
Modern pollen deposition in the tropical lowlands of northeast Queensland, Australia
29
REVIEW OF PALAEOBOTANY AND PALYNOLOGY ELSEVIER Reviewof Palaeobotanyand Palynology 83 (1994)299-327 Modern pollen deposition in the tropical lowlands of northeast Queensland, Australia G.M. Crowley, J. Grindrod, A.P. Kershaw Department of Geography and Environmental Science, Monash University, Clayton, Vic. 3168, Australia (Received July 27, 1993; revised and accepted January 25, 1994) Abstract Surface sediments of mangrove, freshwater wetland and rainforest sites in northeast Queensland were sampled to obtain pollen signatures from a range of climatic and vegetational settings as a basis for interpretation of fossil pollen diagrams. Maximum terrestrial pollen diversity was predicted by curve fitting using the Putter No. 1 growth curve. Taxonomic diversity was found to be a better indicator of rainfall zone than the presence or absence of any one taxon. However, the presence of Chenopodiaceae pollen and a general lack of rainforest pollen types are characteristic of low-rainfall environments. High values for pteridophytes indicate fluvial conditions, while high-altitude taxa were found in lowland sites fed by streams draining upland vegetation. Local habitat indicators provide good evidence for the type of depositional environment, in keeping with other published studies. 1. Introduction Modern pollen studies of the coastal plains of northeast Queensland have so far been concen- trated around fossil study sites (Grindrod and Rhodes, 1984; Grindrod, 1985). Steep climatic gradients in the region, particularly in relation to rainfall, are reflected in a broad range of vegetation community types. Shifts in climate during the Pleistocene and Holocene are likely to have resulted in marked shifts in vegetation. Palynological studies of vegetation history there- fore warrant a more comprehensive survey of present pollen dispersal and deposition. To this end a broad-ranging modern pollen study, based on analysis of surface mud and moss samples, has been undertaken as part of an ongoing study of the vegetation history of the lowlands and conti- nental shelf of northeast Queensland. This paper draws together modern pollen data from three mangrove communities in contrasting climatic and 0034-6667/94/$7.00 © 1994ElsevierScienceB.V. All rights reserved SSDI 0034-6667 (94) 00022-C physical settings, one lowland freshwater swamp and three lowland rainforest localities, in an attempt to characterise various vegetational com- munities by their pollen signatures. 2. Environmental setting The study area lies in the 300 km long coastal corridor between Townsville and Cairns in north- east Queensland (Fig. 1). It includes an extensive coastal plain up to 50 km wide, bounded to the west by escarpments and steep mountainous ter- rain reaching to 1600 m altitude in the Bellenden Ker Range. The coastal plain is predominantly Quaternary alluvium from the granitic and meta- morphic uplands, with long shore-parallel dunes backed by swamps at its seaward margin. The climate of the region is typified by warm rainy summers and cool dry winters (Nix and Kalma, 1972). However, the amount and seasonal-
Transcript of Modern pollen deposition in the tropical lowlands of northeast Queensland, Australia

REVIEW OF
PALAEOBOTANY AND
PALYNOLOGY E L S E V I E R Review of Palaeobotany and Palynology 83 (1994) 299-327
Modern pollen deposition in the tropical lowlands of northeast Queensland, Australia
G.M. Crowley, J. Grindrod, A.P. Kershaw
Department of Geography and Environmental Science, Monash University, Clayton, Vic. 3168, Australia
(Received July 27, 1993; revised and accepted January 25, 1994)
Abstract
Surface sediments of mangrove, freshwater wetland and rainforest sites in northeast Queensland were sampled to obtain pollen signatures from a range of climatic and vegetational settings as a basis for interpretation of fossil pollen diagrams. Maximum terrestrial pollen diversity was predicted by curve fitting using the Putter No. 1 growth curve. Taxonomic diversity was found to be a better indicator of rainfall zone than the presence or absence of any one taxon. However, the presence of Chenopodiaceae pollen and a general lack of rainforest pollen types are characteristic of low-rainfall environments. High values for pteridophytes indicate fluvial conditions, while high-altitude taxa were found in lowland sites fed by streams draining upland vegetation. Local habitat indicators provide good evidence for the type of depositional environment, in keeping with other published studies.
1. Introduction
Modern pollen studies of the coastal plains of northeast Queensland have so far been concen- trated around fossil study sites (Grindrod and Rhodes, 1984; Grindrod, 1985). Steep climatic gradients in the region, particularly in relation to rainfall, are reflected in a broad range of vegetation community types. Shifts in climate during the Pleistocene and Holocene are likely to have resulted in marked shifts in vegetation. Palynological studies of vegetation history there- fore warrant a more comprehensive survey of present pollen dispersal and deposition. To this end a broad-ranging modern pollen study, based on analysis of surface mud and moss samples, has been undertaken as part of an ongoing study of the vegetation history of the lowlands and conti- nental shelf of northeast Queensland. This paper draws together modern pollen data from three mangrove communities in contrasting climatic and
0034-6667/94/$7.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0034-6667 (94) 00022-C
physical settings, one lowland freshwater swamp and three lowland rainforest localities, in an attempt to characterise various vegetational com- munities by their pollen signatures.
2. Environmental setting
The study area lies in the 300 km long coastal corridor between Townsville and Cairns in north- east Queensland (Fig. 1). It includes an extensive coastal plain up to 50 km wide, bounded to the west by escarpments and steep mountainous ter- rain reaching to 1600 m altitude in the Bellenden Ker Range. The coastal plain is predominantly Quaternary alluvium from the granitic and meta- morphic uplands, with long shore-parallel dunes backed by swamps at its seaward margin.
The climate of the region is typified by warm rainy summers and cool dry winters (Nix and Kalma, 1972). However, the amount and seasonal-

300 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
2000
146°E
0 25 50 t I I
Kilornetres
145°E
\ Princess
150°E
Bay 15°S
2000 ,QUEENSLAND
25°S
/1
2500 V
.3750
atchem Inlet
h Wwud Swamp
• INNISFAIL
~2500
,2000
Study sites Modern pollen studies • )(- Present study
[] Kershaw (1973)
• Kershaw and Hyland (1975)
z~ Kershaw and Strickland (1991)
• Grindrod and Rhodes (1984)
0 Grindrod (1985)
Fossil pollen studies
• QuatemaPt
• Tertiary
~1000-,' mm rainfall isohyet
; ~ Land over 200m altitude
18os -
TOWNSVILLE Q
Fig. 1. The study area, showing mean annual isohyets, summarised topography, and the locations of published and present palynological sites.

G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299 327 301
ity of precipitation vary considerably along the coast according to local orographic effects. Hence, rainfall is highest and seasonality least marked where high coastal ranges intercept moisture-laden SE trade winds from the Coral Sea (Nix and Kalma, 1972). Mean daily temperatures through- out the area vary from 17 to 29°C (Tracey, 1982).
At a regional scale the vegetation has been mapped by Murtha (1972), Tracey and Webb (1975) and Tracey (1982). The most humid loca- tions contain a diverse suite of rainforest communi- ties with structural and floristic variations according to altitude, climate and soils. Typically the highest peaks are covered with simple micro- phyll vine thicket (or montane forest), while simple notophyll vine forest (or submontane forest) and mesophyll vine forest (or lowland forest) predomi- nate at mid-altitude and on the lower slopes, respectively. Complex mesophyll vine forest (or lowland forest) blanketed much of the humid areas of coastal plain north of Ingham prior to cultiva- tion for sugar cane. Rainforest remnants display high floristic diversity, with species of the Myrtaceae, Lauraceae, Euphorbiaceae, Rutaceae, Sapindaceae and Proteaceae particularly abundant.
South of Ingham on the coastal plain, seasonal aridity is pronounced and rainforests are replaced by floristically poorer open sclerophyll woodlands dominated by Eucalyptus drepanophylla, E. alba and Melaleuca viridiflora with grassy understories.
Mangrove communities line the tidal reaches of coasts and estuaries. They are particularly luxuri- ant and species rich where sheltered physiographic settings combine with high rainfall and an absence of seasonal aridity. As many as 45 mangrove tree and shrub species are recorded for the area (Bunt et al., 1982), with members of the Rhizophoraceae (Rhizophora, Bruguiera and Ceriops spp.) being the most abundant. On sandy coasts, mangroves are replaced by Casuarina equisetifolia above high- water mark. Beach strand vegetation, in which Eucalyptus tessellaris, E. pellita, Scaevola sericea, Argusia argentia and C. equisetifolia are major components, covers the higher coastal dunes. Extensive near-coastal dune and swale systems are covered by Melaleuca woodland. Only in the drier,
eucalypt-dominated habitats does fire play an important role in vegetation regulation.
3. Site descriptions and sample locations
Seven sites have been analysed for modern pollen deposition (see Fig. 1). Three marine sites, Mutchero Inlet, Hinchinbrook Channel and Cleveland Bay, encompass the range of climatic and physiographic settings for mangrove in the study area. Non-marine sites include Wyvuri Swamp at a humid coastal location, and 3 lowland rainforest sites (R1, R2 and R3) up to 30 km from the sea.
3.1 Mutchero Inlet
Mutchero Inlet (17°14'S, 145°58'E) is the common tidal estuary of the Russell and Mulgrave rivers which drain the Atherton Tableland, Bellenden Ker Range and adjacent coastal plains (see Fig. 1). It receives approximately 4000 mm annual mean rainfall. The catchment of the Russell River is still largely vegetated by native forests, while that of the Mulgrave is mostly cleared for cultivation and pasture. Native vegetation in the area includes open eucalpyt forests on the steeper slopes, and a diversity of rainforest community types on both the ranges and coastal lowlands. Melaleuca forest and mesophyll palm vine forest (or swamp forest), are common on poorly drained lowlands sites. The inlet contains a diverse, closed- canopy mangrove forest with an areal extent of approximately 5 km z (Fig. 2). Rhizophora apicu- lata, R. stylosa, Bruguiera parviflora, B. gymnorr- hiza and Avicennia marina are prominent trees at seaward locations where tidal influence is strong. Further landward common mangrove trees include Xylocaryous granatum, Heritiera littoralis and Lumnitzera racemosa. The understorey shrub Aegiceras corniculatum, the ground fern Acrostichum aureum and epiphytic ferns including Platycerium bifurcatum and Drynaria rigidula are also prominent. A narrow fringing mangrove com- munity dominated by Sonneratia caseolaris with an understorey of A. corniculatum occurs along riverbanks in the upper reaches of the estuary.

302 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
0 1 2 I I I
Kilometres
: : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : " : :
• i i i i i ! i i i i i
Bellenden Ker
o Miriwinni Bartle Frere w /
~ T ~ Mesophyll/complex mesophyll vine forest
~ Vine forest with sclerophyll emergents
~ Swampy coastal plains: Paperbark swamp forest (Mela/euca quinquenetvia) with raised islands of vine forest
~ Paperbark swamp forest [ Cleared land or bare sand (Me/aleuca quinquenervia)
~ Coastal beachndges . . . . . . Lowtide level (.--~'_~ Intertidal muo'flat
~ Mangrove M31D Modern pollen sample
Fig. 2. The Mutchero Inlet study site, showing summarised vegetation and surface sample locations.
Except where clearing has occurred, the mangroves grade into Melaleuca swamp forest, complex low- land rainforest or palm-dominated forest.
Fifteen surface mud samples were collected at Mutchero Inlet, from seaward sites near the con- fluence of the Russell and Mulgrave rivers to the inland limit of saltwater penetration, including one sample from the S. caseolaris community 9 km upstream from the sea (see Fig. 2 for sample locations, and Table 1 for vegetation summaries at each site).
3.2 Hinchinbrook Channel
The Hinchinbrook Channel ( 18 ° 19'S, 146°07'E)is a marine corridor about 6 km wide between Hinchinbrook Island and the mainland (Fig. 3). Limited rainfall records for nearby
Missionary Bay indicate that the area receives between 2000 and 3000 mm/yr (Bunt et al., 1982). Abrupt, mountainous terrain to over 1000 m alti- tude to the east, and steep slopes to the west, provide a wide range of vegetational environments in close proximity to the Channel. Melaleuca open woodland and low eucalypt woodland occur within 2 km of the western shore, while woodlands domi- nated by Allocasuarina littoralis and rainforest occur to the east (Tracey and Webb, 1978).
The tidal reaches of the channel contain extens- ive mangrove communities. Monospecific stands of Rhizophora stylosa command the broad mid- channel shoals, while more diverse communities containing R. stylosa, Bruguiera gymnorrhiza, Sonneratia alba, Avicennia marina, Ceriops tagal, Lumnitzera rosea, Excoecaria agallocha and Aegiceras corniculatum fringe more shoreward

G.M. Crowley et aL /Review of Palaeobotany and Palynology 83 (1994) 299-327 303
Table 1 Local vegetation of surface sample sites from Cleveland Bay, Hinchinbrook Channel, Mutchero Inlet, Wyvuri Swamp
Study Location Site description/vegetation type Taxa recorded Sample Number (height in m) (5 m 2 quadrat at sample site)
Cleveland Bay CI, C 2 Supra-tidal mudflat/sparse Halosarcia indica
chenopod shrubland (<0.4 m), and bare ground
C3 High-tide mangrove zone/mangrove shrubland (< 1.0 m)
C4 Intertidal mangrove/tall Avicennia marina mangrove shrubland (< 3.0 m)
C5, C6, C7, C8, C9 Intertidal mangrove/mangrove Rhizophora stylosa closed forest (5-6 m)
C10 Intertidal mangrove/mangrove Avicennia marina woodland (3-5 m)
C11, C12. C13 Low-tide mudflat, 100 to 300 m seaward of mangrove fringe/unvegetated
Hinchinbrook Channel H1 Freshwater swamp/woodland
(5-7 m) H2 High-tide mangrove/open
shrubland (2-3 m) H3, H4 Intertidal mangrove/shrubland
(2-3 m) H5 Intertidal mangrove/forest (5 6
m) H6, H7, H8, H9 Intertidal mangrove/closed
forest (5 6 m) H10, H11 Intertidal mangrove/closed
forest (5-6 m) H12, H13 Tidal to supratidal
riverbank/riparian shrubland (2-3 m)
H14, H15 Tidal riverbank/riparian woodland (3-6 m)
H16, H17, H18 Tidal riverbank/riparian woodland (4-6 m)
Mutchero Inlet M1, M2 Supra-tidal swamp/swamp forest
(5-6 m) M3 Tidal riverbank/gallery forest
( 15-20 m) M4, M5 Supra-tidal to high-tide
mangrove transition/closed forest (8-12 m)
Avicennia marina/Aegialitis annulata/Ceriops tagal
Melaleuca viridiflora/Cyperaceae spp./Pandanus spp.
Bruguiera gymnorrhiza/Ceriops tagal/Osbornia octodonta/Aegialitis annulata
Bruguiera gymnorrhiza/Ceriops tagal/Lumnitzera littorea
Rhizophora stylosa/Bruguiera gymnorrhiza/Ceriops tagal/Avicennia marina
Rhizophora stylosa
Sonneratia alba/Avicennia marina/Aegiceras corniculatum
Acrostichum speciosum/Clerodendrum inerme/Dalbergia candenatensis/Dendrobium spp./Heritiera littoralis/Hibiscus tiliaceus/Lumnitzera littorea/Osbornia octodonta/Pandanus tectorius
Acanthus ilicifolius/Acrostichum speciosum/Aegiceras corniculatum/Avicennia marina/Bruguiera gymnorrhiza/Dalbergia candenatensis/Excoecaria agallocha/Heritiera littoralis/Platycerium bifurcatum/Rhizophora stylosa
Acrostichum speciosum/Bruguiera gymnorrhiza/Excoecaria agallocha/Flagellaria indica/Gagnia aspera/Heritiera littoralis/Pandanus tectorius/Platycerium bifurcatum/Rhizophora stylosa/Xylocarpus granatum
Acrostichum speciosum/Melaleuca dealbata/Pandanus tectorius/Parsonsia straminea/Stenochlaena palustris
Aegiceras corniculatum/Cassia retusa/Dendrobium spp./Drynaria rigidula/Sonneratia caseolaris
Acacia aulacoearpa/Acrostichum speciosum/Asplenium nidus/Bruguiera gymnorrhiza/Bruguiera sexangular/Cassia retusa/Heritiera littoralis/Lygodium reticulatum/Melaleuca dealbata/Pandanus teetorius/Randia fitzlanii/Schefftera actinophylla/Stenochlaena palustris/Xylocarpus granatum

304
Table l (continued)
G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
Study Location Site description/vegetation type Taxa recorded Sample Number (height in m) (5 m 2 quadrat at sample site)
M6, M7, M8 Intertidal mangrove/closed tbrest (7 9 m)
M9, MI0, Ml l Intertidal mangrove/closed forest (7 9 m)
M 12, M 13 Intertidal mangrove/closed forest (7-9 m)
M 14, M 15 Intertidal mangrove/closed/ forest (10 m)
Wyvuri Swamp W1 Coastal dune/woodland (12 m)
W2 Swamp/Restionaceae fenland (0.4 m)
W3, W4 Swamp/open woodland (5 7 m) W5, W6 Swamp/closed forest (5 8 m)
Acanthus ilicifolius/Acrostichum speciosum/Aegiceras corniculatum/Bruguiera gymnorrhiza/Cassia retusa/Crinum pedunculatum/Drynaria rigidula/Heritiera littoralis/Platycerium biJurcatum/Sonneratia alba
Rhizophora stylosa
Acrostichum speciosum/Xylocarpus granatum
Avicennia marina/Bruguiera gymnorrhiza/Rhizophora so'losa
Allocasuarina littoralis/Cassytha spp./Grevillea pinnatifida/Lygodium microphyllum/Stephania japonica/Syzygium forte/Wilkiea uegeliana
Leptocarpus ramosus
Cyperaceae spp./ Melaleu~ a viridiflora Drynaria rigidula/ Melaleuca viridiflora/Pandanus
tectorius/Platycerium bifurcatum/Poaceae spp./Schefflera actinophylla/Stenochlaena palustris
localities. At their landward limit mangroves are replaced by coastal woodlands characterised by Melaleuca viridiflora, Pandanus spp., Clerodendrum inerme, Dalbergia candenatensis and Allocasuarina littoralis.
Eighteen samples were analysed from Hinchinbrook Channel. These represent a range of vegetational environments including mixed mangrove communities on the western shore, monospecific mid channel communities, Sonneratia-dominated mangrove forest and non- mangrove riparian communities along brackish creeks to the east (Fig. 3; Table 1).
3.3 Cleveland Bay
Cleveland Bay (19°lYS, 146°45'E), lies on the low relief coast near Townsville (Fig. 4). Rainfall is approximately 1200 mm/yr, strongly biased to the summer period, and winter aridity is pro- nounced. Terrestrial vegetation is open woodland dominated by Eucalyptus drepanophylla and E. alba with grassy understories. Samphire communi- ties, characterised by the chenopods Halosarcia spp., Tecticornia spp. and Suaeda arbusculoides
cover broad high-tide mud flats. Beach ridge vege- tation, heavily infested with the introduced shrub Parkinsonia aculeata covers the long chenier com- plex running east from Sandfty Creek. Mangrove communities less than 150 m wide fringe the coast in its mid-tidal reaches. These are neatly zoned, with a landward community dominated by stunted Avicennia marina with Ceriops tagal and Aegialitis annulata, a middle community of tall Rhizophora stylosa, and a narrow, seaward border of Avicennia marina. Surface samples analysed for Cleveland Bay represent low-tide mudflat and mangrove envi- ronments (Fig. 4; Table 1.)
3.4 Wyvuri Swamp
Wyvuri Swamp (17°21'S, 146°01'E) is part of a 7 km 2 freshwater wetland impounded by coastal dune ridges (Fig. 5). The site receives approxi- mately 4000 mm annual precipitation. The dune ridges to the east support a narrow seaward fringe of Casuarina equisitifolia, and coastal dune com- munities with Eucalyptus tessellaris, Scaevola seri- cea, Terminalia catappa, Syzyguim rubiginosum and Eugenia hemilampra. Older, more landward ridges

G. 31. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327 305
. . . . . Approximate catchment ~ Paperbark swamp forest [ ] Mangrove bounda i 7 ~-"J::::= (Melaleuca quinquenervia)
Stunted paperbark swamp 1 '" H igh tide level |~-.::~:] forest (Melaleuca viridiflora) Samphire
HtOI-qModempo,en=.~,s ~-~W~la=¢E~usd,~J ~]Jnte.=~m~
~st~ Mt
Dlamant ina
~ham /
Fig. 3. The Hinchinbrook Channel study site, showing summarised vegetation (after Tracey and Webb, 1978), and surface sample locations.
support Eucalyptus intermedia open forest. A mosaic of vegetation communities covers the swamp surface. Melaleuca quinquinervia open forest with an understorey of Scleria sp., Gahnia sp. and Rhyncospora corymbosa occurs at water depths to approximately 1 m, while monospecific fenlands of Leptocarpus ramosus occur where peaty substrates accumulate above the dry season water level. Five sample site locations from within Wyvuri Swamp and one on the beach ridge barrier are shown in Fig. 5, while details of vegetation at each are given in Table 1.
A list of emergent, canopy and sub-canopy species appears in Appendix 1. All sites contain a high diversity of species, representing a large number of families. The families with the greatest species presence are Lauraceae (RI: 11, R2: 13, R3: 12) Myrtaceae (7, 2, 10), Sapindaceae (8, 9, 6), Rutaceae (6, 7, 3), Moraceae (2, 7, 3), Meliaceae (6, 5, 2), Monimiaceae (2, 5, 1), Proteaceae (2, 4, 4), and Sapotaceae (4, 1, 2).
4. Methods
3.5 Rainforest sites
The three lowland rainforest sites, R1 (17°51'S, 146°04'E), R2 (17°25'S, 145°54'E) and R3 (17°39S, 145°52'E), are located on Fig. 6. These are situated on the 3 major soil types in the region (Table 2).
All pollen-analytical results are based on surface sediment samples. The long periodicity of flower- ing of regional rainforest plants and year to year variation in flowering intensity (Kershaw and Strickland, 1990) precluded a viable pollen trap- ping study. Chemical preparation of all samples

306 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
0 1 I I
Kilometres
Magnetic Island
~ Woodland (Eucalyptus alba, E. drepanophyl/a)
~ Coastal beach ridges (Parkingsonia)
~ Mangrove (Avicennia marina, Ceriops taga/, Rhizophora sty
- - Aegia/itis annu/ata)
Samphire
~ - - ~ Intertidal mudflats
High tide level . . . . . . Low tide level
[ ] Modem pollen rain site
Fig. 4. The Cleveland Bay study site, showing summarised vegetation and surface sample locations.
followed Fa~gri and Iversen (1975). Identifications were made under a light microscope by comparison with reference specimens held in the Department of Geography and Environmental Science at Monash University. Where possible, a minimum of 150 pollen grains was counted in each sample. The pollen sum, used to construct the pollen diagrams, includes all identified pollen grains and excludes pteridophyte spores. The pollen diagrams include well represented pollen types only. Pollen and spore taxa are grouped according to habitat and habit (Appendix 2) in each diagram. Composite diagrams for selected taxa are also presented.
Between-site comparisons of taxonomic diver- sity are based on asymptotic calculations applied
to combined pollen assemblage data. For each site, samples were randomly ordered and cumulative calculations made of terrestrial pollen count and terrestrial pollen diversity. This was done three times, each time using a different random order. The three sets of data thus generated were then pooled to calculate the curve of best fit for the asymptotic Putter no. 1 growth curve (Ricker, 1979):
D ~ Doo - - o (lnD~ kC)
in which C is the terrestrial pollen sum, k is a constant, D is the terrestrial pollen diversity, and Do~ is the theoretical maximum terrestrial pollen diversity, an asympote, i.e. D at C is ~ .
Curve fitting was done manually by choosing a

G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327 307
0 1 2 t I I
Kilometres
C O R A L
SEA
Bramston Beach
~ ' - ~ Mesophyll vine forest
~ Vine forest with sclerophyll ernergents
~ Coastal beach ridge oomrnunitles
l Paperbark swamp forest (Melaleuca quinquenervia)
~ Mangrove
Restiad swamp (Leptocarpus ramosus)
Intertidal mudflat
W3 [] Modem pollen sample
Fig. 5, The Wyvuri Swamp study site, showing summarised vegetation and surface sample locations.
Table 2 Characteristics of rainforest surface sample sites
Site Altitude Vegetation Rainfall Parent material Soil type (m) (mm/yr)
R1 60 Complex mesophyll vine forest 4300 Granite dominated by Backhousia bancroflii
R2 230 Complex mesophyll vine forest 3500 Basalt R3 30 Mesophyll evergreen vine forest 3 1 7 0 Metamorphics
Alluvium
Basaltic red earth Brown earth
A complete species list is presented in Appendix 1.
value for D and calculating the r value for regres- sion o f the equation:
l n ( O o ~ - D ) - l n ( D o ~ ) v s . C
fitted th rough the origin (Steel and Torrie, 1960).
The value o f D~o which gave the highest r 2 value was selected as the mos t likely theoretical maxi- m u m value for terrestrial pollen diversity. As the r 2 values and probabil i ty statistics would have been artificially elevated by the generation o f three

308 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299 327
Fig. 6. Location and summarised vegetation at the lowland rainforest study sites.
sets of data from the same initial sets or data, these values are not presented here.
The use of asymptotic equations predicting taxo- nomic diversity avoids the problems of the compar- ison of taxonomic diversity of samples caused by the clearly dependent and varying relationship between taxonomic diversity and pollen count size. However, caution must be exercised in interpreting the results. In this case a simple comparison is made between the regional vegetation of different rainfall zones as represented by terrestrial pollen taxa. Pteridophyte spores and wetland pollen types are omitted because inherent vagaries of riverine dispersal and localised deposition may detract from the regional picture.
4.1 Taxonomic diversity
The equations which best described the relation- ship between D~ and C were:
(a) Mutchero Inlet D = 4 4 - - e t3"78°2- o.oo3oc) i.e. Do~ = 44
(b) Hinchinbrook D = 3 4 - - e ~3"5264- 0"°°16cl
Channel (c) Cleveland Bay D = 11 - e ~2"3979 o.oo85c) (d) Wyvuri Swamp D=41-e ~3"7136-°'°°17c) (e) Rainforest sites D=47-e 13"sS°l-°'°°z°c)
i.e. Do~ = 34
i.e. D~=ll i.e. Do~ =41 i.e. D~=47
These equations are presented in Fig. 7 along with individual sample data points and totalled sample points for D against C. It should be noted that the equations produced to estimate maximum ter- restrial taxonomic diversity of each region are merely curves of best fit and that these and other values of Do~ produced significant results. However, the asymptotic values generated here increase with rainfall corresponding with increas- ing botanical diversity, with Do~ in the lowest- rainfall zone being 11, in the medium-rainfall zone 34, and in the highest-rainfall zone ranging from 41 to 47. The total number of terrestrial taxa recorded from all but one site was within 3 taxa of the Do~ values obtained. The total terrestrial taxa recorded for Wyvuri Swamp was by contrast only 35, lower than would be expected when

G.M Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327 309
E
(a) MI.~CIIIERO INLET 50. ' S 40'
130'
20'
10'
, , , , , , w , • , , , , ' , • , , , , ,
50 ; (b) HINCHINBROOK CHANNEL
40"
3 0 : ~
20 ~
lO i • i , , , , , I , , I , i i i * | J , i i
502 (¢) CLEVELAND BAY
40
30 ~
20 .
. . . . . . . . . . . . . . . . . .
0 200 400 600 800 1000 1200 1400 1600 1800 2000
(~ WY~JRI SWAMP
, , , • , • , , , , , , , , , , • , , ,
e) RAIN'FOREST SITES
(f) (e)
• . - . . . . . . . . . . . . (a)
0 200 400 600 800 1000 1200 1400 1600 1800 2000 Terrestrial pollen count
Fig. 7. Predicted taxonomic diversity for terrestrial pollen assemblages at each study site, using calculated curves of best fit for the asymptotic Putter No. I growth curve (Ricker, 1979).
compared to the other sites in the high-rainfall region and despite the high terrestrial pollen counts achieved. However, the curve in Fig. 7 clearly shows that, with further counting, taxonomic diversity levels comparable to the other sites are expected.
5. Pollen spectra
5.1 Mutchero Inlet
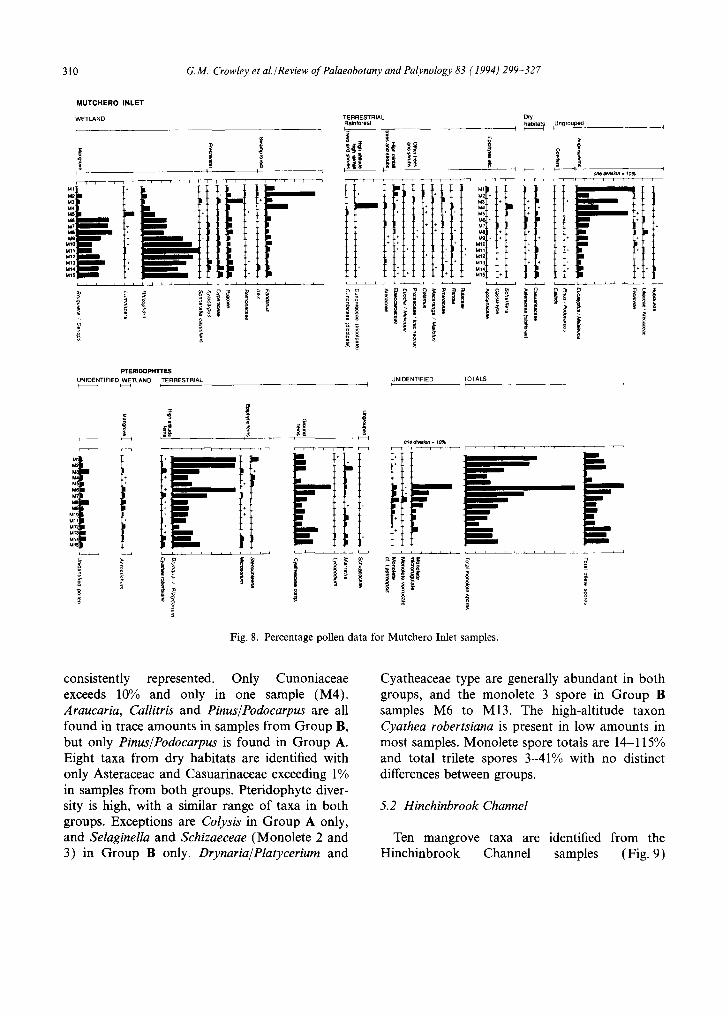
Pollen spectra from Mutchero Inlet fall into two groups (Fig. 8): samples from swamp forests or mangroves of low salinity sites at the limit of saltwater influence (Group A: M1-Mb), and samples from sites within the Rhizophoraceae- dominated mangrove system (Group B: M6-M15). Group A samples are dominated by Pandanus or Eucalyptus/Melaleuca. Mangrove values are up to 31% with Bruguiera 1-6% and
Rhizophora 4-13%. Lumnitzera, usually less than 1% reached 10.5% in M5. Group B samples are dominated by Rhizophoraceae, with Bruguiera most abundant in samples M6-M8, and Rhizophora in samples M9-M15. Eucalyptus/ Melaleuca have values of 5-14%. Despite differ- ences in the relative importance of individual taxa between the two groups, the range of taxa repre- sented is remarkably consistent. Of the five mangrove species recorded, four are found in Group A samples and five in Group B samples. In both groups, saltmarsh, represented only by Chenopodiaceae, is less than I%. Six fresh water swamp species are identified, with a similar range of taxa in both groups. Swamp taxa of both groups are dominated by Poaceae, which reached 17% in M3, a site adjacent to a cane field. Pandanus and Ilex are the only swamp forest taxa identified in both groups. Although 23 rainforest taxa are iden- tified, most occurring in both groups, none is
310 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
M U T C H E R O I N L E T
WETLAND TERRESTRIAL Rainforesl I
,
~ i i L i i i i ~ i i i i i i ,
L ~.12 *
M S +
M ~ O
M 1 2
M 1 3
• M I 4 +
Dry t ~ t u n g r ° u l ~
I'F-- ! PTEIRIDOPHYTES
UNIDE NTIFIEO WETLAND TERRESTRIAL I U FNIDENTIF IED I TOTALS F
M10 Ml l M12 M~3 Mr4
C =
a ~
o n e a ~ . ~ 0 ~ , T , , , r T i IIr-
, i , L i i ,
l •
Fig. 8. Percentage pollen data for Mutchero Inlet samples.
consistently represented. Only Cunoniaceae exceeds 10% and only in one sample (M4). Araucaria, Callitris and Pinus/Podocarpus are all found in trace amounts in samples from Group B, but only Pinus/Podocarpus is found in Group A. Eight taxa from dry habitats are identified with only Asteraceae and Casuarinaceae exceeding 1% in samples from both groups. Pteridophyte diver- sity is high, with a similar range of taxa in both groups. Exceptions are Colysis in Group A only, and Selaginella and Schizaeceae (Monolete 2 and 3) in Group B only. Drynaria/Platycerium and
Cyatheaceae type are generally abundant in both groups, and the monolete 3 spore in Group B samples M6 to M13. The high-altitude taxon Cyathea robertsiana is present in low amounts in most samples. Monolete spore totals are 14-115% and total trilete spores 3-41% with no distinct differences between groups.
5.2 Hinchinbrook Channel
Ten mangrove taxa are identified from the Hinchinbrook Channel samples (Fig. 9)

G. M, Crowley et al./ Review of Palaeobotany and Palynology 83 (1994) 299-327 311
HINCHINBROOK C H A N N E L
H i "
H 5 + ~ H 6 ÷ ~ H7~
H10" + H~11
H13i r m m u
H16 H17
W E T L A N D TERRESTRIAL
Rainforest Dry habitats
r ~ i , [ , i i , r ; i ~ ~ i [ I r , i r r i i [ i i , m 1 I , , , , , i r ,
m -u m + iN - j '+ +
m ' +
- I II
m
i m u m - - m m
] •
i -
i i i I , i t , i l i i t I I I I I I l
\ m
m
H I N C H I N B R O O K CHANNEL
TERRESTRIAL (cont.) I I
Ungmuped I i
) ) I
, r , ,
H 2 - , ~ H3 H4 H5 H6
H10 I"111 H12 H13
H15' H16 H17' H18
1 , , i , 1 ~ r
¢
+t +
+ _ + +
+ _ + + _ +
.+ I [
I I I
P T E R I D O P H Y T E S
UNIDENTIF IED TERRESTRIAL
I [ I l I i i
.1. +
t ! t ] ..t. 4
i i i J i i i i I f
¢
J ) I I
, - , , , . . . .
-+ +
"+
+ +
+
L i i i i i i i i i i I i
° I
TOTALS t
i
m
, l l l l l l l 1 , , I
E
m
Fig. 9. Percentage pollen data for Hinchinbrook Channel samples.

312 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
Rhizophoraceae exceeds 40% in the downstream samples H3-Hl l . Rhizophora is generally the dominant taxon, followed by Bruguiera/Ceriops, except in H8, a mid-channel sample in which Bruguiera/Ceriops reaches 6 7 % . However, Bruguiera/Ceriops is recorded with values as low as 1% in sites where this taxon is present in the vegetation. Of the remaining three mangrove taxa which reached 1%, Avicennia and Lumnitzera are found at background levels of 5% or less. Highest values of these taxa do not correspond to the sites in which these species are present in the vegetation. Sonneratia alba reached 5% only at the two sites from which it is recorded in the vegetation (H10, H11 ). Poaceae is the most consistently represented freshwater swamp taxon and no freshwater swamp species were recorded from the vegetation at the time of sampling. Of the swamp forest taxa only Pandanus, is recorded in the pollen spectra and this is at levels less than 5%, except in H16-H18 in which Pandanus is found growing and in which the taxon reaches 30-52% of the pollen sum. Fourteen rainforest and allied taxa are represented in the samples in trace amounts and samples from sites adjacent to rainforest in Deluge Inlet (H 12-H 18) have the highest rainforest representa- tion. Calamus and Proteaceae are recorded at levels of less than 4% and are not recorded in the vegetation of the sites. Nothofagus, is not recorded at all for the vegetation of north Queensland. The presence of this pollen type may be attributed to laboratory contamination or the re-working of older deposits. The conifers Pinus/Podocarpus, not recorded in the vegetation of the sites, have back- ground levels. Drier habitats are represented at trace levels by five taxa and at levels of less than 3% by Banksia and Haloragis type, neither of which have parent plants recorded in the site vegetation. Casuarinaceae has background levels of 2-11% in downstream sites H6-Hl l and achieves levels of up to 37% in the upstream sites. Such high values may be explained by influx from large areas of Allocasuarina littoralis and A. toru- losa growing on the escarpments and drier uplands of both the Cardwell Range to the west of the Channel and on Hinchinbrook Island to the east (Tracey, 1982). Of the ungrouped taxa, Acacia
maintains low representation in all samples from the eastern side of the channel, but is not recorded in the vegetation of any site, and Eucalyptus/ Melaleuca has variable values. The decline of Eucalyptus/Melaleuca from 42%0 in H1, in which Melaleuca viridiflora was growing, to 8% in sample H5, suggests that in these samples the pollen type was mostly derived from this species. Fluctuating values of 4-11% in the downstream sites H6 and Hl l and 11-18% in the upstream sites, probably reflect regional vegetation in which Eucalyptus species are prevalent. Seven pteridophyte spore types (summarised into 4 categories in Fig. 9) are identified in the samples. Only the samples adjacent to rainforest in Deluge Inlet (H12-H18) contain pteridophyte values greater than 2%. In H12-H15, pteridophyte representation ranges from 35 to 129% of the pollen sum, with Drynaria/Platycerium being the major contributor.
5.3 Cleveland Bay
Six mangrove taxa are recorded from Cleveland Bay samples, four reaching more than trace levels (Fig. 10). Of these Rhizophora and Bruguiera/ Ceriops are the most abundant. Rhizophora values exceed 50% in all samples from sites in which Rhizophora is found growing, and Bruguiera/ Ceriops exceeds 25% in all samples from sites in which Ceriops tagal is growing. However, both taxa have high values in adjacent sites. Avicennia representation ranges from 0 to 8% both inside and outside stands of Avicennia marina. Excoecaria, Lumnitzera and Xylocarpus, all found in the pollen spectra in trace amounts, are not recorded in the local vegetation. Aegialitis, which is present at many sample sites, is not recorded in the pollen spectra.
Chenopodiaceae is the only saltmarsh taxon recorded, reaching 24% in C1 at the landward mangrove fringe where Halosarcia indica is abun- dant. However, at C2, also in this vegetation community, values are only 4%. Chenopodiaceae values from sites in which the taxon is absent from the vegetation range between 0 and 8%, the highest value in the offshore sample C13. Four freshwater swamp taxa are recorded; Restionaceae and

G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327 313
CLEVELAND BAY WETLAND
I.- ~ = l~ f ° r es t D ry ha io i ta ts , Ung rouped I I . - -UNIDENTtFIED, ~ I - - - 4
• ~ . i1 | o r~ d~v~on . ~o%
TERRESTRIAL
L I II LLI Li ~ ~ ~g .
i i ~ , L L i J L , ~ i i i i i L L i ,
PTERIDOPHYTES WETLAND TERRESTRIAL TOTALS
=_
o3 s
Fig. 10. Percentage pollen data for Cleveland Bay samples.
Triglochin in trace amounts only. Highest values of Poaceae are found on the seaward side of the mangroves and across the unvegetated intertidal mudflat.
Values of Cyperaceae of up to 7% are recorded from many of the samples. Pandanus, the only swamp forest taxon represented and itself absent from the local vegetation is also found in trace amounts. Three rainforest and allied taxa are recorded in the pollen counts, and possible parent plants of each are found in the area but outside rainforest communities. These are Arecaceae which may be derived from strand line introduced Cocos nucifera, Elaeocarpaceae, which are most likely derived from riparian scrub Elaeocarpus spp., and Trema, a rainforest or sclerophyll element also found in strandline scrub. Argusia type, probably derived from Argusia argentea, but possibly from Terminalia catappa, both strandline species, is found in one sample at 4%, although its parent plants are also not recorded in the local vegetation. Eucalyptus/Melaleuca representation shows sim- ilarity to that of Poaceae, remaining at low levels through the landward mangrove samples, and increasing in the seaward mangrove and offshore samples, although no parent taxon is found within the sampling sites. Both taxa are therefore indica- tive of regional vegetation. The introduced taxon
Plantago occurs at 4% in C13, and is not recorded in the local vegetation. The high level of uniden- tified pollen in samples Cll-C13 reflects many highly degraded grains from the exposed mudflats. Total pteridophyte spore values are lower than 3% in all samples. Only five taxa are represented, none of which exceeds 1% of the pollen sum.
5.4 Wyvuri Swamp
Samples from Wyvuri Swamp show considerable variation reflecting the widely different habitats sampled (Fig. 11). Wl, from dune woodland, is dominated by Eucalyptus/Melaleuca type and con- tains no mangrove or saltmarsh, taxa. The only other taxa represented are Pandanus, Arecaceae, Flagellaria, Asteraceae, Casuarinaceae, Acacia, Colysis and Drynaria/Platycerium. W2, from monospecific Leptocarpus swamp, is dominated by Restionaceae. A wide range of other taxa from all habitats is represented, but only Casuarinaceae and Drynaria/Platycerium exceed trace amounts. W3 and W4, from Melaleuca swamp forest, are dominated by Cyperaceae, Poaceae and Eucalyptus/Melaleuca type. Although a large number of taxa are represented, Cladium typ6, Casuarinaceae and Fabaceae are the only other taxa to exceed trace values. W5 and W6, from tall

314 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
W Y V U R I S W A M P
WETLAND
-13
I one d iv i s ion = 10%
r l l l l l l [ l l l l l l l l
W +
W I I I l t l l l l l l [ l l l l
I
* i I i r r , , i I t i i 1 i
I: I I I I [ I I I I I I 1 I I I ~ l l
PTERIDOPHYTES
TERRESTRIAL k
TERRESTRIAL TOTALS I I I ,I
W5 W6
L L
o
t
r i ,
I I I
~ n = 1 ~ i l l l l
i'i
m ¢ - -
I I
I I I t I I
O O
~ g
Fig. 11. Percentage pollen data for Wyvuri Swamp samples.
Melaleuca swamp forest, are dominated by Poaceae, Pandanus and Eucalyptus/Melaleuca type with Ilex, Casuarinaceae, Drynaria/Platycerium
and Schizaeceae the only other taxa to exceed trace values. Cyathea robertsiana is present in trace amounts in W5 and W6 only.

G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327 315
5.5 Rain forest sites
No mangroves are represented in the rainforest samples and the only saltmarsh taxon, Chenopodiaceae, has values of less than 1% (Fig. 12). Poaceae is the only freshwater swamp taxon recorded, with values between 2 and 15%. Rainforest and allied taxa comprise over 80% of the pollen sum. No pollen is recorded from high- altitude elements. Values for high-rainfall tree taxa range from 4 to 27% for Euodia/Melicope, 1-14% for Elaeocarpaceae, and 0-3% for Prunus. The best represented of the ungrouped rainforest trees and shrubs are Calamus, Eugenia type and Macaranga/Mallotus. The rainforest vines Entada and Flagellaria attain values of 1-4% and 4-6%, respectively. Four rainforest edge taxa, Acacia aulococarpa, Maesa, Trema and the introduced Lantana, are represented by values up to 6%. None of these is recorded in the local vegetation. Similarly the drier habitat components Casuarinaceae and Asteraceae, and the ungrouped taxa Solanum, Muehlenbeckia type, Eucalyptus/ Melaleuca, Clematis comp. and Acronychia are absent from the local vegetation, but represented by pollen values up to 10%. Prominent pterido- phytes in the pollen spectra are the epiphytes Drynaria (3-8%) and Platycerium (2-15%), the groundferns Nephrolepis and Pellaea, and the treef- ern group Cyatheaceae.
5.6 Wetland~terrestrial ratios
Sites from Mutchero Inlet fall into two cate- gories with regard to wetland/terrestrial ratios (Fig. 13). Samples from mangrove habitats have mangrove representation exceeding 50% and up to 80%. Sites in mangrove/terrestrial habitats have mangrove representation of less than 31% except site M3 which was collected from within a bank of Sonneratia caseolaris less than 10 m wide fring- ing the Mulgrave River on the edge of a cane field. Sonneratia pollen is typically under-represented in Australian pollen species (Grindrod, 1988a). So a low total mangrove pollen count could be expected from this site. A high wetland component in this sample, is largely composed of Poaceae pollen, which in this case may be derived from weeds of
cultivation or sugar cane. In all other samples the freshwater wetland component is below 15%. Generally the Mutchero Inlet mangrove sites have swamp forest values of below 6% while the mangrove/terrestrial sites have values of 7-54%. The only exception is M5, a site within extensive mangrove forest with adjacent Melaleuca dealbata whose pollen has not been separated from more terrestrial Melaleuca and Eucalyptus species. Swamp forest, in this case Pandanus, exceeds 9% only in sites in which P. tectorius is growing. The terrestrial component is less than 30% in all man- grove sites (M6-M15), but is also low in M2 (28%) in which Pandanus dominates the pollen sum. At other mangrove/terrestrial sites the ter- restrial component is inflated (61-80%) by high Melaleuca/Eucalyptus values. Saltmarsh is negligi- ble (< 1%) in all samples.
All samples from Wyvuri swamp have negligible mangrove (<1%) and saltmarsh (<1%) values, reflecting the low salinity of the sites sampled. Site W1, from the dune woodland is dominated by terrestrial pollen (97%), and site W2, from within the Leptocarpus swamp, is dominated by fresh- water wetland taxa (96%). Three swamp-forest sites, W3-W5, have 64 to 71% freshwater wetland taxa and 28-35% terrestrial taxa. In these samples the terrestrial component is probably inflated by pollen from local swamp Melaleuca spp., whose pollen is not differentiated from the terrestrial Melaleuca/Eucalyptus category. Site W6, also in swamp-forest and adjacent to W5, is dominated by swamp-forest taxa (83%), predominantly Pandanus. This, and the differences in Pandanus representation between sites M 1 and M2, indicates that the Pandanus pollen type has marked localized distribution.
The three rainforest sites, although widely sepa- rated, have similar spectra with no mangrove and negligible saltmarsh and swamp forest representa- tion. The spectra are dominated by terrestrial taxa (85-98%) with the remainder of the count largely composed of swamp taxa, principally Poaceae (2-15%).
Pollen spectra of the Hinchinbrook sites reflect the distance to terrestrial vegetation on either side of the channel. Sites towards the centre of the channel (H5-H11 ), all have mangrove representa

316 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
RAINFOREST SITES P O L L E N
WETLAND TERRESTRIAL
Rain forest
~ ~ -~.
I i one d iv is ion = 10%
Dry habitat,'
R2" + R3"
[ J I I I [ I I I I I I I I I I I i I i i I I •
o
Ungrouped P T E R I D O P H Y T E S
TERRESTRIAL I
U N I D E N T I F I E D TOTALS ~ I
m "~-. Q
"I I I
, , i J l I ~ i I i l i i
R2 R2 R3 + R3
L I I I I I L------ .d I I I t
c t~ "o
~ ~ a= ~
i i
8
r ~
o
3.
t i o n e d iv is ion = 10%
J f I
t I I 1
2 o
3 8
Fig. 12. Percentage pollen data for lowland rainforest samples.
tion over 70% and terrestrial representation of less than 25%. On the western shore, terrestrial repre- sentation generally increases landward to 73% in
HI, this component being mainly composed of Casuarinaceae and Eucalyptus/Melaleuca. The freshwater wetland component also increases mar-

G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327 317
MUTCHERO INLET
WYVURISWAMP
100% H1
100%
H3
149
2 r l ' l l l l l l l l l l l l "
HINCHINBROOK CHANNEL
f / J f J / J / / f f J J A
RAINFOREST TREES lOO% AND SHRUBS
~ High altitude, high rainfall taxa
~ High rainfall taxa
B Other taxa
OTHER RAINFOREST TAXA ~ Epiphytas,
herni-epiphytes and lianes
~ Rainforest edge taxa
RAINFOREST SITES 100%
Fig. 13. Wetland/terrestrial pollen ratios for all sites.
ginally (up to 10% in Hl l ) , with increases in Poaceae being largely responsible for this rise. On the eastern shore, the pattern is more complex with spectra reflecting the mosaic of vegetation types. Samples H 16-H 18, have high swamp-forest values dominated by Pandanus from nearby sources. Sites H14 and H15 situated in upstream mangrove have high terrestrial and mangrove values. Sites H12 and H13 situated in mangrove/ riparian scrub have high terrestrial values, and, while mangroves are well represented in H12, they are less well represented in H13.
At Cleveland Bay mangrove representation is highest in the centre of the Rhizophora forest zone (98%), decreasing gradually with distance land- ward and seaward. Lowest mangrove values (below 40%) are found at the seaward extremity of the intertidal mudflats. However, this gradual change is not compensated for by a consistent increase in any one component of the pollen sum. The pollen spectra vary gradually with distance
from the central mangrove zone, and can be described in five separate groups. C1-C3 have mangrove values of 53-71%, freshwater swamp of 10-14% and terrestrial of 11-14%. C4-C7 have mangrove values of 78-88%, freshwater swamp of 2-8% and terrestrial of 6-13%. C8 has mangrove values of 45%, freshwater swamp of 18% and terrestrial of 35%. C9 and C10 have mangrove values of 60 and 61%, freshwater swamp of 79 and 22% and terrestrial of 13 and 20%, respectively. Cll-C13 have mangrove values of 15 and 38%, freshwater swamp of 32 and 39% and terrestrial of 25 to 39%.
5. 7 Terrestrial components
Rainforest and allied taxa represent 86-92% of the terrestrial pollen at the three rainforest sites (Fig. 14). At most other sites, this component varies from 0% to about 15%, and up to 61% at Mutchero Inlet. Eucalyptus/Melaleuca accounts

318 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299 327
M10' Ml l
MUTCHERO INLET 100% 0
H1 H2 H3 H4 H5 H6 H7 H8
WYVURI S W A M P 0 100% !
W . ] ' ' ' j
;J w, t | o,i W5 ~ C2 W6 ~ C3
' " ' C4' 0 RAINFOREST SITES 100% C5' R~j ' , - - - c61
R2 C7 R3 ~ C8
• " C 9
C10 Cl1' C12' C13"
H I N C H I N E R O O K C H A N N E L 100% POLLEN
, , i ' •1~ ~ ] Terrestrial T a x a
• PTERIDOPHYTES I I I Unnnlofo ~n~r=¢
I
I
C L E V E L A N D BAY 100%
I
Fig. 14. A summary of terrestrial components in the pollen spectra for all sites.
for 3-8% in the rainforest sites, while at other sites this component accounts for between 25-96%. Casuarinaceae varies from 0 to 2% in the rainforest sites and 0 to 33% in all other sites except W2 and fifteen of the eighteen Hinchinbrook Channel sites. At no Hinchinbrook Channel site does Casuarinaceae fall below 17%. In W2 and ten Hinchinbrook Channel sites Casuarinaecae exceeds Eucalyptus/Melaleuca in value. The reverse is true in all other surface samples. At all sites, other dry taxa comprise less than 12% of the terrestrial count.
5.8 Rain forest components
Fig. 15 provides a breakdown of components in rainforest pollen spectra. The three localities at which high-altitude rainforest taxa are represented, Mutchero Inlet, Wyvuri Swamp and Hinchinbrook
Channel, are all fed by streams originating at high elevations. Epiphytic angiosperm taxa are totally absent from the Cleveland Bay samples, but pre- sent at all other sites. With these exceptions, no single rainforest component predominates consis- tently within any site or rainfall zone.
5. 9 Terrestrial pollen/ptertidophyte spore ratios
The terrestrial pollen component of the ter- restrial pollen/pteridophyte spore composite sum exceeds 70% in all samples from Cleveland Bay, Wyvuri Swamp and the rainforest sites (Fig. 16). This is also the case for Hinchinbrook Channel in all but four samples (H12-H15), taken from stream-side mangrove and riparian scrub. At Mutchero Inlet pteridophyte values are generally much stronger, reflecting the occurrence of Drynaria and Platycerium in the local vegetation,

0
Mt M2 M3 M4 M5 M6
M7 M8 M9
MIO M l l
M12 M13 M14 M15
G M. Crowley et aL/Review of Palaeobotany and Palynology 83 (1994) 299-327
MUTCHERO INLET HINCHINBROOK CHANNEL 100% 0 100%
H7
I Other dry habitat I I taxa
F ~ Eucalyptus/ Melaleuca type
319
WYVURI SWAMP CLEVELAND BAY 0 I O0% 0 100%
w l ; , - . - . . . . . i
w 4 / I w l i I I ws "!
RAINFOREST SITES 0 100%
R1 R2 R3
1 ; ; .* ! ! ,~ I I I I I I
Fig. 15. A summary of rainforest components in the pollen spectra for all sites (excluding Cleveland Bay).
and fiver-borne spores (Cyathaceae) from upland catchments.
5.10 Identification of local habitats
The different local environments have strong pollen signatures. Identification of mangrove habi- tat by the domination by Rhizophoraceae with total mangrove values exceeding 50% is consistent with observations made elsewhere (Grindrod and Rhodes, 1984; Chappell and Grindrod, 1985; Grindrod, 1985, 1988a; Clark and Guppy, 1988) as is the indication of presence and indeed domi- nance of Sonneratia spp. by Sonneratia values as low as 3% (Grindrod, 1988a). While occurrence of the Rhizophoraceae and Sonneratia spp. could be
identified in the pollen spectra, all other mangrove taxa show no close relationship between vegetation and pollen occurrence. This contrasts with earlier results from Princess Charlotte Bay (Grindrod, 1985) where high local values were recorded for Ceriops/Bruguiera and Avicennia pollen types. Values of Chenopodicaeae exceeding trace levels indicate presence of chenopods in the local or extra-local vegetation, but absence could not be assumed from lower values.
Vegetation in the mangrove/terrestrial transition zone and in freshwater swamp forest samples tend to be dominated by Pandanus and/or Eucalyptus/ Melaleuca derived from swamp Melaleuca spp. A small proportion of the Pandanus pollen rain appears to be widely dispersed, but most is locally

320 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299 327
MUTCHERO INLET 0 100%
~;.;-;-; . ; . ; . ; . ; . ; , m ~ : . : . : . : . : . : . : . : . : . : m f~-~,-o-o-,t-,t-~,-t+t+t÷o-t+(, . . . . . . . . . . . . . . . m . ~ : , ; . : . : . : . ; . ; . : •
, . . . . . . . . . + . + . . . . . . . . . + . . . , m ) J l ' , "I.16+I+i' @ l
HINCHINBROOK CHANNEL
H2 m ~ H4 t ~
H5" H6 H7
H10 Hll H12 H13 H14 H15 H16
100% WETLAND TAXA
~ Mangrove
m Swampfore=
TERRESTRIALTAXA
[ ~ ] T o t a l
o w l ) W2 W3 W4 W5 W6
o
R2
.311
WYVURISWAMP
RAINFOREST SITES
100% 0 C1" C2" C3" CA" C5. C6 C7"
I 0 0%
I ] I I
CLEVELAND BAY 100%
Fig. 16. A summary representation of terrestrial pollen and pteridophyte spores at all sites.
distributed and where present in the vegetation Pandanus often dominates pollen assemblages. However, this patchy distribution indicates that low values should not be interpreted as the absence of this taxon Sites in which Restionaceae and Cyperaceae are prominent in the vegetation are clearly identified by the domination of the pollen spectra by these taxa. Domination of spectra by Eucalyptus/Melaleuca, with or without Poaceae, also occurs away from swampy habitats. The ter- restrial sample W1 is devoid of other wetland taxa, while the offshore samples from Cleveland Bay have a sprinkling of taxa from all habitat types Clearly Eucalyptus and Poaceae are among the more widespread taxa in Australian vegetation and little detailed information can be gleaned from their occurrence in the pollen spectra, in terms of local habitat identification.
6. Identification of climatic zones
There is a clear relationship between rainfall zone and taxonomic diversity of the regional vege- tation component as represented by the terrestrial pollen counts. There is a general increase in the number of taxa as rainfall increases, with specific dry vegetation indicators having similarly low values across the three rainfall zones. Low-rainfall samples are devoid of epiphytic angiosperms and the only possible rainforest taxa present in them might also have been derived from other habitats. Wetland taxa shed little light on regional condi- tions. Exceptions are the occurrence of higher than trace chenopod values in some low-rainfall zone samples, the presence of Ilex in samples from the high-rainfall zone only, although this latter taxon was not recorded in the local vegetation at any

G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327 321
site. Other recognised high-rainfall taxa found at more than one site in the high-rainfall zone and absent from the other zones, are Cunoniaceae, Euodia/Melicope and Proteaceae/Icacinaceae. Otherwise the number of taxa within any habitat grouping is more important in identifying climatic zones, than the presence or absence of any one taxon.
Other potential climatic indicators may in fact be more indicative of the nature of pollen trans- port, with pteridophyte spores being more abun- dant where there is opportunity for fluvial pollen transport. Angiosperm and pteridophyte taxa from high altitudes show signs of being water- transported and should not be considered indica- tive of cold climates where there has been the potential for such transport.
Acknowledgements
This study was supported by an ARC National Research Fellowship to John Grindrod and Gay Crowley. Some additional funding was provided by an ARC Grant to Peter Kershaw and a Marine Science and Technology grant to David Johnson, James Cook University. Permission to conduct fieldwork and collect samples in national parks at Hinchinbrook Channel and Mutchero Inlet was provided by the Queensland National Parks and Wildlife Service. The authors wish to thank David Johnson and Bob Carter for ongoing encourage- ment and direction, Garry Swinton and Phil Scamp for drafting the figures, Liz Anderson for preparing the manuscript, Stephen Garnett, Peter Walbran and Mike Townsend for field assistance, Merna McKenzie for assistance with sample preparation and Len Webb, Jeff Tracey and Betsy Jackes for plant identifications. Jeff Tracey kindly provided lowland rainforest site vegetation descriptions.
Appendix 1. Plant lists for rainforest sites (emergent, canopy and sub-canopy species, with sites of occurrence)
Alangiaceae (Alangium villosum subsp, polyosmoides: R2), Annonaceae (Polyalthia michaelii: R2; P. sp. 126149: R3), Apocynaceae (Alstonia muellerana R3; A. scholaris: RI, R2; Cerbera floribunda: R 1 ), Araliaceae (Delarbrea michieana: R3; Polyscias australiana: R1, R3; P. elegans: R1, R2, R3; P. mollis:
R2; P. murrayi: R2), Arecaceae (Archontophoenix alexandrae: R2, R3; Licuala ramsayi: R3; Linospadix palmeranus: R1, R2; Ptychosperma elegans: R3), Caesalpiniaceae (Pilidiostigma trop- icum: RI, R2, R3), Celastraceae (Siphonodon membranaceus: R1, R2), Clusiaceae (Garcinia warrenii: R1), Cunoniaceae (Davidsonia pruriens: R2, R3; Gillbeea adenopetala: R2), Dichapetalaceae (Dichapetalun papuanum: R2), Ebenaceae (Diospyros cupulosa: R1, R3; D. sp. 176907: R2), Elaeocarpaceae (Elaeocarpus grandis: R1, R2; E. sp. 162569: R3; Sloanea langii: R1, R3; S. macbrydei: R1, R2, R3), Erythroxylaceae (Erythroxylum ecarinatum: R2, R3), Euphorbiaceae (Antidesma erostre: R3; Claoxylon angustifol- ium: R1; Glochidion ferdinandi var. ferdinandi: R2; G. harveya- hum: R1; Macaranga subdentata: R3: Mallotus polyadenos R3; Rockinghamia angustifolia: R2), Fabaceae (Castanospermum australe R1, R2), Hamamelidaceae (Ostrearia australiana: R2), Icacinaceae (Apodytes brachystylis: R1, R2; Citronella smythii: R1, R2, R3), Lauraceae (Beilschmieda bancroftii: R2, R3; B. sp. 138203: R2; B. sp. B.P. Hyland 288: R1; Cinnamomum laubatii: R2; Cryptocarya angulata: R2; C. corrugata: R1; C. hypoglauca: R3; C. mackinnoniana: R1, R2, R3; C. oblata: R2, R3; C. pleurosperma: RI, R2, R3; C. sp. 152693: R3; C. sp. 160374: R1, R2; Endiandra cowleyana: R1, R3; E. hypotephra: R1, R3; E. montana: R2, R3; E. pubens: R1, R2; E. sankeyana: R1, R2, R3; E. looram: R1, R3; Litsea leefeana: R3; L. reticulala: R2; N. dealbala: R 1, R2), Lecythidaceae (Barringtonia racemosa: R1, R2), Leeaceae (Leea indica: R1), Meliaceae (Aglaia ferruginea: R2; Chisocheton longistipitatus: R1; Dysoxylum arborescens: RI, R2; D. klanderi: R3; D. micranthum: R2; D. oppositifolium: R1, R2, R3; D. pettigrewia- num: R1, R2; D. schiffneri: R1; D. sp. 160385: R1), Mimosaceae (Archidendron sp. 189472: R1, R2), Monimiaceae (Daphnandra repandula: R2; Doryphora aromatica: R1, R2; Levieria acumi- nata: R2; Tetrasynandra lariflora: R2; 72 pubescens: R1, R2, R3), Moraceae (Ficus copiosa: R2; F. destruens: R2; F fraseri: R2; F pleurocarpa: RI, R2; F racemosa: R2, R3; F septica: R1, R2; F seplica var. cauliflora: R3; F virgata: R2, R3), Myristicaceae (Myristica muelleri: R1, R2; Ardisia bifaria: R2), Myrtaceae (Acmena divaricata: R1; A. graveolens R1, R2, R3; Austromyrtus minutiflora: R3; Backhousia bancroftii R1; Decaspermun fruticosum: R3; Eugenia corrniflora: R1, R2, R3; Eugenia erythrodoxa: R1; E. kuranda: R3; E. luehmannii: R3; E. tiernyana: R3; E. wilsonii: R3; E. sp. 126045: R2, R3; Rhodamnia sessiliflora: R3; Rhodomyrtus macrocarpa: R1; Syzygium dictyophlevium: R 1 ), Nyctaginaceae (Pisonia umbellif- era: R 1 ), Ochnaceae (Brackenridgea australiana: R3), Pittosporaceae (Pittosporum rubiginosum: R1, R3), Podocarpaceae (Podocarpus dispermus: R1, R3), Proteaceae (Cardwellia sublimis: R3; Carnarvonia araliifolia: R2, R3; Helicia nortoniana: R1, R2, R3; Hollandaea sayeriana: R2; Macadamia whelanii: R1; Musgravea stenostachya: R3; Opistiolepis hetero- phylla: R2), Rhamnaceae (Schistocarpaea johnsonii: R3), Rhizophoraceae (Carallia brachiata: R1, R3), Rosaceae (Prunus turneriana: R1, R2, R3), Rubiaceae (Antirhea tenuiflora: R1, R2; Canthium sp. 1 04951: R 1; Gardenia merikin: R2; Psychotria sp. 241599: R1, R2; R. hirta: R3), Rutaceae (Acronychia vestita: R1, R2; Brombya platynema R2, R3; Euodia bonwickff: R2; E.

322 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299 327
haplophylla: R1, R2; E. xanthoxyloides: R2; Flindershl acumi- nata R1, R2; F bourjotiana: R1, R3; F pimenteliana: R1; Zanthoxylum brachyaeanthum: R1, R2, R3), Sapindaceae (Arvtera divaricata: R1; Castanospora alphandi: R2; Diplogottis subglabra: R1, R2; Ganophylum faleatum: R1; Guioa acutifolia: R3; G.lasinoneura: R1, R2, R3; Harpullia fruteseens. R1, R2; Mischocarpus exangulatus: R3; M. lachnocarpus: R3; Rhysotoechia robertsonii: R1; Sapindaceae 169299: R2; Sarcoptervx martyana: R2, R3; Sarcotoechia protracta: R2; Synima cordieri: R1, R2, R3; Toechima erythrocarpum: R1, R2), Sapotaceae (Chrysophyllum chartaceum: R1, R2; Palaquium galatoxylum: R1, R3; Planchonella chartacea: R1,
R3; P. obovoidea R1 ), Saxifragaceae (Polyosma alangh~cea: R2, R3), Sterculiaceae (Argyrodendron peralatum: R1, R2; Commersonia bartramia: R1; Sterculia laurifolia: R3), Symplocaceae (Symplocos cochinchinensis subsp, thwaitesii: R3; S. paucistaminea: R2, R3), Theaceae (Ternstroemia cherryi: R2), Thymelaeaceae (Phaleria clerodendron: R2), Ulmaceae ( Trema amboinensis: R2), Urticaceae (Dendrocnide cordata: R2; D. moroides: R1 ), Verbenaceae (Clerodendrum traceyanum: R 1, R2; Gmelina fasciculiflora: R1, R3; Vitex acuminata: R1), Winteraceae (Bubbia semecarpoides: R2), Xanthophyllaceae (Xanthophyllum octandrum: R1, R2, R3), Zamiaceae (Lepidozamia hopei: R3).
Appendix 2 Pollen and spore taxa and their likely source plants
Taxon (family) Likely constituent taxa Sites"
(A) POLLEN TAXA
(1) Wetland taxa
1.1 Mangrove taxa Aegiceras (Myrsinaceae) Avieennia ( Verbenaceae) Bruguiera/Ceriops (Rhizophoraceae) Excoecaria (Euphorbiaceae) Lumnitzera (Combretaceae) Osbornia (Myrtaceae) Rhizophora (Rhizophoraceae) Schyphiphora (Rubiaceae) Sonneratia alba (Sonnetatiaceae) S. caseolaris (Sonnetatiaceae) Xylocarpus (Meliaceae)
1.2 Saltmarsh taxa Chenopodiaceae Portulacaceae
1.3 Freshwater wetland taxa Aesehynomone type (Fabaceae)
Brasenia (Cambombaceae) Cladium type (Cyperaceae) Cyperaceae
Myriophyllum (Haloragaceae) Poaceae Restionaceae Sesbania (Fabaceae) Triglochin (Juncaginaceae) Typha (Typhaceae)
1.4 Swamp Jbrest taxa Barringtonia (Lecythidiaceae) llex (Aquifoliaceae) Pandanus (Pandanaceae)
A. corniculatum A. marina, A. officinalis Bruguiera, Ceriops E. agallocha Lumnitzera O. octodonta Rhizophora S. hydrophyllaeea S. alba S. caseolaris ~ X. granatum, X. australasieum
Chenopodium, Einadia, Halosarc&, Tecticornia, Sueda Portulacaceae
Pollen of this type is consistent with Aeschynomone and Pongamia, but may include other Fabaceae
B. schreberi Clad&m, and other Cyperaceae Cyperaceae, principally Gahnia, Lepironia, Schoenus,
Scleria, Rhynchospora Myriophyllum spp. Poaceae (also includes some dryland taxa) Restionaceae, principally Leptocarpus Sesbania Triglochin T. oriental&
Barringtonia I. arnhemensis, L sp. Pandanus
m w
m H C M w H C m h c M H C
h M w H C
h H
M h M w h c
m w r
m w
W M W r
M W R M W
W
m m w
w
M W r M W r
h C h
H C
h H C H C
C h
H C

G M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327 323
Appendix 2 (continued)
Taxon (family) Likely constituent taxa Sites a
(2) Terrestrial pollen taxa 2.1 Rain forest and allied taxa 2.1.1 High-rainfall, high-altitude (> 500 m) rain forest trees and shrubs Balanops (Balanopaceae) Cunoniaceae-dicolpate Cunoniaceae-tricolpate
Orania (Arecaceae) Quint inia (Saxifragaceae)
2.1.2 Other high-rainfall, rain forest trees and shrubs Arecaceae Elaeocarpaceae Euodia/Melicope (Rutaceae) Ophiorrhiza type (Rubiaceae) Proteaceae/Icacinaceae Prunus turnerana (Rosaceae) Symploeos (Symplocaceae) Xanthophyllum (Xanthophyllaceae)
2.1.3 Other rainforest trees and shrubs Alangium (Alangiaceae) Argyrodendron (Sterculiaceae) Balanophora (Balanophoraceae) Calamus (Arecaceae) Emmenosperma (Rhamnaceae) Eugenia type (Myrtaceae) Ganophyllum (Burseraceae) Homolanthus type (Euphorbiaceae) Linociera comp. (Oleaceae) Macaranga/Mallotus (Euphorbiaceae) Meliaceae Myristica (Myristicaceae) Myrsinaceae Nothofagus (Fagaceae) Polyscias (Alaliaceae) Vroteaceae
Randia (Rubiaceae) Rutaceae Sapindaceae-syncolpate Sapotaceae Seolopia comp. (Flacoutiaceae) Urticaceae/Moraceae
2.1.4 Lianes, epiphytes and hemi-epihytes Apocynaceae Cassia type (Caesalpiniaceae) Entada (Mimosaceae) Faradaya (Verbenaceae) Flagellaria (Flagellariaceae) Fr eycinetia (Pandanaceae) Loranthaceae Menispermaceae Scheffiera (Araliaceae)
B. australiana Ceratopetalum, Schizomeria Ackama, Caldcluvia, (Davidsonia?) Geissois,
Pseudoweinmannia, Pullea, Spiraeantheum O. appendiculata Quintinia
Arecaceae, excluding Calamus, Linospadix, Orania Elaeocarpaceae Euodia, Melicope Ophiorrhiza, but may include Cassine (Celastraceae) Apodytes, Irvingbaileya, Opisthiolepis Prunus turnerana Symplocos X. octandrum
A. villosum A rgyrodendron B. fungosa Calamus E. alphitonioides Eugenia, Lophostemon, Tristania G. faleatum Homolanthus, Euphorbia Linoeiera Maearanga, Mallotus Meliaceae, principally Dysoxylon, Toona M. muelleri Myrsinaceae, particularly Myrsine Genus not recorded in area Polyscias Proteaceae (excluding Banksia, Grevillea, Musgravea,
Opisthiolepis, Persoonia) Randia (particlarly P~ fitzalanii) Rutaceae, principally Flindersia Rainforest Sapindaceae Planehonella, Chrysophyllum Scolopia braunii Urticaceae/Moraceae
Alyxia, Parsonsia (Alstonia?) Probably C retusa E. phaseoloides F splendida Flagellaria Freycinetia Loranthaceae Menispermaceae S. actinophylla
h M w M w
h m
M w r h M w R h M w R
r
M r R r
r
r
w R h r
M w R H R R R
W r
r
M w R w R h
r
r h H
r
M w R H
M M W R m w R h m w r h
R m r
M M
R h
m w R h m w h m w r
w r
M r

324
Appendix 2 (continued)
Taxon (family)
G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
Likely constituent taxa Sites a
2.1.5 Rainforest edge taxa Acacia aulacocarpa (Mimosaceae) A lphitonia (Rhamnaceae) Celtis ( Ulmaceae) Glochidion (Euphorbiaceae) Lantana (Verbenaceae) Maesa (Myrsinaceae) Melastomataceae Trema ( Ulmaceae)
A. aulacocarpa Alphitonia Celtis Glochidion Z. camara c Maesa Melastomataceae Trema
2.2 Dry habitat taxa (including open forest, woodland, beach strand and heath taxa) Alectryon type (Sapindaceae) Argusia type(Boraginaceae) Asteraceae-liguliflorae Asteraceae-tubuliflorae Banksia (Proteaceae) Breynia (Euphorbiaceae) Casuarinaceae Choriceras ( Euphorbiaceae) Dodonaea (Sapindaceae) Exocarpus (Santalaceae) Halor agis (Haloragaceae) Leptospermum (Myrtaceae) Petalostigma ( Euphorbiaceae) Triumfetta (Tiliaceae)
2.3 Ungrouped taxa 2.3.1 Conifers mraucariaceae Callitris (Cupressaceae) Pinus (Pinaceae)/Podocarpus (Podocarpaceae)
2.3.2 Angiosperms Acacia (Mimosaceae) Acronychia (Rutaceae) Clematis comp. (Ranunculaceae) Dianella type (Liliaceae) Epacridaceae Eucalyptus/ Melaleuca (Myrtaceae) Fabaceae Gardenia (Rubiaceae) Grevillea (Proteaceae) Lamiaceae Liliaceae/Arecaceae Lomandra (Xanthorrhoeaceae) Muehlenbeckia (Polygonaceae) Plantago (Plantaginaceae) Rubiaceae Solanum (Solanaceae)
(3) Unidentified taxa Unidentified
Alectryon, possibly other Sapindaceae Argusia, possibly also Terminalia (Combretaceae) Asteraceae-liguliflorae Asteraceae-tubuliflorae B. integrifolia, B. plagiocarpa Breynia Allocasuarina, Casuarina Choriceras Dodonaea E. latifolius Haloragis, Gonoearpus Leptospermum Petalostigma Triumf etta
Agathis, Araucaria Callitris Pinus c, Podocarpus
Acacia (excluding A. aulacocarpa) Aeronychia Unknown, this family not recorded in the area Dianella Dracophyllum, Monotoea Eucalyptus, Melaleuca (also includes some wetland taxa) Fabaceae Gardenia Grevillea Lamiaceae Liliaceae/Arecaceae Lomandra Muehlenbeckia Plantago Rubiaceae, possibly Timonius Solanum
Unknown
R w r
m w r r R R r
m w R c
m m w C
R M w h C
H w h
m W R H C m h M
r m w r H
c m h
h
m r M w h M w H
m w H R h R
w m M W R H C M W h c m r m
w r M w h
r R
C M w h
R
M W R H C

G.M. Crowley et al,/Review of Palaeobotany and Palynology 83 (1994) 299-327 325
Appendix 2 (continued)
(B) PTERIDOPHYTES
(4) Wetland pteridophyte taxa Taxon (family) Likely constituent taxa Sites a
4.1 Mangrove ferns Acrostichum (Pteridaceae)
4.2 Freshwater wetland ferns Ceratopteris (Parkeriaceae) Cyclosorus ( Thelypteridaceae)
A. speciosum, A. areum (trilete)
C. thalictroides (trilete) Cyclosorus (trilete)
M
C h
(5) Terrestrial pteridophyte taxa
5.1 High-altitude ferns Cyathea robertsiana (Cyatheaceae)
5.2 Epiphytic and hemi-epiphytic ferns Asplenium (Aspleniaceae) Colysis (Polypodiaceae) Davallia (Davalliaceae) Drynaria (Polypodiaceae) Drynaria/Platycerium (Polypodiaceae) Platycerium (Polypodiaceae) Microsorium (Polypodiaceae) Polypodiaceae Stenochlaena (Blechnaceae)
C. robertsiana (trilete)
Asplenium (monolete) C. ampla, C. sayeri (monolete) Davallia (monolete) Drynaria (monolete) Drynaria/Platycerium Platycerium (monolete) Microsorium (monolete) Polypodiaceae S. palustris (monolete)
M w
m W
M W
M w
M w
R R
H R
h R
5.3 Ground-dwelling pteridophytes Blechnum (Blechnaceae) Botryehium (Ophioglossaceae) Cyatheaceae comp.
Dennstaedtia (Dennstaedtiaceae) Gleichenia (Gleicheniaceae) L ycopodium (Lycopodiaceae) Marattia (Marattiaceae) Nephr olepis (Oleandraceae) Pellaea (Sinopteridaceae) Selaginella (Selaginellaceae)
5.4 Ungrouped terrestrial ferns Schizaceae (trilete)
(6) Unidentified taxa MI (monolete, cf. Lastreopsis) M2 (monolete, verrucate) M3 (monolete, microrugulate) M4 (monolete, psilate) Monolete, other Trilete, other Eroded
Blechnum (monolete) B. australe (trilete) Mostly Cyatheaceae, excludes Cyathea robertsoniana (trilete) D. davallioides (monolete) Gleichenia (trilete) L ycopodium (trilete) 3/1. salicina (monolete) Nephrolepis (monolete) Pellaea (trilete) Selaginella (trilete)
Lygodium, Schizaea (trilete)
Unknown Unknown Unknown Unknown Unknown Unknown Unknown
M w
r
r
R H C
r
h M r M w r
R R
m
M W r h c
M M M
m w h M w h m w
Taxa are listed according to habitat and habit as determined from the literature (Kershaw, 1973; Jones and Clemesha, 1976; Aston, 1977; Lear and Turner, 1977; Williams, 1980, 1984; Airy Shaw, 1981; Francis, 1981; Flora of Australia, 1982, 1984a,b, 1985, 1986, 1987, 1989, 1990; Hyland, 1982; Tracey, 1982; Morley and Toelken, 1983; Cribb and Cribb, 1985). aM/m=Mutchero Inlet; W/w=Wyvuri Swamp; R/r=rainforest sites; H/h=Hinchinbrook Channel; C/c=Cleveland Bay. Upper case indicates major taxa (representation > 1%); lower case indicates minor taxa (representation < 1% i.e. trace levels). bs. lanceolata in Crowley et al. (1990). Clntroduced taxa.

326 G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327
Appendix 3 Range of mangrove values found in wetland vegetation in northeast Queensland
Vegetation type Values (% of total pollen)
Rhizophora Bruguiera/Ceriops Other mangroves Rhizophora + Total mangrove Bruguiera/ Ceriops
Min Max Min Max Min Max Min Max Min Max
Cleveland (Low rainfall ) Avicennia mangrove 24 53 25 27 3 8 49 78 53 81 Rhizophora mangrove 35 71 9 20 0 6 44 82 45 88 Avicennia mangrove a 44 12 3 56 59 Low-tide mudflat 4 29 7 12 0 8 16 35 16 37
Hinchinhrook (Medium rainfall) Avicennia, Sonneratia man- 48 61 13 23 5 7 70 74 75 80 grove (no Rhizophora) Rhizophora mangrove 70 81 6 11 0 1 82 87 83 87 Mangrove (Rhizophora 22 44 15 28 < 1 5 41 72 46 73
minor or absent) Mangrove/swamp forest 0 24 1 26 0 2 1 33 1 35 Swamp forest a 10 2 < 1 12 12 Mid-hannel (upstream)" 14 67 1 81 82 Mid-channel (downstream)" 71 12 < 1 83 83
Mutehero/Wyvuri (High rainfall) Rhizophora mangrove 36 52 15 25 0 4 58 73 62 74 Mangrove (Rhizophora 22 52 23 37 0 5 58 75 58 75
minor or absent) Mangrove/swamp forest 3 13 2 5 0 13 7 18 10 31 Swamp forest 0 5 0 6 0 1 0 11 0 11 Swamp < 1 < 1 < 1 < 1 < 1
aOne sample only
References
Airy Shaw, H.K., 1971. A partial synopsis of Euphorbiaceae-Platylobae of Australia (excluding Phyllanthus, Euphorbia and Calyeopeplus). Kew Bull., 35: 577-700.
Aston, H.I., 1977. Aquatic Plants of Australia. Melbourne Univ. Press, Melbourne, 368 pp.
Bunt, J.S., Williams, W.T. and Duke N.C., 1982. Mangrove distribution in northeastern Australia. J. Biogeogr., 9: 111 120.
Chappell, J. and Grindrod J., 1985. Pollen analysis: A key to past mangrove communities and successional changes in North Australian coastal environments. In: K.N. Bardsley, J.D.S. Davie and C.D. Woodroffe (Editors), Coasts and Tidal Wetlands of the Australian Monsoon Region. Aust. Natl. Univ. North Aust. Res. Unit, Darwin, pp. 225-36.
Clark, R.L. and Guppy, J.C., 1988. A transition from mangrove forest to freshwater wetland in the monsoon tropics of Australia. J. Biogeogr., 15: 665-84.
Cribb, A.B. and Cribb, J.W., 1985. Plant Life of the Great
Barrier Reef and Adjacent Shores. Univ. Queensl. Press, St. Lucia, 294 pp.
F~egri, K. and Iversen, J., 1975. Textbook of Pollen Analysis. Blackwell, Oxford, 3rd ed., 237 pp.
Flora of Australia, 1982. Vol. 8: Lecythidales to Batales. Aust. Gov. Print. Serv., Canberra, 420 pp.
Flora of Australia, 1984a. Vol. 4: Phytolaccaceae to Chenopodiaceae. Aust. Gov. Print. Serv., Canberra, 354 pp.
Flora of Australia, 1984b. Vol. 22: Rhizophorales to Celastrales. Aust. Gov. Print. Serv., Canberra, 239 pp.
Flora of Australia, 1985. Vol. 25: Melianthaceae to Simaroubaceae Aust. Gov. Print. Serv., Canberra, 215 pp.
Flora of Australia, 1986. Vol. 46: Iridaceae to Dioscoreaceae. Aust. Gov. Print. Serv., Canberra, 247 pp.
Flora of Australia, 1987. Vol. 45: Hydatellaceae to Liliaceae. Aust. Gov. Print. Serv., Canberra, 521 pp.
Flora of Australia, 1989. Vol. 3: Hamamelidales to Casuarinales. Aust. Gov. Print. Serv., Canberra, 219 pp.
Flora of Australia, 1990. Vol. 18: Podostemaceae to Combretaceae. Aust. Gov. Print. Serv., Canberra, 349 pp.
Francis, W.D., 1981. Australian Rainforest Trees. Aust Gov. Publ. Serv., 468 pp.

G.M. Crowley et al./Review of Palaeobotany and Palynology 83 (1994) 299-327 327
Grindrod, J., 1985. The palynology of mangroves on a prograded shore, Princess Charlotte Bay, North Queensland, Australia. J. Biogeogr., 12: 32348.
Grindrod, J., 1988a. Holocene Mangrove History of the South Alligator River Estuary, Northern Territory, Australia. Ph.D Thesis. Aust. Nat. Univ., Canberra (unpubl).
Grindrod, J., 1988b. The palynology of Holocene mangrove and saltmarsh sediments, particularly in Northern Australia. Rev. Palaeobot. Palynol., 55: 229-245.
Grindrod, J. and Rhodes, E.G., 1984. Holocene sea-level history of a tropical estuary: Missionary Bay, North Queensland. In: B.G. Thorn (Editor), Coastal Geomorphology in Australia. Academic, New York, NY, pp. 151-178.
Hyland, B.P.M., 1982. A Revised Card Key to Rainforest Trees of North Queensland. CSIRO, Melbourne, 103 pp.
Jones, D.L. and Clemesha, S.C., 1976. Australian Ferns and Fern Allies Currawong Press, Frenchs Forest, 232 pp.
Kershaw, A.P., 1973. Late Quaternary Vegetation of the Atherton Tableland, North-East Queensland, Australia. Ph.D thesis. Aust. Nat. Univ., Canberra (unpubl), 439 pp.
Kershaw, A.P. and Hyland, B.P.M., 1975. Pollen transfer and periodicity in a rainforest situation. Rev. Palaeobot. Palynol., 19:129 138.
Kershaw, A.P. and Strickland, K.M., 1990. A ten year pollen trapping record from rainforest in northeastern Queensland, Australia. Rev. Palaeobot. Palynol., 64, 281-288.
Lear, R. and Turner, T., 1977. Mangroves of Australia. Univ. Queensl. Press, St. Lucia, 84 pp.
Morley, B.D. and Toelken, H.B., 1983. Flowering Plants in Australia. Rigby, Adelaide, 416 pp.
Murtha, G.G., 1972. Soils and Land Use on the Southern Section of the Townsville Plain, North Queensland. CSIRO, Melbourne.
Nix, H.A. and Kalma, J.D., 1972. Climate as a dominant control in the biogeography of northern Australia and New Guinea. In: D. Walker (Editor), Bridge and Barrier: the Natural and Cultural History of Torres Strait. Aust. Natl. Univ., Canberra, Bio. Geo. Publ., BG/3, pp. 61-92.
Ricker, W.E., 1979. Growth rates and models. In: W.S. Hoar, D.J. Randall and J.R. Brett (Editors), Fish Physiology, V8. Bioenergetics and Growth. Academic, New York, NY, pp. 677-743.
Steel, R.G.D and Torrie, J.H., 1960. Principles and Procedures in Statistics with Special Reference to the Biological Sciences. MacGraw Hill, New York, NY, 481 pp.
Tracey, J.G., 1982. The Vegetation of the Humid Tropical Region of North Queensland. CSIRO, Melbourne, 124 pp.
Tracey, J.G. and Webb, L.J., 1975. Vegetation of the Humid Tropical Regional of North Queensland, Map Series. CSIRO, Indooroopilly.
Tracey, J.G. and Webb, L.J., 1978. The vegetation of Hinchinbrook Island. Queensl. Nat., 22: 1-4.
Williams, K.A.W., 1980. Native Plants of Queensland, Vol. 1. Williams, North Ipswitch, 288 pp.
Williams, K.A.W., 1984. Native Plants of Queensland, Vol. 2. Williams, North Ipswitch, 316 pp.




















