
Microalgae, Macroalgae, and Bivalves as Biofilters in Land-Based Mariculture in Israel

Marine Pollution Bulletin 61 (2010) 554–567
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Metal and metalloid bioconcentration capacity of two tropical bivalvesfor monitoring the impact of land-based mining activities in theNew Caledonia lagoon
Laetitia Hédouin a,b,c, Miguel Gomez Batista d, Marc Metian a, Emmanuel Buschiazzo a, Michel Warnau a,c,*
a International Atomic Energy Agency-Marine Environment Laboratories (IAEA-MEL), 4 Quai Antoine Ier, MC-98000, Monacob Institut de Recherche pour le Développement (IRD-Noumea Center), BP A5, 98848 Noumea Cedex, New Caledoniac Littoral, Environnement et Sociétés (LIENSs), UMR 6250 CNRS-Université de La Rochelle, 2 rue Olympe de Gouges, F-17042 La Rochelle Cedex, Franced Centro de Estudios Ambientales de Cienfuegos (CEAC), Ciudad Nuclear, 59350 Cienfuegos, Cuba
a r t i c l e i n f o
Keywords:MetalConcentration effectBioindicatorRadiotracer techniques
0025-326X/$ - see front matter � 2010 Elsevier Ltd.doi:10.1016/j.marpolbul.2010.06.036
* Corresponding author. Present address: InternatTechnical Cooperation Department, Division for AfrBox 100, A-1400 Vienna, Austria.
E-mail address: [email protected] (M. W
a b s t r a c t
The clam Gafrarium tumidum and the oyster Isognomon isognomon have been proposed as potential bio-monitor species of metal contamination in the New Caledonia lagoon. The influence of dissolved concen-trations of As, Cd, Co, Cr, Mn, and Zn on uptake and depuration kinetics, tissue and subcellulardistribution of these elements was investigated in both species. Results indicate that both bivalves takeup elements proportionally to the dissolved concentration for Cd, Cr and Mn in the surrounding waterover the entire range of concentrations tested (three orders of magnitude), and up to the second and thirdhighest added metal concentration tested for Co (23 ng l�1) and Zn (700 ng l�1), respectively. All elementswere efficiently retained in bivalve tissues (estimated Tb1/2 ranging from 16 d to1), suggesting that bothspecies should be able to preserve a record of contamination events over a long period of time. Consid-ering the specific range of concentrations examined here, G. tumidum and I. isognomon would thereforeserve as adequate biomonitor species to monitor dissolved metal contamination in the New Caledonialagoon waters.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
The coral barrier of New Caledonia encloses the second largestlagoon of the world after the Australian Great Barrier Reef(Labrosse et al., 2000). Beside a rich biodiversity and a high ende-mism (approximately 5% of the 15,000 species described so far;Richer de Forges et al., 2000; Bouchet et al., 2002), the New Caledo-nia lagoon is also subjected to a high degree of metal contamina-tion pressure. Indeed, about 25% of the world nickel stocks arepresent in New Caledonia (Dalvi et al., 2004) and ores have beenlocally exploited since the end of the nineteenth century, mainlyto extract Ni. Nowadays, mining activities constitute the mostimportant economical resource of the Territory. Recent develop-ment of more efficient extraction processes based on acid extrac-tion (viz. lixiviation) has now made possible extraction from oreswith lower Ni content than those currently exploited (Mihaylovet al., 2000; Goro-Nickel, 2001; Dalvi et al., 2004). Although thisnew hydrometallurgical process will lead to a more efficient
All rights reserved.
ional Atomic Energy Agency,ica, Wagramer Strasse 5, PO
arnau).
extraction of Ni from laterite ore (>90%; Mihaylov et al., 2000),the acidic solubilization of metals is obviously not restricted toNi which is exploited, but also concerns all other ore-containedby-product metals which are mostly not exploited. Therefore, thereis a non-negligible risk that application of the lixiviation processwill eventually result in increasing discharges of dissolved by-product metals into the environment (Goro-Nickel, 2001; Baroudiet al., 2003). In particular, Co, Cr and Mn (i.e., the most abundantby-products; Mihaylov et al., 2000) in New Caledonia mines shouldbe considered with special caution.
The increase of metal contamination in coastal waters is a pri-mary threatening consequence of mining activities, and an increas-ing attention has been given to the assessment of the metalcontamination status in marine organisms and to the possible im-pacts on the local marine ecosystems of the New Caledonia lagoon(e.g., Dalto et al., 2006; Hédouin et al., 2008a; Metian et al., 2008b;Chouvelon et al., 2009).
Among the common approaches used to study environmentalcontamination, the use of biomonitor species has proven to bevaluable and informative (e.g., Phillips, 1980; Rainbow, 1995; War-nau et al., 1998; Coteur et al., 2003; Danis et al., 2004). This ap-proach has received much scientific interest, mainly in thetemperate zone, with for example the successful implementation

L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567 555
of several large scale monitoring programmes such as the US andEuropean ‘‘Mussel Watches” (e.g., Goldberg et al., 1978; Warnauand Acuña, 2007; Thébault et al., 2008). Yet, only few studies havebeen dedicated to biomonitoring development and implementa-tion in tropical zones. Although studies have proposed bivalvessuch as Saccostrea spp., Crassostrea spp. or Perna spp. as potentialtropical bioindicators (e.g., Phillips, 1985; Rainbow and Phillips,1993), none of these species are present in sufficient abundancein the New Caledonia lagoon to be used as bioindicators. In thiscontext, the infaunal clam Gafrarium tumidum and the oyster Isogn-omon isognomon have been proposed as a potential target species(e.g., Hédouin et al., 2006, 2007, 2008a). Indeed, these species arewidely distributed in the New Caledonia lagoon and would allowproviding ecotoxicological information on the two main types ofenvironments that are found in the lagoon. G. tumidum is an edibleclam that lives buried in muddy to sandy coastal areas, whereas I.isognomon lives on rocks and dead coral skeletons in sandy androcky areas (e.g., Hédouin et al., 2006, 2007, 2008a). Both speciesrespond to most criteria that should be met by a biomonitor spe-cies (Hédouin et al., 2006, 2007, 2008a). Among these pre-requi-sites (see e.g., Moore, 1966; Phillips, 1980, 1990), the speciesselected should display a simple relationship between the concen-trations of a contaminant accumulated in its tissues and the con-centrations of this contaminant in the environment. Althoughthis recommendation is of major importance, the effect of realisticambient metal concentration on the bioconcentration capacities oforganisms is far from being assessed systematically in biomonitorcandidates (e.g., Borchardt, 1983; Talbot, 1985; Bjerregaard, 1988;Warnau et al., 1997; Metian and Warnau, 2008). In the New Cale-donian lagoon this is of particular concern, since very wide rangesof metal concentrations have been reported in coastal waters (upto two orders of magnitude; Fernandez et al., 2002).
Therefore, the purpose of this study was to investigate the effectof dissolved element concentration on element bioconcentrationby the infaunal clam G. tumidum and the oyster I. isognomon ex-posed to realistic concentration ranges of As, Cd, Co, Cr, Mn andZn in seawater. Although metal bioaccumulation in marine bi-valves is known to occur simultaneously from several pathways(seawater, suspended particles and food), the present study fo-cuses on the direct uptake from seawater. Indeed, recent reportsindicate that the metals in the effluent from the new lixiviationprocess that will be implemented soon in New Caledonia aremainly present as dissolved forms (Goro-Nickel, 2003; Stauberet al., 2003). In addition, as La Havannah Canal (i.e., the locationwhere the effluent is to be released) carries relatively low particleloads and has a strong hydrodynamism, with net currents orientedtowards the area of Noumea City (Goro-Nickel, 2003; Fernandez,J.M., unpublished results), there is a non-negligible risk that animportant fraction of the lixiviated metals will remain at leastpartly under dissolved forms over a long distance. Particular atten-tion was given to the determination of the influence of realistic ele-ment concentrations on bioconcentration, tissue distribution andsubcellular distribution of the considered elements.
Table 1Concentrations of the 6 elements investigated and activity of their corresponding c-emitting radiotracers added in seawater for preparing the increasing exposureconcentrations (C0–C4).
Radiotracer (kBq l�1) Added stable metal concentrations (ng l�1)
C0 C1 C2 C3 C4
Cr 1.5 0 10 50 250 1250Mn 0.5 0 10 50 250 1250Co 0.5 0 8 23 248 998Zn 0.5 0 70 250 700 1750As 0.5 0 2000 5000 10,000 –Cd 1.0 0 2 10 50 250
2. Materials and methods
2.1. Organism collection and acclimation
The clams G. tumidum (n = 300; whole-body wet wt from 18 to28 g) were collected by hand-picking in October 2003 in an inter-tidal station (Dumbéa Bay), whereas the oysters I. isognomon(n = 300; whole-body wet wt from 22 to 42 g) were collected bySCUBA diving during the same week in a subtidal station (MaaBay). Both stations are located within 20 km north of Noumea City,New Caledonia. Body size is well known to affect bioaccumulation
of metals in marine organisms (e.g., Boyden, 1974). Therefore,according to recommendations from preliminary studies (see Me-tian, 2003; Hédouin et al., 2006, 2008a), only organisms with ashell width greater than 35 mm (G. tumidum) and shell lengthlonger than 70 mm (I. isognomon) were selected.
After collection, oysters and clams were shipped to the IAEA-MEL premises in Monaco where they were acclimated to laboratoryconditions (open circuit aquarium; water renewal rate: 30% h�1;salinity: 36 psu; temperature T� = 25 ± 0.5 �C; pH 8.0 ± 0.1; light/dark cycle: 12 h/12 h) simulating the prevailing conditions in theNew Caledonia lagoon. During the 2 months of acclimation, bi-valves were fed phytoplankton (Isochrysis galbana; 104 cells ml�1)and recorded mortality was lower than 5%.
2.2. Stable elements, radiotracers and radioanalysis
The elements investigated (As, Cd, Co, Cr, Mn and Zn) wereintroduced into the experimental microcosms both as stable ele-ments and radiotracers (73As, 109Cd, 57Co, 51Cr, 54Mn and 65Zn) inorder to detect element bioaccumulation with high sensitivity(Warnau and Bustamante, 2007). Stable elements were introducedas HNO3 salts (synthesis quality) purchased from Merck, France.Radiotracers of high specific activity were purchased fromAmersham, UK (51Cr as Na2CrO4, T1/2 = 27.7 d; 57Co as CoCl2, T1/2 =271.8 d), Isotope Product Lab., USA (109Cd as CdCl2, T1/2 = 426.6 d;65Zn as ZnCl2; T1/2 = 243.9 d; 54Mn as MnCl2, T1/2 = 312.2 d),Hyenryk Niewodniczanski Institute of Nuclear Physics, Poland(73As as HAsO3, T1/2 = 80.3 d). Radioactivity was measured usinga high-resolution c-spectrometer system composed of fourGermanium – N or P type – detectors (EGNC 33-195-R, Canberra�
and Eurysis�) connected to a multichannel analyser and a com-puter equipped with a spectra analysis software (Interwinner�
6). The radioactivity of samples was determined by comparisonwith standards of known activities and of appropriate geometry.Measurements were corrected for counting efficiency, backgroundand radioactive decay. Counting time was adapted to obtain count-ing rates with propagated errors less than 5% (Rodriguez y Baenaet al., 2006a).
2.3. Experimental procedure
For each element tested, 5 groups of 9 clams (shell width from35 to 44 mm), and 5 groups of 9 oysters (shell length from 74 to112 mm) were placed in 5 aquaria containing 50 l of 0.45-lmfiltered natural Mediterranean seawater (constantly aerated; salin-ity: 36 psu; T� = 25 ± 0.5 �C; pH 8.0 ± 0.1). Bivalves were subse-quently exposed for 14 d to five increasing added concentrationsof a given element, up to 250 ng Cd l�1 (2.2 nM), 998 ng Co l�1
(17 nM) 1250 ng Cr l�1 (24 nM), 1250 ng Mn l�1 (23 nM),1750 ng Zn l�1 (27 nM) and 10,000 ng As l�1 (134 nM) (Table 1).These concentrations were made using increasing amounts of thestable element and a fixed activity of the corresponding
556 L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567
radiotracer. The concentrations were selected in order to cover thewhole ranges of dissolved concentrations representative of thoseexisting in the New Caledonia lagoon (Fernandez et al., 2002; Fer-nandez, J.M., unpublished results). Spikes of radiotracer were: 73As(0.5 kBq l�1), 109Cd (1 kBq l�1), 57Co (0.5 kBq l�1), 51Cr (1.5 kBq l�1),54Mn (0.5 kBq l�1) and 65Zn (0.5 kBq l�1). In terms of stable ele-ment additions, these activities correspond to the following con-centrations: 2.30 � 10�39 ng As l�1, 4.84 � 10�20 ng Cd l�1, 2.75 �10�20 ng Co l�1, 1.39 � 10�19 ng Cr l�1, 3.55 � 10�19 ng Mn l�1 and5.99 � 10�18 ng Zn l�1.
No change in pH was detectable after radiotracer and stable ele-ment additions. Seawater and spikes were renewed daily for 5 d,then every second day in order to keep exposure activities con-stant. Concentrations of the elements in seawater were checkeddaily, before and after each seawater renewal, using the corre-sponding radiotracer activity. This allowed calculation of the actualstable element concentrations based on the ratio between nominalconcentration of the element and the nominal radioactivity of thecorresponding tracer. The maximum decrease in seawater radioac-tivity between two successive seawater renewals (n = 28) was11 ± 10% for 73As, 13 ± 11% for 109Cd, 25 ± 11% for 57Co, 10 ± 6%for 51Cr, 23 ± 11% for 54Mn and 21 ± 12% for 65Zn.
During the experiments, oysters and clams were allowed to feedbriefly (30 min) on phytoplankton Isochrysis galbana (104 cells
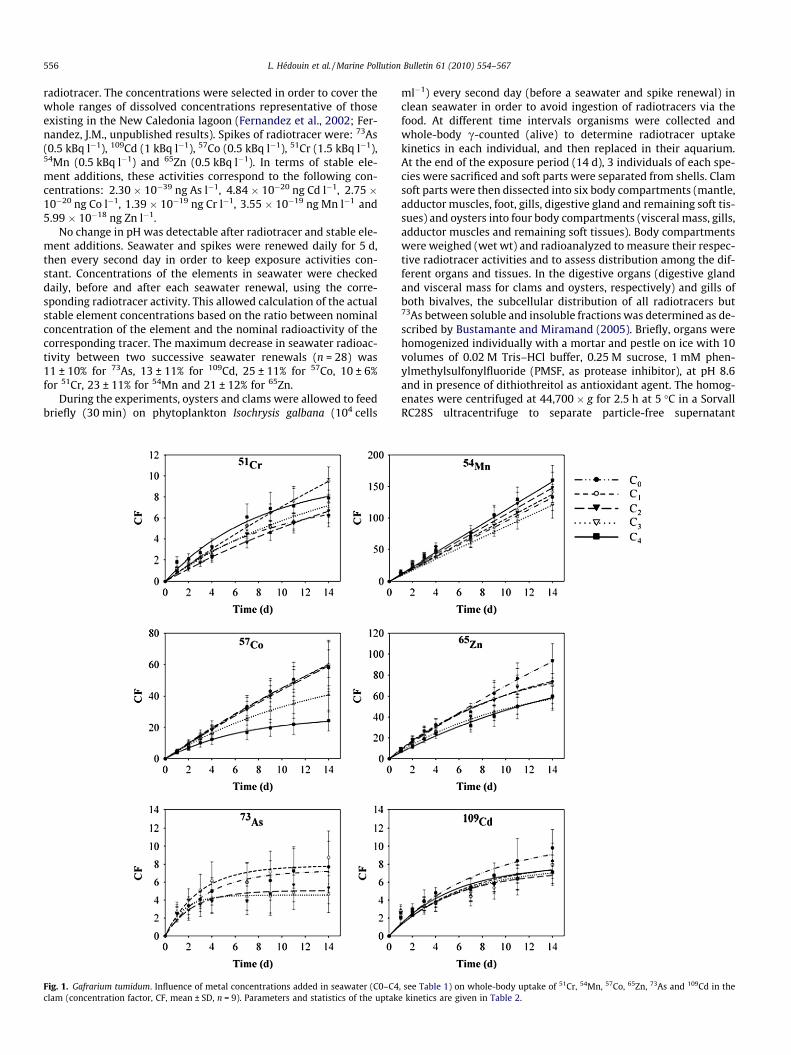
Fig. 1. Gafrarium tumidum. Influence of metal concentrations added in seawater (C0–C4clam (concentration factor, CF, mean ± SD, n = 9). Parameters and statistics of the uptak
ml�1) every second day (before a seawater and spike renewal) inclean seawater in order to avoid ingestion of radiotracers via thefood. At different time intervals organisms were collected andwhole-body c-counted (alive) to determine radiotracer uptakekinetics in each individual, and then replaced in their aquarium.At the end of the exposure period (14 d), 3 individuals of each spe-cies were sacrificed and soft parts were separated from shells. Clamsoft parts were then dissected into six body compartments (mantle,adductor muscles, foot, gills, digestive gland and remaining soft tis-sues) and oysters into four body compartments (visceral mass, gills,adductor muscles and remaining soft tissues). Body compartmentswere weighed (wet wt) and radioanalyzed to measure their respec-tive radiotracer activities and to assess distribution among the dif-ferent organs and tissues. In the digestive organs (digestive glandand visceral mass for clams and oysters, respectively) and gills ofboth bivalves, the subcellular distribution of all radiotracers but73As between soluble and insoluble fractions was determined as de-scribed by Bustamante and Miramand (2005). Briefly, organs werehomogenized individually with a mortar and pestle on ice with 10volumes of 0.02 M Tris–HCl buffer, 0.25 M sucrose, 1 mM phen-ylmethylsulfonylfluoride (PMSF, as protease inhibitor), at pH 8.6and in presence of dithiothreitol as antioxidant agent. The homog-enates were centrifuged at 44,700 � g for 2.5 h at 5 �C in a SorvallRC28S ultracentrifuge to separate particle-free supernatant
, see Table 1) on whole-body uptake of 51Cr, 54Mn, 57Co, 65Zn, 73As and 109Cd in thee kinetics are given in Table 2.
L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567 557
(cytosol; soluble fraction) from the pellet (insoluble fraction).Homogenate aliquots, cytosols and pellets were then radioanalyzed.
The remaining bivalves were then placed in a non-contami-nated medium (50-l aquaria with flowing, 0.45-lm filtered, naturalseawater, flux: 50% h�1, salinity: 36 psu; T� = 25 ± 0.5 �C; light/darkcycle: 12 h/12 h) for 21 d to follow the depuration kinetics of radio-tracers from the organisms. As during the exposure phase, oystersand clams were fed (30 min) phytoplankton Isochrysis galbana(104 cells ml�1) every second day. At the end of the depuration per-iod, all individuals were dissected to determine the distribution ofthe elements among the body compartments as described above.
2.4. Data treatment
Uptake of the five investigated radiotracers was expressed interms of concentration factor (CF, ratio between activity of theradiotracer in the whole organism or in a body compartment –Bq g�1 wet wt – and time-integrated activity of radiotracer in sea-water – Bq g�1 –) (Warnau et al., 1996; Rodriguez y Baena et al.,2006b). Radiotracer uptake kinetics were described using a simplelinear regression model (Eq. (1)), or by a saturation exponential ki-netic model (Eq. (2)) if the observed kinetics tended to reach a stea-dy-state equilibrium:
CFt ¼ kut ð1ÞCFt ¼ CFssð1� e�ketÞ ð2Þ
Table 2Gafrarium tumidum. Estimated parameters of the whole-body uptake kinetics and depuratiadded in seawater (C0–C4; see Table 1) and then maintained for 21 d in clean conditions. Ustate; ku: uptake rate constant (d�1); ASE: asymptotic standard error; R2: determination coabsorption efficiency (%); ke, depuration rate constant (d�1); Tb1/2, biological half-life (d) (windicated).
Tracer Conc. A – Uptake period
Model ku ± ASE CFss ± ASE R2
51Cr C0 E 0.86 ± 0.05d 8.3 ± 0.6d 0.94C1 E 0.79 ± 0.05d 34 ± 14a 0.95C2 E 0.63 ± 0.04d 13 ± 3d 0.93C3 E 0.78 ± 0.05d 12 ± 2d 0.93C4 E 1.2 ± 0.1d 10 ± 1d 0.85
54Mn C0 L 9.8 ± 0.2d 0.93C1 L 10.1 ± 0.2d 0.94C2 L 11.1 ± 0.2d 0.94C3 L 9.3 ± 0.2d 0.90C4 L 11.5 ± 0.2d 0.94
57Co C0 E 6.3 ± 0.4d 121 ± 23d 0.90C1 E 6.1 ± 0.6d 129 ± 40b 0.86C2 E 5.7 ± 0.5d 141 ± 44b 0.91C3 E 5.6 ± 0.6d 55 ± 7d 0.86C4 E 5.3 ± 0.2d 26 ± 2d 0.75
65Zn C0 E 8.0 ± 0.6d 294 ± 129a 0.92C1 E 9.6 ± 0.6d 97 ± 9d 0.92C2 E 9.3 ± 1.0d 106 ± 18d 0.82C3 E 7.4 ± 0.5d 77 ± 7d 0.91C4 E 6.1 ± 0.1d 105 ± 24d 0.87
73As C0 E 2.2 ± 0.3d 7.2 ± 0.5d 0.66C1 E 2.7 ± 0.4d 7.8 ± 0.4d 0.65C2 E 2.2 ± 0.4d 5.0 ± 0.3d 0.53C3 E 3.0 ± 0.6d 4.6 ± 0.2d 0.48
109Cd C0 E 1.4 ± 0.2d 11 ± 1d 0.80C1 E 1.3 ± 0.1d 8.3 ± 0.6d 0.87C2 E 1.3 ± 0.1d 7.4 ± 0.5d 0.85C3 E 1.3 ± 0.2d 7.8 ± 1.0d 0.68C4 E 1.5 ± 0.2d 7.8 ± 0.6d 0.73
a p < 0.05.b p < 0.01.c p < 0.001.d p < 0.0001.
* Not significant (p > 0.05).
where CFt and CFss are the concentration factors at time t (d) and atsteady state, respectively; ku and ke are the uptake and depurationrate constants (d�1), respectively (Whicker and Schultz, 1982;Warnau et al., 1996). Linearity of the uptake kinetics was testedby a linearity test for regression with replication (Zar, 1996). Modelconstants (CFss, ku and ke) were estimated by iterative adjustment ofthe model using the nonlinear curve-fitting routines in the Statisti-ca� software 5.2.1.
Depuration of radiotracers was expressed as the percentage ofremaining radioactivity (radioactivity at time t divided by the ini-tial radioactivity measured in the organism at the beginning of thedepuration period). The depuration kinetics of the radiotracerswere best fitted using either a single-component (Eq. (3)) or a dou-ble-component (Eq. (4)) exponential model:
At ¼ A0 e�ket ð3ÞAt ¼ A0s e�kest þ A0l e�kelt ð4Þ
where At and A0 are the remaining activities (%) at time t (d) and 0,respectively; ke is the depuration rate constant (d�1); ‘s’ and ‘l’ arethe subscripts for the ‘short-lived’ and ‘long-lived’ components,respectively. The short-lived component represents the loss kineticsof the radiotracer fraction that is weakly associated to the organ-isms and rapidly eliminated, whereas the long-lived component de-scribes the loss kinetics of the radiotracer fraction that is tightlybound to the organism (Warnau et al., 1996). For each exponentialcomponent (s and l), a biological half-life can be calculated (Tb1/2s
on kinetics of 6 radiotracers in clams exposed for 14 d to 5 increasing concentrationsptake period: linear (L) or exponential (E) model; CFss, concentration factors at steadyefficient. Depuration period: single- (S) or two- (T) component exponential model; A0,
hen depuration follows a T Model, only parameters of the long-lived component are
B – Depuration period
Model A0 ± ASE ke ± ASE Tb1/2 ± ASE R2
S 97 ± 1d 0.017 ± 0.001d 42 ± 4 0.72S 97 ± 1d 0.018 ± 0.001d 39 ± 3 0.83S 96 ± 2d 0.014 ± 0.002d 50 ± 6 0.59S 97 ± 1d 0.017 ± 0.001d 40 ± 3 0.76S 95 ± 1d 0.012 ± 0.001d 56 ± 6 0.64
S 101 ± 1d 0.022 ± 0.001d 32 ± 1 0.94S 100 ± 1d 0.020 ± 0.001d 35 ± 1 0.96S 101 ± 1d 0.019 ± 0.005d 37 ± 1 0.96S 101 ± 1d 0.020 ± 0.001d 34 ± 1 0.91S 101 ± 1d 0.019 ± 0.001d 36 ± 1 0.91
S 101 ± 1d 0.027 ± 0.001d 25 ± 1 0.96S 102 ± 1d 0.021 ± 0.001d 34 ± 1 0.95S 101 ± 1d 0.023 ± 0.001d 30 ± 1 0.93S 100 ± 1d 0.026 ± 0.001d 27 ± 1 0.97S 98 ± 1d 0.021 ± 0.001d 32 ± 2 0.83
T 86 ± 2d 0.015 ± 0.002d 48 ± 6 0.91T 87 ± 6d 0.014 ± 0.004b 48 ± 15 0.77T 87 ± 3d 0.015 ± 0.002d 45 ± 7 0.83T 82 ± 5d 0.015 ± 0.004c 45 ± 13 0.79T 78 ± 5d 0.012 ± 0.004b 56 ± 19 0.85
T 51 ± 7d 0.037 ± 0.009c 19 ± 4.7 0.95T 53 ± 7d 0.043 ± 0.009d 16 ± 3.4 0.92T 36 ± 13b 0.013 ± 0.022* 54 ± 92 0.93T 54 ± 10d 0.036 ± 0.012b 19 ± 7 0.85
T 79 ± 4d 0.004 ± 0.003* 177 ± 152 0.71T 70 ± 4d 0.002 ± 0.004* 347 ± 693 0.60T 77 ± 8d 0.001 ± 0.007* 1226 ± 14,381 0.45T 69 ± 64* 0.003 ± 0.052* 231 ± 4005 0.70T 77 ± 5d 0.005 ± 0.004* 139 ± 122 0.62
558 L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567
and Tb1/2l) from the corresponding depuration rate constant (kes andkel, respectively) according to the relation Tb1/2 = ln 2/ke.
Possible differences among uptake and depuration kinetics ofeach radiotracer over the concentration range considered weretested using the Linear Mixed Models (LMM) procedure imple-mented in SPSS v12.0 software. This procedure expands the gen-eral linear model so that the error terms and random effects arepermitted to exhibit correlated and non-constant variability. TheLMM, therefore, provides the flexibility to model not only the meanof a response variable, but its covariance structure as well. Timewas specified as the repeated effect and the total dissolved concen-trations (stable + radioactive) of As, Cd, Co, Cr, Mn and Zn as thefixed effect.
Kinetic parameters (ku, ke, CFss, A0s, A0l, kes, kel) estimated for agiven element were plotted against the concentration of the ele-ment (stable + radioactive) dissolved in seawater and were fittedusing simple linear regression.
The level of significance for statistical analyses was always setat a = 0.05.
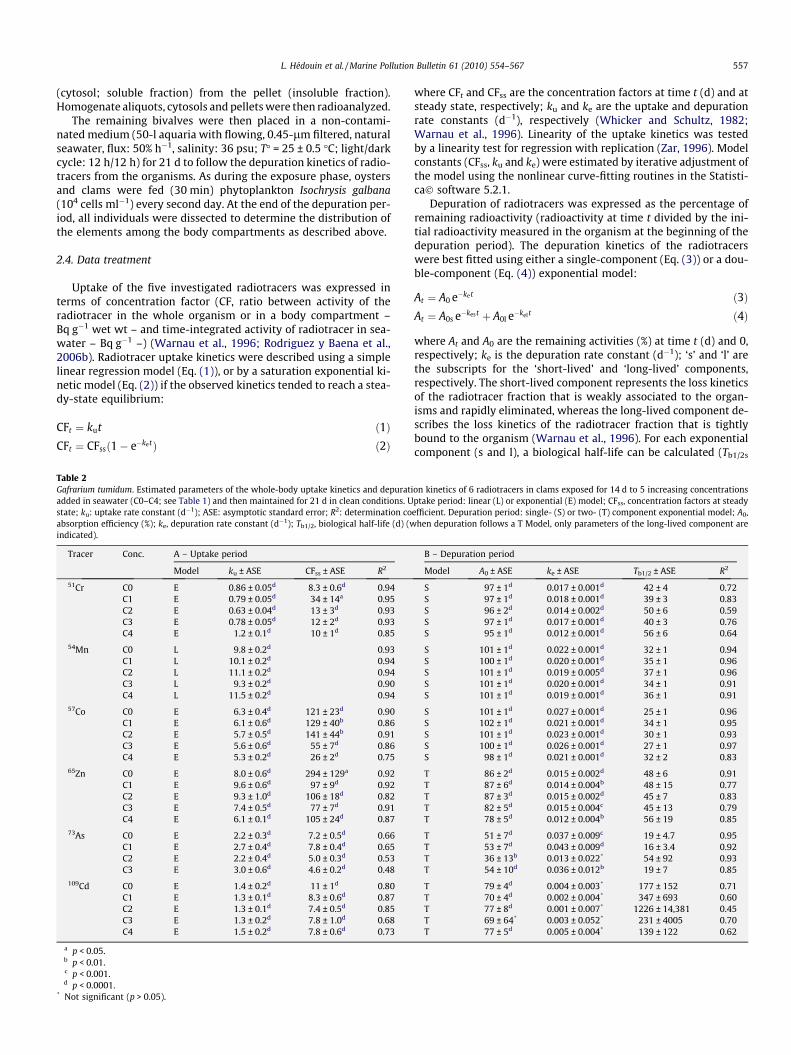
Fig. 2. Gafrarium tumidum. Body distribution (%, mean ± SD, n = 3) of 51Cr, 54Mn, 57Coconcentrations added in seawater (C0–C4, see Table 1).
3. Results
3.1. Clam G. tumidum
During the 14-d exposure period of the clams, whole-body up-take kinetics of all radiotracers but 54Mn displayed saturationkinetics for all concentrations tested (R2: 0.82–0.95 for 51Cr and65Zn; 0.68–0.91 for 109Cd and 57Co; and 0.48–0.66 for 73As)(Fig. 1, Table 2). In the case of 54Mn, the tracer was accumulatedaccording to linear uptake kinetics (R2: 0.90–0.94) (Fig. 1, Table2). Comparisons among the rates at which elements were biocon-centrated in the whole organisms (ku) indicated that the six radio-tracers ranked according to the following decreasing order:54Mn > 65Zn > 57Co > 73As > 109Cd > 51Cr. The estimated steady-state concentration factors (CFss) indicated that the radiotracerswere concentrated to different extent by G. tumidum, accordingto the following order: 57Co � 65Zn > 51Cr, 109Cd > 73As (Table 2).
Linear Mixed Model (LMM) analyses indicated that uptakekinetics of 109Cd, 51Cr and 54Mn did not significantly differ
, 65Zn, 73As and 109Cd in the clams after a 14-d exposure to 5 increasing metal
L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567 559
(pSPSS > 0.05) over the range of concentrations tested. In contrast,the exposure concentration in seawater did affect significantlythe uptake kinetics of 73As, 57Co and 65Zn in clams (pSPSS < 0.0001,except 73As for which pSPSS = 0.03). For Co concentrations higherthan 23 ng l�1 (C2) the depuration rate constant (ke) increased sig-nificantly (p < 0.001), whereas CFss decreased (p < 0.05). Effects onuptake kinetics of As were observed for dissolved As concentra-tions exceeding 5000 ng l�1 (C2), with a significant increase(p < 0.005) of the depuration rate constant (ke). For Zn, a decreasein ke value by one order of magnitude was observed between thecontrol concentration and the other concentrations tested.
The distributions of radiotracers among the clam body compart-ments and between the soluble and insoluble fraction of digestivegland and gill cells were determined at the end of the exposureperiod for the five conditions tested. The body distribution wasslightly affected by exposure concentration for all elements, exceptCo for which substantial changes were observed (Fig. 2). Indeed,when the dissolved Co concentration was higher than 23 ng l�1
(C2), the amount of Co in the digestive gland decreased by 50+%and concomitantly increased in the mantle and gills.
Except for 51Co in the digestive gland, no major difference wasobserved in the subcellular distribution of the radiotracers indigestive and gill cells of clams exposed to the five different ele-ment concentrations. In both digestive gland and gill cells, 109Cdwas mainly associated to the cytosolic fraction (ranging from 93%
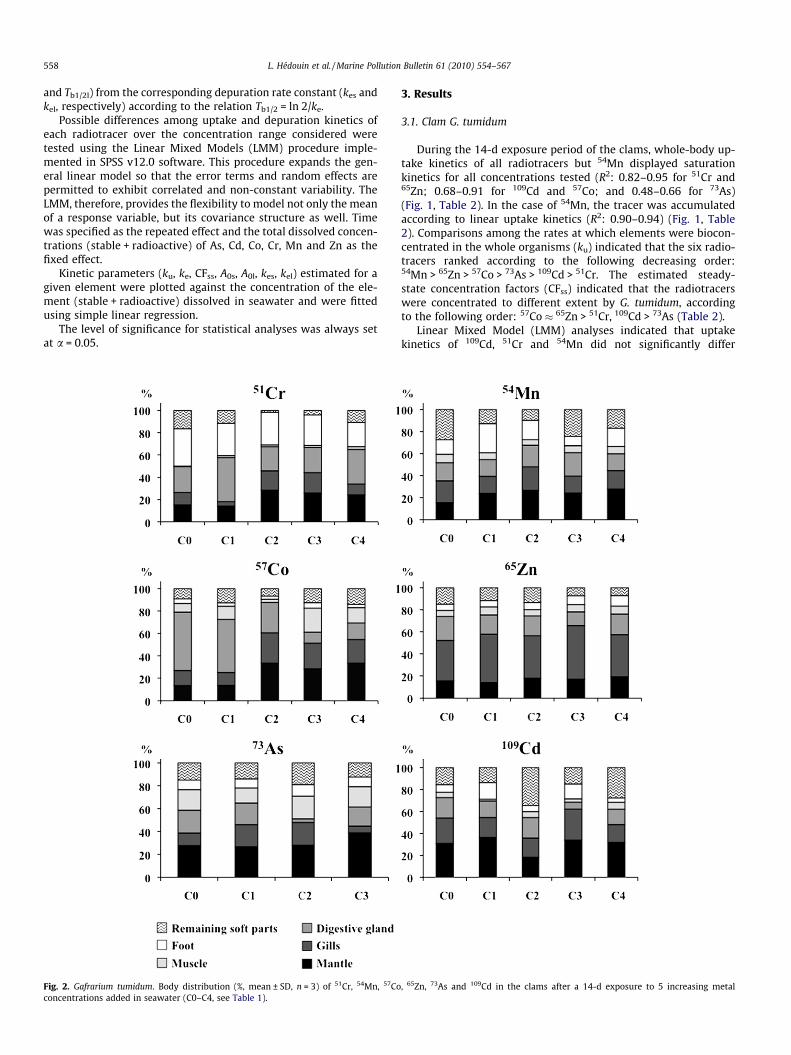
Fig. 3. Gafrarium tumidum. Influence of metal concentrations added in seawater (C0–C4clams (% remaining activities, A(%), mean ± SD, n = 6). Parameters and statistics of the d
to 95% and 73% to 81%, respectively), whereas 65Zn was mostlybound to insoluble compounds (56–74% in the digestive glandand 73–85% in the gills). The major part of 54Mn was associatedwith the insoluble fraction in both digestive gland (50–66%) andgills (64–72%). The major part of 51Cr was associated with theinsoluble fraction in the gills (from 60% to 68%), whereas in thedigestive gland, 51Cr was mainly (54–71%) distributed in the cyto-sol. In the gill cells, 57Co was distributed rather equally betweenthe soluble and insoluble fractions (% insoluble fraction rangingfrom 42% to 63%), whereas it was mainly associated with the solu-ble fraction in the digestive gland. However, it the latter organ,57Co proportion in the soluble fraction decreased when exposureCo concentration increased (from 77% at 0 ng added Co l�1 to 49%at 998 ng added Co l�1).
After the 14-d exposure period, clean flowing seawater condi-tions were restored and depuration kinetics of the radiotracerswere followed for 21 d in the bivalves. Depuration kinetics of57Co, 51Cr and 54Mn were best fitted by a single-component expo-nential model for all concentrations tested (Fig. 3, Table 2) andthose of 73As, 109Cd and 65Zn were best described by a two-compo-nent exponential model. No significant difference (pSPSS > 0.05) wasobserved among the different depuration kinetics determined overthe concentration range examined. The long-lived depuration com-ponent of 109Cd and 65Zn represented a large proportion of theradioactivity (accounting for 69–79% for Cd and 78–87% for Zn)
, see Table 1) on whole-body depuration of 51Cr, 54Mn, 57Co, 65Zn, 73As and 109Cd inepuration kinetics are given in Table 2.
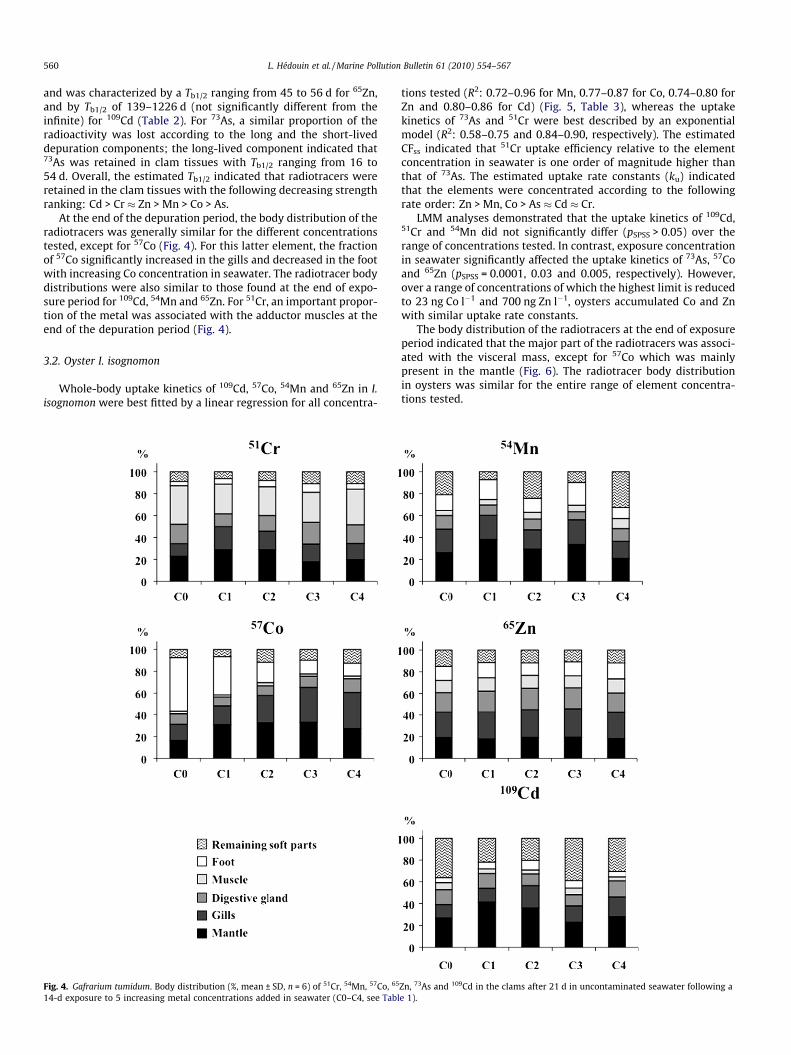
560 L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567
and was characterized by a Tb1/2 ranging from 45 to 56 d for 65Zn,and by Tb1/2 of 139–1226 d (not significantly different from theinfinite) for 109Cd (Table 2). For 73As, a similar proportion of theradioactivity was lost according to the long and the short-liveddepuration components; the long-lived component indicated that73As was retained in clam tissues with Tb1/2 ranging from 16 to54 d. Overall, the estimated Tb1/2 indicated that radiotracers wereretained in the clam tissues with the following decreasing strengthranking: Cd > Cr � Zn > Mn > Co > As.
At the end of the depuration period, the body distribution of theradiotracers was generally similar for the different concentrationstested, except for 57Co (Fig. 4). For this latter element, the fractionof 57Co significantly increased in the gills and decreased in the footwith increasing Co concentration in seawater. The radiotracer bodydistributions were also similar to those found at the end of expo-sure period for 109Cd, 54Mn and 65Zn. For 51Cr, an important propor-tion of the metal was associated with the adductor muscles at theend of the depuration period (Fig. 4).
3.2. Oyster I. isognomon
Whole-body uptake kinetics of 109Cd, 57Co, 54Mn and 65Zn in I.isognomon were best fitted by a linear regression for all concentra-
Fig. 4. Gafrarium tumidum. Body distribution (%, mean ± SD, n = 6) of 51Cr, 54Mn, 57Co, 65
14-d exposure to 5 increasing metal concentrations added in seawater (C0–C4, see Tabl
tions tested (R2: 0.72–0.96 for Mn, 0.77–0.87 for Co, 0.74–0.80 forZn and 0.80–0.86 for Cd) (Fig. 5, Table 3), whereas the uptakekinetics of 73As and 51Cr were best described by an exponentialmodel (R2: 0.58–0.75 and 0.84–0.90, respectively). The estimatedCFss indicated that 51Cr uptake efficiency relative to the elementconcentration in seawater is one order of magnitude higher thanthat of 73As. The estimated uptake rate constants (ku) indicatedthat the elements were concentrated according to the followingrate order: Zn > Mn, Co > As � Cd � Cr.
LMM analyses demonstrated that the uptake kinetics of 109Cd,51Cr and 54Mn did not significantly differ (pSPSS > 0.05) over therange of concentrations tested. In contrast, exposure concentrationin seawater significantly affected the uptake kinetics of 73As, 57Coand 65Zn (pSPSS = 0.0001, 0.03 and 0.005, respectively). However,over a range of concentrations of which the highest limit is reducedto 23 ng Co l�1 and 700 ng Zn l�1, oysters accumulated Co and Znwith similar uptake rate constants.
The body distribution of the radiotracers at the end of exposureperiod indicated that the major part of the radiotracers was associ-ated with the visceral mass, except for 57Co which was mainlypresent in the mantle (Fig. 6). The radiotracer body distributionin oysters was similar for the entire range of element concentra-tions tested.
Zn, 73As and 109Cd in the clams after 21 d in uncontaminated seawater following ae 1).
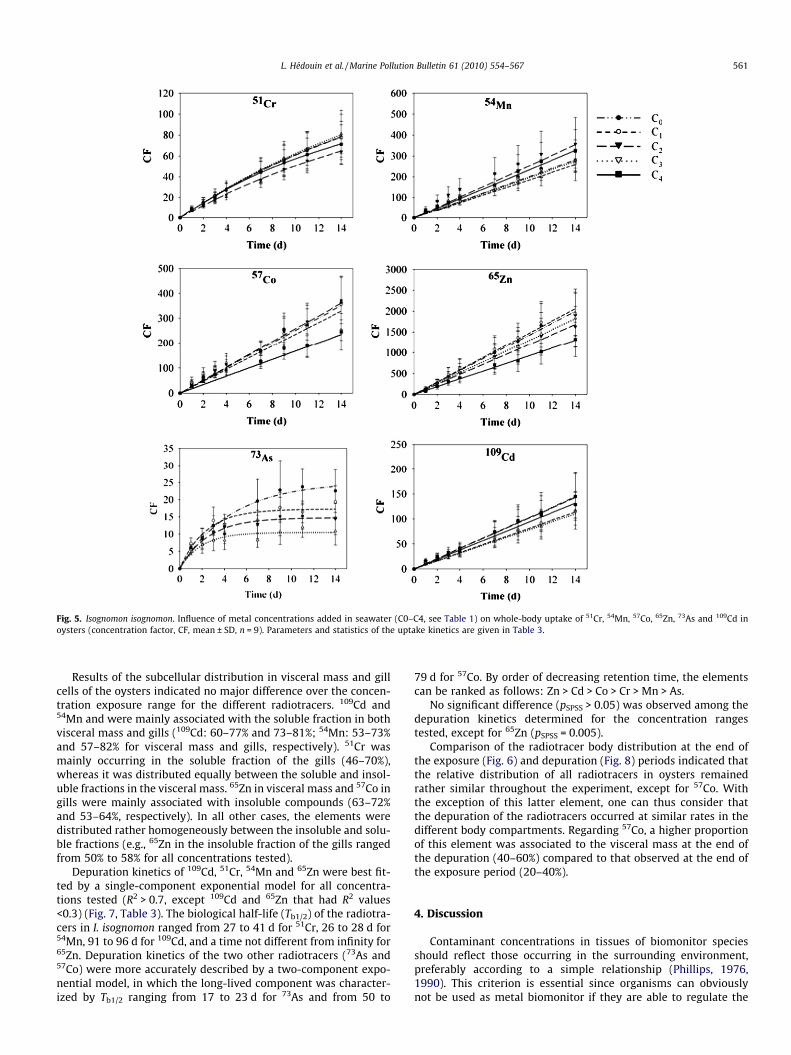
Fig. 5. Isognomon isognomon. Influence of metal concentrations added in seawater (C0–C4, see Table 1) on whole-body uptake of 51Cr, 54Mn, 57Co, 65Zn, 73As and 109Cd inoysters (concentration factor, CF, mean ± SD, n = 9). Parameters and statistics of the uptake kinetics are given in Table 3.
L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567 561
Results of the subcellular distribution in visceral mass and gillcells of the oysters indicated no major difference over the concen-tration exposure range for the different radiotracers. 109Cd and54Mn and were mainly associated with the soluble fraction in bothvisceral mass and gills (109Cd: 60–77% and 73–81%; 54Mn: 53–73%and 57–82% for visceral mass and gills, respectively). 51Cr wasmainly occurring in the soluble fraction of the gills (46–70%),whereas it was distributed equally between the soluble and insol-uble fractions in the visceral mass. 65Zn in visceral mass and 57Co ingills were mainly associated with insoluble compounds (63–72%and 53–64%, respectively). In all other cases, the elements weredistributed rather homogeneously between the insoluble and solu-ble fractions (e.g., 65Zn in the insoluble fraction of the gills rangedfrom 50% to 58% for all concentrations tested).
Depuration kinetics of 109Cd, 51Cr, 54Mn and 65Zn were best fit-ted by a single-component exponential model for all concentra-tions tested (R2 > 0.7, except 109Cd and 65Zn that had R2 values<0.3) (Fig. 7, Table 3). The biological half-life (Tb1/2) of the radiotra-cers in I. isognomon ranged from 27 to 41 d for 51Cr, 26 to 28 d for54Mn, 91 to 96 d for 109Cd, and a time not different from infinity for65Zn. Depuration kinetics of the two other radiotracers (73As and57Co) were more accurately described by a two-component expo-nential model, in which the long-lived component was character-ized by Tb1/2 ranging from 17 to 23 d for 73As and from 50 to
79 d for 57Co. By order of decreasing retention time, the elementscan be ranked as follows: Zn > Cd > Co > Cr > Mn > As.
No significant difference (pSPSS > 0.05) was observed among thedepuration kinetics determined for the concentration rangestested, except for 65Zn (pSPSS = 0.005).
Comparison of the radiotracer body distribution at the end ofthe exposure (Fig. 6) and depuration (Fig. 8) periods indicated thatthe relative distribution of all radiotracers in oysters remainedrather similar throughout the experiment, except for 57Co. Withthe exception of this latter element, one can thus consider thatthe depuration of the radiotracers occurred at similar rates in thedifferent body compartments. Regarding 57Co, a higher proportionof this element was associated to the visceral mass at the end ofthe depuration (40–60%) compared to that observed at the end ofthe exposure period (20–40%).
4. Discussion
Contaminant concentrations in tissues of biomonitor speciesshould reflect those occurring in the surrounding environment,preferably according to a simple relationship (Phillips, 1976,1990). This criterion is essential since organisms can obviouslynot be used as metal biomonitor if they are able to regulate the
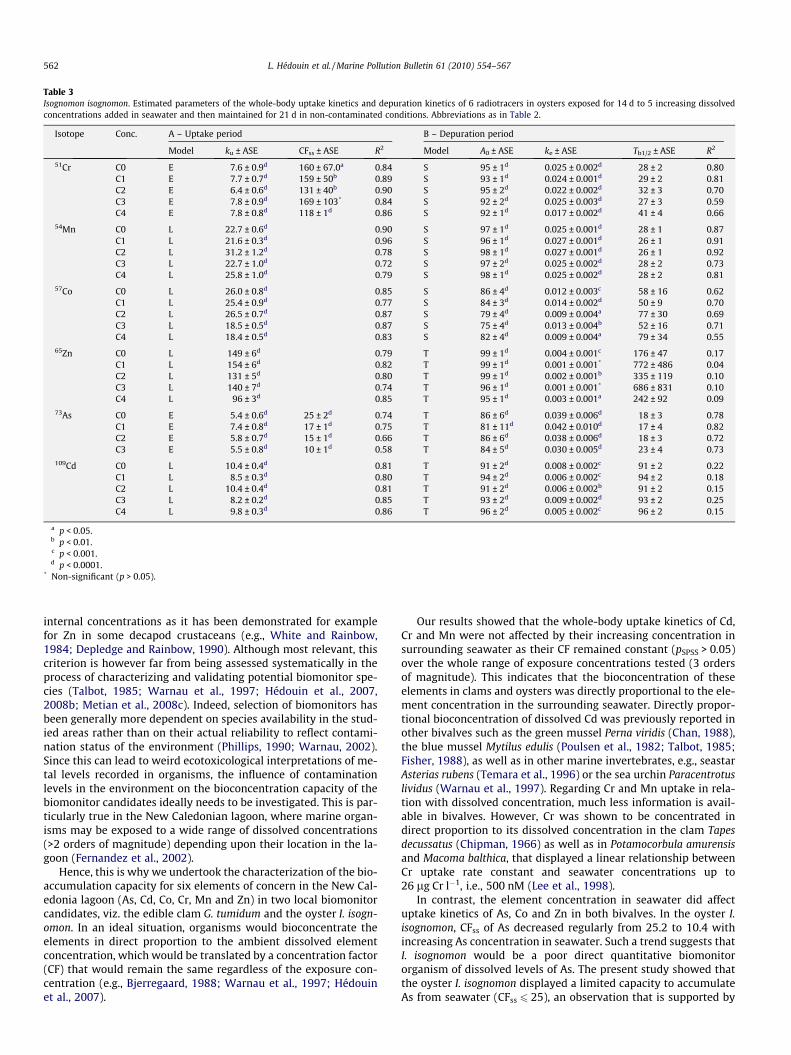
Table 3Isognomon isognomon. Estimated parameters of the whole-body uptake kinetics and depuration kinetics of 6 radiotracers in oysters exposed for 14 d to 5 increasing dissolvedconcentrations added in seawater and then maintained for 21 d in non-contaminated conditions. Abbreviations as in Table 2.
Isotope Conc. A – Uptake period B – Depuration period
Model ku ± ASE CFss ± ASE R2 Model A0 ± ASE ke ± ASE Tb1/2 ± ASE R2
51Cr C0 E 7.6 ± 0.9d 160 ± 67.0a 0.84 S 95 ± 1d 0.025 ± 0.002d 28 ± 2 0.80C1 E 7.7 ± 0.7d 159 ± 50b 0.89 S 93 ± 1d 0.024 ± 0.001d 29 ± 2 0.81C2 E 6.4 ± 0.6d 131 ± 40b 0.90 S 95 ± 2d 0.022 ± 0.002d 32 ± 3 0.70C3 E 7.8 ± 0.9d 169 ± 103* 0.84 S 92 ± 2d 0.025 ± 0.003d 27 ± 3 0.59C4 E 7.8 ± 0.8d 118 ± 1d 0.86 S 92 ± 1d 0.017 ± 0.002d 41 ± 4 0.66
54Mn C0 L 22.7 ± 0.6d 0.90 S 97 ± 1d 0.025 ± 0.001d 28 ± 1 0.87C1 L 21.6 ± 0.3d 0.96 S 96 ± 1d 0.027 ± 0.001d 26 ± 1 0.91C2 L 31.2 ± 1.2d 0.78 S 98 ± 1d 0.027 ± 0.001d 26 ± 1 0.92C3 L 22.7 ± 1.0d 0.72 S 97 ± 2d 0.025 ± 0.002d 28 ± 2 0.73C4 L 25.8 ± 1.0d 0.79 S 98 ± 1d 0.025 ± 0.002d 28 ± 2 0.81
57Co C0 L 26.0 ± 0.8d 0.85 S 86 ± 4d 0.012 ± 0.003c 58 ± 16 0.62C1 L 25.4 ± 0.9d 0.77 S 84 ± 3d 0.014 ± 0.002d 50 ± 9 0.70C2 L 26.5 ± 0.7d 0.87 S 79 ± 4d 0.009 ± 0.004a 77 ± 30 0.69C3 L 18.5 ± 0.5d 0.87 S 75 ± 4d 0.013 ± 0.004b 52 ± 16 0.71C4 L 18.4 ± 0.5d 0.83 S 82 ± 4d 0.009 ± 0.004a 79 ± 34 0.55
65Zn C0 L 149 ± 6d 0.79 T 99 ± 1d 0.004 ± 0.001c 176 ± 47 0.17C1 L 154 ± 6d 0.82 T 99 ± 1d 0.001 ± 0.001* 772 ± 486 0.04C2 L 131 ± 5d 0.80 T 99 ± 1d 0.002 ± 0.001b 335 ± 119 0.10C3 L 140 ± 7d 0.74 T 96 ± 1d 0.001 ± 0.001* 686 ± 831 0.10C4 L 96 ± 3d 0.85 T 95 ± 1d 0.003 ± 0.001a 242 ± 92 0.09
73As C0 E 5.4 ± 0.6d 25 ± 2d 0.74 T 86 ± 6d 0.039 ± 0.006d 18 ± 3 0.78C1 E 7.4 ± 0.8d 17 ± 1d 0.75 T 81 ± 11d 0.042 ± 0.010d 17 ± 4 0.82C2 E 5.8 ± 0.7d 15 ± 1d 0.66 T 86 ± 6d 0.038 ± 0.006d 18 ± 3 0.72C3 E 5.5 ± 0.8d 10 ± 1d 0.58 T 84 ± 5d 0.030 ± 0.005d 23 ± 4 0.73
109Cd C0 L 10.4 ± 0.4d 0.81 T 91 ± 2d 0.008 ± 0.002c 91 ± 2 0.22C1 L 8.5 ± 0.3d 0.80 T 94 ± 2d 0.006 ± 0.002c 94 ± 2 0.18C2 L 10.4 ± 0.4d 0.81 T 91 ± 2d 0.006 ± 0.002b 91 ± 2 0.15C3 L 8.2 ± 0.2d 0.85 T 93 ± 2d 0.009 ± 0.002d 93 ± 2 0.25C4 L 9.8 ± 0.3d 0.86 T 96 ± 2d 0.005 ± 0.002c 96 ± 2 0.15
a p < 0.05.b p < 0.01.c p < 0.001.d p < 0.0001.
* Non-significant (p > 0.05).
562 L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567
internal concentrations as it has been demonstrated for examplefor Zn in some decapod crustaceans (e.g., White and Rainbow,1984; Depledge and Rainbow, 1990). Although most relevant, thiscriterion is however far from being assessed systematically in theprocess of characterizing and validating potential biomonitor spe-cies (Talbot, 1985; Warnau et al., 1997; Hédouin et al., 2007,2008b; Metian et al., 2008c). Indeed, selection of biomonitors hasbeen generally more dependent on species availability in the stud-ied areas rather than on their actual reliability to reflect contami-nation status of the environment (Phillips, 1990; Warnau, 2002).Since this can lead to weird ecotoxicological interpretations of me-tal levels recorded in organisms, the influence of contaminationlevels in the environment on the bioconcentration capacity of thebiomonitor candidates ideally needs to be investigated. This is par-ticularly true in the New Caledonian lagoon, where marine organ-isms may be exposed to a wide range of dissolved concentrations(>2 orders of magnitude) depending upon their location in the la-goon (Fernandez et al., 2002).
Hence, this is why we undertook the characterization of the bio-accumulation capacity for six elements of concern in the New Cal-edonia lagoon (As, Cd, Co, Cr, Mn and Zn) in two local biomonitorcandidates, viz. the edible clam G. tumidum and the oyster I. isogn-omon. In an ideal situation, organisms would bioconcentrate theelements in direct proportion to the ambient dissolved elementconcentration, which would be translated by a concentration factor(CF) that would remain the same regardless of the exposure con-centration (e.g., Bjerregaard, 1988; Warnau et al., 1997; Hédouinet al., 2007).
Our results showed that the whole-body uptake kinetics of Cd,Cr and Mn were not affected by their increasing concentration insurrounding seawater as their CF remained constant (pSPSS > 0.05)over the whole range of exposure concentrations tested (3 ordersof magnitude). This indicates that the bioconcentration of theseelements in clams and oysters was directly proportional to the ele-ment concentration in the surrounding seawater. Directly propor-tional bioconcentration of dissolved Cd was previously reported inother bivalves such as the green mussel Perna viridis (Chan, 1988),the blue mussel Mytilus edulis (Poulsen et al., 1982; Talbot, 1985;Fisher, 1988), as well as in other marine invertebrates, e.g., seastarAsterias rubens (Temara et al., 1996) or the sea urchin Paracentrotuslividus (Warnau et al., 1997). Regarding Cr and Mn uptake in rela-tion with dissolved concentration, much less information is avail-able in bivalves. However, Cr was shown to be concentrated indirect proportion to its dissolved concentration in the clam Tapesdecussatus (Chipman, 1966) as well as in Potamocorbula amurensisand Macoma balthica, that displayed a linear relationship betweenCr uptake rate constant and seawater concentrations up to26 lg Cr l�1, i.e., 500 nM (Lee et al., 1998).
In contrast, the element concentration in seawater did affectuptake kinetics of As, Co and Zn in both bivalves. In the oyster I.isognomon, CFss of As decreased regularly from 25.2 to 10.4 withincreasing As concentration in seawater. Such a trend suggests thatI. isognomon would be a poor direct quantitative biomonitororganism of dissolved levels of As. The present study showed thatthe oyster I. isognomon displayed a limited capacity to accumulateAs from seawater (CFss 6 25), an observation that is supported by

Fig. 6. Isognomon isognomon. Body distribution (%, mean ± SD, n = 3) of 51Cr, 54Mn, 57Co, 65Zn, 73As and 109Cd in the oysters after a 14-d exposure to 5 increasing metalconcentrations added in seawater (C0–C4, see Table 1).
L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567 563
a previous study on the Mediterranean mussel Mytilusgalloprovincialis (Ünlü and Fowler, 1979). However, very high Asconcentrations have been measured in the field in G. tumidum(441 lg As g�1 dry wt) and I. isognomon (77 lg As g�1 dry wt)(Hédouin et al., 2008a). This indicates that As is actually taken upby these bivalves, but that As is most probably predominantly bio-accumulated through a pathway other than the dissolved phase.This would support the hypothesis of Sanders et al. (1989) whoproposed the dietary pathway as the main source of As in marineorganisms.
Concentration factors of Co in both bivalves remained similarup to the second highest concentration (23 ng Co l�1) added in sea-water, and then decreased above that threshold exposure concen-tration. Results indicate that this decreasing trend in both specieswas likely due to a decrease in the uptake rate in oysters and toan increase in depuration rate in clams, when exposed to the high-est dissolved Co concentrations tested. This in turn suggests theoccurrence of protective regulatory mechanisms that reduce theuptake (in oysters) or increase the depuration (in clams) efficiencyof this element under high contamination pressure. The latterhypothesis regarding the clam can also be put in relation withthe changes observed for Co body distribution. Indeed, at the endof the exposure period to the two highest Co concentrations tested,the proportion of Co associated with the digestive gland was 50–
80% less than for the lower exposure concentrations, maybe dueto partial saturation of binding sites. As the digestive gland isknown to be the main organ involved in metal sequestration inmolluscs (e.g., Simkiss and Mason, 1984; Soto et al., 1996; Busta-mante and Miramand, 2005), a preferential distribution of Co inother body compartments could result in a higher fraction of Cothat can be excreted more easily, which would explain the increasein Co depuration efficiency that was observed in clams exposed tothe highest concentrations.
Bioconcentration of Zn in the oysters displayed the same trendas Co although the threshold dissolved concentration was muchhigher (third highest concentration tested); i.e., a net decrease inku was observed only for an added Zn concentration higher than700 ng Zn l�1 (11 nM) instead of 23 ng l�1 (0.4 nM) as observedfor Co. As a comparison, Zn bioconcentration in the gills of themussel M. edulis has been shown to display a linear relationshipwith ambient dissolved Zn up to 1.05 mg Zn l�1 (i.e., 1600 nM)(Vercauteren and Blust, 1996).
The occurrence of these threshold dissolved concentrationsindicate that G. tumidum and I. isognomon could be used as abiomonitor species of Co and Zn contamination if care is taken tolimit their use over a restricted range of dissolved concentrations(viz. our background concentrations + 23 ng added Co l�1 and700 ng added Zn l�1). Dissolved concentrations up to 20 ng Co l�1

Fig. 7. Isognomon isognomon. Influence of metal concentrations added in seawater (C0–C4, see Table 1) on whole-body depuration of 51Cr, 54Mn, 57Co, 65Zn, 73As and 109Cd inoysters (% remaining activities, A(%), mean ± SD, n = 6). Parameters and statistics of the depuration kinetics are given in Table 3.
564 L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567
and 1500 ng Zn l�1 have been reported in highly contaminatedcoastal zones of the New Caledonia lagoon (Fernandez et al.,2002; Goro-Nickel, 2004). This means that the experimental con-centration range up to 23 ng added Co l�1 covers the one that canbe encountered in the field in New Caledonia. With respect to Zn,the background dissolved concentration in the natural seawaterused in our experiments ranged between 330 and 590 ng Zn l�1
(Ferrier-Pagès et al., 2005). This indicates that bivalves took upZn proportionally to the ambient dissolved concentration up to1030–1290 ng Zn l�1 (background Zn concentration in experimen-tal seawater + added Zn concentration), which nearly matches theZn concentration range encountered in the New Caledonianlagoon.
At the end of the exposure period, the organisms were placedin non-contaminating conditions and the depuration kineticparameters were determined for both species. Under low contam-ination pressure (restoration of uncontaminated conditions), thedepuration rate of all the elements in both bivalve species wasgenerally not significantly affected by the previous exposure con-centration in seawater (pSPSS > 0.05), except for Zn in the oyster.Similar results have been reported for Cd in the sea urchin P. livi-dus (Warnau et al., 1997) and the green mussel P. viridis (Black-more and Wang, 2002). For example, the depuration rateconstant of Cd (0.007–0.012 d�1) in the green mussel was not sig-
nificantly different after a 7-d exposure to 0 and 100 lg l�1 ofadded Cd (Blackmore and Wang, 2002). Although unrealistic, thelatter very high exposure concentration indicated that this non-dependence on Cd concentration should hold true for the entirerange of Cd concentrations that can be encountered in the marineenvironment. Our study further showed that all elements testedwere efficiently retained in both bivalve species (Tb1/2 > 20 d), sug-gesting that these molluscs are able to preserve informationregarding their contamination history for a relatively long periodof time (several weeks to months). This is particularly true inthe case of Cd and Zn, for which the highest retention capacityhas been observed.
The particularly strong retention of Zn in I. isognomon (estimatedTb1/2 not different from 1) and its high uptake rate (ku = 150 d�1)emphasize the capacity of the oyster to store large amounts of Znin its tissues. Concordant field data has been previously reportedfor the genus Isognomon (e.g., 4010 lg Zn g�1 in I. alatus fromDominican Republic, Sbriz et al., 1998; 12,160 lg Zn g�1 from Gua-deloupe, Ifremer/RNO-Antilles, unpublished work) as well as forseveral other oyster species (e.g., Ruddell and Rains, 1975; Engeland Brouwer, 1982). Subcellular distribution results have shownthat Zn was mainly associated with the organelle and membranefraction, suggesting that a large proportion of Zn in I. isognomoncould be sequestered in insoluble granules, as was previously
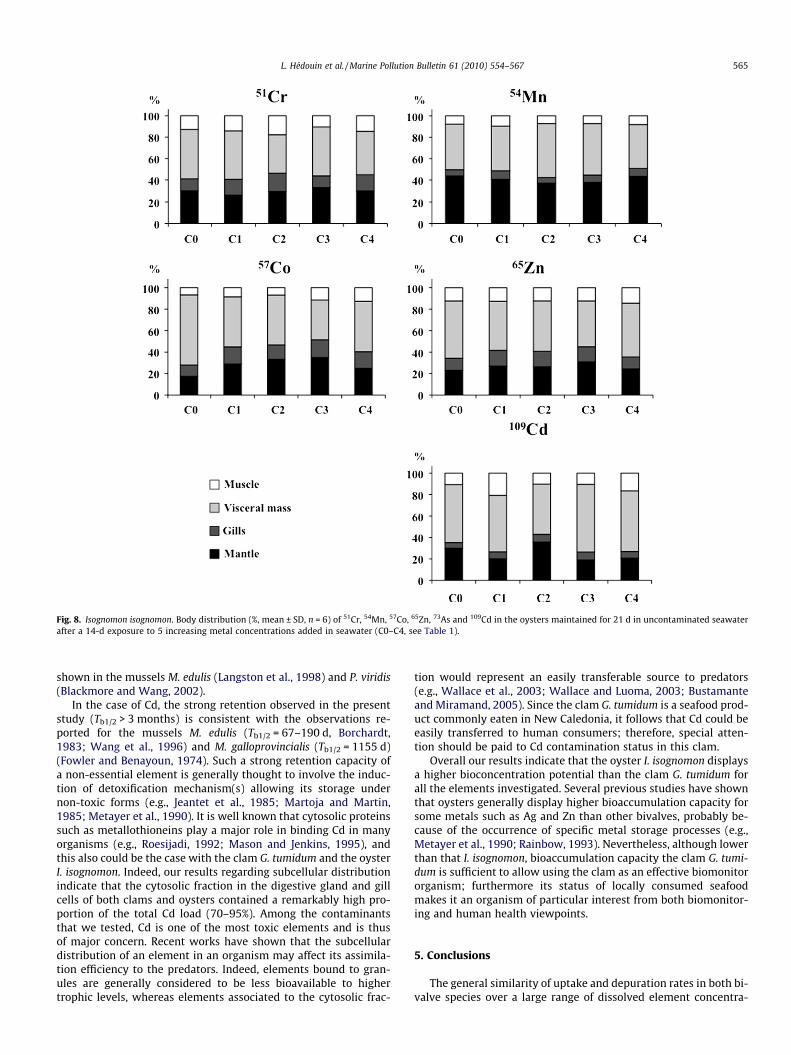
Fig. 8. Isognomon isognomon. Body distribution (%, mean ± SD, n = 6) of 51Cr, 54Mn, 57Co, 65Zn, 73As and 109Cd in the oysters maintained for 21 d in uncontaminated seawaterafter a 14-d exposure to 5 increasing metal concentrations added in seawater (C0–C4, see Table 1).
L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567 565
shown in the mussels M. edulis (Langston et al., 1998) and P. viridis(Blackmore and Wang, 2002).
In the case of Cd, the strong retention observed in the presentstudy (Tb1/2 > 3 months) is consistent with the observations re-ported for the mussels M. edulis (Tb1/2 = 67–190 d, Borchardt,1983; Wang et al., 1996) and M. galloprovincialis (Tb1/2 = 1155 d)(Fowler and Benayoun, 1974). Such a strong retention capacity ofa non-essential element is generally thought to involve the induc-tion of detoxification mechanism(s) allowing its storage undernon-toxic forms (e.g., Jeantet et al., 1985; Martoja and Martin,1985; Metayer et al., 1990). It is well known that cytosolic proteinssuch as metallothioneins play a major role in binding Cd in manyorganisms (e.g., Roesijadi, 1992; Mason and Jenkins, 1995), andthis also could be the case with the clam G. tumidum and the oysterI. isognomon. Indeed, our results regarding subcellular distributionindicate that the cytosolic fraction in the digestive gland and gillcells of both clams and oysters contained a remarkably high pro-portion of the total Cd load (70–95%). Among the contaminantsthat we tested, Cd is one of the most toxic elements and is thusof major concern. Recent works have shown that the subcellulardistribution of an element in an organism may affect its assimila-tion efficiency to the predators. Indeed, elements bound to gran-ules are generally considered to be less bioavailable to highertrophic levels, whereas elements associated to the cytosolic frac-
tion would represent an easily transferable source to predators(e.g., Wallace et al., 2003; Wallace and Luoma, 2003; Bustamanteand Miramand, 2005). Since the clam G. tumidum is a seafood prod-uct commonly eaten in New Caledonia, it follows that Cd could beeasily transferred to human consumers; therefore, special atten-tion should be paid to Cd contamination status in this clam.
Overall our results indicate that the oyster I. isognomon displaysa higher bioconcentration potential than the clam G. tumidum forall the elements investigated. Several previous studies have shownthat oysters generally display higher bioaccumulation capacity forsome metals such as Ag and Zn than other bivalves, probably be-cause of the occurrence of specific metal storage processes (e.g.,Metayer et al., 1990; Rainbow, 1993). Nevertheless, although lowerthan that I. isognomon, bioaccumulation capacity the clam G. tumi-dum is sufficient to allow using the clam as an effective biomonitororganism; furthermore its status of locally consumed seafoodmakes it an organism of particular interest from both biomonitor-ing and human health viewpoints.
5. Conclusions
The general similarity of uptake and depuration rates in both bi-valve species over a large range of dissolved element concentra-

566 L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567
tions confirms that element concentrations measured in clams andoysters would directly reflect the relative degree of contaminationoccurring in seawater. Furthermore, except for Co, the body- andsubcellular distribution of the elements was not influenced bythe element concentration in seawater. Bioconcentration capaci-ties (ku and Tb1/2) of both bivalves indicate that G. tumidum and I.isognomon are able to rapidly reveal the extent of the ambient dis-solved contamination and to preserve the information over rela-tively long periods of time (several weeks or months). The resultsof this study bear out the usefulness of G. tumidum and I. isognomonas biomonitor species of dissolved element contamination for sur-veying and monitoring water quality in tropical areas, and in par-ticular in the New Caledonia lagoon waters. The range of dissolvedconcentrations tested in this study covers those actually encoun-tered in the New Caledonia lagoon. Indeed, with the few limita-tions indicated previously (As and, to a lesser extent, Zn), theconcentration of the elements measured in both bivalve specieswould generally be proportional to the ambient dissolved concen-trations. Nevertheless, care should be taken when applying theseresults to other situations. Indeed, since some concentration effectswere demonstrated, results may only be valid for the range of con-centrations considered here. It has also to be kept in mind that inthe field, bivalves are exposed to several metals simultaneously.This could lead possibly to some synergetic and/or antagonist ef-fects on metal bioaccumulation as it has been previously reportedfor some metals (e.g., Hewitt and Anderson, 1978; Widdows andDonkin, 1992). However, complementary experiments carried outin the field (New Caledonia lagoon) have shown that the bioaccu-mulation of e.g., Co, Cr, Mn and Ni in the two selected bivalves wasnot influenced by the co-occurrence of other metals at high levelscompared to the present laboratory results (Hédouin et al., 2007,2008a). This suggests that the simple relationship between metalconcentration in the organisms and in seawater observed underthe present laboratory conditions could hold true in complex expo-sure conditions. Furthermore, although dissolved metal releases(via acidic lixiviation process) is currently the main concern inNew Caledonia, several studies have shown that food is often animportant uptake pathway for metals (e.g., Sanders et al., 1989;Wang et al., 1996; Reinfelder et al., 1998; Metian et al., 2008a).Therefore, food-related influences on contaminant bioaccumula-tion in these biomonitor species should be also further investigatedin order to complement the results of this study.
Acknowledgements
Authors thank J.L. Teyssié, O. Cotret and E. Giron (IAEA-MEL) forskilful technical assistance and D. Ponton (IRD-Noumea) for hishelp in SPSS statistical treatments. L.H. was beneficiary of a PhDgrant (CIFRE, France) supported by the Goro-Nickel Company,New Caledonia. M.W. is an Honorary Senior Research Associateof the National Fund for Scientific Research (NFSR, Belgium) andhas benefited from a 2008–2009 Invited Expert position at LIENSs(CNRS-Université de La Rochelle), supported by the Conseil Région-al de Poitou-Charentes. This work was supported by the IAEA, IRDand PNEC Programme (Chantier ‘‘Nouvelle-Calédonie”). The IAEA isgrateful for the support provided to its Marine Environment Labo-ratories by the Government of the Principality of Monaco.
References
Baroudi, H., Bureau, J., Rollin, C., 2003. Analyse critique de l’acceptabilité du niveaude rejet de manganèse dans le milieu marin. Rapport final Goro-Nickel INERISInstitut National de l’Environnement Industriel et des Risques, 37pp. (inFrench).
Bjerregaard, P., 1988. Effect of selenium on cadmium uptake in selected benthicinvertebrates. Marine Ecology Progress Series 48, 17–28.
Blackmore, G., Wang, W.-X., 2002. Uptake and efflux of Cd and Zn by the greenmussel Perna viridis after metal preexposure. Environmental Science &Technology 36, 989–995.
Borchardt, T., 1983. Influence of food quantity on the kinetics of cadmium uptakeand loss via food and seawater in Mytilus edulis. Marine Biology 79, 67–76.
Bouchet, P., Lozouet, P., Maestrati, P., Heros, V., 2002. Assessing the magnitude ofspecies richness in tropical marine environments: exceptionally high numbersof molluscs at a New Caledonia site. Biological Journal of the Linnean Society 75,421–436.
Boyden, C.R., 1974. Trace elements contents and body size in molluscs. Nature 251,311–314.
Bustamante, P., Miramand, P., 2005. Subcellular and body distributions of 17 traceelements in the variegated scallop Chlamys varia from the French coast of theBay of Biscay. Science of the Total Environment 337, 59–73.
Chan, H.M., 1988. Accumulation and tolerance to cadmium, copper, lead and zinc bythe green mussel Perna viridis. Marine Ecology Progress Series 48, 295–303.
Chipman, W.A., 1966. Uptake and accumulation of chromium-51 by the clam Tapesdecussatus, in relation to physical and chemical form. In: IAEA (Ed.), Disposal ofRadioactive Wastes into Seas, Oceans and Surface Waters. Proceedings of aSymposium, SM-72/35, Vienna, pp. 571–582.
Chouvelon, T., Warnau, M., Churlaud, C., Bustamante, P., 2009. Hg concentrationsand related risk assessment in coral reef crustaceans, molluscs and fish fromNew Caledonia. Environmental Pollution 157, 331–340.
Coteur, G., Gosselin, P., Wantier, P., Chambost-Manciet, Y., Danis, B., Pernet, P.,Warnau, M., Dubois, P., 2003. Echinoderms as bioindicators, bioassays, andimpact assessment tools of sediment-associated metals and PCBs in the NorthSea. Archives of Environmental Contamination and Toxicology 45, 190–202.
Dalto, A.G., Grémare, A., Dinet, A., Fichet, D., 2006. Muddy-bottom meiofaunaresponses to metal concentrations and organic enrichment in New CaledoniaSouth-West Lagoon. Estuarine, Coastal and Shelf Science 64, 629–644.
Dalvi, A.D., Bacon, W.G., Osborne, R.C., 2004. The past and the future of nickellaterites. PDAC 2004 International Convention, Trade Show & InvestorsExchange, 27pp.
Danis, B., Wantier, P., Dutrieux, S., Flammang, R., Dubois, P., Warnau, M., 2004.Contaminant levels in sediments and asteroids (Asterias rubens L.,Echinodermata) from the Belgian coast and Scheldt estuary: polychlorinatedbiphenyls and heavy metals. Science of the Total Environment 333, 149–165.
Depledge, M.H., Rainbow, P.S., 1990. Models of regulation and accumulation of tracemetals in marine invertebrates. Comparative Biochemistry and Physiology C:Comparative Pharmacology and Toxicology 97, 1–7.
Engel, D.W., Brouwer, M., 1982. Detoxification of accumulated trace metals by theAmerican oyster, Crassostrea virginica: laboratory vs. environment. In: Vernberg,W.B., Calabrese, A., Thurberg, F.P., Vernberg, F.J. (Eds.), PhysiologicalMechanisms of Marine Pollutant Toxicity. Academic Press, New York, pp. 89–107.
Fernandez, J.-M., Breau, L., Cabon, M., Fichez, R., McCorist, G., Magand, O., Moreton,B., Peck, G.A., Szymczak, R., 2002. The fate of metals in New Caledonia.Programme National des Ecosystèmes Côtiers, Banyuls, France.
Ferrier-Pagès, C., Houlbrèque, F., Wyse, E., Richard, C., Allemand, D., Boisson, F.,2005. Bioaccumulation of zinc in the scleractinian coral Stylophora pistillata.Coral Reefs 24, 636–645.
Fisher, H., 1988. Mytilus edulis as a quantitative indicator of dissolved cadmium.Final study and synthesis. Marine Ecology Progress Series 48, 163–174.
Fowler, S.W., Benayoun, G., 1974. Experimental studies on cadmium flux throughmarine biota. Comparative Studies of Food and Environmental Contamination,Vienna, IAEA SM 175/10, pp. 159–178.
Goldberg, E.D., Bowen, V.T., Farrington, J.W., Harvey, G., Martin, J.H., Parker, P.L.,Risebrough, R.W., Robertson, W., Schneider, E., Gamble, E., 1978. The MusselWatch. Environmental Conservation 5, 101–125.
Goro-Nickel, 2001. Projet Goro Nickel. Evaluation environnementale, 52pp. (inFrench).
Goro-Nickel, 2003. Annexe 5.1 Goro Nickel SA – Rapport de synthèse sur l’impact durejet en mer de l’effluent – 1 mars 2004, Document No. 47910-000-30R-GEH-40002, 36pp. (in French).
Goro-Nickel, 2004. Annexe 2.1 IRD, Rapport intermédiaire – Caractéristiquesphysico-chimiques des eaux – mars 2004, 16pp. (in French).
Hédouin, L., Bustamante, P., Churlaud, C., Pringault, O., Fichez, R., Warnau, M.,2008a. Trends in concentrations of selected metalloid and metals in twobivalves from the coral reefs in the SW lagoon of New Caledonia. Ecotoxicologyand Environmental Safety 72, 372–381.
Hédouin, L., Bustamante, P., Fichez, R., Warnau, M., 2008b. The tropical brown algaLobophora variegata as a bioindicator of mining contamination in the NewCaledonia lagoon: a field transplantation study. Marine Environmental Research66, 438–444.
Hédouin, L., Metian, M., Teyssié, J.-L., Fowler, S.W., Fichez, R., Warnau, M., 2006.Allometric relationships in the bioconcentration of heavy metals by the edibletropical clam Gafrarium tumidum. Science of the Total Environment 366, 154–163.
Hédouin, L., Pringault, O., Metian, M., Bustamante, P., Warnau, M., 2007. Nickelbioaccumulation in bivalves from the New Caledonia lagoon: seawater and foodexposure. Chemosphere 66, 1449–1457.
Hewitt, L.A., Anderson, P.D., 1978. Aspects of supra additive interactions betweencadmium and copper in fish exposed to lethal mixtures. Pharmacology 21, 210.
Jeantet, A.Y., Ballan-Dufrançais, C., Martin, J.L., 1985. Recherche des mécanismes dedétoxication du cadmium par l’huître Crassostrea gigas (Mollusque, bivalve). II.Sites intracellulaires d’accumulation du métal dans les organes absorbants et

L. Hédouin et al. / Marine Pollution Bulletin 61 (2010) 554–567 567
excréteurs. Comptes Rendus de l’Academie des Sciences Serie III Sciences de laVie 301, 177–182 (in French).
Labrosse, P., Fichez, R., Farman, R., Adams, T., 2000. New Caledonia. In: Sheppard,C.R.C. (Ed.), Seas at the Millenium: An Environmental Evaluation. Pergamon,Amsterdam, pp. 723–736.
Langston, W.J., Bebianno, M.J., Burt, G.R., 1998. Metal handling strategies inmollusks. In: Langston, W.J., Bebianno, M.J. (Eds.), Metal Metabolism inAquatic Environments. Chapman and Hall, London, UK, pp. 219–283.
Lee, B.-G., Wallace, W.G., Luoma, S.N., 1998. Uptake and loss kinetics of Cd, Cr andZn in the bivalves Potamocorbula amurensis and Macoma balthica: effect of sizeand salinity. Marine Ecology Progress Series 175, 177–189.
Martoja, R., Martin, J.L., 1985. Recherche des mécanismes de détoxication ducadmium par l’huître Crassostrea gigas (Mollusque, bivalve). I. Mise en évidenced’une protéine sulphydrilée de complexation du métal dans les amoebocytes àzinc et cuivre. Comptes Rendus de l’Academie des Sciences Serie III Sciences dela Vie 300, 549–554 (in French).
Mason, A.Z., Jenkins, K.D., 1995. Metal detoxification in aquatic organisms. In:Tessier, A., Turner, D.R. (Eds.), Metal Speciation and Bioavailability in AquaticSystems. John Wiley & Sons, Chichester, pp. 479–608.
Metayer, C., Amiard-Triquet, C., Baud, J.-P., 1990. Variations inter-spécifiques de labioaccumulation et de la toxicité de l’argent à l’égard de trois mollusquesbivalves marins (Species-related variations of silver bioaccumulation andtoxicity to three marine bivalves). Water Research 24, 995–1001.
Metian, M., 2003. Bioaccumulation des métaux lourds chez 4 espèces marines dulagon de Nouvelle Calédonie: caractérisation de leur potentiel bioindicateurpour le monitoring des activités minières locales. Master Thesis, IAEA-MEL,Monaco/Université Libre de Bruxelles, Belgium, 44pp. (in French).
Metian, M., Bustamante, P., Cosson, R.P., Hédouin, L., Warnau, M., 2008a.Investigation of Ag in the king scallop Pecten maximus using field andlaboratory approaches. Journal of Experimental Marine Biology and Ecology367, 53–60.
Metian, M., Bustamante, P., Hédouin, L., Warnau, M., 2008b. Accumulation of ninemetals and one metalloid in the tropical scallop Comptopallium radula fromcoral reefs in New Caledonia. Environmental Pollution 152, 543–552.
Metian, M., Giron, E., Borne, V., Hédouin, L., Teyssié, J.-L., Warnau, M., 2008c. Thebrown alga Lobophora variegata, a bioindicator species for surveying metalcontamination in tropical marine environments. Journal of ExperimentalMarine Biology and Ecology 362, 49–54.
Metian, M., Warnau, M., 2008. The tropical brown alga Lobophora variegata(Lamouroux) Womersley: a prospective bioindicator for Ag contamination intropical coastal waters. Bulletin of Environmental Contamination andToxicology 81, 455–458.
Mihaylov, I., Krause, E., Colton, D.F., Okita, Y., Duterque, J.-P., Perraud, J.-J., 2000. Thedevelopment of a novel hydrometallurgical process for nickel and cobaltrecovery from Goro laterite ore. Canadian Mining and Metallurgical Bulletin(Canada) 93, 124–130.
Moore, N.W., 1966. A pesticide monitoring system with special reference to theselection of indicator species. Journal of Applied Ecology 3, 261–269.
Phillips, D.J.H., 1976. The common mussel Mytilus edulis as an indicator of pollutionby zinc, cadmium, lead and copper. I. Effects of environmental variables onuptake of metals. Marine Biology 38, 59–69.
Phillips, D.J.H., 1980. Quantitative Aquatic Biological Indicators: Their Use toMonitor Trace Metal and Organochlorine Pollution. Chapman and Hall, London.488pp..
Phillips, D.J.H., 1985. Organochlorines and trace metals in green-lipped musselsPerna viridis from Hong Kong waters: a test of indicator ability. Marine EcologyProgress Series 21, 251–258.
Phillips, D.J.H., 1990. Use of macroalgae and invertebrates as monitors ofmetal levels in estuaries and coastal waters. In: Furness, R.W., Rainbow, P.S.(Eds.), Heavy Metals in the Marine Environment. CRC Press, Boca Raton, pp. 81–99.
Poulsen, E., Riisgaard, H.U., Møhlenberg, F., 1982. Accumulation of cadmium andbioenergetics in the mussel Mytilus edulis. Marine Biology 68, 25–29.
Rainbow, P.S., 1993. Biomonitoring of marine heavy metal pollution and itsapplication in Hong-Kong waters. In: Morton, B. (Ed.), The Marine Biology of theSouth China Sea. Hong Kong University Press, Hong Kong, pp. 235–250.
Rainbow, P.S., 1995. Biomonitoring of heavy metal availability in the marineenvironment. Marine Pollution Bulletin 31, 183–192.
Rainbow, P.S., Phillips, D.J.H., 1993. Cosmopolitan biomonitors of trace metals.Marine Pollution Bulletin 26, 593–601.
Reinfelder, J.R., Fisher, N.S., Luoma, S.N., Nichols, J.W., Wang, W.-X., 1998. Traceelement trophic transfer in aquatic organisms: a critique of the kinetic modelapproach. Science of the Total Environment 219, 117–135.
Richer de Forges, B., Koslow, J.A., Poore, G.C.B., 2000. Diversity and endemism of thebenthic seamount fauna in the southwest Pacific. Nature 405, 944–947.
RNO-Antilles, unpublished work. Réseau National d’Observation de la Qualité duMilieu Marin-Antilles, Ifremer et Ministère de l’Ecologie et du DéveloppementDurable (in French).
Rodriguez y Baena, A.M., Miquel, J.C., Masqué, P., Povinec, P., La Rosa, J., 2006a. Asingle vs. double spike approach to improve the accuracy of 234Thmeasurements in small-volume seawater samples. Marine Chemistry 100,269–281.
Rodriguez y Baena, A.M., Metian, M., Teyssié, J.L., De Broyer, C., Warnau, M., 2006b.Experimental evidence for 234Th bioaccumulation in three Antarcticcrustaceans: potential implications in particle flux studies. Marine Chemistry100, 354–365.
Roesijadi, G., 1992. Metallothioneins in metal regulation and toxicity in aquaticanimals. Aquatic Toxicology 22, 81–114.
Ruddell, C.L., Rains, D.W., 1975. The relationship between zinc, copper, and thebasophils of two Crassostreid oysters, C. gigas and C. virginica. ComparativeBiochemistry and Physiology 51A, 558–591.
Sanders, J.G., Osman, R.W., Riedel, G.F., 1989. Pathways of arsenic uptake andincorporation in estuarine phytoplankton and the filter-feeding invertebratesEurytemora affinis, Balanus improvisus and Crassostrea virginica. Marine Biology103, 319–325.
Sbriz, L., Aquino, M.R., Alberto de Rodriguez, N.M., Fowler, S.W., Sericano, J.L., 1998.Levels of chlorinated hydrocarbons and trace metals in bivalves and nearshoresediments from the Dominican Republic. Marine Pollution Bulletin 36, 971–979.
Simkiss, K., Mason, A.Z., 1984. Cellular responses of molluscan tissues toenvironmental metals. Marine Environmental Research 14, 103–118.
Soto, M., Cajaraville, M.P., Marigomez, I., 1996. Tissue and cell distribution ofcopper, zinc and cadmium in the mussel Mytilus galloprovincialis determined byautometallography. Tissue Cell 28, 557–568.
Stauber, J.L., Binet, M.T., Adams, M.S., 2003. Toxicité chronique du lixiviat de nickelsur le biote marin et dulciole. CSIRO, Centre for Advanced Analytical ChemistryEnergy Technology, Inco Australia Management Pty Ltd, 184pp. (in French).
Talbot, V., 1985. Relationship between cadmium concentrations in seawater andthose in the mussel Mytilus edulis. Marine Biology 85, 51–54.
Temara, A., Ledent, G., Warnau, M., Paucot, H., Jangoux, M., Dubois, P., 1996.Experimental cadmium contamination of Asterias rubens (Echinodermata).Marine Ecology Progress Series 140, 83–90.
Thébault, H., Rodriguez y Baena, A.M., Andral, B., Barisic, D., Albaladejo, J.B., Bologa,A.S., Boudjenoun, R., Delfanti, R., Egorov, V.N., El Khoukhi, T., Florou, H.,Kniewald, G., Noureddine, A., Patrascu, V., Pham, M.K., Scarpato, A., Stokozov,N.A., Topcuoglu, S., Warnau, M., 2008. 137Cs baseline levels in the Mediterraneanand Black Sea: a cross-basin survey of the CIESM Mediterranean Mussel WatchProgramme. Marine Pollution Bulletin 57, 801–806.
Ünlü, M.Y., Fowler, S.W., 1979. Factors affecting the flux of arsenic through themussel Mytilus galloprovincialis. Marine Biology 51, 209–219.
Vercauteren, K., Blust, R., 1996. Bioavailability of dissolved zinc to the commonmussel Mytilus edulis in complexing environments. Marine Ecology ProgressSeries 137, 123–132.
Wallace, W.G., Lee, B.G., Luoma, S.N., 2003. Subcellular compartmentalization of Cdand Zn in two bivalves. I. Significance of metal-sensitive fractions (MSF) andbiologically detoxified metal (BDM). Marine Ecology Progress Series 249, 183–197.
Wallace, W.G., Luoma, S.N., 2003. Subcellular compartmentalization of Cd and Zn intwo bivalves. II. Significance of trophically available metal (TAM). MarineEcology Progress Series 257, 125–137.
Wang, W.-X., Fisher, N.S., Luoma, S.N., 1996. Kinetic determinations of traceelement bioaccumulation in the mussel Mytilus edulis. Marine Ecology ProgressSeries 140, 91–113.
Warnau, M., 2002. Bioaccumulation of heavy metals and radionuclides in thePosidonia oceanica meadow, an endemic Mediterranean ecosystem. In: Metaland Radionuclide Bioaccumulation in Marine Organisms, CIESM WorkshopMonographs No. 19, Monaco.
Warnau, M., Acuña, O., 2007. Is there a need for developing an integrated basin-wide monitoring network to enhance sustainable development in theMediterranean Sea? Rapport de la Commission internationale de la MerMéditerranée 38, 42pp.
Warnau, M., Bustamante, P., 2007. Radiotracer techniques: a unique tool in marineecotoxicological studies. Environmental Bioindicators 2, 217–218.
Warnau, M., Biondo, R., Temara, A., Bouquegneau, J.-M., Jangoux, M., Dubois, P.,1998. Distribution of heavy metals in the echinoid Paracentrotus lividus (lmk)from the Mediterranean Posidonia oceanica ecosystem: seasonal andgeographical variations. Journal of Sea Research 39, 267–280.
Warnau, M., Teyssié, J.-L., Fowler, S.W., 1996. Biokinetics of selected heavy metalsand radionuclides in the common Mediterranean echinoid Paracentrotus lividus:seawater and food exposure. Marine Ecology Progress Series 141, 83–94.
Warnau, M., Teyssié, J.-L., Fowler, S.W., 1997. Cadmium bioconcentration in theechinoid Paracentrotus lividus: influence of the cadmium concentration inseawater. Marine Environmental Research 43, 303–314.
Whicker, F.W., Schultz, V., 1982. In: Schultz, V. (Ed.), Radioecology: Nuclear Energyand the Environment. CRC Press, Boca Raton, FL, USA. 320pp..
White, S.L., Rainbow, P.S., 1984. Regulation of zinc concentration by Palaemonselegans (Crustacea: Decapoda): zinc flux and effect of temperature, zincconcentration and moulting. Marine Ecology Progress Series 16, 135–147.
Widdows, J., Donkin, P., 1992. Mussels and environmental contaminants:bioaccumulation and physiological aspects. In: Gosling, E. (Ed.), The MusselMytilus: Ecology, Physiology, Genetics and Culture. Elsevier, Amsterdam, pp.383–424.
Zar, J.H., 1996. Biostatistical Analysis, third ed. Prentice-Hall, Upper Saddle River, NJ.662pp.
Copyright © 2022 FDOKUMEN




















