Microdyromys remmerti, sp. Nov., a new Gliridae (Rodentia, Mammalia) from the Aragonian type area...
17
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [García-Paredes, Israel][Museo Nal Ciencias Naturales] On: 16 September 2010 Access details: Access Details: [subscription number 918400093] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Journal of Vertebrate Paleontology Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t917000010 Microdyromys remmerti, sp. nov., a new Gliridae (Rodentia, Mammalia) from the Aragonian type area (Miocene, Calatayud-Montalbán basin, Spain) ISRAEL GARCÍA-PAREDES ab ; PABLO PELÁEZ-CAMPOMANES a ; M.ÁNGELES ÁLVAREZ-SIERRA c a Museo Nacional de Ciencias Naturales, CSIC, Madrid, Spain b Nationaal Natuurhistorisch Museum Naturalis, RA Leiden, The Netherlands c Departamento de Paleontología, Facultad de Ciencias Geológicas, Universidad Complutense de Madrid, Unidad de Investigación en Paleontología, Instituto de Geología Económica, CSIC, Madrid, Spain Online publication date: 15 September 2010 To cite this Article GARCÍA-PAREDES, ISRAEL , PELÁEZ-CAMPOMANES, PABLO and ÁLVAREZ-SIERRA, M.ÁNGELES(2010) 'Microdyromys remmerti, sp. nov., a new Gliridae (Rodentia, Mammalia) from the Aragonian type area (Miocene, Calatayud-Montalbán basin, Spain)', Journal of Vertebrate Paleontology, 30: 5, 1594 — 1609 To link to this Article: DOI: 10.1080/02724634.2010.501453 URL: http://dx.doi.org/10.1080/02724634.2010.501453 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript of Microdyromys remmerti, sp. Nov., a new Gliridae (Rodentia, Mammalia) from the Aragonian type area...

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [García-Paredes, Israel][Museo Nal Ciencias Naturales]On: 16 September 2010Access details: Access Details: [subscription number 918400093]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Vertebrate PaleontologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t917000010
Microdyromys remmerti, sp. nov., a new Gliridae (Rodentia, Mammalia)from the Aragonian type area (Miocene, Calatayud-Montalbán basin,Spain)ISRAEL GARCÍA-PAREDESab; PABLO PELÁEZ-CAMPOMANESa; M.ÁNGELES ÁLVAREZ-SIERRAc
a Museo Nacional de Ciencias Naturales, CSIC, Madrid, Spain b Nationaal Natuurhistorisch MuseumNaturalis, RA Leiden, The Netherlands c Departamento de Paleontología, Facultad de CienciasGeológicas, Universidad Complutense de Madrid, Unidad de Investigación en Paleontología, Institutode Geología Económica, CSIC, Madrid, Spain
Online publication date: 15 September 2010
To cite this Article GARCÍA-PAREDES, ISRAEL , PELÁEZ-CAMPOMANES, PABLO and ÁLVAREZ-SIERRA,M.ÁNGELES(2010) 'Microdyromys remmerti, sp. nov., a new Gliridae (Rodentia, Mammalia) from the Aragonian typearea (Miocene, Calatayud-Montalbán basin, Spain)', Journal of Vertebrate Paleontology, 30: 5, 1594 — 1609To link to this Article: DOI: 10.1080/02724634.2010.501453URL: http://dx.doi.org/10.1080/02724634.2010.501453
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Journal of Vertebrate Paleontology 30(5):1594–1609, September 2010© 2010 by the Society of Vertebrate Paleontology
ARTICLE
MICRODYROMYS REMMERTI, SP. NOV., A NEW GLIRIDAE (RODENTIA, MAMMALIA) FROMTHE ARAGONIAN TYPE AREA (MIOCENE, CALATAYUD-MONTALBAN BASIN, SPAIN)
ISRAEL GARCIA-PAREDES,*,1,3 PABLO PELAEZ-CAMPOMANES,1 and M. ANGELES ALVAREZ-SIERRA2
1Museo Nacional de Ciencias Naturales, CSIC, C/ Jose Gutierrez Abascal 2, 28006, Madrid, Spain, [email protected];[email protected];
2Departamento de Paleontologıa, Facultad de Ciencias Geologicas, Universidad Complutense de Madrid, Unidad de Investigacionen Paleontologıa, Instituto de Geologıa Economica, CSIC, C/ Jose Antonio Novais 2 28040, Madrid, Spain, [email protected];
3Nationaal Natuurhistorisch Museum Naturalis, P.O. Box 9517 2300, RA Leiden, The Netherlands
ABSTRACT—A new species of Gliridae (Rodentia, Mammalia), Microdyromys remmerti, sp. nov., from the Aragonian typearea in the Calatayud-Montalban basin (Miocene, Spain) is described. This species is characterized by combining a relativelylarge size and a moderately complicated to very complicated and irregular dental morphology. It has a wide and continuousrange of morphological variation with intermediate forms filling all the range. In the studied localities, M. remmerti, sp. nov.,displays a general increase in the complexity of the dental pattern through time, showing a unique pattern of addition of extraridges in the lower molars. Comparisons with morphologically similar species are discussed; a taxonomical and nomenclatorialissue regarding two named species of the genus is clarified. The combination of the morphometrical features that character-ize M. remmerti, sp. nov., have never been found in any other glirid material outside the Calatayud-Montalban basin, thusreinforcing the idea of the endemic character of the Gliridae faunas from the Iberian Miocene. Microdyromys koenigswaldiis proposed as most probable ancestor of M. remmerti, sp. nov. The divergent evolution of the latter species is discussed in itspaleoecological context. The functional consequences of this morphological divergence could be related also with the openingof new ecological niches during the middle Aragonian (Middle Miocene).
INTRODUCTION
The availability of a rich and dense fossil record from a re-stricted area and the accurate age control of the rock successionthat yielded the record provide the framework necessary to re-construct the history and evolution of faunas and ecosystems ingreat detail. The Aragonian type area situated in the Calatayud-Montalban basin (Spain) fulfills these requirements.
After numerous field campaigns that started in the 1950s, theCalatayud-Montalban basin can be considered as having one ofthe richest and better known micromammal fossil records forthe Miocene; it contains several well-exposed sections with su-perposed fossil sites. These sections have been correlated mag-netostratigraphically, allowing the precise assignation of numer-ical ages for the localities. Apart from the classical taxonomi-cal and paleoecological works (see Daams et al., 1999c, for his-torical review of micromammal paleontology in the Calatayud-Montalban basin), the updated and new paleontological informa-tion has allowed detailed studies on long-term evolution of mi-cromammal faunas (e.g., Daams et al., 1999a; Van der Meulenet al., 2005; Van Dam et al., 2006; Van der Meulen and Pelaez-Campomanes, 2007). However, this kind of analysis of the fos-sil record needs systematic and taxonomical reviews of differentgroups (e.g., Pelaez-Campomanes, 2001; Van der Meulen et al.,2003; Garcıa-Paredes, 2006; Lopez Guerrero et al., 2008; OliverPerez et al., 2008; Garcıa-Paredes et al., 2009; Van den Hoek Os-tende et al., 2009) as an integrated research strategy due to thenew available material and information.
One of the groups recently revised is the Gliridae (Rodentia,Mammalia) from the Early and Middle Miocene of the Arag-onian type area (Garcıa-Paredes, 2006). During this time, the
*Corresponding author.
Gliridae became the most important group in European rodentfaunas, reaching its maximum diversity and relative abundance(Daams and De Bruijn, 1995). The evolution towards new den-tal morphologies played an important role in this rise of the di-versity, and it probably represented an increase in niches occu-pied by this family in the Early and Middle Miocene. Among thenew morphotypes are the evolution towards a more simple den-tal morphology that occurred during the Early Miocene, mainlyin the Iberian Peninsula (Garcıa-Paredes et al., 2009), but alsothe evolution towards a more complicated dental pattern. This isthe case of Microdyromys remmerti, sp. nov., from the Spanishmiddle Aragonian (Middle Miocene).
GEOLOGICAL SETTING
The Calatayud-Montalban basin is situated in the northeastsector of Spain (Fig. 1). It is filled by Neogene and Holocene de-posits and it is surrounded by Paleozoic reliefs. The basin con-tains the stratotypes of two continental stages: the Aragonian(Daams et al., 1977) and the Ramblian (Daams et al., 1987).
The material described in this paper belongs to a rich anddense micromammal fossil record from several sites located inthe Aragonian type area (Province of Zaragoza, Spain). It is asmall and restricted area of about 2 km2 (Fig. 1) that allows ac-curate stratigraphical control for more than 100 fossil sites situ-ated in four main sections. The sections have been firmly corre-lated lithologically, biostratigraphically, and magnetostratigraph-ically, and form the framework of the locality succession. For adetailed description of the sections, (bio)stratigraphical and sed-imentological information we refer to Daams et al., 1999c. Thesesections have been correlated to the Geomagnetic Polarity TimeScale (GPTS), allowing the estimation of numerical ages for thelocalities and the determination of sediment accumulation rates
1594
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

GARCIA-PAREDES ET AL.—NEW MICRODYROMYS FROM THE SPANISH MIOCENE 1595
(Krijgsman et al., 1994, 1996; Daams et al., 1999b). The numeri-cal ages of the localities have been recently refined in Van Damet al., 2006, based on calibrations of the new Astronomical TunedNeogene Time Scale (Lourens et al., 2004).
MATERIALS AND METHODS
The fossil assemblages were collected by screen washing thefossiliferous layers throughout the above-mentioned stratigraph-ical sections. The sampling coverage of the fossil record from theAragonian type area has been calculated by Van der Meulen etal. (2005) using the number of upper and lower first and secondmolars of rodents from 96 localities of the Miocene sequence.Several localities from the Daroca area, situated less than 12 kmto the southeast from the Aragonian type area, were includedin the calculations. The cumulative curve of sample sizes of thetemporally ordered localities yields a good fit with the linear re-gression line (R2 = 0.98), indicating that the sections have beensampled quite evenly (Van der Meulen et al., 2005:fig. A1).
FIGURE 1. Location of the Calatayud-Montalban basin in Spain andsynthetic geology of the region. Area for studied localities (Aragoniantype area) is indicated with a square mark. Modified after Alvarez Sierraet al. (2003).
FIGURE 2. Nomenclature of parts of the occlusal surface of the cheekteeth of Gliridae following De Bruijn (1966) and Daams (1981) with someadditions. Modified after Garcıa-Paredes (2006).
Table 1 lists the material studied of Microdyromys remmerti,sp. nov., from localities of the Aragonian type area (Calatayud-Montalban basin), including the chronological framework andcorrelations used in this paper: Series; Continental Stages; Mam-mal Neogene Zones (MN zones) proposed by Mein (1975, 1990)for Europe and revised by De Bruijn et al. (1992); local biostrati-graphical zones after Daams et al. (1999c) and numerical ages ofthe localities after Van Dam et al. (2006); abbreviations for local-ities and institutions where collections are stored (see bellow).
The nomenclature for parts of the occlusal surface of the cheekteeth of Gliridae (Fig. 2) is modified after De Bruijn (1966) andDaams (1981). Notation P/p and M/m is used for upper/lower pre-molars and upper/lower molars, respectively.
The photographs of specimens were made on the Environ-mental Scanning Electron Microscope FEI Quanta 200 in envi-ronmental mode at the Museo Nacional de Ciencias Naturales,CSIC (Madrid, Spain). All the teeth shown in Figures 3–6 areat same magnification and figured as if they are from the leftside.
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

1596 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 30, NO. 5, 2010
TABLE 1. Material of Microdyromys remmerti, sp. nov., from the Aragonian type area (Calatayud-Montalban basin, Spain).
Microdyromys remmerti, sp. nov.,
BiozoneAge(Ma)
Localities andabbreviations Collections P4 M1 M2 M3 p4 m1 m2 m3 Total
Dd 14.50 Valdemoros 3F VA3F MNCN 3 4 3 1 2 3 2 3 2114.53 Valdemoros 3E VA3E RGM — 2 2 5 — 1 7 2 1914.53 Valdemoros 6A VA6A MNCN — — 2 — — 1 — — 314.55 Valdemoros 7C VA7C MNCN — — — — 1 3 2 — 614.61 Valdemoros 1A VA1A MNCN — — — — — 1 — — 114.71 Vargas 8C VR8C MNCN 3 6 14 4 10 11 14 7 6914.73 Vargas 8B VR8B MNCN 6 13 17 8 9 14 11 8 8614.81 VARGAS 7 VR7 MNCN 18 55 50 33 25 56 33 38 308
Dc 14.82 Valdemoros 3D VA3D RGM — 1 2 — — 1 2 2 814.84 Valdemoros 3B VA3B IAUU;
RGM— — — 2 1 — — 1 4
15.49 Villafeliche 4B VL4B MNCN;IAUU
4 4 5 6 5 6 6 1 37
15.50 Villafeliche 4A VL4A MNCN;RGM;IAUU
15 25 24 9 11 31 23 10 148
Total 49 110 119 68 64 128 100 72 710
Chronological framework and correlations used in this paper: Miocene (Middle) Series Epoch; Aragonian (Middle) Continental Stage; MammalNeogene Zones (MN 5) proposed by Mein (1975, 1990) for Europe and revised by De Bruijn et al. (1992); local biostratigraphical zones (biozone)after Daams et al. (1999a) and numerical ages of the localities after Van Dam et al. (2006); localities, abbreviations for localities, and institutions wherecollections are stored (see Institutional Abbreviations). Type locality in upper case.
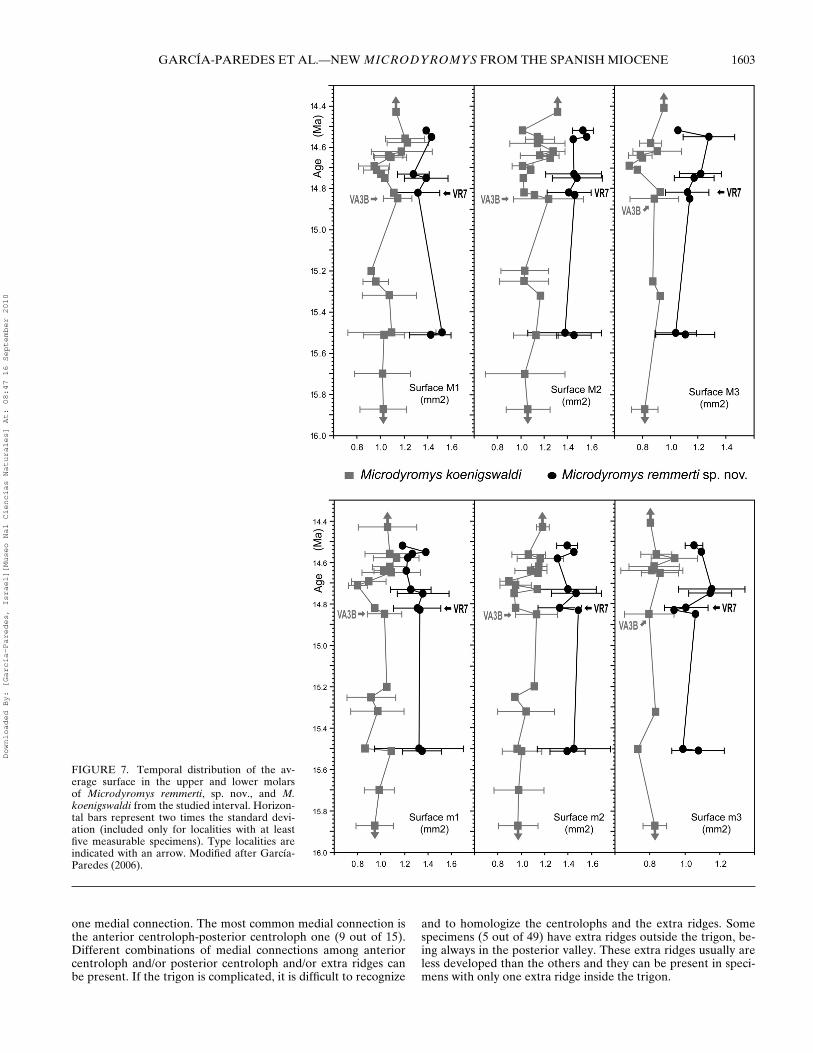
The measurements of specimens have been taken with a NikonMeasuroscope 10 microscope with digital micrometer in mmunits. ‘Width’ represents the maximum width of the crown, takenparallel to the contact facet of the posterior wall, in case ofpremolars and first molars, or the anterior wall in second andthird molars. ‘Length’ represents the maximum length of thecrown and has been measured at right angle to width. Wheneverpossible, two indexes have been calculated: ‘surface’ as lengthmultiplied by width in mm2, and the length/width ratio, a non-dimensional index which gives an idea of the slenderness of thetooth. Measurements and descriptive statistics of studied assem-blages of M. remmerti, sp. nov., are given in Tables 2 and 3. Thetemporal distribution of the average surface in the upper andlower molars of M. remmerti, sp. nov., and M. koenigswaldi fromthe studied interval are shown in Figure 7.
Analysis of variance (ANOVA) and post hoc test (displayed ashomogeneous subsets) have been carried out for size of M. rem-merti, sp. nov. The results are included in Supplementary Data(Tables S1 and S2; www.vertpaleo.org/jvp/JVPcontents.html).Levene test for homogeneity of variance was performed prior toANOVA.
Additional descriptive statistics for measurements of the com-pared species of Microdyromys are listed in Supplementary Data(Tables S3 and S4). Size differences between samples of Micrody-romys remmerti, sp. nov., and M. koenigswaldi from their type lo-calities have been evaluated using Student’s t-tests (Supplemen-tary Data, Table S5).
The number of ridges has been counted in every specimen, ex-cept for those partially broken. The accessory and less-developedridges have been counted independently of their size. Whenthere are two non-aligned structures in a valley, they have beencounted as different ridges; if they are aligned they have beencounted as the same, but interrupted, ridge. The frequencies ofthe number of ridges are given in Tables 4 and 5; based on thisinformation, schematic drawings have been used to show the po-sition layout and the number of ridges in the lower molars, andtheir frequencies have been calculated (Table 6). Correlationsbetween number of ridges and length per dental element havebeen tested using non-parametric correlations Spearman’s Rhotest (Supplementary Data, Tables S6–S8).
All the calculations and statistical analysis have been car-ried out with SPSS v. 11.5.1 and SPSS v. 16.0 (SPSS, 2002,2007).
Institutional Abbreviations—IAUU, Instituut voor Aard-wetenschappen, Utrecht University, Utrecht, The Netherlands;MNCN, Museo Nacional de Ciencias Naturales, CSIC, Madrid,Spain; RGM, Nationaal Natuurhistorisch Museum Naturalis, Ge-ology Department (formerly Rijksmuseum van Geologie en Min-eralogie), Leiden, The Netherlands.
SYSTEMATIC PALAEONTOLOGY
Order RODENTIA Bowdich, 1821Family GLIRIDAE Muirhead, 1819
MICRODYROMYS De Bruijn, 1965 [“1966”]
Original Diagnosis—“Small Gliridae with cheek teeth thathave a concave occlusal surface. Lingual outside of upper cheekteeth ornamented. The four main ridges of M1, M2, and M3 areconnected by an endoloph. Anterior centroloph of M1 and M2longer than posterior centroloph. P4/p4 and M3/m3 relativelylarge compared to other genera. The accessory ridges are lowerthan the main ridges” (De Bruijn, 1965 [“1966”]:11).
Emended Diagnosis—“Small Gliridae with a slightly or mod-erately concave occlusal surface and regular, parallel crest. Lin-gual wall of upper molars often crenulated. Tendency to forma complete endoloph in the older species, complete endolophpresent in the younger ones. Both centrolophs long, the precen-troloph being longer than the postcentroloph. Centrolophs gen-erally not interconnected. Extra crest outside the trigone rare”(Freudenthal and Martın-Suarez, 2007b:266).
Remarks on Microdyromys
Genus Microdyromys De Bruijn, 1965 [“1966”] (see noteon citation below), was defined in Miocene localities from theCalatayud-Montalban basin (Spain) and it is widely knownfrom Spain, and also from Portugal, France, Belgium, Germany,Switzerland, Austria, Poland, Czech Republic, Slovakia, Hun-gary, Italy, Greece, Turkey, Serbia, Georgia, and China. The
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

TA
BL
E2.
Des
crip
tive
stat
isti
csof
Mic
rody
rom
ysre
mm
erti,
sp.n
ov.,
uppe
rch
eek
teet
hfr
omth
elo
calit
ies
ofth
eA
rago
nian
type
area
(Cal
atay
ud-M
onta
lban
basi
n).
Len
gth
(L)
Wid
th(W
)Su
rfac
e(L
×W
)R
atio
L/W
NM
in.
Mea
nM
ax.
SDN
Min
.M
ean
Max
.SD
NM
in.
Mea
nM
ax.
SDN
Min
.M
ean
Max
.SD
P4
VA
3F2
0.81
0.82
0.83
20.
990.
990.
992
0.80
0.81
0.82
20.
820.
830.
84V
R8C
30.
700.
730.
773
0.80
0.87
0.92
30.
560.
630.
713
0.82
0.84
0.88
VR
8B6
0.73
0.79
0.87
0.04
66
0.87
0.95
1.05
0.05
86
0.64
0.76
0.91
0.09
06
0.81
0.83
0.85
0.01
3V
R7
170.
710.
770.
820.
028
170.
860.
931.
020.
044
170.
640.
710.
790.
049
170.
750.
830.
890.
040
VL
4B2
0.76
0.78
0.79
21.
001.
021.
032
0.76
0.79
0.81
20.
760.
760.
77V
L4A
140.
670.
780.
840.
049
150.
910.
971.
050.
040
140.
610.
760.
880.
073
140.
720.
800.
840.
036
M1
VA
3F4
1.09
1.11
1.13
41.
221.
251.
284
1.38
1.39
1.40
40.
850.
890.
93V
A3E
21.
081.
101.
122
1.29
1.31
1.32
21.
431.
441.
442
0.82
0.84
0.87
VR
8C5
1.01
1.08
1.12
0.04
33
1.19
1.21
1.22
31.
201.
281.
323
0.85
0.88
0.90
VR
8B12
1.02
1.12
1.17
0.04
912
1.10
1.23
1.30
0.05
511
1.20
1.39
1.52
0.09
411
0.86
0.90
0.94
0.02
8V
R7
520.
991.
081.
180.
041
421.
141.
221.
330.
048
421.
131.
321.
540.
093
420.
830.
890.
940.
027
VA
3D1
1.06
VL
4B2
1.17
1.17
1.17
21.
291.
301.
312
1.51
1.52
1.53
20.
890.
900.
91V
L4A
241.
041.
121.
190.
043
231.
191.
271.
360.
044
231.
291.
421.
580.
088
230.
830.
880.
930.
031
M2
VA
3F3
1.12
1.16
1.19
31.
281.
331.
383
1.48
1.53
1.57
30.
810.
870.
91V
A3E
21.
131.
171.
202
1.31
1.34
1.37
21.
481.
561.
642
0.86
0.87
0.88
VA
6A2
1.02
1.06
1.09
11.
331
1.45
10.
82V
R8C
121.
021.
121.
190.
053
121.
221.
301.
390.
056
121.
241.
451.
650.
120
120.
820.
860.
930.
031
VR
8B14
1.05
1.15
1.25
0.05
814
1.22
1.30
1.36
0.04
411
1.31
1.48
1.65
0.10
711
0.83
0.88
0.95
0.03
7V
R7
471.
011.
111.
200.
040
371.
171.
271.
400.
052
361.
201.
411.
560.
094
360.
790.
880.
940.
031
VA
3D2
1.14
1.17
1.20
21.
241.
251.
262
1.44
1.46
1.49
20.
900.
940.
97V
L4B
51.
021.
081.
180.
065
51.
221.
271.
400.
074
51.
261.
381.
650.
158
50.
800.
850.
890.
036
VL
4A23
1.01
1.10
1.14
0.03
519
1.27
1.32
1.38
0.03
219
1.30
1.46
1.56
0.07
419
0.78
0.83
0.88
0.02
6M
3V
A3F
10.
901
1.17
11.
051
0.77
VA
3E5
0.90
0.99
1.05
0.05
84
1.28
1.29
1.31
41.
161.
281.
384
0.70
0.77
0.80
VR
8C4
0.98
1.00
1.03
41.
151.
221.
284
1.15
1.22
1.30
40.
770.
820.
87V
R8B
60.
911.
011.
070.
057
41.
161.
201.
234
1.08
1.17
1.25
40.
760.
820.
88V
R7
310.
910.
981.
080.
046
291.
071.
151.
250.
041
281.
011.
121.
270.
078
280.
800.
850.
930.
039
VA
3B2
0.95
0.96
0.97
21.
161.
191.
212
1.10
1.14
1.17
20.
800.
810.
82V
L4B
60.
850.
880.
910.
021
41.
111.
171.
224
0.98
1.04
1.11
40.
740.
770.
79V
L4A
90.
850.
920.
980.
045
81.
151.
211.
360.
070
80.
991.
111.
320.
105
80.
710.
760.
840.
045
Abb
revi
atio
ns:M
ax,m
axim
umva
lues
;Min
,min
imum
valu
es;N
,num
ber
ofsp
ecim
ens;
SD,s
tand
ard
devi
atio
n.F
orab
brev
iati
ons
oflo
calit
ies,
thei
rlo
calb
iost
rati
grap
hica
lzon
es,a
ndth
eir
num
eric
alag
esse
eT
able
1.
1597
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

TA
BL
E3.
Des
crip
tive
stat
isti
csof
Mic
rody
rom
ysre
mm
erti,
sp.n
ov.,
low
erch
eek
teet
hfr
omth
elo
calit
ies
ofth
eA
rago
nian
type
area
(Cal
atay
ud-M
onta
lban
basi
n).
Len
gth
(L)
Wid
th(W
)Su
rfac
e(L
×W
)R
atio
L/W
NM
in.
Mea
nM
ax.
SDN
Min
.M
ean
Max
.SD
NM
in.
Mea
nM
ax.
SDN
Min
.M
ean
Max
.SD
p4V
A3F
20.
860.
890.
922
0.83
0.87
0.90
20.
710.
770.
832
1.02
1.03
1.04
VA
7C1
0.83
10.
891
0.74
10.
93V
R8C
90.
830.
880.
950.
039
100.
760.
820.
870.
035
90.
640.
730.
800.
056
91.
021.
071.
130.
039
VR
8B8
0.82
0.87
0.93
0.04
18
0.79
0.85
0.92
0.04
38
0.65
0.74
0.84
0.06
38
0.93
1.02
1.10
0.04
6V
R7
240.
810.
850.
890.
025
240.
750.
840.
910.
035
240.
620.
720.
810.
045
240.
951.
011.
090.
036
VL
4B3
0.80
0.87
0.94
50.
780.
820.
920.
057
30.
630.
740.
863
1.01
1.03
1.05
VL
4A11
0.81
0.86
0.91
0.02
611
0.79
0.85
0.90
0.03
111
0.64
0.74
0.81
0.04
511
0.98
1.01
1.05
0.02
3m
1V
A3F
11.
132
1.05
1.07
1.08
11.
191
1.08
VA
3E1
1.20
11.
151
1.38
11.
04V
A6A
11.
151
1.10
11.
271
1.05
VA
7C2
1.10
1.12
1.13
31.
081.
101.
112
1.22
1.23
1.24
20.
991.
011.
03V
A1A
11.
061
1.15
11.
221
0.92
VR
8C11
1.07
1.15
1.20
0.03
811
1.03
1.09
1.15
0.04
111
1.10
1.25
1.38
0.08
711
1.03
1.06
1.10
0.01
6V
R8B
111.
121.
191.
270.
052
101.
051.
151.
200.
051
101.
181.
361.
520.
111
100.
981.
041.
070.
030
VR
747
1.04
1.16
1.29
0.05
345
1.04
1.12
1.21
0.04
543
1.10
1.31
1.56
0.10
143
0.95
1.03
1.11
0.04
2V
A3D
11.
211
1.10
11.
331
1.10
VL
4B4
1.11
1.18
1.29
41.
061.
121.
234
1.18
1.33
1.59
41.
051.
051.
07V
L4A
271.
131.
201.
300.
036
261.
041.
131.
240.
047
241.
221.
351.
610.
083
241.
001.
071.
200.
045
m2
VA
3F1
1.22
21.
141.
171.
191
1.39
11.
07V
A3E
61.
191.
211.
230.
017
51.
161.
201.
220.
024
41.
391.
451.
484
0.98
1.01
1.03
VA
7C2
1.14
1.16
1.17
21.
131.
141.
142
1.30
1.31
1.32
21.
001.
021.
04V
R8C
131.
141.
201.
250.
038
121.
111.
161.
210.
033
121.
271.
401.
500.
079
121.
011.
041.
070.
020
VR
8B10
1.17
1.23
1.27
0.03
310
1.14
1.20
1.25
0.03
710
1.33
1.47
1.58
0.07
910
1.00
1.03
1.06
0.02
0V
R7
271.
111.
181.
250.
035
271.
061.
131.
230.
039
251.
211.
331.
490.
072
250.
981.
041.
090.
031
VA
3D1
1.28
21.
161.
171.
181
1.48
11.
10V
L4B
51.
191.
241.
290.
036
51.
121.
171.
260.
057
51.
331.
451.
630.
109
51.
021.
071.
110.
032
VL
4A20
1.12
1.21
1.26
0.04
021
1.08
1.15
1.22
0.03
420
1.27
1.39
1.52
0.07
720
0.97
1.05
1.10
0.03
1m
3V
A3F
31.
061.
081.
103
0.97
0.98
0.98
31.
031.
051.
083
1.09
1.10
1.12
VA
3E2
1.01
1.03
1.05
21.
051.
071.
082
1.09
1.10
1.10
20.
940.
971.
00V
R8C
71.
041.
101.
180.
054
60.
991.
041.
110.
049
61.
031.
151.
290.
095
60.
951.
061.
120.
060
VR
8B8
1.04
1.09
1.13
0.03
68
1.00
1.06
1.11
0.03
58
1.05
1.15
1.23
0.06
28
0.97
1.03
1.09
0.03
9V
R7
380.
921.
021.
120.
049
370.
910.
991.
060.
035
370.
861.
011.
140.
063
370.
871.
031.
110.
054
VA
3D2
0.95
0.97
0.99
20.
940.
971.
002
0.93
0.94
0.95
20.
951.
001.
05V
A3B
11.
051
1.01
11.
061
1.04
VL
4B1
0.99
11.
001
0.99
10.
99V
L4A
100.
981.
051.
100.
043
100.
941.
021.
080.
046
100.
961.
081.
190.
076
100.
921.
031.
100.
053
Abb
revi
atio
ns:M
ax,m
axim
umva
lues
;Min
,min
imum
valu
es;N
,num
ber
ofsp
ecim
ens;
SD,s
tand
ard
devi
atio
n.F
orab
brev
iati
ons
oflo
calit
ies,
thei
rlo
calb
iost
rati
grap
hica
lzon
es,a
ndth
eir
num
eric
alag
esse
eT
able
1.
1598
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010
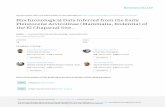
GARCIA-PAREDES ET AL.—NEW MICRODYROMYS FROM THE SPANISH MIOCENE 1599
FIGURE 3. Microdyromys remmerti, sp. nov.,from type locality Vargas 7. A, left P4, VR7-602 (seven ridges); B, left M1, VR7-638 (eightridges); C, left M2, VR7-622 (eight ridges); D,left M3, VR7-726 (eight ridges); E, invertedright P4, VR7-611 (seven ridges); F, invertedright M1, VR7–649 (eight ridges); G, left M2,VR7–673 (ten ridges); H, left M3, VR7–728(nine ridges); I, inverted right P4, VR7–614(seven ridges); J, inverted right M1, VR7–665(seven ridges); K, inverted right M2, VR7–697(nine ridges); L, inverted right M3, VR7–732(eleven ridges); M, inverted right P4, VR7–618(five ridges); N, inverted right M1, VR7–669(ten ridges); O, inverted right M2, VR7–700(holotype; nine ridges); P, inverted right M3,VR7–749 (eleven ridges). Scale bar equals 1mm.
designed type species for genus is Microdyromys koenigswaldiDe Bruijn, 1965 [“1966”].
The suprageneric status of Microdyromys has recentlychanged, has been transferred from the subfamily Dryomyinae(De Bruijn, 1967) to the subfamily Bransatoglirinae (Daams andDe Bruijn, 1995) by Freudenthal and Martın-Suarez (2007b).This question is out of the scope of this paper and is not discussedhere.
According to Freudenthal and Martın-Suarez (2007a), the old-est record of the genus is Microdyromys sp. from the Late Eoceneof Aguaton 2D (province of Teruel, Spain), reported as M.aff. misonnei in Freudenthal, 1997. These authors included theOligocene species Bransatoglis misonnei Vianey-Liaud, 1994, andB. heissigi Uhlig, 2001, in Microdyromys and give the above-mentioned emended diagnosis for the genus. This taxonomicalinterpretation fills the absence of record of Microdyromys fromLate Eocene and Early Oligocene; it was already pointed outby Freudenthal (1996), but remains controversial (e.g., Vianey-Liaud, 2003). As the suprageneric status of the genus, this is-sue is not discussed here. In contrast, some questions can be
pointed out regarding the emended diagnosis of Microdyromysby Freudenthal and Martın-Suarez (2007b). In our opinion, thereis a problem in the restriction of Microdyromys to small sizeGliridae, because it includes M. sinuosus (Alvarez Sierra, 1986,in Alvarez Sierra and Garcıa-Moreno, 1986), which can be con-sidered a medium-sized Gliridae (Table S3). Another questionis the presence of extra ridges outside the trigone (or trigon),which is rare in the species of Microdyromys from the Oligocene(sensu Freudenthal and Martın-Suarez, 2007a, 2007b), but it iscommon, even a diagnostic feature, in some Miocene species, asis discussed later.
Microdyromys sinuosus from the Late Miocene of Ampudia 3(MN 10, Duero basin, Spain) is the last occurrence of the genus.Franzen and Storch (1975) reported Microdyromys sp. from theTurolian (MN 11) locality Dorn-Durkheim 1 in Germany, but itwas assigned latter to Dryomys sp. by Daams (1981) and Daamsand De Bruijn (1995).
Note—We follow the Recommendation 22A of the Interna-tional Code of Zoological Nomenclature (ICZN, 1999), includingthe citation of both the actual date of publication and the imprint
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010
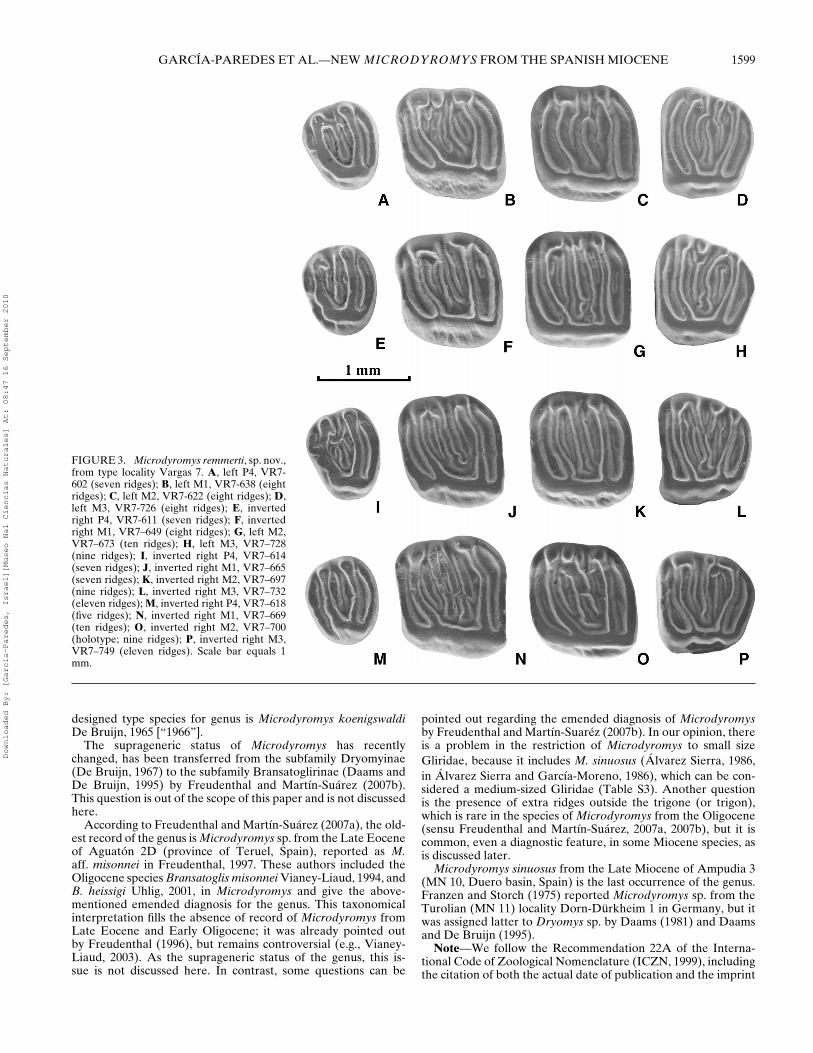
1600 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 30, NO. 5, 2010
FIGURE 4. Microdyromys remmerti, sp. nov.,from type locality Vargas 7. A, left p4,VR7–757 (eight ridges); B, left m1, VR7–790(nine ridges); C, left m2, VR7–846 (sevenridges); D, left m3, VR7–872 (seven ridges);E, left p4, VR7–761 (seven ridges); F, left m1,VR7–805 (nine ridges); G, inverted right m2,VR7–855 (nine ridges); H, inverted right m3,VR7–895 (seven ridges); I, left p4, VR7–771(seven ridges); J, left m1, VR7–809 (nineridges); K, left m2, VR7–850 (ten ridges); L,inverted right m3, VR7–896 (seven ridges);M, inverted right p4, VR7–777 (seven ridges);N, inverted right m1, VR7–813 (eight ridges);O, inverted right m2, VR7–857 (eight ridges);P, inverted right m3, VR7–900 (seven ridges).Scale bar equals 1 mm.
date. This is due to a nomenclatorial issue involving one speciesof this genus, which is discussed later.
MICRODYROMYS REMMERTI, sp. nov.(Figs. 3–6)
Holotype—Right M2, VR7–700 (Fig. 3O).Etymology—Dedicated to the memory of our colleague and
glirid specialist Dr. Remmert Daams.Type Locality and Horizon—Vargas 7, province of Zaragoza,
Calatayud-Montalban basin, Spain (local zone Dd, MN 5, middleAragonian, Middle Miocene).
Paratypes—P4: VR7–601 to VR7–605, VR7–607 to VR7–619;M1: VR7–620 to VR7–659, VR7–661 to VR7–672, VR7–682,VR7–686, VR7–938; M2: VR7–660, VR7–673 to VR7–681,VR7–683 to VR7–685, VR7–687 to VR7–699, VR7–701 toVR7–722, VR7–940; M3: VR7–723 to VR7–756; p4: VR7–757to VR7–765, VR7–767 to VR7–782; m1: VR7–783 to VR7–829,VR7–831 to VR7–837, VR7–848, VR7–854; m2: VR7–830,
VR7–838 to VR7–847, VR7–849 to VR7–853, VR7–855 toVR7–871; m3: VR7–872 to VR7–900, VR7–926 to VR7–934.
Repository Institution—Museo Nacional de Ciencias Natu-rales, CSIC (Madrid, Spain).
Diagnosis—Microdyromys characterized by: relatively largesize (for Microdyromys); moderately to very complicated andirregular dental pattern (seven to eleven ridges); shape, posi-tion, and/or connections of the ridges may be irregular and vari-able; the ornamentation on the lingual side of the upper molarsis strongly marked, forming a fold; M1 and M2 with long cen-trolophs and at least one extra ridge inside the trigon; protoloph,centrolophs, and metaloph with prominent and cusp-like labialend; centrolophs may be labially isolated in most complicatedspecimens; thick lingual end of the anterior centroloph, whichmay be connected medially to the posterior centroloph and/orto the anterior extra ridge of the trigon; lingual border of M2longer than the labial one; M3 with higher frequency of extra andlonger ridges outside the trigon than M1 and M2. Lower molarswith long centrolophid and well-developed extra ridges in the an-terior and posterior valleys; mesolophid and posterolophid with
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

GARCIA-PAREDES ET AL.—NEW MICRODYROMYS FROM THE SPANISH MIOCENE 1601
FIGURE 5. Microdyromys remmerti, sp. nov.,from Villafeliche 4A. A, left P4, VL4A-36 (fiveridges); B, left M1, VL4A-7 (nine ridges); C,left M2, VL4A-17 (eight ridges); D, left M3,VL4A-626 (nine ridges); E, left P4, VL4A-601 (five ridges); F, inverted right M1, RGM-390077 (eight ridges); G, left M2, VL4A-614(seven ridges); H, inverted right M3, VL4A-627 (nine ridges); I, inverted right p4, VL4A-51 (seven ridges); J, inverted right m1, VL4A-64 (seven ridges); K, inverted right m2, VL4A-79 (eight ridges); L, left m3, VL4A-91 (sevenridges); M, left p4, VL4A-628 (six ridges); N,left m1, VL4A-632 (nine ridges); O, left m2,VL4A-86 (eight ridges); P, left m3, VL4A-96(seven ridges). Scale bar equals 1 mm.
strong forward inflexion at labial border; centrolophid and pos-terior extra ridge may be labially connected to mesolophid andposterolophid, respectively; on the average, the order of additionof extra ridges in m1 and m2 is interpreted as occurs from the pos-terior valley to the anterior one, having up to three extra ridgesin the posterior valley; m3 simpler and more regular than m1 andm2.
Differential Diagnosis—Microdyromys remmerti, sp. nov., dif-fers from M. hildebrandti Werner, 1994, M. koenigwaldi DeBruijn, 1965 [“1966”], M. legidensis Daams, 1981, M. monspelien-sis Aguilar, 1977, M. orientalis Wu, 1986, and M. praemurinus(Freudenberg, 1941) in: larger size; higher number of ridges;more complex and irregular dental pattern. Differs from M. com-plicatus De Bruijn, 1965 [“1966”], in: larger size; more irreg-ular dental pattern; higher frequency of labially isolated cen-trolophs; lower frequency of extra ridges outside the trigon;in m1–m2 with more than seven ridges additional ridges oc-cur generally first in the posterior valley than in the more an-terior ones. Differs from M. sinuosus (Alvarez Sierra, 1986, inAlvarez Sierra and Garcıa-Moreno, 1986) in: smaller size; more
labially isolated centrolophs; ‘Spanish fan’ or ‘fish tail’ shapeof the M3; absence of a complete endolophid in the p4 andm1. Differs from Oligocene species (according to Freudenthaland Martın-Suarez, 2007a, 2007b) M. puntarronensis Freudenthaland Martın-Suarez, 2007a, M. missonnei (Vianey-Liaud, 1994),and M. heissigi (Uhlig, 2001) in: larger size; more complex den-tal pattern; presence of a complete endoloph in the upper mo-lars.
Stratigraphical Distribution—Local zone Dc (MN 5, middleAragonian, Middle Miocene) to local zone Dd (MN 5, middleAragonian, Middle Miocene).
Material and Measurements—See Tables 1–3.
Description of the Material from the Type Locality Vargas 7
P4—Oval outline and concave occlusal surface. The numberof ridges varies from five to seven (Table 4). The anteroloph isusually reduced (15 out of 17) and situated in the labial half ofthe occlusal surface. Specimens with intermediate degrees of re-duction of the anteroloph can be found. Two out of 17 specimens
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010
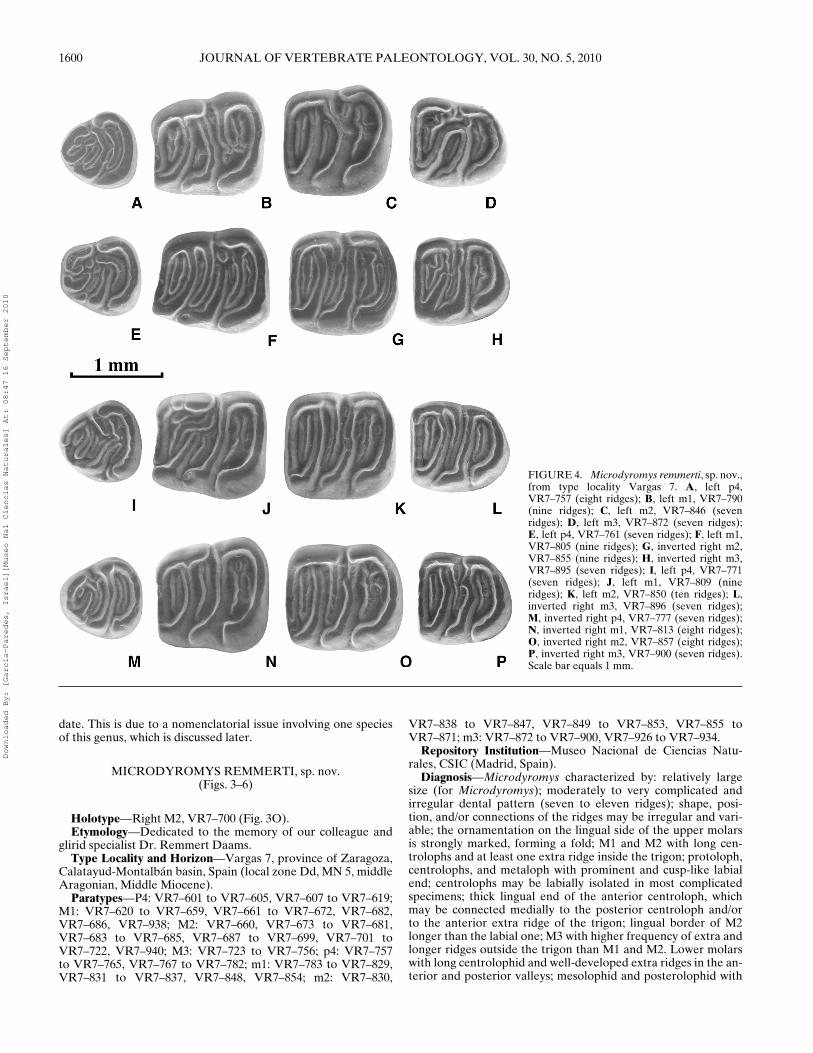
1602 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 30, NO. 5, 2010
FIGURE 6. Microdyromys remmerti, sp. nov.,from Vargas 8B. A, left P4, VR8B-187 (sixridges); B, left M1, VR8B-192 (ten ridges);C, inverted right M2, VR8B-207 (nine ridges);D, inverted right M3, VR8B-221; E, invertedright P4, VR8B-189 (seven ridges); F, invertedright M1, VR8B-194 (nine ridges); G, invertedright M2, VR8B-210 (nine ridges); H, invertedright M3, VR8B-224 (eight ridges); I, invertedright p4, VR8B-233 (seven ridges); J, invertedright m1, VR8B-237 (nine ridges); K, left m2,VR8B-252 (ten ridges); L, left m3, VR8B-262(seven ridges); M, left p4, VR8B-234 (sevenridges); N, inverted right m1, VR8B-245 (tenridges); O, left m2, VR8B-255 (ten ridges); P,inverted right m3, VR8B-269 (seven ridges).Scale bar equals 1 mm.
have a complete anteroloph. The trigon is relatively symmetricaland central. All the specimens have at least one centroloph thatis longer than half width of the occlusal surface. Eleven out of17 specimens show two centrolophs and the remaining ones haveone centroloph that cannot be homologized with either anterioror posterior one. Ten out of 16 specimens have an extra ridge(anterior or posterior) inside the trigon. In two of them, the ex-tra ridge is not interpreted as a second centroloph because of itsmorphology and situation. Connections inside the trigon are vari-able but, in general, if there are two centrolophs, the smaller oneis labially isolated. Medial connections between both centrolophsand/or with the extra ridge may be present.
M1—The M1 has a trapezium-like but rounded outline and aconcave occlusal surface. The number of ridges varies from sevento ten (Table 4). The lingual connection between anteroloph andprotoloph is curved and the end of the anteroloph can be dis-tinguished as part of the endoloph. The lingual wall has a verystrong fold-like ornamentation and it stands out as a furrow.The labial part of the metaloph is bent anteriorly in most spec-
imens (47 out of 49) and it is straight in only two specimens. Thelabial part of the posteroloph is bent anteriorly in all specimens.The trigon is asymmetrical U-shaped, and the vertical arms ofthe U appear to be bent anteriorly. Both centrolophs and ante-rior extra ridge of the trigon are always present. Centrolophs arelong, extending backwards and tend to occupy the entire valley.Anterior centroloph is longer than posterior one in most speci-mens (53 out of 54). The labial end of centrolophs is cusp-like.More than half of the specimens (27 out of 49) have both an-terior centroloph-protoloph and posterior centroloph-metalophlabial connections, 10 out of 49 specimens show only anteriorcentroloph-protoloph labial connection, 8 out of 49 specimensonly have posterior centroloph-metaloph, and 4 out of 49 spec-imens do not have labially connected centrolophs. The lingualend of anterior centroloph usually is bulky and it may have a lon-gitudinal extension connecting to posterior centroloph and/or tothe extra ridge. In 34 out of 49 specimens there are not medialconnections. Medial connections are always inside the trigon andthey are quite variable. In 15 out of 49 specimens there is at least
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010
GARCIA-PAREDES ET AL.—NEW MICRODYROMYS FROM THE SPANISH MIOCENE 1603
FIGURE 7. Temporal distribution of the av-erage surface in the upper and lower molarsof Microdyromys remmerti, sp. nov., and M.koenigswaldi from the studied interval. Horizon-tal bars represent two times the standard devi-ation (included only for localities with at leastfive measurable specimens). Type localities areindicated with an arrow. Modified after Garcıa-Paredes (2006).
one medial connection. The most common medial connection isthe anterior centroloph-posterior centroloph one (9 out of 15).Different combinations of medial connections among anteriorcentroloph and/or posterior centroloph and/or extra ridges canbe present. If the trigon is complicated, it is difficult to recognize
and to homologize the centrolophs and the extra ridges. Somespecimens (5 out of 49) have extra ridges outside the trigon, be-ing always in the posterior valley. These extra ridges usually areless developed than the others and they can be present in speci-mens with only one extra ridge inside the trigon.
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

1604 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 30, NO. 5, 2010
TABLE 4. Frequency, percentage (in parentheses), and mean number of ridges of the upper cheek teeth of Microdyromys remmerti, sp. nov.,from localities of the Aragonian type area (Calatayud-Montalban basin, Spain).
Number of ridges
5 6 7 8 9 10 11 12 N Mean
P4 VA3F 1 (33) 2 (67) 3 5.67VR8C 1 (33) 2 (67) 3 5.67VR8B 1 (25) 3 (75) 4 6.75VR7 2 (13) 6 (38) 8 (50) 16 6.38VL4B 1 (100) 1 5.00VL4A 10 (71) 4 (29) 14 5.29Total 15 (37) 15 (37) 11 (27) 41 5.90
M1 VA3F 2 (67) 1 (33) 3 7.33VA3E 1 (50) 1 (50) 2 7.50VR8C 3 (100) 3 7.00VR8B 3 (30) 1 (10) 2 (20) 4 (40) 10 8.70VR7 37 (76) 10 (20) 1 (2) 1 (2) 49 7.31VL4B 3 (100) 3 7.00VL4A 18 (78) 3 (13) 2 (9) 23 7.30Total 67 (72) 16 (17) 5 (5) 5 (5) 93 7.44
M2 VA3F 2 (100) 2 8.00VA3E 1 (50) 1 (50) 2 8.00VA6A 1 (100) 1 8.00VR8C 1 (9) 3 (27) 5 (45) 2 (18) 11 8.73VR8B 3 (23) 3 (23) 5 (38) 1 (8) 1 (8) 13 8.54VR7 25 (63) 9 (23) 4 (10) 2 (5) 40 7.58VA3D 1 (100) 1 9.00VL4B 1 (50) 1 (50) 2 7.50VL4A 12 (71) 4 (24) 1 (6) 17 7.35Total 45 (49) 23 (25) 17 (19) 5 (5) 1 (1) 91 7.84
M3 VA3F 1 (100) 1 7.00VA3E 1 (20) 4 (80) 5 8.80VR8C 2 (50) 2 (50) 4 9.50VR8B 2 (25) 4 (50) 1 (13) 1 (13) 8 10.13VR7 9 (29) 15 (48) 5 (16) 1 (3) 1 (3) 31 8.10VA3B 1 (50) 1 (50) 2 8.00VL4B 2 (67) 1 (33) 3 7.33VL4A 5 (63) 1 (13) 2 (25) 8 7.63Total 18 (29) 18 (29) 16 (26) 6 (10) 2 (3) 2 (3) 62 8.39
Abbreviations: N, number of specimens. For abbreviations of localities, their biostratigraphical zones, and their numerical ages see Table 1.
M2—This element is somewhat more complicated than M1 butshows similar characters. It has square but rounded outline. Thenumber of ridges varies from seven to ten (Table 4). The labialpart of the metaloph is bent anteriorly in most of specimens (35out of 40) but it is not as outstanding as in M1. Trigon is lessoblique than M1 and it has longer and straighter centrolophs andextra ridges. In most specimens (46 out of 48) the anterior cen-troloph is larger than posterior one, being the opposite in onespecimen and both centrolophs are equal in the remaining spec-imen. Connections inside the trigon are quite variable. In 22 outof 39 specimens only anterior centroloph-protoloph labial con-nection is present. Ten out of 39 specimens show both anteriorcentroloph-protoloph and posterior centroloph-metaloph labialconnections and five specimens show no one of them. The tworemaining specimens only have posterior centroloph-metalophlabial connection. Medial connections inside the trigon are morefrequent than in M1 and there are one (13 out of 41), two (3 outof 41), or three (1 out of 41), being most common the anteriorcentroloph–posterior centroloph one (11 out of 17). Extra ridgesoutside the trigon are present in the anterior valley (5 out of 40),in the posterior one (4 out of 40), or in both (2 out of 40).
M3—The morphological pattern is more variable than in M1and M2. The outline is ‘Spanish fan’ or ‘fish tail’ shape; the labialborder is rounded and it is longer than the lingual one. The num-ber of ridges varies from seven to twelve (Table 4). The frequencyof specimens with more than seven ridges (22 out of 31) is higherthan in M1 and M2, and extra ridges are more developed. Thetrigon is more symmetrical and central than in M1 and M2. Theornamentation on the lingual side is markedly strong between
lingual ends of protoloph and metaloph. Longer centroloph isthe anterior one (22 out of 32), the posterior (7 out of 32), orboth are equal (3 out of 32). In 12 out of 30 specimens eitheranterior centroloph-protoloph or posterior centroloph-metalophlabial connections are absent and only two specimens have bothconnections. Specimens with only anterior centroloph-protolophlabial connection (9 out of 30) or posterior centroloph-metalophlabial connection (7 out of 30) are present. Medial connections in-side the trigon are more frequent than in M1 and M2, being one(11 out of 31), two (5 out of 31), or three (4 out of 31). The mostcommon medial connection is between the anterior centrolophand posterior centroloph (11 out of 20). Extra ridges outside thetrigon are present in the anterior valley (6 out of 33) or in bothanterior and posterior ones (1 out of 33).
p4—The outline is oval and the occlusal surface is concave,which is mainly constituted by posterior part of the tooth. Themorphological structures of the anterior part are very close to-gether. The number of ridges can be six, seven, or eight (Ta-ble 5); the different connections among ridges are variable. An-terolophid and metalophid are reduced. In most specimens (14out of 17) there is a small extra ridge in the anterior valley. Allspecimens have a centrolophid and an extra ridge in the poste-rior valley; both ridges are well developed and situated in themedial part. Mesolophid and posterolophid are long comparedto the width of the occlusal surface; their labial and lingual partsare usually bent anteriorly. The mesolophid extends along thelabial border and it can be connected to the metalophid and/orcentrolophid. Mesolophid can be divided and connected to theanterior part of the lingual border through central valley.
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

GARCIA-PAREDES ET AL.—NEW MICRODYROMYS FROM THE SPANISH MIOCENE 1605
TABLE 5. Frequency, percentage (in parentheses), and mean number of ridges of the lower cheek teeth of Microdyromys remmerti, sp. nov.,from localities of the Aragonian type area (Calatayud-Montalban basin, Spain).
Number of ridges
5 6 7 8 9 10 11 N Mean
p4 VA3F 2(100) 2 7.00VR8C 1 (17) 5 (83) 6 6.83VR8B 5 (100) 5 7.00VR7 3 (18) 12 (71) 2 (12) 17 6.94VL4B 4 (100) 4 6.00VL4A 1 (10) 4 (40) 5 (50) 10 6.40Total 1 (2) 12 (27) 29 (66) 2 (5) 44 6.73
m1 VA3F 1 (100) 1 10.00VA3E 1 (100) 1 10.00VA7C 1 (50) 1 (50) 2 8.50VA1A 1 (100) 1 9.00VR8C 1 (10) 4 (40) 2 (20) 3 (30) 10 9.70VR8B 1 (10) 3 (30) 4 (40) 2 (20) 10 9.70VR7 9 (21) 14 (33) 12 (28) 6 (14) 2 (5) 43 8.49VL4B 1 (33) 2 (67) 3 7.67VL4A 6 (27) 14 (64) 2 (9) 22 7.82Total 16 (17) 33 (35) 23 (25) 14 (15) 7 (8) 93 8.60
m2 VA3F 1 (50) 1 (50) 2 9.50VA3E 1 (14) 3 (43) 3 (43) 7 9.29VA7C 1 (50) 1 (50) 2 8.00VR8C 2 (20) 4 (40) 3 (30) 1 (10) 10 9.30VR8B 4 (40) 2 (20) 4 (40) 10 8.40VR7 14 (56) 5 (20) 3 (12) 2 (8) 1 (4) 25 7.84VL4B 2 (67) 1 (33) 3 7.33VL4A 17 (89) 2 (11) 19 7.11Total 38 (48) 14 (18) 12 (15) 13 (16) 2 (3) 79 8.08
m3 VA3F 3 (100) 3 7.00VA3E 2 (100) 2 7.00VR8C 4 (67) 2 (33) 6 7.33VR8B 7 (88) 1 (13) 8 7.13VR7 30 (91) 3 (9) 33 7.09VA3D 1 (100) 1 7.00VA3B 1 (100) 1 7.00VL4A 2 (20) 8 (80) 10 6.80Total 2 (3) 58 (88) 6 (9) 66 7.06
Abbreviations: N, number of specimens. For abbreviations of localities, their biostratigraphical zones, and their numerical ages see Table 1.
m1—The outline is trapezoidal and the anterior part is muchnarrower than posterior one. The occlusal surface is concave andthe labial border is quite prominent. The number of ridges variesfrom seven to eleven (Table 5). The m1 is the dental elementhaving more variability and its pattern can be the most irregu-lar and complicated. All specimens have a long centrolophid andvery well developed extra ridges in the anterior and posterior val-ley. The centrolophid can be connected labially to mesolophid (4out of 43) or metalophid (2 out of 43). From this simplest mor-phology, the number of additional extra ridges increases fromthe posterior valley to the anterior one. In 20 out of 43 spec-imens there are at least two extra ridges in the posterior val-ley. There are specimens (8 out of 43) with three ridges betweenmesolophid and posterolophid, having in some cases medial con-nections among these ridges. Labial part of mesolophid and pos-terolophid extends anteriorly along the labial border; the labialend of the posterior valley can be almost closed. In the posteriorvalley the main extra ridge is labially connected to posterolophidin 12 out of 43 specimens. In some specimens the dental patternis irregular and chaotic in the shape, position, and connectionsof ridges, making its description difficult. In these irregular andcomplicated specimens it is hard to recognize and to homologizeeven parts of main ridges.
m2—The outline is almost rectangular with anterior and pos-terior parts having similar width. The occlusal surface is concaveand the labial border is quite prominent. In general, m2 has samecharacters as the m1 but it is less variable and less complicated.The number of ridges varies from seven to eleven (Table 5). All
specimens have a long centrolophid and well-developed extraridges in the anterior and posterior valley. Although there arecertain variabilities from the simplest general morphology, thenumber of ridges increases from the posterior valley to the moreanterior ones. There are specimens having irregular morphologybut not as extreme and variable as in m1.
m3—The occlusal surface is concave and the labial border isprominent. The posterior part is rounded and of similar widththan anterior one. The number of ridges is seven or eight (Ta-ble 5). All specimens have at least an extra ridge in the anteriorand the posterior valley showing variable development. The cen-trolophid is long and can be connected to the mesolophid in thelabial border. In most specimens, the centrolophid is connectedto lingual border at right angle. Additional extra ridge is placedat the posterior valley in all specimens with eight ridges. Usuallythis ridge is less developed or even incipient. Some specimensshow certain irregularity concerning mainly medial connectionsamong ridges.
COMPARISONS AND DISCUSSION
Daams (1981) gave detailed description of Microdyromys fromseveral localities in Germany, Switzerland, France, and Spain, an-alyzing the distribution of standard morphotypes of the cheekteeth. Most of the localities (17 out of 34) studied by this authorbelong to the Miocene of the Calatayud-Montalban basin. Ac-cording to Daams (1981:60), the rigorous distinction of all spec-imens among the different species of Microdyromys from the
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

1606 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 30, NO. 5, 2010
TABLE 6. Frequency and percentage (in parentheses) of number and position of ridges in the lower molars of Microdyromys remmerti, sp.nov., from localities of the Aragonian type area (Calatayud-Montalban basin, Spain).
N
m1 VA3F 1 (100) 1VA3E 1 (100) 1VA7C 1 (50) 1 (50) 2VA1A 1 (100) 1VR8C 1 (10) 2 (20) 2 (20) 2 (20) 3 (30) 10VR8B 1 (13) 1 (13) 2 (25) 1 (13) 2 (25) 1 (13) 8VR7 9 (21) 4 (9) 10 (23) 4 (9) 4 (9) 4 (9) 4 (9) 2 (4) 1 (2) 1 (2) 43VL4B 1 (33) 2 (67) 3VL4A 6 (27) 14 (64) 2 (9) 22Total 16 (17) 21 (23) 12 (13) 6 (7) 9 (10) 6 (7) 8 (9) 7 (8) 5 (6) 1 (1) 91
m2 VA3F 1 (50) 1 (50) 2VA3E 1 (14) 2 (29) 1 (14) 3 (33) 7VA7C 1 (50) 1 (50) 2VR8C 2 (20) 1 (10) 1 (10) 1 (10) 2 (20) 2 (20) 1 (10) 10VR8B 4 (40) 2 (20) 2 (20) 2 (20) 10VR7 14 (56) 3 (12) 4 (16) 1 (4) 2 (8) 1 (4) 25VL4B 2 (67) 1 (33) 3VL4A 17 (89) 2 (11) 19Total 38 (48) 8 (10) 8 (10) 1 (1) 4 (5) 3 (4) 5 (6) 10 (13) 2 (3) 79
m3 VA3F 3 (100) 3VA3E 2 (100) 2VR8C 4 (67) 2 (34) 6VR8B 7 (88) 1 (13) 8VR7 30 (91) 1 (3) 2 (6) 33VA3D 1 (100) 1VA3B 1 (100) 1VL4A 2 (20) 8 (80) 10Total 2 (3) 58 (88) 1 (2) 5 (8) 66
Abbreviations: N, number of specimens. For abbreviations of localities, their biostratigraphical zones, and their numerical ages see Table 1.
Miocene is quite difficult because the large overlap in size andmorphology. This overlapping among species has been pointedout by Garcıa-Paredes (2006) as well.
Since Daams’ paper on Microdyromys, the number of local-ities, their temporal succession, and the number of specimensfrom the Aragonian type area have increased enormously. Thesefacts allow us to better define the different species and evaluatetheir differences in size and morphology among the various as-semblages. The observed metrical and morphological variation ofM. remmerti, sp. nov., and comparisons with co-occurring and/ormorphologically similar species are discussed below.
Microdyromys remmerti, sp. nov., from Localities in the StudiedArea
All the studied material of Microdyromys remmerti, sp. nov.is listed in Table 1; the measurements and some morphologicalcharacters are given in Tables 2–6. Microdyromys remmerti, sp.nov., is characterized by combining a relatively large size and amoderately complicated to very complicated dental morphology.One of the observed features is a gradient in complication of theupper cheek teeth because, in general, the number of ridges in-creases from the P4 through the M3 (Table 4).
Although the stratigraphical distribution of M. remmerti, sp.nov., is relatively short, being restricted to the middle Aragonian(Middle Miocene), it displays important changes in morphology:a general increase in the complexity of the dental pattern throughtime (Tables 4–6), showing a unique pattern in the addition ofextra ridges in lower molars. This species shows a wide range ofmorphological variation with intermediate forms filling the con-tinuous range. One of the extremes of the morphological range isrepresented by specimens with a chaotic and irregular pattern inshape, position, and connections among ridges; in these cases it isdifficult to recognize and to homologize the different ridges (e.g.,Fig. 6G).
The first record of M. remmerti, sp. nov., comes from Villafe-liche 4A (local biozone Dc). This locality contains abundant ma-terial and all the typical features of this species are present, butin lower frequency than in younger assemblages (e.g., number ofextra ridges, number of longitudinal connections among ridges,labially isolated centrolophs). The morphology, in general, is stillrelatively simpler and less irregular (Fig. 5) than the type mate-rial (Figs. 3, 4), thus representing a less derived assemblage, moresimilar in morphology to that of M. koenigswaldi, as is discussedlatter.
From Villafeliche 4A (local zone Dc) to Vargas 8C (local zoneDd) there is an increase in the complexity of the dental pattern.The number of ridges and the specimens with a chaotic and ir-regular pattern in shape, position, and connections among ridgeshave the highest frequency in Vargas 8C for lower molars andVargas 8B for upper ones (Tables 4–5; Fig. 6). After this localitythe number of specimens is very low and a clear increase in thenumber of ridges cannot be observed; however, some cases showhigher number of connections among ridges, which can also beunderstood as an increase in the complexity of the dental pattern.
One of the important morphological features characterizingMicrodyromys remmerti, sp. nov., is the position of additionalextra ridges in the m1 and m2. From the simplest morphologywith seven ridges, the addition of more extra ridges takes placefirst in the posterior valley (Table 6). Within the total m1 of M.remmerti, sp. nov., 57 out of 91 specimens have two or three ex-tra ridges in the posterior valley. These specimens represent 76%of the total m1 with more than seven ridges, the remaining 24%are specimens having the additional ridges at one or both sidesof the centrolophid, but not in the posterior valley. The simplestseven ridges morphology in the m2 is more frequent (38 out of79 specimens) than in the m1. Within the specimens with morethan seven ridges, the 78% (32 out of 41 specimens) have two orthree extra ridges in the posterior valley and the 22% have anextra ridge at one or both sides of the centrolophid (Table 6).
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

GARCIA-PAREDES ET AL.—NEW MICRODYROMYS FROM THE SPANISH MIOCENE 1607
The m3 have a simpler morphology than the m1 and m2. Themost complicated m3 have more longitudinal connections amongridges than the simple ones; but only 6 out of 66 specimens havemore than seven ridges (Table 5). Regarding the size of Mi-crodyromys remmerti, sp. nov., a detailed analysis of the aver-age length, width, surface, and length/width ratio of the cheekteeth from the Aragonian type area (Tables 2 and 3) have beenperformed. The results of ANOVA test, considering together allstudied assemblages, show generalized significant differences (P< 0.05) for the metrical variables (Table S1). Particular differ-ences among localities have been evaluated by means of posthoc test, which are displayed as homogeneous subsets combiningnon-significantly different means for groups in the same column(Table S2). The results are not consistent with any generalizedpattern in the different dental elements and/or the different met-rical variables. Depending on the dental element and the metri-cal variable, differences are significant among different localities.Similarly, the temporal distribution of the average surface for up-per and lower molars in M. remmerti, sp. nov., from the Arago-nian type area (Fig. 7) shows a random walk rather than a generaltrend towards size increase or decrease.
Microdyromys koenigswaldi
Microdyromys remmerti, sp. nov., always co-occurs with M.koenigswadi in the studied area. The morphological differencesbetween the two species are conspicuous due to the more com-plicated and irregular dental pattern of M. remmerti, sp. nov. Al-though, older assemblages of M. remmerti, sp. nov., from the bio-zone Dc are morphologically closer to M. koenigswaldi, there areclear differences in size (Fig. 7) and dental pattern, as has beenpointed out before in the discussion of the material from Villafe-liche 4A. We must say that in this locality (among others, e.g., Vil-lafeliche 4B, Valdemoros 3B, Valdemoros 3D, Vargas 8C, Valde-moros 7C, Valdemoros 3F) also co-occur the smaller and sim-pler M. monspeliensis sensu Daams, 1981 (Garcıa-Paredes, 2006),which is not discussed here. Size differences between Micrody-romys koenigswaldi and M. remmerti, sp. nov., have been evalu-ated by performing Student’s t-test in the type localities (exceptfor P4 and p4). The differences (Table S5) are highly significant(P < 0.01) in length, width, and surface for all molars, M. rem-merti, sp. nov., being the larger one. The length/width ratio onlydiffers in the M1 (P < 0.01), element which is more elongated inM. remmerti, sp. nov., than in M. koenigswaldi.
We can conclude that, in the studied area, there is a divergentevolution of M. remmerti, sp. nov., in relation to the more conser-vative M. koenigswaldi. First, there is a divergence in size (Fig. 7),later in morphology due to the general increase in the complexityof M. remmerti, sp. nov.
Microdyromys sinuosus
Among the species of the genus, the assemblages of Micrody-romys remmerti, sp. nov., with more complicated morphologymost closely resembles M. sinuosus. This species is only knownfrom the Late Miocene (MN 10) of the Duero basin (Spain).However, because of the scarcity of the available material of M.sinuosus (Table S3), its morphological and metrical ranges arenot well known. Both species have a complicated and somewhatirregular dental pattern, but the lower cheek teeth of M. rem-merti, sp. nov., do not display an endolophid, the upper molarsusually have labially isolated centrolophs, and low frequency ofextra ridges outside the trigon; the pattern of addition of extraridges in the lower molars cannot be observed in M. sinuosus.The few available m1 and m2 of M. sinuosus described in AlvarezSierra and Garcıa Moreno (1986) have three extra ridges in theposterior valley but also two well-developed extra ridges at bothsides of the centrolophid, being more similar to that of M. compli-
catus (as far as the position of extra ridges is concerned). More-over, the size of the cheek teeth is smaller in M. remmerti, sp.nov., than in M. sinuosus (Table S3).
Microdyromys complicatus, M. miocaenicus, and aNomenclatorial Issue
Microdyromys complicatus De Bruijn, 1965 [“1966”], was de-scribed in the Middle Miocene locality Armantes 7 (MN 6), sit-uated in the Calatayud-Montalban basin. Although this speciesis present in the studied area in MN 5 (Garcıa-Paredes, 2006), itdoes not co-occur with M. remmerti, sp. nov., in any locality. Mi-crodyromys complicatus has a complicated dental morphology,which can display a number of ridges similar to M. remmerti, sp.nov., and somewhat overlapping in the morphology of the lowermolars. Nevertheless, M. complicatus differs by its smaller size(Table S3), its more regular dental pattern, and having alwayswell-developed extra ridges outside the trigon in the upper mo-lars. In M. remmerti, sp. nov., the lingual end of the anterior cen-troloph may be connected medially with the protoloph and thereis higher frecuency of both centrolophs labially isolated. If the m1and m2 of M. remmerti, sp. nov., have more than seven ridges, theadditional ridges occurs first in the posterior valley and onwards,whereas in M. complicatus additional ridges are always both sidesof centrolophid.
Outside the Iberian Peninsula, material of Microdyromys withcomplicated morphology, similar to that of M. remmerti, sp. nov.,has been described as M. complicatus or M. miocaenicus (Baude-lot, 1966 [“1965”]) from several Miocene localities of Europe(e.g., Baudelot, 1972; Engesser, 1972; Mayr, 1979; Wu, 1990), buta nomenclatural and taxonomical consideration regarding bothnames must be pointed out. Engesser (1972), Mayr (1979), andWu (1990) agree in considering same species the material de-scribed in Sansan (type locality of M. miocaenicus) and Armantes7 (type locality of M. complicatus); they stated that M. miocaeni-cus has priority over M. complicatus due to the imprint date (1965and 1966, respectively). However, as discussed by Daams (1981),Daams and De Bruijn (1995), and Wu (1993), the name M. com-plicatus has priority because Baudelot’s paper came out in 1966with imprint date of 1965, but De Bruijn’s paper came out in1965 with imprint date of 1966. Due to the difference betweenthe actual date and the imprint date of both papers, the priorityof the name has remained confusing. More recent papers (e.g.,Kalin, 1997; Kalin, 2003; Kalin and Kempf, 2009; Sach, 1999)still retain the name M. miocaenicus to describe Microdyromysmaterial from central Europe. Given that, we propose to followRecommendation 22A.2 of the International Code of ZoologicalNomenclature (ICZN, 1999:24). Thus, including the citation ofboth the actual and the imprint date, the priority of the name isclear and the citation for the bibliographic reference is not con-fusing: Microdyromys complicatus De Bruijn, 1965 [“1966”], orM. complicatus De Bruijn, 1965 [imprint 1966], versus M. mio-caenicus (Baudelot, 1966 [“1965”]).
FINAL REMARKS AND CONCLUSIONS
Although some of the morphological characters defining M.remmerti, sp. nov., are present in other species of Gliridae,its unique combination of morphometrical features have neverbeen found in any other glirid material outside the Calatayud-Montalban basin. The present information suggests that thisspecies can be considered an endemic form. This idea is in accor-dance with the endemic character of the Gliridae faunas from theIberian Miocene, with several genera and species almost exclu-sive to the Iberian Peninsula (see Daams and De Bruijn, 1995).The morphological similarities between M. koenigswaldi and theolder and less derived assemblages of M. remmerti, sp. nov.,indicates a probable ancestor-descendant relationship. The twospecies co-occur in the studied area and M. remmerti, sp. nov.,
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

1608 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 30, NO. 5, 2010
shows a divergent evolution in relation to the more conservativeM. koenigswaldi by its larger size and more complicated dentalpattern. In general, this species shows an increase in the complex-ity of the dental pattern through time without a clear change insize. We understand that underlying the general increase in com-plexity of the morphology of the m1 and m2 there is a ‘structural’pattern in the addition of extra ridges. In our opinion, this pat-tern can be related with the morphogenetic processes involvedin the development of the teeth and we have assigned it a sys-tematic value. The position of the extra ridges has already beenused in Gliridae as species-level diagnostic character (De Bruijn,1966; 1967), and it has been discussed in Van der Meulen and DeBruijn (1982).
On the other hand, the morphological change of a feature re-lated functionally with the use/exploitation of trophic resourcesreduces the competitiveness between species by reducing theniches overlap and it facilitates the co-existence. In terms of func-tional morphology, it is important to evaluate whether the ob-served morphological changes of M. remmerti, sp. nov., are re-lated to tooth size. Therefore, non-parametric correlations be-tween ‘number of ridges’ and ‘length’ have been performed. Theresults are highly significant (P < 0.01) for the upper molars whenall the assemblages are analyzed together (Table S6), whereascorrelation for the lower cheek teeth is not significant. In thissense, as in the pattern of addition of extra ridges, upper andlower cheek teeth seem to be developed in a different way. How-ever, if the analysis is performed per locality, the correlationsonly are significant (P < 0.05) in Villafeliche 4A and Vargas 7for the M1, Vargas 7 for the M3, Villafeliche 4A for the m1,and Vargas 8C for the m3 (Tables S7 and S8). These results donot allow us to conclude that there is a generalized correlationbetween tooth length and number of ridges, especially in thelower cheek teeth. If the increase in complexity and number ofridges occur without the increase in size of dental elements (espe-cially in length), then the ridge spacing decreases. This divergentmorphology between M. remmerti, sp. nov., and M. koenigswaldigives more evidence of a change in exploitation of trophic re-sources than a simple concurrent increase in both occlusal surfaceand ridge number.
In fact, the stratigraphical distribution of M. remmerti, sp. nov.,is coincident with the Mid-Miocene Climate Optimum (Zachos etal., 2001). According to Van Dam et al. (2006), the beginning ofthe local biozone Dc in the Iberian Peninsula is after a period ofeccentricity minima and it falls within one of the turnover peaksthat represent a more humid period. Bohme et al. (2006) useda method for qualitatively estimating paleoprecipitation basedon fossil herpetological communities. The estimation of paleo-precipitation gives significantly low, arid to semiarid values forthe Calatayud-Daroca Basin during the early Middle Miocene ingeneral (15.9–15.3 Ma), which are in accordance with sedimento-logical data of the Calatayud-Daroca basin (Daams et al., 1999c).This study indicates the existence of a significant increase in pre-cipitation in the locality Villafeliche 4A, corresponding with thefirst record of M. remmerti, sp. nov., All these data indicate anincrease in the environmental variability during the middle Arag-onian (Middle Miocene). Thus, the availability of new ecologicalniches can be related with the evolution towards new morpho-types exemplified by M. remmerti, sp. nov.
ACKNOWLEDGMENTS
We thank all the colleagues and students who helped us in thefield work for many years, L. W. van den Hoek Ostende and A.J. van der Meulen for their supportive comments and the transla-tion of various texts from German, and S. K. Donovan for valu-able suggestions on the English style of the manuscript and thenomenclatorial issues. We acknowledge the Nationaal Natuurhis-torisch Museum Naturalis, Department of Earth Sciences of the
Utrecht University (The Netherlands), and Museo Nacional deCiencias Naturales, CSIC (Madrid, Spain), for the provided fa-cilities to study their collections. Editor R. Asher, A. R. Evans,and an anonymous reviewer made constructive suggestions forimprovement of the manuscript. We acknowledge financial sup-port by the projects PB 98–0691-C03–02, CGL2004–02094/BTE,CGL2007–65208/BTE, CGL2008–04200/BTE, Research GroupUCM 910607, and FECYT-MICIIN postdoctoral contract toI.G.-P.
LITERATURE CITED
Aguilar, J.-P. 1977. Les gisements continentaux de Plaissan et de la nou-velle faculte de medecine (Herault); leur position stratigraphique.Geobios 10:81–101.
Alvarez Sierra, M. A., and E. Garcıa-Moreno. 1986. New Gliridae andCricetidae from the middle and upper Miocene of the Duero basin,Spain. Studia Geologica Salmanticensia 22:145–189.
Alvarez Sierra, M. A., J. P. Calvo, J. Morales, A. Alonso-Zarza, B.Azanza, I. Garcıa Paredes, M. Hernandez Fernandez, A. J. van derMeulen, P. Pelaez-Campomanes, V. Quiralte, M. J. Salesa, I. M.Sanchez, and D. Soria. 2003. El transito Aragoniense–Vallesienseen el area de Daroca-Nombrevilla (Zaragoza, Espana). Coloquiosde Paleontologıa Volumen Extraordinario 1:25–33.
Baudelot, S. 1965. Complement a l’etude des rongeurs de Sansan: lesGlirides. Bulletin de la Societe Geologique de France 7:758–764.
Baudelot, S. 1972. Etude des chiropteres, insectivores et rongeurs duMiocene de Sansan. Ph.D. dissertation, Universite Paul Sabatier deToulouse, Toulouse, 364 pp.
Bowdich, T. E. 1821. An Analysis of the Natural Classification of Mam-malia for the Use of Students and Travellers. J. Smith, Paris, 115pp.
Bohme, M., A. Ilg, A. Ossig, and H. Kuchenhoff. 2006. New method toestimate paleoprecipitation using fossil amphibians and reptiles andthe middle and late Miocene precipitation gradients in Europe. Ge-ology 34:425–428.
De Bruijn, H. 1966. Some new Miocene Gliridae (Rodentia, Mammalia)from the Calatayud area (prov. Zaragoza, Spain). Proceedings of theKoninklijke Nederlandse Akademie Van Wetenschappen Series B69:58–78.
De Bruijn, H. 1967. Gliridae, Sciuridae y Eomyidae (Rodentia, Mam-malia) miocenos de Calatayud (provincia de Zaragoza, Espana) ysu relacion con la bioestratigrafıa del area. Boletın del InstitutoGeologico y Minero de Espana 78:189–373.
De Bruijn, H., R. Daams, G. Daxner-Hock, V. Fahlbusch, L. Ginsburg, P.Mein, and J. Morales. 1992. Report of the RCMNS working groupon fossil mammals, Reisensburg 1990. Newsletters on Stratigraphy26;65–118.
Daams, R. 1981. The dental pattern of the Dormice Dryomys, My-omimus, Microdyromys and Peridyromys. Utrecht Micropaleonto-logical Bulletins, Special Publication 3:1–113.
Daams, R., and H. de Bruijn. 1995. A classification of the Gliridae (Ro-dentia) on the basis of dental morphology. Hystrix (n. s.) 6:3–50.
Daams, R., M. Freudenthal, and M. A. Alvarez Sierra. 1987. Ramblian: anew stage for continental deposits of Early Miocene age. Geologieen Mijnbouw 65:297–308.
Daams, R., M. Freudenthal, and A. van de Weerd. 1977. Aragonian, anew stage for continental deposits of Miocene age. Newsletters onStratigraphy 6:42–55.
Daams, R., A. J. Van Der Meulen, P. Pelaez-Campomanes, and M.A. Alvarez-Sierra. 1999a. Trends in rodent assemblages from theAragonian (early-middle Miocene) of the Calatayud–Daroca Basin,Aragon, Spain; pp. 127–139 in J. Agustı, L. Rook, and P. Andrews(eds.), Hominoid Evolution and Climatic Change in Europe. Vol-ume 1. The Evolution of Neogene Terrestrial Ecosystems in Europe.Cambridge University Press, Cambridge, U.K.
Daams, R., A. J. Van Der Meulen, M. A. Alvarez Sierra, P. Pelaez-Campomanes, and W. Krijgsman. 1999b. Aragonian stratigraphyreconsidered, and a re-evaluation of the middle Miocene mam-mal biochronology in Europe. Earth and Planetary Science Letters165:287–294.
Daams, R., A. J. Van Der Meulen, M. A. Alvarez Sierra, P. Pelaez-Campomanes, J. P. Calvo, M. A. Alonso Zarza, and W. Krijgsman.1999c. Stratigraphy and sedimentology of the Aragonian (Early to
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010

GARCIA-PAREDES ET AL.—NEW MICRODYROMYS FROM THE SPANISH MIOCENE 1609
Middle Miocene) in its type area (North-Central Spain). Newsletterson Stratigraphy 37:103–139.
Engesser, B. 1972. Die obermiozane Saugetierfauna von Anwil (Basel-land). Tatigkeitsberichte der Naturforschenden Gesellschaft Basel-land 28:37–363.
Franzen, J. L., and G. Storch. 1975. Die unterpliozane (turolis-che) Wirbeltierfauna von Dorn-Durkheim, Rheinhessen (SW-Deutschland): I. Entdeckung, Geologie, Mammalia: Carnivora, Pro-boscidea, Rodentia—Grabungsergebnisse 1972–1973. Senckenber-giana lethaea 56:233–303.
Freudenberg, H. 1941. Die oberoligocanen Nager von Gaimersheim beiIngolstadt und ihre Verwandten. Palaeontographica A92:99–164.
Freudenthal, M. 1996. The Early Oligocene rodent fauna of Olalla 4A(Teruel, Spain). Scripta Geologica 112:1–67.
Freudenthal, M. 1997. Paleogene rodent faunas from the province ofTeruel (Spain); pp. 397–415 in J.-P. Aguilar, S. Legendre, and J.Michaux (eds.), Actes du Congres BiochroM’97. Memoires TravauxEcole Pratique des Hautes Etudes, Institut Montpellier 21.
Freudenthal, M., and E. Martın-Suarez. 2007a. Microdyromys (Gliri-dae, Rodentia, Mammalia) from the Early Oligocene of Montalban(Prov. Teruel, Spain). Scripta Geologica 135:179–211.
Freudenthal, M., and E. Martın-Suarez. 2007b. Revision of the subfamilyBransatoglirinae (Gliridae, Rodentia, Mammalia). Scripta Geolog-ica 135:241–273.
Garcıa-Paredes, I. 2006. Patrones evolutivos de los Gliridae (Rodentia,Mammalia) del Mioceno inferior y medio del area tipo del Arago-niense (Cuenca de Calatayud–Montalban). Ph.D. dissertation, Uni-versidad Complutense de Madrid, Madrid, 676 pp.
Garcıa-Paredes, I., P. Pelaez-Campomanes, and M. A. Alvarez Sierra.2009. Gliridae (Rodentia, Mammalia) with a simple dental pattern:a new genus and new species from the European Early and MiddleMiocene. Zoological Journal of the Linnean Society 157:622–652.
International Commission on Zoological Nomenclature. 1999. Interna-tional Code of Zoological Nomenclature. International Trust forZoological Nomenclature, London, 306 pp.
Kalin, D. 1997. The mammal zonation of the upper marine molasse ofSwitzerland reconsidered: a local biozonation of MN 2–MN 5; pp.515–531 in J.-P. Aguilar, S. Legendre, and J. Michaux (eds.), Actesdu Congres BiochroM’97. Memoires et Travaux de l’Ecole pratiquedes Hautes Etudes, Institut Montpellier 21.
Kalin, D. 2003. Der Wellenberg bei Frauend—die jungsten Anteileder Oberen Susswassermolasse der Ostschweiz: biostratigraphischeDaten und tektonische Implikationen. Mitteilungen thurgauischennaturforschenden Gesellschaft 59:125–147.
Kalin, D., and O. Kempf. 2009. High-resolution stratigraphy from thecontinental record of the Middle Miocene Northern Alpine Fore-land Basin of Switzerland. Neues Jahrbuch fur Geologie und Pale-ontologie Abhandlungen 254:177–235.
Krijgsman, W., C. G. Langereis, R. Daams, and A. J. Van Der Meulen.1994. Magnetostratigraphic dating of the middle Miocene climatechange in the continental deposits of the Aragonian type area in theCalatayud-Teruel basin (Central Spain). Earth and Planetary Sci-ence Letters 128:513–526.
Krijgsman, W., M. Garces, C. G. Langereis, R. Daams, J. A. van Dam, A.J. Van Der Meulen, J. Agustı, and L. Cabrera. 1996. A new chronol-ogy for the middle to late Miocene continental record in Spain.Earth and Planetary Science Letters 142:367–380.
Lopez Guerrero, P., M. A. Alvarez-Sierra, R. Lopez-Antonanzas,A. Oliver Perez, and P. Pelaez-Campomanes. 2008. Cricetodon-tini (Cricetidae, Rodentia) del Aragoniense superior (MiocenoMedio) de Nombrevilla 2 (Zaragoza, Espana). PalaeontologicaNova 8:259–271.
Lourens, L. J., F. J. Hilgen, J. Laskar, N. J. Shackleton, and D. Wilson.2004. The Neogene Period; pp. 409–440 in F. M. Gradstein, J. G.Ogg, and A. G. Smith (eds.), A Geological Time Scale 2004. Cam-bridge University Press, Cambridge, U.K.
Mayr, H. 1979. Gebissmorphologische Untersuchungen an miozanenGliriden (Mammalia, Rodentia) Suddeutschlands. Ph.D. disserta-tion, Ludwig-Maximilians University, Munich, 380 pp.
Mein, P. 1975. Resultats du Groupe de Trevail des Vertebres; pp.78–81 in J. Senes (ed.), Report on activity of RCMNS Working
group (1971–1975). Regional Committee on Mediterranean Neo-gene Stratigraphy, Bratislava.
Mein, P. 1990. Updating of MN zones; pp. 73–90 in E. H. Lindsay,V. Fahlbusch, and P. Mein (eds.), European Neogene MammalChronology. Plenum Press, New York.
Muirhead, L. 1819. Mazology [sic], pp. 393–486 [pls. 353–358] in D. Brew-ster (ed.), The Edinburgh Encyclopaedia. Fourth ed. William Black-wood, Edinburgh, 13:1–744, pls. 347–371, 1830.
Oliver Perez, A., P. Lopez Guerrero, and P. Pelaez-Campomanes. 2008.Primer representante del genero Megacricetodon de la Cuencade Calatayud–Daroca (Zaragoza, Espana). Palaeontologica Nova8:317–329.
Pelaez-Campomanes, P. 2001. Revision of the Aragonian (Miocene) At-lantoxerus (Sciuridae). Journal of Paleontology 75:418–426.
Sach, V. J. 1999. Litho- und biostratigraphische Untersuchungen inder Oberen Sußwassermolasse des Landkreises Biberach a. d.Riß (Oberschwaben). Stuttgarter Beitrage zur Naturkunde Serie B276:1–167.
SPSS. 2002. Statistical Package for the Social Sciences for Windows, Rel.11.5.1. SPSS Inc., Chicago.
SPSS. 2007. Statistical Package for the Social Sciences for Windows, Rel.16.0. SPSS Inc., Chicago.
Uhlig, U. 2001. The Gliridae (Mammalia) from the Oligocene (MP24) ofGroben 3 in the folded Molasse of southern Germany. Palaeoverte-brata 30:151–187.
Van Dam, J. A., H. Abdul Aziz, M. A. Alvarez Sierra, F. J. Hilgen, L.W. van den Hoek Ostende, L. J. Lourens, P. Mein, A. J. van derMeulen, and P. Pelaez-Campomanes. 2006. Long-period astronomi-cal forcing of mammal turnover. Nature 443:687–691.
Van Den Hoek Ostende, L. W., M. Furio, and I. Garcıa-Paredes. 2009.New data on Paenelimnoecus from the Middle Miocene of Spain insupport of the subfamily Allosoricinae (Soricidae, Mammalia). ActaPalaeontologica Polonica 54:159–164.
Van Der Meulen, A. J., and H. de Bruijn. 1982. The mammals from theLower Miocene of Aliveri (Island of Evia, Greece). Part 2. TheGliridae. Proceedings of the Koninklijke Nederlandse AkademieVan Wetenschappen Series B 85:485–524.
Van Der Meulen, A. J., and P. Pelaez-Campomanes. 2007. Residents andtransients in the fossil record; pp. 1–17 in S. A. Levin (ed.), Ency-clopaedia of Biodiversity. Elsevier, Amsterdam.
Van Der Meulen, A. J., P. Pelaez-Campomanes, and R. Daams. 2003. Re-vision of medium-sized Cricetidae from the Miocene of the Daroca-Villafeliche area in The Calatayud-Teruel basin (Zaragoza, Spain).Coloquios de Paleontologıa Volumen Extraordinario 1:385–441.
Van Der Meulen, A. J., P. Pelaez-Campomanes, and S. A. Levin. 2005.Age structure, residents, and transients of Miocene rodent commu-nities. The American Naturalist 165:E108–E125.
Vianey-Liaud, M. 1994. La radiation des Gliridae (Rodentia) al’Eocene superieur en Europe Occidentale et sa descendanceOligocene. Munchner Geowissenschaftliche Abhandlungen A 26:117–160.
Vianey-Liaud, M. 2003. Gliridae (Mammalia, Rodentia) de l’Oligoceneeuropeen: origine de trois genres miocenes. Coloquios de Paleon-tologıa Volumen Extraordinario 1:669–698.
Werner, J. 1994. Beitrage zur Biostratigraphie der Unteren Sußwasser-Molasse Suddeutschlands—Rodentia und Lagomorpha (Mam-malia) aus den Fundstellen der Ulmer Gegend. Stuttgarter Beitragezur Naturkunde Serie B 200:1–263.
Wu, W. 1986. The Aragonian vertebrate fauna of Xiacaowan, Jiansu—4.Gliridae (Rodentia, Mammalia). Vertebrata Palasiatica 24:32–42.
Wu, W. 1990. Die Gliriden (Mammalia, Rodentia) aus der Oberen Suss-wasser Molasse von Puttenhausen (Niederbayern). Mitteilungen derBayerischen Staatssamlung fur Palaontologie und historische Ge-ologie 30:65–105.
Wu, W. 1993. Neue Gliridae (Rodentia, Mammalia) aus untermiozanen(orleanischen) Spaltenfullungen Suddeutschlands. Documenta Nat-urae 81:1–149.
Zachos, J., M. Pagani, L. Sloan, E. Thomas, and K. Billups. 2001. Trends,rhythms, and aberrations in global climate 65 Ma to present. Science292:686–693.
Submitted November 23, 2009; accepted May 14, 2010.
Downloaded By: [García-Paredes, Israel][Museo Nal Ciencias Naturales] At: 08:47 16 September 2010