
Microbial Metabolism and Community Structure in Response to Bioelectrochemically Enhanced...
9
Microbial Metabolism and Community Structure in Response to Bioelectrochemically Enhanced Remediation of Petroleum Hydrocarbon-Contaminated Soil Lu Lu, † Tyler Huggins, † Song Jin, ‡ Yi Zuo, § and Zhiyong Jason Ren* ,† † Department of Civil, Environmental, and Architectural Engineering, University of Colorado Boulder, Boulder, Colorado 80309, United States ‡ Department of Civil and Architectural Engineering, University of Wyoming, Laramie, Wyoming 82071, United States § Chevron Energy Technology Company, San Ramon, California 94583, United States * S Supporting Information ABSTRACT: This study demonstrates that electrodes in a bioelectrochemical system (BES) can potentially serve as a nonexhaustible electron acceptor for in situ bioremediation of hydrocarbon contaminated soil. The deployment of BES not only eliminates aeration or supplement of electron acceptors as in contemporary bioremediation but also significantly shortens the remediation period and produces sustainable electricity. More interestingly, the study reveals that microbial metabolism and community structure distinctively respond to the bioelectrochemically enhanced remediation. Tubular BESs with carbon cloth anode (CCA) or biochar anode (BCA) were inserted into raw water saturated soils containing petroleum hydrocarbons for enhancing in situ remediation. Results show that total petroleum hydrocarbon (TPH) removal rate almost doubled in soils close to the anode (63.5-78.7%) than that in the open circuit positive controls (37.6-43.4%) during a period of 64 days. The maximum current density from the BESs ranged from 73 to 86 mA/m 2 . Comprehensive microbial and chemical characterizations and statistical analyses show that the residual TPH has a strongly positive correlation with hydrocarbon-degrading microorganisms (HDM) numbers, dehydrogenase activity, and lipase activity and a negative correlation with soil pH, conductivity, and catalase activity. Distinctive microbial communities were identified at the anode, in soil with electrodes, and soil without electrodes. Uncommon electrochemically active bacteria capable of hydrocarbon degradation such as Comamonas testosteroni, Pseudomonas putida, and Ochrobactrum anthropi were selectively enriched on the anode, while hydrocarbon oxidizing bacteria were dominant in soil samples. Results from genus or phylum level characterizations well agree with the data from cluster analysis. Data from this study suggests that a unique constitution of microbial communities may play a key role in BES enhancement of petroleum hydrocarbons biodegradation in soils. ■ INTRODUCTION Petroleum hydrocarbon contamination in soil and groundwater is a widespread environmental problem, especially in areas with high industrial activities. 1 Remediation of hydrocarbon contaminants using in situ bioremediation, such as biosparging, biostimulation, and bioaugmentation, is usually considered cost-effective and environmentally friendly compared to physical and chemical remedies. 2,3 However, the performance of bioremediation is primarily governed by the interplay between the abundance of electron acceptor (e.g., nitrate, sulfate, Fe(III), or oxygen) 4,5 and hydrocarbon contaminant as the electron donor and carbon source for microorganisms. A common challenge for bioremediation is the limitation of electron acceptor availability, especially in the contaminated subsurface where natural electron acceptors are usually depleted. The delivery of oxygen or alternative electron acceptors such as nitrate could be limited by matrix permeability and require relatively high energy and material input. Alternative electron acceptors may also cause secondary contaminations depending on specific site conditions. 6 It was recently discovered that electrodes can be used as the electron acceptor by electrochemically active bacteria, also known as exoelectrogens, electricigens, or anode-respiring bacteria 5,7,8 during their anaerobic oxidation of substrates. In recently developed bioelectrochemical systems (BESs), 8 such as “microbial fuel cells” (MFCs), the electrons collected on the anode are transferred to the air-cathode for oxygen reduction, Received: December 29, 2013 Revised: February 26, 2014 Accepted: March 5, 2014 Article pubs.acs.org/est © XXXX American Chemical Society A dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX-XXX
Transcript of Microbial Metabolism and Community Structure in Response to Bioelectrochemically Enhanced...

Microbial Metabolism and Community Structure in Response toBioelectrochemically Enhanced Remediation of PetroleumHydrocarbon-Contaminated SoilLu Lu,† Tyler Huggins,† Song Jin,‡ Yi Zuo,§ and Zhiyong Jason Ren*,†
†Department of Civil, Environmental, and Architectural Engineering, University of Colorado Boulder, Boulder, Colorado 80309,United States‡Department of Civil and Architectural Engineering, University of Wyoming, Laramie, Wyoming 82071, United States§Chevron Energy Technology Company, San Ramon, California 94583, United States
*S Supporting Information
ABSTRACT: This study demonstrates that electrodes in abioelectrochemical system (BES) can potentially serve as anonexhaustible electron acceptor for in situ bioremediation ofhydrocarbon contaminated soil. The deployment of BES notonly eliminates aeration or supplement of electron acceptors asin contemporary bioremediation but also significantly shortensthe remediation period and produces sustainable electricity.More interestingly, the study reveals that microbial metabolismand community structure distinctively respond to thebioelectrochemically enhanced remediation. Tubular BESswith carbon cloth anode (CCA) or biochar anode (BCA)were inserted into raw water saturated soils containingpetroleum hydrocarbons for enhancing in situ remediation.Results show that total petroleum hydrocarbon (TPH) removal rate almost doubled in soils close to the anode (63.5−78.7%)than that in the open circuit positive controls (37.6−43.4%) during a period of 64 days. The maximum current density from theBESs ranged from 73 to 86 mA/m2. Comprehensive microbial and chemical characterizations and statistical analyses show thatthe residual TPH has a strongly positive correlation with hydrocarbon-degrading microorganisms (HDM) numbers,dehydrogenase activity, and lipase activity and a negative correlation with soil pH, conductivity, and catalase activity. Distinctivemicrobial communities were identified at the anode, in soil with electrodes, and soil without electrodes. Uncommonelectrochemically active bacteria capable of hydrocarbon degradation such as Comamonas testosteroni, Pseudomonas putida, andOchrobactrum anthropi were selectively enriched on the anode, while hydrocarbon oxidizing bacteria were dominant in soilsamples. Results from genus or phylum level characterizations well agree with the data from cluster analysis. Data from this studysuggests that a unique constitution of microbial communities may play a key role in BES enhancement of petroleumhydrocarbons biodegradation in soils.
■ INTRODUCTION
Petroleum hydrocarbon contamination in soil and groundwateris a widespread environmental problem, especially in areas withhigh industrial activities.1 Remediation of hydrocarboncontaminants using in situ bioremediation, such as biosparging,biostimulation, and bioaugmentation, is usually consideredcost-effective and environmentally friendly compared tophysical and chemical remedies.2,3 However, the performanceof bioremediation is primarily governed by the interplaybetween the abundance of electron acceptor (e.g., nitrate,sulfate, Fe(III), or oxygen)4,5 and hydrocarbon contaminant asthe electron donor and carbon source for microorganisms. Acommon challenge for bioremediation is the limitation ofelectron acceptor availability, especially in the contaminatedsubsurface where natural electron acceptors are usuallydepleted. The delivery of oxygen or alternative electron
acceptors such as nitrate could be limited by matrixpermeability and require relatively high energy and materialinput. Alternative electron acceptors may also cause secondarycontaminations depending on specific site conditions.6
It was recently discovered that electrodes can be used as theelectron acceptor by electrochemically active bacteria, alsoknown as exoelectrogens, electricigens, or anode-respiringbacteria5,7,8 during their anaerobic oxidation of substrates. Inrecently developed bioelectrochemical systems (BESs),8 such as“microbial fuel cells” (MFCs), the electrons collected on theanode are transferred to the air-cathode for oxygen reduction,
Received: December 29, 2013Revised: February 26, 2014Accepted: March 5, 2014
Article
pubs.acs.org/est
© XXXX American Chemical Society A dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX−XXX

and at the same time electric current can be produced.7 Such anapproach is hypothesized to stimulate and enhance hydro-carbon degradation accompanied by energy production,because the electrode pair serves as a nonexhaustible electronacceptor, which eliminates the necessity of aeration in thesubsurface but sustains an aerobic-like metabolic pathway onthe cathode. Previous studies showed the feasibility of using anelectrode approach for the removal of different hydrocarbons,such as crude oil,9 diesel,4 refinery wastewater,10 petroleumsludge,11 and aromatic compounds5,12 through direct exoelec-trogenic oxidation5 or syntrophic interactions4 among microbialconsortia. However, most studies were conducted usingsynthetic hydrocarbon substrates in sediment or groundwater,which are very different from actual soil contaminationconditions due to the great variety in soil composition,permeability, and water content.13−17 For example, our earlystudies show that a lab scale two-chamber MFC reactorincreased diesel carbon degradation by 165% (from 31% to82%) than the open circuit control within 21 days ofoperation.4 Another study demonstrated that the influenceradius of the anode of a U-tube MFC and soil moisturesignificantly affected the degradation rate, and the totalpetroleum hydrocarbon (TPH) removal was enhanced by120% in saturated condition (33% water content) and close tothe anode (<1 cm).9 While these studies demonstrated theengineering feasibility, to our best knowledge there has been noreport on the microbial metabolic activity and communitystructure and their changes during the bioelectrochemicallyenhanced remediation. Such understanding is important,because it helps reveal the relationships between the twoconcurrent degradation activities, natural attenuation using thenative electron acceptor in the soil and exoelectrogen consortiataking advantage of the electrode(s) as the electron acceptor,and consequently to better exploit appropriate remediationstrategies.In this study, we developed BESs with tubular configuration
for remediation, which was designed for its future installation inthe groundwater piezometers and monitoring wells. We usedpetroleum hydrocarbon contaminated soil and comparedsystem performance between two anode materials (biocharand carbon cloth). Biochar has been used as soil amendmentsto improve water and nutrient retention and recently served aselectrode materials in microbial fuel cells.18 Moreover, for thefirst time we closely tracked the changes of microbialcommunity structure and quantified the hydrocarbon-degradingmicroorganisms (HDM) numbers and relevant enzymeactivities. We correlated the microbial and enzymaticinformation with the soil chemical and biological parametersand revealed how microbial metabolism as well as anodebiofilm and indigenous microbial communities responded tothe bioelectrochemically enhanced remediation of petroleumhydrocarbon in soil.
■ MATERIALS AND METHODSSoil Sample Collection and Characterization. Soil
samples were collected near Lincoln, Nebraska, USA. Thesampling location has a known history of diesel and engine oilcontamination. Contaminated soil samples with TPH concen-tration of 11.46 g/kg-dry soil (DS) as well as uncontaminatedsoil samples from surrounding areas were taken from 5 to 10cm below ground surface and transported on ice to the labovernight. After being partially air-dried, soil samples weresieved to <2 mm to remove plant roots and rocks and stored at
4 °C before use. The main characteristics of the soil samples aresummarized in Supporting Information (SI) Table S1. The soilwas saturated (immersed) with deionized water during regularexperiments without adding buffer solution or additionalhydrocarbons.
Soil Tubular BES Construction and Operation. TubularBESs were constructed by wrapping an assembly of anode,separator, and cathode layers around a perforated PVC tube (L20 cm × D 3.5 cm), with the cathode layer facing inside andexposed to air, and the anode facing outside exposed to soil (SIFigure S1). Activated carbon cloth (Chemviron Carbon Ltd.,PA) with a Pt/C catalyst layer (0.1 mg Pt/cm2) and four PTFEdiffusion layers19 on carbon support was served as the aircathode, and the effective cathode projected area was 110 cm2.The anode was either made of carbon cloth (11 cm × 12 cm,Fuel Cell Earth LLC, MA) or biochar, which was manufacturedin our lab as described in Huggines et al.18 and the SI. Fourlayers of nonconductive permeable glass fiber were sandwichedbetween the electrodes as the separator to prevent short circuit.The carbon cloth anode (CCA) was wrapped together withseparators and the cathode, while the biochar anode (BCA) wasconstructed by packing biochar particles in between theseparator layer and an outermost layer of nylon mesh, withan average thickness of 1 cm (Figure S1B). A graphite rod (L 6cm × D 0.5 cm) was inserted into the biochar pack (stackingvolume of 168 cm3) as a current collector. The anode andcathode were connected using a 100 Ω external resistor.The tubular BESs were inserted into 15 cm (H) × 16 cm
(D) buckets containing saturated hydrocarbon-contaminatedsoil. The top of each bucket was covered to prevent evaporationof water and volatile hydrocarbons, and water loss was made upby deionized water to maintain the saturation condition. In onecarbon cloth reactor, a nonionic surfactant Triton X-100 (500mg/L) was mixed with soil (CCA-S) to examine if the additionof surfactant would improve desorption and transfer ofhydrocarbons from soil toward the anode reaction zone.20
Table S2 (SI) summarizes the abbreviations of the differentreactor setups and sample conditions. Soil samples wereroutinely extracted from the top, middle, and bottom of the soillayer in each sampling event in triplicates, and the compositesoil samples in vertical orientation were used for chemical andbiological analyses. Noncontaminated soil (NCS) obtainedfrom pristine background and original hydrocarbon contami-nated soil (OHCS) were also be sampled from the site foranalyses. Anode biofilms and soil samples collected from CCA,BCA, and CCA-S reactors were used for bacterial communityanalysis. Indigenous microbes remained in all systems withoutany external inoculation. All reactors were operated at roomtemperature of 20−22 °C without light irradiation.
Chemical, Biological, and Electrochemical Analyses.All soil samples were characterized for total petroleumhydrocarbon (TPH) by following the EPA Method 8015D21
and MA EPH.22 Briefly, 5 g of the soil sample was blended withanhydrous Na2SO4 until the soil flows freely, then the mixturewas mixed with 10 mL of dichloromethane followed byextraction using vortex and sonication, each 5 min. Thesuspension was centrifuged at 8,000 × g for 5 min, and thesupernatant was decanted. This procedure was repeated, andthe combined extractants were concentrated to 1 mL andpurified by using silica gel. TPH was measured using a ThermoTRACE gas chromatograph equipped with a Restek Rxi-1 mscolumn (20 m × 0.18 mm ID, 0.18 μm film thickness) andflame ionization detector. Aliphatic hydrocarbon calibration
Environmental Science & Technology Article
dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX−XXXB
standards were prepared by diluting an n-alkane standardmixture (C8−C40, Sigma-Aldrich Co. Ltd.) at five concen-trations. The calibration factor (CF) is the ratio of GC peakarea response to the mass of injected analyte. TPH was definedas the collective concentration of all hydrocarbon compoundseluting from n-nonane (C9) to n-hexatriacontane (C36)22 andwas calculated using a collective CF of all component standardsin that fraction (C9−C36 n-alkane, 28 components) at differentcalibration standard concentrations.A method with a combination of 96-well microtiter plate
most probable number (MPN) procedure and positiveidentification by iodonitrotetrazolium violet (INT) reductionwas used to enumerate the hydrocarbon-degrading micro-organisms (HDM) using number 2 fuel oil as the selectivegrowth substrate.23 Soil analytical results were reported on adry-weight basis. The pH and conductivity of soil weremeasured in a 1:5 (w/v) soil:deionized water mixture. Soilparticle-size distribution was evaluated using the methoddescribed by Kettler et al. (2001).24 The soil organic carbon(SOC) was determined by the Walkley-Black procedure,25 andsoil organic matter (SOM) was measured by the mass loss onignition.26 Available nitrate, phosphate, sulfate, and ammonianitrogen in soil were extracted as described by Mussa et al.(2009)27 and determined according to standard methods.28
Voltage (U) across the external resistor (R = 100 Ω) wasrecorded every minute by using a data acquisition system(model 2700, Keithley, Inc.). Current density (J, mA/m2) andpower density (P, mW/m2) were normalized to the effectivecathode projected area (A = 0.011 m2) and calculated as J = U/RA and P = JU, respectively.29 A potentiostat (PC4/300,Gamry Instruments, NJ) was used to perform linear sweepvoltammetry (LSV) (0.1 mV/s scan rate) for measurements ofpolarization curve, which was transformed to power curves toobtain maximum power density.Soil Enzyme Activity Characterization and Bacterial
Community Structure Analysis. Soil dehydrogenase activitywas measured by an improved method of triphenyltetrazoliumchloride (TTC) reduction30 and was expressed as μg TPF-reduction/g-DS using a TPF calibration curve. Soil catalaseactivity was measured by back-titrating residual H2O2 withKMnO4
31 and was expressed as mg H2O2-consumption/g-DS.Soil lipase activity was determined based on standard assays oftitration32 and was expressed as mL KOH-consumption/g-DS.One blank control experiment was conducted for each soilsample and enzyme activity characterization. Details aboutanalyses of soil enzyme activities can be found in the SI.The bacterial community structure was analyzed by using
high-throughput 454 GS-FLX pyrosequencing of the 16SrRNA. Soil samples were collected from the field samplinglocations as well as 1 or 5 cm from the anodes in active andopen circuit control reactors. Biofilm samples were also takenfrom the anode electrodes of all reactors. The pyrosequencingand bioinformatics analysis were carried out according topreviously described methods,33,34 and the details are providedin the SI. Raw sequencing data were deposited to the NCBISequence Read Archive (SRA) with accession No. SRP026155.Statistical Data Analysis. All analyses described were
conducted in triplicates, and the means were reported. Pearsonproduct-moment correlation coefficients between two soilparameters were analyzed by SPSS 16.0 software. Correlationswere considered significant at P < 0.05.
■ RESULTS AND DISCUSSIONCurrent Production Profiles from Different BESs.
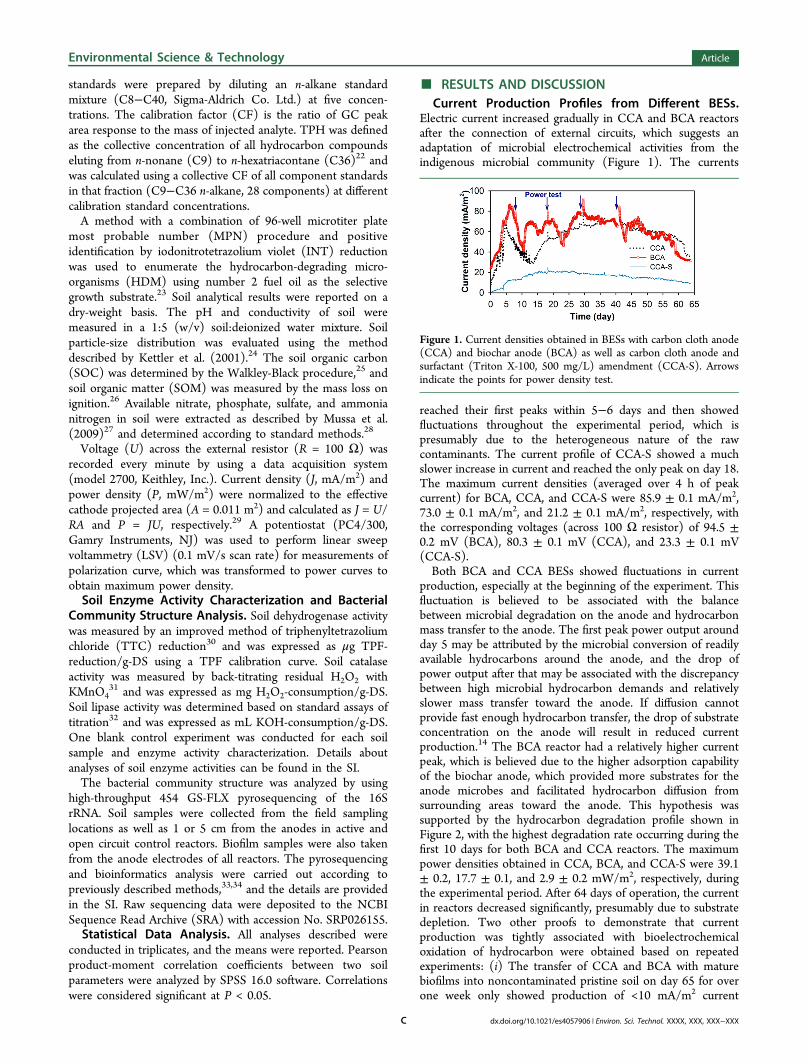
Electric current increased gradually in CCA and BCA reactorsafter the connection of external circuits, which suggests anadaptation of microbial electrochemical activities from theindigenous microbial community (Figure 1). The currents
reached their first peaks within 5−6 days and then showedfluctuations throughout the experimental period, which ispresumably due to the heterogeneous nature of the rawcontaminants. The current profile of CCA-S showed a muchslower increase in current and reached the only peak on day 18.The maximum current densities (averaged over 4 h of peakcurrent) for BCA, CCA, and CCA-S were 85.9 ± 0.1 mA/m2,73.0 ± 0.1 mA/m2, and 21.2 ± 0.1 mA/m2, respectively, withthe corresponding voltages (across 100 Ω resistor) of 94.5 ±0.2 mV (BCA), 80.3 ± 0.1 mV (CCA), and 23.3 ± 0.1 mV(CCA-S).Both BCA and CCA BESs showed fluctuations in current
production, especially at the beginning of the experiment. Thisfluctuation is believed to be associated with the balancebetween microbial degradation on the anode and hydrocarbonmass transfer to the anode. The first peak power output aroundday 5 may be attributed by the microbial conversion of readilyavailable hydrocarbons around the anode, and the drop ofpower output after that may be associated with the discrepancybetween high microbial hydrocarbon demands and relativelyslower mass transfer toward the anode. If diffusion cannotprovide fast enough hydrocarbon transfer, the drop of substrateconcentration on the anode will result in reduced currentproduction.14 The BCA reactor had a relatively higher currentpeak, which is believed due to the higher adsorption capabilityof the biochar anode, which provided more substrates for theanode microbes and facilitated hydrocarbon diffusion fromsurrounding areas toward the anode. This hypothesis wassupported by the hydrocarbon degradation profile shown inFigure 2, with the highest degradation rate occurring during thefirst 10 days for both BCA and CCA reactors. The maximumpower densities obtained in CCA, BCA, and CCA-S were 39.1± 0.2, 17.7 ± 0.1, and 2.9 ± 0.2 mW/m2, respectively, duringthe experimental period. After 64 days of operation, the currentin reactors decreased significantly, presumably due to substratedepletion. Two other proofs to demonstrate that currentproduction was tightly associated with bioelectrochemicaloxidation of hydrocarbon were obtained based on repeatedexperiments: (i) The transfer of CCA and BCA with maturebiofilms into noncontaminated pristine soil on day 65 for overone week only showed production of <10 mA/m2 current
Figure 1. Current densities obtained in BESs with carbon cloth anode(CCA) and biochar anode (BCA) as well as carbon cloth anode andsurfactant (Triton X-100, 500 mg/L) amendment (CCA-S). Arrowsindicate the points for power density test.
Environmental Science & Technology Article
dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX−XXXC
densities in both BESs. (ii) Current density rebounded to amaximum of 135 ± 3 mA/m2 after 21.44 g-TPH/kg-DS ofdiesel being added into noncontaminated soil in a BCA reactoron day 75 (SI Figure S4). Surfactant amendment in soil had anegative effect on the electricity production in BES, as shownby the CCA-S profile. The desorption test (SI Figure S2)indicates that the dose of surfactant used in this study (500 mg/L) appears to increase the surfactant sorption onto soil, whichresulted in increased TPH partitioning onto soil particles. Theenhancement of phase transfer of hydrophobic hydrocarbonsfrom the pseudoaqueous to the soil-adsorbed phase decreasedthe availability of substrates for anode utilization and reducedthe performance of CCA-S BES.
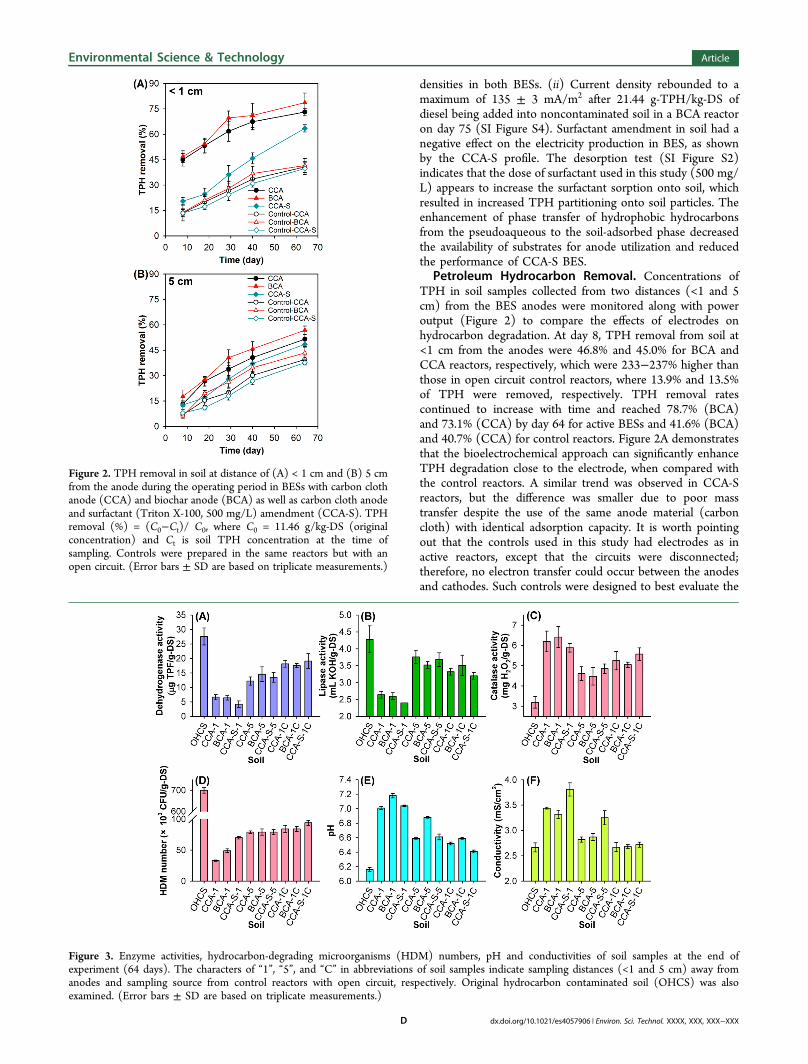
Petroleum Hydrocarbon Removal. Concentrations ofTPH in soil samples collected from two distances (<1 and 5cm) from the BES anodes were monitored along with poweroutput (Figure 2) to compare the effects of electrodes onhydrocarbon degradation. At day 8, TPH removal from soil at<1 cm from the anodes were 46.8% and 45.0% for BCA andCCA reactors, respectively, which were 233−237% higher thanthose in open circuit control reactors, where 13.9% and 13.5%of TPH were removed, respectively. TPH removal ratescontinued to increase with time and reached 78.7% (BCA)and 73.1% (CCA) by day 64 for active BESs and 41.6% (BCA)and 40.7% (CCA) for control reactors. Figure 2A demonstratesthat the bioelectrochemical approach can significantly enhanceTPH degradation close to the electrode, when compared withthe control reactors. A similar trend was observed in CCA-Sreactors, but the difference was smaller due to poor masstransfer despite the use of the same anode material (carboncloth) with identical adsorption capacity. It is worth pointingout that the controls used in this study had electrodes as inactive reactors, except that the circuits were disconnected;therefore, no electron transfer could occur between the anodesand cathodes. Such controls were designed to best evaluate the
Figure 2. TPH removal in soil at distance of (A) < 1 cm and (B) 5 cmfrom the anode during the operating period in BESs with carbon clothanode (CCA) and biochar anode (BCA) as well as carbon cloth anodeand surfactant (Triton X-100, 500 mg/L) amendment (CCA-S). TPHremoval (%) = (C0−Ct)/ C0, where C0 = 11.46 g/kg-DS (originalconcentration) and Ct is soil TPH concentration at the time ofsampling. Controls were prepared in the same reactors but with anopen circuit. (Error bars ± SD are based on triplicate measurements.)
Figure 3. Enzyme activities, hydrocarbon-degrading microorganisms (HDM) numbers, pH and conductivities of soil samples at the end ofexperiment (64 days). The characters of “1”, “5”, and “C” in abbreviations of soil samples indicate sampling distances (<1 and 5 cm) away fromanodes and sampling source from control reactors with open circuit, respectively. Original hydrocarbon contaminated soil (OHCS) was alsoexamined. (Error bars ± SD are based on triplicate measurements.)
Environmental Science & Technology Article
dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX−XXXD

performance of BES; however, the presence of electrodeswithout complete circuits has been shown to enhancebiodegradation of contaminants compared to natural attenu-ation without electrode insert.5,14 Therefore the controls in thisstudy probably established an elevated baseline that wouldunderestimate the performance of BESs.The TPH removal rates decreased with the distance from the
anode (Figure 2B). At 5 cm from the anode, the TPH removalon day 8 was 17.7% (BCA) and 13.2% (CCA), respectively,and on day 64, the removal increased to 56.8% (BCA) and51.5% (CCA). The influence of the electrodes decreases alongwith distance, due to less microbial electrochemical activitiesand pollutant mass transfer. However, the TPH degradationrates in active BESs were consistently 31−37% higher thanthose in the control reactors, indicating the positive influence ofBES on degradation even further from the electrodes. Theactual measurement of the radius of influence (ROI) for theBESs was limited to the configuration of the lab reactor;however, theoretical ROIs would be extrapolated based on thedata. Actual ROIs and other engineering parameters areexpected to be obtained from a field pilot. Hydrocarbonresidues of all BES anodes were less than 15% of TPH in soilsclose to these electrodes at the end of the test, indicating thatthe majority of contaminants were biodegraded instead of beingadsorbed by the electrodes.Changes and Correlations of Soil Chemical and
Biological Characteristics. Soil enzyme activities, hydro-carbon-degrading microorganisms (HDM) counts, pH, andconductivities are key parameters to confirming and quantifyingmicrobial catalyzed TPH removal (Figure 3). Figure 3A and 3Bshow that at the end of the experiment (day 64), the activitiesof dehydrogenase and lipase close to the active anode (CCA-1,BCA-1, and CCA-S-1) were only 15−62% of the originalhydrocarbon contaminated soil (OHCS), and the disconnectedcontrol reactors (CCA-1C, BCA-1C, and CCA-S-1C) alsoshowed an 18−36% decrease due to biodegradation at thepresence of BES hardware. The 5 cm samples showed a 12−56% drop from the OHCS. Dehydrogenase and lipase aremajor microbial catalysts that are directly involved in organicmatter decomposition and xenobiotics detoxification,35 andtheir activity decline correlated to less microbial metabolismwhen hydrocarbons had been degraded. This was supported bystatistical analysis of correlations between enzyme activities andTPH concentrations (Figure 2), although no solid proof
showed that dehydrogenase and lipase are specific functionalenzymes directly involved in hydrocarbon degradation in thisstudy. Many studies on traditional bioremediation of hydro-carbon contaminated soil also observed that dehydrogenase andlipase activities increase temporarily in response to freshhydrocarbon contamination and decline rapidly after thedepletion of the carbon source.35−37 Different from dehydro-genase or lipase, catalase activities significantly increased by84−101% for soils close to the anodes in the active BESs whencompared to OHCS (Figure 3C). This is consistent with otherstudies that the activity of catalase is inhibited by hydrocarbonsand therefore negatively correlated with hydrocarbon concen-tration in soil.37 The HDM number in different soil samplesalso reflected the changes of TPH concentration, especially theresidual TPH content. As shown in Figure 3D, HDM of CCA-1was 33 × 103 CFU/g-DS, which was 4.7% of the initial HDM inthe OHCS. The HDM numbers in other samples alsodemonstrated the similar trend, which is in agreement withprevious studies that the HDM number usually drops whensubstrate concentration is low,36 even though immediately afterthe substrate depletion there might be a spike in HDM due totemporary reserves in the cells.9 The decreased number ofHDM is also consistent with the reduced activities ofdehydrogenase and lipase. However, it should be noted thatthe aerobic HDM numbers may not reveal anaerobic microbialcommunities, such as Deltaproteobacteria and Fermicutesdetected from high throughput sequencing. There are alsospecific enzymes critical for anaerobic degradation of hydro-carbons or metabolites were not characterized due toexperiment limitation.The soil pH increased from 6.16 at the beginning of the
experiment to 6.59−7.18 at the end of the study (Figure 3E).Such increase could be attributed to the depletion of existingorganic acids produced from indigenous biodegradation ofhydrocarbon in aged soil by electrochemically active bacteria onthe anode. Another possible reason for pH increase might bebicarbonate accumulation resulting from biodegradation ofhydrocarbon. In addition, the membrane-less tubular config-uration of the electrode assembly in the saturated soil conditionprovided a good environment for proton and ion transfer, andno proton accumulation was observed. A similar trend wasobserved for conductivity, as shown in Figure 3F. The soilconductivities in active BESs increased by 6−43% but remainedconsistent in the controls. This was probably caused by the
Table 1. Correlation Matrix (Coefficient and Significance Level) between Soil TPH, pH, Conductivity and Soil BiologicalParametersa
TPH pH conductivity HDM dehydrogenase activity catalase activity
pH −0.940 /0.000d
conductivity −0.679 0.774 /0.031b 0.009c
HDM 0.837 −0.651 −0.365 /0.003c 0.042b 0.299
dehydrogenase activity 0.940 −0.928 −0.853 0.725 /0.000d 0.000d 0.002c 0.018b
catalase activity −0.847 0.763 0.612 −0.764 −0.797 /0.002c 0.010b 0.060 0.010b 0.006c
lipase activity 0.819 −0.835 −0.759 0.634 0.841 −0.9360.004c 0.003c 0.011b 0.049b 0.002c 0.000d
aCorrelation: Strong (positive, 0.5 to 1.0 or negative, −1.0 to −0.5). Medium (positive, 0.3 to 0.5 or negative, −0.5 to −0.3). Small (positive, 0.1 to0.3 or negative, −0.3 to −0.1). None (positive, 0.0 to 0.09 or negative, −0.09 to −0.0). Sample number = 30. bP < 0.05. cP < 0.01. dP < 0.001.
Environmental Science & Technology Article
dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX−XXXE

formation of ionic species such as intermediate compoundsduring the biodegradation pathways, resulting in an increase ofconductivity in soil.38
Statistically, the parameters analyzed clearly indicate that thecloser the soil is to the active BESs anode, the higher microbialhydrocarbon degradation activities and faster removal of TPHwould occur. As summarized in Table 1, the concentration ofTPH negatively correlated with the soil pH (P < 0.001),conductivity (P < 0.05), and catalase activity (P < 0.01) butpositively correlated with hydrocarbon-degrading microorgan-isms (HDM) numbers (P < 0.01), dehydrogenase activity (P <0.001), and lipase activity (P < 0.01). Such correlations,accompanied by BES current detection and TPH degradation,provide a comprehensive profile on how BES enhanceshydrocarbon remediation, harvests electrical energy, and shapesthe ecological restoration of the soil.Differences of Bacterial Community Structures and
Bacterial Phylogenetic Identification. One of the keyfindings in this study is the shifts of microbial communitystructure during the BES-enhanced degradation of petroleumhydrocarbons. Using high-throughput culture-independent 454GS-FLX pyrosequencing, different bacterial communities wereidentified in the soils and anode biofilms. From thirteengenerated libraries, a total of 122,143 high-quality 16S rRNAgene sequences with an average length of 464 bp were obtained(SI Table S3). These sequences were assigned into 35,460 and28,878 OTUs with a distance limit of 0.03 and 0.05,respectively. Although the rarefaction curves did not reach aplateau (SI Figure S3), the Good’s Coverage estimatorsindicated that the sizes of libraries were sufficient to cover60−92% of the bacterial communities (Table S3). TheShannon diversity index (H′) includes the information ofrichness (i.e., the number of species present) and how theabundance of each species is distributed (the evenness of thespecies). Noncontaminated soil obtained from the site showedthe highest biodiversity (H′ = 7.96 at 0.03 OTU distance)(Table S3) among all communities, while the H′ was smaller incontaminated soil (H′ = 6.99), consistent with the trendpreviously reported.39,40 The diversity in soils decreased to anaverage of 6.62 for active BESs and 6.15 for the disconnectedcontrol reactors. The biofilms on the BES anodes were of lowbiodiversities (average H′ = 5.05), suggesting a selectiveenrichment of specific communities during the biodegradationof petroleum hydrocarbons.Distinct clusters of bacterial communities were presented by
Weighted Fast UniFrac Principal Coordinates Analysis (PCoA)in Figure 4. Thirteen different community structures werefound forming three distinct clusters: (a) bacterial groups onthe anode biofilms (CCA, BCA, and CCA-S); (b) groups insoils including noncontaminated and contaminated soil (NCSand OHCS); and (c) groups in soils that associated with BES,including those from active reactors (CCA-1 to CCA-S-5) andthose from the control reactors (BCA-1C and CCA-S-1C)(Figure 4A). This cluster analysis results well match thefunctional data based microbial community distribution. Thebacterial community structure associated with electrochemicalelectron transfer on the anode (lower left cluster) is very distantfrom the communities in soil despite their same origin. Bacteriain soil associated with electrode enhanced remediation areclustered together (top right cluster) but are very different fromthe communities in pristine soils (lower right cluster) regardlessof the initial contamination level. Furthermore, as shown inFigure 4B, even within the cluster of group c, the dendogram
constructed on the basis of community phylogenetic lineagesindicates that soil communities from the control reactors arehighly grouped together regardless of different reactors. Thesefindings demonstrate that even from the same origin of soilwith the same microbial consortia, the bacterial communitystructures drastically diverged after the BES remediationexperiments, associated with metabolic functions need bydifferent groups.The phylogenetic analysis of sequences revealed a high
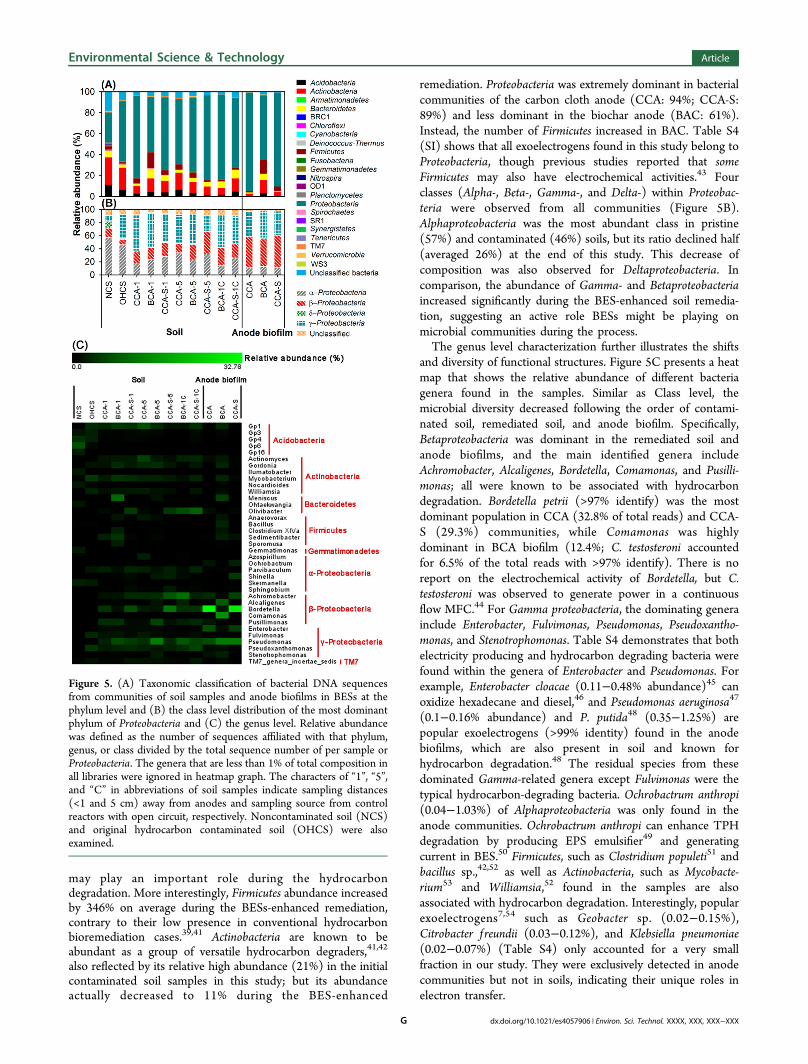
bacterial diversity in the soil samples. Twenty-two bacterialphyla were observed from thirteen libraries (Figure 5A). Theabsolute majority of sequences belonged to Proteobacteria andaccounted for 28.7−94.0% of the total reads in each bacterialcommunity. This agrees with previous studies on hydrocarboncontaminated soil bacterial communities.39,41,42 Other majorbacterial phyla included Actinobacteria, Bacteroidetes, Firmicutes,and Acidobacteria, as well as a certain amount of unclassifiedsequences at the phylum level.Bacterial species in uncontaminated soil exhibits relatively
even distribution at phylum level among all communities withProteobacteria (29%) and Actinobacteria (26%) as the two mostabundant phyla (Figure 5A). Hydrocarbon contaminationappeared to have resulted in the rise of Proteobacteriaabundance (57%), and the proportion rose even higher to anaverage of 69% after remediation, suggesting that Proteobacteria
Figure 4. Weighted Fast UniFrac (A) Principal Coordinates Analysis(PCoA) and (B) cluster of the bacterial communities of soil samplesand anode biofilms in BESs on the basis of phylogenetic lineages thatsamples contain. The characters of “1”, “5”, and “C” in abbreviations ofsoil samples indicate sampling distances (<1 and 5 cm) away fromanodes and sampling source from control reactors with open circuit,respectively. Noncontaminated soil (NCS) obtained from pristinebackground site and original hydrocarbon contaminated soil (OHCS)were also examined. (p-values for all samples together <0.001 in the ptest).
Environmental Science & Technology Article
dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX−XXXF
may play an important role during the hydrocarbondegradation. More interestingly, Firmicutes abundance increasedby 346% on average during the BESs-enhanced remediation,contrary to their low presence in conventional hydrocarbonbioremediation cases.39,41 Actinobacteria are known to beabundant as a group of versatile hydrocarbon degraders,41,42
also reflected by its relative high abundance (21%) in the initialcontaminated soil samples in this study; but its abundanceactually decreased to 11% during the BES-enhanced
remediation. Proteobacteria was extremely dominant in bacterialcommunities of the carbon cloth anode (CCA: 94%; CCA-S:89%) and less dominant in the biochar anode (BAC: 61%).Instead, the number of Firmicutes increased in BAC. Table S4(SI) shows that all exoelectrogens found in this study belong toProteobacteria, though previous studies reported that someFirmicutes may also have electrochemical activities.43 Fourclasses (Alpha-, Beta-, Gamma-, and Delta-) within Proteobac-teria were observed from all communities (Figure 5B).Alphaproteobacteria was the most abundant class in pristine(57%) and contaminated (46%) soils, but its ratio declined half(averaged 26%) at the end of this study. This decrease ofcomposition was also observed for Deltaproteobacteria. Incomparison, the abundance of Gamma- and Betaproteobacteriaincreased significantly during the BES-enhanced soil remedia-tion, suggesting an active role BESs might be playing onmicrobial communities during the process.The genus level characterization further illustrates the shifts
and diversity of functional structures. Figure 5C presents a heatmap that shows the relative abundance of different bacteriagenera found in the samples. Similar as Class level, themicrobial diversity decreased following the order of contami-nated soil, remediated soil, and anode biofilm. Specifically,Betaproteobacteria was dominant in the remediated soil andanode biofilms, and the main identified genera includeAchromobacter, Alcaligenes, Bordetella, Comamonas, and Pusilli-monas; all were known to be associated with hydrocarbondegradation. Bordetella petrii (>97% identify) was the mostdominant population in CCA (32.8% of total reads) and CCA-S (29.3%) communities, while Comamonas was highlydominant in BCA biofilm (12.4%; C. testosteroni accountedfor 6.5% of the total reads with >97% identify). There is noreport on the electrochemical activity of Bordetella, but C.testosteroni was observed to generate power in a continuousflow MFC.44 For Gamma proteobacteria, the dominating generainclude Enterobacter, Fulvimonas, Pseudomonas, Pseudoxantho-monas, and Stenotrophomonas. Table S4 demonstrates that bothelectricity producing and hydrocarbon degrading bacteria werefound within the genera of Enterobacter and Pseudomonas. Forexample, Enterobacter cloacae (0.11−0.48% abundance)45 canoxidize hexadecane and diesel,46 and Pseudomonas aeruginosa47
(0.1−0.16% abundance) and P. putida48 (0.35−1.25%) arepopular exoelectrogens (>99% identity) found in the anodebiofilms, which are also present in soil and known forhydrocarbon degradation.48 The residual species from thesedominated Gamma-related genera except Fulvimonas were thetypical hydrocarbon-degrading bacteria. Ochrobactrum anthropi(0.04−1.03%) of Alphaproteobacteria was only found in theanode communities. Ochrobactrum anthropi can enhance TPHdegradation by producing EPS emulsifier49 and generatingcurrent in BES.50 Firmicutes, such as Clostridium populeti51 andbacillus sp.,42,52 as well as Actinobacteria, such as Mycobacte-rium53 and Williamsia,52 found in the samples are alsoassociated with hydrocarbon degradation. Interestingly, popularexoelectrogens7,54 such as Geobacter sp. (0.02−0.15%),Citrobacter f reundii (0.03−0.12%), and Klebsiella pneumoniae(0.02−0.07%) (Table S4) only accounted for a very smallfraction in our study. They were exclusively detected in anodecommunities but not in soils, indicating their unique roles inelectron transfer.
Figure 5. (A) Taxonomic classification of bacterial DNA sequencesfrom communities of soil samples and anode biofilms in BESs at thephylum level and (B) the class level distribution of the most dominantphylum of Proteobacteria and (C) the genus level. Relative abundancewas defined as the number of sequences affiliated with that phylum,genus, or class divided by the total sequence number of per sample orProteobacteria. The genera that are less than 1% of total composition inall libraries were ignored in heatmap graph. The characters of “1”, “5”,and “C” in abbreviations of soil samples indicate sampling distances(<1 and 5 cm) away from anodes and sampling source from controlreactors with open circuit, respectively. Noncontaminated soil (NCS)and original hydrocarbon contaminated soil (OHCS) were alsoexamined.
Environmental Science & Technology Article
dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX−XXXG

■ ASSOCIATED CONTENT
*S Supporting InformationFour additional tables and four figures. This material is availablefree of charge via the Internet at http://pubs.acs.org.
■ AUTHOR INFORMATION
Corresponding Author*Phone: (303) 492-4137. Fax: (303) 492-7217. E-mail: [email protected].
NotesThe authors declare no competing financial interest.
■ ACKNOWLEDGMENTS
This work was supported by the Chevron Energy TechnologyCompany. We appreciate feedback from Chevron and thereviewers.
■ REFERENCES(1) Du, W.; Wan, Y.; Zhong, N.; Fei, J.; Zhang, Z.; Chen, L.; Hao, J.Status quo of soil petroleum contamination and evolution ofbioremediation. Pet. Sci. 2011, 8 (4), 502−514.(2) Das, N.; Chandran, P. Microbial degradation of petroleumhydrocarbon contaminants: an overview. Biotech. Res. Int. 2010, 2011,1−13.(3) Tyagi, M.; da Fonseca, M. M. R.; de Carvalho, C. C. C. R.Bioaugmentation and biostimulation strategies to improve theeffectiveness of bioremediation processes. Biodegradation 2011, 22(2), 231−241.(4) Morris, J. M.; Jin, S.; Crimi, B.; Pruden, A. Microbial fuel cell inenhancing anaerobic biodegradation of diesel. Chem. Eng. J. 2009, 146(2), 161−167.(5) Zhang, T.; Gannon, S. M.; Nevin, K. P.; Franks, A. E.; Lovley, D.R. Stimulating the anaerobic degradation of aromatic hydrocarbons incontaminated sediments by providing an electrode as the electronacceptor. Environ. Microbiol. 2010, 12 (4), 1011−1020.(6) Brown, R. A.; Hinchee, R. E.; Norris, R. D.; Wilson, J. T.Bioremediation of petroleum hydrocarbons: A flexible, variable speedtechnology. Rem. J. 1996, 6 (3), 95−109.(7) Logan, B. E. Exoelectrogenic bacteria that power microbial fuelcells. Nat. Rev. Microbiol. 2009, 7 (5), 375−381.(8) Wang, H.; Ren, Z. J. A comprehensive review of microbialelectrochemical systems as a platform technology. Biotechnol. Adv.2013, 31 (8), 1796−1807.(9) Wang, X.; Cai, Z.; Zhou, Q.; Zhang, Z.; Chen, C.Bioelectrochemical stimulation of petroleum hydrocarbon degradationin saline soil using U-tube microbial fuel cells. Biotechnol. Bioeng. 2012,109 (2), 426−433.(10) Morris, J. M.; Jin, S. Feasibility of using microbial fuel celltechnology for bioremediation of hydrocarbons in groundwater. J.Environ. Sci. Health, Part A: Environ. Sci. Eng. 2008, 43 (1), 18−23.(11) Mohan, S. V.; Chandrasekhar, K. Self-induced bio-potential andgraphite electron accepting conditions enhances petroleum sludgedegradation in bio-electrochemical system with simultaneous powergeneration. Bioresour. Technol. 2011, 102 (20), 9532−9541.(12) Rakoczy, J.; Feisthauer, S.; Wasmund, K.; Bombach, P.; Neu, T.R.; Vogt, C.; Richnow, H. H. Benzene and sulfide removal fromgroundwater treated in a microbial fuel cell. Biotechnol. Bioeng. 2013,110 (12), 3104−3113.(13) Morris, J. M.; Jin, S. Enhanced biodegradation of hydrocarbon-contaminated sediments using microbial fuel cells. J. Hazard. Mater.2012, 213-214 (30), 474−477.(14) Williams, K. H.; Nevin, K. P.; Franks, A.; Englert, A.; Long, P.E.; Lovley, D. R. Electrode-based approach for monitoring in situmicrobial activity during subsurface bioremediation. Environ. Sci.Technol. 2010, 44 (1), 47−54.
(15) Huang, D.-Y.; Zhou, S.-G.; Chen, Q.; Zhao, B.; Yuan, Y.;Zhuang, L. Enhanced anaerobic degradation of organic pollutants in asoil microbial fuel cell. Chem. Eng. J. 2011, 172 (2), 647−653.(16) Yuan, Y.; Zhou, S.; Zhuang, L. A new approach to in situsediment remediation based on air-cathode microbial fuel cells. J. SoilsSediments 2010, 10 (7), 1427−1433.(17) An, J.; Moon, H.; Chang, I. S. Multiphase electrode microbialfuel cell system that simultaneously converts organics coexisting inwater and sediment phases into electricity. Environ. Sci. Technol. 2010,44 (18), 7145−7150.(18) Huggins, M. T.; Wang, H.; Kearns, J.; Jenkins, P.; Ren, Z. J.Biochar as a sustainable electrode material for electricity production inmicrobial fuel cells. Bioresour. Technol. 2014, 157, 114−119.(19) Wang, H.; Davidson, M.; Zuo, Y.; Ren, Z. Recycled tire crumbrubber anodes for sustainable power production in microbial fuel cells.J. Power Sources 2011, 196 (14), 5863−5866.(20) Sahoo, D.; Smith, J. A. Enhanced trichloroethene desorptionfrom long-term contaminated soil using Triton X-100 and pHincreases. Environ. Sci. Technol. 1997, 31 (7), 1910−1915.(21) U.S. Environmental Protection Agency. Non-HalogenatedOrganics using GC/FID, Method 8015D. In June 2003.(22) Massachusetts Department of Environmental Protection.Method for the Determination of Extractable Petroleum Hydro-carbons (EPH). In May 2004.(23) Wrenn, B. A.; Venosa, A. D. Selective enumeration of aromaticand aliphatic hydrocarbon degrading bacteria by a most-probable-number procedure. Can. J. Microbiol. 1996, 42 (3), 252−258.(24) Kettler, T. A.; Doran, J. W.; Gilbert, T. L. Simplified method forsoil particle-size determination to accompany soil-quality analyses. SoilSci. Soc. Am. J. 2001, 65 (3), 849−852.(25) Walkley, A.; Black, I. A. An examination of the Degtjareffmethod for determining soil organic matter, and a proposedmodification of the chromic acid titration method. Soil Sci. 1934, 37(1), 29−38.(26) Bianchi, S. R.; Miyazawa, M.; Oliveira, E. L. D.; Pavan, M. A.Relationship between the mass of organic matter and carbon in soil.Braz. Arch. Biol. Technol. 2008, 51 (2), 263−269.(27) Mussa, S. A. B.; Elferjani, H. S.; Haroun, F. A.; Abdelnabi, F. F.Determination of available nitrate, phosphate and sulfate in soilsamples. Int. J. PharmTech Res. 2009, 1 (3), 598−604.(28) American Public Health Association (APHA), American WaterWorks Association, and Water Environment Federation. StandardMethods for the Examination of Water and Wastewater, 20th ed.; InWashington, DC, 1998.(29) Park, J.-D.; Ren, Z. Hysteresis controller based maximum powerpoint tracking energy harvesting system for microbial fuel cells. J.Power Sources 2012, 205, 151−156.(30) Friedel, J. K.; Molter, K.; Fischer, W. R. Comparison andimprovement of methods for determining soil dehydrogenase activityby using triphenyltetrazolium chloride and iodonitrotetrazoliumchloride. Biol. Fertil. Soils 1994, 18 (4), 291−296.(31) Johnson, J. L.; Temple, K. L. Some variables affecting themeasurement of ″catalase activity″ in soil. Soil Sci. Soc. Am. J. 1964, 28(2), 207−209.(32) Hankin, L.; Hill, D. E.; Stephens, G. R. Effect of mulches onbacterial populations and enzyme activity in soil and vegetable yields.Plant Soil 1982, 64 (2), 193−201.(33) Lu, L.; Xing, D.; Ren, N. Bioreactor performance andquantitative analysis of methanogenic and bacterial communitydynamics in microbial electrolysis cells during large temperaturefluctuations. Environ. Sci. Technol. 2012, 46 (12), 6874−6881.(34) Lu, L.; Xing, D.; Ren, N.; Logan, B. E. Syntrophic interactionsdrive the hydrogen production from glucose at low temperature inmicrobial electrolysis cells. Bioresour. Technol. 2012, 124, 68−76.(35) Margesin, R.; Zimmerbauer, A.; Schinner, F. Monitoring ofbioremediation by soil biological activities. Chemosphere 2000, 40 (4),339−346.
Environmental Science & Technology Article
dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX−XXXH

(36) Margesin, R.; Schinner, F. Bioremediation (natural attenuationand biostimulation) of diesel-oil-contaminated soil in an alpine glacierskiing area. Appl. Environ. Microbiol. 2001, 67 (7), 3127−3133.(37) Achuba, F. I.; Peretiemo-Clarke, B. O. Effect of spent engine oilon soil catalase and dehydrogenase activities. Int. Agrophys. 2008, 22(1), 1−4.(38) Allen, J. P.; Atekwana, E. A.; Atekwana, E. A.; Duris, J. W.;Werkema, D. D.; Rossbach, S. The microbial community structure inpetroleum-contaminated sediments corresponds to geophysicalsignatures. Appl. Environ. Microbiol. 2007, 73 (9), 2860−2870.(39) Popp, N.; Schlomann, M.; Mau, M. Bacterial diversity in theactive stage of a bioremediation system for mineral oil hydrocarbon-contaminated soils. Microbiology 2006, 152 (11), 3291−3304.(40) Rosano-Hernandez, M. C.; Ramírez-Saad, H.; Fernandez-Linares, L. Petroleum-influenced beach sediments of the CampecheBank, Mexico: diversity and bacterial community structure assessment.J. Environ. Manage. 2012, 95, S325−S331.(41) Militon, C.; Boucher, D.; Vachelard, C.; Perchet, G.; Barra, V.;Troquet, J.; Peyretaillade, E.; Peyret, P. Bacterial community changesduring bioremediation of aliphatic hydrocarbon-contaminated soil.FEMS Microbiol. Ecol. 2010, 74 (3), 669−681.(42) Zhang, D.-C.; Mortelmaier, C.; Margesin, R. Characterization ofthe bacterial archaeal diversity in hydrocarbon-contaminated soil. Sci.Total Environ. 2012, 421, 184−196.(43) Wrighton, K. C.; Agbo, P.; Warnecke, F.; Weber, K. A.; Brodie,E. L.; DeSantis, T. Z.; Hugenholtz, P.; Andersen, G. L.; Coates, J. D. Anovel ecological role of the Firmicutes identified in thermophilicmicrobial fuel cells. ISME J. 2008, 2 (11), 1146−1156.(44) Juang, D.-F.; Yang, P.-C.; Chou, H.-Y.; Chiu, L.-J. Effects ofmicrobial species, organic loading and substrate degradation rate onthe power generation capability of microbial fuel cells. Biotechnol. Lett.2011, 33 (11), 2147−2160.(45) Rezaei, F.; Xing, D.; Wagner, R.; Regan, J. M.; Richard, T. L.;Logan, B. E. Simultaneous cellulose degradation and electricityproduction by Enterobacter cloacae in a microbial fuel cell. Appl.Environ. Microbiol. 2009, 75 (11), 3673−3678.(46) Hua, X.; Wu, Z.; Zhang, H.; Lu, D.; Wang, M.; Liu, Y.; Liu, Z.Degradation of hexadecane by Enterobacter cloacae strain TU thatsecretes an exopolysaccharide as a bioemulsifier. Chemosphere 2010, 80(8), 951−956.(47) Rabaey, K.; Boon, N.; Siciliano, S. D.; Verhaege, M.; Verstraete,W. Biofuel cells select for microbial consortia that self-mediate electrontransfer. Appl. Environ. Microbiol. 2004, 70 (9), 5373−5382.(48) Friman, H.; Schechter, A.; Nitzan, Y.; Cahan, R. Effect ofexternal voltage on Pseudomonas putida F1 in a bio electrochemical cellusing toluene as sole carbon and energy source. Microbiology 2012,158 (2), 414−423.(49) Calvo, C.; Silva-Castro, G. A.; Uad, I.; Fandino, C. G.; Laguna,J.; Gonzalez-Lopez, J. Efficiency of the EPS emulsifier produced byOchrobactrum anthropi in different hydrocarbon bioremediation assays.J. Ind. Microbiol. Biotechnol. 2008, 35 (11), 1493−1501.(50) Zuo, Y.; Xing, D. F.; Regan, J. M.; Logan, B. E. Isolation of theexoelectrogenic bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell. Appl. Environ. Microbiol. 2008, 74 (10), 3130−3137.(51) Gao, M.; Li, L.; Liu, J. Simultaneous removal of hydrogen sulfideand toluene in a bioreactor: Performance and characteristics ofmicrobial community. J. Environ. Sci. 2011, 23 (3), 353−359.(52) Obuekwe, C. O.; Al-Jadi, Z. K.; Al-Saleh, E. S. Hydrocarbondegradation in relation to cell-surface hydrophobicity among bacterialhydrocarbon degraders from petroleum-contaminated Kuwait desertenvironment. Int. Biodeterior. Biodegrad. 2009, 63 (3), 273−279.(53) Cheung, P.-Y.; Kinkle, B. K. Mycobacterium diversity and pyrenemineralization in petroleum-contaminated soils. Appl. Environ. Micro-biol. 2001, 67 (5), 2222−2229.(54) Zhang, J.; Zhang, E.; Scott, K.; Burgess, J. G. Enhancedelectricity production by use of reconstituted artificial consortia ofestuarine bacteria grown as biofilms. Environ. Sci. Technol. 2012, 46(5), 2984−2992.
Environmental Science & Technology Article
dx.doi.org/10.1021/es4057906 | Environ. Sci. Technol. XXXX, XXX, XXX−XXXI




















