos en Uruguay[Situation of the administration of fur seals and sea lions in Uruguay], pp. 577–584. In:Menafra R. et al., Eds. Bases para la conservación y el manejo de la costa uruguaya [Basisfor the conservation and management of the Uruguayan coast]. Montevideo: Vida SilvestreUruguay.
PHILLIPS A.V. 2003. Behavioral cues used in reunions between mother and pup South Americanfur seals. Journal of Mammalogy 84: 524–535.
ROULIN A. 2002. Why do lactating females nurse alien offspring? A review of hypotheses andempirical evidence. Animal Behaviour 63: 201–208.
SOKAL R.R. & ROHLF F.J. 1981. Biometry, 2nd ed. San Francisco: W.H. Freeman & Company.SOTO K.H., TRITES A.W. & ARIAS-SCHREIBER M. 2004. The effects of prey availability on pup mor-
tality and the timing of birth of South American sea lions (Otaria flavescens) in Perú. Jour-nal of Zoology, London 264: 419–428.
TRILLMICH F. 1981. Mutual mother-pup recognition in Galapagos fur seals and sea lions: cuesused and functional significance. Behaviour 78: 21–42.
VAZ-FERREIRA R. 1981. South American sea lion Otaria flavescens (Shaw, 1800), pp. 39–65. In:Ridgway S.H. & Harrison R.J., Eds. Handbook of marine mammals, 2nd ed. Vol. 1: Thewalrus, sea lions, fur seals and sea otter. London: Academic Press.
VAZ-FERREIRA R. & ACHAVAL F. 1979. Relación y reconocimiento materno-filial en Otaria flavescens(Shaw) “lobo de un pelo”, y reacciones de los machos subadultos ante los cachorros [Rela-tionship and mother-pup recognition in Otaria flavescens (Shaw) “Southern sea lion”, andthe reactions of subadult males to the pups]. Acta Zoologica Lilloana 35: 295–302.
Downloaded By: [Trimble, Micaela] At: 16:54 9 August 2010




















