
Morphoproteomic Profile of mTOR, Ras/Raf Kinase/ERK, and NF-κB Pathways in Human Gastric...
15
Morphoproteomic Profile of mTOR, Ras/Raf Kinase/ERK, and NF-κ B Pathways in Human Gastric Adenocarcinoma Wei Feng, 1 Robert E. Brown, 1 Camtu D. Trung, 2 Wei Li, 1 Liwei Wang, 2 aer Khoury, 2 Sadir Alrawi, 2 James Yao, 2 Keping Xia, 2 and Dongfeng Tan 2 1 e University of Texas Medical School–Houston and 2 e University of Texas MD Anderson Cancer Center, Houston, Texas Abstract. Preclinical studies using human gastric adenocarcinoma (GAC) cell lines have shown that the mammalian target of rapamycin (mTOR) inhibitor, rapamycin, can inhibit tumor growth and that the extracellular signal-regulated kinase (ERK) of the Ras/Raf kinase/ERK pathway is related to chemoresistance and apoptosis. We examined the state of activation of components of mTOR, Ras/Raf kinase/ERK, and nuclear factor (NF)-κB signal transduction pathways, as well as cell cycle protein analyte correlates in GAC cases. Formalin-fixed paraffin-embedded tissue microarray blocks containing samples from 210 cases of GAC were examined. Immunohistochemistry was utilized to detect the following antigens: S100P, upstream stimulator of ERK, and NF-κB pathways; phosphorylated (p)-mTOR (Ser 2448), p-ERK-1/2 (r 202/Tyr 204), and one of their common down-stream effectors, p-p70S6K(r 389); p-NF-κBp65(Ser 536); and cell cycle associated proteins, Ki-67, and S phase kinase-associated protein (Skp)2. Immuno- reactivity (0 to 4+) of protein expression and compartmentalization were assessed by bright-field microscopy. e majority of cases showed positive (1+ to 4+) cytoplasmic/plasmalemmal p-mTOR (88%), and moderate- strong (2+ to 4+) nuclear p-p70S6K (93%) and nuclear S100P (81%) expression. A subset of cases exhibited moderate-strong nuclear p-ERK-1/2 (15%) and p-NF-κBp65 (36%) expression. e majority of cases showed concomitant moderate-strong (2+ to 4+) nuclear Ki-67 (71%) and Skp2 (68%). Nuclear expression levels of p-ERK-1/2 and p-NF-κBp65, of p-p70S6K and p-NF-κB, and of Ki-67 and Skp2, respectively, showed significant linear correlations in GAC (p <0.001). Additionally, there were statistically significant differences in the mean expression levels of p-ERK-1/2 and p-NF-κBp65 in diffuse vs intestinal types of GAC, with higher levels of both in the diffuse type ( p = 0.001 and p <0.0001, respectively). In summary, morphoproteomic analysis reveals constitutive activation of mTOR and to some extent, Ras/Raf kinase and NF-κB pathways in GAC, as evidenced by increased cytoplasmic p-mTOR, nuclear translocation of p-p70S6K and p-ERK-1/2 phosphorylated at putative sites of activation (Ser 2448, r 389, and r 202/ Tyr 204, respectively), as well as correlative expression of cell cycle analytes, Ki-67, and Skp2. ese results suggest that a prospective study is warranted to evaluate the use of morphoproteomic profiling of individual patients with GAC in order to design combinatorial treatment strategies that target the mTOR, Ras/Raf kinase/ERK, and/or NF-κB pathways. Keywords: morphoproteomics, gastric adenocarcinoma, mTOR, ERK, NF-κB Address correspondence to Dongfeng Tan, M.D., Department of Pathology, M.D. Anderson Cancer Center, 1515 Holcombe Bvld, Houston, TX 77030, USA; tel 713 745 4977; fax 713 745 1105; e-mail [email protected]; or to Robert E. Brown, M.D., Department of Pathology and Laboratory 0091-7370/08/0300-0195. $4.50. © 2008 by the Association of Clinical Scientists, Inc. Available online at www.annclinlabsci.org Annals of Clinical & Laboratory Science, vol. 38, no. 3, 2008 Medicine, University of Texas Medical School–Houston, 6431 Fannin Street, MSB 2.286, Houston, TX 77030, USA; tel 713 500 5332; fax 713 500 0695; e-mail robert.brown@ uth.tmc.edu. 195
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Morphoproteomic Profile of mTOR, Ras/Raf Kinase/ERK, and NF-κB Pathways in Human Gastric...

Morphoproteomic Profile of mTOR, Ras/Raf Kinase/ERK, and NF-κB Pathways in Human Gastric Adenocarcinoma
Wei Feng,1 Robert E. Brown,1 Camtu D. Trung,2 Wei Li,1 Liwei Wang,2 Thaer Khoury,2 Sadir Alrawi,2 James Yao,2 Keping Xia,2 and Dongfeng Tan2
1The University of Texas Medical School–Houston and 2The University of Texas MD Anderson Cancer Center, Houston, Texas
Abstract. Preclinical studies using human gastric adenocarcinoma (GAC) cell lines have shown that the mammalian target of rapamycin (mTOR) inhibitor, rapamycin, can inhibit tumor growth and that the extracellular signal-regulated kinase (ERK) of the Ras/Raf kinase/ERK pathway is related to chemoresistance and apoptosis. We examined the state of activation of components of mTOR, Ras/Raf kinase/ERK, and nuclear factor (NF)-κB signal transduction pathways, as well as cell cycle protein analyte correlates in GAC cases. Formalin-fixed paraffin-embedded tissue microarray blocks containing samples from 210 cases of GAC were examined. Immunohistochemistry was utilized to detect the following antigens: S100P, upstream stimulator of ERK, and NF-κB pathways; phosphorylated (p)-mTOR (Ser 2448), p-ERK-1/2 (Thr 202/Tyr 204), and one of their common down-stream effectors, p-p70S6K(Thr 389); p-NF-κBp65(Ser 536); and cell cycle associated proteins, Ki-67, and S phase kinase-associated protein (Skp)2. Immuno-reactivity (0 to 4+) of protein expression and compartmentalization were assessed by bright-field microscopy. The majority of cases showed positive (1+ to 4+) cytoplasmic/plasmalemmal p-mTOR (88%), and moderate-strong (2+ to 4+) nuclear p-p70S6K (93%) and nuclear S100P (81%) expression. A subset of cases exhibited moderate-strong nuclear p-ERK-1/2 (15%) and p-NF-κBp65 (36%) expression. The majority of cases showed concomitant moderate-strong (2+ to 4+) nuclear Ki-67 (71%) and Skp2 (68%). Nuclear expression levels of p-ERK-1/2 and p-NF-κBp65, of p-p70S6K and p-NF-κB, and of Ki-67 and Skp2, respectively, showed significant linear correlations in GAC (p <0.001). Additionally, there were statistically significant differences in the mean expression levels of p-ERK-1/2 and p-NF-κBp65 in diffuse vs intestinal types of GAC, with higher levels of both in the diffuse type ( p = 0.001 and p <0.0001, respectively). In summary, morphoproteomic analysis reveals constitutive activation of mTOR and to some extent, Ras/Raf kinase and NF-κB pathways in GAC, as evidenced by increased cytoplasmic p-mTOR, nuclear translocation of p-p70S6K and p-ERK-1/2 phosphorylated at putative sites of activation (Ser 2448, Thr 389, and Thr 202/Tyr 204, respectively), as well as correlative expression of cell cycle analytes, Ki-67, and Skp2. These results suggest that a prospective study is warranted to evaluate the use of morphoproteomic profiling of individual patients with GAC in order to design combinatorial treatment strategies that target the mTOR, Ras/Raf kinase/ERK, and/or NF-κB pathways.
Keywords: morphoproteomics, gastric adenocarcinoma, mTOR, ERK, NF-κB
Address correspondence to Dongfeng Tan, M.D., Department of Pathology, M.D. Anderson Cancer Center, 1515 Holcombe Bvld, Houston, TX 77030, USA; tel 713 745 4977; fax 713 745 1105; e-mail [email protected]; or to Robert E. Brown, M.D., Department of Pathology and Laboratory
0091-7370/08/0300-0195. $4.50. © 2008 by the Association of Clinical Scientists, Inc.
Available online at www.annclinlabsci.org
Annals of Clinical & Laboratory Science, vol. 38, no. 3, 2008
Medicine, University of Texas Medical School–Houston, 6431 Fannin Street, MSB 2.286, Houston, TX 77030, USA; tel 713 500 5332; fax 713 500 0695; e-mail [email protected].
195

Introduction
Gastric cancer is the second leading cause of cancer-related death and the fourth most common cancer world-wide [1]. Gastric adenocarcinoma (GAC) has a dismal outcome since a high percentage of cases present with advanced disease. Surgical resection remains the mainstay of gastric cancer treatment. However, the rate of local tumor recurrence and lymph node metastasis after surgical treatment is high, ranging from 45% to 87.5% depending on the type of resection and lymph node dissection [2,3]. Standard chemotherapy regimens of cisplatin, 5-fluorouracil (5-FU), and epirubicin have low and frequently short-lived response rates (20 to 40%) [4]. The lack of effective treatment has led to intensive studies to examine molecular regulators of cellular growth and angiogenesis, such as inhibitors of receptor tyrosine kinases (RTK’s) and matrix metalloproteinases (MMPI’s) in efforts to improve present therapeutic regimens. There are several main convergent and inter-connected pathways that are involved in the plethora of molecular regulators (such as RTK’s) that are presently under investigation. They are the mammalian target of rapamycin (mTOR) pathway, the Ras/Raf kinase/ERK pathway, and the nuclear factor (NF)-κB pathway [5]. The mTOR pathway is known to regulate protein synthesis, cell cycle progression, metabolism, and angiogenesis [6]. It is regulated via sequential activation of phosphatidyl-inositol 3’-kinase (PI3’-K) and then Akt, which in turn leads to phosphorylative activation of mTOR at serine 2448 [7,8]. The raptor mTOR protein complex phosphorylatively activates its down-stream effector, p70S6K, at threonine 389 [5,7,9-14]. Preclinical study of human gastric cancer cell lines has demonstrated in vitro inhibition of the mTOR pathway by rapamycin through analysis of down-stream effectors such as hypoxia-inducible factor (HIF)-1α, as well as cellular proliferation rate and angiogenesis [15,16], indicating a role for the mTOR pathway in GAC carcinogenesis. The mTOR pathway is an appealing target clinically because it has an identified inhibitor, rapamycin, an analogue of which (temsirolimus) has been used in phase II and III clinical trials for metastatic renal cell carcinoma [17,18]. In addition to gastric adeno-
carcinomas, the mTOR pathway has been shown in preclinical studies to play a role in carcinogenesis of the breast, ovary, and prostate as well as other gastrointestinal malignancies, including pancreatic and colon adenocarcinoma and in ALK-positive anaplastic large cell lymphoma [15,19-25]. Studies in other tumors have also demonstrated phosphoryl-ative activation of p70S6K via the mTOR pathway, resulting in increased G1 cell cycle progression, cell survival, and tumor cell proliferation [21,22,25]. However, the extent of involvement of the mTOR pathway in cancer growth in individual patients is unclear, as reflected in mixed results in a phase II trial with the mTOR inhibitor, temsirolimus [17]. The Ras/Raf kinase/ERK pathway consists of the Ras oncogene and its direct effector in mammalian cells, Raf-1 serine/threonine kinase, which in turn activates a series of kinases including the extra-cellular signal-regulated kinase (ERK) [5,26-29]. ERK is responsible for phosphorylating p70S6K, which mediates cell growth and cell cycle entry [21,30,31]. ERK may also activate the raptor mTOR complex via inactivation of tuberin (TSC2) [32]. Preclinical studies have shown increased cell survival and proliferation with respect to expression level of ERK in gastric carcinoma cell lines [33,34]. Activation of the transcription factor NF-κB signaling pathway is involved in cellular prolifer-ation, apoptosis, and tumor suppression and its constitutive activation is related to progression of various cancers, including renal cell carcinoma, cervical cancer, and esophageal cancer [35]. Its role in gastric carcinoma is unclear, with conflicting reports of the prognostic significance of NF-κB expression [36-39]. NF-κB has been shown to be activated by the same Akt pathway that activates mTOR and by the Ras/Raf kinase/ERK pathway [5,40-44]. S100P is a 95-amino acid protein that was first purified from placenta and that has a restricted cellular distribution. Its expression has been reported be GI malignancies, such as colon and pancreatic adenocarcinoma, and may have adverse prognostic significance in lung cancer [45-47]. S100P functions in an autocrine manner via the receptor for advanced glycation end-product (RAGE) to activate ERK and NF-κB pathways and to promote cell proliferation and survival [48,49].
Annals of Clinical & Laboratory Science, vol. 38, no. 3, 2008196

The primary purpose of this study was to assess the state of activation of the mTOR, Ras/Raf kinase/ERK, and NF-κB pathways in GAC, using a morphoproteomic approach [5], which combines aspects of histopathology, molecular biology, and protein chemistry in an attempt to define the protein circuitry in signal transduction pathways of convergence in GAC.
Materials and Methods
Study groups. After Institutional Review Board (IRB) approval, a total of 215 total and partial gastrectomy specimens with adequate material were retrieved from the files of the Department of Pathology at University of Texas MD Anderson Cancer Center. The gastrectomies had been performed with curative intent over the span of 20 yr. To avoid potential treatment effects, specimens with preoperative treatment were not included. All tissues had been fixed in 10% neutral buffered formalin, routinely processed, and embedded in paraffin. All hematoxylin-eosin (H&E)-stained sections from each case were reviewed and two sections with an adequate amount of gastric adenocarcinoma from each case were selected for inclusion in the tissue microarrays.
Construction of tissue microarrays. Tissue microarrays (TMAs) were constructed using a manual tissue arrayer device from Beecher Instruments (Sun Prairie, WI). Areas with an adequate amount of gastric adenocarcinoma were marked in the paraffin blocks, and three 1-mm diameter cores from each case were included in respective arrays. Thirty-eight 1-mm diameter cores of non-neoplastic gastric tissues from 10 cases with normal gastric mucosa, gastritis, or intestinal metaplasia, but no dysplasia, were also assembled into a tissue microarray to serve as non-neoplastic controls. External controls were provided by an additional slide containing a “sausage” section of different tissues.
Immunohistochemistry using phosphospecific probes. Tissue microarray sections, 4-μm thick, were deparaffinized in xylene and rehydrated in a graded series of ethanols. Heat-induced epitope retrieval was performed. Phospho-specific probes included antibodies directed against mTOR, p70S6K, ERK-1/2, and NF-κBp65, phosphorylated at putative sites of activation, serine (Ser) 2448, threonine (Thr) 389, Thr 202/ tyrosine (Tyr) 204, and Ser 536, respectively (Cell Signaling Technology, Inc., Beverly, MA). The tissue was treated with 3% H2O2 and rinsed with Tween-20 (TBST) buffer. A few drops of diluted normal blocking serum were placed on the tissue and incubated at room temperature. The serum was blotted off and the slides were incubated with primary antibody overnight at 4ºC. The rest of the staining procedure took place on a DAKO Autostainer programmed to treat each slide with diluted biotinylated secondary antibody for 30 min. The slides were rinsed and incubated with DAB (3,3’-diaminobenzidine chromogen solution, EnVision+ System Kit, DAKO) for 10 min. The slides were rinsed again and
counterstained with Gill II hematoxylin, treated with xylene, and cover-slipped. The remaining immunohistochemical staining techniques for Ki-67 (1:500, 30 min incubation at room temperature, M7240, DAKO), Skp2 (1:50, 45 min at room temperature, sc-7164, Santa Cruz Biotechnology, Santa Cruz, CA), and S100P (1:20, 30 min at room temperature MAB2957, R&D Systems, Minneapolis, MN) were performed with a DAKO Autostainer. Positive and negative controls were noted to react appropriately.
Scoring of immunoreactivity of gastric adenocarcinoma, non-neoplastic gastric epithelial cells and cellular compartment-alization of chromogenic signal. Expression levels of the p-mTOR (Ser 2448), p-p70S6K (Thr 389), p-ERK-1/2 (Thr 202/Tyr 204), p-NF-κBp65 (Ser 536), S100P, Ki-67, and Skp2 protein analytes in target cells were determined using bright-field microscopy and applying a scoring system on a scale of 0 to 4+ as follows: 0 for no detectable chromogenic signal; 1+ for focal weak staining; 2+ for partial intermediate or focal strong target cells staining; 3+ for >50% strong or intermediate staining, and 4+ for diffuse strong staining. The cellular compartmentalization of each signal was indicated as predominantly cytoplasmic/plasmalemmal or nuclear.
Statistical analyses. Correlation of semiquantitative staining expression levels between markers with respective Pearson’s r was determined using Excel. Student’s t-test was used to determine significance of correlations of staining expression levels between markers, as well as differences in staining expression levels between different variables such as tumoral subtype. Two-tailed p values were used to assess statistical significance, with the threshold set at p <0.05.
Results
Two-hundred-ten (210) cases had adequate material on the microarrays for at least one immuno-histochemical stain and were included in the analysis. The patients ranged in age from 29 to 89 yr, with a mean of 63. Of the 210 patients, 78 were female and 132 were male. The length of survival ranged from 11 days to 19 yr, with a mean of 27 mo. The survival curves correlated with pathologic staging (pTNM) as well as tumor stage groupings.
Semi-quantitative assessment of p-mTOR and p-p70S6K expression in gastric adenocarcinoma and non-neoplastic gastric epithelium. One hundred-eighty-four (184) and 186 of the total cases of GAC demonstrated good quality, reproducible staining for p-mTOR (Ser 2448) and p-p70S6K (Thr 389), respectively. Ninty-three (93) of 184 GACs (50.5%) exhibited moderate to strong (>2+) cytoplasmic/plasmalemmal expression of p-mTOR and 174 of
Morphoproteomics and gastric cancer 197

186 (93.5%) showed similar moderate to strong intranuclear expression of p-p70S6K. In non-neoplastic gastric epithelium the most intense cytoplasmic/plasmalemmal expression of p-mTOR appeared to be present predominately within the deep foveolar pits, isthmus, and areas of intestinal metaplasia. The overall expression in the tumors appeared to be increased vis-a-vis non-neoplastic
gastric mucosal epithelium. The expression levels of p-p70S6K in nuclei of the non-neoplastic gastric epithelium appeared comparable to that seen in the nuclei of GAC. Moreover, the expression levels were compared in subsets of cases divided according to Lauren’s classification of diffuse and intestinal subtypes. Moderate to strong cytoplasmic/plasma-lemmal expressions of p-mTOR were seen in 58.1%
Fig.1. Representative hematoxylin-eosin (H&E) depictions of chronic gastritis with intestinal metaplasia, diffuse type gastric adenocarcinoma (GAC), and intestinal type GAC, respectively (top row). Phosphospecific probes for mammalian target of rapamycin, mTOR (phosphorylated on serine 2448) and p70S6Kinase (phosphorylated on threonine 389) show intense [DAB] brown chromogen for p-mTOR in the cytoplasm and plasmalemmal aspect and for p-p70S6K in the nuclei and to a lesser extent, cytoplasm of metaplastic and foveolar epithelia and tumor cells (middle and bottom rows, respectively). Note that the interstitial inflammatory cells also express p-p70S6K. (Original magnification x400.)
Annals of Clinical & Laboratory Science, vol. 38, no. 3, 2008198

of the diffuse and 46.7% of the intestinal subtypes, respectively. Moderate to strong expression nuclear expressions of p-p70S6K were seen in 96.9% of the
diffuse and 91.7% of the intestinal subtypes, respectively. These findings are illustrated in Fig. 1 and summarized in Tables 1 and 2.
Fig. 2. S100P expression (top row) in chronic gastritis with intestinal metaplasia reveals strong nuclear and to some extent cytoplasmic signal for this protein analyte in foveolar epithelium and occasional goblet cells, but absence from most of the metaplastic epithelium. Tumoral epithelia in diffuse and intestinal type gastric adenocarcinoma (GAC) also show a similar pattern of compartmental expression, strong nuclear and lesser cytoplasmic expression. Phosphospecific probes for extracellular signal-regulated kinase (ERK)-1/2 (phosphorylated on threonine 202/tyrosine 204) and nuclear factor (NF)-κBp65 (phosphorylated on serine 536), the downstream effectors of either S100P signaling through receptor for advanced glycation end-products (RAGE) and/or signaling through the canonical Ras/Raf kinase/ERK and PI3 -̀K/Akt pathways (see Fig. 4) reveal: similar and strong pERK-1/2 expression in the nuclei of foveolar epithelial cells and in occasional goblet cells and variable nuclear expression in both diffuse and intestinal type tumoral epithelial cells, noting only scattered positivity in the intestinal type; and nuclear and to a lesser extent, cytoplasmic signal for p-NF-κBp65 in the foveolar and metaplastic epithelia of chronic gastritis and variable nuclear expression in diffuse and intestinal type tumoral cells (middle and bottom rows, respectively). Note that the interstitial inflammatory cells also express nuclear p-NF-κBp65. (Original magnification x 400.)
Morphoproteomics and gastric cancer 199

S100P and p-ERK-1/2 in gastric adenocarcinoma and non-neoplastic gastric epithelium. One hundred-eighty-three (183) and 191 of the total cases of GAC demonstrated good quality, reproducible staining for S100P, p-ERK (Thr 202/Tyr 204), and p-NF-κBp65 (Ser 536), respectively. One-hundred-sixty-six (166) of 183 GACs (80.9%) exhibited moderate to strong (>2+) intranuclear expression of S100P, while a smaller subgroup, 32 of 191 (15.2%) showed similar moderate to strong intranuclear expression of p-ERK-1/2. In non-neoplastic gastric epithelium, intranuclear express-ion of S100P and p-ERK-1/2 was present primarily within the foveolar pits and isthmus but not in the gastric glands (see Table 1). Expression levels of these protein analytes were compared in subsets of GAC cases divided according to Lauren’s classification of diffuse or intestinal subtypes. Moderate to strong intranuclear expression of S100P was seen in 85.1% of diffuse and 78.1% of intestinal subtypes. Moderate to strong intranuclear expression of p-ERK-1/2 was noted in 27.8% of diffuse and 10.3% of intestinal subtypes. These
findings are illustrated in Fig. 2 and summarized in Tables 1 and 2.
p-NF-κBp65 in gastric adenocarcinoma and non-neoplastic gastric epithelium. One hundred-ninety-two (192) of the total cases of GAC demonstrated good quality, reproducible staining for p-NF-κBp65 (Ser 536). Sixty-nine (69) of 192 cases (35.9%) showed moderate to strong intranuclear expression of p-NF-κBp65. In non-neoplastic gastric epithelium the intranuclear expression of p-NF-κBp65 appeared comparable to that seen in the nuclei of GAC. When intranuclear expression levels of this protein in GAC were determined according to the Lauren’s classification, 61.5% of cases in the diffuse and 22.2% of cases in the intestinal subtype showed moderate to strong intranuclear expression. These findings are illustrated in Fig. 2 and summarized in Tables 1 and 2.
Ki-67 and Skp2 in gastric adenocarcinoma and non-neoplastic gastric epithelium. One-hundred-seventy-seven (177) and 182 of the total cases of
Table 1. Expression of p-mTOR, p-p70S6K, S100P, p-ERK, p-NF-κBp65, Ki-67, and Skp2 in gastric adenocarcinomas (GAC) and non-neoplastic gastric mucosa (NNGM)
Immunohisto- mTOR pathway Ras/Raf/Kinase NF-κB pathway Proliferation markers chemical stain & RAGE pathway p-mTOR* p70S6K† S100P† p-ERK-1/2† p-NF-κBp65† Ki-67† Skp2†
Total positive GAC cases 162 (88%) 186 (100%) 166 (90.7%) 95 (45.2%) 144 (75%) 153 (86.4%) 166 (91.2%)
Weak positive GAC cases (1+ to 2+) 69 (37.7%) 12 (6.5%) 18 (9.8%) 64 (30.5%) 75 (39.1%) 27 (15.3%) 43 (23.6%)
Strong positive GAC cases (>2+) 93 (50.5%) 174 (93.5%) 148 (80.9%) 32 (15.2%) 69 (35.9%) 126 (71.2%) 123 (67.6%)
Total positive NNGM cases 16+ (94.1%) 16 (100%) 16‡ (100%) 9‡ (64.3%) 16 (94.1%) 13‡ (86.7%) 12‡ (75%)
Weak positive NNGMcases (1+ to 2+) 9+ (52.9%) 3 (18.8%) 3‡ (18.8%) 8‡ (57.1%) 8 (47.1%) 10‡ (66.7%) 10‡ (62.5%)
Strong positive NNGM cases (>2+) 7+ (41.2%) 13 (81.3%) 13‡ (81.3%) 1‡ (7.2%) 8 (47.1%) 3‡ (20%) 2‡ (12.5%)
*cytoplasmic/plasmalemmal staining.†nuclear staining.‡Positivity primarily involving the foveolar pits and isthmus and occasional goblet cells. +Positivity most intense at the foveolar pits and isthmus.
Annals of Clinical & Laboratory Science, vol. 38, no. 3, 2008200
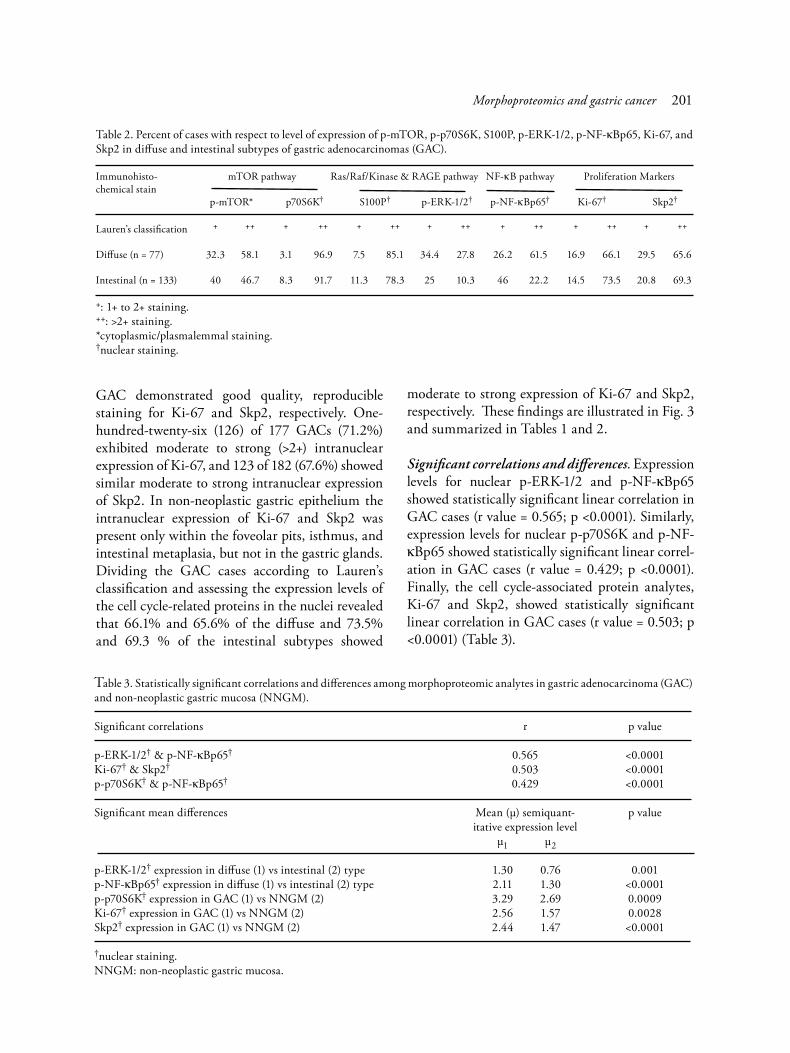
GAC demonstrated good quality, reproducible staining for Ki-67 and Skp2, respectively. One-hundred-twenty-six (126) of 177 GACs (71.2%) exhibited moderate to strong (>2+) intranuclear expression of Ki-67, and 123 of 182 (67.6%) showed similar moderate to strong intranuclear expression of Skp2. In non-neoplastic gastric epithelium the intranuclear expression of Ki-67 and Skp2 was present only within the foveolar pits, isthmus, and intestinal metaplasia, but not in the gastric glands. Dividing the GAC cases according to Lauren’s classification and assessing the expression levels of the cell cycle-related proteins in the nuclei revealed that 66.1% and 65.6% of the diffuse and 73.5% and 69.3 % of the intestinal subtypes showed
moderate to strong expression of Ki-67 and Skp2, respectively. These findings are illustrated in Fig. 3 and summarized in Tables 1 and 2.
Significant correlations and differences. Expression levels for nuclear p-ERK-1/2 and p-NF-κBp65 showed statistically significant linear correlation in GAC cases (r value = 0.565; p <0.0001). Similarly, expression levels for nuclear p-p70S6K and p-NF-κBp65 showed statistically significant linear correl-ation in GAC cases (r value = 0.429; p <0.0001). Finally, the cell cycle-associated protein analytes, Ki-67 and Skp2, showed statistically significant linear correlation in GAC cases (r value = 0.503; p <0.0001) (Table 3).
Table 2. Percent of cases with respect to level of expression of p-mTOR, p-p70S6K, S100P, p-ERK-1/2, p-NF-κBp65, Ki-67, and Skp2 in diffuse and intestinal subtypes of gastric adenocarcinomas (GAC).
Immunohisto- mTOR pathway Ras/Raf/Kinase & RAGE pathway NF-κB pathway Proliferation Markerschemical stain p-mTOR* p70S6K† S100P† p-ERK-1/2† p-NF-κBp65† Ki-67† Skp2†
Lauren’s classification + ++ + ++ + ++ + ++ + ++ + ++ + ++
Diffuse (n = 77) 32.3 58.1 3.1 96.9 7.5 85.1 34.4 27.8 26.2 61.5 16.9 66.1 29.5 65.6
Intestinal (n = 133) 40 46.7 8.3 91.7 11.3 78.3 25 10.3 46 22.2 14.5 73.5 20.8 69.3
+: 1+ to 2+ staining.++: >2+ staining.*cytoplasmic/plasmalemmal staining.†nuclear staining.
Table 3. Statistically significant correlations and differences among morphoproteomic analytes in gastric adenocarcinoma (GAC) and non-neoplastic gastric mucosa (NNGM).
Significant correlations r p value
p-ERK-1/2† & p-NF-κBp65† 0.565 <0.0001Ki-67† & Skp2† 0.503 <0.0001p-p70S6K† & p-NF-κBp65† 0.429 <0.0001
Significant mean differences Mean (μ) semiquant- p value itative expression level μ1 μ2 p-ERK-1/2† expression in diffuse (1) vs intestinal (2) type 1.30 0.76 0.001p-NF-κBp65† expression in diffuse (1) vs intestinal (2) type 2.11 1.30 <0.0001p-p70S6K† expression in GAC (1) vs NNGM (2) 3.29 2.69 0.0009Ki-67† expression in GAC (1) vs NNGM (2) 2.56 1.57 0.0028Skp2† expression in GAC (1) vs NNGM (2) 2.44 1.47 <0.0001
†nuclear staining.NNGM: non-neoplastic gastric mucosa.
Morphoproteomics and gastric cancer 201

Differences in mean expression levels for nuclear p-ERK-1/2 and p-NF-κBp65 were signif-icantly higher in the diffuse subtype vs the intestinal subtype of GAC by t-test with 2-tailed p values of 0.001 and <0.0001, respectively. Mean expression levels for p-p70S6K, Ki-67, and Skp2 were significantly increased in GAC vs the chronic gastritis non-neoplastic gastric mucosa (p values of 0.0009, 0.0028, and <0.0001, respectively). These data are summarized in Table 3.
Discussion
This morphoproteomic study demonstrates the constitutive activation of mTOR and, to a lesser extent, Ras/Raf kinase/ERK and NF-κB pathways in gastric adenocarcinomas. The mTOR pathway is activated in GAC as evidenced by positive expression of p-mTOR and p-p70S6K, phosphorylated at
putative sites of activation, Ser 2448 and Thr 389, respectively and by the nuclear translocation of p-p70S6K [5,7-14,50]. This accords with an in vivo study that reported strong expression by immuno-histochemistry of p-mTOR in a subset of human GAC and by marked reduction in gastric cancer growth and tumor vascularization with rapamycin treatment in a xenograft model [15]. However, the expression patterns of the mTOR activation markers do not always coincide. For example, in the present study there was relatively poor correlation between p-mTOR (Ser 2448) and its down-stream effector, p-p70S6K (Thr 389), the latter of which shows more frequent moderate to strong positive expression (93.5% vs 50.5%, with an r value of only 0.193). A possible explanation for this expression discrepancy between p-mTOR and p-p70S6K is that there is an alternate activation pathway rather than the conventional PI3’-K/Akt
Fig. 3. Cell cycle-associated protein analytes to include Ki-67 (G1, S, G2, and M phase-associated [top row]) and Skp2 (S phase-associated [bottom row]) in chronic gastritis with intestinal metaplasia and in diffuse and intestinal type gastric adenocarcinoma (GAC) reveal variable nuclear (brown chromogen) immunopositivity in foveolar, metaplastic, and tumoral nuclei. (Original magnification x 400.)
Annals of Clinical & Laboratory Science, vol. 38, no. 3, 2008202

pathway. One alternative would be activation mediated by phospholipase D (PLD) via phosphatidic acid (PA), guanosine triphosphatase Cdc42, and the lysophosphatidic acid pathway [19, 20,51-53]. In this context, it is noteworthy that the
mean phospholipase D activity in human gastric carcinoma has been shown to be significantly higher than in adjacent noncancerous mucosa. Moreover, the ratio in cancerous to adjacent noncancerous gastric tissues increased with larger
Fig. 4. Morphoproteomic correlates (*) with variable expression by immunohistochemistry as demonstrated by us and others in gastric adenocarcinoma (GAC) [15,36-39,55,65-70,87-89], when considered in the context of signal transduction pathways of convergence illustrate the potential for: upstream signaling by tyrosine kinases to include VEGFR1-3, PDGFRalpha, EGFR, tie-1/axl, and c-Met through the PI3’-K/Akt pathway (as evidenced by p-Akt expression) and/or the Ras/Raf kinase/ERK pathway (as evidenced by p-ERK 1/2 expression and nuclear translocation) [73-82]; direct activation of ERK by the S100P/RAGE pathway [46,48,49] (again as evidenced by p-ERK-1/2 expression with nuclear translocation in some GAC cases) and of p70S6K by the product of phospholipase D catalysis, namely phosphatidic acid [19,20,51-54] (as evidenced by p-p70S6K nuclear expression that is disproportionate to p-mTOR expression, one of its putative upstream activators [see Results]); convergent signaling of the PI3’-K/Akt and Ras/Raf kinase/ERK pathways to activate the NF-κB pathway through the activation of IKK by both p-Akt and p-ERK 1 /2 [41-44,56,90,91] resulting in p-NF-κBp65 expression with nuclear translocation in some GAC cases; convergent signaling by p-p70S6K and p-ERK-1/2 resulting in G1 to S cell cycle progression as evidenced by Skp2 expression [60] and with a potential influence by p-NF-κB to facilitate the G2/M phase progression [21,61-63,92] leading to tumoral cell proliferation in GAC. The opportunity for complex cross-talk between the PI3’-K/Akt and Ras/Raf kinase/ERK pathways of convergence is inherent in the role of Rheb in activating the mTOR pathway [93-95], while Rheb also interferes in the activation of ERK by inhibiting B-Raf kinase [96-98]. Similarly, there is potential interplay between the NF-κB and ERK pathways and the activation of mTOR via the respective roles of IKK in inhibiting TSC1 [41-44,56,57] and p-ERK-1/2 in inhibiting TSC2 [99], the latter of which forms a complex (TSC1 + TSC2) to inhibit Rheb [94]. Such an interplay along with the variable expression of upstream tyrosine kinases including the inhibitory tyrosine kinase, EphA7 [71,72] allows for variation among individual patients and speaks to the need for morphoproteomic analysis of individual tumors.
Abbreviations: VEGFR: vascular endothelial growth factor receptor; PDGFR: platelet-derived growth factor receptor; EGFR: epidermal growth factor receptor; PI3’-K: phosphatidylinositol 3’-kinase; TSC1; tuberous sclerosis complex 1 (hamartin); TSC2: tuberous sclerosis complex 2 (tuberin); Rheb: Ras homolog enriched in brain; IKK; inhibitor kappa kinase; ERK; extra-cellular signal-regulated kinase; NF-κB: nuclear factor-κB; mTOR: mammalian target of rapamycin; RAGE: receptor for advanced glycation end-products; Skp2: S phase kinase-associated protein2.
Morphoproteomics and gastric cancer 203

tumors, suggesting that such activity plays a role in the promotion of gastric carcinoma [54]. Such an alternative pathway may also explain, at least in part, discrepant and unexpected mixed responses to rapamycin therapy in cancer trials [17,20]. In addition, the strong baseline p-p70S6K expression in gastric epithelium, as shown in the present study, where p70S6K showed diffuse expression in non-neoplastic gastric foveolar and glandular epithelium unlike the more often localized foveolar expression of p-mTOR (see Table 1), also suggests more than one means of activation of p70S6K. S100P, a ligand for RAGE, is expressed in colon and pancreatic adenocarcinomas and has been shown to activate ERK and NF-κB [46-49]. The present study reports its expression in GACs as well as its localized expression within non-neoplastic proliferative foveolar-pit gastric epithelium. This latter finding is consistent with the moderate to strong expression of S100P in the majority of the diffuse subtype of GAC, which is thought to be comprised in part of foveolar cell types. Expression of the receptor for advanced glycation end-products (RAGE), the signal transducer of S100P stimulation, has been reported in gastric cancer. Moreover, it is closely associated with the invasive and metastatic activity of this tumor [55]. Applying a morpho-proteomic perspective, it is notable that the nuclear expression of p-ERK-1/2, when occuring in the present series, is associated in most cases with concurrent expression of S100P, a known upstream activator of the receptor for advanced glycation end-products and in turn ERK-1/2 phosphorylation and NF-κB activity [46,48,49]. Downstream positive signaling by p-ERK-1/2 on the NF-κB pathway is supported by the expression p-NF-κBp65 and by the fact that both of these activated molecules are expressed at statistically greater mean levels in Lauren’s diffuse subtype of GAC. The logic of this perspective is strengthened by the strong statistical correlation between p-ERK-1/2 and p-NF-κBp65 in our series, consistent with known activation of the latter by the former [41-44]. Parenthetically, the highly significant correlation between p-p70S6K and p-NF-κBp65 in our series is consistent with a report in the literature of cross-talk between the NF-κB and mTOR pathways. Thus, inhibitor kappa kinase
(IKK) beta, a component of the NF-κB pathway [56], physically interacts with and phosphorylatively inactivates and suppresses TSC1, which leads to activation of the mTOR pathway [57]. Finally, a preclinical study showed that the activity of NF-κB and the expression of certain of its downstream effectors was reduced in a human gastric adenocarcinoma cell line, along with tumor growth and capacity for invasion, following exposure to caffeic acid phenethyl ester [58]. This finding has pathogenetic significance regarding the role of the NF-κB pathway in GAC. Caffeic acid phenethyl ester is a suppressor of the activation of κB kinase [59] and thereby inhibits the NF-κB pathway. An end result of the activation of these three major pathways of convergence is the promotion of cell growth and proliferation, and we have shown this correlation in our series of GAC cases in the form of the moderate to strong expression of the cell cycle-associated protein analytes, Ki-67 and Skp2, in the majority of cases. These pathways and convergent signaling are depicted in Fig. 4. Such expression far exceeds that seen in the non-neoplastic gastric mucosa where the nuclear immunopositivity was limited to the foveolar pits and isthmus. Our observations are strengthened by the strong correlation between Ki-67 and Skp2 expression. The potential clinical importance of the Skp2 expression lies in the association of this protein analyte with the S phase of the cell cycle [60] indicating that the majority of GAC cases are progressing from the G1 to the S phase and therefore, such cases may be candidates for an S phase active chemotherapeutic agent. Moreover, it coincides with the fact that both the ERK and mTOR pathways promote such progression [21, 61-63]. It is of potential clinical import that the mTOR inhibitor, rapamycin, significantly decreases Skp2 mRNA and protein levels in a dose- and time-dependent fashion (depending on the sensitivity of the cancer cell line to rapamycin) and that the decrease in Skp2 levels is followed by cell growth arrest at G1 [64]. The morphoproteomic analysis of pathways of convergence incorporates immunohistochemical (IHC) data from the literature on the correlative expression of protein analytes involved in signal transduction that potentially could result in,
Annals of Clinical & Laboratory Science, vol. 38, no. 3, 2008204

contribute to, or moderate their constitutive activation. In the context of this study on GAC, a computer-assisted search of the National Library of Medicine’s PubMed database revealed several studies relevant to upstream signaling. Specifically, there are multiple receptor tyrosine kinases (RTK’s) in GAC, variably expressed and detected by IHC. These include the following: EphA7 receptor tyrosine kinase [65], cytoplasmic vascular endo-thelial growth factor receptors (VEGFR)1-3 and platelet-derived growth factor receptor (PDGFR) alpha, and plasmalemmal (membranous) epidermal growth factor receptor (EGFR)1 [66,67], receptor protein tyrosine kinases, tie-1 and axl [68,69], and c-Met [70]. To expand on these, EphA7 could act as a moderator by inhibiting the Ras/mitogen-activated protein kinase (MAPK) cascade and attenuating activation of MAPK by receptors for platelet-derived growth factor, epidermal growth factor, and vascular endothelial growth factor [70] and also inhibiting basal ERK-1/2 activity [71]. There is evidence from other studies that VEGFR, PDGFRalpha, EGFR, axl, and c-Met activate the ERK pathway [72-81]. Additionally, activation of the Akt pathway has been reported as a result of downstream signaling by VEGFR [72-74], PDGFR-alpha [76,77], EGFR [79,80], and c-Met [81]. In short, there is correlative, morphoproteomic expression of tyrosine kinases in GAC that occurs upstream of the Ras/Raf kinase/ERK and Akt/mTOR pathways of convergence demonstrated in this study. These are illustrated in Fig. 4. From a therapeutic standpoint, the plethora of upstream tyrosine kinases in GAC suggests that designing therapies to target the pathways of convergence may be a logical approach in individual patients depending on their morphoproteomic profile. Finally, our study demonstrates constitutive activation of ERK-1/2 and NF-κBp65 in compan-ionate, non-neoplastic tissues when present in the series and specifically in the foveolar epithelium, and when present, metaplastic intestinal epithelium from mucosa involved by chronic gastritis. This coincides with other studies reported in the literature [83-85]. Moreover, it provides a stimulus for future studies to assess the impact of the downregulation of such pathways in patients, diagnosed with this precursor lesion, as a possible
preventative measure against malignant trans-formation. In summary, morphoproteomic analysis in this study confirmed the constitutive activation of components of the mTOR pathway, p-mTOR (Ser 2448), and p-p70S6K (Thr 389) with nuclear translocation; activation of the downstream component of the Ras/Raf kinase/ERK pathway, p-ERK-1/2 (Thr 202/Tyr 204); and activation of the effector component of the NF-κB pathway, p-NF-κBp65 (Ser 536), within human gastric adeno-carcinoma. These results along with the unique finding of correlative expression of an upstream component to ERK and NF-κB activation in the form of S100P validates the expression of several potential targets of therapy for gastric adeno-carcinomas. It is important to note that although there is some commonality of expression of the mTOR pathway component, p-p70S6K, there is hetero-geneity of expression of the Ras/Raf kinase and NF-κB pathway components, particularly along the lines of diffuse and intestinal subtypes. This exemplifies that morphoproteomic profiling of an individual patient’s tumor may be necessary to uncover potential targets in signal transduction pathways that may be contributing to the tumor’s proliferative capacity, invasive potential, and chemoradioresistance. Such profiling may allow for development of combinatorial therapeutic regimens that incorporate signal transduction pathway inhibitors and cell cycle-specific cytotoxic therapies appropriate to an individual patient’s gastric adenocarcinoma (ie, personalized medicine). Acknowledgments
The authors thank Pamela K. Johnston, HT (ASCP), for technical assistance; Mannie Steglich (ASCP) for construction of TMAs; and Bheravi Patel for secretarial support and graphic skills. This study was partially supported by the faculty development fund of the University of Texas MD Anderson Cancer Center.
References
1. The World Cancer Report–the major findings. Cent Eur J Public Health 2003;11:177-179.
Morphoproteomics and gastric cancer 205

2. Hartgrink HH, van de Velde CJ, Putter H, Bonenkamp JJ, Klein Kranenbarg E, Songun I, Welvaart K, van Krieken JH, Meijer S, Plukker JT, van Elk PJ, Obertop H, Gouma DJ, van Lanschot JJ, Taat CW, de Graaf PW, von Meyenfeldt MF, Tilanus H, Sasako M. Extended lymph node dissection for gastric cancer: who may benefit? Final results of the randomized Dutch gastric cancer group trial. J Clin Oncol 2004;22:2069-2077.
3. Gunderson LL, Sosin H. Adenocarcinoma of the stomach: areas of failure in a re-operation series (second or symptomatic look) clinicopathologic correlation and implications for adjuvant therapy. Int J Radiat Oncol Biol Phys 1982;8:1-11.
4. Rivera F, Vega-Villegas ME, Lopez-Brea MF. Chemo-therapy of advanced gastric cancer. Cancer Treat Rev 2007;33:315-324.
5. Brown RE. Morphoproteomics: exposing protein circuit-ries in tumors to identify potential therapeutic targets in cancer patients. Expert Rev Proteomics 2005; 2:337-348.
6. Shaw RJ, Cantley LC. Ras, PI(3)K and mTOR signalling controls tumour cell growth. Nature 2006;441:424-430.
7. Cao X, Kambe F, Moeller LC, Refetoff S, Seo H. Thyroid hormone induces rapid activation of Akt/protein kinase B-mammalian target of rapamycin-p70S6K cascade through phosphatidylinositol 3-kinase in human fibroblasts. Mol Endocrinol 2005;19:102-112.
8. Navé BT, Ouwens M, Withers DJ, Alessi DR, Shepherd PR. Mammalian target of rapamycin is a direct target for protein kinase B: identification of a convergence point for opposing effects of insulin and amino-acid deficiency on protein translation. Biochem J 1999;344: 427-431.
9. Pullen N, Dennis PB, Andjelkovic M, Dufner A, Kozma SC, Hemmings BA, Thomas G. Phosphorylation and activation of p70s6k by PDK1. Science 1998; 279:707-710.
10. Dennis PB, Pullen N, Kozma SC, Thomas G. The principal rapamycin-sensitive p70 (s6k) phosphorylation sites, T-229 and T-389, are differentially regulated by rapamycin-insensitive kinase kinases. Mol Cell Biol 1996;16:6242-6251.
11. Valentinis B, Navarro M, Zanocco-Marani T, Edmonds P, McCormick J, Morrione A, Sacchi A, Romano G, Reiss K, Baserga R. Insulin receptor substrate-1, p70S6K, and cell size in transformation and differentiation of hemopoietic cells. J Biol Chem 2000;275:25451-25459.
12. Xu G, Zhang W, Bertram P, Zheng XF, McLeod H. Pharmacogenomic profiling of the PI3K/PTEN-AKT-mTOR pathway in common human tumors. Int J Oncol 2004;24:893-900.
13. Ali SM, Sabatini DM. Structure of S6 kinase 1 determines whether raptor-mTOR or rictor-mTOR phosphorylates its hydrophobic motif site. J Biol Chem 2005;280:19445-19448.
14. Brown RE, Zhang PL, Lun M, Zhu S, Pellitteri PK, Riefkohl W, Law A, Wood GC, Kennedy TL. Morpho-proteomic and pharmacoproteomic rationale for mTOR
effectors as therapeutic targets in head and neck squamous cell carcinoma. Ann Clin Lab Sci 2006;36:273-282.
15. Lang SA, Gaumann A, Koehl GE, Seidel U, Bataille F, Klein D, Ellis LM, Bolder U, Hofstaedter F, Schlitt HJ, Geissler EK, Stoeltzing O. Mammalian target of rapamycin is activated in human gastric cancer and serves as a target for therapy in an experimental model. Int J Cancer 2007;120:1803-1810.
16. Urano N, Fujiwara Y, Doki Y, Tsujie M, Yamamoto H, Miyata H, Takiguchi S, Yasuda T, Yano M, Monden M. Overexpression of hypoxia-inducible factor-1 alpha in gastric adenocarcinoma. Gastric Cancer 2006;9:44-49.
17. Atkins MB, Hidalgo M, Stadler WM, Logan TF, Dutcher JP, Hudes GR, Park Y, Liou SH, Marshall B, Boni JP, Dukart G, Sherman ML. Randomized phase II study of multiple dose levels of CCI-779, a novel mammalian target of rapamycin kinase inhibitor, in patients with advanced refractory renal cell carcinoma. J Clin Oncol 2004;22:909-918.
18. Hudes G, et al. A phase III, randomized, 3 arm study of temsirolimus (TEMSR) or interferon-alpha (IFN) or the combination of TEMSR + IFN in the treatment of first-line, poor risk patients with advanced renal cell carcinoma. Proc Am Soc Clin Oncol 2006;1(24):LBA4.
19. Chen Y, Rodrik V, Foster DA. Alternative phospholipase D/mTOR survival signal in human breast cancer cells. Oncogene 2005;24:672-679.
20. Chen Y, Zheng Y, Foster DA. Phospholipase D confers rapamycin resistance in human breast cancer cells. Oncogene 2003;22:3937-3942.
21. Gao N, Flynn DC, Zhang Z, Zhong XS, Walker V, Liu KJ, Shi X, Jiang BH. G1 cell cycle progression and the expression of G1 cyclins are regulated by PI3K/AKT/mTOR/p70S6K1 signaling in human ovarian cancer cells. Am J Physiol Cell Physiol 2004;287:C281-291.
22. Gao N, Zhang Z, Jiang BH, Shi X. Role of PI3K/AKT/mTOR signaling in the cell cycle progression of human prostate cancer. Biochim Biophys Res Commun 2003; 310:1124-1132.
23. Guba M, von Breitenbuch P, Steinbauer M, Koehl G, Flegel S, Hornung M, Bruns CJ, Zuelke C, Farkas S, Anthuber M, Jauch KW, Geissler EK. Rapamycin inhibits primary and metastatic tumor growth by antiangiogenesis: involvement of vascular endothelial growth factor. Nat Med 2002;8:128-135.
24. Stephan S, Datta K, Wang E, Li J, Brekken RA, Parangi S, Thorpe PE, Mukhopadhyay D. Effect of rapamycin alone and in combination with antiangiogenesis therapy in an orthotopic model of human pancreatic cancer. Clin Cancer Res 2004;10:6993-7000.
25. Vega F, Medeiros LJ, Leventaki V, Atwell C, Cho-Vega JH, Tian L, Claret FX, Rassidakis GZ. Activation of mammalian target of rapamycin signaling pathway contributes to tumor cell survival in anaplastic lymphoma kinase-positive anaplastic large cell lymphoma. Cancer Res 2006;66:6589-6597.
26. Chow S, Patel H, Hedley DW. Measurement of MAP kinase activation by flow cytometry using phospho-
Annals of Clinical & Laboratory Science, vol. 38, no. 3, 2008206

specific antibodies to MEK and ERK: potential for pharmacodynamic monitoring of signal transduction inhibitors. Cytometry. 2001; 46(2):72-78.
27. Wilhelm SM, Carter C, Tang L, Wilkie D, McNabola A, Rong H, Chen C, Zhang X, Vincent P, McHugh M, Cao Y, Shujath J, Gawlak S, Eveleigh D, Rowley B, Liu L, Adnane L, Lynch M, Auclair D, Taylor I, Gedrich R, Voznesensky A, Riedl B, Post LE, Bollag G, Trail PA. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res 2004;64:7099-7109.
28. Adnane L, Trail PA, Taylor I, Wilhelm SM. Sorafenib (BAY 43-9006, Nexavar), a dual-action inhibitor that targets RAF/MEK/ERK pathway in tumor cells and tyrosine kinases VEGFR/PDGFR in tumor vasculature. Methods Enzymol 2005;407:597-612.
29. Murphy DA, Makonnen S, Lassoued W, Feldman MD, Carter C, Lee WM. Inhibition of tumor endothelial ERK activation, angiogenesis, and tumor growth by sorafenib (BAY43-9006). Am J Pathol 2006;169:1875-1885.
30. Iijima Y, Laser M, Shiraishi H, Willey CD, Sundarava-divel B, Xu L, McDermott PJ, Kuppuswamy D. c-Raf/MEK/ERK pathway controls protein kinase C-mediated p70S6K activation in adult cardiac muscle cells. J Biol Chem 2002;277:23065-23075.
31. Sawhney RS, Cookson MM, Sharma B, Hauser J, Brattain MG. Autocrine transforming growth factor alpha regulates cell adhesion by multiple signaling via specific phosphorylation sites of p70S6 kinase in colon cancer cells. J Biol Chem 2004;279:47379-47390.
32. Ma L, Chen Z, Erdjument-Bromage H, Tempst P, Pandolfi PP. Phosphorylation and functional inactivation of TSC2 by Erk implications for tuberous sclerosis and cancer pathogenesis. Cell 2005;121:179-193.
33. Liu N, Sun LG, Yu HY, Zhang Y, Yang Q, Wang XP. The function of ERK on Fas-mediated apoptotic signaling pathway of human gastric carcinoma cells. Zhonghua Zhong Liu Za Zhi 2005;27:201-203.
34. Verma A, Atten MJ, Attar BM, Holian O. Seleno-methionine stimulates MAPK (ERK) phosphorylation, protein oxidation, and DNA synthesis in gastric cancer cells. Nutr Cancer 2004;49:184-190.
35. Gilmore TD, Koedood M, Piffat KA, White DW. Rel/NF-kappaB/IkappaB proteins and cancer. Oncogene 1996;13:1367-1378.
36. Lee BL, Lee HS, Jung J, Cho SJ, Chung HY, Kim WH, Jin YW, Kim CS, Nam SY. Nuclear factor-kappaB activation correlates with better prognosis and Akt activation in human gastric cancer. Clin Cancer Res 2005;11:2518-2525.
37. Sasaki N, Morisaki T, Hashizume K, Yao T, Tsuneyoshi M, Noshiro H, Nakamura K, Yamanaka T, Uchiyama A, Tanaka M, Katano M. Nuclear factor-kappaB p65 (RelA) transcription factor is constitutively activated in human gastric carcinoma tissue. Clin Cancer Res 2001; 7:4136-4142.
38. Chang MS, Lee HS, Jung EJ, Kim CW, Lee BL, Kim WH. Cell-cycle regulators, bcl-2 and NF-kappaB in Epstein-Barr virus-positive gastric carcinomas. Int J Oncol 2005;27:1265-1272.
39. Yang GF, Deng CS, Xiong YY, Luo J, Wang BC, Tian SF, Xu K. Expression of NFkappaB p65 and its target genes in gastric cancer and precancerous lesions. Zhonghua Zhong Liu Za Zhi 2004;26:551-553.
40. Agarwal A, Das K, Lerner N, Sathe S, Cicek M, Casey G, Sizemore N. The AKT/I kappa B kinase pathway promotes angiogenic/metastatic gene expression in colorectal cancer by activating nuclear factor-kappa B and beta-catenin. Oncogene 2005;24:1021-1031.
41. Hennig B, Lei W, Arzuaga X, Ghosh DD, Saraswathi V, Toborek M. Linoleic acid induces proinflammatory events in vascular endothelial cells via activation of PI3K/Akt and ERK1/2 signaling. J Nutr Biochem 2006; 17:766-772.
42. Lappas M, Permezel M, Rice GE. Advanced glycation endproducts mediate pro-inflammatory actions in human gestational tissues via nuclear factor-kappaB and extracellular signal-regulated kinase 1/2. J Endocrinol 2007;193:269-277.
43. Kim KW, Kim SH, Lee EY, Kim ND, Kang HS, Kim HD, Chung BS, Kang CD. Extracellular signal-regulated kinase/90-KDA ribosomal S6 kinase/nuclear factor-kappa B pathway mediates phorbol 12-myristate 13-acetate-induced megakaryocytic differentiation of K562 cells. J Biol Chem 2001;276:13186-13191.
44. Rangaswami H, Bulbule A, Kundu GC. Nuclear factor-inducing kinase plays a crucial role in osteopontin-induced MAPK/IkappaBalpha kinase-dependent nuclear factor kappaB-mediated promatrix metallo-proteinase-9 activation. J Biol Chem 2004;279:38921-38935.
45. Bartling B, Rehbein G, Schmitt WD, Hofmann HS, Silber RE, Simm A. S100A2-S100P expression profile and diagnosis of non-small cell lung carcinoma: impairment by advanced tumour stages and neoadjuvant chemotherapy. Eur J Cancer 2007;43:1935-1943.
46. Fuentes MK, Nigavekar SS, Arumugam T, Logsdon CD, Schmidt AM, Park JC, Huang EH. RAGE activation by S100P in colon cancer stimulates growth, migration, and cell signaling pathways. Dis Colon Rectum 2007;50:1230-1240.
47. Ohuchida K, Mizumoto K, Egami T, Yamaguchi H, Fujii K, Konomi H, Nagai E, Yamaguchi K, Tsuneyoshi M, Tanaka M. S100P is an early developmental marker of pancreatic carcinogenesis. Clin Cancer Res 2006; 12:5411-5416.
48. Arumugam T, Simeone DM, Schmidt AM, Logsdon CD. S100P stimulates cell proliferation and survival via receptor for activated glycation end products (RAGE). J Biol Chem 2004;279:5059-5065.
49. Ishihara K, Tsutsumi K, Kawane S, Nakajima M, Kasaoka T. The receptor for advanced glycation end-products (RAGE) directly binds to ERK by a D-domain-like docking site. FEBS Lett 2003;550:107-113.
Morphoproteomics and gastric cancer 207

50. Kim SJ, Kahn CR. Insulin stimulates p70 S6 kinase in the nucleus of cells. Biochem Biophys Res Commun 1997;234:681-685.
51. Fang Y, Park IH, Wu AL, et al. PLD1 regulates mTOR signaling and mediates Cdc42 activation of S6K1. Curr Biol 2003;13:2037-2044.
52. Chen J, Fang Y. A novel pathway regulating the mammalian target of rapamycin (mTOR) signaling. Biochem Pharmacol 2002;64:1071-1077.
53. Lehman N, Ledford B, Di Fulvio M, Frondorf K, McPhail LC, Gomez-Cambronero J. Phospholipase D2-derived phosphatidic acid binds to and activates ribosomal p70 S6 kinase independently of mTOR FASEB J 2007;21:1075-1087.
54. Uchida N, Okamura S, Kuwano H. Phospholipase D activity in human gastric carcinoma. Anticancer Res 1999;19:671-675.
55. Kuniyasu H, Oue N, Wakikawa A, Shigeishi H, Matsutani N, Kuraoka K, Ito R, Yokozaki H, Yasui W. Expression of receptors for advanced glycation end-products (RAGE) is closely associated with the invasive and metastatic activity of gastric cancer. J Pathol 2002; 196:163-170.
56. Douillette A, Bibeau-Poirier A, Gravel SP, Clément JF, Chénard V, Moreau P, Servant MJ. The proinflammatory actions of angiotensin II are dependent on p65 phosphorylation by the IkappaB kinase complex. J Biol Chem 2006;281:13275-13284.
57. Lee DF, Kuo HP, Chen CT, Hsu JM, Chou CK, Wei Y, Sun HL, Li LY, Ping B, Huang WC, He X, Hung JY, Lai CC, Ding Q, Su JL, Yang JY, Sahin AA, Hortobagyi GN, Tsai FJ, Tsai CH, Hung MC. IKK beta suppression of TSC1 links inflammation and tumor angiogenesis via the mTOR pathway. Cell 2007;130:440-455.
58. Wu CS, Chen MF, Lee IL, Tung SY. Predictive role of nuclear factor-kappaB activity in gastric cancer: a promising adjuvant approach with caffeic acid phenethyl ester. J Clin Gastroenterol 2007;41:894-900.
59. Choi K, Choi C. Differential regulation of c-jun N-terminal kinase and NF-kappaB pathway by caffeic acid phenethyl ester in astroglial and monocytic cells. J Neurochem 2008;105:557-564.
60. Chiarle R, Fan Y, Piva R, Boggino H, Skolnik J, Novero D, Palestro G, De Wolf-Peeters C, Chilosi M, Pagano M, Inghirami G. S-phase kinase-associated protein 2 expression in non-Hodgkin’s lymphoma inversely correlates with p27 expression and defines cells in S phase. Am J Pathol 2002;160:1457-1466.
61. Fingar DC, Richardson CJ, Tee AR, Cheatham L, Tsou C, Blenis J. mTOR controls cell cycle progression through its cell growth effectors S6K1 and 4E-BP1/eukaryotic translation initiation factor 4E. Mol Cell Biol 2004;24:200-216.
62. Godeny MD, Sayeski PP. ERK1/2 regulates ANG II-dependent cell proliferation via cytoplasmic activation of RSK2 and nuclear activation of elk1. Am J Physiol Cell Physiol 2006;291:C1308-1317.
63. Meloche S, Pouysségur J. The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transition. Oncogene 2007;26:3227-3239.
64. Shapira M, Kakiashvili E, Rosenberg T, Hershko DD. The mTOR inhibitor rapamycin down-regulates the expression of the ubiquitin ligase subunit Skp2 in breast cancer cells. Breast Cancer Res 2006;8:R46.
65. Wang J, Li G, Ma H, Bao Y, Wang X, Zhou H, Sheng Z, Sugimura H, Jin J, Zhou X. Differential expression of EphA7 receptor tyrosine kinase in gastric carcinoma. Hum Pathol 2007;38:1649-1656.
66. Drescher D, Moehler M, Gockel I, Frerichs K, Müller A, Dünschede F, Borschitz T, Biesterfeld S, Holtmann M, Wehler T, Teufel A, Herzer K, Fischer T, Berger MR, Junginger T, Galle PR, Schimanski CC. Coexpression of receptor-tyrosine-kinases in gastric adenocarcinoma–a rationale for a molecular targeting strategy? World J Gastroenterol 2007;13:3605-3609.
67. Takehana T, Kunitomo K, Suzuki S, Kono K, Fujii H, Matsumoto Y, Ooi A. Expression of epidermal growth factor receptor in gastric carcinomas. Clin Gastroenterol Hepatol 2003;1:438-445.
68. Lin WC, Li AF, Chi CW, Chung WW, Huang CL, Lui WY, Kung HJ, Wu CW. tie-1 protein tyrosine kinase: a novel independent prognostic marker for gastric cancer. Clin Cancer Res 1999;5:1745-1751.
69. Wu CW, Li AF, Chi CW, Lai CH, Huang CL, Lo SS, Lui WY, Lin WC. Clinical significance of AXL kinase family in gastric cancer. Anticancer Res 2002;22:1071-1078.
70. Kaji M, Yonemura Y, Harada S, Liu X, Terada I, Yamamoto H. Participation of c-met in the progression of human gastric cancers: anti-c-met oligonucleotides inhibit proliferation or invasiveness of gastric cancer cells. Cancer Gene Ther 1996;3:393-404.
71. Miao H, Wei BR, Peehl DM, Li Q, Alexandrou T, Schelling JR, Rhim JS, Sedor JR, Burnett E, Wang B. Activation of EphA receptor tyrosine kinase inhibits the Ras/MAPK pathway. Nat Cell Biol 2001;3:527-530.
72. Miao H, Nickel CH, Cantley LG, Bruggeman LA, Bennardo LN, Wang B. EphA kinase activation regulates HGF-induced epithelial branching morphogenesis. J Cell Biol 2003;162:1281-1292.
73. Kilic E, Kilic U, Wang Y, Bassetti CL, Marti HH, Hermann DM. The phosphatidylinositol-3 kinase/Akt pathway mediates VEGF’s neuroprotective activity and induces blood brain barrier permeability after focal cerebral ischemia. FASEB J 2006;20:1185-1187.
74. Namiecińska M, Marciniak K, Nowak JZ. VEGF as an angiogenic, neurotrophic, and neuroprotective factor. Postepy Hig Med Dosw 2005;59:573-583.
75. Takahashi M, Matsui A, Inao M, Mochida S, Fujiwara K. ERK/MAPK-dependent PI3K/Akt phosphorylation through VEGFR-1 after VEGF stimulation in activated hepatic stellate cells. Hepatol Res 2003;26:232-236.
76. Grako KA, Ochiya T, Barritt D, Nishiyama A, Stallcup WB. PDGF (alpha)-receptor is unresponsive to PDGF-AA in aortic smooth muscle cells from the NG2 knockout mouse. J Cell Sci 1999;112:905-915.
Annals of Clinical & Laboratory Science, vol. 38, no. 3, 2008208

77. Chen QP, Giannobile WV. Adenoviral gene transfer of PDGF downregulates gas gene product PDGFalphaR and prolongs ERK and Akt/PKB activation. Am J Physiol Cell Physiol 2002;282:C538-544.
78. Kang DE, Yoon IS, Repetto E, Busse T, Yermian N, Ie L, Koo EH. Presenilins mediate phosphatidylinositol 3-kinase/AKT and ERK activation via select signaling receptors. Selectivity of PS2 in platelet-derived growth factor signaling. J Biol Chem 2005;280:31537-31547.
79. Roberts PJ, Der CJ. Targeting the Raf-MEK-ERK mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene 2007;26:3291-3310.
80. Jiang Q, Zhou C, Bi Z, Wan Y. EGF-induced cell migration is mediated by ERK and PI3K/AKT pathways in cultured human lens epithelial cells. J Ocul Pharmacol Ther 2006;22:93-102.
81. Fridell YW, Jin Y, Quilliam LA, Burchert A, McCloskey P, Spizz G, Varnum B, Der C, Liu ET. Differential activation of the Ras/extracellular-signal-regulated protein kinase pathway is responsible for the biological consequences induced by the Axl receptor tyrosine kinase. Mol Cell Biol 1996;16:135-145.
82. Zhang YH, Wei W, Xu H, Wang YY, Wu WX. Inducing effects of hepatocyte growth factor on the expression of vascular endothelial growth factor in human colorectal carcinoma cells through MEK and PI3K signaling pathways. Chin Med J 2007;120:743-748.
83. Tétreault MP, Chailler P, Beaulieu JF, Rivard N, Ménard D. Epidermal growth factor receptor-dependent PI3K-activation promotes restitution of wounded human gastric epithelial monolayers. J Cell Physiol 2008; 214:545-557.
84. Kacar F, Meteoğlu I, Yasa H, Levi E. Helicobacter pylori-induced changes in the gastric mucosa are associated with mitogen-activated protein kinase (MAPK) activation. Appl Immunohistochem Mol Morphol 2007; 15:224-228.
85. Wang W, Luo HS, Yu BP. Expression of NF-kappaB and human telomerase reverse transcriptase in gastric cancer and precancerous lesions. World J Gastroenterol 2004; 10:177-181.
86. Doger FK, Meteoglu I, Ozkara E, Erkul ZK, Okyay P, Yükselen V. Expression of NF-kappaB in Helicobacter pylori infection. Dig Dis Sci 2006;51:2306-2309.
87. Nam SY, Lee HS, Jung GA, Choi J, Cho SJ, Kim MK, Kim WH, Lee BL. Akt/PKB activation in gastric carcinomas correlates with clinicopathologic variables and prognosis. APMIS 2003;111:1105-1113.
88. Kobayashi I, Semba S, Matsuda Y, Kuroda Y, Yokozaki H. Significance of Akt phosphorylation on tumor growth and vascular endothelial growth factor expression in human gastric carcinoma. Pathobiology 2006; 73:8-17.
89. Dragovich T, McCoy S, Fenoglio-Preiser CM, Wang J, Benedetti JK, Baker AF, Hackett CB, Urba SG, Zaner KS, Blanke CD, Abbruzzese JL. Phase II trial of erlotinib in gastroesophageal junction and gastric adenocarcin-omas: SWOG 0127. J Clin Oncol 2006;24:4922-4927.
90. Tong Q, Zheng L, Lin L, Li B, Wang D, Huang C, Li D. VEGF is upregulated by hypoxia-induced mitogenic factor via the PI-3K/Akt-NF-kappaB signaling pathway. Respir Res 2006;7:37.
91. Ouyang W, Li J, Ma Q, Huang C. Essential roles of PI-3K/Akt/IKKbeta/NFkappaB pathway in cyclin D1 induction by arsenite in JB6 Cl41 cells. Carcinogenesis 2006;27:864-873.
92. Brignole C, Marimpietri D, Pastorino F, Nico B, Di Paolo D, Cioni M, Piccardi F, Cilli M, Pezzolo A, Corrias MV, Pistoia V, Ribatti D, Pagnan G, Ponzoni M. Effect of bortezomib on human neuroblastoma cell growth, apoptosis, and angiogenesis. J Natl Cancer Inst 2006;98:1142-1157.
93. Scheele JS, Marks RE, Boss GR. Signaling by small GTPases in the immune system. Immunol Rev 2007; 218:92-101.
94. Tee AR, Manning BD, Roux PP, Cantley LC, Blenis J. Tuberous sclerosis complex gene products, Tuberin and hamartin control mTOR signaling by acting as a GTPase-activating protein complex toward Rheb. Curr Biol 2003;13:1259-1268.
95. Cai SL, Tee AR, Short JD, Bergeron JM, Kim J, Shen J, Guo R, Johnson CL, Kiguchi K, Walker CL. Activity of TSC2 is inhibited by AKT-mediated phosphorylation and membrane partitioning. J Cell Biol 2006;173:279-289.
96. Karbowniczek M, Cash T, Cheung M, Robertson GP, Astrinidis A, Henske EP. Regulation of B-Raf kinase activity by tuberin and Rheb is mammalian target of rapamycin (mTOR)-independent. J Biol Chem 2004; 279:29930-29937.
97. Zhu Y, Zhong X, Zheng S, Du Q, Xu W. Transformed immortalized gastric epithelial cells by virulence factor CagA of Helicobacter pylori through Erk mitogen-activated protein kinase pathway. Oncogene 2005; 24:3886-3895.
98. Karbowniczek M, Robertson GP, Henske EP. Rheb inhibits C-raf activity and B-raf/C-raf heterodimerization. J Biol Chem 2006;281:25447-25456.
99. Ma L, Teruya-Feldstein J, Bonner P, Bernardi R, Franz DN, Witte D, Cordon-Cardo C, Pandolfi PP. Identi-fication of S664 TSC2 phosphorylation as a marker for extracellular signal-regulated kinase mediated mTOR activation in tuberous sclerosis and human cancer. Cancer Res 2007;67:7106-7112.
.
Morphoproteomics and gastric cancer 209




















