
Microbial community structure and methane-cycling activity of subsurface sediments at Mississippi...
9
Microbial community structure and methane-cycling activity of subsurface sediments at Mississippi Canyon 118 before the Deepwater Horizon disaster Sarah Underwood a , Laura Lapham b , Andreas Teske a , Karen G. Lloyd a,n a Department of Marine Sciences, University of North Carolina at Chapel Hill, NC, USA b Chesapeake Biological Laboratory, USA article info Keywords: Methane Methanogenesis Sulfate reduction Anoxic sediments Deepwater horizon abstract The Deepwater Horizon disaster caused a shift in microbial communities in Gulf of Mexico seawater, but less is known about the baseline for microbial communities in the underlying sediments. We compared 16S rRNA and functional gene sequences deriving from DNA and RNA with geochemical profiles (sulfate and methane concentrations, δ 13 C of methane and carbon dioxide, and chloride concentrations) of a sediment gravitycore from the upper continental slope of the northwestern Gulf of Mexico (MC118) in 2008, 15 km from the spill site. The highest number of archaeal sequences were ANME-1 and ANME-2 archaea in the sulfate-reducing upper core segments (12 and 42 cmbsf), ANME-1 and Methanomicro- biales in the middle methanogenic depths (200 and 235 cmbsf), and ANME-1 at the deepest depths (309, 400, and 424 cmbsf). The presence of mcrA gene transcripts showed that members of the ANME-1 group are active throughout the core and transcribe the mcrA gene, a key gene of methanogenesis and anaerobic methane oxidation. The bacterial community consists mostly of members of the Deltaproteo- bacteria, Chloroflexi, Cytophaga, Epsilonproteobacteria, and the Japan Sea Group 1 throughout the core. The commonly detected genera of gammaproteobacterial hydrocarbon-degrading bacteria in the water column are not found in this sediment survey, indicating that the benthic sediment is an unlikely reservoir for these aerobes. However, the sediments contain members of the sulfate-reducing families Desulfobulbaceae and Desulfobacteraceae, some members of which degrade and completely oxidize aromatic hydrocarbons and alkanes, and the Desulfobacterium anilini lineage of obligately aromatics- degrading sulfate reducers. Thus, the benthic sediments are the most likely reservoir for the active deltaproteobacterial populations that were observed repeatedly after the Deepwater Horizon spill in the fall of 2010. & 2015 Elsevier Ltd. All rights reserved. 1. Introduction Hydrocarbon seeps on the continental slope of the northern Gulf of Mexico contain a combination of methane and other light hydrocarbons resulting from thermal cracking of deeply buried ancient organic matter, augmented by methane produced by the biological remineralization of organic matter in shallower depths with cooler, microbially-compatible temperatures (Sassen et al., 2004). In areas of active hydrocarbon seepage, upward flow of these deeply-sourced fluids is met by sulfate-rich seawater diffus- ing into surficial sediments, where sulfate is quickly consumed by microbial sulfate reduction with multiple carbon sources and elec- tron donors, including methane (Lloyd et al., 2006, 2010). In most cases, the rates of sulfate-dependent anaerobic oxidation of meth- ane (AOM) at hydrocarbon seeps in the Gulf of Mexico and other locations are at least an order of magnitude lower than sulfate reduction rates with other substrates that are available in situ at the same sites (Bowles et al., 2011). In surficial Gulf of Mexico cold seep sediments, the archaea and bacteria that drive sulfate- dependent AOM are the methanogen-affiliated archaea ANME- 1a/b, ANME2a/b, and ANME-2c co-occurring with deltaproteobac- terial sulfate-reducing bacteria; these AOM communities are consistently detectable in surficial seep sediments in the Gulf of Mexico (Lloyd et al., 2006, 2010). However, in addition to well- studied and highly active seep sites that are often associated with conspicuous microbial mats and chemosynthetic invertebrate communities, many sediments of the slope of the Gulf of Mexico Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/dsr2 Deep-Sea Research II http://dx.doi.org/10.1016/j.dsr2.2015.01.011 0967-0645/& 2015 Elsevier Ltd. All rights reserved. n Corresponding author. Current address: Department of Microbiology, University of Tennessee at Knoxville, USA. Tel.: þ1 865 974 3441. E-mail address: [email protected] (K.G. Lloyd). Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments at Mississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎
Transcript of Microbial community structure and methane-cycling activity of subsurface sediments at Mississippi...

Microbial community structure and methane-cycling activityof subsurface sediments at Mississippi Canyon 118 beforethe Deepwater Horizon disaster
Sarah Underwood a, Laura Laphamb, Andreas Teske a, Karen G. Lloyd a,n
a Department of Marine Sciences, University of North Carolina at Chapel Hill, NC, USAb Chesapeake Biological Laboratory, USA
a r t i c l e i n f o
Keywords:MethaneMethanogenesisSulfate reductionAnoxic sedimentsDeepwater horizon
a b s t r a c t
The Deepwater Horizon disaster caused a shift in microbial communities in Gulf of Mexico seawater, butless is known about the baseline for microbial communities in the underlying sediments. We compared16S rRNA and functional gene sequences deriving from DNA and RNA with geochemical profiles (sulfateand methane concentrations, δ13C of methane and carbon dioxide, and chloride concentrations) of asediment gravity core from the upper continental slope of the northwestern Gulf of Mexico (MC118) in2008, 15 km from the spill site. The highest number of archaeal sequences were ANME-1 and ANME-2archaea in the sulfate-reducing upper core segments (12 and 42 cmbsf), ANME-1 and Methanomicro-biales in the middle methanogenic depths (200 and 235 cmbsf), and ANME-1 at the deepest depths (309,400, and 424 cmbsf). The presence of mcrA gene transcripts showed that members of the ANME-1 groupare active throughout the core and transcribe the mcrA gene, a key gene of methanogenesis andanaerobic methane oxidation. The bacterial community consists mostly of members of the Deltaproteo-bacteria, Chloroflexi, Cytophaga, Epsilonproteobacteria, and the Japan Sea Group 1 throughout the core.The commonly detected genera of gammaproteobacterial hydrocarbon-degrading bacteria in the watercolumn are not found in this sediment survey, indicating that the benthic sediment is an unlikelyreservoir for these aerobes. However, the sediments contain members of the sulfate-reducing familiesDesulfobulbaceae and Desulfobacteraceae, some members of which degrade and completely oxidizearomatic hydrocarbons and alkanes, and the Desulfobacterium anilini lineage of obligately aromatics-degrading sulfate reducers. Thus, the benthic sediments are the most likely reservoir for the activedeltaproteobacterial populations that were observed repeatedly after the Deepwater Horizon spill in thefall of 2010.
& 2015 Elsevier Ltd. All rights reserved.
1. Introduction
Hydrocarbon seeps on the continental slope of the northernGulf of Mexico contain a combination of methane and other lighthydrocarbons resulting from thermal cracking of deeply buriedancient organic matter, augmented by methane produced by thebiological remineralization of organic matter in shallower depthswith cooler, microbially-compatible temperatures (Sassen et al.,2004). In areas of active hydrocarbon seepage, upward flow ofthese deeply-sourced fluids is met by sulfate-rich seawater diffus-ing into surficial sediments, where sulfate is quickly consumed by
microbial sulfate reduction with multiple carbon sources and elec-tron donors, including methane (Lloyd et al., 2006, 2010). In mostcases, the rates of sulfate-dependent anaerobic oxidation of meth-ane (AOM) at hydrocarbon seeps in the Gulf of Mexico and otherlocations are at least an order of magnitude lower than sulfatereduction rates with other substrates that are available in situ atthe same sites (Bowles et al., 2011). In surficial Gulf of Mexico coldseep sediments, the archaea and bacteria that drive sulfate-dependent AOM are the methanogen-affiliated archaea ANME-1a/b, ANME2a/b, and ANME-2c co-occurring with deltaproteobac-terial sulfate-reducing bacteria; these AOM communities areconsistently detectable in surficial seep sediments in the Gulf ofMexico (Lloyd et al., 2006, 2010). However, in addition to well-studied and highly active seep sites that are often associated withconspicuous microbial mats and chemosynthetic invertebratecommunities, many sediments of the slope of the Gulf of Mexico
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/dsr2
Deep-Sea Research II
http://dx.doi.org/10.1016/j.dsr2.2015.01.0110967-0645/& 2015 Elsevier Ltd. All rights reserved.
n Corresponding author. Current address: Department of Microbiology, Universityof Tennessee at Knoxville, USA. Tel.: þ1 865 974 3441.
E-mail address: [email protected] (K.G. Lloyd).
Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments atMississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i
Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎

are characterized by moderate or low seepage activity. Such sed-iments lack the macrofaunal and microbial mat indicators ofsurficial seepage, and the sulfate–methane transition zone whereAOM can be sustained by overlapping methane and sulfate inthe sediment column is often found meters into the sediments(Lapham et al., 2008).
The role of ANME archaea in sediments with a deep (more than afew tens of cm below the seafloor) methane–sulfate transition zone isuncertain, since ANME archaea are often not detected by 16S rRNAand functional gene sequencing in deep sediments (Biddle et al., 2006,2011; Briggs et al., 2012; Inagaki et al., 2006; Sørensen and Teske,2006). Likewise, the detection of other methanogen-affiliated archaeabecomes challenging and spotty in deep subsurface sediments (Briggset al., 2012; Colwell et al., 2008; Inagaki et al., 2006; Lever, 2013). Inresponse to these difficulties, some uncultured archaeal groups werespeculatively linked with the methane cycle (i.e., Sørensen and Teske,2006). However, plausible alternate candidates for sulfate-dependentmethane oxidation or methanogenesis in the deep subsurface havenot been identified or verified; the detection problems are currentlyviewed as sensitivity issues that should be tackled by improved primerdesign (Lever, 2008) and more sensitive DNA or cell isolation tech-niques (Kallmeyer et al., 2008).
We addressed the detection and diversity of ANME and othermethanogen-affiliated archaea in subsurface sediments with a deta-iled study of an intermediate model system, a gravity core from thevicinity (10 m) of methane seeps at Mississippi Canyon lease block 118(MC118) with a methane–sulfate transition zone between ca. 1 and1.5 m depth. The scale of this core (4.55 m depth) places it into theintermediate subsurface interval between small push cores that arerecovered from active seafloor seeps by ROV, submersible or Multi-corer (ca. 0.5 m length), and deep subsurface ODP and IODP coreswhich retrieve spatially extended methane–sulfate gradients and deepmethane–sulfate transition zones from organic-rich continental mar-gin sediments on a scale of 50 m and up (for example, D’Hondt et al.,2004). One goal of this study was to analyze the composition ofmethanogenic and methane-oxidizing microbial communities in theintermediate subsurface, and to compare them to highly activesurficial seeps and deep subsurface sediment. Specifically, we identi-fied the depth distribution of ANME archaea as well as other pote-ntially methane-cycling groups and sulfate-reducing groups by analyz-ing bacterial and archaeal 16S rRNA genes and key functional genes(dsrAB, mcrA) of dissimilatory sulfate reduction (Zverlov et al., 2005;Klein et al., 2001) and AOM andmethanogenesis (Springer et al., 1995;Luton et al., 2002; Hallam et al., 2004), respectively. Since DNA can bewell-preserved in anoxic cold environments (Willerslev and Cooper,2005), we also analyzed archaeal and bacterial 16S rRNA transcripts, aswell as dsrAB andmcrAmRNA transcripts that may be more indicativeof the active population.
This sediment core was obtained two years before the Deep-water Horizon oil spill from the northwestern margin of WoolseyMound at MC118, approx. 15 km northwest of the Macondo well-head. Woolsey Mound is a transient, thermogenic, fault-controlledgas hydrate system characterized by abundant gas hydrate/carbo-nate outcrops, bubble plumes, clam shells and microbial mats(Macelloni et al., 2013; Lloyd et al., 2010). Yet, information on thesedimentary microbial community below 15 cmbsf is absent. Thisstudy therefore opens up a second scientific goal, to provide asample of the sedimentary microbial community composition ofthe upper continental slope before the Deepwater Horizon blow-out. Previously, microbial community analyses in the deep Gulf ofMexico have focused on brine lakes and hydrocarbon seeps (Joyeet al., 2009), Beggiatoa mats at seep sites (Mills et al., 2004), highlyactive cold seep sediments (Lloyd et al., 2006, 2010; Bowles et al.,2011), and methane hydrate deposits (Lanoil et al., 2001; Millset al., 2003). These case studies of extreme habitats do not providea suitable baseline for the wide range of benthic sediments in the
Gulf of Mexico that have been affected by the Macondo blowout.The broad spectrum of all bacterial and archaeal phylotypes fromthis core will contribute to a general baseline for microbialcommunity structure in moderately active upper continental slopesediments in the Gulf of Mexico.
Since the continental slope of the Gulf of Mexico experiencessubstantial natural oil seepage (Kennicutt et al., 1988) and is likelyto constitute a natural reservoir of oil-degrading microbial popula-tions, we finally address a third scientific goal which examineswhether the deep sedimentary community could possibly providea source of microbial populations that are primed to metabolizehydrocarbon contamination from the spill. To do this, the abun-dant sequence datasets from the Gulf of Mexico after the Deep-water Horizon blowout (Redmond and Valentine, 2012; Yang et al.,2014; Arnosti et al., this issue) were mined for comparativeidentification of possible hydrocarbon-degrading microorganisms.
2. Materials and methods
2.1. Core collection and pore-water chemistry
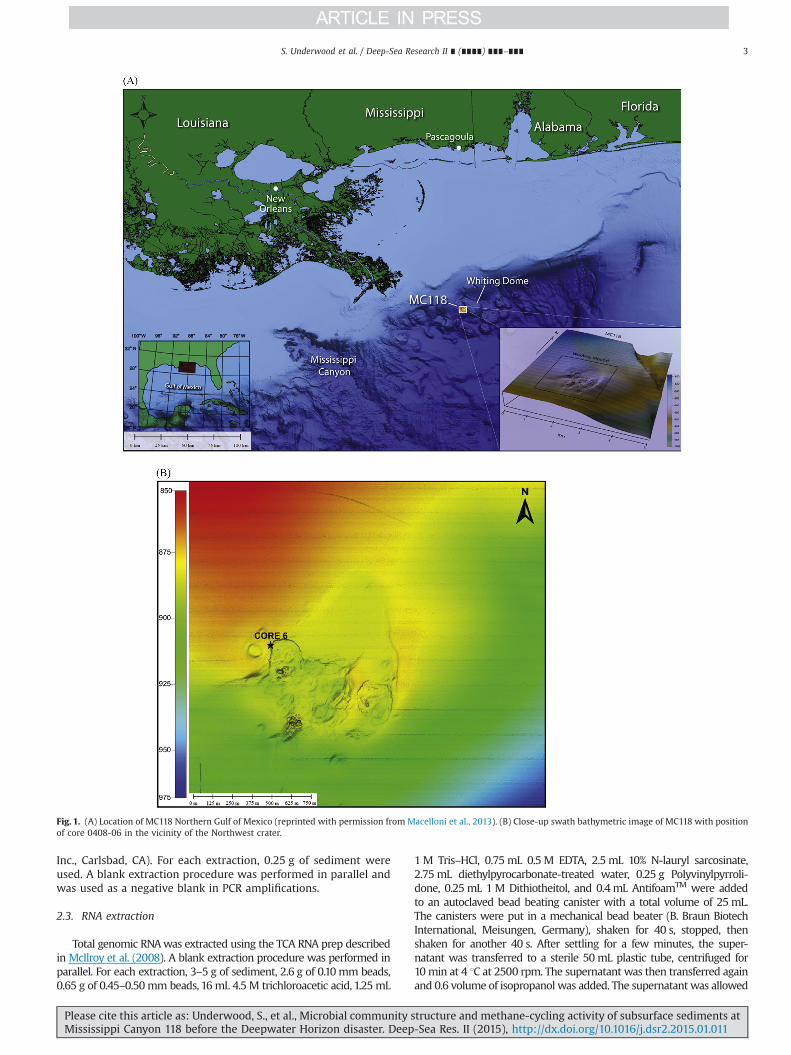
Gravity core 0408-06 was taken by R/V Pelican on April 28, 2008,at 12:58 pm, Latitude 28151027.8996″N, Longitude 88129040.3390″ W,at 879 m water depth to the west of the Northwest Crater (Fig. 1), aregion with active seepage and microbial mats (Lloyd et al., 2010).Upon retrieval of the core on deck, it was cut into 1.5 m sections, splitlengthwise into two halves and immediately sub-sampled formethane, porewater ions, dissolved inorganic carbon, and molecularbiology. Horizontal gas cracks in the sediment column introducedempty spaces between different core sections that were removedfrom the reported depths for all core analyses. For methane mea-surements, 6 mL sediment was added to a serum vial containing1 mL 1 M KOH and immediately crimp sealed with a butyl stopperand shaken to stop biological activity. Methane concentrations weredetermined from the headspace of sediment aliquots at Florida StateUniversity using a Shimadzu mini-2 gas chromatograph equippedwith a flame ionization detector. Methane and dissolved inorganiccarbon (DIC) isotopes were measured by directly injecting micro-volumes of headspace aliquots into a continuous flow Hewlett-Packard 5890 GC equipped with a 6 m Poroplot Q column at 35 1Cand a Finnigan Mat Delta S (Bremen, Germany). Isotope ratios werereported using the standard “del” notation, δ13C (‰)¼(R(sample)/R(PDBstandard)�1)n1000, where R is the ratio of the heavy to light isotope(13C:12C). For porewater analysis, plastic 15 ml tubes were filledcompletely with sediment and centrifuged. The resulting porewaterwas filtered at 0.2 mm, acidified with 10% HCl, and frozen at �20 1Con the ship. An unacidified portion was also frozen for chlorideconcentrations. Sulfate and chloride concentrations were measuredat Florida State University by diluting samples 1:1000 with the eluentbuffer and directly injecting into a 2010i Dionex ion chromatograph(Sunnyvale, CA). For molecular biology, �20 mL samples were placedinto sterile 50 mL plastic tubes and immediately frozen in liquidnitrogen, but had to be transferred to �20 1C in the ship's freezerbefore the end of the cruise. They were returned to the lab packed ina large quantity of dry ice and stored at �80 1C. Due to the constrainsof shipboard sampling and sample demand for other analyses,molecular biology samples were not evenly spaced throughout thesediment column but turned out to represent two samples from thesulfate-rich upper sediment, and five samples from the methane-richdeep sediment layers.
2.2. DNA extraction
Total genomic DNA was extracted using the PowerSoil DNA kitaccording to the manufacturers specifications (MoBio Laboratories,
S. Underwood et al. / Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎2
Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments atMississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i
Inc., Carlsbad, CA). For each extraction, 0.25 g of sediment wereused. A blank extraction procedure was performed in parallel andwas used as a negative blank in PCR amplifications.
2.3. RNA extraction
Total genomic RNAwas extracted using the TCA RNA prep describedin McIlroy et al. (2008). A blank extraction procedure was performed inparallel. For each extraction, 3–5 g of sediment, 2.6 g of 0.10mm beads,0.65 g of 0.45–0.50mm beads, 16mL 4.5 M trichloroacetic acid, 1.25mL
1M Tris–HCl, 0.75mL 0.5M EDTA, 2.5 mL 10% N-lauryl sarcosinate,2.75mL diethylpyrocarbonate-treated water, 0.25 g Polyvinylpyrroli-done, 0.25mL 1M Dithiotheitol, and 0.4mL AntifoamTM were addedto an autoclaved bead beating canister with a total volume of 25mL.The canisters were put in a mechanical bead beater (B. Braun BiotechInternational, Meisungen, Germany), shaken for 40 s, stopped, thenshaken for another 40 s. After settling for a few minutes, the super-natant was transferred to a sterile 50mL plastic tube, centrifuged for10min at 4 1C at 2500 rpm. The supernatant was then transferred againand 0.6 volume of isopropanol was added. The supernatant was allowed
Fig. 1. (A) Location of MC118 Northern Gulf of Mexico (reprinted with permission from Macelloni et al., 2013). (B) Close-up swath bathymetric image of MC118 with positionof core 0408-06 in the vicinity of the Northwest crater.
S. Underwood et al. / Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎ 3
Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments atMississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i

to sit for 2–4 h at -20 1C to precipitate. The tubes were then centrifugedfor 1 h at 4 1C at 3400 rpm. The supernatant was removed and 1.5 mL70% ethanol was added and the tubes were centrifuged for 15min at4 1C at 3400 rpm. The ethanol was added again and the tubes werecentrifuged under the same conditions. The ethanol was poured off andthe pellet allowed to air dry for 3–5 h. The pellet containing RNA wasthen resuspended in 100 mL diethylpyrocarbonate-treated water. To100 mL suspended nucleic acid, 2 mL TURBO DNase and 11 mL 10�TURBO DNase Buffer were added (Ambion Inc., Austin, TX). The mixturewas incubated for 30min at 37 1C. 2 mL DNase Inactivation Reagentwere then added and flicked to mix for 2 min. The mixture wascentrifuged and the supernatant was removed and saved foramplification.
2.4. PCR conditions
For each DNA extraction, a PCR and reverse transcription-PCR(RT-PCR) amplification was attempted for DNA or RNA dilutions of1:1, 1:10, 1:100, 1:1000, a water blank and a DNA positive control,as well as the blank extraction control in a BioRad iCycler ThermalCycler (Bio-Rad Laboratories, Inc., Hercules, CA). A sample withoutreverse transcriptase was included to check the RT-PCR samplesfor DNA contamination. DNA extractions were amplified usingprimers A8F and A1492R for archaeal, and B8F and B1492R forbacterial 16S rRNA genes (Table 1). Each PCR mixture contained2.5 mL TaKaRa FB110X Buffer, 2.0 mL TaKaRa 2.5 mM dNTP mixture,1 mL 10 mg/mL Bovine Serum Albumin, 0.25 mL TaKaRa SpeedSTARHot Start Polymerase (TaKaRa Bio, Inc., Otsu, Japan), 2.0 mL A8f orB8f primer (10 mM), 2.0 mL A1492r or B1492r primer (10 mM), 1 mLextracted DNA and sterile water to a total volume of 25 mL. Onedenaturation cycle at 94 1C for 2 min was followed by 30 cyclesconsisting of 15 s denaturation at 98 1C, 15 s annealing at 58 1C,and 20 s elongation at 72 1C. A final elongation step at 72 1C for10 min was followed by storage of the PCR products at �20 1C.
McrA genes were amplified using the primers ANME-1f, ANME-1r, mcrIRDf and mcrIRDf in each PCR reaction (Table 1). Each PCRcontained all four of these primers mixed together, using the samereaction mixture as stated above. Touchdown PCR was performedusing the following procedure. A denaturation cycle of 94 1C for2 min was followed by 20 cycles, each one consisting of 5 s dena-turation at 95 1C, 15 s of annealing at 60 1C (touchdown, decreas-ing 0.51 per cycle), and 20 s elongation at 72 1C. Thirty additionalPCR cycles were run at constant annealing temperature, andconsisted of 5 s denaturation at 95 1C, 15 s annealing at 55 1C,and 20 s elongation at 72 1C, before concluding with a final 2 minat 72 1C. McrA genes were also amplified using only the mcrIRDfand mcrIRDr primers. DsrAB genes were amplified using the sameprotocol, except the primers Dsr1f and Dsr1r were used (Table 1).
RNA was reverse transcribed and amplified using the sameprimers as were used for DNA amplification (Table 1). Each RT-PCRmixture contained 12.5 mL 2� One Step RNA PCR Buffer, 2 mL of
each primer (10 mM), 0.5 mL Rnase Inhibitor, 0.5 mL TaKaRa Ex TaqHS, 0.5 mL Reverse Transcriptase XL (AMV), 1 mL sample and sterilewater to a total volume of 25 mL. The same temperature cycleswere followed as described above except that a Reverse Tran-scriptase step of 42 1C for 15 min was added to the beginning. Thesame protocol was followed for mcrA and dsrAB genes except thata reverse transcription step was added to the touchdown PCRprotocol described above. All PCR products were stained withethidium bromide dye and viewed on an agarose gel.
2.5. Cloning and sequencing
Gel purification was performed for PCR and RT-PCR productswith the following method. Approximately 24 μL of PCR productwith 5 μL of blue/orange loading dye (Promega) and 2 μl 50�SYBR green dye was run in a 1.8% TAE low melting point agarosegel for 40 min at 100 V in 1� TAE buffer. The correct band wasextracted on a Dark Reader Transilluminator (Clare ChemicalResearch, Inc., Dolores, CO). The extracted band was cleaned usingthe Ultra Clean Gel Spin DNA Purification kit according to themanufacturer's specifications with two exceptions (MoBio Labora-tories, Inc., Carlsbad, CA). At step 15, the spin filter containsethanol, and was centrifuged for 5 min instead of 30 s to ensureethanol removal. At step 17, 30 μL of Elution Buffer was addedinstead of 50 μL to obtain a more concentrated product.
PCR products were ligated into a PCR 2.1 TOPO Cloning Vectorand transformed into chemically competent TOP-10 E. coli cellsusing the TOPO TA Cloning Kit according to the manufacturer'sprotocol with one exception (Invitrogen, Inc., Carlsbad, CA). Thevector was allowed to incubate with the PCR product and saltsolution for 30 min at room temperature instead of 5 min. The OneShot Chemical Transformation procedure was followed accordingto the manufacturer's protocol (Invitrogen, Inc., Carlsbad, CA).25 μL and 100 μL of transformed cells were then plated onto twominimal media agar plates containing bromo-chloro-indolyl-galactopyranoside (X-GAL) and Kanamycin for blue/white screen-ing. After 24 h of incubation at 37 1C, white colonies were picked.The colonies were re-plated after another 24 h for sequencing(GENEWIZ, Inc., South Plainfield, NJ) using M13 (�20) forwardprimer and M13 (�20) reverse primer.
Sequences were edited using Sequencher 4.7 (Gene CodesCorporation, Ann Arbor, MI). Operational taxonomic units (OTUs)were determined using MOTHUR (Schloss et al., 2009). OTUswhere all members had multiple ambiguous sites at the overlapbetween forward and reverse reads were removed due to low qu-ality. Chimeras were identified using Pintail, Greengenes (DeSantiset al., 2006) and also by Blasting 50 and 30 ends separately to checkfor agreement. Bacterial and archaeal 16S genes and cDNA werealigned using Arb (www.arb-home.de), mcrA genes were alignedusing MUSCLE (Edgar, 2004), and dsrAB genes were aligned to anArb database of dsrAB genes (Loy et al., 2009). Full-length
Table 1Primers used in PCR and RT-PCR amplification.
Primer Target cDNA Predicted target group Sequence (50–30) Annealing temp (1C) Ref
A8f 16S Archaea TCC GGT TGA TCC TGC C 58 Teske et al. (2002)A1492r 16S Archaea GGC TAC CTT GTT ACG ACT T 58 Teske et al. (2002)B8f 16S Bacteria AGR GTT TGA TCC TGG CTC AG 58 Teske et al. (2002)B1492r 16S Bacteria CGG CTA CCT TGT TAC GAC TT 58 Teske et al. (2002)mcrIRDf mcrA Methanogens/ANME TWYGACCARATMTGGYT 60 Lever (2008)McrIRDr mcrA Methanogens/ANME ACRTTCATBGCRTARTT 60 Lever (2008)ANME-1f mcrA ANME-1 GACCAGTTGTGGTTCGGAAC 60 Lever (2008)ANME-1r mcrA ANME-1 ATCTCGAATGGCATTCCCTC 60 Lever (2008)Dsr1f dsrA SRB ACS CAY TGG AAG CAC G 54 Wagner et al. (1998)Dsr4r dsrB SRB GTG TAG CAG TTA CCG CA 54 Wagner et al. (1998)
S. Underwood et al. / Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎4
Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments atMississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i
representatives of each 16S OTU were included in the phylogenetictrees. Out of 1741 total clones sequences, 1097 were of highenough quality to be included in the analysis. All sequences shownin trees were deposited at Genbank with accession numbersKP814566-KP814679.
3. Results and discussion
3.1. Geochemistry
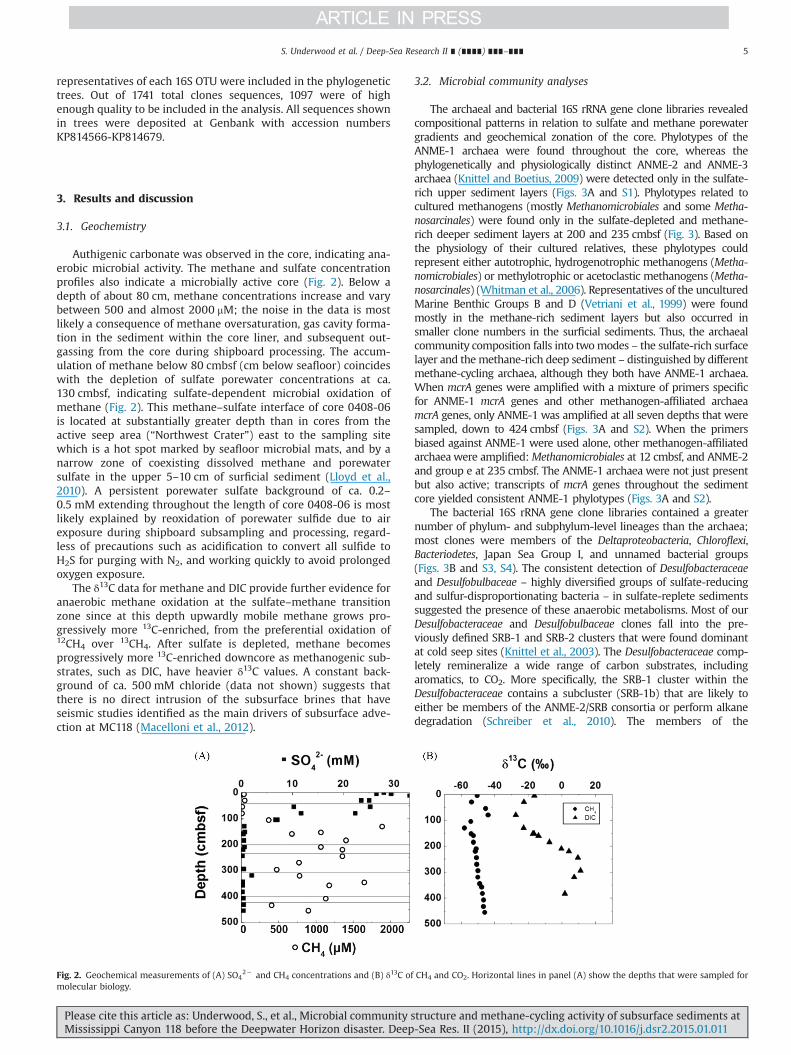
Authigenic carbonate was observed in the core, indicating ana-erobic microbial activity. The methane and sulfate concentrationprofiles also indicate a microbially active core (Fig. 2). Below adepth of about 80 cm, methane concentrations increase and varybetween 500 and almost 2000 mM; the noise in the data is mostlikely a consequence of methane oversaturation, gas cavity forma-tion in the sediment within the core liner, and subsequent out-gassing from the core during shipboard processing. The accum-ulation of methane below 80 cmbsf (cm below seafloor) coincideswith the depletion of sulfate porewater concentrations at ca.130 cmbsf, indicating sulfate-dependent microbial oxidation ofmethane (Fig. 2). This methane–sulfate interface of core 0408-06is located at substantially greater depth than in cores from theactive seep area (“Northwest Crater”) east to the sampling sitewhich is a hot spot marked by seafloor microbial mats, and by anarrow zone of coexisting dissolved methane and porewatersulfate in the upper 5–10 cm of surficial sediment (Lloyd et al.,2010). A persistent porewater sulfate background of ca. 0.2–0.5 mM extending throughout the length of core 0408-06 is mostlikely explained by reoxidation of porewater sulfide due to airexposure during shipboard subsampling and processing, regard-less of precautions such as acidification to convert all sulfide toH2S for purging with N2, and working quickly to avoid prolongedoxygen exposure.
The δ13C data for methane and DIC provide further evidence foranaerobic methane oxidation at the sulfate–methane transitionzone since at this depth upwardly mobile methane grows pro-gressively more 13C-enriched, from the preferential oxidation of12CH4 over 13CH4. After sulfate is depleted, methane becomesprogressively more 13C-enriched downcore as methanogenic sub-strates, such as DIC, have heavier δ13C values. A constant back-ground of ca. 500 mM chloride (data not shown) suggests thatthere is no direct intrusion of the subsurface brines that haveseismic studies identified as the main drivers of subsurface adve-ction at MC118 (Macelloni et al., 2012).
3.2. Microbial community analyses
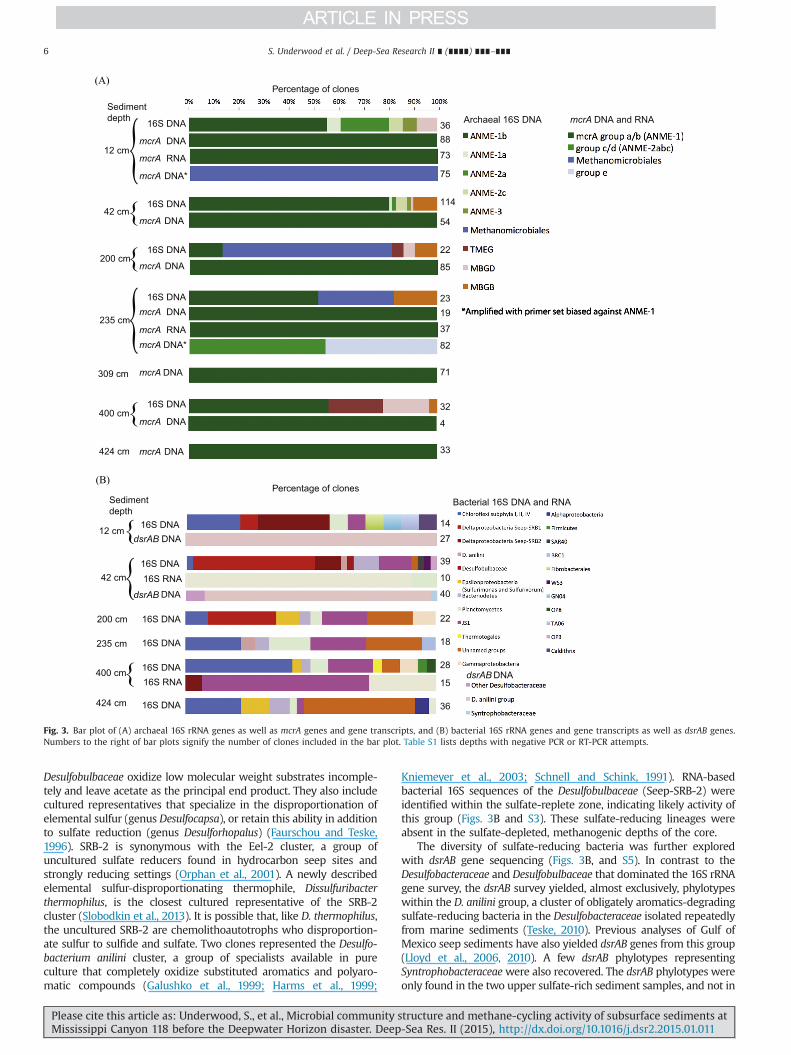
The archaeal and bacterial 16S rRNA gene clone libraries revealedcompositional patterns in relation to sulfate and methane porewatergradients and geochemical zonation of the core. Phylotypes of theANME-1 archaea were found throughout the core, whereas thephylogenetically and physiologically distinct ANME-2 and ANME-3archaea (Knittel and Boetius, 2009) were detected only in the sulfate-rich upper sediment layers (Figs. 3A and S1). Phylotypes related tocultured methanogens (mostly Methanomicrobiales and some Metha-nosarcinales) were found only in the sulfate-depleted and methane-rich deeper sediment layers at 200 and 235 cmbsf (Fig. 3). Based onthe physiology of their cultured relatives, these phylotypes couldrepresent either autotrophic, hydrogenotrophic methanogens (Metha-nomicrobiales) or methylotrophic or acetoclastic methanogens (Metha-nosarcinales) (Whitman et al., 2006). Representatives of the unculturedMarine Benthic Groups B and D (Vetriani et al., 1999) were foundmostly in the methane-rich sediment layers but also occurred insmaller clone numbers in the surficial sediments. Thus, the archaealcommunity composition falls into twomodes – the sulfate-rich surfacelayer and themethane-rich deep sediment – distinguished by differentmethane-cycling archaea, although they both have ANME-1 archaea.When mcrA genes were amplified with a mixture of primers specificfor ANME-1 mcrA genes and other methanogen-affiliated archaeamcrA genes, only ANME-1 was amplified at all seven depths that weresampled, down to 424 cmbsf (Figs. 3A and S2). When the primersbiased against ANME-1 were used alone, other methanogen-affiliatedarchaea were amplified:Methanomicrobiales at 12 cmbsf, and ANME-2and group e at 235 cmbsf. The ANME-1 archaea were not just presentbut also active; transcripts of mcrA genes throughout the sedimentcore yielded consistent ANME-1 phylotypes (Figs. 3A and S2).
The bacterial 16S rRNA gene clone libraries contained a greaternumber of phylum- and subphylum-level lineages than the archaea;most clones were members of the Deltaproteobacteria, Chloroflexi,Bacteriodetes, Japan Sea Group I, and unnamed bacterial groups(Figs. 3B and S3, S4). The consistent detection of Desulfobacteraceaeand Desulfobulbaceae – highly diversified groups of sulfate-reducingand sulfur-disproportionating bacteria – in sulfate-replete sedimentssuggested the presence of these anaerobic metabolisms. Most of ourDesulfobacteraceae and Desulfobulbaceae clones fall into the pre-viously defined SRB-1 and SRB-2 clusters that were found dominantat cold seep sites (Knittel et al., 2003). The Desulfobacteraceae comp-letely remineralize a wide range of carbon substrates, includingaromatics, to CO2. More specifically, the SRB-1 cluster within theDesulfobacteraceae contains a subcluster (SRB-1b) that are likely toeither be members of the ANME-2/SRB consortia or perform alkanedegradation (Schreiber et al., 2010). The members of the
Fig. 2. Geochemical measurements of (A) SO42� and CH4 concentrations and (B) δ13C of CH4 and CO2. Horizontal lines in panel (A) show the depths that were sampled for
molecular biology.
S. Underwood et al. / Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎ 5
Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments atMississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i
Desulfobulbaceae oxidize low molecular weight substrates incomple-tely and leave acetate as the principal end product. They also includecultured representatives that specialize in the disproportionation ofelemental sulfur (genus Desulfocapsa), or retain this ability in additionto sulfate reduction (genus Desulforhopalus) (Faurschou and Teske,1996). SRB-2 is synonymous with the Eel-2 cluster, a group ofuncultured sulfate reducers found in hydrocarbon seep sites andstrongly reducing settings (Orphan et al., 2001). A newly describedelemental sulfur-disproportionating thermophile, Dissulfuribacterthermophilus, is the closest cultured representative of the SRB-2cluster (Slobodkin et al., 2013). It is possible that, like D. thermophilus,the uncultured SRB-2 are chemolithoautotrophs who disproportion-ate sulfur to sulfide and sulfate. Two clones represented the Desulfo-bacterium anilini cluster, a group of specialists available in pureculture that completely oxidize substituted aromatics and polyaro-matic compounds (Galushko et al., 1999; Harms et al., 1999;
Kniemeyer et al., 2003; Schnell and Schink, 1991). RNA-basedbacterial 16S sequences of the Desulfobulbaceae (Seep-SRB-2) wereidentified within the sulfate-replete zone, indicating likely activity ofthis group (Figs. 3B and S3). These sulfate-reducing lineages wereabsent in the sulfate-depleted, methanogenic depths of the core.
The diversity of sulfate-reducing bacteria was further exploredwith dsrAB gene sequencing (Figs. 3B, and S5). In contrast to theDesulfobacteraceae and Desulfobulbaceae that dominated the 16S rRNAgene survey, the dsrAB survey yielded, almost exclusively, phylotypeswithin the D. anilini group, a cluster of obligately aromatics-degradingsulfate-reducing bacteria in the Desulfobacteraceae isolated repeatedlyfrom marine sediments (Teske, 2010). Previous analyses of Gulf ofMexico seep sediments have also yielded dsrAB genes from this group(Lloyd et al., 2006, 2010). A few dsrAB phylotypes representingSyntrophobacteraceaewere also recovered. The dsrAB phylotypes wereonly found in the two upper sulfate-rich sediment samples, and not in
12 cm
42 cm
200 cm
16S DNA
16S RNA16S DNA
16S DNA
16S DNA
dsrAB DNA
dsrAB DNA
400 cm
424 cm
16S DNA16S RNA
16S DNA
235 cm
{
{
{
Bacterial 16S DNA and RNA
dsrAB DNA
Percentage of clones
27
40
14
39
10
22
18
28
15
36
Sediment depth
12 cm
42 cm
200 cm
400 cm
424 cm
235 cm
309 cm
16S DNA
16S DNA
16S DNA
16S DNA
16S DNA
mcrA DNA
mcrA DNA
mcrA DNA
mcrA DNA
mcrA DNA
mcrA DNA
mcrA DNA
mcrA RNA
mcrA RNA
mcrA DNA*
mcrA DNA*
{
{
{
{
{
Archaeal 16S DNA
Percentage of clones
mcrA DNA and RNA36
114
54
75
73
88
32
71
23
22
82
19
85
37
33
4
Sediment depth
Fig. 3. Bar plot of (A) archaeal 16S rRNA genes as well as mcrA genes and gene transcripts, and (B) bacterial 16S rRNA genes and gene transcripts as well as dsrAB genes.Numbers to the right of bar plots signify the number of clones included in the bar plot. Table S1 lists depths with negative PCR or RT-PCR attempts.
S. Underwood et al. / Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎6
Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments atMississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i

the methane-rich, sulfate-depleted sediment layers where dsrABamplification was unsuccessful (Table S1).
After the Deltaproteobacteria, phylotypes of the Chloroflexi werefound in high clone abundance at MC118 (Figs. 3B and S3).Following the previously proposed subdivision of the Chloroflexiinto four subphyla (Hugenholtz et al., 1998), most phylotypes fromMC118 fall into subdivisions I and II. Subdivision I has beenpopulated over time with cultured genera and species of hetero-trophic, anaerobic filamentous bacteria, and was renamed as theproposed class “Anaerolineae” (Yamada et al., 2006). Subdivision IIcontains the cultured dehalogenating anaerobe Dehalococcoidesethenogenes and related dehalogenating isolates (Moe et al., 2009),as well as phylotypes from shallow and deep marine sedimentsand the terrestrial subsurface.
Another group of bacterial phylotypes detected in the MC118subsurface sediments were members of the phylum Planctomycetes,mostly composed of uncultured phylotypes within the Phycisphaer-aceae, a family proposed on the basis of a phylogenetically distinct,heterotrophic and facultatively anaerobic isolate, Phycisphaera mikur-ensis, from a marine alga (Fukunaga et al., 2009). Anammox bacteriawere not detected. Bacteroidetes, Gammaproteobacteria, and the JS-1group were the most frequently detected phylum-level lineages froma total of about 20 mutually exclusive bacterial lineages in the MC118core (Figs. S3 and S4).
3.3. Methodological issues and RNA-based community analysis
No dsrAB transcripts were detected from any depth, althoughnested amplification was performed. Bacterial 16S rRNA tran-scripts amplified at all depths except 12 cmbsf (Table S1). Archaeal16S rRNA transcripts did not amplify at any depth, but this is notlikely to indicate that archaea are inactive because mcrA tran-scripts were amplified from the same RNA aliquots at depths of 12and 235 cmbsf (Table S1). The lack of archaeal 16S rRNA amplifica-tion was most likely due to our failure to denature rRNA moleculesbefore the reverse transcription step (Sambrook and Russell,2001). It is possible that the reverse archaeal 16S rRNA primersmust access a section of the ribosomal rRNA that is unavailable inits native conformation.
In samples where RNA was available, the results generally sup-ported the communities seen using DNA analysis. At 12 cm and235 cm, mcrA RNA sequences were all from the ANME-1 group, thesame as the mcrA DNA sequences.
4. Conclusions
4.1. Comparing core 0408-06 to other deep sediment cores
The MC118 sediment core 0408-06 provides an example whereANME-1 archaea remain detectable into the subsurface at 4 mdepth, using conventional 16S rRNA gene PCR and cloning as wellas mcrA transcript sequencing. The presence of ANME-1 and othermethanogen-affiliated archaea at MC118 stands in contrast tocloning and sequencing surveys of deep marine subsurface sedi-ment with sulfate–methane transition zones below 30 m sedimentdepth where methanogen-affiliated phylotypes have been elusive(Inagaki et al., 2006; Sørensen and Teske, 2006; Biddle et al.,2006). Subsurface mcrA gene signatures of Methanobrevibacter sp.(Parkes et al., 2005) have been questioned as possible contamina-tion issues (discussed in detail by Lever (2013)). It is likely thatcore 0408-06, with a �1.5 m depth sulfate–methane transitionzone, has a more active methane cycle than deep subsurface cores,and therefore its methane cycling communities are dense enoughto be detected by PCR methods, aided by improved mcrA primerswith better coverage (Lever, 2008). Following the empirical
categorization of cored sediments at MC118, the core 0408-06samples would fall into the “high microbial activity” categorybased on methane–sulfate transition depth and methane pore-water concentrations (Lapham et al., 2008). The combination ofgeochemical indicators and consistent molecular detection ofmethane-cycling microorganisms indicates that core 0408-06 ismore similar to active seepage sediments than to continentalmargin sediment without seepage component.
4.2. Physiological potential of ANME-1 archaea
Previous molecular and geochemical studies of marine andestuarine sediments had provided evidence that the ANME-1archaea were able to persist in methane-rich, sulfate-depletedsediments, and grow as methanogenic archaea if required (Lloydet al., 2011). The possibility of multiple roles and capabilities forthis frequently detected archaeal lineage in the marine methanecycle is intriguing. Here, ANME-1 archaea remain prominentlydetectable throughout the sediment column, and are transcrip-tionally active in both the sulfate-rich surface sediments and themethane-rich deeper sediments. The high concentrations of pore-water methane, the δ13C signature of methanogenesis, the pre-sence of diverse Methanomicrobiales and Methanosarcinales, andthe absence of key genes of sulfate reduction in the deep sedimentindicates that this layer is indeed the methanogenic zone wheresulfate reducers are excluded and ANME-1 archaea would have toadapt their metabolic mode accordingly.
4.3. Sulfate-reducing bacteria as oil spill responders
The bacterial baseline of the sediments that bear the impact ofthe Deepwater Horizon-derived fallout shows evidence for prim-ing of hydrocarbon-degrading potential. The sulfate-reducingbacterial community in the gravity core samples from MC118 iscomprised of members of the Desulfobacteraceae (evidenced by16S rRNA and dsrAB genes), the Desulfobulbaceae (16S rRNA genes)and the D. anilini lineage (the majority of dsrAB genes and a single16S rRNA gene clone). Although the transcription of 16S rRNA fromthese groups indicates viability, they may not have been veryactive since their dsrAB gene transcripts were below detectionlimit. These sulfate-reducing families and clades include numerousbacterial strains and species that respond to hydrocarbon avail-ability by complete oxidation of alkanes and aromatics with sulfateas the electron acceptor (Teske, 2010). It has been shown that theoil-derived fallout on the seafloor near the Macondo wellheadstimulated the sulfate-reducing community in the surficial sedi-ments and catalyzed in situ enrichment of sulfate-reducing bac-teria in fall 2010. For example, the Desulfobacterales proportion ofmetagenomes from sediment collected in September and October2010 near the wellhead increased in relation to distant controlsites (Kimes et al., 2013). Within an extensive monitoring timelinefrom May 2010 to July 2011, clone library detection of members ofthe Desulfobacteraceae and Desulfobulbaceae peaked in mid-October 2010 for sediments collected near the wellhead (Yanget al., 2014). Inferences that connect our pre-2010 sediment phy-lotypes of Desulfobacteraceae and Desulfobulbaceae from theMC118 gravity core with the 2010 oil spill responders have to bequalified since hydrocarbon-degrading capabilities occur onlyscattered throughout these two families. Yet, the cultured mem-bers of the D. anilini lineage are so far exclusively specializedin sulfate-reducing aromatics degradation, and therefore repres-ent the strongest case for an inducible sedimentary reservoir ofspecifically hydrocarbon-degrading sulfate reducers. Salt wedge-induced upward sediment advection, subsurface brine flow, and –
on a smaller scale – tidal pumping and bioturbation can facilitate
S. Underwood et al. / Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎ 7
Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments atMississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i

the exchange between the subsurface microbial reservoir and thesediment–water interface.
4.4. Outlook: Beyond the Deltaproteobacteria
These considerations on microbial hydrocarbon-degrading func-tion have focused on the deltaproteobacterial proportion of bacteriain the MC118 sediments. However, it is important to consider the fulltaxonomic range of benthic microorganisms where relevant func-tionalities could be hiding in plain sight. Degradation of complexbiopolymers and cleavage of strong covalent bonds (C–halogen or C–C) is a recurring theme among the major microbial lineages in theMC118 sediment samples. Members of the Chloroflexi are active inthe production of polysaccharide-degrading exoenzymes (Kragelundet al., 2007), in the fermentation of carbohydrate polymers (Yamadaet al., 2006), and in the reductive dehalogenation of halogenatedcompounds (Moe et al., 2009). Members of the Planctomycetes andBacteroidetes stand out by their genomic potential for carbohydratepolymer utilization and also by cell attachment and capture ofpolysaccharides (Martinez-Garcia et al., 2012). Lineages within theuncultured benthic archaea MCG and MBG-D likely catalyze thehydrolysis and fermentative degradation of proteins and amino acids(Lloyd et al., 2013). Syntrophic associations of hydrogenotrophicmethanogens and bacteria (members of the deltaproteobacterialfamily Syntrophobacteraceae) are capable of complete methanogenicremineralization of n-alkanes in a process of anaerobic microbialalkane cracking at low temperatures (Zengler et al., 1999). The sedi-ments of the Gulf of Mexico and their resident microbial populationshave been exposed to hydrocarbon seepage for millions of years, andthe wide range of uncultured phyla in this survey strongly cautionsagainst closing the book on their anaerobic hydrocarbon degradationcapabilities.
Acknowledgments
We thank the Captain and crew of R/V Pelican for their expertsupport of our sampling campaign at MC118, and the science crew forteamwork and help under challenging circumstances. This work wassupported in part through the BP/Gulf of Mexico Research Initiative tosupport consortium research entitled “Ecosystem Impacts of Oil andGas Inputs into the Gulf (ECOGIG)” administered by the University ofMississippi. During different phases of this project, Sarah Underwood,Laura Lapham, Andreas Teske and Karen Lloyd were also supportedthrough the Gulf of Mexico Gas Hydrate Research Consortium at theUniversity of Mississippi, and by NSF (Rapid Response: The microbialresponse to the Deepwater Horizon Oil Spill, NSF-OCE 1045115). TheGRIIDC dataset IDs for this manuscript are R1.x132.134:0058 and R1.x132.139.005. This is UMCES contribution XXX.
Appendix A. Supporting information
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.dsr2.2015.01.011.
References
Arnosti, C., Ziervogel, K., Yang, T., Teske, A., 2015. Oil-derived marine aggregates –
hot spots of polysaccharide degradation by specialized bacterial communities.Deep-Sea Res. II, in this issue.
Biddle, J.F., Lipp, J.S., Lever, M.A., Lloyd, K.G., Sørensen, K.B., Anderson, R., Fredricks,H.F., Elvert, M., Kelly, T.J., Schrag, D.P., Sogin, M.L., Brenchley, J.E., Teske, A.,House, C.H., Hinrichs, K.-U., 2006. Heterotrophic Archaea dominate sedimen-tary subsurface ecosystems off Peru. Proc. Natl. Acad. Sci. USA 103, 3846–3851.
Biddle, J.F., White, J.R., Teske, A.P., House, C.H., 2011. Metagenomics of the subsur-face Brazos-Trinity Basin (IODP site 1320): comparison with other sedimentand pyrosequenced metagenomes. ISME J. 5, 1038–1047.
Bowles, M.W., Samarkin, V.A., Bowles, K.M., Joye, S.B., 2011. Weak coupling betweensulfate reduction and the anaerobic oxidation of methane in methane-richseafloor sediments during ex-situ incubation. Geochim. Cosmochim. Acta 75,500–519.
Briggs, B.R., Inagaki, F., Morono, Y., Futagami, T., Huguet, C., Rosell-Mele, A.,Lorenson, T.D., Colwell, F.S., 2012. Bacterial dominance in subseafloor sedimentscharacterized by methane hydrates. FEMS Microbiol. Ecol. 81, 88–98.
Colwell, F.S., Boyd, S., Delwiche, M.E., Reed, D.W., Phelps, T.J., Newby, D.T., 2008.Estimates of biogenic methane production rates in deep marine sediments atHydrate Ridge, Cascadia Margin. Appl. Environ. Microbiol. 74, 3444–3452.
DeSantis, T.Z., Hugenholtz, P., Larsen, N., Rojas, M., Brodie, E.L., Keller, K., et al., 2006.Greengenes, a chimera-checked 16S rRNA gene database and workbenchcompatible with ARB. Appl. Environ. Microbiol. 72, 5069–5072.
D’Hondt, S., Jørgensen, B.B., Miller, D.J., Batzke, A., Blake, R., Cragg., B.A., et al., 2004.Distributions of microbial activities in deep subseafloor sediments. Science 306,2216–2221.
Edgar, R.C., 2004. MUSCLE: multiple sequence alignment with high accuracy andhigh throughput. Nucleic Acids Res. 32, 1792–1797.
Faurschou, M.I., Teske, A., 1996. Desulforhopalus vacuolatus gen. nov., sp. nov., a newmoderately psychrophilic sulfate-reducing bacterium with gas vacuoles iso-lated from a temperate estuary. Arch. Microbiol. 166, 160–168 (S).
Fukunaga, Y., Kurahashi, M., Sakiyama, Y., Ohuchi, M., Yokota, A., Harayama, S.,2009. Phycisphaera mikurensis, gen. nov., sp. nov., isolated from a marine alga,and proposal of Phycispaeraceae fam.nov., Phycisphaerales ord. nov., and Phyci-sphaerae classis nov. in the phylum Planctomycetes. J. Gen. Appl. Microbiol. 55,267–275.
Galushko, A., Minz, D., Schink, B., Widdel, F., 1999. Anaerobic degradation ofnaphthalene by a pure culture of a novel type of marine sulphate-reducingbacterium. Environ. Microbiol. 1, 415–420.
Hallam, S.J., Putnam, N., Preston, C.M., Detter, J.C., Rokhsar, D., Richardson, P.,DeLong, E.F., 2004. Reverse methanogenesis: testing the hypothesis withenvironmental genomics. Science 305, 1457–1462.
Harms, G., Zengler, K., Rabus, R., Aeckersberg, F., Minz, D., Rossello-Mora, R.,Widdel, F., 1999. Anaerobic oxidation of o-xylene, m-xylene, and homologousalkylbenzenes by new types of sulfate-reducing bacteria. Appl. Environ. Micro-biol. 65, 999–1004.
Hugenholtz, P., Goebel, B.M., Pace, N.R., 1998. Impact of culture-independentstudies on the emerging phylogenetic view of bacterial diversity. J. Bacteriol.180, 4765–4774.
Inagaki, F., Nunoura, T., Nakagawa, S., Teske, A., Lever, M.A., Lauer, A., Suzuki, M.,Takai, K., Delwiche, M., Colwell, F.S., Nealson, K.H., Horikoshi, K., D’Hondt, S.L.,Jørgensen, B.B., 2006. Biogeographical distribution and diversity of microbes inmethane hydrate-bearing deep marine sediments on the Pacific Ocean Margin.Proc. Natl. Acad. Sci. USA 103, 2815–2820.
Joye, S.B., Samarkin, V.A., Orcutt, B.N., MacDonald, IR., Hinrichs, K.-U., Elvert, M.,Teske, A., Lloyd, K.G., Lever, M.A., Montoya, J.P., Meile, C.D., 2009. Metabolicvariability in seafloor brines revealed by carbon and sulphur dynamics. Nat.Geosci. 2, 349–354.
Kallmeyer, J., Smith, D.C., Spivack, A.J., D’Hondt, S., 2008. New cell extractionprocedure applied to deep subsurface sediments. Limnol. Oceanogr. Methods 6,236–245.
Kennicutt II, M.C., Brooks, J.M., Denoux, G.J., 1988. Leakage of deep, reservoiredpetroleum to the near surface of the Gulf of Mexico continental slope. Mar.Chem. 24, 39–59.
Kimes, N.E., Callaghan, A.V., Aktas, D.F., Smith, W.L., Sunner, J., Golding, B.T.,Drozdowska, M., Hazen, T.C., Suflita, J.M., Morris, P.J., 2013. Metagenomicanalysis and metabolite profiling of deep-sea sediments from the Gulf ofMexico following the Deepwater Horizon oil spill. Front. Microbiol. 4, 50.
Klein, M., Friedrich, M., Roger, A.J., Hugenholtz, P., Fishbain, S., et al., 2001. Multiplelateral transfers of dissimilatory sulfite reductase genes between major lineagesof sulfate-reducing prokaryotes. J. Bacteriol. 183, 6028–6035.
Kniemeyer, O., Fischer, T., Wilkes, H., Glöckner, F.O., Widdel, F., 2003. Anaerobicdegradation of ethylbenzene by a new type of marine sulfate-reducingbacterium. Appl. Environ. Microbiol. 69, 760–768.
Knittel, K., Boetius, A., Lemke, A., Eilers, H., Lochte, K., Pfannkuche, O., Linke, P.,Amann, R., 2003. Activity, distribution, and diversity of sulfate reducers andother bacteria in sediments above gas hydrate (Cascadia Margin, OR). Geomi-crobiol. J. 20, 269–294.
Knittel, K., Boetius, A., 2009. Anaerobic oxidation of methane: progress with anunknown process. Annu. Rev. Microbiol. 63, 311–334.
Kragelund, C., Levantesi, C., Borger, A., Thelen, K., Eikelboom, D., Tandoi, V., et al.,2007. Identity, abundance and ecophysiology of filamontous Chloroflexi speciespresent in activated sludge treatment plants. FEMS Microbiol. Ecol. 59, 671–682.
Lanoil, B.D., Sassen, R., La Duc, M.T., Sweet, S.T., Nealson, K.H., 2001. Bacteria andarchaea physically associated with Gulf of Mexico gas hydrates. Appl. Environ.Microbiol. 67, 5143–5153.
Lapham, L.L., Chanton, J.P., Martens, C.S., Sleeper, K., Woolsey, J.R., 2008. Microbialactivity in surficial sediments overlying acoustic wipeout zones at a Gulf ofMexico cold seep. Geochem. Geophys. Geosyst. 9, Q06001. http://dx.doi.org/10.1029/2008GC001944.
Lever, M.A., 2008. Anaerobic Carbon Cycling Pathways in the Deep SubseafloorInvestigated via Functional Genes, Chemical Gradients, Stable Carbon Isotopes,and Thermodynamic Calculations. University of North Carolina, Chapel Hill, NC.
Lever, M.A., 2013. Functional gene surveys from ocean drilling expeditions – areview and perspective. FEMS Microbiol. Ecol. 84, 1–23.
S. Underwood et al. / Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎8
Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments atMississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i

Lloyd, K.G., Lapham, L., Teske, A., 2006. An anaerobic methane-oxidizing commu-nity of ANME-1b archaea in hypersaline Gulf of Mexico sediments. Appl.Environ. Microbiol. 72, 7218–7230.
Lloyd, K.G., Albert, D., Biddle, J.F., Chanton, L., Pizarro, O., Teske, A., 2010. Spatialstructure and activity of sedimentary microbial communities underlying aBeggiatoa spp. mat in a Gulf of Mexico hydrocarbon seep. PLoS One 5 (1), e8738.http://dx.doi.org/10.1371/journal.pone.0008738.
Lloyd, K.G., Alperin, M., Teske, A., 2011. Environmental evidence for net methaneproduction and oxidation in putative Anaerobic MEthanotrophic (ANME)archaea. Environ. Microbiol. 13, 2548–2564.
Lloyd, K.G., Schreiber, L., Petersen, D.G., Kjeldsen, K.U., Lever, M.A., Steen, A.D.,Stepanauskas, R., Richter, M., Kleindienst, S., Lenk, S., Schramm, A., Jørgensen,B.B., 2013. Predominant archaea in marine sediments degrade detrital proteins.Nature 496, 215–218.
Loy, A., Duller, S., Baranyl, C., Mussman, M., Ott, J., Beja, O., et al., 2009. Reversedissimilatory sulfite reductase as phylogenetic marker for a subgroup of sulfur-oxidizing prokaryotes. Environ. Microbiol. 11, 289–299.
Luton, P.E., Wayne, J.M., Sharp, R.J., Riley, P.W., 2002. The mcrA gene as analternative to 16S rRNA in the phylogenetic analysis of methanogen populationsin landfill. Microbiology 148, 3521–3530.
Martinez-Garcia, M., Brazel, D.M., Swan, B.K., Arnosti, C., Chain, P.S.G., et al., 2012.Capturing single cell genomes of active polysaccharide degraders: an unex-pected contribution of Verrucomicrobia. PLoS One 7 (4), 335314. http://dx.doi.org/10.1371/journal.pone.0035314.
Macelloni, L., Simonetti, A., Knapp, J.H., Knapp, C.C., Lutken, C.B., Lapham, L.L., 2012.Multiple resolution seismic imaging of a shallow hydrocarbon plumbing system,Woolsey Mound, Northern Gulf of Mexico. Mar. Petrol. Geol. 38, 128–142.
Macelloni, L., Brunner, C.A., Caruso, S., Lutken, C.B., D’Emidio, M., Lapham, L.L., 2013.Spatial distribution of seafloor geo-biological and geochemical processes asproxies of fluid flow regime and evolution of a carbonate/hydrates mound inthe northern Gulf of Mexico. Deep-Sea Res. I 74, 25–38.
McIlroy, S., Porter, K., Seviour, R.J., Tillett, D., 2008. Method for SimultaneousIsolation of Microbial RNA and DNA from problematic populations. Appl.Environ. Microbiol. 74, 6806–6807.
Mills, H.J., Martinez, R.J., Story, S., Sobecky, P.A., 2004. Identification of members ofthe metabolically active microbial populations associated with Beggiatoaspecies mat communities from Gulf of Mexico cold-seep sediments. Appl.Environ. Microbiol. 70, 5447–5458.
Mills, H.J., Hodges, C., Wilson, K., MacDonald, I.R., Sobecky, P.A., 2003. Microbialdiversity in sediments associated with surface-breaching gas hydrate moundsin the Gulf of Mexico. FEMS Microbiol. Ecol. 46, 39–52.
Moe, W.M., Yan, J., Nobre, M.F., da Costa, M.S., Rainey, F.A., 2009. Dehalogenimonaslykanthroporepellens gen. nov., sp. nov. a reductively dehalogenating bacteriumisolated from chlorinated solvent-contaminated groundwater. J. Int. Syst. Evol.Microbiol 59, 2692–2697.
Orphan, V.J., Hinrichs, K.-U., Ussler III, W., Paull, C.K., Taylor, L.T., Sylva, S.P., et al.,2001. Comparative analysis of methane-oxidizing archaea and sulfate-reducingbacteria in anoxic marine sediments. Appl. Environ. Microbiol. 67, 1922–1934.
Redmond, M.C., Valentine, D.L., 2012. Natural gas and temperature structured amicrobial community response to the Deepwater Horizon oil spill. Proc. Natl.Acad. Sci. USA 109, 20292–20297.
Sambrook, J., Russell, D.W., 2001. Molecular Cloning: A Laboratory Manual. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY.
Sassen, R., Roberts, H.H., Carney, R., Milkov, A.V., DeFreitas, D.A., Lanoil, B., Zhang, C.,2004. Free hydrocarbon gas, gas hydrate, and authigenic minerals in chemo-synthetic communities of the northern Gulf of Mexico continental slope:relation to microbial processes. Chem. Geol. 205, 195–217.
Schnell, S., Schink, B., 1991. Anaerobic aniline degradation via reductive deamina-tion of 4-aminobenzoyl-CoA in Desulfobacterium anilini. Arch. Microbiol. 155,183–190.
Schloss, P.D., Westcott, S.L., Ryabin, T., Hall, J.R., Hartmann, M., Hollister, E.B., et al.,2009. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl.Environ. Microbiol. 75, 7537–7541.
Schreiber, L., Holler, T., Knittel, K., Meyerdierks, A., Amann, R., 2010. Identification ofthe dominant sulfate-reducing bacterial partner of anaerobic methanotrophs ofthe ANME-2 clade. Environ. Microbiol. 12, 2327–2340.
Slobodkin, A.I., Reysenbach, A.-L., Slobodkina, G.B., Kolganova, T.V., Kostrikina, N.A.,Bonch-Osmolovskaya, E.A., 2013. Dissulfuribacter thermophilus gen. nov., sp.nov., a thermophilic, autotrophic, sulfur-disproportionating, deeply branchingdeltaproteobacterium from a deep-sea hydrothermal vent. Int. J. Syst. Evol.Microbiol. 63, 1967–1971.
Springer, E., Sachs, M.S., Woese, C.R., Boone, D.R., 1995. Partial gene sequences forthe A subunit of methyl-coenzyme M reductase (mcrA) as a phylogenetic toolfor the family Methanosarcinaceae. Int. J. Syst. Bacteriol. 45, 554–559.
Sørensen, K.B., Teske, A., 2006. Stratified communities of active archaea in deepmarine subsurface sediments. Appl. Environ. Microbiol. 72, 4596–4603.
Teske, A., Hinrichs, K.-U., Edgcomb, V., de Vera Gomez, A., Kysela, D., Sylva, S.P.,Sogin, M.L., Jannasch, H.W., 2002. Microbial diversity in hydrothermal sedi-ments in the Guaymas Basin: evidence for anaerobic methanotrophic commu-nities. Appl. Environ. Microbiol. 68, 1994–2007.
Teske, A., 2010. Sulfate-reducing and methanogenic hydrocarbon-oxidizing microbialcommunities in the marine environment. Part 21: Microbial communities basedon hydrocarbons, oils and fats: Natural habitats. In: Timmis, Kenneth (Ed.), Hand-book of Hydrocarbon and Lipid Microbiology. Springer, Heidelburg, Germany,pp. 2203–2223. http://dx.doi.org/10.1007/978-3-540-77587-4_160.
Vetriani, C., Jannasch, H.W., MacGregor, B.J., Stahl, D.A., Reysenbach, A.-L., 1999.Population structure and phylogenetic characterization of marine benthicarchaea in deep-sea sediments. Appl. Environ. Microbiol. 65, 4375–4384.
Wagner, M., Roger, A.J., Flax, J.L., Brusseau, G.A., Stahl, D.A., 1998. Phylogeny ofdissimilatory sulfite reductases supports an early origin of sulfate respiration.J. Bacteriol. 180, 2975–2982.
Whitman, W.B., Bowen, T.L., Boone, D.R., 2006. The methanogenic bacteria. 3rdedition In: Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt,E. (Eds.), The Prokaryotes, vol. 3, 3. Springer, New York, pp. 165–207.
Willerslev, E, Cooper, A., 2005. Ancient DNA. Proc. R. Soc. B 272, 3–16.Yamada, T., Sekiguchi, Y., Hanada, S., Imachi, H., Ohashi, A., Harada, H., Kamagata, Y.,
2006. Anaerolinea thermolimosa sp. nov., Levilinea saccharolytica gen. nov, sp.nov., and Leptolinea tardivitalis gen. nov., sp. nov., novel filamentous anaerobes,and description of the new classes Anaerolinea classis nov., and Caldilineaeclassis nov., in the bacterial phylum Chloroflexi. Int. J. Syst. Evol. Microbiol. 56,1331–1340.
Yang, T., Nigro, L.M., Gutierrez, T., D’Ambrosio, L., Joye, S.B., Highsmith, R., Teske, A.,2014. Pulsed blooms and persistent oil-degrading bacterial populations in thewater column during and after the Deepwater Horizon blowout. Deep-Sea Res.II , http://dx.doi.org/10.1016/j.dsr2.2014.01.014 (early online).
Yang, T., Speare, K., Joye, S., Teske. A., 2014. Bacterial community dynamics in oil-polluted seafloor sediment (May 2010–July 2011). Poster Presentation at the2014 Gulf of Mexico Oilspill and Ecosystem Science Conference, January 27–30,Mobile, AL.
Zengler, K., Richnow, H.H., Rosselló-Mora, R., Michaelis, W., Widdel, F., 1999.Methane formation from long-chain alkanes by anaerobic microorganisms.Nature 401, 266–269.
Zverlov, V., Klein, M., Lücker, S., Friedrich, M.W., Kellermann, J., Stahl, D.A., Loy, A.,Wagner, M., 2005. Lateral gene transfer of dissimilatory (bi)sulfite reductaserevisited. J. Bacteriol. 187, 2203–2208.
S. Underwood et al. / Deep-Sea Research II ∎ (∎∎∎∎) ∎∎∎–∎∎∎ 9
Please cite this article as: Underwood, S., et al., Microbial community structure and methane-cycling activity of subsurface sediments atMississippi Canyon 118 before the Deepwater Horizon disaster. Deep-Sea Res. II (2015), http://dx.doi.org/10.1016/j.dsr2.2015.01.011i




















