
A unified Lower-Middle Cambrian chronostratigraphy for West Gondwana
Upload
univ-lille1Category
view
6download
0
5 (2006) 255–265www.elsevier.com/locate/sedgeo
Sedimentary Geology 18
Microbial crusts as indicators of stratigraphic diastems in theCambrian Brèche à Micmacca, Atlas Mountains of Morocco
J. Javier Álvaro a,b,⁎, Sébastien Clausen b
a Departamento Ciencias de la Tierra, Universidad de Zaragoza, 50009-Zaragoza, Spainb Laboratoire de Paléontologie et Paléogéographie du Paléozoïque, UMR 8014 CNRS,
Université des Sciences et Technologies de Lille, 59655-Villeneuve d'Ascq, France
Abstract
The presence of microbial crusts interrupting high-energy clastic sediments in carbonate and mixed (carbonate–siliciclastic)platforms can be considered as distinct indicators of stratigraphic diastems, characterized by either very low rates of deposition ornon-deposition. This analysis is based on a microfacies study of the Cambrian Brèche à Micmacca (Atlas Mountains, Morocco),which contains some stratigraphically condensed limestones characterized by: (i) disarticulation and fragmentation of skeletons; (ii)abundance of volcanigenic debris; (iii) rarity of muddy fraction; (iv) dominance of a sessile, epibenthic shelly fauna dominated byechinoderms, chancelloriids and sponge spicules; and (v) a complex diagenetic history due to erosion and reworking of lithifiedsubstrates. The Brèche à Micmacca limestones are composite event-concentration, low-relief shoal complexes composed ofparautochthonous and allochthonous bioclastic assemblages. These units can be described as ‘hiatal accumulations’ because of thesubdivision of the coquinas by minor discontinuities or hiatal surfaces, and the composite, multiple-event nature of the grainstones/packstones.
The growth of microbial mats can be used as a record of time interrupting background-sedimentation patterns characterized bythe amalgamation of high-energy events. The Brèche àMicmacca was deposited in an environment in which some intervals of non-deposition (or under extremely low-sedimentation rates) alternated with the succession of sedimentary events. The biologicalresponse of microbial communities to coat stratigraphic discontinuities can be considered as an integral part of event stratigraphy.They enhance the preservation and identification potential of interruptions in the background sedimentation on substrates devoid ofburrowing activity.
A distinct event of non-deposition is also recognized, based on the presence of microbial fabrics, at the top of the Brèche àMicmacca. Its microkarst structures reflect a major fall in the relative sea level of a reduced region of the platform resulting indissolution and colonization of coelobiontic cavities by microbial mats and branching filaments.© 2006 Elsevier B.V. All rights reserved.
Keywords: Microbialites; Shell beds; Fe-oxides; Cambrian; Morocco
⁎ Corresponding author. Departamento Ciencias de la Tierra,Universidad de Zaragoza, 50009-Zaragoza, Spain. Fax: +34976761088.
E-mail addresses: [email protected] (J.J. Álvaro),[email protected] (S. Clausen).
0037-0738/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.sedgeo.2005.12.025
1. Introduction
The sedimentary record represents a succession ofepisodic or discontinuous depositional events, in somecases difficult to decipher due to burrowing, diageneticoverprints and/or differential compaction. The interplayof magnitude and frequency of sedimentary events,

256 J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
relative to background or normal conditions, and therecovery time for a given environment integrate todetermine the preservation potential of episodic events.In addition, these are commonly bounded by surfaces ofomission, which may result either from (i) high-intensityerosion, (ii) non-deposition, or (iii) very low rate ofdeposition. Manganese crusts, phosphate and glauconiteminerals, mineralised hardgrounds, highly burrowedintervals, paleosols and polygonally cracked emergencesurfaces are considered to be potential indicators of non-deposition. In contrast, very low depositional rates areidentifiable by two kinds of stratigraphic markers:unconformities and condensed sections. As summarizedby Shanmugam (1988), the major differences betweenboth stratigraphic markers are that: (i) unconformitiesare surfaces whereas condensed sections are intervals;and (ii) unconformities represent major breaks in thestratigraphic record, whereas condensed sections com-prise a more or less complete stratigraphic record, butwhich may contain many relatively minor sedimentarybreaks or diastems.
Monty (1976) pointed out that physical disconti-nuities play a decisive role in the occurrence and build-up of microbial fabrics on some carbonate and mixedplatforms: e.g., at least four days of non-deposition areneeded to re-establish well-preserved individual micro-bial laminae in modern stromatolites (Gerdes andKrumbein, 1994). As rapid syngenetic cementation ofmicrobial fabrics is a plausible way to withstand erosiveforces under turbulent waters (Pratt, 1982; Riding,1991b; Álvaro et al., 1998; among others), the episodicdevelopment of microbial crusts may represent markerbeds of either non-deposition or very low rate ofdeposition both in high-energy and quiescent environ-ments. In addition, early lithification of biofilms andthromboids promoted by calcification of cyanobacteriamay have significantly influenced the preservationpotential of microbial deposits by making them lesssusceptible to early diagenetic alteration, such asdolomitization (Glumac and Walker, 1997). As a result,microbial fabrics can be exceptionally preserved in asedimentary succession highly neomorphosed or epi-genically replaced by other minerals.
The aim of this paper is to recognize the factors thataccount for the temporal and paleoenvironmentaldistribution and preservation of microbial crusts in astratigraphic unit (the so-called ‘Brèche à Micmacca’ orMicmacca Breccia of Cambrian age, in the AtlasMountains, Morocco) characterized by breccia/grain-stone/packstone textures related to high-energy condi-tions and stratigraphic condensation (Destombes et al.,1985). Although the Brèche à Micmacca consists of
heterolithic strata, only the volcaniclastic and bioclasticlimestones will be studied below as they representcompound-event strata embedded in homogeneousshales. The analysis of microbial crusts in the BrècheàMicmacca limestones is proposed here as a tool for therecognition of sharp hiatuses interrupting backgroundsedimentation patterns.
2. Geologic setting and stratigraphy
The Moroccan Atlas comprises three distinctgeographic and geologic chains, named the Middle-,High- and Anti-Atlas (Fig. 1). The latter is boundedby the Souss valley, which has given its name to oneof the Neoproterozoic–early Paleozoic basins postu-lated in northwestern Africa: the Souss Basin (Geyerand Landing, 1995; Álvaro et al., 2003). Itslithostratigraphic units spanning the Lower–MiddleCambrian transition are the Tazlaft/Asrir formationsand the Brèche à Micmacca, proposed by Geyer(1989) as a member of the Jbel Wawrmast Formation(Fig. 2). The Tazlaft and Asrir formations (30–160 mthick) are heterolithic successions of sandstones,conglomerates and thin volcanic ashes, the formercropping out in the east–central and eastern Anti-Atlas, and the latter along the margin of the Anti-Atlasand in the High Atlas (Geyer and Landing, 1995). TheBrèche à Micmacca (up to 300 m thick; Destombes etal., 1985; Van Looy, 1985) consists of reddish tobrownish, volcaniclastic and bioclastic, bedded andnodular limestones that alternate with shale orsandstone strata. The member crops out from theHigh Atlas to the Sarhro Jbel and the El Graara massif(Fig. 1). Finally, the upper part of the Jbel WawrmastFormation consists of fine-grained sandstones andgreen (and minor purple) shales, with isolated ashintercalations and bioclastic limestone beds andnodules. The formation crops out from the westernHigh Atlas to the eastern and west–central Anti-Atlas.
A volcanic episode is recognized across the Lower–Middle Cambrian transition. Boudda and Choubert(1972), Destombes et al. (1985), and Geyer andLanding (1995) indicated that thin (up to 5 cm thick)volcanic ashes (frequently K-bentonites) are inter-bedded within the Brèche à Micmacca strata in thecentral and western Anti-Atlas, the Sarhro Jbel, and theHigh Atlas, and local pillowed flows lie locally in theSarhro Jbel (Geyer and Landing, 1995). The centres ofvolcanism are believed to have been in the westernHigh Atlas or somewhat farther north and northwest inthe latest Early Cambrian (Boudda et al., 1979;Buggisch and Siegert, 1988).
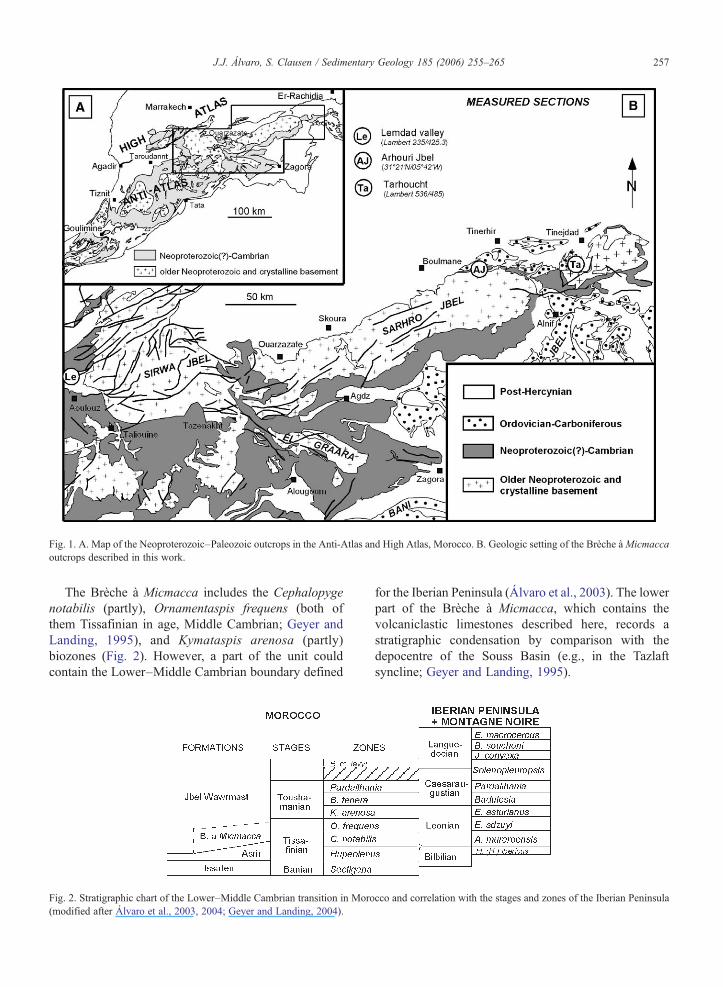
Fig. 1. A. Map of the Neoproterozoic–Paleozoic outcrops in the Anti-Atlas and High Atlas, Morocco. B. Geologic setting of the Brèche àMicmaccaoutcrops described in this work.
257J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
The Brèche à Micmacca includes the Cephalopygenotabilis (partly), Ornamentaspis frequens (both ofthem Tissafinian in age, Middle Cambrian; Geyer andLanding, 1995), and Kymataspis arenosa (partly)biozones (Fig. 2). However, a part of the unit couldcontain the Lower–Middle Cambrian boundary defined
Fig. 2. Stratigraphic chart of the Lower–Middle Cambrian transition in Moro(modified after Álvaro et al., 2003, 2004; Geyer and Landing, 2004).
for the Iberian Peninsula (Álvaro et al., 2003). The lowerpart of the Brèche à Micmacca, which contains thevolcaniclastic limestones described here, records astratigraphic condensation by comparison with thedepocentre of the Souss Basin (e.g., in the Tazlaftsyncline; Geyer and Landing, 1995).
cco and correlation with the stages and zones of the Iberian Peninsula

258 J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
3. Facies of the Brèche à Micmacca limestones
Facies with microbial textures occur at numerousexposures of the Brèche àMicmacca limestones, mainlyoverlying erosive surfaces. The microbial fabrics arecommon at three selected sites: the Lemdad valley,Tarhoucht, and the Arhouri Jbel (Figs. 1B and 3). Theyare patchly distributed on discrete bedding-planelimestone exposures, which are variable in number,and are in places condensed into a single bed or evendisappear laterally (Fig. 4A–B). The limestones occurboth overlying older rocks (even pre-Pan-Africaninliers) and interbedded with shales, and typicallyshow sharp crosscutting relationships. Stylolites andsolution-seam structures, locally abundant, enhance theapparent ‘brecciated’ appearance of these beds. Theirgreenish to reddish colours are directly related to theabundance and preservation of volcaniclastic debris.When they are unsorted, up to 2 cm across, and morecompletely replaced by chlorite, the whole rock isgreenish. By contrast, when the volcaniclastic fragmentsare relatively sorted (less than 0.5 cm in size), and
Fig. 3. Geometric relationships of the fac
coated by iron oxide, the rock is reddish in outcrop.Hematite affects as well the bioclastic content.
The limestones (up to 1.2 m thick) containamalgamated units up to 0.4 m thick (Fig. 4A). Thebase of each is defined by erosive surfaces that arenearly horizontal to broadly concave–convex. They arelined with lag concentrations, less than 20 cm thick, ofsiliciclastic (mainly sand, siltstone, claystone, andvolcaniclasts) and intraformational, limestone-dominatedclasts (Fig. 4B). Larger clasts are dominantly intrafor-mational and angular in shape, whereas the granule-sized clasts are subangular to subrounded in shape. Thebreccia lenses that directly cover the scour surfacesshow chaotic lithoclast orientations, rarely parallel tostratification. The upper parts of the individual units arepackstones (Fig. 4C) passing both vertically andlaterally into local grainstones and breccia levels, insome cases forming low-angle laminae. They containdisarticulated to broken trilobite debris and a widediversity of echinoderm ossicles (such as ceratocystids,ctenocystids, edrioasteroids, eocrinoid thecal plates andcolumnals, encrusting pelmatozoan holdfasts, etc.), and
ies in Tarhoucht and Arhouri Jbel.

Fig. 4. A. Crosscutting relationships of erosive discontinuities capped by microbial crusts (T2 /8). B. Detail of previous picture (boxed area) witharrowed scour surfaces covered by granule lags and stromatolite crusts. C. Volcaniclastic and bioclastic grainstone rich in echinoderm ossicles andtrilobite sclerites with characteristic dark aspect of polyphase skeletons after selective hematite replacement (Le1 /14). D. Microchannel truncating thetop of a microbial crest (partly replaced by iron oxides), filled by silt-sized quartz grains and an oncolite; both the stromatolitic mat and the channelinfill were subsequently eroded and overlain by bioclastic grainstones (Le11 /27). E. Extraclast encrusted by a compound agglutinated stromatolite(showing successive onlapping laminae) embedded in a bioclastic grainstone (Le1 /27). F. Sharp transition from wavy to columnar stromatolites, thewhole framework selectively replaced by iron oxide (Le1 /15). G. Subvertical crack cutting a vertical amalgamation of microbial crusts and silt-sizedlaminae (the top replaced by iron oxide), fill with the overlying bioclastic grainstone (Le1 /27). Scale bars are 1 mm long (except E=3 mm).
259J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
secondary heteractinid and hexactinellid sponge spi-cules, chancelloriid sclerites, brachiopods and helcio-nellids (Clausen, 2004). Skeletons are usuallydisorganized and locally subparallel to bedding. Com-
mon multiphase clasts contain cores of first-generationpebbles with accretionary sediment and erosion sur-faces. Both litho- and bioclasts were cracked duringtheir winnowing: the identification of minute intra-

260 J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
bioclastic, V-shaped brittle fractures is favoured by thedifferent colour of their filling, composed of inter-clastsediment and hematite cement. Hematite typicallyoccurs as a thin coating around bio- and lithoclasts,but it also stains red infiltrated mineral heterogeneities,such as cleavage planes of feldspars and skeletalarticulations (e.g., in trilobites), in some cases com-pletely replacing the skeletons.
Matrix, where observed, varies from mixtures ofcalcite micrite and microsparite, detrital dolomitic andterrigenous silt and sand in variable proportions. Micriteoccurs in scattered pockets and lenses (up to 1 cm long)commonly inclined at various angles to the inferreddepositional horizontal, and sometimes filling intrapar-ticular geopetal structures discordant with shelterfillings and stratification. Spar-filled voids have irreg-ular walls and a primary skeletal support. Primaryshelter pores, commonly formed beneath elongatedtrilobite and rarer brachiopod debris, were partially towholly occluded by one or several generations of calcitecements: the first one invariably of fibrous crystals (ca.0.5×0.1–0.2 mm), sometimes differentiated into twofibrous phases, and a later bladed sparry phase (ca. 0.1–0.3 mm across). Evidence of mechanical compaction,such as grain breakage and deformed clasts, is scarce.Bioturbation is absent.
Some composite clasts have grains and cementstruncated at borders indicating that some sedimentswere lithified when reworked. Subrounded to sub-angular shapes of volcaniclastic grains indicatevariable lateral transport during resedimentation.Multiphase clasts suggest multiple depositional, ce-mentation and erosive events. Intraclasts indicatereworking and ‘cannibalistic’ processes affecting theown carbonate seafloor. The bioclastic packstones andgrainstones are interpreted as derived from washingand reworking of bioclasts and other allochems fromthe surrounding seafloor generating low-angle shoalsunder high-energy conditions, which were amalgam-ated vertically by the episodic activity of high-energypulses. The presence of a selective diageneticprecipitation of iron oxides (mainly hematite; Fig.4C) allows differentiation into parautochthonous(unaffected) and allochthonous (secondarily reworkedafter hematite replacement) skeletons, as polyphaselitho- and bioclasts are coated or replaced by ironoxides, a cement that generally occluded the primary(intraparticular) porosity. Rapid cementation and henceinternal stabilization of the facies would haveprevented mechanical compaction. The packstones,grainstones and breccias are not earthquake-triggeredsediment gravity-flow deposits because units are
bedded and relatively thin (up to 0.4 m thick), clastsare small, and inferred depositional slopes are low.
4. Marker beds of sharp hydrodynamic fluctuations
Stromatolites form crusts overlying and interbeddedwithin the Brèche àMicmacca limestones (Fig. 3). Theyare up to 8 cm thick, although usually do not exceed 3cm. Although the thickness of the laminae is notconstant, in some cases they can be followed more than6 m laterally. The morphology of the crusts varies fromwrinkled to domal, and columnar features. Thestromatolites grew perpendicularly to substrate surfaces,and generally followed their irregularities. Two kinds ofstromatolite nucleation are described below: (i) coatingreworked intra- and extraclasts, and (ii) encrustingerosive surfaces and steeped walls of synsedimentaryfractures.
(i) Some sedimentary and volcanigenic extraclastsare incompletely coated by compound microbialcrusts (Fig. 4E). Cross-sections show crinkled towavy laminae, which incorporated both bio- andlithoclasts, medium-to-very coarse sand in size.Several generations of stromatolitic growth can bedistinguished, separated by onlapping geometries,which progressively reduced their lateral distribu-tion; as a result, the whole stromatolite frameworkshows common subtriangular cross-sections,sharply truncated by bioclastic grainstones.
(ii) Other compound stromatolites occur encrustingboth erosive surfaces and steep walls of centi-metre-thick, V-shaped cracks (up to 1 cm deep;Fig. 4G). Their internal fabric is a combinationof shapes. The first microbial mats grewperpendicularly to the substrate displayingwavy to crinkled, millimetre-thick laminae oflime mud, locally discontinuous and, in somecases, bearing a microscopic fenestral fabric.Lamination commonly passes upward into later-ally linked domal and hemispheroidal stromato-lites, with diameters ranging from 0.4 to 15 mmand internal laminae often wavy or definingsmaller, laterally linked hemispheroids. Finally,some stromatolite frameworks pass upward intocolumnar shapes, 0.3–0.8 mm wide and up to0.8 mm high, subparallel to slightly divergentbranching, and with rare lateral linkage (Fig.4F). Space between columns was subsequentlyfilled by bio- and lithoclasts. Well-defined light-dark microlaminae, occur locally defined by thin(50–200 μm), gently curved micritic layers. High-

261J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
energy pulses are episodically recorded as smallchannels (0.5 cm in diameter and deep) erodingboth intra-stromatolitic laminae and stromatolitictops, subsequently covered by a millimetre-thicksandstone, rich in bioclastic and volcaniclasticdebris, cemented by hematite and partiallyencrusted by biofilms. Finally, the whole stromat-olitic crusts were truncated and covered by eithermillimetre-thick quartz-rich sandstones or bio-clastic packstones/grainstones.
Although the stromatolitic microfabric was largelythe consequence of in situ calcite precipitation, there isevidence of detritus incorporation into the structures.Sediment consisting of quartz, feldspar, corroded micas,and volcaniclastic sand- and silt-sized grains wasepisodically trapped and bound into the surface.Terrigenous material is present in almost all the laminaeincreasing upward (in a wavy-to-columnar trend)between 10% to 60%. Along one lamina, the grainsconcentrate in small areas outlining micritic masses witha certain relief surrounded by clastic pockets, apparentlydeposited in stromatolite depressions (some of themgenerated by erosive microchannels). However, al-though it was introduced or reworked intermittently,by current or wave action, the silt and sand content ofthe water was not sufficient to prevent the growth ofmicrobial communities. Loose, unstabilized silt andsand deposited on the mat surface was frequentlyrippled by currents, which were sometimes able to erodeforming channel-like structures, subsequently over-grown by mats. No evidence of grazing and boringorganisms was found.
The first colonization phase of microbial laminatedmats lacking embedded siliciclastic material suggeststhat these substrates were temporarily exposed to quietwater conditions where thin microbial mats coveredthe substrates. These mats buried a variety of inheritedmicroreliefs related to the irregularities of a shelled/grainy substrate, such as erosive microchannels (Fig.4D), lenticular, lags and walls of V-shaped cracks(Fig. 4G). Smooth mats are common in areas of lowhydraulic energy (Gerdes and Krumbein, 1994),although a rapid syngenetic cementation allowsmicrobial mats to withstand erosive forces in high-energy environments. As we found no evidence ofgeneral subaerial exposure and dessication of thesemicrobial mats (except local development of karsticmicrocavities: see below), the aforementioned micro-bial cracks must be related to substrate instability afterearly lithification of mats. Subsequent winnowing andpartial reworking of the substrate (as suggested by the
abundance of polyphase clasts) would have favouredthe formation of irregularities on the substrate thatwould have induced the instability of lateral early-lithified microbial crusts.
The formation of laterally linked and stacked convexstromatolites is probably related to turbulence andresultant scour that would have promoted doming.Domes would have maintained by greater accretion ontheir crests. Columnar stromatolites probably formedwhen hemispheroidal stromatolites grew at a relativelyrapid pace because of high sedimentation rate both onmat surfaces and around them, maintaining a constantlylow synoptic relief. The stromatolites were progressive-ly subjected to moderate bottom currents, and theycoalesced to form an extensive, current-resistant fabric.The progressive modification of the stromatolitic shapesmay have developed as an adaptation to the continualexposure to such currents, their episodic erosive activityand the slight increase in siliciclastic input. Although theintra-bed, domal and columnar stromatolites achievedsynoptic relief of a few centimetres, they were (finally)truncated and overlain by scours and microchannels. Bycontrast, those that covered the top of the Brèche àMicmacca limestones were overlain by greenish shales,and preserve their complete 3D morphology. In this latecase, oncolitic accumulations, up to 3 cm thick, occurlaterally to the stromatolitic crusts and commonly fillshallow depressions.
5. Indicators of dissolution and karstic interruptions
The presence of pitted truncation surfaces at the topof the youngest volcaniclastic–bioclastic limestone ofthe Brèche à Micmacca in some exposures of theLemdad valley (e.g., section Le11 after Geyer andLanding, 1995) and Tarhoucht is marked by anotherdistinct development of microbial fabrics. Millimetre-sized, secondary cavities are easily recognisable incross-section, and consist of a series of bowl- andfunnel-shaped pits and vugs, in a packstone–grainstonetexture where burrowing is absent. Pitted truncationsurfaces are continuous with subjacent vugs. Vuggyinfill generally comprises less than 10% of rockvolume close to the top of the bed, and decreasesnotably 3 cm downward. Vugs and inter-connectedpores offer distinct crosscutting relationships, truncat-ing both skeletal grains and sparry cements. Complexsecondary cavities display three types of fill (Fig. 5A):(i) the ceilings are lined by a 0.2–0.3 mm-thick bandof fibrous calcite cements, (ii) the second fill compriseslayered quartz silt-sized grains with admixed clays, and(iii) the third fill is an iron-oxide (hematite) cement,

262 J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
which fills the remaining pore space. On the contrary,simple secondary voids were only filled by hematite,although some of them previously recorded microbialcolonizations, composed of stromatolitic crusts (subse-quently replaced by iron oxides; Fig. 5B), and branchingmicro-organisms (replaced at present by quartz mosaics)living in coelobiontic or low-light habitats (Fig. 5C–E).The pendent and sideway growth of branching organismsfrom the walls of microcavities indicates that there was anetwork of open spaceswith early-cementedwalls. Thesemicro-organisms show a bundles of filaments, develop-ingupwardbranching filaments. Individual growth formsrange from 1.5 to 3.5 mm long; filaments are circular incross-section and average 30–40 μm in diameter. Theyare straight or slightly undulating, and are characterizedby either a simple, regular, and dichotomously branchedhabit, or asymmetrically branching patternsmaking acuteangles (less than 20°). Based on their size andmorphology (branched clusterswith fan-like longitudinal
Fig. 5. Microphotographs of microkarstic cavities. A. Cavity with three typequartz grains on the lowest part (s), and iron-oxide cements (io) (T3 /1b). B. Ccavity, with wall composed of skeletons and cements of the enclosed packstdetail in D and E; Le11 /16). Scale bars=1 mm (except C=2.5 mm).
section, radial erect growth, and separate–not juxta-posed–adjacent filaments), thesemicrobial filaments canbe assigned to the Hedstroemia Group (Riding, 1991b).These coelobites were probably unrelated to photosyn-thetic activities.
Contrasts in depositional environment, diagenetichistory and microbial colonization across pitted surfacesindicate that the top of the Brèche à Micmacca in theLemdad and Tarhoucht areas was probably subaeriallyexposed or in contact with undersaturated waters. As aresult, a pot-holed surface was produced prior to micro-bial colonization and subsequent hematite impregnation.
6. Paleoecological implications and sedimentarycontrols
The benthic community recorded in the Brèche àMicmacca limestones exhibits high-diversity patterns(Clausen, 2004). Echinoderm-chancelloriid-sponge
s of fill: a band of fibrous calcite cement (fc) on the ceiling, silt-sizedavity walls encrusted by stromatolitic laminae (Le11 /16). C. Irregular
one, filled with hematite and microbial filaments (boxed and shown in

263J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
meadows were subjected to high-energy conditions thatdestroyed them almost as quickly as they formed. Thecoarsest fraction of the skeletons was transported shortdistances, while the smallest grains were redistributedby currents across the platform resulting in episodicsheets of loose hydrodynamic bioclastic sediment. Thispeak in biodiversity (when comparing with underlyingand overlying strata) may seemingly be interpreted asabrupt changes in population or community structurerelated to short-term expansions of benthic communi-ties. However, the comparative study of diageneticpatterns and microbial fabrics allows the recognition ofa complex sedimentary history, incompletely con-strained due to the allochthonous character of theshelly assemblage. The irregular hydrodynamic regimeof its background sedimentation is reflected by theepisodic presence of intra-bed erosive surfaces (insome cases laterally amalgamated) and microbialfabrics. The limestone beds represent ecologicallymixed assemblages and display peaks in biodiversitypatterns related to: (i) the episodic shallow-watererosive reworking; (ii) the taphonomic feedback duringshell accumulation (e.g indicated by the presence ofencrusting pelmatozoan holdfasts); and (iii) the tapho-nomically complex histories reflected by the selectivereplacement and coating of iron oxides and by thepolyphase clasts. The limestones have recorded asuccession of epibenthic communities, which colonizedfirm sediments giving rise to limited and scatteredechinoderm-sponge-chancelloriid meadows, whose re-lief above the surrounding seafloor was slight andlimited. Intensive bottom current abrasion and winnow-ing affected the substrate in which the microbes werethe only organisms preserved in their original growthposition. Microbial installation and growth indicate thatthe bottom energy was episodically lower, at least thetime span necessary to allow colonization by theseopportunistic biofilms and mats, so that the biomats canbe considered as stratigraphic markers of the irregularhydrodynamic regime on some substrates. Mat-bearingsurfaces may have withstood episodic high-energysedimentary pulses if they (or probably the degradingmaterial under the living surface) lithified rapidly andincorporated thin lenses and laminae of siliciclasticdebris, implying either rapid microbial motility or high-growth rates.
Growth of microbial crusts was induced by benthicmicrobial communities, either from biologically in-duced precipitation around micro-organisms or viadecomposition of their organic matter that providemicro-environments conductive to calcite precipitation.The microbial communities formed surficial mats across
the substrate irregularities enhanced by skeletal fabrics.Fine skeletal debris and silt washed on to the surfacebecame trapped and bound by the microbial communi-ties in a manner analogous to many modern microbialmats generating ‘agglutinated stromatolites’ (sensuRiding, 1991a, 1999). The main hydrodynamic factorsthat controlled biomat development were:
(i) The rate of sediment accumulation: a lowsediment accumulation rate permitted coloniza-tion of wide substrates by microbial communities.Once established, the microbial communitiesexpanded and coalesced. The ideal verticalsequence from wrinkled to domal and low-reliefcolumnar stromatolites may reflect the upwardincrease of sedimentation rates (Dromart et al.,1994; Hofmann et al., 1999).
(ii) Early lithification: several observations suggestthat the crusts were lithified early, as evidenced bytheir erosion and truncation of their tops preserv-ing scours and microchannels filled by quartz andvolcaniclastic-dominated silts and fine-grainedsandstones.
(iii) Substrate stability: although the development ofmicrobial crusts requires presence of suitablesubstrates, in this case the microtopography of theshelly/grainy seafloor provided growth space,where the cemented shelled surface (mainlycomposed of bioclastic grainstones and early-cemented packstones) was permanently suitablefor microbial nucleation. The lack of intra-bedoncolites, relatively common on the top of thelimestones, suggests a relative stability of thesubstrates. As clay flakes are absent embedded orunder the microbial crusts, development of clayeysubstrates from short-term intervals of quiescencecannot be interpreted.
(iv) Sediment input: as rapid and frequent sedimen-tation impedes primary production, in situ syn-thesis of organic matter corresponds to intervalsof sediment starvation (Gerdes et al., 1991).Therefore, the silt and clay content of the waterwas not sufficient to prevent the growth ofmicrobial communities, although they were intro-duced or reworked intermittently as reflected bydevelopment of agglutinated stromatolites. How-ever, the vertical growth of microbial mats (passingupward from crinkled to low-relief columnarshapes) and the vertical increase in siliciclasticdebris trapped between the laminae and columns,indicate a progressive increase of water turbu-lence and siliciclastic input.

264 J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
In summary, the limestones of the Brèche àMicmacca are composite event-concentration, low-relief shoal complexes composed of parautochthonousand allochthonous bioclastic assemblages. These unitscan be described as ‘hiatal accumulations’ (sensuKidwell, 1991; Kidwell and Bosence, 1991) becauseof (i) the subdivision of the coquinas by minordiscontinuities or hiatal surfaces, and (ii) the composite,multiple-event nature of the packstones/grainstones.Although hiatal accumulations are not necessarily con-densed, because skeletons can accumulate in significantthicknesses that equal or even exceed aggradation inadjacent environments, the Brèche àMicmacca is strati-graphically condensed when comparing with shale-dominated lithofacies (Destombes et al., 1985; Geyerand Landing, 1995).
Finally, under what conditions do not microbialcrusts not form during depositional hiatuses? Based onfield and laboratory studies (e.g., Golubic and Browne,1996; Steneck et al., 1998; Reid et al., 2003), the timespan related to hiatuses is another ecological control:brief hiatuses can favour development of microbiallaminated crusts, whereas prolonged episodes or longerhiatuses can be characterized by microboring activityassociated with growth of complex and diversifiedmicrobial (climax) communities. As a result, there is alimit to the time span related to these hiatal processes, aslonger hiatal episodes can result in development ofeukaryotic algal communities, which do not formlaminated structures.
7. Conclusions
With this paper we wish to draw attention to animportant indicator of stratigraphic diastems: the pres-ence of microbial crusts interrupting high-energy clasticsediments in carbonate and mixed (carbonate–siliciclas-tic) platforms. This analysis is based on a microfaciesstudy of the limestones of the Cambrian Brèche àMicmacca, the lower member of the Jbel WawrmastFormation in the AtlasMountains, Morocco. The Brècheà Micmacca contains some stratigraphically condensedlimestones characterized by: (i) disarticulation andfragmentation of skeletons; (ii) abundance of volcani-genic debris; (iii) rarity of muddy fraction; (iv)dominance of a sessile, epibenthic shelly fauna domi-nated by echinoderms, chancelloriids and spongespicules; and (v) a complex diagenetic history due toerosion and reworking of lithified substrates.
Abrupt changes in hydrodynamic patterns areindicated by the presence of microbial mats, whichreflect in situ production of organic matter and
interruption of background sedimentation. Whereas thelatter is recognized by the abundance of disarticulatedand fragmented skeletons derived from different sources(leading to ecologically mixed assemblages), the micro-bial mats are autochthonous. This alternation allowsidentification of two kinds of sedimentation rates: aspointed out by Gerdes et al. (1991), a microbial crustonly a few millimetres thick may correspond to a timespan of many years, while the much thicker bioclasticaccumulations they bound may have been deposited in afew hours. The microbial mats appear to have beenrestricted to particular microenvironmental conditions,episodically developed on irregular substrates ofreduced clastic influence and critical combination ofturbulence, episodic high-energy pulses, and time spanrelated to the hiatuses. They were also influenced by thelocal hydrodynamic regime, as shown by scours andsmall channels on mat surfaces. Although the micro-environmental or ecological changes that resulted in thislocalized and episodic, microbial colonization areunknown in detail, some major control factors can beadvanced, such as decreased sedimentation rates,reduction in siliciclastic supply, decreased water energy,or a combination of these.
A distinct non-deposition event is also recognized,based on the presence of microbial fabrics, at the top ofthe youngest limestone bed of the Brèche à Micmacca.Although the presence of some hematite patches mayseemingly be interpreted as reworked iron-rich extra-clasts and impregnated intraclasts, the presence ofmicrobial crusts (replaced by iron oxides) and branchingfilaments (replaced by silica mosaics) allows the(proper) identification of dissolution processes, micro-bial colonization of coelobiontic or low-light habitats,and final impregnation and filling with iron-oxidecements. The crosscutting border of the karstic pitsand vugs (affecting both skeletons and cements), and theradial growth of the microbial filaments from the cavitywalls allow distinct recognition of dissolution processes;in addition, the direct encrusting patterns of microbialfilaments adapted to the wall irregularities is a furtherindicator of dissolution predating microbial coloniza-tion. The microkarst structures described at the top ofthe Brèche à Micmacca reflect a fall in the relative sealevel of a reduced region of the platform resulting in theformation of dissolution and colonization of coelobion-tic cavities. Finally, the definitive flooding of the wholeplatform led to the establishment of clayey substrates.
The growth of microbial mats can be used as a recordof time interrupting background-sedimentation patternscharacterized by the amalgamation of high-energyevents. The Brèche à Micmacca was deposited in an

265J.J. Álvaro, S. Clausen / Sedimentary Geology 185 (2006) 255–265
environment in which some intervals of non-deposition(or extremely low-sedimentation rates) alternated withthe amalgamation of millimetre- to centimetre-scale,sedimentary events. The biological response of micro-bial communities to coat stratigraphic discontinuities canbe considered as an integral part of event stratigraphy.They enhance the preservation and identificationpotential of interruptions in the background sedimenta-tion on substrates devoid of burrowing activity.
Acknowledgements
This paper is a contribution to ECLIPSE project‘Evolution of biogeochemical cycles from Archean toRecent environments’ funded by CNRS. Helpfulcomments and advice from Rachel Wood and BrianPratt, and editorial work by Gilbert Camoin have greatlyimproved the manuscript.
References
Álvaro, J.J., Vennin, E., Vizcaïno, D., 1998. Depositional controls onEarly Cambrian microbial carbonates from the Montagne Noire,southern France. Trans. R. Soc. Edinb. Earth Sci. 89, 135–143.
Álvaro, J.J., Elicki, O., Geyer, G., Rushton, A.W.A., Shergold, J.H.,2003. Palaeogeographical controls on the Cambrian trilobiteimmigration and evolutionary patterns reported in the westernGondwana margin. Palaeogeogr. Palaeoclimat. Palaeoecol. 195,5–35.
Álvaro, J.J., Vizcaïno, D., Kordule, V., Fatka, O., Pillola, G.L., 2004.Some solenopleurine trilobites from the Languedocian (late MidCambrian) of western Europe. Geobios 37, 135–147.
Boudda, A., Choubert, G., 1972. Sur la limite inférieure du Cambrienau Maroc. C.R. Acad. Sci. Paris 275, 5–8.
Boudda, A., Choubert, G., Faure-Muret, A., 1979. Essai destratigraphie de la couverture sédimentaire de l'Anti-Atlas:Adoudounien, Cambrien inférieur. Notes Mém. Serv. Géol.Maroc. 271, 1–96.
Buggisch, W., Siegert, R., 1988. Paleogeography and facies of the‘grès terminaux’ (uppermost Lower Cambrian, Anti-Atlas,Morocco). In: Jacobshagen, V.H. (Ed.), The Atlas System ofMorocco. Studies on its Geodynamic Evolution. Lect. NotesEarth Sci., vol. 15. Springer-Verlag, Berlin, pp. 107–121.
Clausen, S., 2004. The non-reefal echinoderm-sponge meadows of theLower–Middle Cambrian transition in the western Mediterraneanregion. PhD, University of Lille I, France. [unpublished].
Destombes, J., Hollard, H., Willefert, S., 1985. Lower Palaeozoicrocks of Morocco. In: Holland, C.H. (Ed.), Lower PalaeozoicRocks of the World. Lower Palaeozoic of North-Western andWest Central Africa, vol. 4. John Wiley and Sons, Chichester,pp. 157–184.
Dromart, G., Gaillard, C., Jansa, L.F., 1994. Deep-marine microbialstructures in the Upper Jurassic of western Tethys. In: Bertrand-Sarfati, J., Monty, C. (Eds.), Phanerozoic Stromatolites II. KluwerAcad. Publ., Dordrecht, pp. 295–318.
Gerdes, G., Krumbein, W.E., 1994. Peritidal potential stromatolites—asynopsis. In: Bertrand-Sarfati, J., Monty, C. (Eds.), PhanerozoicStromatolites II. Kluwer Acad. Publ., Dordrecht, pp. 101–129.
Gerdes, G., Krumbein, W.E., Reineck, H.E., 1991. Biolaminations —ecological versus depositional dynamics. In: Einsele, G., Ricken,W., Seilacher, A. (Eds.), Cycles and Events in Stratigrahy.Springer-Verlag, Berlin, pp. 592–607.
Geyer, G., 1989. Late Precambrian to early Middle Cambrianlithostratigraphy of southern Morocco. Beringeria 1, 115–143.
Geyer, G., Landing, E. (Eds.), 1995. Morocco'95. The Lower–Middle Cambrian standard of Gondwana. Beringeria, Sp. Iss.,vol. 2, pp. 1–171.
Geyer, G., Landing, E., 2004. A unified Lower–Middle Cambrianchronostratigraphy for West Gondwana. Acta Geol. Pol. 54,179–218.
Glumac, B., Walker, K., 1997. Selective dolomitization of Cambrianmicrobial carbonate deposits: a key to mechanisms and environ-ments of origin. Palaios 12, 98–110.
Golubic, S., Browne, K.M., 1996. Schizothrix gebeleinii sp. novabuilds subtidal stromatolites, Lee Stocking Island. AlgologicalStudies 83, 273–290.
Hofmann, H.J., Grey, K., Hickman, A.H., Thorpe, R.I., 1999. Origin of3.45 Ga coniform stromatolites in Warrawoona Group, WesternAustralia. Geol. Soc. Am. Bull. 111, 1256–1262.
Kidwell, S.M., 1991. Taphonomic feedback (live/dead interactions)in the genesis of bioclastic beds: key for reconstructing sedi-mentary dynamics. In: Einsele, G., Ricken, W., Seilacher, A.(Eds.), Cycles and Events in Stratigrahy. Springer-Verlag, Berlin,pp. 268–282.
Kidwell, S.M., Bosence, D.W.J., 1991. Taphonomy and time-averaging of marine shelly faunas. In: Allison, P.A., Briggs, D.E.G. (Eds.), Taphonomy: Releasing the Data Locked in theFossil Record. Topics in Geobiology. Plenum Press, New York,pp. 115–209.
Monty, C.L.V., 1976. The origin and development of cryptalgalfabrics. In: Walter, M.R. (Ed.), Stromatolites. Development inSedimentology, vol. 20. Elsevier, Amsterdam, pp. 193–249.
Pratt, B., 1982. Stromatolite framework of carbonate mud mounds.J. Sediment. Petrol. 53, 1203–1227.
Reid, P., Dupraz, C.D., Visscher, P.T., Sumner, D.Y., 2003. Microbialprocesses forming marine stromatolites. In: Krumbein, W.E.,Paterson, D.M., Zavarzin, G.A. (Eds.), Fossil and Recent Biofilms:A Natural History of Life on Earth. Kluwer Acad. Publ.,Dordrecht, pp. 103–118.
Riding, R., 1991a. Classification of microbial carbonates. In: Riding,R. (Ed.), Calcareous Algae and Stromatolites. Springer-Verlag,Berlin, pp. 21–51.
Riding, R., 1991b. Calcified cyanobacteria. In: Riding, R. (Ed.),Calcareous Algae and Stromatolites. Springer-Verlag, Berlin,pp. 55–87.
Riding, R., 1999. The term stromatolite: towards an essentialdefinition. Lethaia 32, 321–330.
Shanmugam, G., 1988. Origin, recognition, and importance oferosional unconformities in sedimentary basins. In: Kleinspehn,K., Paola, C. (Eds.), New Perspectives in Basin Analysis. SpringerVerlag, Berlin, pp. 83–107.
Steneck, R.S., Miller, T.E., Reid, R.P., Macintyre, I.G., 1998.Ecological controls on stromatolite development in a modernreef environment: a test of the ecological refuge paradigm.Carbonates and Evaporites 13, 48–65.
Van Looy, J., 1985. Het kaartblad Tazenakht 1 /100.000, Anti-Atlas,Marokko. Kartiering, Lithostratigrafie, Biostratigrafie van Pre-cambrium tot Tremadoc. Proefschrift Fac. Wetenschappen, Kath.Univ. Leuven.
Copyright © 2022 FDOKUMEN




















