
Membrane bioreactor wastewater treatment plants reveal diverse yeast and protist communities of...
12
Membrane bioreactor wastewater treatment plants reveal diverse yeast and protist communities of potential significance in biofouling Raquel Liébana, Lucía Arregui*, Ignacio Belda, Luis Gamella, Antonio Santos, Domingo Marquina and Susana Serrano Department of Microbiology III, Complutense University, Madrid, Spain (Received 15 July 2014; accepted 7 December 2014) The yeast community was studied in a municipal full-scale membrane bioreactor wastewater treatment plant (MBR-WWTP). The unexpectedly high diversity of yeasts indicated that the activated sludge formed a suitable environ- ment for them to proliferate, with cellular concentrations of 2.2 ± 0.8 × 10 3 CFU ml −1 . Sixteen species of seven genera were present in the biological reactor, with Ascomycetes being the most prevalent group (93%). Most isolates were able to grow in a synthetic wastewater medium, adhere to polyethylene surfaces, and develop biofilms of variable complexity. The relationship between yeast populations and the protists in the MBR-WWTP was also studied, revealing that some protist species preyed on and ingested yeasts. These results suggest that yeast populations may play a role in the food web of a WWTP and, to some extent, contribute to membrane biofouling in MBR systems. Keywords: wastewater treatment; membrane bioreactor; biofouling; yeast biofilms; protists; predation Introduction Although microorganisms are considered to be directly responsible for wastewater purification in activated sludge treatment processes, the complexity of microbio- logical communities is often underestimated during the design of wastewater treatment plants (WWTPs). A full understanding of the ecological, physiological, and bio- chemical activities of the microbiota is necessary for optimal operational control (Cabrol & Malhautier 2011; Nielsen et al. 2012). Bacterial populations are mainly responsible for degrading organic matter in the wastewater (Ju et al. 2014), while protists and micro- metazoa are the main consumers in food chains (Madoni et al. 1993). However, the importance of other microbial communities, such as yeast or filamentous fungi, has either not been considered or has been underestimated (Watanabe et al. 2009; Yang et al. 2011, 2013; Basak et al. 2014). Previous studies have shown that yeast populations could help to treat different types of sewage through the production of hydrolytic enzymes such as lipases or pro- teases (Jarboui et al. 2012, 2013; Yang et al. 2013). However, doubts have been raised about whether such yeasts could survive in an activated sludge system in a WWTP for a long time. Yeast populations in wastewater and their possible ecological role in WWTPs have received little attention (Lopandic et al. 2013). The few studies in this field have considered the presence of mycotic communities in conventional WWTPs as poten- tial pathogens, focusing mainly on filamentous fungi, with yeast populations not being adequately studied (Awad & Kraume 2010, 2011). Furthermore, none of these studies have considered the presence of yeasts in membrane bioreactor (MBR)-WWTP systems. In MBR systems, liquid/solid separation is performed by filtration through a membrane (Wu & Lee 2011). The drawback of this system is fouling of the membranes, in particular biofouling, where a biofilm develops on the membrane surface. This phenomenon decreases the over- all efficiency of the process, ultimately leading to an increase in operating costs (Meng et al. 2009; Feng et al. 2011; Sweity et al. 2011; Oh et al. 2012). Since micro- bial biofilm development has been recognized as being involved in membrane biofouling, the structure and composition of the microbiota and the relationships and interactions between its members have been subject to in-depth investigation in MBR systems. Most of these investigations have focused on bacterial biofilms, with less information so far on the contribution by other microbial communities such as yeasts or protists, as either single-species biofilms or as part of multi-species complex biofilms. Yeast cells can quickly adapt their adhesion proper- ties to new environments, which is of considerable eco- nomic importance for food-processing companies, because adherent yeast can form highly resistant biofilms in industrial installations. Adherence is a complex pro- cess, mediated in yeast by cell wall-bound adhesins. These proteins have sugars or amino acid residues that permit adherence to biotic and abiotic surfaces, and thus *Corresponding author. Email: [email protected] © 2015 Taylor & Francis Biofouling, 2015 Vol. 31, No. 1, 71–82, http://dx.doi.org/10.1080/08927014.2014.998206 Downloaded by [Biblioteca Universidad Complutense de Madrid], [Antonio Santos] at 01:25 17 January 2015
Transcript of Membrane bioreactor wastewater treatment plants reveal diverse yeast and protist communities of...

Membrane bioreactor wastewater treatment plants reveal diverse yeast and protistcommunities of potential significance in biofouling
Raquel Liébana, Lucía Arregui*, Ignacio Belda, Luis Gamella, Antonio Santos, Domingo Marquina and Susana Serrano
Department of Microbiology III, Complutense University, Madrid, Spain
(Received 15 July 2014; accepted 7 December 2014)
The yeast community was studied in a municipal full-scale membrane bioreactor wastewater treatment plant(MBR-WWTP). The unexpectedly high diversity of yeasts indicated that the activated sludge formed a suitable environ-ment for them to proliferate, with cellular concentrations of 2.2 ± 0.8 × 103 CFU ml−1. Sixteen species of seven generawere present in the biological reactor, with Ascomycetes being the most prevalent group (93%). Most isolates were ableto grow in a synthetic wastewater medium, adhere to polyethylene surfaces, and develop biofilms of variable complexity.The relationship between yeast populations and the protists in the MBR-WWTP was also studied, revealing that someprotist species preyed on and ingested yeasts. These results suggest that yeast populations may play a role in the foodweb of a WWTP and, to some extent, contribute to membrane biofouling in MBR systems.
Keywords: wastewater treatment; membrane bioreactor; biofouling; yeast biofilms; protists; predation
Introduction
Although microorganisms are considered to be directlyresponsible for wastewater purification in activatedsludge treatment processes, the complexity of microbio-logical communities is often underestimated during thedesign of wastewater treatment plants (WWTPs). A fullunderstanding of the ecological, physiological, and bio-chemical activities of the microbiota is necessary foroptimal operational control (Cabrol & Malhautier 2011;Nielsen et al. 2012). Bacterial populations are mainlyresponsible for degrading organic matter in thewastewater (Ju et al. 2014), while protists and micro-metazoa are the main consumers in food chains (Madoniet al. 1993). However, the importance of other microbialcommunities, such as yeast or filamentous fungi, haseither not been considered or has been underestimated(Watanabe et al. 2009; Yang et al. 2011, 2013; Basaket al. 2014).
Previous studies have shown that yeast populationscould help to treat different types of sewage through theproduction of hydrolytic enzymes such as lipases or pro-teases (Jarboui et al. 2012, 2013; Yang et al. 2013).However, doubts have been raised about whether suchyeasts could survive in an activated sludge system in aWWTP for a long time. Yeast populations in wastewaterand their possible ecological role in WWTPs havereceived little attention (Lopandic et al. 2013). The fewstudies in this field have considered the presence ofmycotic communities in conventional WWTPs as poten-tial pathogens, focusing mainly on filamentous fungi,
with yeast populations not being adequately studied(Awad & Kraume 2010, 2011). Furthermore, none ofthese studies have considered the presence of yeasts inmembrane bioreactor (MBR)-WWTP systems.
In MBR systems, liquid/solid separation is performedby filtration through a membrane (Wu & Lee 2011). Thedrawback of this system is fouling of the membranes, inparticular biofouling, where a biofilm develops on themembrane surface. This phenomenon decreases the over-all efficiency of the process, ultimately leading to anincrease in operating costs (Meng et al. 2009; Feng et al.2011; Sweity et al. 2011; Oh et al. 2012). Since micro-bial biofilm development has been recognized as beinginvolved in membrane biofouling, the structure andcomposition of the microbiota and the relationships andinteractions between its members have been subject toin-depth investigation in MBR systems. Most of theseinvestigations have focused on bacterial biofilms, withless information so far on the contribution by othermicrobial communities such as yeasts or protists, aseither single-species biofilms or as part of multi-speciescomplex biofilms.
Yeast cells can quickly adapt their adhesion proper-ties to new environments, which is of considerable eco-nomic importance for food-processing companies,because adherent yeast can form highly resistant biofilmsin industrial installations. Adherence is a complex pro-cess, mediated in yeast by cell wall-bound adhesins.These proteins have sugars or amino acid residues thatpermit adherence to biotic and abiotic surfaces, and thus
*Corresponding author. Email: [email protected]
© 2015 Taylor & Francis
Biofouling, 2015Vol. 31, No. 1, 71–82, http://dx.doi.org/10.1080/08927014.2014.998206
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

the expression of these proteins allows yeast to adaptquickly and to form biofilms. The expression of thesegenes is activated by various environmental events, suchas carbon or nitrogen deficiency, pH changes, or changesin ethanol levels, which can all promote a change in theyeast phenotype from non-adherent to adherent as anadaptive response to stress (Verstrepen & Klis 2006). InMBR systems, adhesion and biofilm formation on thesurface of the filtration membrane can severely hinderthe filtration process.
The protist populations described for MBR systemsdiffer from those found in other conventional (Arévaloet al. 2009; Wu et al. 2011) or advanced nutrientremoval systems (Pérez-Uz et al. 2010). These changesin the protist populations are important, since severalbiotic indexes are based on the amount and presence ofprotists and micrometazoa in the reactor and theseindexes are used as bioindication tools to assess theeffectiveness of the treatment process (Madoni 1994).Moreover, protist populations play fundamental ecologi-cal roles both in natural environments (Hülsmann &Galil 2001; Caron et al. 2012) and in artificial ones suchas a WWTP. Their predation capacity has been reportedas having unexpected consequences on microbial biofilmdevelopment (Matz & Kjelleberg 2005; Matz et al.2008).
Given that yeast and protist communities have notbeen studied a great deal in MBR systems, in this paperyeast populations inhabiting the mixed liquor of a full-scale MBR plant and their adhesion and biofilm-develop-ment capacities were studied in an attempt to determinetheir potential impact on MBR biofouling. The role ofyeast in food chains was subsequently assessed by inves-tigating yeast–protist relationships.
Materials and methods
Wastewater treatment plant
The study was carried out in a full-scale MBR-WWTPlocated in Barcelona (Spain). The plant had two differentlines, one of which included a MBR system designed totreat 32,000 m3 day−1 of wastewater, with 58,400 m2 ofactive surface distributed in six parallel trains (seven cas-settes per train, 44 modules per cassette and 31.6 m2 permodule). The domestic and pre-treated industrial waste-waters were biologically treated through an activatedsludge process involving anoxic and aerobic phases forthe elimination of nitrogen. The treated water was regen-erated by ultrafiltration through a membrane module(polyvinylidene difluoride hollow fiber membranes witha 0.04 μm pore size). The system was designed to oper-ate under a permeate flux of 22.8–41.1 l (m2 h)−1 and amaximum transmembrane pressure of –0.55 bar. A flowdiagram of the system is shown in Figure S1.
Sampling
Activated sludge samples were collected in the finalstretch of the aerobic zone in the biological reactor, asindicated in Figure S1, and sent to the laboratory at 2–7°C to ensure the biological stability of the protist andyeast analysis.
Yeast isolation procedures
Aliquots of 1 ml were taken from the samples, and addi-tional 10-fold dilutions were made serially to a final dilu-tion of 10−4. Volumes of 0.1 ml were used for plating onSabouraud culture medium supplemented with chloram-phenicol (0.5 g l−1) to inhibit bacterial growth, and withrose bengal (50 mg l−1) to restrict the growth of filamen-tous fungi. Three plates of each dilution were incubatedat 30°C for 2–5 days for the development and completedifferentiation of yeast colony types. Different macro-scopic characteristics were used for the selection ofstrains (eg colony morphology, aspect, pigmentation androughness). Repeated restreaking was performed on thesame medium to obtain pure cultures. Yeast strains weremaintained on yeast mold agar (YMA) slants containing0.3% (w/v) yeast extract (Difco, Detroit, MI, USA),0.3% (w/v) malt extract (Difco), 0.5% (w/v) proteasepeptone No. 3 (Difco), 1% (w/v) glucose, and 2% (w/v)agar.
Identification of yeast species and phylogenetic analysisof cultured yeasts
Yeast isolates were identified by the sequence analysis ofthe 26S large subunit rRNA gene. Total genomic DNAwas extracted using the isopropanol method (Querol et al.1992), and the DNA for sequencing was amplified usingan Eppendorf Mastercycler apparatus, as described byKurtzman and Robnett (1991) with forward NL-1 primer(5′’-GCA TAT CAA TAA GCG GAG GAA AAG-3’′),and reverse NL-4 primer (5’′-GGT CCG TGT TTC AAGACG G’′). PCRs were conducted in a 50 μl reaction vol-ume in thin-walled 0.2 ml tubes. Briefly, 1 μl of DNAtemplate (100 ng μl−1 quantified with spectrophotometer)was added to a 25 μl master reaction mixture (Ready-MixTM Taq PCR Reaction Mix, Sigma-Aldrich, Madrid,Spain), 1 μl of NL1 and NL4 primers, and 23 μl of steriledistilled water. The thermal cycling parameters were asfollows: initial denaturation at 95°C for 5 min, followedby 30 cycles consisting of denaturation at 95°C for1 min, annealing at 56°C for 1.5 min and extension at72°C for 2 min, followed by a final extension at 72°C for8 min. The PCR-amplified fragments were analyzed byelectrophoresis in 1.2% of agarose in TAE buffer andstaining with gelRed®. The amplification efficiency wasanalyzed under UV light, and images were taken with a
72 R. Liébana et al.
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

digital camera. The amplified D1/D2 fragments werepurified with the UltraCleanTM PCR Clean-Up PCR puri-fication kit (MOBIO Laboratories, Carlsbad, CA, USA),according to the manufacturer′s protocol. The sequencesobtained for identifying the yeasts were analyzed andcompared by BLAST-search (GenBank; www.ncbi.nlm.nih.gov). Partial 26S rRNA gene sequences of all the iso-lates were submitted to GenBank under the followingaccession numbers: KF667506 and KF912805 toKF912866 (Table S1, Figure 1). All the nucleotidesequences were analyzed and manually edited using theBioEdit version 7.0.0 software editor (Ibis Biosciences,Carlsbad, CA, USA). The sequences were aligned usingCLUSTAL W, and pairwise distances were computedwith MEGA version 5.2.2 (Tamura et al. 2011), using theneighbor-joining method. A maximum parsimony analy-sis was performed to construct a phylogenetic tree usingPAUP 4b10 (Swofford 2002). Maximum parsimony treeswere generated using heuristic search methods, and gapswere encoded as missing data. Multiple equal-length par-simony trees were collapsed into 50% majority rule con-
sensus trees. The confidence limits for the phylogenetictree were estimated from bootstrap analyses (1,000 repli-cations) and Microsporidium sp. GU124636.1 as an out-group. Sequences obtained from the following isolates:M3.1 (KF667506), M3.37 (KF912865), M3.41(KF912830), M3.47 (KF912861), M3.52 (KF912831),M3.58 (KF912849), M3.68 (KF912852) and M3.69(KF912853) were not considered because their sequenceswere too short for the phylogenetic analysis (Figure S2).
Detection of yeast killing activity
Each yeast isolate was tested for its ability to kill theselected target strain (Saccharomyces cerevisiae HansenBY4741). The isolates to be tested were inoculated in ±1 cm diameter concentrated zones on plates withYMAMB medium (Llorente et al. 1997) that had beenpreviously covered by a lawn (4 × 105 cells ml−1) of theyeast to be tested for sensitivity, which had been grownin YMA medium and suspended in sterile water shortlybefore inoculation. The plates were incubated for seven
Figure 1. Distribution of yeast species in the full-scale MBR WWTP used for this study. The data obtained show that 16 speciesbelonging to seven genera were found in the biological reactor, with Candida being the most abundant genus (52%), followed bySaccharomyces (18%) and Debaryomyces (16%).
Biofouling 73
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

days at 20°C. Killer strains were surrounded by a clearzone of inhibition with a dark blue boundary.
Yeast growth in a synthetic wastewater medium
Yeast isolates were inoculated in a synthetic wastewatermedium (513.1 mg l−1 peptone, 708.6 mg l−1 tryptone,407.4 mg l−1 NaCl, 44.6 mg l−1 Na2SO4, 44.6 mg l−1
K2HPO4, 3.7 mg l−1 MgCl2·6H2O, 4.5 mg l−1
FeCl2·4H2O, 3.7 mg l−1 CaCl2, 0.06 mg l
−1 MnSO4·H2O,0.037 mg l−1 Na2MoO4·2H2O, 0.008 mg l−1 NaOH,0.056 mg l−1 ZnSO4·H2O, 0.09 mg l−1 CoSO4·7H2O,0.12 mg l−1 CuSO4·5H2O) at a cellular concentration of~ 106 cells ml−1, and incubated at 20°C. Growth wasassessed spectrophotometrically at OD600 nm and deter-mined as positive when an increase in OD600 nm wasobserved after incubation. Those strains with the bestgrowth ability were selected for further study.
Analysis of yeast adherence capabilities
Eighteen yeast strains were cultured in flasks overnight at20°C (150 rpm) in sterile synthetic wastewater, and thendiluted to an OD600 nm of 0.1 in the same medium. Sub-sequently, 100 μl of each diluted culture were pipettedinto each one of eight wells of a sterile non-treated poly-ethylene 96-well microtiter plate (Corning® Costar® cul-ture plates, Sigma-Aldrich). Sterile culture medium onlywas used as a negative control. The plates were preparedin duplicate, and incubated at 20°C, with shaking at100 rpm. The adherence capabilities of the tested strainswere quantified at 24, 48 and 72 h according to the crys-tal violet method described by Merritt et al. (2011).Absorbance was measured at 590 nm at room tempera-ture using an iEMS Reader MF (Labsystems, Helsinki,Finland). In this assay, eight readings were recorded foreach strain and time interval, and repeated twice (inde-pendent studies) using fresh overnight cultures and theresults were averaged. The strains were classified by theiradherence capabilities according to Stepanovic et al.(2000). By applying these criteria, the strains studiedwere classified as non-adherent when Abs > Abscontrol,weakly adherent (Abscontrol < Abs < 2 × Abscontrol), mod-erately adherent (2 × Abscontrol < Abs < 4 × Abscontrol)and strongly adherent (Abs > 4 × Abscontrol). In addition,a mixture of all the tested strains was used by combiningequal volumes of the diluted (OD600 = 0.1) overnightpure cultures. The results obtained in the experiment wereexamined by multifactor analysis of variance followed bythe Kruskal–Wallis test to identify significant differencesbetween strains in terms of their adherence capabilities.Statistical tests were performed using Statgraphics Centu-rion XVI.I (Statpoint Technologies Inc., Warrenton, VA,USA) (Figure 2).
Confocal microscope analysis of yeast biofilms
Seven strongly adherent yeast strains and a moderatelyadherent strain, along with a mixture of all the testedstrains, were selected for the analysis of biofilm develop-ment over a timeframe of 24 to 72 h by using confocallaser scanning microscopy (CLSM). Cells were grown onflat bottom sterile non-treated polyethylene 24-well plates(Thermo Fisher Scientific Inc., Waltham, MA, USA) insynthetic wastewater, as described above. After incuba-tion, the wells were washed with a synthetic wastewatermedium, and the biofilm was stained with Syto9/propidi-um iodide (LIVE/DEAD® FungaLight™ Yeast ViabilityKit, Life Technologies SA, Alcobendas, Madrid, Spain).Three image stacks of each plate were acquired from ran-dom positions using a 20× lens, so a total area of 1.13 ×106 μm2 (1.88 × 105 μm2 per image stack multiplied bysix images stacks) was investigated. Images were taken at2 μm intervals from the surface of the plate bottom to thetop of the biofilm, and the number of images in eachstack varied according to the thickness of the biofilm. Allthe images had a pixel resolution of 1,024 × 1,024, andwere acquired by the same operator to assure a similarvalue for the gray-level pixel background. Images wereprocessed by the program bioImage_L v.2.1 (Chávez dePaz 2009). The output data from the bioImage analysiscontain the following information: total field area (μm2),percentage of the total field area covered by cells, totalcellular biomass (μm2), cellular biomass (μm2) ofcells stained fluorescent green, percentage of the totalcells stained fluorescent green, cellular biomass (μm2) ofcells stained fluorescent red, percentage of the total cellsstained fluorescent red, and 3-D reconstructions of eachstack with horizontal and perpendicular views (Table 1).
Characterization of protists and micrometazoacommunities
Microscope analysis was performed within 24 h aftersampling. Two 25 μl replicates were used to directlycount the protists under a phase contrast microscope(Olympus BX50, Olympus Iberia SAU, Barcelona,Spain). Photomicrographs of living or stained specimens(silver carbonate impregnation, Fernández-Galiano 1976)were also collected using differential interference contrast(DIC) microscopy and bright field microscopy (NikonEclipse 80i with a Nikon Digital Sight camera, Izasa,Barcelona, Spain). Naked amoebae were assigned tothree classes, as recommended by Madoni (1991) forstudies in WWTP. The identification was performedaccording to the criteria proposed by Lee et al. (2000)and Serrano et al. (2008). The terminology on protistecological groups was also used for comparison purposes(Madoni 1991, 1994).
74 R. Liébana et al.
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

Protist isolation
Euplotes sp. and Paramecium sp. were isolated fromthe activated sludge of an MBR and an A2O (anaero-bic-anoxic-aerobic phases) WWTP, respectively, bymicromanipulation using a sterile tissue-treated polyeth-ylene 24-well microtiter plate (TPP®, Ibian Technolo-gies) in a wheat medium. Both cultures were fedweekly with 2 μl of an early stationary phase cultureof Escherichia coli CECT 515 in TSB (PanreacQuímica SA, Barcelona, Spain). Once isolated andcloned in microtiter plates, the protists were maintainedin a growth chamber at 18°C, being subsequentlytransferred to sterile cell culture polyethylene tubes(Greiner Bio-One Francisco Soria Melguizo SA,Madrid, Spain).
Protist predation on yeast cells
A study was conducted on the ability of the protists toingest yeasts, using cell overnight cultures of Candidapalmioleophila M3-28 and S. cerevisiae M3-38 in YMBmedium at 30°C. The yeasts (106 cells ml−1) were washedtwice by centrifugation at 7,000 rpm and resuspended inmineral water (prepared by sequential filtration throughGF/F and 0.2 μm polycarbonate filters and autoclaved).Twenty-five protist cells were then directly captured bymicromanipulation and deposited in 2 ml of the previouslyprepared yeast cell suspensions. The protists and yeastswere maintained at 18°C for 24–48 h in sterile Petri dishes.Finally, the protists were captured by micromanipulationand directly observed by DIC microscopy (Nikon Eclipse80i microscope with a Nikon Digital Sight camera).
Figure 2. Adherence capabilities at 24 h (white bars), 48 h (gray bars) and 72 h (black bars) of yeast isolates determined by thecrystal violet assay method. Dashed lines indicate levels of adherence used to classify yeasts as weakly adherent, moderately adherent,and strongly adherent (Stepanovic et al. 2000).
Table 1. Analysis of biofilms developed in a synthetic wastewater medium by eight strongly adherent yeast isolates.
Yeaststrains
Substratumarea (μm2)
Cell areacoverage (μm2)
Coverage(%)
Totalbiovolume
(μm3)Viable cell
biovolume (%)Dead cell
biovolume (%)Not segmentedbiovolume(%)
M3-8 1.13 × 106 6.10 × 105 54.20 8.99 × 107 99.12 0.52 0.35M3-12 1.13 × 106 4.44 × 105 39.39 1.09 × 107 99.76 0.11 0.12M3-16 1.13 × 106 7.56 × 104 6.71 1.06 × 107 99.78 0.03 0.17M3-18 1.13 × 106 7.48 × 105 66.47 7.04 × 107 99.06 0.83 0.09M3-21 1.13 × 106 7.35 × 105 65.26 7.23 × 107 99.70 0.17 0.12M3-28 1.13 × 106 6.55 × 105 58.17 6.27 × 107 99.85 0.12 0.02M3-31 1.13 × 106 5.95 × 105 52.82 9.29 × 107 95.85 3.78 0.36M3-42 1.13 × 106 8.92 × 105 79.25 1.34 × 108 98.84 0.74 0.41aMix 1.13 × 106 4.71 × 105 41.85 4.89 × 107 97.22 2.73 0.04
aMixture of all the tested yeast strains.
Biofouling 75
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

Results
Identification and characterization of yeast isolates andphylogenetic analysis of cultured yeasts
A total of 67 strains of yeasts were isolated from themixed liquor, with cellular densities ranging from 1.4 to3 × 103 CFU ml−1. Ascomycetes were the major popula-tion, with Basidiomycetes making up only 7% of thepopulation. The results indicated the presence of 16 dif-ferent species of yeasts, belonging to a total of sevengenera, with a predominance of Candida sorbophila,Debaryomyces hansenii and S. cerevisiae, which jointlyaccounted for 50% of the total of 67 strains identified(Figures 1 and S2). The ratios of cultivable yeast onSabouraud-chloramphenicol-bengal rose to total cultiva-ble bacteria (data not shown) ranged from 1:1.65 × 103
to 1:0.53 × 106, showing that yeasts are a significantmember of the microbial population in a MBR-WWTP.
A detailed analysis of the most relevant physiologicalcharacteristics showed that all the yeast isolates wereable to grow in synthetic wastewater, and their carbohy-drate metabolism enabled them to grow in the aerobicconditions present in the reactor. Moreover, 83% of thestrains were able to form biofilm on the surface of theculture medium (veil) and/or on solid surfaces in micro-titer plates. The determination of the killer activity of theyeast isolates against the sensitive control strain S. cere-visiae BY4741 showed that the killer characteristic waspresent in 18% of the yeast isolates (Table S1).
Yeast adherence capabilities
Based on a species diversity criterion, 18 strains wereselected to test their adherence capabilities and subse-quent surface biofilm development in a synthetic waste-water medium. The isolates M3-8, M3-28 and M3-42,three C. palmioleophila strains, recorded the highestabsorbance values (Figure 2). The multifactor analysis ofvariance confirmed the significant differences betweenspecies in their adherence capabilities (p < 0.05), with ap value of 0 when the medians were compared in theKruskal–Wallis test for each time interval studied.
CLSM analysis of yeast biofilms
Average biofilm thickness values were obtained directlyfrom data recorded in the CLSM sessions, where mea-surements were made of the distance from the well sur-face to the highest point containing biomass in eachimage stack (Figure 3). The observed values rangedbetween 17.35 and 17.68 μm for the 72 h Candidapalmioleophila biofilms (strains M3-8 and M3-42) and39.70 μm for the 24 h Candida parapsilosis M3-12 bio-films. Except for the Candida davisiana M3-31 biofilms,all the other strains recorded the highest thickness values
at 24 h of experimentation. Accordingly, and also consid-ering the earlier period of yeast surface colonization asthe most promising for preventing MBR membrane bio-fouling, subsequent experiments focused on 24 h yeastbiofilms.
A visual inspection of individual CLSM image stacksindicated that the patterns the biofilms formed were dif-ferent. The results calculated for several variables by thebioImage_L software, chosen to characterize yeast bio-films [substratum area (μm2), cell area coverage (μm2),coverage (%), total biovolume (μm3), viable cell biovo-lume (%), dead cell biovolume (%) and not segmentedbiovolume (%)], confirmed these observations (Table 1).Differences between strains in their ability to colonizethe polystyrene surface (total area of 1.13 × 106 μm2)were observed, with C. palmioleophila M3-42 colonizing79.25% of the surface, followed by Rhodotorula muci-laginosa M3-18 and Yarrowia lipolytica M3-21, whichboth recorded values close to 65%. The three aforemen-tioned strains were uniformly distributed over the surface(Figure 3A, B and C); by contrast, M3-16 D. hanseniicolonized only 6.75% of the surface, showing a tendencyto form clusters (Figure 3D). Different cellular morphol-ogies were observed in the biofilms developed byC. palmioleophila M3-28 and C. davisiana M3-31, withthe presence of both individual cells and pseudo-myceliaformation (Figure 3E). Multi-specific biofilms (Figure 3F)were formed by all of the highly adherent strains, and allrecorded high cell mortality, indicating the presence ofinterspecific relationships such as competitive factors likethe killer phenotype shown by D. hansenii M3-16.
The highest values recorded for biofilm biovolume(μm3 covered by the total biofilm population, and theresult of multiplying the total added biomass by theoverall z-level) were for C. palmioleophila M3-42(1.34 × 108 μm3) and C. parapsilosis M3-12 (1.09 ×108 μm3). The former recorded the highest value for totaladded biomass and the latter the highest biofilm thick-ness (Table 1).
A common feature for all the 24 h yeast biofilmsinvestigated was that > 95% of the population remainedviable (green fluorescent subpopulation), and thereforemetabolically active. The vertical distribution of thesmall percentage of non-viable cells (red fluorescent sub-population) was similar to that of the green fluorescentsubpopulations in two strains of C. palmioleophila,M3-8 and M3-28. However, in the case of C. parapsilo-sis M3-12, Y. lipolytica M3-21, C. davisiana M3-31 andC. palmioleophila M3-42 biofilms, dead cells appearedin lower layers than the green fluorescent stained ones(probably where nutrients were less accessible). Lastly,24 h biofilms formed by D. hansenii M3-16 andR. mucilaginosa M3-18 had some dead cells at the upperlevels, close to the biofilm surface.
76 R. Liébana et al.
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

A mixed culture of different strains was also used todetermine whether the interactions between various pop-ulations had any influence on the formation of the bio-film. The mixture of strains formed a biofilm with anaverage thickness of 25.7 μm, with a substratum cover-age < 50%, which was one of the lowest values involv-ing biofilm biovolume (4.89 × 107 μm3), with 2.73% ofthe dead cells located in deeper biofilm layers than thoseoccupied by the predominantly viable subpopulation.The images showed dense cell clusters and a slight ten-dency to form filaments.
Characterization of the population of protists andmicrometazoa
The identification and quantification of protists and mi-crometazoa showed that in terms of overall abundancethe Vorticella aquadulcis complex and Epistylis sp. werekey members of the community, with 8 × 102 and 7.2 ×102 individuals ml−1, respectively. With intermediate
values, and in order of decreasing density, were the fol-lowing: Acineria sp. (4.2 × 102 individuals ml−1), Op-ercularia sp. (4.0 × 102 individuals ml−1),amoeba > 50 μm (3.2 × 102 individuals ml−1), amoeba20–50 μm (2.6 × 102 individuals ml−1), rotifers (2.0 × 102
individuals ml−1) and Euplotes sp. (1.4 × 102 individualsml−1); with the least representative members beingPseudovorticella sp. (4.0 × 101 individuals ml−1), Vorti-cella microstoma complex (2.0 × 101 individuals ml−1)and Litonotus sp. (2.0 × 101 individuals ml−1). In otherwords, the community of protists was represented mainlyby both sessile and crawling ciliates associated with thefloc, together with amoeboid organisms.
The dominant group was the ciliates (77%), followedby amoeba (17%), with the minority group being micro-metazoa (6%). With respect to the ciliates, the majoritygroup was sessile ciliates (77%), followed by crawlingciliates (22%), with the remaining 1% consisting of free-swimming ciliates.
Figure 3. CLSM images (200×) of the 24 h biofilms developed in a synthetic wastewater medium by a selection of several stronglyadherent yeast isolates. The spatial distribution of viable (fluorescent green) and nonviable cells (fluorescent red) varied depending onthe yeast strain. Panel F shows a multi-specific biofilm with high cell mortality, indicating the presence of interspecific relationships.(A) Candida palmioleophila M3-42; (B) Rhodotorula mucilaginosa M3-18; (C) Yarrowia lipolytica M3-21; (D) Debaryomyceshansenii M3-16; (E) Candida palmioleophila M3-28; (F) multi-specific biofilm developed by all the strains tested (M3-8, M3-12,M3-16, M3-18, M3-21, M3-28, M3-31 and M3-42).
Biofouling 77
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

Protist ability to ingest yeast
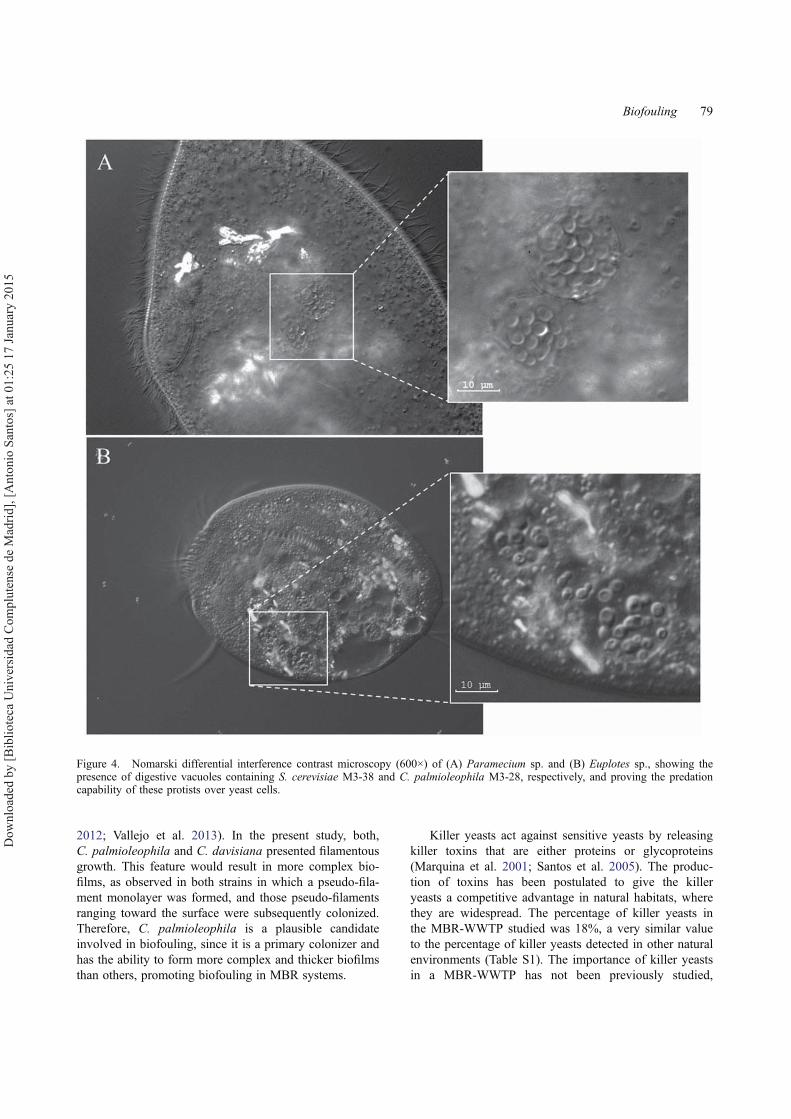
The trophic relationship between protists and yeasts wasstudied under laboratory conditions, firstly by usingC. palmioleophila M3-28 and Euplotes sp. The resultsindicate that Euplotes sp., which was observed to crawlon the floc surface (moving its ventral cirri) or swim-ming freely between flocs, was able to ingest C. palmi-oleophila M3-28, a yeast strain characterized by itsstrong adherence capability and its effectiveness in sur-face colonization (58.17%). Similarly, Paramecium sp.,selected as representative of the bacterivorous free-swim-ming ciliates, revealed its ability to ingest S. cerevisiae.The results showed several intracellular vacuoles locatedclose to the proximal end of the oral area of both ciliatesthat contained yeast cells, where cell digestion was prob-ably taking place, indicating that these ciliates have anoral funnel that is large enough to ingest yeast (Figure 4).
Discussion
Identification and characterization of isolated yeast
It was found here that the yeast community present inthe mixed liquor of a WWTP with MBR systems isunexpectedly diverse and abundant. While in naturalbacterial communities < 1% of the species may be cul-tured, some authors report that the number of cultivableyeasts in wastewater (Yang et al. 2011) and soil environ-ments (Stringini et al. 2008) is very high, and may besimilar to that obtained with independent cultivationmethods. The research presented here provides informa-tion about the capacity of several yeast strains to attachto a surface and their ability to develop biofilm, with theaim being to investigate their potential contribution tomembrane fouling. Nonculturable yeasts could thereforenot be considered. The data on viable yeast cells wereconsistent with those obtained in other studies thatreported yeast concentrations ranging between 102 and105 CFU g−1 sludge (Yang et al. 2011). Although theyeast community is not as significant as the bacterial orprotist community in an activated sludge ecosystem, it isvery diverse in a WWTP, as observed in recent studies(Yang et al. 2011, 2013). The results obtained in thisstudy are in line with this. The diversity of yeast generaamong the 67 isolates was significant, with Candidabeing the predominant genus, a fact that has also beenreported previously in activated sludge studies. Othergenera detected, such as Rhodotorula, Debaryomyces,Saccharomyces, Yarrowia and Filobasidium, were alsomentioned (Yang et al. 2011). The characteristics of theWWTP and the physico-chemical nature of the actualwastewater could determine the composition of the yeastcommunity and the prevalence of certain species overothers depending on their metabolic capabilities (Yanget al. 2013).
Many yeast species are known to thrive in simpleaqueous media at neutral or slightly acidic pH values, aswas observed in synthetic residual water. A WWTP con-tains nutrients that yeasts could use for survival andgrowth. Yeasts are extremely diverse in their nutritionalrequirements, and this diversity should be considered ina WWTP. In nature, yeasts function as natural decom-posers, assimilating carbon and nitrogen sources withvarying degrees of effectiveness. Yeasts are able to breakdown complex carbon compounds and to assimilate agreat variety of simple inorganic and organic nitrogensources. The presence of bacterial communities in con-centrations that are hundreds or thousands of timeshigher than those of yeast may relegate the role of yeastsas decomposers to a secondary level. However, yeastsshould not be overlooked or underestimated as they dohave diverse metabolic capabilities (Hagler & Aheard1987).
Characterization of yeast isolates
Different yeast strains have recently been studied todetermine the production of hydrolyzing enzymes andthe degradation of certain contaminants in a WWTP(Jarboui et al. 2012; Yang et al. 2013). Due to the partic-ular problem concerning biofouling in harboring MBRsystems, different features, such as surface adhesion, bio-film formation or killer toxin production, have been ana-lyzed using yeast isolates. Yeasts are known to be ableto adhere to abiotic surfaces and develop biofilms, whichexemplifies their impressive phenotypic plasticity, allow-ing them to adapt quickly to stressful environments(Verstrepen & Klis 2006). Moreover, the adhesionbehavior of closely related strains can be substantiallydifferent (Verstrepen et al. 2004). The control of biofilmformation on membrane surfaces should focus upon pio-neer populations rather than on the total microbial com-munity present in the mixed liquor of MBR systems.Identifying the populations firmly attached onto themembrane surfaces during the early period of filtrationprovides a list of candidates for future surface coloniza-tion and biofilm research (Zhang et al. 2006). In thissense, the yeasts isolated and characterized were able toadhere to the surface assayed, which has similar chemi-cal and physical properties to the MBR membrane. Fur-thermore, species such as C. palmioleophila, R.mucilaginosa and Y. lipolytica colonized > 50% of thesurface in 24 h, keeping cells metabolically active, so itmay be affirmed that these species could be initial colo-nizers of the filtration membrane. Moreover, C. palmi-oleophila is not only a pioneer in the colonization, as itis also able to form thick biofilms at 24 h.
Traditionally, filamentous growth has been interpretedas a strategy adopted by yeast as a dispersion mechanismto obtain access to more resources (Cullen & Sprague
78 R. Liébana et al.
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015
2012; Vallejo et al. 2013). In the present study, both,C. palmioleophila and C. davisiana presented filamentousgrowth. This feature would result in more complex bio-films, as observed in both strains in which a pseudo-fila-ment monolayer was formed, and those pseudo-filamentsranging toward the surface were subsequently colonized.Therefore, C. palmioleophila is a plausible candidateinvolved in biofouling, since it is a primary colonizer andhas the ability to form more complex and thicker biofilmsthan others, promoting biofouling in MBR systems.
Killer yeasts act against sensitive yeasts by releasingkiller toxins that are either proteins or glycoproteins(Marquina et al. 2001; Santos et al. 2005). The produc-tion of toxins has been postulated to give the killeryeasts a competitive advantage in natural habitats, wherethey are widespread. The percentage of killer yeasts inthe MBR-WWTP studied was 18%, a very similar valueto the percentage of killer yeasts detected in other naturalenvironments (Table S1). The importance of killer yeastsin a MBR-WWTP has not been previously studied,
Figure 4. Nomarski differential interference contrast microscopy (600×) of (A) Paramecium sp. and (B) Euplotes sp., showing thepresence of digestive vacuoles containing S. cerevisiae M3-38 and C. palmioleophila M3-28, respectively, and proving the predationcapability of these protists over yeast cells.
Biofouling 79
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

although this aspect could be an interesting new area ofresearch in such an artificial habitat. In microenviron-ments, such as those present in the flocs of the activatedsludge or in the biofilms attached to the filtration mem-branes of the MBR systems, the production of killer tox-ins might imply a competitive advantage between yeastpopulations. By contrast, among the factors that areknown to affect toxin activity, pH and temperature wereshown to be the most important (Santos et al. 2009).Their stability at only acidic pH values (< 5.0) and lowtemperatures (< 25°C) compromises their potential roleas competitive factors during microbial development andbiofilm formation in activated sludge.
Characterization of the population of protists andmicrometazoa
The MBR configuration features certain differences toconventional systems: the amount of biomass in the reac-tor is greater due to the higher retention time of thesludge, and the system therefore operates with a lowersludge loading, which also affects aeration efficiency.Studies on the protist populations in MBR systems arescarce. A prevalence of ciliates was found in the reactor,with the species identified being commonly detected in aWWTP (Serrano et al. 2008; Seviour & Nielsen 2010).This prevalence of sessile and crawling ciliates has in allcases been related to good operational conditions in acti-vated sludge plants (Martín-Cereceda et al. 1996).Amoebae and flagellates also constitute significant popu-lations within these systems, as reported by Arévaloet al. (2009). Thus, the distribution of protist groups inthe MBR WWTP studied could be compared with thoseconventional activated sludge systems operating withhigh solid retention times in the reactor, and this findingwill also explain the high density of micrometazoadetected (Salvadó 1994).
Protist ability to ingest planktonic yeast cells
Predation by protists is considered to be one of the majorcauses of the decline in the number of bacteria in aquaticand terrestrial ecosystems (Jürgens & Matz 2002; Sherr& Sherr 2002). Protists can clarify media with high bac-terial density due to their mobility, small size and abun-dance (Matz & Kjelleberg 2005). Predation by ciliatestakes place via specific strategies of phagotrophic uptake,with factors such as the size, quality, behavior and mor-phology of the prey, as well as environmental conditions,determining the predatory capacity of protists (Pfister &Arndt 1998; Jezbera et al. 2005).
Several authors have studied the influence of protistpredation on bacteria in a WWTP (Salvadó et al. 1995;Ratsak et al. 1996; Decamp & Warren 1998). However,
very few studies have addressed the capacity of protiststo ingest yeast, with the potential use of these microor-ganisms to clear yeast from media being of great interest.The two genera of protists analyzed, frequently found inthe biological reactors of WWTPs, could be consideredexcellent candidates for yeast predation, since both havea well-developed oral cavity. The results of this workshow that yeasts are devoured by Euplotes sp. and Para-mecium sp., as the yeast cells can be observed inside thedigestive vacuoles of the ciliates. Predation by protistscauses a selective pressure on the prey on which theyfeed, so different phenotypic changes, such as anincrease in cell size, filamentation or biofilm formation,could occur in response to this biological activity (Matz& Kjelleberg 2005).
In conclusion, activated sludge obtained from a full-scale MBR contained a diverse yeast community. Theisolated yeasts were able to adhere to polymeric sub-strata, similar to those found in membrane modules ofWWTPs. Their adhesion capabilities varied widely, sug-gesting that some yeast strains may contribute more thanother strains to the biofouling of membranes in MBRsystems. In this study the ecological role of yeasts withinthe microbial community inhabiting the MBR was alsoconsidered. It was found that in addition to their reportedhydrolytic capabilities which could contribute to waste-water degradation, yeasts were predated by the protistsEuplotes sp. (crawling ciliate) and Paramecium sp.(swimming ciliate). Therefore yeasts should be consid-ered as being part of the food chain and it can be statedthat their role and importance in wastewater treatmenthas not yet been fully understood.
AcknowledgementsThe authors thank Dr Antonio Murciano (Applied MathematicsDepartment, Complutense University of Madrid) and DrHumbert Salvadó (Animal Biology Department, University ofBarcelona) for helpful discussions. They are also grateful to theProteomic and Genomic Centre and the Cytometry and Fluores-cence Microscopy Centre (Complutense University of Madrid)for technical assistance.
Conflict of interest disclosure statementNo potential conflict of interest was reported by the author(s).
FundingThis work was supported by Spain’s Ministry of Science andInnovation [grant CTQ2011-29231-C05-01].
Supplemental materialThe supplemental material for this paper is available online athttp://dx.doi10.1080/08927014.2014.998206.
80 R. Liébana et al.
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

ReferencesArévalo J, Moreno B, Pérez J, Gómez MA. 2009. Applicability
of the sludge biotic index (SBI) for MBR activated sludgecontrol. J Hazard Mater. 167:784–789.
Awad MF, Kraume M. 2010. The occurrence of fungi in acti-vated sludge from MBRs. World Acad Sci Eng Technol.71:561–564.
Awad MF, Kraume M. 2011. Fungal diversity in activatedsludge from membrane bioreactors in Berlin. Can J Micro-biol. 57:693–698.
Basak B, Bhunia B, Dutta S, Chakraborty S, Dey A. 2014.Kinetics of phenol biodegradation at high concentration bya metabolically versatile isolated yeast Candida tropicalisPHB5. Environ Sci Pollut Res. 21:1444–1454.
Cabrol L, Malhautier L. 2011. Integrating microbial ecology inbioprocess understanding: the case of gas biofiltration. ApplMicrobiol Biotechnol. 90:837–849.
Caron DA, Countway PD, Jones AC, Kim DY, Schnetzer A.2012. Marine protistan diversity. Ann Rev Mar Sci. 4:467–493.
Chávez de Paz LE. 2009. Image analysis software based oncolor segmentation for characterization of viability andphysiological activity of biofilms. Appl Environ Microbiol.75:1734–1739.
Cullen PJ, Sprague GF Jr. 2012. The regulation of filamentousgrowth in yeast. Genetics. 190:23–49.
Decamp O, Warren A. 1998. Bacterivory in ciliates isolatedfrom constructed wetlands (reedbeds) used for wastewatertreatment. Water Res. 32:1989–1996.
Feng L, Li X, Song P, Du G, Chen J. 2011. Surface interac-tions and fouling properties of Micrococcus luteus with mi-crofiltration membranes. Appl Biochem Biotechnol.165:1235–1244.
Fernández-Galiano D. 1976. Silver impregnation of ciliatedprotozoa: procedure yielding good results with the pyrini-dated silver carbonated method. Trans Am Microsc Soc.95:557–560.
Hagler AN, Aheard DG. 1987. The yeasts. In: Rose AH,Harrison JS, editors. Ecology of aquatic yeasts Vol 2.London: Academic Press; p. 181–205.
Hülsmann N, Galil BS. 2001. The effects of freshwater flushingon marine heterotrophic protists – implications for ballastwater management. Mar Pollut Bull. 42:1082–1086.
Jarboui R, Baati H, Fetoui F, Gargouri A, Gharsallah N,Ammar E. 2012. Yeast performance in wastewater treat-ment: case study of Rhodotorula mucilaginosa. EnvironTechnol. 33:951–960.
Jarboui R, Magdich S, Ayadi RJ, Gargouri A, Gharsallah N,Ammar E. 2013. Aspergillus niger P6 and Rhodotorula mu-cilaginosa CH4 used for olive mill wastewater (OMW) bio-logical treatment in single pure and successive cultures.Environ Technol. 34:629–636.
Jezbera J, Hornak K, Simek K. 2005. Food selection by bacte-rivorous protists: insight from the analysis of the food vac-uole content by means of fluorescence in situ hybridization.FEMS Microbiol Ecol. 52:351–363.
Ju F, Xia Y, Guo F, Wang Z, Zhang T. 2014. Taxonomic relat-edness shapes bacterial assembly in activated sludge ofglobally distributed wastewater treatment plants. EnvironMicrobiol. doi:10.1111/1462-2920.12355.
Jürgens K, Matz C. 2002. Predation as a shaping force for thephenotypic and genotypic composition of planktonic bacte-ria. Antonie van Leeuwenhoek. 81:413–434.
Kurtzman CP, Robnett CJ. 1991. Phylogenetic relationshipsamong species of Saccharomyces, Schizosaccharomyces,
Debaryomyces and Schwanniomyces determined from par-tial ribosomal RNA sequences. Yeast. 7:61–72.
Lee JJ, Leedale GF, Bradbury P. 2000. The illustrated guide tothe protozoa. Society of Protozoologists. 2nd ed. Lawrence(KS): Allen Press.
Llorente P, Marquina D, Santos A, Peinado JM, Spencer-MartinsI. 1997. Effect of salt on the killer phenotype of yeasts fromolive brines. Appl Environ Microbiol. 63:1165–1167.
Lopandic K, Rentsendorj U, Prillinger H, Sterflinger K. 2013.Molecular characterization of the closely related Debary-omyces species: proposition of D. vindobonensis sp. nov.from a municipal wastewater treatment plant. J Gen ApplMicrobiol. 59:49–58.
Madoni P. 1991. Role of protozoans and their indicator valuein the activated sludge process. In: Biological approach tosewage treatment process: current status and perspectives.Perugia: Centro Bazzucchi; p. 21–27.
Madoni P. 1994. A sludge biotic index (SBI) for the evaluationof the biological performance of activated sludge plantsbased on the microfauna analysis. Water Res. 28:67–75.
Madoni P, Davoli D, Chierici E. 1993. Comparative analysis ofthe activated sludge microfauna in several sewage treatmentworks. Water Res. 27:1485–1491.
Marquina D, Barroso J, Santos A, Peinado JM. 2001. Produc-tion and characteristics of Debaryomyces hansenii killertoxin. Microbiol Res. 156:387–391.
Martín-Cereceda M, Serrano S, Guinea A. 1996. A comparativestudy of ciliated protozoa communities in activated sludgeplants. FEMS Microbiol Ecol. 21:267–276.
Matz C, Kjelleberg S. 2005. Off the hook – how bacteria sur-vive protozoan grazing. Trends Microbiol. 13:302–307.
Matz C, Webb JS, Schupp PJ, Phang SY, Penesyan A, SuhelenE, Steinberg P, Kjelleberg S. 2008. Marine biofilm bacteriaevade eukaryotic predation by targeted chemical defense.PLoS ONE. 3:e2744.
Meng F, Chaeb SR, Drewsc A, Kraumec M, Shind HS, YangF. 2009. Recent advances in membrane bioreactors(MBRs): membrane fouling and membrane material. WaterRes. 43:1489–1512.
Merritt JH, Kadouri DE, O’Toole GA. 2011. Growing and ana-lyzing static biofilms. Curr Protoc Microbiol. doi:10.1002/9780471729259.mc01b01s00.
Nielsen PH, Saunders AM, Hansen AA, Larsen P, Nielsen JL.2012. Microbial communities involved in enhanced biologi-cal phosphorus removal from wastewater – a model systemin environmental biotechnology. Curr Opin Biotechnol.23:452–459.
Oh HS, Yeon KM, Yang CS, Kim SR, Lee CH, Park SY, HanJY, Lee JK. 2012. Control of membrane biofouling inMBR for wastewater treatment by quorum quenching bac-teria encapsulated in microporous membrane. Environ SciTechnol. 46:4877–4884.
Pérez-Uz B, Arregui L, Calvo P, Salvadó H, Fernández N,Rodríguez E, Zornoza A, Serrano S. 2010. Assessment ofplausible bioindicators for plant performance in advancedwastewater treatments. Water Res. 44:5059–5069.
Pfister G, Arndt H. 1998. Food selectivity and feeding behaviorin omnivorous filter-feeding ciliates: a case study for Styl-onychia. Eur J Protistol. 34:446–457.
Querol A, Barrio E, Huerta T, Ramón D. 1992. Molecularmonitoring of wine fermentations conducted by active dryyeast strains. Appl Environ Microbiol. 58:2948–2953.
Ratsak CH, Maarsen KA, Kooijman SALM. 1996. Effects ofprotozoa on carbon mineralization in activated sludge.Water Res. 30:1–12.
Biofouling 81
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015

Salvadó H. 1994. Effect of mean cellular retention time on cili-ated protozoan populations in urban wastewater treatmentplants based on proposed model. Water Res. 28:1315–1321.
Salvadó H, Gracia MP, Amigó JM. 1995. Capability of ciliatedprotozoa as indicators of effluent quality in activated sludgeplants. Water Res. 29:1041–1050.
Santos A, Del Mar Alvarez M, Mauro MS, Abrusci C,Marquina D. 2005. The transcriptional response of Saccha-romyces cerevisiae to Pichia membranifaciens killer toxin.J Biol Chem. 280:41881–41892.
Santos A, San Mauro M, Bravo E, Marquina D. 2009. PMKT2,a new killer toxin from Pichia membranifaciens, and itspromising biotechnological properties for control of thespoilage yeast Brettanomyces bruxellensis. Microbiology.155:624–634.
Serrano S, Arregui L, Pérez-Uz B, Calvo P, Guinea A. 2008.Guidelines for the identification of ciliates in wastewatertreatment plants. London: IWA; p. 112.
Seviour R, Nielsen PH. 2010. Microbial ecology of activatedsludge. London: IWA; p. 667.
Sherr EB, Sherr BF. 2002. Significance of predation by protistsin aquatic microbial food webs. Antonie van Leeuwenhoek.81:293–308.
Stepanovic S, Vukovic D, Dakic I, Savic B, Svabic-VlahovicM. 2000. A modified microtiter-plate test for quantificationof staphylococcal biofilm formation. J Microbiol Methods.40:175–179.
Stringini M, Comitini F, Taccari M, Ciani M. 2008. Yeastdiversity in crop-growing environments in Cameroon. Int JFood Microbiol. 127:184–189.
Sweity A, Ying A, Ali-Shtayeh MS, Yang F, Bick A, Oron G,Herzberg M. 2011. Relation between EPS adherence, visco-elastic properties, and MBR operation: biofouling studywith QCM-D. Water Res. 45:6430–6440.
Swofford DL. 2002. PAUP. Phylogenetic analysis using parsi-mony (and other methods). Sunderland (MA): Sinauer.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, KumarS. 2011. MEGA5: molecular evolutionary genetics analysisusing maximum likelihood, evolutionary distance, and max-imum parsimony methods. Mol Biol Evol. 28:2731–2739.
Vallejo JA, Sánchez-Pérez A, Martínez JP, Villa TG. 2013. Cellaggregations in yeasts and their applications. Appl Micro-biol Biotechnol. 97:2305–2318.
Verstrepen KJ, Klis FM. 2006. Flocculation, adhesion and bio-film formation in yeasts. Mol Microbiol. 60:5–15.
Verstrepen KJ, Reynolds TB, Fink GR. 2004. Origins of variationin the fungal cell surface. Nat Rev Microbiol. 2:533–540.
Watanabe T, Ikeda H, Masaki K, Fujii T, Iefuji H. 2009. Clon-ing and characterization of a novel phytase from wastewa-ter treatment yeast Hansenula fabianii J640 and expressionin Pichia pastoris. J Biosci Bioeng. 108:225–230.
Wu SC, Lee CM. 2011. Correlation between fouling propensityof soluble extracellular polymeric substances and sludgemetabolic activity altered by different starvation conditions.Bioresour Technol. 102:5375–5380.
Wu B, Yi S, Fane AG. 2011. Microbial behaviors involved incake fouling in membrane bioreactors under different solidsretention times. Bioresour Technol. 102:2511–2516.
Yang Q, Angly FE, Wang Z, Zhang H. 2011. Wastewater treat-ment systems harbor specific and diverse yeast communi-ties. Biochem Eng J. 58-59:168–176.
Yang Q, Zhang H, Li X, Wang Z, Xu Y, Ren S, Chen X, XuY, Hao H, Wang H. 2013. Extracellular enzyme productionand phylogenetic distribution of yeasts in wastewater treat-ment systems. Bioresour Technol. 129:264–273.
Zhang K, Choi H, Dionysiou DD, Sorial GA, Oerther DB.2006. Identifying pioneer bacterial species responsible forbiofouling membrane bioreactors. Environ Microbiol.8:433–440.
82 R. Liébana et al.
Dow
nloa
ded
by [B
iblio
teca
Uni
vers
idad
Com
plut
ense
de
Mad
rid],
[Ant
onio
San
tos]
at 0
1:25
17
Janu
ary
2015




















