
Bioconcentration, metabolism and neurotoxicity of the organophorous flame retardant 1,3-dichloro...
8
Aquatic Toxicology 158 (2015) 108–115 Contents lists available at ScienceDirect Aquatic Toxicology j o ur na l ho me pag e: www.elsevier.com/locate/aquatox Bioconcentration, metabolism and neurotoxicity of the organophorous flame retardant 1,3-dichloro 2-propyl phosphate (TDCPP) to zebrafish Qiangwei Wang a,b,1 , James Chung-Wah Lam c,d,e,1 , Yin-Chung Man c,d,e , Nelson Lok-Shun Lai c,d,e , Karen Ying Kwok c,d,e , Yong yong Guo a , Paul Kwan-Sing Lam c,d,e , Bingsheng Zhou a,∗ a State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China b University of Chinese Academy of Sciences, Beijing 100049, China c State Key Laboratory in Marine Pollution; Department of Biology and Chemistry, City University of Hong Kong, Kowloon, Hong Kong SAR, China d Research Centre for the Oceans and Human Health, City University of Hong Kong, Shenzhen Research Institute Building, Shenzhen 518057, China e Department of Biology and Chemistry, City University of Hong Kong, Kowloon, Hong Kong SAR, China a r t i c l e i n f o Article history: Received 5 August 2014 Received in revised form 23 October 2014 Accepted 1 November 2014 Available online 7 November 2014 Keywords: TDCPP Bioconcentration and metabolism Neurotoxicity Gender-specific toxicity Zebrafish a b s t r a c t Organophosphate flame retardants are ubiquitous environmental contaminants; however, knowledge is limited regarding their environmental health risks and toxicity. Here, we investigated the effects of acute and long-term exposure to tris(1,3-dichloro-2-propyl) phosphate (TDCPP) to the nervous system of zebrafish. Zebrafish embryos (2 h post-fertilization) were exposed to TDCPP (0–100 g/L) for 6 months up until sexual maturation. Concentrations of TDCPP and its metabolic product (bis(1,3-dichloro-2-propyl) phosphate, BDCPP) were measured in the tissues of 5 day post-fertilization (dpf) larvae. There was no effect on locomotion, acetylcholinesterase activity, levels of the neurotransmitters dopamine and sero- tonin, and expression of mRNAs and proteins related to central nervous system development (e.g., myelin basic protein, ˛1-tubulin) in any exposure group. However, in adult fish, reductions of dopamine and serotonin levels were detected in the brains of females but not males. Downregulation of nervous sys- tem development genes was observed in both the male and female brain tissues. TDCPP concentrations were measured in adult fish tissues including the brain, and greater levels were detected in females. Our results showed that females are more sensitive to TDCPP stress than males in terms of TDCPP-induced neurotoxicity. We demonstrate that long-term exposure to lower concentrations of TDCPP in fish can lead to neurotoxicity. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Organophosphorus flame retardants (OPFRs) have been used as additives in various household and industrial products, such Abbreviations: AChE, acetylcholinesterase; BDCPP, bis(1,3-dichloro-2-propyl) phosphate; OP, organophosphorus; dpf, days post-fertilization; d21-TPrP, deuter- ated tri-n-propyl phosphate; d9-TMP, deuterated trimethyl phosphate; DMSO, dimethylsulfoxide; gfap, glial fibrillary acidic protein; hpf, hours post-fertilization; LC, liquid chromatograph; mbp, myelin basic protein; OPFRs, organophosphorus flame retardants; PBDEs, polybrominated diphenyl ethers; SEM, standard error; syn2a, synapsin IIa; TH, thyroid hormone; TCPP, tris(2-chloro-1-methylethyl) phos- phate; TDCPP, tris(1,3-dichloro-2-propyl) phosphate. ∗ Corresponding author. Tel.: +86 27 68780042; fax: +86 27 68780123. E-mail address: [email protected] (B. Zhou). 1 Co-first author. as electronic equipment, plastic products, textiles, furniture and building materials (Reemtsma et al., 2008). Due to the adverse health effects caused by brominated flame retardants, such as poly- brominated diphenyl ethers (PBDEs), many industrial countries have banned or phased-out PBDEs, and OPFRs have been proposed as alternatives (Stapleton et al., 2009). Consequently, the produc- tion and usage of OPFRs has increased and now they are ubiquitous environmental contaminants and they are also detectable in biota samples (Stapleton et al., 2009; Van der Veen and de Boer, 2012). In river water, OPFR concentrations are generally at ng–g/L lev- els (Martinez-Carballo et al., 2007; Regnery and Püttmann, 2010; Cristale et al., 2013), and highest concentrations of tris(2-chloro-1- methylethyl) phosphate (TCPP) were reported in United Kingdom rivers (up to 26 g/L) (Cristale et al., 2013). OPFRs have been detected in river sediments too, and here they can reach up to 1300 g/kg as reported for the River Schwechat in Austria http://dx.doi.org/10.1016/j.aquatox.2014.11.001 0166-445X/© 2014 Elsevier B.V. All rights reserved.
Transcript of Bioconcentration, metabolism and neurotoxicity of the organophorous flame retardant 1,3-dichloro...

Bo(
QNPa
b
c
d
e
a
ARRAA
KTBNGZ
1
a
padLflsp
h0
Aquatic Toxicology 158 (2015) 108–115
Contents lists available at ScienceDirect
Aquatic Toxicology
j o ur na l ho me pag e: www.elsev ier .com/ locate /aquatox
ioconcentration, metabolism and neurotoxicity of therganophorous flame retardant 1,3-dichloro 2-propyl phosphateTDCPP) to zebrafish
iangwei Wanga,b,1, James Chung-Wah Lamc,d,e,1, Yin-Chung Manc,d,e,elson Lok-Shun Lai c,d,e, Karen Ying Kwokc,d,e, Yong yong Guoa,aul Kwan-Sing Lamc,d,e, Bingsheng Zhoua,∗
State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, ChinaUniversity of Chinese Academy of Sciences, Beijing 100049, ChinaState Key Laboratory in Marine Pollution; Department of Biology and Chemistry, City University of Hong Kong, Kowloon, Hong Kong SAR, ChinaResearch Centre for the Oceans and Human Health, City University of Hong Kong, Shenzhen Research Institute Building, Shenzhen 518057, ChinaDepartment of Biology and Chemistry, City University of Hong Kong, Kowloon, Hong Kong SAR, China
r t i c l e i n f o
rticle history:eceived 5 August 2014eceived in revised form 23 October 2014ccepted 1 November 2014vailable online 7 November 2014
eywords:DCPPioconcentration and metabolismeurotoxicityender-specific toxicity
a b s t r a c t
Organophosphate flame retardants are ubiquitous environmental contaminants; however, knowledgeis limited regarding their environmental health risks and toxicity. Here, we investigated the effects ofacute and long-term exposure to tris(1,3-dichloro-2-propyl) phosphate (TDCPP) to the nervous system ofzebrafish. Zebrafish embryos (2 h post-fertilization) were exposed to TDCPP (0–100 �g/L) for 6 months upuntil sexual maturation. Concentrations of TDCPP and its metabolic product (bis(1,3-dichloro-2-propyl)phosphate, BDCPP) were measured in the tissues of 5 day post-fertilization (dpf) larvae. There was noeffect on locomotion, acetylcholinesterase activity, levels of the neurotransmitters dopamine and sero-tonin, and expression of mRNAs and proteins related to central nervous system development (e.g., myelinbasic protein, ˛1-tubulin) in any exposure group. However, in adult fish, reductions of dopamine andserotonin levels were detected in the brains of females but not males. Downregulation of nervous sys-
ebrafish tem development genes was observed in both the male and female brain tissues. TDCPP concentrationswere measured in adult fish tissues including the brain, and greater levels were detected in females. Ourresults showed that females are more sensitive to TDCPP stress than males in terms of TDCPP-inducedneurotoxicity. We demonstrate that long-term exposure to lower concentrations of TDCPP in fish canlead to neurotoxicity.
. Introduction
Organophosphorus flame retardants (OPFRs) have been useds additives in various household and industrial products, such
Abbreviations: AChE, acetylcholinesterase; BDCPP, bis(1,3-dichloro-2-propyl)hosphate; OP, organophosphorus; dpf, days post-fertilization; d21-TPrP, deuter-ted tri-n-propyl phosphate; d9-TMP, deuterated trimethyl phosphate; DMSO,imethylsulfoxide; gfap, glial fibrillary acidic protein; hpf, hours post-fertilization;C, liquid chromatograph; mbp, myelin basic protein; OPFRs, organophosphorusame retardants; PBDEs, polybrominated diphenyl ethers; SEM, standard error;yn2a, synapsin IIa; TH, thyroid hormone; TCPP, tris(2-chloro-1-methylethyl) phos-hate; TDCPP, tris(1,3-dichloro-2-propyl) phosphate.∗ Corresponding author. Tel.: +86 27 68780042; fax: +86 27 68780123.
E-mail address: [email protected] (B. Zhou).1 Co-first author.
ttp://dx.doi.org/10.1016/j.aquatox.2014.11.001166-445X/© 2014 Elsevier B.V. All rights reserved.
© 2014 Elsevier B.V. All rights reserved.
as electronic equipment, plastic products, textiles, furniture andbuilding materials (Reemtsma et al., 2008). Due to the adversehealth effects caused by brominated flame retardants, such as poly-brominated diphenyl ethers (PBDEs), many industrial countrieshave banned or phased-out PBDEs, and OPFRs have been proposedas alternatives (Stapleton et al., 2009). Consequently, the produc-tion and usage of OPFRs has increased and now they are ubiquitousenvironmental contaminants and they are also detectable in biotasamples (Stapleton et al., 2009; Van der Veen and de Boer, 2012).In river water, OPFR concentrations are generally at ng–�g/L lev-els (Martinez-Carballo et al., 2007; Regnery and Püttmann, 2010;Cristale et al., 2013), and highest concentrations of tris(2-chloro-1-
methylethyl) phosphate (TCPP) were reported in United Kingdomrivers (up to 26 �g/L) (Cristale et al., 2013). OPFRs have beendetected in river sediments too, and here they can reach upto 1300 �g/kg as reported for the River Schwechat in Austria
oxicol
(O
iuaTetBiiTaIp
2ec0WbRtt1p(i1t
pdVsaeadhoeeiiTTc(cld2tnaielpt
tf
Q. Wang et al. / Aquatic T
Martinez-Carballo et al., 2007). In marine fish, concentrations ofPFRs can reach 2000 ng/g lipid weight (Kim et al., 2011).
Tris(1,3-dichloro-2-propyl) phosphate (TDCPP) is an OPFR thats commonly used as an additive in polyurethane foam paddingsed in furniture, children’s foam products and in upholstery forutomobiles (Dishaw et al., 2011; Stapleton et al., 2011, 2012).DCPP is found in indoor dust at high concentrations (Stapletont al., 2009), while in the aquatic environment, TDCPP concentra-ions in surface waters are typically <100 ng/L (Van der Veen and deoer, 2012). However, TDCPP at up to several �g/L can be detected
n water at waste disposal sites; for example, 3 �g/L was detectedn effluent from a sewage treatment plant (Marklund et al., 2005).DCPP is not readily degraded in water and it tends to persist andccumulate in the environment, which maybe bioavailable to fish.ndeed, up to 140 ng/g lipid weight of TDCPP has been measured inerch (Sundkvist et al., 2010).
In China, more than 70,000 tons of OPFRs were produced in007, and this accounted for 35% of global production (Reemtsmat al., 2008). In China, limited data show that the surface wateroncentration of TDCPP is 2.5–40 ng/L in the Songhua River and.62–5.54 �g/kg in sediment from Taihu Lake (Cao et al., 2012;ang et al., 2011). Up to 251 �g/kg lipid weight of TDCPP has
een reported in muscle samples of freshwater fish from the Pearliver Delta region in South China (Ma et al., 2013), where tribu-oxyethyl phosphate is the dominant OPFR and this can reach upo 8842 �g/kg lipid weight. Total OPFRs were detected at up to312 �g/kg dry weight in sludge samples collected from munici-al wastewater treatment plants in the same region of South ChinaZeng et al., 2014). OPFRs have also been detected in tap watern China (85.1–325 ng/L) (Li et al., 2014) and in office air (up to47.7 ng/m3) (Yang et al., 2014). Thus, the production and applica-ion of OPFRs has resulted in environmental contamination.
Toxicological studies have shown that exposure to OPFRs has theotential to cause neurological, adverse reproductive, endocrineisruptive and systemic effects in animals (Dishaw et al., 2011;an der Veen and de Boer, 2012). In vitro reporter gene assayshowed that TDCPP has potential endocrine-disrupting effectsnd can act as an androgen receptor (AR) antagonist (Kojimat al., 2013). TDCPP may have thyroid endocrine-disrupting effectss concentrations of this chemical in house dust correlate withecreased concentrations of circulating thyroid hormone (TH) inumans (Meeker and Stapleton, 2009). TDCPP can cause devel-pmental abnormalities during zebrafish embryogenesis (Mcgeet al., 2012; Fu et al., 2013; Liu et al., 2013a), including thyroidndocrine-disruption activity, as it reduced the levels of circulat-ng T4 levels in zebrafish larvae (Wang et al., 2013). Moreover,n cultured chicken embryos TDCPP caused altered expression ofH-responsive genes (Crump et al., 2012; Farhat et al., 2013).DCPP also disrupted sex hormone levels in human adrenocorticalarcinoma cells (H295R) and impaired reproduction in zebrafishLiu et al., 2012, 2013b). A recent DNA microarray study inhicken embryos injected with TDCPP demonstrated toxicity toipid metabolism, including cholesterol, as well as a transcriptionalysregulation of genes involved in immune response (Farhat et al.,014). In vitro studies show that TDCPP exhibits similar neuro-oxic actions to organophosphorus (OP) pesticides (e.g., alteredeurodifferentiation) (Dishaw et al., 2011) and exerts cytotoxicnd neurotoxic effects to PC12 cells (e.g., decreased cell growth,ncreased apoptosis, altered cell morphology, and changes in genexpression) (Ta et al., 2014). However, knowledge of the toxico-ogical and environmental health effects of TDCPP remains limited,articularly the effects of long-term exposures to low concentra-
ions and whether TDCPP causes neurotoxicity in vivo.The objectives of the present study were to investigate poten-ial neurotoxicity of TDCPP in fish. As public health concerns haveocused primarily on the potential hazardous effects resulting
ogy 158 (2015) 108–115 109
from long-term exposure, especially to infants and young children(Staskal et al., 2006), we examined the neurotoxicity of TDCPPexposure at an early developmental stage of zebrafish (5 dayspost-fertilization [dpf]) and after long-term exposure (6 months).Moreover, early developmental stages are typically more sen-sitive to toxicant stress, particularly the developing brain (Riceand Barone, 2000), thus we examined the expression of severalgenes that are expressed exclusively in the nervous system dur-ing early developmental stages of zebrafish (Brosamle and Halpern,2002; Udvadia et al., 2001). These candidate genes can serveas biomarkers for developmental neurotoxicity by demonstratingtheir responsiveness to neurotoxicants (Fan et al., 2010). Acetyl-cholinesterase (AChE) activity is widely used as a biomarker forthe presence of neurotoxicants in aquatic environments, and thisassay has been used to detect OP pesticides (Payne et al., 1996).The evaluation of neurotransmitter parameters has emerged asan important strategy to assess neurochemical, behavioral andtoxicological phenotypes in zebrafish (Rico et al., 2011), and thelocomotion abilities of larvae can also indicate toxicity (Rao et al.,2005). Therefore, we also examined the neurotransmitter sys-tem, including AChE activity and neurotransmitter concentrations(e.g., dopamine and serotonin). Taken together, this informationwill further elucidate the environmental risks of OPFRs to aquaticorganisms. Since little information is available regarding biocon-centrations of TDCPP and its metabolism in fish, we also assess thechemical accumulation of TDCPP in the whole body and the brainof zebrafish.
2. Materials and methods
2.1. Reagents
Reagents were purchased from the following sources: TDCPP(CAS#13674-87-8; >95.6% purity) from TCI Tokyo ChemicalIndustry Co., Ltd. (Tokyo, Japan); bis(1,3-dichloro-2-propyl)phosphate (BDCPP, >97% purity) from Wellington Laborato-ries, Ontario, Canada; deuterated tri-n-propyl phosphate (d21-TPrP, 99.2%), deuterated trimethyl phosphate (d9-TMP, 99.9%)from CDN Isotopes Inc. (Pointe-Claire, Quebec, Canada), 3,4-dihydroxybenzylamine (DHB, >98.0% purity), dopamine (>98.5%purity) and serotonin (>98.0% purity) from Sigma–Aldrich, St. Louis,USA. TDCPP stock solutions and serial dilutions were preparedin dimethylsulfoxide (DMSO) (purity >99.9%; Amresco, Solon, OH,USA). All other chemicals used were of analytical grade.
2.2. Zebrafish maintenance and experimental design
Adult zebrafish (AB strain) were maintained and embryos wereexposed to toxicants according to a published protocol (Yu et al.,2011). Briefly, normal embryos were randomly selected at theblastula stage (2 h post-fertilization [hpf]) and transferred intoglass beakers containing 500 mL of TDCPP solution (0, 4, 20 and100 �g/L). Experiments were carried out in triplicate for eachexposure concentration and each beaker contained approximately300 embryos. The selection of the lowest exposure concentration(4 �g/L) was based on a previous study (Wang et al., 2013), wheredecreased T4 levels in zebrafish larvae were observed after short-term exposure (144 h) to 10 �g/L TDCPP, and this concentrationis also similar to that reported in sewage effluent (e.g., 3 �g/L)(Marklund et al., 2005). A subset of the 5-dpf larvae was sampledat random to measure locomotion activity, AChE activity, neu-
rotransmitter levels, gene and protein expression, and chemicalconcentrations. Fish (10 days old) were placed in 20-L aquaria (50larvae per aquaria) and at 40 dpf the larvae were transferred into30-L aquaria (40 fry per aquaria). The fish were aerated, kept at
1 oxicol
2dpcimZbapctN
2a
fu(wmtwes(r
2
a(tdtT28w�amfAawcK(
2
a(wip−s5t
10 Q. Wang et al. / Aquatic T
8 ◦C and exposed to an artificial day-night cycle (14 h light, 10 hark). During all the experimental period, exposure media wererepared with dechlorinated carbon-filtered water and replacedompletely with freshly prepared media daily. Control and exper-mental treatment groups received 0.001% (v/v) DMSO. After 6
onths of exposure, survival and growth (weight) were recorded.ebrafish were euthanized using an overdose of MS-222 (300 mg/L)y prolonged immersion until cessation of opercular movementnd total weights and snout-to-vent lengths were recorded. Sam-les of brain were collected and preserved at −80 ◦C for laterhemical analyses. All studies were conducted in accordance withhe guidelines for the care and use of laboratory animals of theational Institute for Food and Drug Control of China.
.3. Quantitative real-time polymerase chain reaction (qRT-PCR)ssay
Adult fish were euthanized as above and brains were removedrom the skull by dissection. Three brains were pooled to besed as one replicate for each treatment. The brain samplesn = 3 replicates) and larval samples (30 larvae, n = 3 replicates)ere collected and preserved with RNAiso Plus according to theanufacturer’s instructions (Takara, Dalian, China). RNA extrac-
ion, purification, quantification and first-strand cDNA synthesisere performed following the protocols described by Chen
t al. (2012). Oligonucleotide primers specific to each of theelected genes were identified using the online Primer 3 programhttp://frodo.wi.mit.edu/) (Supplementary material, Table S1). Thepl8 gene was selected as the internal standard.
.4. Protein extraction and Western blotting
A Western blot was performed as previously described usingpproximately 150 larvae and brain samples for each replicaten = 3 replicates; Chen et al., 2012). Briefly, 50 �g of each pro-ein sample was separated by electrophoresis on a 12% sodiumodecylsulfate polyacrylamide gel. Following electrophoresis, pro-eins were transferred to polyvinylidene difluoride membranes.he membranes were further blocked with 5% fat-free milk for
h in tris-buffered saline (TBS) (10 mM tris, 150 mM NaCl, pH.0) and 0.1% Tween 20, before being incubated overnight at 4 ◦Cith primary antibodies against myelin basic protein (mbp; 1:300),1-tubulin (1:100) or �-actin (1:1000). Both rabbit ¦Á1-tubulinntibody (Abcam, Cambridge, UK) and rabbit mbp (AnaSpec, Fre-ont, CA) have previously been verified as reactive and suitable
or zebrafish studies (Thatcher et al., 2008; Buckley et al., 2010).fter incubation with horseradish peroxidase-conjugated goatnti-rabbit IgG (1:50,000) for 2 h at room temperature, the signalas detected by enhanced chemiluminescence. The relative opti-
al density of each band was analyzed using Kodak film (Eastmanodak Co., Rochester, NY) and Quantity One version 4.3 software
Bio-Rad, USA).
.5. Neurotransmitter measurements
Monoamine extraction from both larvae (50, n = 3 replicates)nd from adult fish brains was performed as previously describedWang et al., 2012; Sari et al., 2010). Before extraction, 1 ng of DHBas added as internal standard (Sari et al., 2010). After weigh-
ng and homogenizing in 400 �L of 1% formic acid dissolved inre-cooled acetonitrile, the samples were incubated for 15 min at
20 ◦C, before centrifuging at 12,000 × g for 20 min at 4 ◦C. Theupernatant was freeze-dried and reconstituted by vortexing in0 �L of 50% acetonitrile/water (v/v). Samples were further cen-rifuged at 12,000 × g for 20 min at 4 ◦C and the supernatant was
ogy 158 (2015) 108–115
transferred to a new tube and stored at −20 ◦C until chemical anal-yses were performed.
Identification and quantification of analytes were carriedout using an Agilent 1200 (Agilent, USA) liquid chromatographequipped with a QQQ (Agilent, USA) tandem mass spectrometer.Dopamine and serotonin were used as the analytical standards.Agilent ZorbaxXDB-C18 (2.1 × 50 mm, 1.8 �m) was used for LC sep-aration. Data were acquired with MassLynx 4.0 and processed withQuanLynx software for calibration and for quantification of ana-lytes.
2.6. AChE activity in larvae and in adult fish brains
Adult fish were euthanized and brains removed as above. Threebrains were pooled to be used as one replicate for each treat-ment. The brains (n = 3 replicates) and larvae (50, n = 3 replicates)were homogenized on ice in 60 volumes (v/w) of tris-citrate buffer(50 mM tris, 2 mM EDTA, 2 mM EGTA; pH 7.4, adjusted with cit-ric acid) (Chen et al., 2012), and then centrifuged at 3000 × g for10 min at 4 ◦C. AChE enzyme activity was measured by a commer-cial kit following the manufacturer’s instructions (Nanjing KeygenBiotech, Co, Ltd.). Protein concentration was measured using theBradford method.
2.7. Locomotion activity measurement
Quantification of larval locomotion activity was performedusing the Video-Track system (ViewPoint Life Sciences, Inc., Mon-treal, Canada) following a previously described method (Chen et al.,2012). The 5-dpf zebrafish larvae were placed into 24-well platewells (1 larva per well) to measure swimming activity. Larvae wereallowed to acclimate for 10 min before the test started. Swim-ming behavior was monitored for 20 min in continuous light andalso in response to 40 min dark-to-light photoperiod stimulation(cycle of 10 min visible light followed by 10 min infrared light). Data(frequency of movements, distance traveled, and total duration ofmovement) were collected from 30 larvae per treatment every 60 s,and further analyzed using custom Open Office.Org 2.4 software.
2.8. Quantification of TDCPP and BDCPP
2.8.1. Extraction and clean-upActual TDCPP concentrations in the exposure solutions were
measured at the embryo, larval and adult stages before and afterrenewal of the water solution (n = 3 replicate tanks). TDCPP wasextracted and analyzed as described previously (Wang et al., 2011).
TDCPP and BDCPP concentrations were determined in 5-dpf lar-vae (100, n = 3 replicates), in adult fish (one individual fish) andbrain tissue (three, n = 3 replicates). d21-TPrP and d9-TMP wereused as internal standards.
Zebrafish larvae samples, adult fish and brain tissues wereweighed and spiked with 100 ng of d21-TPrP and d9-TMP as inter-nal standards for TDCPP and BDCPP, and then 4 mL acetonitrile wasadded. Samples were homogenized, sonicated for 30 min, and cen-trifuged at 3000 × g for 5 min. The supernatant was collected andthe extraction was repeated two additional times using 3 mL ace-tonitrile. The supernatants were combined and then concentratedto 1 mL under nitrogen. The extract was reconstituted in 125 mLwater, transferred to solid-phase extraction cartridges (Bond Elut-PPL 200 mg, 3 mL), eluted with 2 ml acetonitrile and 2 mL methanol,and then diluted to 10 mL with methanol. The sample was mixedin 100 �L methanol:water (1:1) before instrumental analysis.
2.8.2. LC–MS/MS analysisTDCPP and BDCPP identification and quantification was per-
formed using a LC-tandem mass spectrometer system consisting
oxicology 158 (2015) 108–115 111
oc
2
twdtasadtweBT7
2
irDeTCc
3
3
cSow(2o
us1(8bt
3
dTtesftoarg
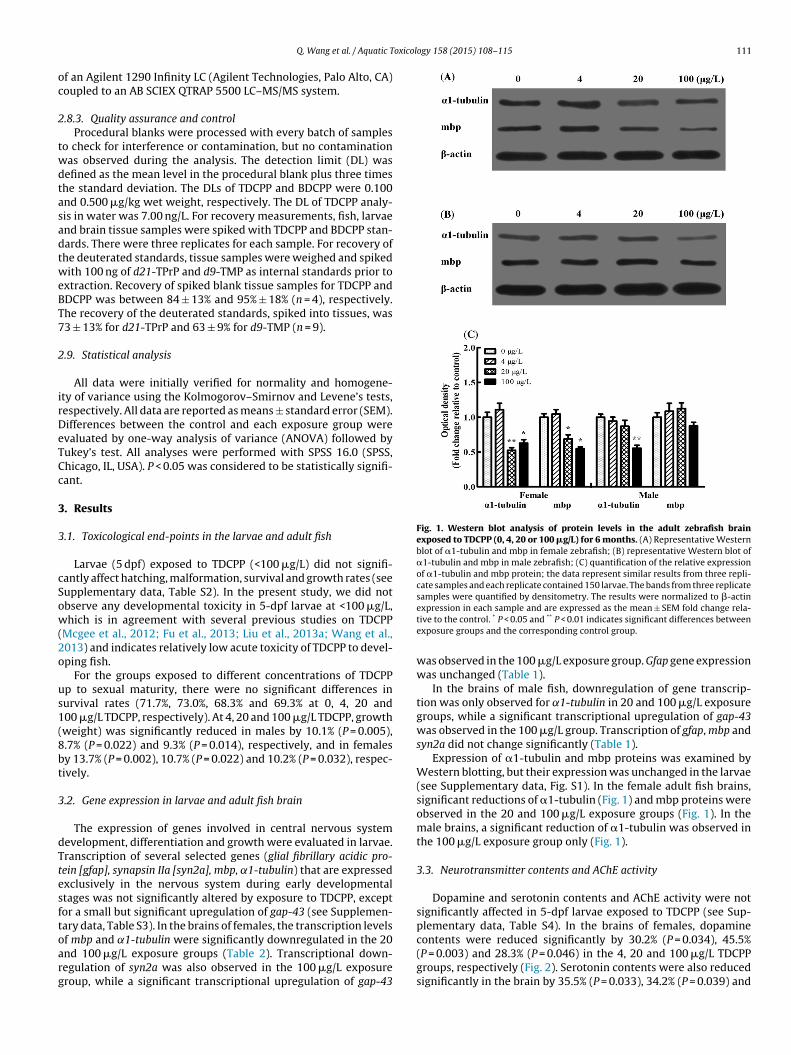
Fig. 1. Western blot analysis of protein levels in the adult zebrafish brainexposed to TDCPP (0, 4, 20 or 100 �g/L) for 6 months. (A) Representative Westernblot of �1-tubulin and mbp in female zebrafish; (B) representative Western blot of�1-tubulin and mbp in male zebrafish; (C) quantification of the relative expressionof �1-tubulin and mbp protein; the data represent similar results from three repli-cate samples and each replicate contained 150 larvae. The bands from three replicatesamples were quantified by densitometry. The results were normalized to �-actin
Q. Wang et al. / Aquatic T
f an Agilent 1290 Infinity LC (Agilent Technologies, Palo Alto, CA)oupled to an AB SCIEX QTRAP 5500 LC–MS/MS system.
.8.3. Quality assurance and controlProcedural blanks were processed with every batch of samples
o check for interference or contamination, but no contaminationas observed during the analysis. The detection limit (DL) wasefined as the mean level in the procedural blank plus three timeshe standard deviation. The DLs of TDCPP and BDCPP were 0.100nd 0.500 �g/kg wet weight, respectively. The DL of TDCPP analy-is in water was 7.00 ng/L. For recovery measurements, fish, larvaend brain tissue samples were spiked with TDCPP and BDCPP stan-ards. There were three replicates for each sample. For recovery ofhe deuterated standards, tissue samples were weighed and spikedith 100 ng of d21-TPrP and d9-TMP as internal standards prior to
xtraction. Recovery of spiked blank tissue samples for TDCPP andDCPP was between 84 ± 13% and 95% ± 18% (n = 4), respectively.he recovery of the deuterated standards, spiked into tissues, was3 ± 13% for d21-TPrP and 63 ± 9% for d9-TMP (n = 9).
.9. Statistical analysis
All data were initially verified for normality and homogene-ty of variance using the Kolmogorov–Smirnov and Levene’s tests,espectively. All data are reported as means ± standard error (SEM).ifferences between the control and each exposure group werevaluated by one-way analysis of variance (ANOVA) followed byukey’s test. All analyses were performed with SPSS 16.0 (SPSS,hicago, IL, USA). P < 0.05 was considered to be statistically signifi-ant.
. Results
.1. Toxicological end-points in the larvae and adult fish
Larvae (5 dpf) exposed to TDCPP (<100 �g/L) did not signifi-antly affect hatching, malformation, survival and growth rates (seeupplementary data, Table S2). In the present study, we did notbserve any developmental toxicity in 5-dpf larvae at <100 �g/L,hich is in agreement with several previous studies on TDCPP
Mcgee et al., 2012; Fu et al., 2013; Liu et al., 2013a; Wang et al.,013) and indicates relatively low acute toxicity of TDCPP to devel-ping fish.
For the groups exposed to different concentrations of TDCPPp to sexual maturity, there were no significant differences inurvival rates (71.7%, 73.0%, 68.3% and 69.3% at 0, 4, 20 and00 �g/L TDCPP, respectively). At 4, 20 and 100 �g/L TDCPP, growthweight) was significantly reduced in males by 10.1% (P = 0.005),.7% (P = 0.022) and 9.3% (P = 0.014), respectively, and in femalesy 13.7% (P = 0.002), 10.7% (P = 0.022) and 10.2% (P = 0.032), respec-ively.
.2. Gene expression in larvae and adult fish brain
The expression of genes involved in central nervous systemevelopment, differentiation and growth were evaluated in larvae.ranscription of several selected genes (glial fibrillary acidic pro-ein [gfap], synapsin IIa [syn2a], mbp, ˛1-tubulin) that are expressedxclusively in the nervous system during early developmentaltages was not significantly altered by exposure to TDCPP, exceptor a small but significant upregulation of gap-43 (see Supplemen-ary data, Table S3). In the brains of females, the transcription levels
f mbp and ˛1-tubulin were significantly downregulated in the 20nd 100 �g/L exposure groups (Table 2). Transcriptional down-egulation of syn2a was also observed in the 100 �g/L exposureroup, while a significant transcriptional upregulation of gap-43expression in each sample and are expressed as the mean ± SEM fold change rela-tive to the control. * P < 0.05 and ** P < 0.01 indicates significant differences betweenexposure groups and the corresponding control group.
was observed in the 100 �g/L exposure group. Gfap gene expressionwas unchanged (Table 1).
In the brains of male fish, downregulation of gene transcrip-tion was only observed for ˛1-tubulin in 20 and 100 �g/L exposuregroups, while a significant transcriptional upregulation of gap-43was observed in the 100 �g/L group. Transcription of gfap, mbp andsyn2a did not change significantly (Table 1).
Expression of �1-tubulin and mbp proteins was examined byWestern blotting, but their expression was unchanged in the larvae(see Supplementary data, Fig. S1). In the female adult fish brains,significant reductions of �1-tubulin (Fig. 1) and mbp proteins wereobserved in the 20 and 100 �g/L exposure groups (Fig. 1). In themale brains, a significant reduction of �1-tubulin was observed inthe 100 �g/L exposure group only (Fig. 1).
3.3. Neurotransmitter contents and AChE activity
Dopamine and serotonin contents and AChE activity were notsignificantly affected in 5-dpf larvae exposed to TDCPP (see Sup-plementary data, Table S4). In the brains of females, dopamine
contents were reduced significantly by 30.2% (P = 0.034), 45.5%(P = 0.003) and 28.3% (P = 0.046) in the 4, 20 and 100 �g/L TDCPPgroups, respectively (Fig. 2). Serotonin contents were also reducedsignificantly in the brain by 35.5% (P = 0.033), 34.2% (P = 0.039) and
112 Q. Wang et al. / Aquatic Toxicol
Fig. 2. Neurotransmitter content in female brain tissues exposed to TDCPP (0, 4,20 or 100 �g/L) for 6 months. The neurotransmitter (dopamine, serotonin) levelsare expressed as ng/g. ww. All data are expressed as mean ± SEM of three replicatess
3Trb
3
ap
3
cae
ca(0T
wTso2t((
TGpt
amples; each replicate contained 3 brain tissues. * P < 0.05 and ** P < 0.01 indicatesignificant differences between exposure groups and control group.
2.9% (P = 0.047) in the females exposed to 4, 20 and 100 �g/LDCPP, respectively (Fig. 2). Long-term exposure did not affect neu-otransmitter contents in male fish or AChE activity in the adult fishrains (see Supplementary data, Table S4).
.4. Locomotion activity
There were no significant changes in 5-dpf larvae locomotion inny of the exposure groups compared with the control (see Sup-lementary data, Fig. S2).
.5. Quantification of TDCPP and BDCPP
The close correlation between the nominal and measured TDCPPoncentrations (Table 2) indicated the stability of TDCPP in water,nd this observation is consistent with a previous study (Wangt al., 2013). No TDCPP was detected in the control group.
TDCPP contents (�g/kg wet weight) in the larvae exposed to thishemical increased with exposure concentration, while substantialmounts of its metabolite, BDCPP, were detected in exposed larvaeFig. 3A). The ratio of TDCPP: BDCPP was 2.45 ± 0.03, 0.99 ± 0.17 and.84 ± 0.01 in 4, 20 and 100 �g/L exposure groups, respectively. NoDCPP or BDCPP was detected in the control group.
The total body burden and brain contents of TDCPP and BDCPPere measured in adult fish. In males, the total body burden of
DCPP and BDCPP showed a concentration-dependent relation-hip in 4, 20 and 100 �g/L exposure groups (Fig. 3B). The ratiof TDCPP: BDCPP was 2.27 ± 1.22, 6.95 ± 0.8 and 5.56 ± 3.22 in 4,
0 and 100 �g/L exposure groups, respectively. Brain tissue con-ained considerable amounts of TDCPP, but no BDCPP was detectedFig. 3B). Control males contained detectable amounts of TDCPP0.9 ± 0.4 �g/kg), while BDCPP content was <0.5 �g/kg wet weight.able 1ene transcription levels in adult zebrafish brain exposed to TDCPP (0, 4, 20 and 100 �ooled as one replicate sample) and are expressed as fold change relative to control.* P <
he control group.
ZebrafishTDCPP (�g/L)
Female
0 4 20 100
gfap 1.02 ± 0.10 1.17 ± 0.20 1.14 ± 0.18 1.13 ±syn2a 1.01 ± 0.06 1.14 ± 0.16 1.17 ± 0.13 0.55 ±mbp 1.01 ± 0.10 0.88 ± 0.16 0.50 ± 0.10* 0.48 ±˛1-tubulin 1.01 ± 0.09 0.78 ± 0.10 0.51 ± 0.13* 0.54 ±gap-43 1.00 ± 0.04 1.38 ± 0.18 1.75 ± 0.13 2.35 ±
ogy 158 (2015) 108–115
In females, the measured total body burden of TDCPP and BDCPPalso showed a concentration-dependent relationship in the 4, 20and 100 �g/L exposure groups (Fig. 3C). The ratio of TDCPP: BDCPPwas 3.39 ± 0.63, 3.23 ± 2.04 and 8.73 ± 3.83 in 4, 20 and 100 �g/Lexposure groups, respectively. A similar deposition of TDCPP wasalso detected in female fish brain tissue (Fig. 3C). Moreover, a smallamount of BDCPP was detected in the brains from the highest expo-sure group (100 �g/L). Negligible levels of TDCPP (0.8 ± 0.6 �g/kg)were detected, while BDCPP content was <0.5 �g/kg wet weight incontrol females.
4. Discussion
Although several recent studies have described the potentialneurotoxicity of TDCPP to cultured PC12 cells, the potential neu-rotoxicity of TDCPP to animals is currently unknown. In addition,to our knowledge, few studies have been conducted to assess theimpact of long-term exposure to low concentrations of OPFRs infish. In the present study, we observed no acute toxicity to devel-oping zebrafish larvae, but we did observe significant decreases indopamine and seratonin within adult female zebrafish after long-term exposure to TDCPP, which further supports the importanceof assessing the risks of long-term low exposures to environmen-tal OPFRs. Furthermore, we found evidence for tissue burden andmetabolism of TDCPP in the larvae and adult zebrafish, particularlyin the brain.
In the present study, we observed high body burdens ofTDCPP in fish, indicating bioavailability and bioaccumulation ofthis hydrophobic chemical under laboratory exposure conditions(log Kow = 3.76) (OEHHA, 2011). We also observed the metaboliteBDCPP, which is the primary metabolite of TDCPP produced inmammals (Cooper et al., 2011; Carignan et al., 2013). The relativelyhigh content of BDCPP indicates a high TDCPP biotransformationcapacity by enzymes in the fish liver. In addition, unusually highTDCPP contents were detected in brain tissue, suggesting that it isable to cross the blood brain barrier and accumulates in brain sug-gesting that the brain may be a key target organ for TDCPP toxicityand this may have direct effects on nerve cells, leading to neuro-toxicity. Furthermore, BDCPP was also detected in the brain and somore research is warranted concerning the toxic effects of TDCPP onthe central nervous system. It should be mentioned that chemicalanalysis showed lower ratio of TDCPP: BDCPP in larval stages, indi-cating that larvae may have high biotransformation potential. Onthe other hand, higher body burdens of TDCPP and BDCPP in larvaecould also be due to larval fish having a lower capacity to excretethese chemicals. Indeed, a previous study with PBDE bioaccumula-tion in developing mice showed that young animals have a reducedability to excrete PBDEs during development (Staskal et al., 2006).Hence, it is also necessary to investigate whether TDCPP preferen-
tially distributes to lipophilic tissues, as larval stages possess higherlipid contents. Additional studies regarding the bioaccumulationkinetics of TDCPP in developing fish and also potential hazardouseffects to early developmental stages are needed.g/L) for 6 months. The data represent the mean ± SEM of three replicates (3 brains0.05; ** P < 0.01 indicates a significant difference between the exposure groups and
Male
0 4 20 100
0.20 1.04 ± 0.17 1.14 ± 0.12 1.16 ± 0.01 0.95 ± 0.14 0.05* 1.02 ± 0.10 1.22 ± 0.10 0.86 ± 0.15 0.99 ± 0.13 0.11* 1.01 ± 0.09 1.31 ± 0.21 1.06 ± 0.22 1.27 ± 0.22 0.08* 1.01 ± 0.10 0.81 ± 0.08 0.65 ± 0.03* 0.48 ± 0.07**
0.51* 1.01 ± 0.10 1.24 ± 0.21 1.15 ± 0.14 1.89 ± 0.27*

Q. Wang et al. / Aquatic Toxicology 158 (2015) 108–115 113
Table 2Actual TDCPP concentrations in exposure solutions at embryo (2-dpf), larval (42-dpf) and adult (120-dpf) stages. The data (average before and after water changes) areexpressed as mean ± standard error (SEM) of 3 replicate beakers (tanks). LOD: limit of detection.
Nominal TDCPP (�g/L) 0 4 20 100
Measuredconcentrations (�g/L)
Embryo <LOD
Larval <LOD
Adult <LOD
Fig. 3. TDCPP and BDCPP contents in zebrafish and brain tissue after exposureto TDCPP. (A) TDCPP and BDCPP contents in the 5-dpf zebrafish larvae; (B) TDCPPand BDCPP contents in adult male zebrafish; (C) TDCPP and BDCPP contents in adultfemale zebrafish; for adult fish, the values represent the mean of three individualreplicate fish. For larvae, TDCPP and BDCPP were measured in 50 larvae, with threereplicate samples. The data are expressed as mean ± SEM.
5.2 ± 0.6 23.6 ± 0.8 113.0 ± 2.05.2 ± 0.3 24.2 ± 2.2 98.7 ± 6.94.6 ± 0.1 22.8 ± 0.9 102.5 ± 2.6
We observed higher contents of TDCPP in females than inmales, indicating gender-dependent bioconcentration of TDCPP,but the mechanisms underlying this differential bioconcentrationremain unknown. However, gender-dependent bioconcentrationof organic toxicants has often been observed in fish species. Forexample, long-term exposure to low concentrations of PBDEsresulted in higher body burden in male zebrafish (Yu et al.,2011; Chen et al., 2012). This phenomenon was also observedin wild marine dolphins (Stenella attenuate) where males bioac-cumulated significantly higher levels of PBDEs (Ko et al., 2014).In fish species, higher bioaccumulation of other toxicants (e.g.,metals like mercury) is often observed in females (Burger et al.,2007). Hence, gender-specific bioconcentration of different typesof toxicants is common, and this could be due to differences inuptake, metabolism, distribution and elimination, as well as genderdifferences in nutritional needs, feeding behavior and various phys-iological activities influencing contaminant accumulation (Burgeret al., 2007). Furthermore, in vivo gender-specific receptors can alsocontribute to gender differences in the bioaccumulation of pollut-ants (Monteverdi and Di Giulio, 2000). Hence, the gender-specificbioconcentration of toxicants could cause gender-specific sensitiv-ity or toxicity (Burger et al., 2007; Deane et al., 2014). Given thatTDCPP is relatively hydrophobic (log Kow = 3.76), it is possible thatthe differences in uptake between males and females were drivenby differences in the lipid content. In the present study, we observedthat endpoints indicative of neurotoxicity were more sensitive infemales than males, which could be partly explained by gender-specific sensitivity or toxicity.
In our present study, neurotransmitter levels were unchangedin larvae, which may indicate that the developing nervous sys-tem is not a target during acute exposure to low concentrationsof TDCPP. It is well-known that the mechanism of OP toxicity is viainhibition of AChE activity, but we did not observe any effect onAChE activity in larvae and in the adult fish brain. This observationsuggests that, unlike OPs, TDCPP is not a typical neuron toxin. Inaddition to affecting neurotransmitters, we hypothesize that themechanism by which TDCPP causes neurotoxicity is by modifyingproteins in the brain. Thus, we examined several genes and pro-teins that are expressed in the central nervous system during earlydevelopmental stages of the zebrafish and in adult fish brains.
We found that the expression of the selected genes associatedwith the central nervous system remained unchanged in the lar-vae (except for gap-43), but there was significant downregulationof these key genes (e.g., ˛1-tubulin, mbp, SYN2a) and proteins inadult fish. The ˛1-tubulin gene encodes an intermediate filamentprotein that forms an essential part of microtubules (Baas, 1997).Thus, if the transcriptional effect is also seen at protein level, alter-ations to �1-tubulin proteins or their levels would likely affectthe structure and function of microtubules, which may have dra-matic effects on brain architecture and function. It should also bementioned that downregulation of the ˛1-tubulin gene expression(both at mRNA and protein level) was observed in both males andfemales, suggesting that it is very sensitive to TDCPP stress. Mbp,a major protein constituent of the myelin sheath, is encoded by
mbp, also a biomarker of myelination that is expressed in oligo-dendrocytes in the central and peripheral nervous systems (Leeand Fields, 2009). Perturbation of mbp gene expression can impact
1 oxicol
mmoaguasrdta2eri(gi
vcbn2tt
emnabcbtest
C
A
PXtRrRt
A
i1
R
A
14 Q. Wang et al. / Aquatic T
yelination, which may lead to myelination deficiency and ulti-ately disrupt neuronal functions. We observed downregulation
f mbp gene expression only in females, which further indicates gender-specific effect on myelination in oligodendrocytes. Theap-43 gene transcription was upregulated. gap-43 is frequentlysed as a marker in re-inducing axonal growth for regenerationfter damage (Benowitz and Routtenberg, 1997). It is tempting topeculate that the upregulation of gap-43 represents an adaptiveesponse to maintain overall brain growth necessary to offset theirect effects of toxicants (Alm et al., 2008). Our results are consis-ent with previous reports regarding upregulation of gap-43 eithert transcriptional or protein level in the brain of mouse (Alm et al.,006; Viberg et al., 2008) or in zebrafish (Chen et al., 2012) afterxposure to PBDEs, which is a common response and most likelyeflects a compensatory mechanism. SYN2a plays an important rolen both synaptogenesis and neurotransmitter release in mammalsKao et al., 1998). We observed a small downregulation of SYN2aene transcription in females exposed to 100 �g/L TDCPP, suggest-ng that it is not sensitive to TDCPP stress.
Our results show that the locomotion was unchanged in the lar-ae by exposure to TDCPP. Alterations of locomotion can result fromhanges in gene expression in the central nervous system and/ory direct neurotoxic effects on neuronal cells, including changingeurotransmitters and AChE activity (Chen et al., 2012; Irons et al.,013). In our present study, we did not observe any alterations inhese parameters or locomotion, suggesting no acute developmen-al neurotoxicity to fish embryos and larvae.
In summary, the present report demonstrates that long-termxposure to TDCPP can result in neurotoxicity in fish. However, theechanisms underlying alterations of gene expression in central
ervous system, as well as how TDCPP affects neurotransmitters,re unknown. As TDCPP can bioaccumulate in the brain, it shoulde investigated how TDCPP can affect cell signaling pathways in theentral nervous system. Current knowledge on the environmentalehavior and toxicity of OPFRs is limited despite increasing produc-ion and application of these chemicals, which are now commonnvironmental contaminants. Therefore, it is important to under-tand the environmental risks of OPFRs to aquatic ecosystems andheir potential human health effects.
ompeting interest
The authors declare no competing financial interests.
cknowledgments
This work was supported by the Strategic Priority Researchrogram of the Chinese Academy of Sciences (Grant no.DB14040103), the National Natural Science Foundation of China
o BZ (21237005), PL (41276111), JL (41206080), and the Hong Kongesearch Grants Council (CityU 160613) and the National Natu-al Science Foundation of China-Research Grants Committee Jointesearch Scheme (N CityU 127/12). The authors sincerely thank thewo anonymous reviewers for their constructive comments.
ppendix A. Supplementary data
Supplementary data associated with this article can be found,n the online version, at http://dx.doi.org/10.1016/j.aquatox.2014.1.001.
eferences
lm, H., Scholz, B., Fischer, C., Kultima, K., Viberg, H., Eriksson, P., Dencker, L.,Stigson, M., 2006. Proteomic evaluation of neonatal exposure to 2,2,4,4,5-pentabromodiphenyl ether. Environ. Health Perspect. 114, 254–259.
ogy 158 (2015) 108–115
Alm, H., Kultima, K., Scholz, B., Nilsson, A., Andrén, P.E., Fex-Svenningsen, Å., Dencker,L., Stigson, M., 2008. Exposure to brominated flame retardant PBDE-99 affectscytoskeletal protein expression in the neonatal mouse cerebral cortex. Neuro-toxicology 29, 628–637.
Baas, P.W., 1997. Microtubules and axonal growth. Curr. Opin. Cell Biol. 9, 29–36.Benowitz, L.I., Routtenberg, A., 1997. GAP-43: an intrinsic determinant of neuronal
development and plasticity. Trends Neurosci. 20, 84–91.Brosamle, C., Halpern, M.E., 2002. Characterization of myelination in the developing
zebrafish. Glia 39, 47–57.Buckley, C.E., Marguerie, A., Alderton, W.K., Franklin, R.J., 2010. Temporal dynamics
of myelination in the zebrafish spinal cord. Glia 58, 802–812.Burger, J., Fossi, C., McClellan-Green, P., Orlando, E.F., 2007. Methodologies, bioindi-
cators, and biomarkers for assessing gender-related differences in wildlifeexposed to environmental chemicals. Environ. Res. 104, 135–152.
Cao, S., Zeng, X., Song, H., Li, H., Yu, Z., Sheng, G., Fu, J., 2012. Levels and distributionsof organophosphate flame retardants and plasticizers in sediment from TaihuLake, China. Environ. Toxicol. Chem. 31, 1478–1484.
Carignan, C.C., McClean, M.D., Cooper, E.M., Watkins, D.J., Fraser, A.J., Heiger-Bernays,W., Stapleton, H.M., Webster, T.F., 2013. Predictors of tris(1,3-dichloro-2-propyl)phosphate metabolite in the urine of office workers. Environ. Int. 55, 56–61.
Chen, L., Yu, K., Huang, C., Yu, L., Zhu, B., Lam, P.K.S., Lam, J.C.W., Zhou, B., 2012. Prena-tal transfer of polybrominated diphenyl Ethers (PBDEs) results in developmentalneurotoxicity in zebrafish larvae. Environ. Sci. Technol. 46, 9727–9734.
Cooper, E.M., Covaci, A., van Nuijs, A.L.N., Webster, T.F., Stapleton, H.M.,2011. Analysis of the flame retardant metabolites bis(1,3-dichloro-2-propyl)phosphate (BDCPP) and diphenyl phosphate (DPP) in urine using liq-uid chromatography–tandem mass spectrometry. Anal. Bioanal. Chem. 401,2123–2132.
Cristale, J., Katsoyiannis, A., Sweetman, A.J., Jones, K.C., Lacorte, S., 2013. Occurrenceand risk assessment of organophosphorus and brominated flame retardants inthe River Aire (UK). Environ. Pollut. 179, 194–200.
Crump, D., Chiu, S., Kennedy, S.W., 2012. Effects of tris(1,3-dichloro-2-propyl) phos-phate and tris(1-chloropropyl) phosphate on cytotoxicity and mRNA expressionin primary cultures of avian hepatocytes and neuronal cells. Toxicol. Sci. 126,140–148.
Deane, E.E., van de Merwe, J.P., Hui, J.H.L., Wu, R.S.S., Woo, N.Y.S., 2014. PBDE-47exposure causes gender specific effects on apoptosis and heat shock proteinexpression in marine medaka, Oryzias melastigma. Aquat. Toxicol. 147, 57–67.
Dishaw, L.V., Powers, C.M., Ryde, I.T., Roberts, S.C., Seidler, F.J., Slotkin, T.A., Stapleton,H.M., 2011. Is the PentaBDE replacement, tris(1,3-dichloro-2-propyl) phosphate(TDCPP), a developmental neurotoxicant? Studies in PC12 cells. Toxicol. Appl.Pharmacol. 256, 281–289.
Fan, C.Y., Cowden, J., Simmons, S.O., Padilla, S., Ramabhadran, R., 2010. Geneexpression changes in developing zebrafish as potential markers for rapid devel-opmental neurotoxicity screening. Neurotoxicol. Teratol. 32, 91–98.
Farhat, A., Buick, J.K., Williams, A., Yauk, C.L., O’Brien, J.M., Crump, D., Williams,K.L., Chiu, S., Kennedy, S.W., 2014. Tris(1,3-dichloro-2-propyl) phosphateperturbs the expression of genes involved in immune response and lipidand steroid metabolism in chicken embryos. Toxicol. Appl. Pharmacol. 275,104–112.
Farhat, A., Crump, D., Chiu, S., Williams, K.L., Letcher, R.J., Gauthier, L.T., Kennedy,S.W., 2013. In ovo effects of two organophosphate flame retardants-TCPP andTDCPP-on pipping success, development, mRNA expression, and thyroid hor-mone levels in chicken embryos. Toxicol. Sci. 134, 92–102.
Fu, J., Han, J., Zhou, B., Gong, Z., Santos, E.M., Huo, X., Zheng, W., Liu, H., Yu, H.,Liu, C., 2013. Toxicogenomic responses of zebrafish embryos/larvae to tris(1,3-dichloro-2-propyl) phosphate (TDCPP) reveal possible molecular mechanismsof developmental toxicity. Environ. Sci. Technol. 47, 10574–10582.
Irons, T.D., Kelly, P.E., Hunter, D.L., MacPhail, R.C., Padilla, S., 2013. Acute adminis-tration of dopaminergic drugs has differential effects on locomotion in larvalzebrafish. Pharmacol. Biochem. Behav. 103, 792–813.
Kao, H.T., Porton, B., Czernik, A.J., Feng, J., Yiu, G., Haring, M., Benfenati, F., Greengard,P., 1998. A third member of the synapsin gene family. Proc. Natl. Acad. Sci. U.S.A.95, 4667–4672.
Kim, J.W., Isobe, T., Chang, K.H., Amano, A., Maneja, R.H., Zamora, P.B., Siringane, F.P.,Tanabe, S., 2011. Levels and distribution of organophosphorus flame retardantsand plasticizers in fishes from Manila Bay, the Philippines. Environ. Pollut. 159,3653–3659.
Ko, F.C., We, N.Y., Chou, L.S., 2014. Bioaccumulation of persistent organic pollut-ants in stranded cetaceans from Taiwan coastal waters. J. Hazard. Mater. 277,127–133.
Kojima, H., Takeuchi, S., Itoh, T., Iida, M., Kobayashi, S., Yoshida, T., 2013. In vitroendocrine disruption potential of organophosphate flame retardants via humannuclear receptors. Toxicology 314, 76–83.
Lee, P.R., Fields, R.D., 2009. Regulation of myelin genes implicated in psychiatricdisorders by functional activity in axons. Front. Neuroanat. 3, 4.
Li, J., Yu, N., Zhang, B., Jin, L., Li, M., Hu, M., Zhang, X., Wei, S., Yu, H., 2014. Occurrenceof organophosphate flame retardants in drinking water from China. Water Res.54, 53–61.
Liu, C., Wang, Q., Liang, K., Liu, J., Zhou, B., Zhang, X., Liu, H., Giesy, J.P., Yu, H.,2013a. Effects of tris(1,3-dichloro-2-propyl) phosphate and triphenyl phosphate
on receptor-associated mRNA expression in zebrafish embryos/larvae. Aquat.Toxicol. 128, 147–157.Liu, X., Ji, K., Choi, K., 2012. Endocrine disruption potentials of organophosphateflame retardants and related mechanisms in H295R and MVLN cell lines and inzebrafish. Aquat. Toxicol. 114–115, 173–181.

oxicol
L
M
M
M
M
M
M
O
P
R
R
R
R
R
S
Q. Wang et al. / Aquatic T
iu, X., Ji, K., Jo, A., Moon, H.B., Choi, K., 2013b. Effects of TDCPP or TPP on gene tran-scriptions and hormones of HPG axis, and their consequences on reproductionin adult zebrafish (Danio rerio). Aquat. Toxicol. 134, 104–111.
arklund, A., Andersson, B., Haglund, P., 2005. Organophosphorus flame retardantsand plasticizers in Swedish sewage treatment plants. Environ. Sci. Technol. 39,7423–7429.
artinez-Carballo, E., Gonzalez-Barreiro, C., Sitka, A., Scharf, S., Gans, O., 2007. Deter-mination of selected organophosphate esters in the aquatic environment ofAustria. Sci. Total Environ. 388, 290–299.
a, Y., Cui, K., Zeng, F., Wen, J., Liu, H., Zhu, F., Ouyang, G., Luan, T., Zeng, Z., 2013.Microwave-assisted extraction combined with gel permeation chromatographyand silica gel cleanup followed by gas chromatography-mass spectrometry forthe determination of organophosphorus flame retardants and plasticizers inbiological samples. Anal. Chim. Acta 786, 47–53.
cgee, S.P., Cooper, E.M., Stapleton, H.M., Volz, D.C., 2012. Early zebrafish embryo-genesis is susceptible to developmental TDCPP exposure. Environ. HealthPerspect. 120, 1585–1591.
eeker, J.D., Stapleton, H.M., 2009. House dust concentrations of organophosphateflame retardants in relation to hormone levels and semen quality parameters.Environ. Health Perspect. 118, 318–323.
onteverdi, G.H., Di Giulio, R.T., 2000. In vitro and in vivo association of 2,3,7,8-tetrachlorodibenzo-p-dioxin and benzo[a]pyrene with the yolk-precursorprotein vitellogenin. Environ. Toxicol. Chem. 19, 2502–2511.
ffice of Environmental Health Hazard Assessment’s (OEHHA), 2011. Evidenceon the Carcinogenicity of Tris(1,3-Dichloro-2-Propyl) Phosphate. Office ofEnvironmental Health Hazard Assessment’s (OEHHA) http://oehha.ca.gov/prop65/hazard ident/pdf zip/TDCPP070811.pdf
ayne, J.F., Mathieu, A., Melvin, W., Fancey, L.L., 1996. Acetylcholinesterase,an old biomarker with a new future? Field trials in association with twourban rivers and a paper mill in Newfoundland. Mar. Pollut. Bull. 32,225–231.
ao, J.V., Begum, G., Pallela, R., Usman, P.K., Rao, R.N., 2005. Changes in behavior andbrain acetylcholinesterase activity in mosquito fish, Gambusia affinis in responseto the sub-lethal exposure to chlorpyrifos. Int. J. Environ. Res. Public Health 2,478–483.
eemtsma, T., Quintana, J.B., Rodil, R., Garcia-Lopez, M., Rodriguez, I., 2008.Organophosphorus flame retardants and plasticizers in water and air I. Occur-rence and fate. Trends Anal. Chem. 27, 727–737.
egnery, J., Püttmann, W., 2010. Occurrence and fate of organophosphorus flameretardants and plasticizers in urban and remote surface waters in Germany.Water Res. 44, 4097–4104.
ice, D., Barone, S.J., 2000. Critical periods of vulnerability for the developing nervoussystem: evidence from humans and animal models. Environ. Health Perspect.108 (Suppl. 3), 511–533.
ico, E.P., Rosemberg, D.B., Seibt, K.J., Capiotti, K.M., Da Silva, R.S., Bonan, C.D., 2011.Zebrafish neurotransmitter systems as potential pharmacological and toxico-
logical targets. Neurotoxicol. Teratol. 33, 608–617.ari, Y., Hammad, L.A., Saleh, M.M., Rebec, G.V., Mechref, Y., 2010. Alteration ofselective neurotransmitters in fetal brains of prenatally alcohol-treated C57BL/6mice: quantitative analysis using liquid chromatography/tandem mass spec-trometry. Int. J. Dev. Neurosci. 28, 263–269.
ogy 158 (2015) 108–115 115
Stapleton, H.M., Klosterhaus, S., Eagle, S., Fuh, J., Meeker, J.D., Blum, A., Webster, T.F.,2009. Detection of organophosphate flame retardants in furniture foam and U.S.house dust. Environ. Sci. Technol. 43, 7490–7495.
Stapleton, H.M., Klosterhaus, S., Keller, A., Ferguson, P.L., van Bergen, S., Cooper, E.,Webster, T.F., Blum, A., 2011. Identification of flame retardants in polyurethanefoam collected from baby products. Environ. Sci. Technol. 45, 5323–5331.
Stapleton, H.M., Sharma, S., Getzinger, G., Ferguson, P.L., Gabriel, M., Webster, T.F.,Blum, A., 2012. Novel and high volume use flame retardants in US couches reflec-tive of the 2005 pentaBDE phase out. Environ. Sci. Technol. 46, 13432–13439.
Staskal, D.F., Diliberto, J.J., Birnbaum, L.S., 2006. Disposition of BDE 47 in developingmice. Toxicol. Sci. 90, 309–316.
Sundkvist, A.M., Olofsson, U., Haglund, P., 2010. Organophosphorus flame retardantsand plasticizers in marine and fresh water biota and in human milk. J. Environ.Monit. 12, 943–951.
Ta, N., Li, C., Fang, Y., Liu, H., Lin, B., Jin, H., Tian, L., Zhang, H., Zhang, W., Xi, Z., 2014.Toxicity of TDCPP and TCEP on PC12 cell: Changes in CAMKII. GAP43, tubulinand NF-H gene and protein levels. Toxicol. Lett. 227, 164–171.
Thatcher, E.J., Paydar, I., Anderson, K.K., Patton, J.G., 2008. Regulation of zebrafish finregeneration by microRNAs. Proc. Natl. Acad. Sci. U.S.A. 105, 18384–18389.
Udvadia, A.J., Koster, R.W., Skene, J.H., 2001. GAP-43 promoter elements in trans-genic zebrafish reveal a difference in signals for axon growth during CNSdevelopment and regeneration. Development 128, 1175–1182.
Van der Veen, I., de Boer, J., 2012. Phosphorus flame retardants: Properties, pro-duction, environmental occurrence, toxicity and analysis. Chemosphere 88,1119–1153.
Viberg, H., Mundy, W., Eriksson, P., 2008. Neonatal exposure to decabrominateddiphenyl ether (PBDE 209) results in changes in BDNF, CaMKII and GAP-43,biochemical substrates of neuronal survival, growth, and synaptogenesis. Neu-rotoxicology 29, 152–159.
Wang, H.Y., Chung-Davidson, Y.W., Li, K., Scott, A.M., Li, W.M., 2012. Quantificationof monoamine neurotransmitters and melatonin in sea lamprey brain tissues byhigh performance liquid chromatography-electrospray ionization tandem massspectrometry. Talanta 89, 383–390.
Wang, Q., Liang, K., Liu, J., Yang, L., Guo, Y., Liu, C., Zhou, B., 2013. Exposure of zebrafishembryos/larvae to TDCPP alters concentrations of thyroid hormones and tran-scriptions of genes involved in the hypothalamic-pituitary-thyroid axis. Aquat.Toxicol. 126, 207–213.
Wang, X., Liu, J., Yin, Y., 2011. Development of an ultra-high-performance liq-uid chromatography-tandem mass spectrometry method for high throughputdetermination of organophosphorus flame retardants in environmental water.J. Chromatogr. A 1218, 6705–6711.
Yang, F., Ding, J., Huang, W., Xie, W., Liu, W., 2014. Particle size-specific distributionsand preliminary exposure assessments of organophosphate flame retardants inoffice air particulate matter. Environ. Sci. Technol. 48, 63–70.
Yu, L., Lam, J.C.W., Guo, Y., Wu, R.S.S., Lam, P.K.S., Zhou, B., 2011. Parental transferof polybrominated diphenyl ethers (PBDEs) and thyroid endocrine disruption in
zebrafish. Environ. Sci. Technol. 45, 10652–10659.Zeng, X., He, L., Cao, S., Ma, S., Yu, Z., Gui, H., Sheng, G., Fu, J., 2014. Occurrenceand distribution of organophosphate flame retardants/plasticizers in wastewa-ter treatment plant sludges from the Pearl River Delta, China. Environ. Toxicol.Chem. 33, 1720–1725.














![Synthesis, Spectroscopic and Electrochemical Studies of Isomeric Dichloro-bis-[ N (1)-Alkyl-2-(Arylazo)Imidazole]-Osmium(II). Single Crystal X-ray Structures of Blue-Violet Dichloro-Bis-[](https://static.fdokumen.com/doc/165x107/6333de5ece61be0ae50edc8a/synthesis-spectroscopic-and-electrochemical-studies-of-isomeric-dichloro-bis-.jpg)





