
Micro-freshwater Gastropods at Çatalhöyük as Environmental Indicators AND Unio Shells from...
14
Humans and Landscapes of Çatalhöyük Reports from the 2000–2008 Seasons
Transcript of Micro-freshwater Gastropods at Çatalhöyük as Environmental Indicators AND Unio Shells from...

Humans and Landscapes of ÇatalhöyükReports from the 2000–2008 Seasons

Humans and Landscapes of ÇatalhöyükReports from the 2000–2008 Seasons
Çatalhöyük Research Project Series Volume 8
Edited by
Ian Hodder
BRITISH INSTITUTE AT ANKARABIAA Monograph No. 47
Monumenta Archaeologica 29Cotsen Institute of Archaeology Press

Published by the British Institute at Ankarac/o The British Academy, 10 Carlton House Terrace, London SW1Y 5AH
and the Cotsen Institute of Archaeology at UCLA 308 Charles E. Young Dr. N, A163 Fowler, Los Angeles, CA 90095
UK distributor: Oxbow Books, Park End Place, Oxford OX1 1HNUSA distributor: University of New Mexico Press, 1717 Roma Ave. NE, Albuquerque, NM 87106
Library of Congress Cataloging-in-Publication Data
ISBN 978 1 898249 30 6
BIAA Monograph no. 47 ISSN 0969-9007 (British Institute at Ankara) Monumenta Archaeologica 29 (Cotsen Institute of Archaeology at UCLA)
Copyright ©2013 British Institute of Archaeology at Ankara; Regents of the University of CaliforniaAll rights reserved.
The British Institute at Ankara (BIAA), one of the British Academy’s overseas institutes, has promoted world-class research in Turkey and the Black Sea region in the fi elds of history, archaeology and related social sciences since its foundation in 1947. The BIAA publishes scholarly monographs relating to the archaeology and history of Turkey, with a particular emphasis on publishing the results of Institute-funded research.
The British Institute at Ankara Publications Editorial Board
Dr Gülnur Aybet Ms Gina Coulthard Dr Shahina FaridDr Tamar HodosProfessor Roger MatthewsProfessor Stephen Mitchell Dr Claire Norton Dr Lutgarde Vandeput
Typeset by Campbell Archaeological Services, Lymm CheshirePrinted by . . . .

81
Freshwater molluscs, both Gastropods (snails) and bivalves, form the largest component of the malacofauna at Çatal-höyük. Freshwater bivalves are common in many archaeo-logical sites, where they are often found in middens (e.g. Claassen 1998), as is also true at Çatalhöyük (Chapter 16). The occurrence of freshwater gastropods, usually inedible, is more ambiguous. In a few cases they served as shell beads (Kuhn et al. 2004), but more often they were brought to the site inadvertently, either as construction material (Bar-Yosef & Heller 1987) or along with drinking water (Bar-Yosef Mayer 2005).
In sites located on the banks of rivers or lakes, this fauna may serve to reconstruct the natural environment of the site during its existence by studying the changes in composition of the malacofauna, as well as size changes of some shell species whose size responds to climatic changes (White et al. 2008; Mienis & Ashkenazi 2011). Because Çatalhöyük did not lie directly on a riverbank or lake shore, however, the en-vironmental information provided from its molluscan fauna has a somewhat different nature.
The freshwater shells discovered at the site are from the vicinity, including the nearby Çarsamba River, as well as lakes and marshes where mud was collected for construc-tion materials and pottery production. Some of the shells could also have been brought along with macrophyte plants that were abundant at the site (Rosen 2005; Chapter 9). The ecological information attributed to these species enhances the understanding of the environment in the vicinity of the site during its occupation. In this research, we divided the molluscs according to their ecological environment and found two major groups representing two major environ-ments: one of high energy and highly oxygenated water, a lotic environment, and another of low energy and more vari-ably oxygenated water, a lentic environment. We compared this information with the stratigraphic origin of the finds and found that resource exploitation throughout the Çatalhöyük sequence relied on both lentic and lotic environments. This contribution might enhance our understanding of the site’s surrounding during its occupation.
Material and methods
Shells <10mm in size were recovered from flotation of the ex-cavated sediments in 1–4mm mesh screens. These shells were examined under stereoscopic microscope and their identifi-cation was based on comparisons with various publications (Zhadin 1965, Schütt 1965; Schütt & Şeşen 1992; Yıldırım & Schütt 1996; Yıldırım 1999; Glöer & Meier-Brook 2003; Yıldırım et al. 2006a, b, c), as well as to a modern reference collection of the first author’s.
To get a clear idea of the environment in each level of the excavation, we chose to concentrate on a representative sample deriving from secure contexts with clear stratigraphic relations, mostly ‘history houses’ (Hodder 2010). Counting of the species at each of the 14 levels in the South Area (from 17 buildings and two spaces) was based on minimum number of individuals (MNI). MNI was determined by counting only apices of gastropods and ignoring the numerous fragments and broken shells in the samples. It is worth noting that some species included in this study are juvenile specimens of larger gastropods that are within the ‘micro-shell’ size range (e.g. Viviparus, Lymnaea, and Planorbarius). Most other species, however, are just a few millimeters high in their fully-grown adult stage (Fig. 5.1).
Results
In total, 1850 specimens were examined from 14 levels in the South Area belonging to 13 freshwater gastropod species (Fig. 5.2). The species identified were first qualified in terms of their biogeographic origins, then according to their eco-logical preferences, and finally within their archaeological/stratigraphic provenance.
The recent known zoogeographic origin of 54 per cent of the studied species is Palaearctic, 31 per cent is Holarctic, and 15 per cent are endemic shells (Gümüş & Bar-Yosef Mayer in press). Some of the freshwater prosobranch gastropods (those that breathe with gills) represent different habitats. Bythinella turca and Fagotia esperi, which are stenoec mollusks (mean-ing that they have a limited range of tolerance to environ-
Chapter 5
Micro-freshwater Gastropods at Çatalhöyük as Environmental Indicators
Burçin A. Gümüş & Daniella E. Bar-Yosef Mayer

Volume 8: Humans and Landscapes of Çatalhöyük
82
Figu
re 5
.1 T
he m
icro
-fres
hwat
er g
astro
pod
spec
ies i
dent
ified
at Ç
atal
höyü
k. (a
, b, c
, f, g
, i, j
, k, l
: pho
togr
aphe
d by
Jas
on Q
uinl
anr;
d: p
hoto
grap
hed
by P
eter
G
löer
; e: a
fter G
löer
200
2 (D
anub
e Ri
ver)
; h: p
hoto
grap
hed
by G
ümüş
and
De
Cup
ere,
Sag
alas
sos r
esea
rch
Proj
ect).

Chapter 5: Micro-freshwater Gastropods
83
mental changes in their ecosystem), represent high-energy and highly-oxygenated environments, in particular, rivers and floodplain ponds. Bythinella turca, an endemic species of Turkish malacofauna, is described from several perennial springs including Cire (Balkırı), Eğirdir-Isparta and Kuşbaba Spring, Başgöz Spring, Aziziye Köyü, Can Trout Springs and Burdur (Radoman 1976; Yıldırım 1998; Yıldırım et al. 2001). These prosobranch gastropods prefer cold-running waters that are highly oxygenated (i.e. lotic habitats). Another set of species, Galba truncatula, Gyraulus albus, Gyraulus cris-tata, Lymnaea stagnalis, Planorbarius corneus, Planorbis carinatus, Radix auricularia and Stagnicola palustris, which are euryoec mollusks (meaning that they have a wide toler-ance range to environmental changes), are mostly freshwater pulmonates (that breathe with a ‘lung’) that represent low-energy environments with slow-running waters (i.e. lentic habitats) with variable levels of dissolved oxygen such as eu-trophic swamps and lakes (Boycott 1936; Fretter 1968; Pen-nak 1989; Yıldırım & Şeşen 1994; Yıldırım & Schütt 1996; Gümüş & Bar-Yosef Mayer 2011 in press).
The most abundant species in this study are three proso-branch species, Valvata piscinalis (n=908), Viviparus viviparus (n=353), and Theodoxus cf. heldreichi heldreichi (n=215). These three species can represent both lentic and lotic environments, however, and therefore cannot serve as environmental indicators. Valvata piscinalis is reported from a variety of running water environments such as rivers to quite small brooks, canals, ditches and lakes, including a few low-lying mountain lakes (Boycott 1936). It is a common species in running water of all kinds, provided the current is not too fast, and is rare in closed ponds, on the stony shore of lakes, in reed-beds and in submerged vegetation in deeper water (Macan 1949; 1950). This species is also reported from standing or slow moving water with rich vegetation such as mud bottoms (Gittenberger & Janssen 1998).
Viviparus viviparus has been reported from reservoirs with standing water and a considerable accumulation of hu-mous acids (pH: 4.8; Fretter & Alastair 1962), thus belonging to eutrophic lentic freshwater systems (Şahin 2008). Yet other reports describe this species as living preferably in rivers, ca-
Figure 5.2. The abundance of the micro-freshwater gastropods in the studied sample.
Gyraulus (Armiger) crista (Linnaeus, 1758)
Planorbarius corneus (Linnaeus, 1758)
Planorbis carinatus (O.F. Müller, 1774)
Gyraulus albus (O.F. Müller, 1774)
St i l l t i (O F Müll 1774)
Galba truncatula (O.F. Müller, 1774)
Radix auricularia (Linnaeus, 1758)
Fagotia esperi (Férussac, 1823)
Lymnaea stagnalis (Linnaeus, 1758)
Stagnicola palustris (O.F. Müller, 1774)
Bythinella cf. turca (Radoman, 1976)
Valvata piscinalis (O.F. Müller, 1774)
g p ( )
Theodoxus cf. heldreichi heldreichi (Martens, 1879)
Viviparus viviparus (Linnaeus, 1758)
0 200 400 600 800 1000
Fagotia esperi
Stagnicola palustris

Volume 8: Humans and Landscapes of Çatalhöyük
84
Tabl
e 5.
1. N
umbe
r of t
axa,
num
ber o
f spe
cim
ens a
nd h
abita
t pre
fere
nces
of m
icro
-fres
hwat
er g
astro
pods
in th
e st
udy
area
.
Lev
el
Loc
atio
n N
umbe
r of
Tax
a n
Len
tic S
yste
ms*
L
otic
Sys
tem
s**
Len
tic &
Lot
ic S
yste
ms*
**
SOU
TH
T
B.1
0 4
30
0 0.
00%
0
0.00
%
30
100.
00%
SOU
TH
S
B.4
4 11
52
5 58
11
.05%
22
4.
19%
44
5 84
.76%
SOU
TH
R
B.5
6 6
34
2 5.
88%
1
2.94
%
31
91.1
8%
SOU
TH
R
B.4
2 7
93
2 2.
15%
4
4.30
%
87
93.5
5%
SOU
TH
Q
B.6
5 10
19
8 18
9.
09%
0
0.00
%
180
90.9
1%
SOU
TH
Q
B.5
3 3
8 2
25.0
0%
0 0.
00%
6
75.0
0%
SOU
TH
P
B.7
5 9
240
21
8.75
%
17
7.08
%
202
84.1
7%
SOU
TH
O
B.7
6 4
7 0
0.00
%
0 0.
00%
7
100.
00%
SOU
TH
O
B.7
9 3
14
1 7.
14%
0
0.00
%
13
92.8
6%
SOU
TH
O
B.8
0 3
37
3 8.
11%
0
0.00
%
34
91.8
9%
SOU
TH
N
B.8
9 1
1 0
0.00
%
0 0.
00%
1
100.
00%
SOU
TH
?M
B
.8
1 5
0 0.
00%
0
0.00
%
5 10
0.00
%
SOU
TH
?M
B
.24
3 6
2 33
.33%
0
0.00
%
4 66
.67%
SOU
TH
L
B.6
2
14
0 0.
00%
0
0.00
%
14
100.
00%
SOU
TH
K
B.1
7 3
48
2 4.
17%
0
0.00
%
46
95.8
3%
SOU
TH
J
B.2
3 2
2 0
0.00
%
0 0.
00%
2
100.
00%
SOU
TH
J
B.1
8 2
2 0
0.00
%
0 0.
00%
2
100.
00%
SOU
TH
H
Sp.1
99
2 11
0
0.00
%
0 0.
00%
11
10
0.00
%
SOU
TH
G
Sp.1
81
9 57
5 49
8.
52%
40
6.
96%
48
6 84
.52%
TO
TA
L
85
18
50
160
8.65
%
84
4.54
%
1606
86
.81%
*Lym
naea
sta
gnal
is, S
tagn
icol
a pa
lust
ris,
Gal
ba tr
unca
tula
, Rad
ix a
uric
ular
ia, G
yrau
lus
cris
ta, P
lano
rbar
ius
corn
eus
**B
ythi
nella
cf.
turc
a, F
agot
ia e
sper
i
***T
heod
oxus
cf.
held
reic
hi h
eldr
eich
i, V
ivip
arus
viv
ipar
us, V
alva
ta p
isci
nalis
, Pla
norb
is c
arin
atus
, Gyr
aulu
s al
bus

Chapter 5: Micro-freshwater Gastropods
85
Figure 5.3. Frequency of shells from all environment types by level/location.
nals and other moving waters, and rarely in standing water (Gittenberger & Janssen 1998). Thus, we conclude that V. viviparus can be found in both lentic and lotic environments.
Theodoxus cf. heldreichi heldreichi is known from the stony bottoms of both lakes and ponds from Beyşehir, Eğirdir and Hoyran lakes, as well as from the Quaternary sediments of Acıgöl, Ereğli Lake near Çumra, the reed-bed of Hotamış Lake and the channels and ponds of Büyük Gökçeli Spring, Eğirdir-Isparta (Schütt 1991; Schütt & Şeşen 1992; Karaşahin & Yıldırım 1997; Yıldırım 1998).
Table 1 presents the total MNI counts and number of taxa for each level and for the representative buildings. This data is presented according to the two environments that we identi-fied: that of high energy waters and highly oxygenated waters, i.e. lotic environments, and that of low energy or standing wa-ters, i.e. lentic environment. Figure 5.2 shows the total MNI counts of all microshells in our sample, including the dominant species of Viviparus, Valvata and Theodoxus. 86 per cent of the shells we examined can represent either lotic or lentic habitats, and do not show a dominance of one over the other (Fig. 5.3). Yet, when examining only the indicative species (Fig. 5.4), we see that the two environments are more or less equally repre-sented by the microshells. We note that in some of the contexts
examined (e.g. B.20, B.24, B.69, B.85), no shells were present; the reason for this has yet to be determined.
Discussion and conclusions
Çatalhöyük is said to have been surrounded by marshes throughout much of its existence and that these marshes may have had an impact on agricultural practices at the site (Roberts & Rosen 2009). We attempted to contribute to the interpretation of the site’s environment by studying the mol-luscs that were brought to the site as ecofacts, as opposed to artifacts that have been studied separately (Bar-Yosef Mayer & Gümüş 2010; Volume 9, Chapter 19; Chapter 16).
Freshwater snails and bivalves form the largest group of shells at the site. Most of them are minute snails (under 10mm) that are either that size when fully grown, or the ju-veniles of larger specimens such as Viviparus, Lymnaea and Planorbarius. The freshwater gastropod species that were studied here were discovered mainly in the debris of mud-brick and mortar that was used to build the houses of Çatal-höyük. While inspecting some of the bricks and mortar in the field, we identified the molluscs as inclusions and we as-
SOUTH R B 56
SOUTH S B.44
SOUTH T B.10
SOUTH Q B 53
SOUTH Q B.65
SOUTH R B.42
SOUTH R B.56
SOUTH O B 79
SOUTH O B.80
SOUTH P B.75
SOUTH Q B.53
SOUTH ?M B 8
SOUTH N B.89
SOUTH O B.76
SOUTH O B.79Lentic Systems*Lotic Systems**Lentic & Lotic Systems***
SOUTH K B 17
SOUTH L B.6
SOUTH ?M B.24
SOUTH ?M B.8
SOUTH H S 199
SOUTH J B.18
SOUTH J B.23
SOUTH K B.17
0 100 200 300 400 500 600 700
SOUTH G Sp.181
SOUTH H Sp.199

Volume 8: Humans and Landscapes of Çatalhöyük
86
sume that they were inadvertently introduced with the mud from nearby riverbeds. We also assume that bricks and mor-tar were constructed from newly collected materials in each building phase. Micromorphological studies of midden sedi-ments also observed shell fragments within building materi-als (Shillito et al. 2011b; Chapter 3). These shells represent the site’s surroundings; the majority (86 per cent) of micro-shells are species that can tolerate both types of environ-ments; therefore, we cannot use them as indicators for either a lentic or a lotic environment.
It is worth noting that the lakes and ponds that surrounded Çatalhöyük were also the source for aquatic macrophytes, the phytolith remains of which were discovered at the site. In some cases, gastropods live on the macrophytes and it is pos-sible that some of the microshells were brought along with these and not only with mud for bricks.
It is plausible that swamps were visited for the collection of Phragmites sp. (reeds) and Cyperaceae (sedges), and that lentic pulmonates at the site lived on these plants and derived from their collection. The presence of microshells has the po-
Figure 5.4. Frequency of shells indicative of either lotic or lentic environments by level/location.
tential to indicate what types of water bodies the inhabitants of the site chose to collect mud and plants from, and the pro-portion between the numbers of the two groups of gastropods indicates that both environments were equally present (see also Chapter 9).
Acknowledgments
We thank Ian Hodder and Shahina Farid for enabling the study of the Çatalhöyük molluscs. Many of the Çatalhöyük team members consulted and assisted in various aspects of this study. We would like to thank in particular Slobodan Mitrović and Milena Vasić for obtaining the heavy residue samples, and Serena Love, Arlene Rosen, Philippa Ryan, Nurcan Yalman, and Chris Doherty for their valuable advice. We thank Peter Glöer for permission to use his photos of the shells. We are most grateful to Henk K. Mienis (National Col-lections of Natural History, Tel Aviv University) for valuable comments on an earlier version of this paper.
SOUTH R B 56
SOUTH S B.44
SOUTH T B.10
SOUTH Q B 53
SOUTH Q B.65
SOUTH R B.42
SOUTH R B.56
SOUTH O B 79
SOUTH O B.80
SOUTH P B.75
SOUTH Q B.53
SOUTH ?M B 8
SOUTH N B.89
SOUTH O B.76
SOUTH O B.79
Lentic Systems*Lotic Systems**
SOUTH K B 17
SOUTH L B.6
SOUTH ?M B.24
SOUTH ?M B.8
SOUTH H S 199
SOUTH J B.18
SOUTH J B.23
SOUTH K B.17
0 10 20 30 40 50 60 70 80 90 100
SOUTH G Sp.181
SOUTH H Sp.199

87
Excavations at Çatalhöyük have yielded thousands of mol-lusc shell remains. These include shells that were deliberately collected by humans, and thousands of others that were inad-vertently collected along with the mud for the construction of the mudbrick dwellings at the site (Chapter 5; Chapter 16; Love 2010). Shells that were deliberately collected by hu-mans include those collected for the production of ornaments and were brought from the Mediterranean shores and Tau-rus Mountains (Bar-Yosef Mayer et al. 2010). Unio bivalves studied here were also collected from freshwater rivers and lakes in the vicinity of the site, and may have served primar-ily as food, and secondarily as raw material for making or-naments and other artifacts, although it should be noted that some of the collected specimens are juveniles that were inad-vertently collected along with thousands of small gastropods (Chapter 5).
Because Unio is found throughout the sequence of Çatalhöyük, and because the genus lives for several years (Aldridge 1999), we can use the shells to gain information on seasonal climate fluctuations that existed during the Neo-lithic and Chalcolithic periods, and contribute to the debate on the climatic and hydrological variations that may have in-fluenced the economy and society of Neolithic Çatalhöyük. Seasonal activities are expressed at the site in several media, but especially in the faunal and floral remains (Fairbairn et al. 2005a). Subsistence strategies at the site were dictated by arid summers and wet winters (otherwise known as the ‘Mediter-ranean climate’). The site existed during the early stages of agriculture, and the resources exploited by its inhabitants are based on both wild and domesticated fauna and flora (Twiss et al. 2009). Shell-fishing could have been one of the seasonal activities at the site. Other evidence for seasonal activities is expressed in the construction of bins that served as stor-age areas for plant food at the site (Twiss 2008; Bogaard et al. 2009), and activities pertaining to rearing sheep flocks (Russell & Martin 2005). Another enigma concerns the grad-ual abandonment of the East Mound and resettlement on the West Mound; it has been suggested that adverse climatic con-ditions may have caused this change (Marciniak & Czerniak 2007; Biehl & Rosenstock 2009; Roberts & Rosen 2009).
Therefore, a closer view of seasonal climatic conditions dur-ing the site’s occupation is vital for understanding these ac-tivities and their impact on the inhabitants’ economy.
Climatic conditions that prevailed in this area of Turkey during the late Pleistocene and early Holocene were studied by various teams who worked in and around the site. Most conspicuously, Roberts et al. (2001), studying a sedimento-logical core from Lake Eski Acigöl, suggested that there was a rapid rise in precipitation at the beginning of the Holocene followed by a sustained period of moisture (significantly above modern values) until c. 6500 cal BC. Consequently, Roberts & Rosen (2009) suggested that the inhabitants of the site had to grow cereals many kilometres away from the oc-cupation site. While Unio and other freshwater shells found in large numbers at Çatalhöyük confirm that plentiful fresh-water was close to the site (Chapter 5), the interpretation of the appearance of freshwaters near the site (lake, rivers, marshes) and how humans adapted to their surroundings has been contested (Volume 9, Chapter 3). Additional regional climate records of the late Pleistocene and early Holocene around southwestern and south central Turkey were pub-lished over the last decade that include lake isotopic records (Roberts et al. 1999; Leng et al. 1999b; Jones et al. 2002; Eastwood et al. 2007a; Jones et al. 2007; Roberts et al. 2008, Fig. 6.1) with most agreeing that precipitation levels at the beginning of the Holocene were slightly more elevated than today’s records.
Unionidae bivalves can potentially provide a means of assessing seasonal fluctuations and they are particularly important as a potential archive of environmental informa-tion because they store isotopic information in their shells as they grow. These bivalves currently occur in a wide range of freshwater habitats, including rivers, lakes and marshes (e.g., Negus 1966; Aldridge 1999; Tchernov 1975; Heller 2009). Research on unionids in Turkey is very limited (e.g. Modell 1951, Geldiay & Bilgin 1969, Bilgin 1980, Falkner 1994, Öktener 2004, Şereflişan et al. 2009), and the tax-onomy of the region remains poorly resolved (Graf & Cum-mings 2007). The mussels are inactive, or express reduced activity during cold winter periods when their growth may
Chapter 6
Unio Shells from Çatalhöyük: Preliminary Palaeoclimatic Data from Isotopic Analyses
Daniella E. Bar-Yosef Mayer, Melanie J. Leng, David C. Aldridge, Carol Arrowsmith, Burçin A. Gümüş & Hilary J. Sloane

Volume 8: Humans and Landscapes of Çatalhöyük
88
be reduced or cease (Mienis pers. comm.), giving rise to dis-tinct annual growth lines (annuli) within the shell (Goew-ert et al. 2007). In Britain, Unio spp. growth maximum is between April and September (Aldridge pers. observ.; Ne-gus 1966). Glochidia, calcareous larvae of Unio spp., are released on mucus threads in order to attach to host fish on which they must metamorphose in the UK during May and June (Aldridge & McIvor 2003). Unlike most freshwater bivalves, unionids have a relatively long life span (e.g. Ne-gus 1966; Aldridge 1999), with greater investment in shell lengthening in the first few years. The seasonal variations are expressed in the shell and can be measured.
Since the isotopic composition of freshwater bivalve carbonate generally reflects that of water where they reside, shells are potentially useful as tools for investigating changes in lake water related to the local climate (Leng & Marshall 2004; Goewert et al. 2007). Whole-shell isotope data pro-vide an integration of seasonal change over a few years or so, while inter-seasonal information can be gained by sampling at short intervals along the growth axis of the shell (Leng et al. 1999a). Here we present preliminary results of isotopic composition of Unio shells from Çatalhöyük, in an attempt to gain palaeoclimatic data relevant to human activities at and around the site during the critical period of early agriculture. We previously tested the isotope data from a single mod-ern shell of known date and origin which was then used for comparison (Bar-Yosef Mayer et al. 2012). The incremental growth (seasonal) δ13C and δ18O data from the shells were similar to today, and clear seasonal patterns were detected in the shells from the Neolithic and early Chalcolithic periods.
Materials and methods
Subfossil shells were collected from the archaeological site at Çatalhöyük. Unio mancus eucirrus valves were gathered from 18 levels at the site, and while most of the Unio valves from Çatalhöyük were broken at the posterior end, we only collected valves for isotope analysis that were complete from umbo to margin so that isotope samples could be taken from the entire length of the shells representing several years of the mollusc’s growth. Unio valves were measured using a digital calliper. In complete shells (both valves present, n=7) height, length and thickness/convexity were measured. Complete valves (n=64) were measured for height and length. Broken valves were only measured where the entire height was pre-sent (n=260). Minimum Number of Individuals (MNI) was based on counting of umbones of right and left valves, and choosing the larger number of valves (either right or left) in each level.
This study was based on four specimens from representa-tive levels along the sedimentary sequence at the site, which have been dated to early Pottery Neolithic through early Chalcolithic. We selected the best preserved shell specimens (Table 6.1, Fig. 6.2): from the lowermost level of occupa-tion, Level South G at the bottom of the East Mound (5306); from the middle of the sequence in Level South L (1563); from the uppermost levels of the East Mound in Level TP N (17670); and a sample from the Chalcolithic levels in the West Mound (17208) from Trench 5. All shells come from re-liable midden contexts that were not re-deposited. Assuming that these shells were collected as living individuals from the
Figure 6.1. Map of Turkey showing Çatalhöyük and lakes mentioned in the text.

Chapter 6: Unio Shells
89
nearby freshwater sources, they represent four points in time between ca. 9200 cal BC and 7000 cal BC.
The chosen shells show limited signs of surface abrasion or dissolution. They were gently brushed in de-ionized wa-ter to remove adhering sediment. For the incremental growth analysis, between 31 and 66 samples were taken at c. 0.5mm intervals along the shells’ exterior surface from the umbo to the ventral margin, using a 1mm diameter drill. For isotope measurement of the incremental growth, 100μg samples were analyzed using a GV IsoPrime mass spectrometer and multi-prep system together with similarly-sized samples of a labo-ratory calcite standard. Analytical precision (1 SD), based on the laboratory standard, is typically <0.1‰ for both carbon and oxygen isotope ratios. Isotope values (δ13C and δ18O) are reported as per mil (‰) deviations of the isotope ratios (13C/12C, 18O/16O) from VPDB.
Results and discussion
A modern shell that was tested as a control sample validated our procedures and interpretations. The analysis of δ13C and δ18O values of the modern shell (Fig. 6.3) collected from a lake near Adana had δ13C values that range between c. –12 and –5‰ and was consistent with the bivalve utilizing a mixed carbon pool from the water’s inorganic pool, as well as dietary organic carbon (Fritz & Poplawski 1974). The data suggest that the bivalves utilized carbon from both organic (food) and inorganic (debris) sources, but dominantly from the 13C-depleted organic component.
The oxygen isotopic composition of freshwater mus-
sel shells is predominantly dependent on the isotopic com-position of the water in which the carbonate formed and on temperature to a lesser extent (Leng & Marshall 2004). In-ter-seasonal information can be gained by sampling at short intervals across the growth lines of freshwater bivalves (e.g. Koike 2008), although in general, the rivers and open lakes are less sensitive to seasonal variation than ponds and lakes.
Table 6.1. Shells collected from the archaeological site at Çatalhöyük and the modern environment. The sedimentary sequence at Çatalhöyük was sampled in four points along the occupational phases from the lowermost level of occupation, Level South G at the bottom of the East mound, in the mid-dle Level South L, the uppermost levels of the East Mound in the TP Area, as well as one Chalcolithic sample from the West Mound.
Figure 6.2. One of the Unio shells sampled for stable isotopes. Over 30 samples were taken at c. 0.5 mm intervals along the shells outer margin (resulting in a trench like feature) from the umbo to the ventral margin, using a 1mm diameter drill.
Unit Archaeological context Stratigraphic position Date Shell height (mm)
Modern Modern 2008 AD
(17208) Ceramic cluster &
groundstone tools
West Mound (Trench 5) Early Chalcolithic (ECA IV), 6th
millennium BC
(1 date: 5900 cal BC)
25.11
(17670) Midden, faunal remains,
clay balls, 2 broken Unio
‘beads’
Level TP N Late Neolithic (ECA III) 33.46
(1563) Midden, including
phytoliths
Level South L (Sp.115) End Early Pottery Neolithic
(ECA II)
c. 6500 cal BC
33.31
(5306) Midden Level South G (Sp.181) Early Pottery Neolithic (ECA II)
c. 7000 cal BC
31.48
Volume 8: Humans and Landscapes of Çatalhöyük
90
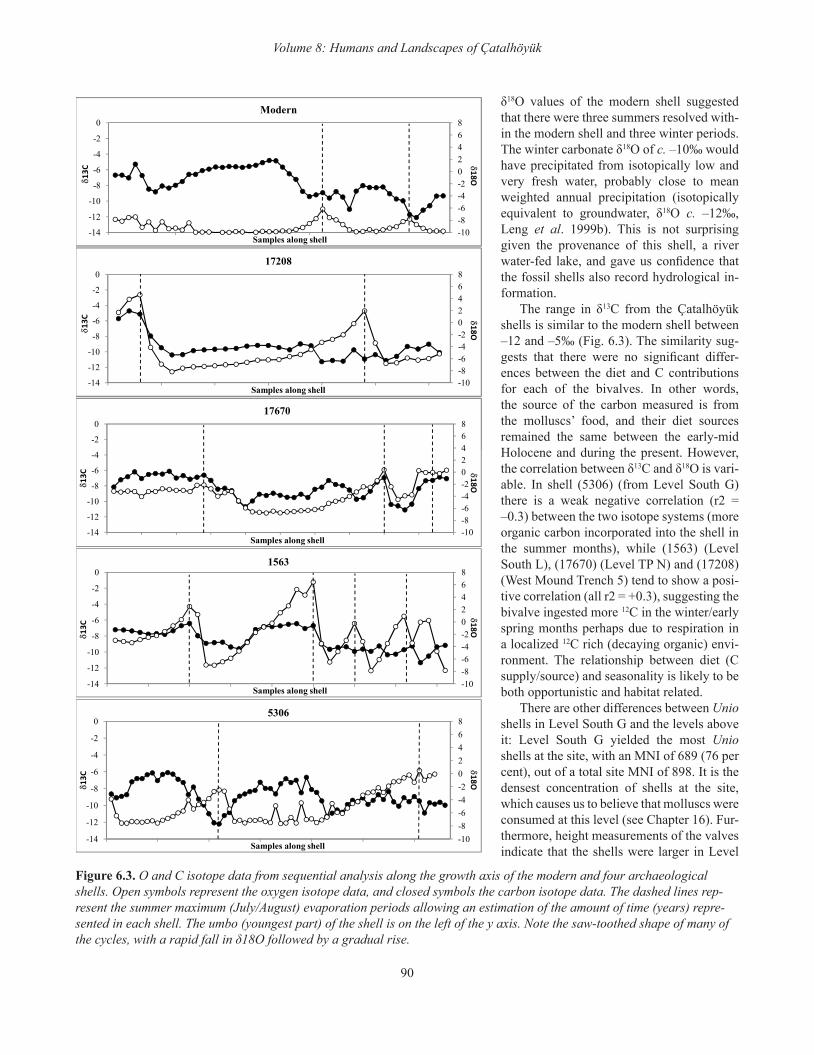
δ18O values of the modern shell suggested that there were three summers resolved with-in the modern shell and three winter periods. The winter carbonate δ18O of c. –10‰ would have precipitated from isotopically low and very fresh water, probably close to mean weighted annual precipitation (isotopically equivalent to groundwater, δ18O c. –12‰, Leng et al. 1999b). This is not surprising given the provenance of this shell, a river water-fed lake, and gave us confidence that the fossil shells also record hydrological in-formation.
The range in δ13C from the Çatalhöyük shells is similar to the modern shell between –12 and –5‰ (Fig. 6.3). The similarity sug-gests that there were no significant differ-ences between the diet and C contributions for each of the bivalves. In other words, the source of the carbon measured is from the molluscs’ food, and their diet sources remained the same between the early-mid Holocene and during the present. However, the correlation between δ13C and δ18O is vari-able. In shell (5306) (from Level South G) there is a weak negative correlation (r2 = –0.3) between the two isotope systems (more organic carbon incorporated into the shell in the summer months), while (1563) (Level South L), (17670) (Level TP N) and (17208) (West Mound Trench 5) tend to show a posi-tive correlation (all r2 = +0.3), suggesting the bivalve ingested more 12C in the winter/early spring months perhaps due to respiration in a localized 12C rich (decaying organic) envi-ronment. The relationship between diet (C supply/source) and seasonality is likely to be both opportunistic and habitat related.
There are other differences between Unio shells in Level South G and the levels above it: Level South G yielded the most Unio shells at the site, with an MNI of 689 (76 per cent), out of a total site MNI of 898. It is the densest concentration of shells at the site, which causes us to believe that molluscs were consumed at this level (see Chapter 16). Fur-thermore, height measurements of the valves indicate that the shells were larger in Level
-10-8-6-4-202468
-14
-12
-10
-8
-6
-4
-2
0
δ18Oδ13C
Samples along shell
Modern
-10-8-6-4-202468
-14
-12
-10
-8
-6
-4
-2
0
δ18O
Samples along shell
δ13C
17208
468
-2
017670
-10-8-6-4-202468
-14
-12
-10
-8
-6
-4
-2
0
δ18Oδ13C
Samples along shell
Modern
-10-8-6-4-202468
-14
-12
-10
-8
-6
-4
-2
0
δ18O
Samples along shell
δ13C
17208
-10-8-6-4-202468
-14
-12
-10
-8
-6
-4
-2
0
δ18Oδ13C
Samples along shell
17670
-10-8-6-4-202468
-14
-12
-10
-8
-6
-4
-2
0
δ18O
δ13C
Samples along shell
1563
-10-8-6-4-202468
-14
-12
-10
-8
-6
-4
-2
0
δ18Oδ13C
Samples along shell
5306
Figure 6.3. O and C isotope data from sequential analysis along the growth axis of the modern and four archaeological shells. Open symbols represent the oxygen isotope data, and closed symbols the carbon isotope data. The dashed lines rep-resent the summer maximum (July/August) evaporation periods allowing an estimation of the amount of time (years) repre-sented in each shell. The umbo (youngest part) of the shell is on the left of the y axis. Note the saw-toothed shape of many of the cycles, with a rapid fall in δ18O followed by a gradual rise.

Chapter 6: Unio Shells
91
South G (average height 32mm, SD 4.09) than in subsequent Neolithic levels (average height 21mm, SD 9.22). It is worth noting that during the Chalcolithic, in the West Mound, the average height increases to 28mm, SD 4.65. These data might suggest that the population of Unio shells around the site was over exploited at the time that Level South G existed, and caused a decrease in their size (a well-known archaeologi-cal phenomenon, e.g. Claassen 1998, 45–49), and that sub-sequently, in the Chalcolithic, the Unio population recovered somewhat and became larger again.
The range in δ18O of the subfossil shells is different from the modern shell, being between –8 and +6‰. However, the lowest value is only slightly higher than the winter δ18O of the modern shell and may indicate that the isotopic composition of the early Holocene winter rainfall (or winter groundwa-ter recharge of the lakes) in the area around Çatalhöyük was similar, but slightly enriched compared with that of today. The high values in the archaeological shells are most likely to be a function of intense summer evaporation, suggesting there was no (or little) summer rainfall, conditions similar to today in this region. When comparing these results to the previously studied cores of Akgöl and Süleymanhaci (Leng et al. 1999b), it seems that the lakes around Çatalhöyük during its occupation had a smaller surface, thus reducing evapora-tion. Alternatively, winter rainfall around the occupation site, or as rain and snowfall in the mountain catchment area, was much higher in the early Holocene allowing for either river flooding and/or a complete winter recharge of the lakes that the bivalves inhabited. Doherty’s reconstruction of a mosaic of many small streams surrounding the site suggests the for-mer (Volume 9, Chapter 3). The clear, saw-toothed shape of the δ18O data displays a rapid fall and a gradual rise in δ18O which may relate to a late season (i.e. late autumn) cessation in growth which then resumes the following spring. These results confirm a winter hiatus in the shells’ growth.
Summary and conclusions
The abundance of Unio shells at Neolithic Çatalhöyük al-lows us to analyze them as a proxy for climate and seasonal fluctuations during the site’s occupation. Incremental growth of bivalve shells provides inter-seasonal environmental data which complements other environmental studies at the site. The shells enable resolution of between three and five sea-sonal cycles each. The carbon isotope variations within all studied shells (modern and archaeological) are similar in most shells, being intermediate between carbonate and or-ganic matter derived C. There is some co-variation in shells (1563), (17208), (17670) from Levels South L, TP N, and West Trench 5, respectively, suggesting a possible link be-
tween molluscan diet and seasonality, i.e. more uptake of organic matter in the winter/early spring months. These iso-tope data complement Roberts & Rosen’s (2009) evidence that water bodies near Çatalhöyük were replenished by river flooding in the springtime. However, while their interpreta-tion suggests water bodies were shallow or standing, and then became progressively smaller and shallower due to seasonal evaporation during the dry summer months, it is also possi-ble that what they see is evidence for winter ponds. Because Unio shells are dependent on constant oxygenation of the wa-ter, even during the winter months when they are inactive, we cannot say that those were collected from shallow or standing water bodies.
The archaeological shells show a distinct seasonality (summer to winter) expressed in all four shells. The winter values suggest high recharge by winter precipitation at the lake or delivered by river floods of the lake(s); this enables lake waters to return to a δ18O composition that is typical of regional groundwater, while the summer values are indicative of intense aridification during the summer months (similar to today). The return of lakes to typical groundwater isotopic composition in winters suggests very high water table levels.
The data presented here are limited; however, in the one level where we feel confident that people were gathering shellfish as a food source, Level South G, the data also sug-gest that the shell was collected in the autumn. Although we are aware that further sampling is necessary, this is a signifi-cant contribution to the study of seasonal activities at Çatal-höyük (Fairbairn et al. 2005a). This preliminary investigation of Unio from Çatalhöyük demonstrates the great potential of palaeoclimatic information from these shells. Bivalve isotope analysis is one of the few methods for investigating seasonal fluctuations in the past, and can provide information on the effects of seasonality on human activity due to climate.
Acknowledgments
This contribution is an abridged version of a paper by the same authors published in Journal of Archaeological Sci-ence (doi: 10.1016/j.jas.2011.09.003). We wish to thank Ian Hodder and Shahina Farid for enabling this study, Henk K. Mienis for providing the modern sample and for consultation on Unionidae, Arek Marciniak for providing the sample from the TP area, and Peter Biehl for providing the sample from the West Mound. We are grateful to Neil Roberts and three anonymous reviewers for providing most valuable comments on the JAS paper. DB-Y thanks the Israeli Ministry of Sci-ence, Culture and Sport for supporting the national collec-tions of natural history at Tel Aviv University as a biodiver-sity, environment, and agriculture research knowledge centre.




















