
Mother-young relationships in captive ungulates: variability and clustering
12
Anim .Behao .,1986,34,134 145 Mother youngrelationshipsincaptiveungulates : variabilityandclustering KATHERINERALLS*,KARLKRANZ*&BARBARALUNDRIGANt *DepartmentofZoologicalResearch,NationalZoologicalPark,SmithsonianInstitution,Washington,D .C . 20008,U.S.A . tMuseum of Zoology,University of Michigan,AnnArbor,Michigan48109,U.S.A . Abstract. Werecordedfourmeasuresofmother youngassociationandthepercentageoftimetheyoung spentlyingduringthefirstweekafterbirthfor59mother youngpairsbelongingto22speciesandseven familiesofungulates .Themeasuresofmother youngassociationwerepositivelycorrelatedwitheach otherandnegativelycorrelatedwiththepercentageoftimetheyoungspentlying .Threeclusteranalyses, basedonvariouscombinationsofmeasures,separatedthebovidsintotwogroupsrecognizableas `followers'and'hiders' .Whenthedataonall22specieswereconsidered,fiveconsistentspeciesgroups occurredinthethreeclusteranalyses .Thetraditionalfollower hiderdichotomyprovidesarough characterizationofmother youngrelationshipsinmanyungulates,butdoesnotadequatelydescribethe behaviouralvariationpresent .Byusingstandardizedquantitativemeasures,wewereabletocharacterize thisvariationandtoestablishaframeworkforfuturestudy . Studiesofungulatemother youngrelationships haveledtothegeneralizationthattheyoungmay beroughlyclassifiedintotwogroups :thosethat initiallystayclosetotheirmothersandthosethat spendthefirstpartoftheirlivesatsomedistance fromtheirmothers,oftenconcealedinvegetation . Thesestrikingbehaviouraldifferencesareapparent toanyonewhohasobservedavarietyofungulate species;thefirstformaldescriptionsappeared duringthe1960s(Fraser1968 ;Talbotetal .1965 ; Walther1961,1964,1966) .Walther(1965)termed thesegroups'NachfolgeTypus'(follower type) and'Abliege Typus'(lying apart type),respect ively .InEnglishthetwobehaviouralpatternsare usuallyreferredtoas`following'and`hiding'(Lent 1974 ;Leuthold1977) . TheobservationsonwhichWaltherbasedthe behaviouraldichotomyweremadeonseveral speciesofAfricanantelopes,andhissubsequent work(summarizedinWalther1979)alsoconcen tratedonbovids .However,theconceptshavebeen appliedtoungulatesinotherfamiliessuchas cervidsandequids(Lent1974 ;Leuthold1977) . Thereareanumberofhypothesesinthelitera tureregardingtheadaptivesignificanceoffollow ingandhiding .Weusetheterms`following'and `hiding'forthetwotypesofspatialrelationship withoutimplicationsastothebehaviouralmecha nismsbywhichtheserelationshipsaremaintained . Followinghasbeenviewedasastrategyfor 134 avoidingpredatorsinopenhabitat(Lent1974 ; Estes1976)andisoftenassociatedwithmigratory habitsand/ordefenceoftheyoungagainstpreda tors .Hidingisthoughttoreducepredationriskin closedhabitats(Lent1974 ;Estes1976),butmay alsoprotectyoungfromintraspecificaggression (Lent1974)orcompetition(Murdocketal .1983) andhelpthemtoconserveenergyandbodywater (Langman1977) . Althoughtheterms'hider'and`follower'are frequentlyusedintheungulateliterature,the adequacyofthisclassificationsystemhasnever beenevaluated .ThecollectionsoftheNational ZoologicalPark(NZP),Washington, D .C ., and theMetropolitanTorontoZoo(MTZ),Scarbor ough,Ontario,provideduswithanopportunityto compareawiderangeofspeciesusingstandardized methods .Thebroadspecies typicalpatternsofthe mother youngrelationshipareknowntobe retainedincaptivity ;indeed,theoriginaldescrip tionsofthefollowerandhidergroupswerelargely basedonobservationsofcaptiveanimals(Walther 1964,1965,1966) . Theprincipalgoalsofourstudyweretoassess theadequacyofthefollower hidermodel, measurethevariabilityamongspeciesreportedly belongingtoeachcategoryanddeterminethe extenttowhichintermediatetypesmightexist, bothwithintheBovidaeandacrossungulatesasa whole .
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Mother-young relationships in captive ungulates: variability and clustering

Anim. Behao ., 1986, 34, 134-145
Mother-young relationships in captive ungulates :variability and clustering
KATHERINE RALLS*, KARL KRANZ* & BARBARA LUNDRIGANt*Department of Zoological Research, National Zoological Park, Smithsonian Institution, Washington, D .C .
20008, U.S .A .tMuseum of Zoology, University of Michigan, Ann Arbor, Michigan 48109, U.S .A .
Abstract. We recorded four measures of mother-young association and the percentage of time the youngspent lying during the first week after birth for 59 mother-young pairs belonging to 22 species and sevenfamilies of ungulates . The measures of mother-young association were positively correlated with eachother and negatively correlated with the percentage of time the young spent lying . Three cluster analyses,based on various combinations of measures, separated the bovids into two groups recognizable as`followers' and 'hiders' . When the data on all 22 species were considered, five consistent species groupsoccurred in the three cluster analyses . The traditional follower-hider dichotomy provides a roughcharacterization of mother-young relationships in many ungulates, but does not adequately describe thebehavioural variation present . By using standardized quantitative measures, we were able to characterizethis variation and to establish a framework for future study .
Studies of ungulate mother-young relationshipshave led to the generalization that the young maybe roughly classified into two groups : those thatinitially stay close to their mothers and those thatspend the first part of their lives at some distancefrom their mothers, often concealed in vegetation .These striking behavioural differences are apparentto anyone who has observed a variety of ungulatespecies; the first formal descriptions appearedduring the 1960s (Fraser 1968 ; Talbot et al . 1965 ;Walther 1961, 1964, 1966) . Walther (1965) termedthese groups 'Nachfolge Typus' (follower-type)and 'Abliege-Typus' (lying-apart-type), respect-ively . In English the two behavioural patterns areusually referred to as `following' and `hiding' (Lent1974; Leuthold 1977) .The observations on which Walther based the
behavioural dichotomy were made on severalspecies of African antelopes, and his subsequentwork (summarized in Walther 1979) also concen-trated on bovids . However, the concepts have beenapplied to ungulates in other families such ascervids and equids (Lent 1974 ; Leuthold 1977) .
There are a number of hypotheses in the litera-ture regarding the adaptive significance of follow-ing and hiding . We use the terms `following' and`hiding' for the two types of spatial relationshipwithout implications as to the behavioural mecha-nisms by which these relationships are maintained .Following has been viewed as a strategy for
134
avoiding predators in open habitat (Lent 1974 ;Estes 1976) and is often associated with migratoryhabits and/or defence of the young against preda-tors . Hiding is thought to reduce predation risk inclosed habitats (Lent 1974 ; Estes 1976), but mayalso protect young from intraspecific aggression(Lent 1974) or competition (Murdock et al . 1983)and help them to conserve energy and body water(Langman 1977) .
Although the terms 'hider' and `follower' arefrequently used in the ungulate literature, theadequacy of this classification system has neverbeen evaluated. The collections of the NationalZoological Park (NZP), Washington, D.C ., andthe Metropolitan Toronto Zoo (MTZ), Scarbor-ough, Ontario, provided us with an opportunity tocompare a wide range of species using standardizedmethods. The broad species-typical patterns of themother-young relationship are known to beretained in captivity ; indeed, the original descrip-tions of the follower and hider groups were largelybased on observations of captive animals (Walther1964, 1965, 1966) .
The principal goals of our study were to assessthe adequacy of the follower-hider model,measure the variability among species reportedlybelonging to each category and determine theextent to which intermediate types might exist,both within the Bovidae and across ungulates as awhole .
Rails et al . : Mother-young relations in ungulates
135
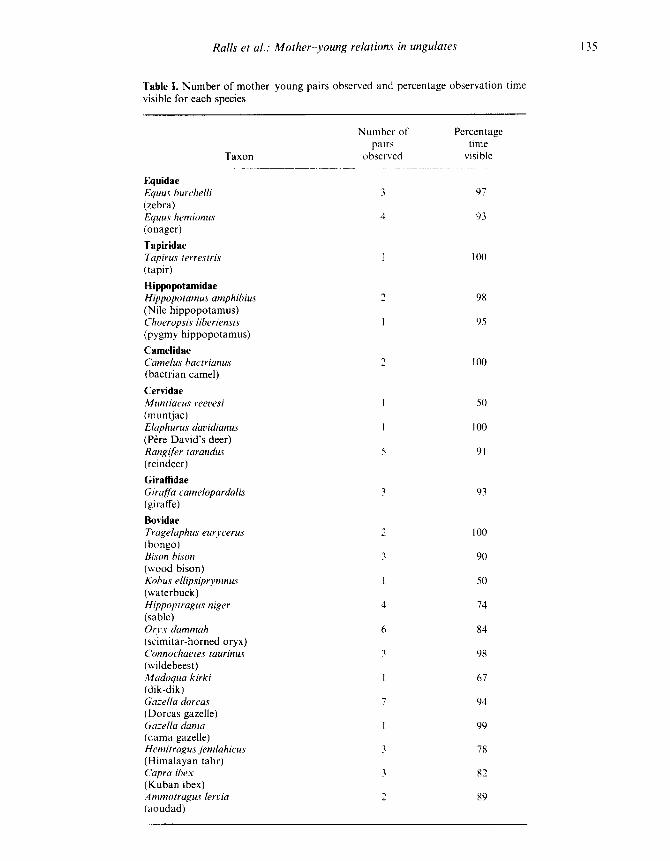
Table 1 . Number of mother-young pairs observed and percentage observation timevisible for each species
Taxon
Number ofpairs
observed
Percentagetime
visible
EquidaeEquus hurchelli 3 97(zebra)Equus hernionus 4 93(onager)TapiridaeTapirus terrestris I 100(tapir)HippopotamidaeHippopotamus amphibius 98(Nile hippopotamus)Choeropsis liberiensis I 95(pygmy hippopotamus)CamelidaeCamelus bactrianus 100(bactrian camel)CervidaeMuntiacus reevesi 1 50(muntjac)Elaphurus davidianus 1 100(Pere David's deer)Rangifer tarandus 5 91(reindeer)GiraffidaeGirnffa camelopardalis I 93(giraffe)BovidaeTragelaphus eurycerus 2 100(bongo)Bison bison 2 90(wood bison)Kobus ellipsiprymnus 1 50(waterbuck)Hippoporagus niger 4 74(sable)Orrx dammah 6 84(scimitar-horned oryx)Connochaetes taurinus 1 98(wildebeest)Madoqua kirki 1 67(dik-dik)Ga:ella dorcas 7 94(Dorcas gazelle)Ga:ella dama 1 99(dama gazelle)Hemitragus jemlahicus 78(Himalayan tahr)Capra ibex 82(Kuban ibex)Ammotragus lervia 2 89(aoudad)

1 3 6
METHODS
Fifty-nine mother-young pairs representing 22species and seven families were observed for a totalof over 1400 h between 1977 and 1982 (Table I) .Most pairs were observed either at NZP or theNZP's Conservation and Research Center (CRC)in Front Royal, Virginia . The bison (Bison bison),tahr (Hemitragus jemlahicus), aoudad (Ammotra-gus lervia) and ibex (Capra ibex) were observed atMTZ and the waterbuck (Kobus ellipsiprymnus) atKing's Dominion in Doswell, Virginia . Enclosuresizes and the number of conspecifics present withthe mother-young pairs varied . The social group-ings in captivity resembled those in the wild .Monogamous species such as dik-dik (Madoquakirki) (Hendrichs & Hendrichs 1971) were kept inpairs and gregarious species, except for the Nilehippopotamus (Hippopotamus amphibius), werekept in herds .
Each pair was observed for at least 6 weeks .However, we considered only first week data in theanalysis reported here because hiding behaviour insome species may begin to decrease in the secondweek after birth (Walther 1979). First week datawere collected by 15 experienced observers, eachassigned to one or more mother-young pairs .Although differences between observers may haveinfluenced our results, we believe this was not aserious problem. Inter-observer differences shouldhave been smaller in the first week, when experi-enced observers collected data, than in subsequentweeks, when a much larger group of volunteersmade many of the observations . Our analysis of themean weekly coefficients of variation over theentire 8-week study period showed that even thesewere not related to the number of observerscontributing data for a particular species (Rails etal . 1983) .
An attempt was made to observe each pair for I hper day, three or more times during week 1, butscheduled observations were occasionally missed .Most observations were made during periods whenanimals were usually most active, either in themorning, between 0800 and 1000 hours, or in lateafternoon between 1600 and 1800 hours . Fifty-sixof the young were singletons and three were twins(one aoudad and two ibex) . In the latter cases, onetwin was chosen for observation throughout thestudy. Data were recorded on a simple check sheetwhich was revised several times to eliminate
Animal Behaviour, 34, 1
measures on which observers could not reach ahigh level of agreement (at least 90°0) .
The most fundamental behavioural differencebetween followers and hiders is usually consideredto be the degree of association between mother andyoung during the first days after birth . We mea-sured degree of association in four ways . Two ofour measures, the percentage of 1-min intervals themother and young were in contact (Contact) andthe percentage of 1-min intervals the mother andyoung were within I mother-length of one another(< I ML), were suggested by Lent's (1974) modelcontrasting the behaviour of followers and hiders .We considered mother and young to be in contactwhen the two were lying or standing with any partof their bodies touching. Recording the averagedistance between mother and young (as suggestedby Lent's model) proved impractical becauseobserver estimates of absolute distance variedwidely. Thus, we recorded distance in 'mother-lengths', a different absolute value for each species .Although he presented no quantitative data ..
Walther (1979) suggested that hider and followerspecies could be easily distinguished by the distancebetween mother and young while both animalswere lying . Our third measure, number of minutesmother and young both lying within I mother-length/total minutes both mother and young lying(Walther's measure), was based on this suggestion .
The final measure of association, the percentageof 1-min intervals the mother was the closestanimal to her young (MCA), was designed toestimate the extent to which a mother was thenearest neighbour to her young. MCA was cor-rected for the number of animals in the enclosure[(observed %MCA-expected %MCA)/(100-expected %MCA) x 100]. This index (MCA index)was scaled so that it would have the same range (0-100) as the other measures .Focal mother-young pair sampling was com-
bined with instantaneous sampling at the end ofeach minute for all measures except Contact .Because some observers had difficulty ignoringinteresting behaviours such as nursing or sniffing,we scored Contact on a 1-0 basis regardless ofwhere in the 1-min interval it occurred . Eachmeasure was expressed as the percentage of I-minintervals during which it was recorded, e .g . < IML=(number of minutes mother and young wererecorded as within one mother-length of eachother/total minutes during which data wererecorded) x 100. We used the number of minutes

during which data were recorded as a denominator,rather than the minutes of observation time,because the mother and young were not alwaysvisible for the entire observation period (Table 1) .
We performed cluster analyses (after Sokal 1961)to obtain a graphic representation of the relation-ships among species. Three cluster analyses wereperformed, each employing a different combina-tion of behavioural measures . The three combina-tions of measures were : (I) only the measures ofmother-young association suggested by the workof Lent (1974) and Walther (1979), i .e. < I ML,Walther's measure and Contact ; (2) all fourmeasures of mother-young association, i .e . thethree above plus MCA index ; and (3) these fourmeasures plus the percentage of 1-min intervals theyoung spent lying (Young Lying) .
The clustering procedure required that we esti-mate missing data points . Data on one measurewere missing for eight species and on threemeasures for one species . The two most common
Rails et al . : Mother-young relations in ungulates
Table 11 . Means and standard deviations for five behavioural measures recorded during the first week after birth
Underlined values are estimates for missing data .Standard deviations were calculated only when three or more pairs were observed .* Members of the family Bovidae.t 'Inactive' rather than 'lying' scored .
1 3 7
reasons for missing data were : (1) the MCA indexcould not be calculated because the mother andyoung were the only animals present in the enclo-sure, and (2) Walther's measure could not becalculated because mother and young were neverobserved both lying. Estimates for missing datapoints were calculated by a least squares regressionanalysis using the Spearman's rank correlation ofcoefficients between the four measures of motheryoung association described above . Estimates ofover 100 were recorded as 100 .
A computer program in the Michigan InteractiveData Analysis System (MIDAS) was used toobtain the cluster diagrams . This program calcu-lates Euclidean distances between species and usesan averaging technique to form clusters in whichspecies (and groups of species) linked at lowdistance values are more similar on the measuresused than those linked at high distance values . Thecophenetic correlation (a measure of agreementbetween the similarity values implied by the den-
Species< I ML
mean (SD)
Walther'smeasure
mean (so)Contactmean (SD)
MCA indexmean (so)
YoungLying
mean (SD)
Hippopotamust 100 100 88 100 61Pygmy hippopotamust 100 100 58 100 91Camel 99 100 32 100 58Onager 98 (3) 100 (-) 36 (2) 100 (0) 32 (11)Zebra 94 (2) 100 (-) 25 (13) 99 (2) 10 (11)Wildebeest* 92 (11) 100 (0) 20 (11) 85 (7) 63 (20)Reindeer 76 (13) 94 (48) 32 (16) 95 (5) 76 (16)Tapir 75 100 18 91 73Bison* 72 (19) 72 (43) 20 (6) 73 (2) 82 (12)Tahr* 64 (13) 71 (43) 36 (11) 69 (6) 64 (9)Aoudad* 64 100 32 65 73Giraffe 56 (20) 58 (-) 8 (8) 61 (0) 18 (5)Ibex* 54 (12) 65 (17) 40 (16) 67 (7) 70 (26)Bongo* 45 46 8 85 74Oryx* 35 (30) 32 (41) 12 (7) 69 (31) 84 (13)Pere David's deer 27 0 5 41 79Dama gazelle* 24 0 2 68 69Dik-dik* 19 0 1 0 85Dorcas gazelle* 12 (4) 0 (0) 4 (4) 45 (18) 82 (8)Sable* 3 (5) 0 (0) 0 (0) 29 (60) 99 (2)Waterbuck* 0 0 0 7 100Muntjac 0 0 0 3 95

1 3 8
Table 111. Spearman's rank correlation coefficients between five behavioural measuresrecorded during the first week after birth
Measures
1
2
3
4
l2
0-94013
0-86594
0.92215
-0-6256
drogram and those of the original similaritymatrix) was used to evaluate clustering perfor-mance.
RESULTS
Means and standard deviations for five beha-vioural measures recorded during the first weekafter birth for the 22 species are given in Table II .Correlation coefficients between the five measuresof mother-young behaviour are shown in TableIII. The four measures of mother-young associ-ation are positively correlated with one another(P<0-01) and negatively correlated with the per-centage of time the young spent lying (P < 0 .01) .These correlation coefficients were also calculatedfor bovids and non-bovids separately . The result-
Animal Behaviour, 34, 1
0-82860-8484
0-7795-0-5399
-0-5375
-0-5766
Measures: (() < 1 ML ; (2) Walther's measure ; (3) Contact ; (4) MCA index; and (5) YoungLying .All values are significant at P<0-01 .
Table IV . Spearman's rank correlation coefficients between five behavioural measures recorded duringthe first week after birth for bovids and non-bovids
(1) < I ML ; (2) Walther's measure ; (3) Contact ; (4) MCA index; and (5) Young Lying .* P<0 . 05; ** P<0 . 01 .
ing matrices (Table IV) reflect the same trends asshown in Table III . However, the correlationcoefficients between Young Lying and the fourmeasures of mother-young association are lowerand non-significant for the non-bovids .
Relationships Among Species : Walther's MeasureIn order to evaluate Walther's idea that follower
and hider species can be distinguished by thedistance between mother and young while bothanimals are lying, we first examined relationshipsamong the bovid species in terms of Walther'smeasure. It clearly separates wildebeest (Conno-chaetes taurinus) and aoudad, which scored 100' ,from dama gazelle (Gazella dama), Dorcas gazelle(Gazella dorcas), dik-dik (Madoqua kirki), sable
Measures 1 2 3 4
Bovids12 0 . 9458**3 0 . 8409** 0 . 8427*4 0 . 7587** 0 . 6678* 0-55425 -0 . 7411** -0-5664 -07063* -06311*
Non-bovids12 0 . 8515**3 0 .9576** 0 . 7879**4 09727** 0 . 8788** 09394**5 -0 . 2333 -0-2727 -0-1788 -0-2970

16000-
11000
6000
1000
0 800-
I
y 600-
400
200 -
'Followers'
'Riders'
m
Rails et al . : Mother-young relations in ungulates
139
0
Figure 1 . Cluster analysis of data for bovids using < IML, Walther's measure and Contact . Cophenetic corre-lation =07491 .
(Hippotragus niger) and waterbuck, all of whichscored 0% (Table II) . However, five other bovids,bison, tahr, ibex, bongo (Tragelaphus eurycerus)and oryx (Oryx dammah), had intermediate scores(range 32-72") .
Walther's measure was much less satisfactory forthe non-bovid species . Adults of the two hippopo-tamus species (Nile hippopotamus and pygmyhippopotamus, Choeropsis liberiensis) rarely, ifever, lie down and we therefore had to baseWalther's measure on the percentage of intervalsboth mother and young were inactive . Further-more, mother and young were never both lyingduring the first week in four other species, giraffe(Giraffa camelopardalis), onager (Equus hemionus),zebra (Equus burchelli) and muntjac (Muntiacusreeresi) . Walther's measure had to be estimatedfrom our other measures in these cases . Walther'smeasure separates the remaining non-bovids intotwo groups, camel (Camelus bactrianus), tapir(Tapirus terrestris) and reindeer (Rangifer taran-dus) which scored 100%, 100% and 94%, respect-ively and Pere David's deer (Elaphurus davidianus)which scored 0
Relationships Among Species : Cluster Analysis
Clustering procedures impose a hierarchical
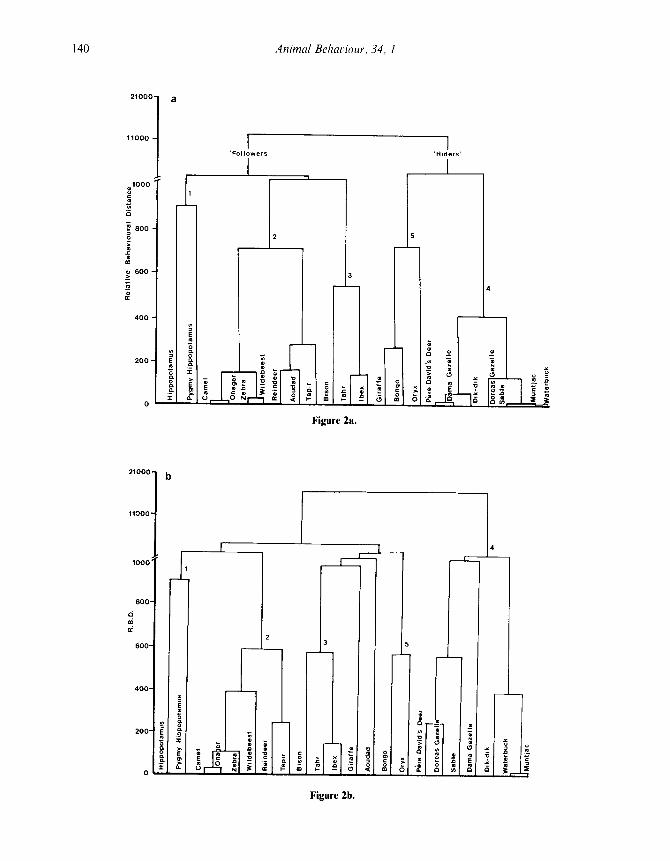
structure on species and species groups so it isnecessary to consider whether the method producesan unacceptable distortion of the data . The reason-ably high cophenetic correlations for our analyses(0. 7416-0 . 7571) indicate that the cluster diagrams(Figs 1, 2) represent an adequate summary ofactual relationships between species . The clusteranalysis based on < 1 ML, Walther's measure andContact separates the bovids into two groups (Fig .1) . The smaller group consists of wildebeest, anAfrican antelope, bison (tribe Bovini), and threeCaprini : tahr, ibex and aoudad . The larger groupconsists entirely of African antelopes : bongo . oryx,dama and Dorcas gazelles, dik-dik, sable andwaterbuck. Cluster analyses using the additionalmeasures. MCA index and both MCA index andYoung Lying, produce the same two major groups(not shown) .
Five consistent species groups can be dis-tinguished in the all species cluster diagrams (Fig .2a, b, c) . The first group includes Nile hippopota-mus and pygmy hippopotamus, which are alwaysassociated with one another and separated fromthe remaining species . The second group containscamel, wildebeest, reindeer, and tapir . which arealways together although other members of thecluster vary . The third group includes the bison,tahr and ibex . This group appears either in associ-ation with the group 2 species (Fig . 2a, c) or as partof a large 'intermediate' cluster (Fig . 2b) . Group 4includes five bovids and two cervids : Dorcas anddama gazelles, dik-dik, waterbuck, sable, muntjacand Pere David's deer. Group 5 contains bongoand onyx; these two species are either looselyassociated with the bovids and cervids of group 4(Fig . 2a) . in the 'intermediate' cluster (Fig . 2b), orloosely associated with the large cluster containinggroups I . 2 and 3 (Fig . 2c) .
DISCUSSION
Methodological Problems
Many of our sample sizes were small, either interms of the number of pairs per species, the totaltime during which data were recorded, or both, butthis did not appear to have a marked effect on theresults . Data for the species with the smallestsample sizes (muntjac, waterbuck and dik-dik)resembled those for other hider species with larger
140
21000-
11000 -
m 100
m
O
400 -
200 -
21000
11000
1000
400-
200-
0
E
0
x
a
x
E
0.
E
E
O a
'Followers'
m mac ° 3O N
a
3
d
ama
Animal Behaviour, 34, 1
Figure 2a.
a
Figure 2b .
M
0
0
0
0 0.
0
O
•
wEa
9
•
0
•
0
u
•
mO

21000-
11000-
1 0
9 800-0a
600-
400-
200-
C
E
0
2
2
E EU
Rails et al. : Mother-young relations in ungulates
141
3 a 00
sample sizes and our analyses placed all three in the`strong' hider group, in agreement with field obser-vations (Table V). Furthermore, data on two pairsof the same species never differed to the extent thatour classification as follower or hider would havechanged had we been able to observe only one ofthe pairs .
Differences in enclosure size and design, and theextent to which observation periods coincided withthe activity periods of different species may haveinfluenced our results in unknown ways . The onlyspecies observed in both a small (0 . 13 ha) and alarge (12 . 14 ha) enclosure was the oryx; we did notdetect any obvious differences on our measuresbetween animals from these two enclosures .
We believe that the main effect of differences inenclosure design was on the percentage of time thatboth mother and young were visible, which tendedto be lower in enclosures where the animals wereable to move out of view . Although abundant coverin an enclosure made it difficult at times for anobserver to locate the young, it is unlikely that theavailability of cover affected the amount of timeyoung of hider species spent `hiding' . Young ofthese species typically hide (i .e . lie motionless at aconsiderable distance from the mother) almost all
0 a 0
0
0
0.
rt
a a
Figure 2c .
Figure 2. Cluster analyses of data for all species using various combinations of measures : (a) < I ML, Walther'smeasure and Contact ; (b) these three measures plus MCA index ; and (c) these four measures plus Young Lying .Cophenetic correlations=07416, 07571 and 0-749 1, respectively .
•
0•
n•
a
the time in the first days after birth (Lent 1974), andwe and others (Walther 1979) have observed thatthey will hide even if little or no cover is available .
Most species tended to be more active in themorning and late afternoon than during mid-dayand our observation periods agreed roughly withthese periods of increased activity. The mostnocturnal species were probably the Nile hippopo-tamus, pygmy hippopotamus and tapir. In thewild, the Nile hippopotamus remains close to heryoung in the water during the day in the earlyweeks, but leaves it behind while she feeds on landat night (H . Klingel, personal communication) .Such a marked day-night difference in the spatialrelationship between mother and young has notbeen reported in any other ungulate species . Thepygmy hippopotamus apparently leaves her youngalone in the water at times, but it is not known ifthis occurs mostly at night (Galat-Luong 1981) .Very little is known about mother-young relation-ships in tapirs, but the young have a disruptivelypatterned pelage, which suggests that they arehiders (Schiirer 1976). Because mother and youngwere usually resting in close proximity during ourdaytime observations, all three of these speciesclustered with the followers (Fig . 2a, b, c) . It may he
142
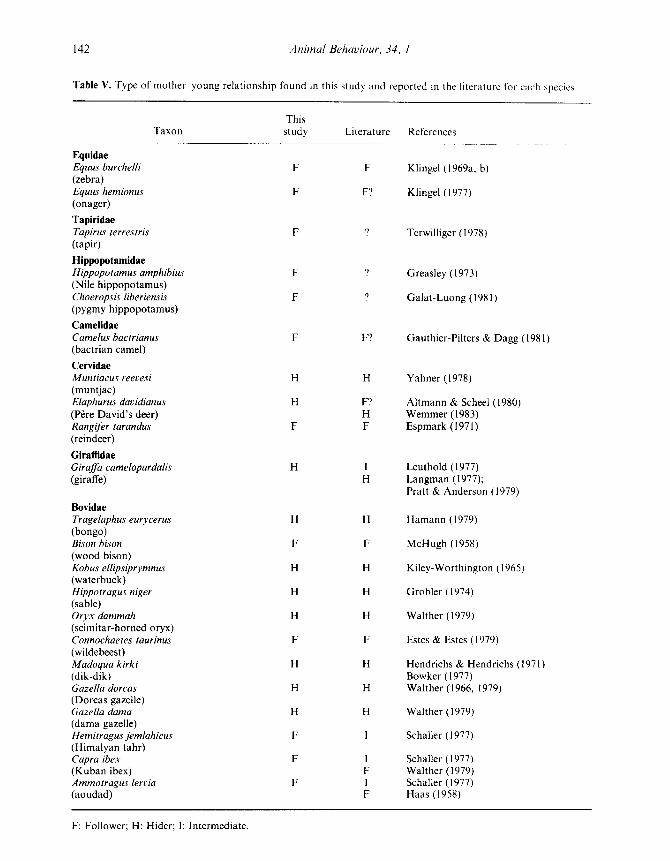
Table V. Type of mother-young relationship found in this study and reported in the literature for each species
Animal Behaviour, 34, l
ThisTaxon study Literature References
EquidaeEquus burchelli F F Klingel (1969a, b)(zebra)Equus hemionus F F? Klingel (1977)(onager)
TapiridaeTapirus terrestris F Terwilliger (1978)(tapir)
HippopotamidaeHippopotamus amphibius F Greasley (1973)(Nile hippopotamus)Choeropsis liberiensis F 9 Galat-Luong (1981)(pygmy hippopotamus)
CamelidaeCamelus bactrianus F F? Gauthier-Pilters & Dagg (1981)(bactrian camel)
CervidaeMuntiacus reevesi H H Yahner (1978)(muntjac)Elaphurus davidianus H F? Altmann & Scheel (1980)(Pere David's deer) H Wemmer (1983)Rangifer tarandus F F Espmark (1971)(reindeer)
GiraffidaeGiraffa camelopardalis H I Leuthold (1977)(giraffe) H Langman (1977) ;
Pratt & Anderson (1979)
BovidaeTragelaphus eurycerus H H Hamann (1979)(bongo)Bison bison F F McHugh (1958)(wood bison)Kobus ellipsiprymnus H H Kiley-Worthington (1965)(waterbuck)Hippotragus niger H H Grobler (1974)(sable)Oryx dammah H H Walther (1979)(scimitar-horned oryx)Connochaetes taurinus F F Estes & Estes (1979)(wildebeest)Madoqua kirki H H Hendrichs & Hendrichs (1971)(dik-dik) Bowker (1977)Gazella dorcas H H Walther (1966, 1979)(Dorcas gazelle)Gazella dama H H Walther (1979)(dama gazelle)Hemitragus jemlahicus F I Schaller (1977)(Himalyan tahr)Capra ibex F I Schaller (1977)(Kuban ibex) F Walther (1979)Ammotragus lervia F 1 Schaller (1977)(aoudad) F Haas (1958)
F: Follower; H : Hider ; I : Intermediate .

that maternal behaviour in these species actuallyresembles that of oryx and bongo ; the mothersometimes rests close to the young but leaves italone when she forages .
The Complexity of Mother-young Relationships inUngulates
Given the diversity in mother-young relation-ships documented in this study, it seems appro-priate to ask whether the traditional division ofungulate species into followers and hiders is ade-quate and whether a modification of this system orsome other classification would prove more useful .Bon ids
When only the bovids are considered, the classi-fication of species as either followers or hidersseems reasonable, particularly if each of these twogroups is subdivided as Walther (1979) suggests .Walther states that the species within a genususually have similar patterns of mother-youngbehaviour and he assigns many bovid genera to oneof four categories : strong follower, weak follower,strong hider or weak hider .
Those species in our study scoring 100°,0 onWalther's measure (wildebeest and aoudad) mightbe regarded as the strong followers and thosescoring 0 (dama and Dorcas gazelles, dik-dik,sable and waterbuck) as strong hiders (Table II) .The cluster diagram (Fig. 1) contains the samestrong follower and hider groups . This classifica-tion agrees with Walther (1979), except that aou-dad (Ammotragus) and waterbuck (Kobus) areplaced in the strong rather than the correspondingweak subcategory .
Five of the bovid species in our study hadintermediate scores (between 0 and 100) onWalther's measure . In the cluster analysis, three ofthese, bison, tahr and ibex, associated with thestrong followers and the other two, bongo andoryx, with the strong hiders . We tentatively labelthese groups the weak followers and weak hiders,respectively . This is in accordance with Walther(1979), except that we place the bongo (Tragela-phus) into the weak rather than the strong hidercategory .
All speciesIf the two large clusters in Fig . 2a are considered
to represent the followers and hiders, our classifica-tion is in near perfect agreement with the literature(Table V). This simple dichotomous classification,however, obscures the considerable variation in
Rails et al . : Motheryoung relations in ungulates 143
mother-young behaviour among the species withineach group (Table II) .
Among the followers, the two hippopotamusspecies displayed a distinct behavioural patternwith very close mother-young association and lowactivity levels . The two equid species, zebra andonager, scored high on the four measures ofassociation, but the young spent considerably lesstime lying down than those of other followers(Table II). As a result, the onager and zebraclustered with the followers when only measures ofassociation were considered (Fig . 2a. b), butformed a separate cluster when Young Lying wasincluded as a variable (Fig . 2c) .
Young giraffe showed a unique pattern : theywere intermediate between followers and hiderswith respect to < I ML, but spent less time lyingthan the young of any other species except zebra(Table II) . Giraffe, therefore, have a different groupassociation in each of the cluster analyses . Depend-ing on which combination of behavioural measureswas considered, they were weakly associated withthe bovid followers (Fig . 2c), with the bovid hiders(Fig. 2a) or in an intermediate cluster (Fig . 2b) .This unusual combination of characteristics mayexplain conflicting comments about giraffe in theliterature (Table V) .The behaviour of the weak bovid hiders, bongo
and oryx, was unusual in that a mother often restedclose to her young although she left it alone forextended periods. Stringham (1974) found a similarbehavioural pattern in the moose (Alces alces), andsuggested three types of mother-young relation-ships in cervids : followers, hiders and 'defenders' .Defenders, such as moose, may leave the youngalone while foraging but rest close to and defend itat other times. Both bongo and oryx possessexceptionally large horns and the tendency forfemales to protect their calves is striking. Duringour study, one female oryx with a small calf brokeher horns attempting to attack an escaped cheetahthrough the fence of her enclosure .
Although we agree with Stringham's view thatthe extent to which the mother can and will protecther young is an important dimension to consider incross-species comparisons of mother-young rela-tionships in ungulates, we doubt that his three-wayclassification can be successfully extended fromcervids to ungulates as a whole . He suggests thatfollowers cope with predators by flight, hiders byconcealment, and defenders through defence .However, followers such as bison may defend their -

144
Animal Behaviour, 34, 1
young vigorously either alone or as part of a socialgroup. Furthermore, many ungulate species whichmight be considered behaviourally intermediatebetween followers and hiders cannot be character-ized as showing a high level of maternal defence .The species in the `intermediate' cluster (Fig . 2b)(bongo, oryx, bison, giraffe, tahr, ibex and aoudad)form a heterogeneous group . Bongo and oryx tendto cluster with the hiders because the mothers oftengraze at considerable distances from their young,while the bison tends to cluster with the followersbecause a close association between mother andyoung is maintained regardless of activity state .The giraffe, as previously noted, shows its ownunique combination of behavioural traits . Thegoats and sheep, tahr, ibex and aoudad, may formanother distinct group . Schaller (1977) suggeststhat most sheep (Ovibovini) and goats (Caprini)are characterized by very short hiding periods andsubsequent following of the mother by the young .We suspect this is true in some species even thoughwe did not detect a distinct shift from hiding tofollowing in those we observed .
In summary, we conclude that the traditionalfollower-hider dichotomy, although useful as arough characterization of mother-young relation-ships in many ungulates, does not adequatelydescribe the entire range of behavioural variationin these relationships . A modification of thisclassification, such as the division of each majorcategory into weak and strong categories, mayprove helpful for some groups . However, a farmore informative representation of similarities anddifferences among species can be attained by usingnumerical scores on standardized measures .
In general, it has been difficult to combine andcompare results of past studies on mother-youngbehaviour in ungulates because data have beencollected in different ways by different observers .We believe that our five behavioural measuresprovide a reasonably good basis for characterizingmother-young relationships in ungulate species .By using standardized quantitative measures suchas these, future investigators should be able toresolve apparent contradictions and to integratenewly studied species into the framework estab-lished by our study .
ACKNOWLEDGMENTS
This project would not have been possible without
the help of the Friends of the National Zoo,Washington, D.C., who provided many of thevolunteer observers as well as financial support . Wethank L . W. Cahill, director of live collections atMTZ, for facilitating observations there and Lee-Ann Hayek and James Craig for help with thestatistical analysis . We are very grateful to all thosewho observed animals or helped in other ways atNZP and MTZ and regret that space does not allowus to recognize them individually .
REFERENCES
Altmann, D . & Scheel, H . 1980 . Geburt, Beginn desSozialverhaltens and erstes Lernen beim Milu, Elaphusdavidianus . Milu ., 5, 146-156 .
Bowker, M. H . 1977 . Behavior of Kirk's dik-dik (Mado-qua kirki hindei) in Kenya . Ph .D. thesis, NorthernArizona University .
Espmark, Y . 1971 . Mother-young relationship and onto-geny of behavior in reindeer (Rangi(er tarandus L .) . Z .Tierpsychol ., 29, 42-81 .
Estes, R . D . 1976 . The significance of breeding synchronyin the wildebeest . E. Afr . Wildl. J., 14, 135--152 .
Estes, R . D . & Estes, R . K . 1979 . The birth and survival ofwildebeest calves . Z. Tierpsvcho/ ., 50, 45 95 .
Fraser, A . F. 1968 . Reproductive Behaviour in Ungulates .London: Academic Press .
Galat-Luong, A. 1981. Quelques observations sur unhippopotame pygmee nouveau-ne (Choeropsis liberien-sis) en foret de Tai, Cote d'Ivoire . Mammalia, 45, 39-41 .
Gauthier-Pilters, H . & Dagg, A . 1 . 1981 . The Camel . ItsEvolution, Ecology, Behavior, and Relationship to Man .Chicago: University of Chicago Press .
Greasley, W . H . 1973 . Protective maternal behavior ofhippopotamus . Puku, 7, 196 .
Grobler, J . H . 1974 . Aspects of the biology, populationecology, and behavior of the sable Hippotragus nigerniger (Harris, 1838) in the Rhodes Matapos NationalPark, Rhodesia . Arnoldia (Rhodesia), 7, 1-36 .
Haas, G . 1958 . Untersuchungen uber angeborene Verhal-tensweisen bei Mahnenspringern (Ammotragus lerviaPallas) . Z. Tierpsvchol., 16, 218-242 .
Hamann, U . 1979 . Beobachtungen zum Verhalten vonBongos (Tragelaphus erveeros Ogilby 1836) im Frank-furter Zoo. Zool. Gart., 49, 319-375 .
Hendrichs, H . & Hendrichs, U . 1971 . Freilanduntersu-chungen zur Okologie and Ethologie der Zwerg-Antilope Madoqua (Rhynchotragus) kirki (Gunter,1880) In : Dik-Dik and E/efanten (Ed. by H . Hendrichs& U. Hendrichs), pp . 9-75 . Munich : Piper .
Kiley-Worthington, 1 . 1965 . The waterbuck (Kobusdefassa Ruppel 1835 and K. ellipsiprymnus Ogilby1833) in East Africa : spatial distribution . A study ofsexual behavior. Mammalia, 29, 177-204 .
Klingel, H . 1969a. Reproduction in the plains zebra,Equus burchelli boehmii : behavior and ecological fac-tors . J. Reprod. Fert ., Suppl ., 6, 339-345 .

Klingel, H . 1969b . The social organization and popula-tion ecology of the plains zebra (Equus quagga) . Zool .Afr., 4, 249-263 .
Klingel, H . 1977 . Observations on social organizationand behaviour of African and Asiatic wild asses (Equusafricanus and E. hemionus) . Z . Tierpsychol., 44, 323-331 .
Langman, V . A . 1977 . Cow-calf relationships in giraffe(Giraf'a camelopardahs giro lit) . Z. Tierpsrchol ., 43,264-286.
Lent, P . C . 1974 . Mother-infant relationships in ungu-lates . In : The Behavior of Ungulates and Its Relation toManagement (Ed. by V . Geist & F . Walther), pp . 14--55 . Morges, Switzerland : International Union for theConservation of Nature and Natural Resources .
Leuthold, W . 1977 . African Ungulates: A ComparativeReview oftheir Ethology and Behavioral Ecology . NewYork: Springer-Verlag .
McHugh. T. 1958 . Social behavior of the Americanbuffalo (Bison bison bison) . Zoologica, 43, 1 -40 .
Murdock, G . K . .. Stine . W. W. & Maple, T . L . 1983 .Observations of maternal-infant interactions in acaptive herd of sable antelope (Hippotragus niger) .Z_ool Biol ., 2, 215 224 .
Pratt, D . M . & Anderson, V . H . 1979 . Giraffe cow calfrelationships and social development of the calf in theSerengeti . Z. Tierpsrchol., 51, 233-251 .
Rails, K . . Lundrigan, B . & Kranz, K . 1983 . Variability ofbehavioral data recorded by volunteer observers . Int .Zoo Yhk, 22, 244 249 .
Schaller, G . B . 1977 . Mountain Monarchs . Chicago :University of Chicago Press .
Schurer, U. 1976 . Beobachtungen an einem neugebore-nen Flachlandtapir, Tapirus terrestris (Linne 1766) .Zool. Gart . 46, 367-370 .
Sokal, R . 1961 . Distance as a measure of taxonomicsimilarity . Svst . Zool ., 10, 70--76 .
Rails et al . : Mother-young relations in ungulates
145
Stringham, S . F . 1974 . Mother-infant relationships inmoose . Nat. Can ., 101, 325-369 .
Talbot . L . M ., Payne, W . J . A ., Leger, H . P ., Verdcourt,L. D. & Talbot, M . H . 1965 . The meat productionpotential of wild animals in Africa . Commonw. Bar.Anim . Breed. Genet ., 16.
Terwilliger, V . J . 1978 . Natural history of Baird's tapir onBarro Colorado Island, Panama, Canal Zone . Biotro-pica, 10, 211 230.
Walther, F. 1961. Einige Verhaltensbeobachtungen amBergwild des Georg von Opel Freigehege . Tier/o -schung, 1960-1961, 53-89 .
Walther, F. 1964 . Verhaltensstudien an der GattungTragelaphus de Blainville (1816) in Gefangenschaftunter besonderer Berucksichtigung des Sozialverhal-tens . Z. Tierpsvchol., 21, 393-467 .
Walther, F . 1965 . Verhaltensstudien an der Grantsgazelle(Ga_ella granti Brooke, 1872) im Ngorongoro-Krater .Z. Tierpsrchol., 22, 167-208 .
Walther, F . 1966 . Mit Horn and Huf Berlin : Paul PareyVerlag .
Walther, F. 1979 . Das Verhalten der Horntrager (Bovi-dae) . Handb. Zool., Band 8, Lieferung 54, 10(30), I184 .
Wemmer, C . 1983 . Sociology and management . In : TheBiology ofan Extinct Species: Pere David's Deer (Ed .by B. Beck & C. Wemmer) pp. 126-132. Park Ridge .New Jersey: Noyes .
Yahner, R . H. 1978 . Some features of mother--youngrelationships in Reeve's muntjac (Muntiacus recresi) .Appl. Anim, Ethol., 4, 379-388 .
(Re(eived 11 September 1984 ; revised 27 December 1984 .MS. number. .44190 )




















