
Mechanisms regulating the recruitment of macrophages into hypoxic areas of tumors and other ischemic...
11
Review article Mechanisms regulating the recruitment of macrophages into hypoxic areas of tumors and other ischemic tissues Craig Murdoch, Athina Giannoudis, and Claire E. Lewis The mechanisms responsible for recruit- ing monocytes from the bloodstream into solid tumors are now well characterized. However, recent evidence has shown that these cells then differentiate into macro- phages and accumulate in large numbers in avascular and necrotic areas where they are exposed to hypoxia. This paral- lels their tendency to congregate in isch- emic areas of other diseased tissues such as atherosclerotic plaques and arthritic joints. In tumors, macrophages appear to undergo marked phenotypic changes when exposed to hypoxia and to switch on their expression of a number of mito- genic and proangiogenic cytokines and enzymes. This then promotes tumor growth, angiogenesis, and metastasis. Here, we compare the various mecha- nisms responsible for monocyte recruit- ment into tumors with those regulating the accumulation of macrophages in hy- poxic/necrotic areas. Because the latter are best characterized in human tumors, we focus mainly on these but also dis- cuss their relevance to macrophage mi- gration in ischemic areas of other dis- eased tissues. Finally, we discuss the relevance of these mechanisms to the development of novel cancer therapies, both in providing targets to reduce the proangiogenic contribution made by hy- poxic macrophages in tumors and in de- veloping the use of macrophages to de- liver therapeutic gene constructs to hypoxic areas of diseased tissues. (Blood. 2004;104:2224-2234) © 2004 by The American Society of Hematology Introduction Macrophages are essential cellular components of the innate immune system. They are released from the bone marrow as immature monocytes and circulate in the blood before extravasat- ing into tissues, where they differentiate into resident macrophages. These cells can be found in almost all tissues of the body and, depending on the local microenvironment, acquire specialized phenotypic characteristics. Macrophages exhibit diverse functions, including phagocytosis, antigen presentation, antimicrobial cytotox- icity, and tissue remodeling as well as the secretion of a wide range of growth factors, cytokines, complement components, prostaglan- dins, and enzymes. 1 The presence of leukocytes in human tumors was first described by Virchow in 1863, who thought they reflected the onset of cancer at sites of previous chronic inflammation. It is now widely recognized that macrophages represent a prominent component of this leukocytic infiltrate in most malignant tumors and in some instances can comprise up to 50% of the cell tumor mass. 2,3 These cells, often called tumor-associated macrophages (TAMs), are thought to be almost entirely derived from peripheral blood monocytes recruited into the tumor from the local circulation (rather than resident macrophages present in the healthy tissue before the tumor developed). 4 The various possible roles of TAMs in tumor angiogenesis and progression have recently been reviewed extensively elsewhere. 5-8 Macrophages can exhibit direct cytotoxicity toward tumor cells in vitro by producing cytotoxic molecules such as tumor necrosis factor-alpha (TNF-), nitric oxide, and reactive oxygen intermedi- ates as well as by stimulating lymphocyte responsiveness via the presentation of tumor-associated antigens and production of inter- leukin-12 (IL-12). Alternatively, TAMs can promote tumor growth by secreting growth factors that promote the proliferation, inva- sion, and metastasis of tumor cells as well as the vascularization of tumors. They also inhibit lymphocyte activity at the tumor site by producing such immunosuppressive cytokines as IL-10 and prosta- noids. 9-11 The protumor functions could explain our finding that the extent of tumor infiltration by macrophages correlates with in- creased tumor angiogenesis and reduced overall and recurrence- free survival in breast cancer. 11 Interestingly, we found that TAM density was highest in avascular and necrotic (ie, hypoxic) areas of these breast tumors and that it was their density in these areas that was most closely linked to tumor angiogenesis and patient outcome. Oxygen microelectrodes and hypoxia-specific markers have demonstrated the presence of many areas of hypoxia and anoxia (complete lack of oxygen) in most forms of human tumor, including those of the breast, brain, cervix, head/neck, and soft tissue sarcomas 12,13 (Figure 1A). Whereas normal tissues typically exhibit median oxygen tensions of 30 to 70 mm Hg, most solid tumors contain multiple areas with a median value of less than 10 mm Hg. These hypoxic/anoxic regions appear because the newly formed blood vessels in tumors are often disorganized with many blind ends, incomplete endothelial linings, and basement mem- branes and have a tendency to collapse. 14 Consequently, blood flow is sluggish and irregular, and the delivery of oxygen and nutrients is poor to many regions of the tumor. The rapid expansion and oxygen consumption of tumor cells around these new blood vessels also contributes to the level of hypoxia formed, although once a From the Tumor Targeting Group, Academic Unit of Pathology, Division of Genomic Medicine, University of Sheffield Medical School, United Kingdom. Submitted March 24, 2004; accepted May 28, 2004. Prepublished online as Blood First Edition Paper, July 1, 2004; DOI 10.1182/blood-2004-03-1109. Supported by the Biotechnology and Biological Sciences Research Council (BBSRC) (C.M.), Yorkshire Cancer Research (YCR) (A.G.), and the Tumor Targeting Group from the BBSRC, Medical Research Council (MRC), Engineering and Physical Sciences Research Council (EPSRC), and the Association for International Cancer Research (AICR) (C.E.L.). Reprints: Claire Lewis, Tumor Targeting Group, Academic Unit of Pathology, Division of Genomic Medicine, University of Sheffield Medical School, Beech Hill Rd, Sheffield S10 2RX, United Kingdom; e-mail: [email protected]. © 2004 by The American Society of Hematology 2224 BLOOD, 15 OCTOBER 2004 VOLUME 104, NUMBER 8
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Mechanisms regulating the recruitment of macrophages into hypoxic areas of tumors and other ischemic...

Review article
Mechanisms regulating the recruitment of macrophages into hypoxic areas oftumors and other ischemic tissuesCraig Murdoch, Athina Giannoudis, and Claire E. Lewis
The mechanisms responsible for recruit-ing monocytes from the bloodstream intosolid tumors are now well characterized.However, recent evidence has shown thatthese cells then differentiate into macro-phages and accumulate in large numbersin avascular and necrotic areas wherethey are exposed to hypoxia. This paral-lels their tendency to congregate in isch-emic areas of other diseased tissues suchas atherosclerotic plaques and arthriticjoints. In tumors, macrophages appear toundergo marked phenotypic changes
when exposed to hypoxia and to switchon their expression of a number of mito-genic and proangiogenic cytokines andenzymes. This then promotes tumorgrowth, angiogenesis, and metastasis.Here, we compare the various mecha-nisms responsible for monocyte recruit-ment into tumors with those regulatingthe accumulation of macrophages in hy-poxic/necrotic areas. Because the latterare best characterized in human tumors,we focus mainly on these but also dis-cuss their relevance to macrophage mi-
gration in ischemic areas of other dis-eased tissues. Finally, we discuss therelevance of these mechanisms to thedevelopment of novel cancer therapies,both in providing targets to reduce theproangiogenic contribution made by hy-poxic macrophages in tumors and in de-veloping the use of macrophages to de-liver therapeutic gene constructs tohypoxic areas of diseased tissues. (Blood.2004;104:2224-2234)
© 2004 by The American Society of Hematology
Introduction
Macrophages are essential cellular components of the innateimmune system. They are released from the bone marrow asimmature monocytes and circulate in the blood before extravasat-ing into tissues, where they differentiate into resident macrophages.These cells can be found in almost all tissues of the body and,depending on the local microenvironment, acquire specializedphenotypic characteristics. Macrophages exhibit diverse functions,including phagocytosis, antigen presentation, antimicrobial cytotox-icity, and tissue remodeling as well as the secretion of a wide rangeof growth factors, cytokines, complement components, prostaglan-dins, and enzymes.1
The presence of leukocytes in human tumors was first describedby Virchow in 1863, who thought they reflected the onset of cancerat sites of previous chronic inflammation. It is now widelyrecognized that macrophages represent a prominent component ofthis leukocytic infiltrate in most malignant tumors and in someinstances can comprise up to 50% of the cell tumor mass.2,3 Thesecells, often called tumor-associated macrophages (TAMs), arethought to be almost entirely derived from peripheral bloodmonocytes recruited into the tumor from the local circulation(rather than resident macrophages present in the healthy tissuebefore the tumor developed).4
The various possible roles of TAMs in tumor angiogenesis andprogression have recently been reviewed extensively elsewhere.5-8
Macrophages can exhibit direct cytotoxicity toward tumor cells invitro by producing cytotoxic molecules such as tumor necrosisfactor-alpha (TNF-�), nitric oxide, and reactive oxygen intermedi-ates as well as by stimulating lymphocyte responsiveness via the
presentation of tumor-associated antigens and production of inter-leukin-12 (IL-12). Alternatively, TAMs can promote tumor growthby secreting growth factors that promote the proliferation, inva-sion, and metastasis of tumor cells as well as the vascularization oftumors. They also inhibit lymphocyte activity at the tumor site byproducing such immunosuppressive cytokines as IL-10 and prosta-noids.9-11 The protumor functions could explain our finding that theextent of tumor infiltration by macrophages correlates with in-creased tumor angiogenesis and reduced overall and recurrence-free survival in breast cancer.11 Interestingly, we found that TAMdensity was highest in avascular and necrotic (ie, hypoxic) areas ofthese breast tumors and that it was their density in these areas thatwas most closely linked to tumor angiogenesis and patient outcome.
Oxygen microelectrodes and hypoxia-specific markers havedemonstrated the presence of many areas of hypoxia and anoxia(complete lack of oxygen) in most forms of human tumor,including those of the breast, brain, cervix, head/neck, and softtissue sarcomas12,13 (Figure 1A). Whereas normal tissues typicallyexhibit median oxygen tensions of 30 to 70 mm Hg, most solidtumors contain multiple areas with a median value of less than 10mm Hg. These hypoxic/anoxic regions appear because the newlyformed blood vessels in tumors are often disorganized with manyblind ends, incomplete endothelial linings, and basement mem-branes and have a tendency to collapse.14 Consequently, blood flowis sluggish and irregular, and the delivery of oxygen and nutrients ispoor to many regions of the tumor. The rapid expansion and oxygenconsumption of tumor cells around these new blood vessels alsocontributes to the level of hypoxia formed, although once a
From the Tumor Targeting Group, Academic Unit of Pathology, Division ofGenomic Medicine, University of Sheffield Medical School, United Kingdom.
Submitted March 24, 2004; accepted May 28, 2004. Prepublished online asBlood First Edition Paper, July 1, 2004; DOI 10.1182/blood-2004-03-1109.
Supported by the Biotechnology and Biological Sciences Research Council(BBSRC) (C.M.), Yorkshire Cancer Research (YCR) (A.G.), and the TumorTargeting Group from the BBSRC, Medical Research Council (MRC),
Engineering and Physical Sciences Research Council (EPSRC), and theAssociation for International Cancer Research (AICR) (C.E.L.).
Reprints: Claire Lewis, Tumor Targeting Group, Academic Unit ofPathology, Division of Genomic Medicine, University of Sheffield MedicalSchool, Beech Hill Rd, Sheffield S10 2RX, United Kingdom; e-mail:[email protected].
© 2004 by The American Society of Hematology
2224 BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8

threshold level of hypoxia is reached, tumor cells in that area stopproliferating and switch to anaerobic glycolysis for energy produc-tion.15 Various studies have shown that the presence of large areasof hypoxia in tumors correlates with poor prognosis.16 This isthought to be because hypoxic tumor cells are relatively resistant tosuch conventional anticancer therapies as radiotherapy and chemo-therapy. Well-oxygenated tumor cells are markedly more respon-sive to radiotherapy than their hypoxic counterparts becauseoxygen-derived free radicals potentiate the protein and DNAdamage induced by the ionizing radiation. Most anticancer chemo-therapeutic agents only kill tumor cells if they are rapidlyproliferating, so the nonproliferative, hypoxic fractions of tumorsare relatively resistant to their effects.
While in this hypoxic, nonproliferative state, tumor cells arealso known to secrete cytokines and enzymes to induce the growthof new blood vessels within the tumor, thereby providing oxygenand nutrients for tumor growth as well as increased exit routes fortumor cells into the general circulation. Hypoxia also exerts aselective pressure on tumor cells because only those with anaggressive phenotype (eg, mutated for the tumor suppressor gene,p53) are able to survive hypoxia and go on to repopulate the tumor
and metastasize to distant sites.14 Because these hypoxic areas arerelatively inaccessible to conventional anticancer drugs and genevectors (due to the absence of a blood supply), recent research hasfocused on the development of novel drug/gene vectors capable ofpenetrating these regions in tumors.
Several studies analyzing macrophage migration into tumorshave reported the presence of TAMs in stromal areas of tumors(Figure 1B).17,18 However, as mentioned previously, there isincreasing evidence to show that TAMs mainly accumulate inavascular, necrotic/hypoxic areas of tumors where they presumablyact to clear necrotic cell debris from these sites (Figure 1C-F).17,19-22 Furthermore, we have also shown that TAMs located inthese sites respond to the hypoxia present with altered geneexpression leading to the development of a distinct protumorphenotype.23 These hypoxic responses of macrophages are unlikelyto be confined to tumors, because increased numbers of macro-phages have also been reported in ischemic areas of dermalwounds,24 atherosclerotic plaques,25 synovia of joints with rheuma-toid arthritis,26 and eyes with proliferative retinopathy.27 It istherefore possible that macrophages may exhibit a similar pheno-type in the hypoxic areas of these diseased tissues.
Here, we outline the various ways in which monocytes arerecruited into tumors and then the potential mechanisms used bythese tissues to attract and entrap TAMs in hypoxic areas. We alsodiscuss the likely relevance of these mechanisms to macrophagebehavior in hypoxic areas of other diseased tissues. Finally, weassess the therapeutic implications of these findings for thedevelopment of novel therapies for cancer and these other diseases.
Recruitment of monocytes into tumors
A number of chemokines have been shown to be involved in therecruitment of monocytes from the bloodstream. These are dividedinto 4 subclasses (CXC, CC, C, and CX3C), and the specific effectsof these molecules on their target cells are mediated by members ofa family of 7-transmembrane, G protein–coupled receptors.28,29
From research carried out over the past 2 decades it has becomeincreasingly evident that chemokines and their receptors play amajor role in all aspects of tumor biology, including recruitment ofleukocyte populations, manipulation of the tumor response, regula-tion of angiogenesis and tumor growth, and control of themovement of tumor cells during metastasis.30-32
Several CC chemokines, particularly CCL2 (formally monocytechemoattractant protein-1, or MCP-1) and CCL5 (RANTES, orregulated on activation normal T cells expressed and secreted),specifically attract and activate mononuclear cells, and it is thesemolecules that have been most heavily implicated in monocyterecruitment to human tumors (Table 1). CCL2 and CCL5 areproduced by tumor cells, fibroblasts, endothelial cells, and evenTAMs themselves, and their expression has been shown topositively correlate with TAM numbers in tumors (Table 1). CCL2gene transfer to xenograft and syngeneic murine tumors promotesmonocyte uptake.51 Moreover, when melanoma cells were trans-duced to express CCL2 and transplanted into severe combinedimmunodeficient (SCID) mice, the level of monocyte uptakecorrelated with the level of their expression of the chemokine.52
As well as being chemoattractants, both CCL2 and CCL5 havealso been shown to directly stimulate monocytes to expressproteins that aid not only in monocyte migration but also tumorprogression. For example, CCL5 stimulates human monocytes toexpress CCL2, CCL3 (macrophage inflammatory protein-1�),
Figure 1. Colocalization of hypoxia and macrophages. Colocalization in humansquamous carcinoma of the uterine cervix (A-D) and human PC3 (prostate carci-noma) xenografted tumors grown in nude mice (E-F). (A) Hypoxic areas (arrow),visualized by immunolabeling of the reductively activated hypoxic-specific markerpimonidazole (red), are observed at a distance from the disorganized blood vessels(BV). (B) In some areas of these tumors, CD68� TAMs (brown) accumulate in stromal(S) rather than tumor (T) areas. (C-D) Colocalization of TAMS (brown) and hypoxia(red) showing TAM accumulation in avascular, perinecrotic, necrotic (N), and hypoxiaareas. (E-F) Serial sections showing F4/80-positive (brown; panel F) TAMs inhypoxic, perinecrotic areas (red; panel E) of PC3 tumors. N indicates necrosis; and T,tumor. The microscope used was a Leitz Orthoplan. Magnification of panel A, � 100;panel B, � 400; panel C, � 160; panel D, � 160; panel E, � 160; panel F, � 160(panels D-F, cropped images). Slight adjustments were made in brightness andcontrast on each whole image. The temperature was room temperature. The imagingmedium was DPX. Chromogens used were DAB (brown) 3,3-diaminobenzene andvector red (red) (Vector Laboratories, Burlingame, CA). The camera was a Fuji HC3002 digital camera. Image processing was performed by Photograb 3002 (Windowsplatform).
RECRUITMENT OF MACROPHAGES INTO TUMORS 2225BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8
CCL4 (macrophage inflammatory protein-1�), and CXCL8 (inter-leukin-8) as well as the chemokine receptor CCR1. This suggeststhat chemokine activation of monocytes may amplify monocyterecruitment as well as that of other leukocyte populations.53
Furthermore, both CCL2 and CCL5 have been found to stimulatemonocytic cell lines, blood monocytes, and brain macrophages(microglia) to secrete matrix metalloproteinases-9 (MMP-9), MMP-19, and urokinase-type plasminogen activator receptor (uPA-R).53-56 These proteases degrade basement membrane and extracel-lular matrix components to aid leukocyte migration into tissues andhave also been implicated in the proteolytic remodeling of theextracellular matrix, which is important at several points duringmultistage progression of tumors.57
Although these data suggest that CCL2 and CCL5 play animportant role in recruiting monocytes into tumors, elevated CCchemokine expression is not found in all malignant tumors. Forexample, in prostate carcinoma, the number of CCL2 mRNA-expressing cells is significantly lower than in benign prostatichyperplasia. CCL2 transcripts were located in the fibromuscularstroma and basal cells of nonneoplastic glands but, in contrast toother tumors, not in cancer cells.58 In a separate study theexpression of CCL2 mRNA has also been shown to be reduced inprostate cancer compared with normal prostate,59 suggesting thatCCL2 may not be an influential factor in the recruitment ofmonocytes into prostate carcinoma.
Other CC chemokines may also be involved in monocyterecruitment to tumors. For example, a number of well-knownmonocyte attractants exist in both ovarian tumors and theirascites,60 including CCL3, CCL4, CCL8 (monocyte chemotacticprotein-2), and CCL22 (macrophage-derived chemokine).61 Thereis also evidence to show that CXC chemokines, particularlyCXCL8, are highly expressed in many human tumors and various
tumor cell lines.30 Although the involvement of CXC chemokinesin directing angiogenesis has been extensively researched,62,63 theirrole in directing the uptake of monocytes by tumors is less welldefined. Monocytes and monocyte-derived macrophages can ex-press low levels of CXCR1 and CXCR2, which bind CXCL8,64,65
and CXCL8 has been shown to mediate the adhesion of monocytesto endothelial cells under flow conditions,66 implying that thischemokine may play a role in monocyte recruitment by tumors.Although usually identified as a lymphocyte chemoattractant,monocytes and macrophages can also respond to the chemotacticeffects of CXCL12 (stromal-derived factor-1�) via their expressionof the receptors for this chemokine, CXCR4.67,68 CXCL12 isconstitutively expressed by a broad range of tissues and hasrecently been implicated in the metastasis of CXCR4-bearingtumor cells to specific organs that have elevated CXCL12 levels,particularly the lung, liver, and bone marrow.69 Although one studyshowed CXCL12 expression in biopsies of ovarian cancer,70 moststudies have found tumor cells to express either very little or noCXCL12,71-74 and thus this CXC chemokine is unlikely to play arole in the attraction of monocytes into tumors.
As well as chemokines, several cytokines have also beenimplicated in the recruitment of monocytes into tumors. One suchfactor is colony-stimulating factor-1 (CSF-1), which is produced bymany cell types, including monocytes and macrophages them-selves.75,76 CSF-1 has a major role in mononuclear phagocytebiology, affecting the growth, differentiation, and cell survival ofthese cells.77,78 It is also a potent chemoattractant for monocytesand macrophages.79 Elevated expression of CSF-1 and its receptor(CSF-1R) has been found in various types of human tumors (Table1), and the fact that CSF-1 attracts macrophages by binding toCSF-1R suggests that it may mediate macrophage recruitment intumors. In support of this theory, Dorsch and colleagues found that
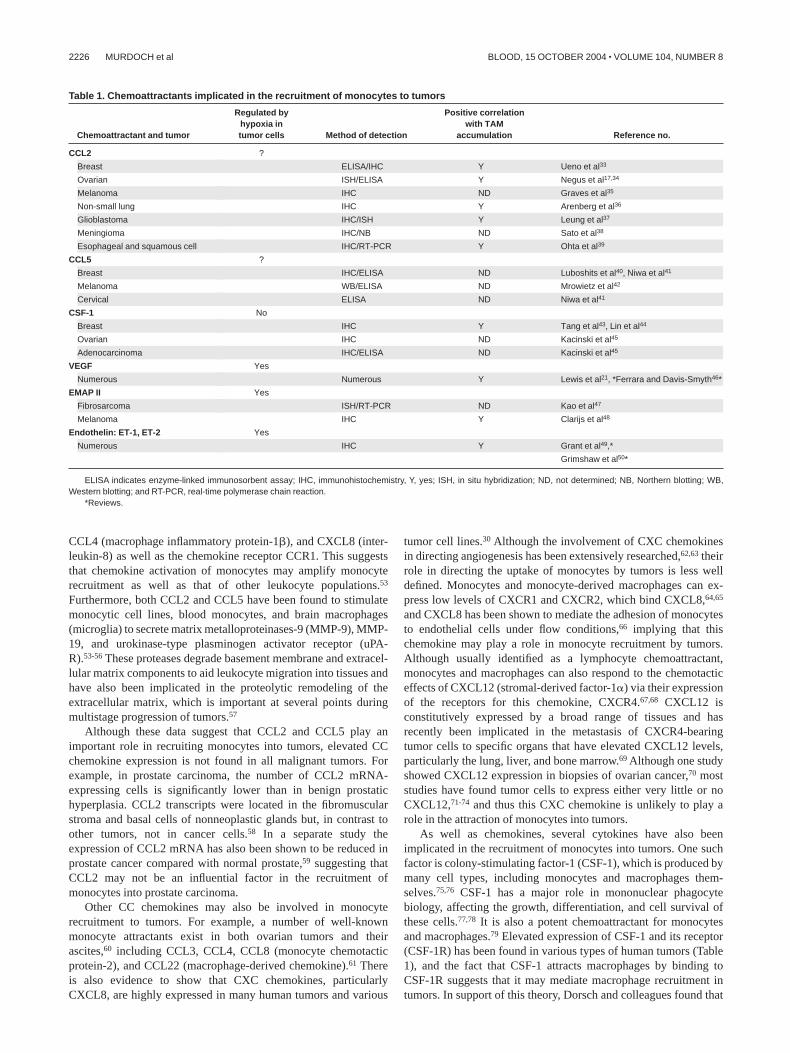
Table 1. Chemoattractants implicated in the recruitment of monocytes to tumors
Chemoattractant and tumor
Regulated byhypoxia intumor cells Method of detection
Positive correlationwith TAM
accumulation Reference no.
CCL2 ?
Breast ELISA/IHC Y Ueno et al33
Ovarian ISH/ELISA Y Negus et al17,34
Melanoma IHC ND Graves et al35
Non-small lung IHC Y Arenberg et al36
Glioblastoma IHC/ISH Y Leung et al37
Meningioma IHC/NB ND Sato et al38
Esophageal and squamous cell IHC/RT-PCR Y Ohta et al39
CCL5 ?
Breast IHC/ELISA ND Luboshits et al40, Niwa et al41
Melanoma WB/ELISA ND Mrowietz et al42
Cervical ELISA ND Niwa et al41
CSF-1 No
Breast IHC Y Tang et al43, Lin et al44
Ovarian IHC ND Kacinski et al45
Adenocarcinoma IHC/ELISA ND Kacinski et al45
VEGF Yes
Numerous Numerous Y Lewis et al21, *Ferrara and Davis-Smyth46*
EMAP II Yes
Fibrosarcoma ISH/RT-PCR ND Kao et al47
Melanoma IHC Y Clarijs et al48
Endothelin: ET-1, ET-2 Yes
Numerous IHC Y Grant et al49,*
Grimshaw et al50*
ELISA indicates enzyme-linked immunosorbent assay; IHC, immunohistochemistry, Y, yes; ISH, in situ hybridization; ND, not determined; NB, Northern blotting; WB,Western blotting; and RT-PCR, real-time polymerase chain reaction.
*Reviews.
2226 MURDOCH et al BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8

transplanted mouse tumors transfected with the CSF-1 geneexhibited an increase in TAM infiltration.80 Moreover, Tang et alfound a strong correlation between CSF-1 expression and TAMinfiltration in breast carcinomas.43 However, the most compellingevidence for CSF-1 regulating monocyte recruitment by tumors hascome from studies using CSF-1 knock-out mice. Lin and col-leagues crossed a transgenic mouse strain that spontaneouslydevelops mammary tumors with a CSF-1 knock-out mouse.44 Theyfound that monocyte infiltration into tumors was markedly re-duced, and this correlated with delayed tumor progression. Introduc-tion of CSF-1 by targeted gene expression produced a significantincrease in TAM levels in mammary tumors, leading to acceleratedtumor progression and metastasis.44
Another cytokine implicated in monocyte recruitment is theheparin-binding glycoprotein vascular endothelial growth factor(VEGF). This growth factor has potent mitogenic effects onendothelial cells and as such plays a major role in both physiologicand pathologic neovascularization. Indeed, VEGF is a key compo-nent of the angiogenic process, and elevated levels of this cytokineare a common feature of many human tumors and diseased tissues(Table 1).46,81 VEGF is also chemotactic for monocytes andmacrophages in vitro via activation of one form of the VEGFreceptor, VEGF-R1 (flt-1).82,83 Furthermore, macrophages frommice deficient in VEGF-R1 showed significantly reduced migra-tion in response to VEGF in a mouse model of embryonicangiogenesis.84 In a recent study, we observed a positive correlationbetween elevated VEGF expression and the number of infiltratingmacrophages in breast tumors,85 suggesting that high levels ofVEGF produced by tumors may attract monocytes into tumors and,more importantly, direct the movement of TAMs within tumors.
A less well-defined proinflammatory cytokine that chemoat-tracts monocytes and macrophages is endothelial monocyte-activating polypeptide II (EMAP II).86 Studies by Knies et alshowed that EMAP II mRNA and its precursor protein, pro-EMAPII, are expressed by many types of human tumor (Table 1), whereasthe mature cytokine is only present in the supernatant of dying cellsin vitro and at sites of apoptosis in the developing mouse embryo.87
This is because processing of the pro-form to the mature protein isdependent upon cleavage by proteases and the release of these isup-regulated during cell death.88 The release of mature EMAP II inthis way may aid the recruitment of macrophages to sites ofprogrammed cell death and necrosis where they are able toremove dead cells and necrotic debris.87 Because many tumorscontain multiple areas of necrosis and tumors with increasedlevels of necrosis contain increased numbers of TAMs,20 it ispossible that mature EMAP II may play a part in monocyterecruitment to some tumors.
Endothelins 1-3 (ET-1, -2, and -3) are small vasoactive andmitogenic peptides that are secreted by a wide array of cell typesand, like chemokines, mediate their effects by binding to 7-trans-membrane G protein–coupled receptors, ET-RA and ET-RB.89
Endothelins and their receptors are highly expressed in severaltypes of human tumor and human tumor cell lines in vitro49,90
(Table 1). ET-1 is chemotactic for human monocytes—a mecha-nism involving binding to ET-RA on these cells91—whereas ET-2 isa chemoattractant for macrophages by binding to ET-RB (which isnot expressed on monocytes, so they do not migrate towardET-2).50 These findings suggest that ET-1 may help to recruitmonocytes into tumors, whereas ET-2 may play a role in theirsubsequent localization within the tumor mass. Notably, much ofthe data linking the role of these chemoattractants to monocyte/macrophage recruitment into tumors has come from descriptive
studies correlating their expression with macrophage accumulationin various forms of human tumors. Studies utilizing knock-outmice or blocking the activity of these molecules with neutralizingantibodies are rare and are now warranted in order to determinewhether each molecule is essential in monocyte recruitment andTAM localization in vivo.
Although such chemoattractants are thought to attract mono-cytes to tumors, it is possible that these phagocytes are alsoattracted by cell debris released upon lysis from tumor cellsundergoing necrosis. Cell debris does not attract phagocytes inculture,92 suggesting that the intracellular components of cellsmay not attract monocytes or macrophages in vivo. However,this has yet to be fully investigated, and it is possible thatsoluble factors released from either apoptotic tumor cells, ordegradation of the local extracellular matrix as tumor cellsundergo necrosis, may attract such cells. For example, partiallydegraded collagen fragments have been found to be chemotacticfor monocytes and macrophages.93
TAM accumulation in hypoxic/necrotic areasin tumors
Various studies have thrown light on the mechanism used to attractor entrap TAMs in hypoxic regions of tumors. One mechanism isthe specific up-regulation of chemoattractants by hypoxia. Follow-ing extravasation into tumors, monocytes may migrate into hy-poxic areas following a hypoxically generated chemoattractantgradient. Indeed, hypoxic induction of various chemoattractants bytumor cells has been observed. Several studies have found elevatedlevels of VEGF in tumor cells and macrophages located inavascular and perinecrotic areas of human tumors.21,94,95 Further-more, a number of gene expression profiling studies have shownthat various tumor cell lines up-regulate VEGF in response tohypoxia.96-98 Moreover, expression of VEGF and HIF-1 (hypoxiainducible factor-1, the main transcription factor activated byhypoxia) colocalizes with hypoxic regions (as detected using thehypoxic cell marker, pimonidazole) in a murine model of Lewislung carcinoma.99 As mentioned previously, more TAMs arepresent in poorly vascularized, VEGF-positive areas of breastcarcinomas than well-vascularized, VEGF-negative sites, suggest-ing that VEGF may exert a chemotactic action on macrophages invivo and help to guide their migration into hypoxic tumor sites.21
Interestingly, Raleigh and colleagues found no correlation betweenhypoxia (using pimonidazole) and the expression of either VEGFor HIF-1 in human uterine or head and neck tumors, suggesting thatVEGF expression in some tumor types may not be solely regulatedby hypoxia.100,101
Matschurat and colleagues showed that high levels of pro-EMAP II expression in both methylcholanthrene (meth A) fibrosar-comas and B16 murine melanomas were found predominantly inperinecrotic areas of tumors, which are known to be hypoxic(Figure 1D). Pro-EMAP II in these areas could be converted to theactive form by proteases released from necrotic cell debris,102
suggesting a role for EMAP II in macrophage recruitment intothese sites. In fact, macrophages have been shown to colocalize toareas of EMAP II expression in uveal melanoma.48 Interestingly,elevated levels of mature EMAP II protein but not mRNA havebeen detected in the supernatants of hypoxic tumor cells in vitro.103
This suggests that such tumor cells can generate elevated levels ofactive EMAP II in response to hypoxia without the need for genetranscription and that this may represent a mechanism by which
RECRUITMENT OF MACROPHAGES INTO TUMORS 2227BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8

macrophages are rapidly recruited to perinecrotic, hypoxic areaswithin tumors. Definitive studies linking the expression of EMAPII and the recruitment of TAMs in hypoxic/necrotic tumor sites arenow warranted.
Endothelins, particularly ET-2, have also been shown to beregulated by hypoxia. Using a murine mammary tumor model,Grimshaw et al demonstrated endothelin ET-2 expression colocaliz-ing with hypoxic areas when using the hypoxia marker nitroimidaz-ole-L-yl-butyl theophylline (NITP).104 Furthermore, subsequent invitro studies using both murine and human breast tumor cell linesshowed hypoxic up-regulation of mRNA for ET-2, ET-RA, andET-RB in an HIF-1�–dependent manner.98,104 Interestingly, bind-ing of ET-1 to ovarian tumor cell lines triggers the activation andstabilization of HIF-1�, which then increases VEGF mRNA andprotein levels in these cells.105 One could therefore speculate thatblood monocytes are attracted to some tumors partly by elevatedlevels of ET-1. Furthermore, because macrophages express ET-RBit is possible that up-regulated ET-2– and ET-1–induced VEGFattracts TAMs into hypoxic tumor areas. In support of this,expression of ET-2– and ET-RB–positive macrophages was foundto colocalize in breast tumors.50,106 Recently, Cramer et al foundthat HIF-1� is crucial for macrophage accumulation in inflamma-tory sites such as the hypoxic synovia or arthritic joints in mice.107
It would be interesting to see whether ET-1 or ET-2 up-regulatesHIF-1� expression by monocytes or macrophages respectively inthe absence of hypoxia.
There is no evidence for hypoxic up-regulation of tumorcell–derived CSF-1 or CC chemokines, although recent data showthat THP-1 cells and murine macrophages down-regulate theexpression of CCL2 in response to hypoxia.108 In contrast, the CXCchemokine CXCL8 has been shown to be up-regulated by hypoxicconditions in several tumor cell types.109,110 Interestingly, produc-tion of CXCL12, which is not up-regulated in tumor cells byhypoxia, is elevated in synovial fibroblasts when cultured underhypoxic conditions.111 Moreover, CXCL12 induced the migrationof U937 cells into human synovial tissue transplanted into SCIDmice.112 These data suggest that CXCL12 may attract macro-phages, monocytes, and lymphocytes to hypoxic areas withinarthritic joints but not tumors. Recently, an alternative hypothesisfor the entrapment of TAMs has been postulated. Initially it wassuggested that, once recruited, TAMs are retained at hypoxic sitesdue to down-regulation of chemokine receptors or chemokines byTAMs or cancer cells, respectively.113,114 In these studies, TAMsisolated from ovarian carcinoma were found to display reducedlevels of CCR2 (the receptor for CCL2) mRNA and surfaceexpression and did not migrate in response to CCL2 in chemotaxisassays. The defect was specific for CCR2, as CCR1 and CCR5were expressed at similar levels in TAMs and monocyte-derivedmacrophages. Furthermore, neutralizing antibody studies sug-gested that defective CCR2 expression in TAMs was largelydependent on local TNF-� production at the tumor site.113 Thus,TNF-� produced in hypoxic areas may down-regulate TAM-expressed CCR2, which would then prevent TAM migration out ofthese areas. In addition, Negus and colleagues investigated theeffect of hypoxia on both CCL2 expression by human ovariantumor cell lines and monocyte migration in vitro. They found thathypoxia had no effect on basal CCL2 protein expression levels butdown-regulated TNF-�–induced CCL2 production by ovariancancer cells by approximately 25%.114 However, these levels arestill 15-fold higher than basal levels and thus still stimulatemonocyte chemotaxis even in the presence of hypoxia.114 Hypoxiaalso inhibited monocyte migration in response to CCL2 at oxygen
tensions of 1% or less, suggesting a specific oxygen-sensingmechanism was responsible for these effects.114 This study, how-ever, failed to examine the effect of hypoxia on TAM chemotaxis,which is the cell type most likely to encounter hypoxic conditionsin vivo.
Further experiments by Turner et al showed that hypoxia notonly inhibited CCL2 and other chemokine-induced migration ofthe promonocytic cell line, THP-1, and primary human macro-phages but also reduced their migration toward the bacterial-derived chemotactic factor fMLP (N-Formyl-Met-Leu-Phe), amonocyte chemoattractant that also signals via a G protein–coupled receptor. Furthermore, hypoxia did not down-regulateCCR2, and the inhibition of migration was not dependent onTNF-� or any other soluble factors.115 As a consequence, Turner etal proposed that inhibition of monocyte migration was due tohypoxia-induced metabolic changes within the cell. However,Grimshaw and Balkwill found that THP-1 cells incubated inreduced pH conditions or with inhibitors of either the electrontransport chain or mitochondria were still able to migrate toCCL2.116 Using RNA arbitrarily primed polymerase chain reaction(PCR) analysis, they found that out of several thousand genesscreened, only mitogen-activated protein kinase phosphatase 1(MKP-1) was consistently up-regulated by hypoxia.116 Althoughsimilar results have been reported using an oxygen-responsivepheochromocytoma cell line, which shows an 8-fold increase inmRNA and protein levels of MKP-1 in response to hypoxia,117
up-regulation of MKP-1 was not detected in microarray geneexpression profiling of hypoxic macrophages.23,118 This suggeststhere may be differences in the hypoxic regulation of MKP-1between different cell types and between primary macrophages andmonocytic cell lines. In light of these findings, it is important todetermine the precise role of MKP-1 in regulating TAM entrap-ment at hypoxic sites by using inhibitory RNA technology, genedeletion, or by generating MKP-1–deficient mice.
Nonetheless, these data present an interesting hypothesis as tohow TAMs might be retained in hypoxic areas of tumors. MKP-1attenuates both p44/p42 mitogen-activated protein kinase (MAPK)(extracellular signal-regulated kinase [ERK 1 and 2, respectively])and p38 MAPK activation and activity.119,120 Phosphorylation andthus activation of p38 MAPK and ERK1/2 are required forchemokine-mediated activation and migration of monocytes andmonocytic cell lines.121,122 These results suggest that dephosphory-lation of phosphorylated MAPK by MKP-1 is required for thehypoxic-induced inhibition of macrophage migration in response tochemokines and possibly other monocyte chemoattractants. More-over, up-regulation of MKP-1 was observed in both unstimulatedand stimulated cells, suggesting that hypoxic regulation of MKP-1is independent of cell activation status.116 Interestingly, TNF-� alsoinduces MKP-1 expression and, in conjunction with simulta-neously down-regulating CCR2 expression, rapidly inhibits mono-cyte migration in response to CCL2.116
At present, the mechanism of MKP-1 induction by hypoxia isnot known. Chemical activators of HIF-1 also inhibited chemokine-induced chemotaxis of THP-1 cells and up-regulated MKP-1 in apheochromocytoma cell line,116,117 suggesting that hypoxic activa-tion of HIF-1 may regulate MKP-1 activation. We have recentlyfound that transcripton factors Ets-1, nuclear factor–�B (NF�B),and activating transcription factor-4 (ATF-4) are also up-regulatedin primary macrophages by hypoxia (L. Elbarghati, C.M., andC.E.L., unpublished observations, January 2004), and so these mayalso be involved in regulating MKP-1 expression.
2228 MURDOCH et al BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8

The finding that activation of MKP-1 by hypoxia leads tochemotaxis arrest due to loss of chemotactic signal transductionmay have important implications for the entrapment of TAMs inhypoxic sites within tumors and diseased tissues by other hypoxi-cally regulated chemoattractants. For example, both VEGF andET-2 are chemotactic for macrophages in vitro via VEGF-R1 (flt-1)and ET-RB, respectively.50,82,83 Relatively little is known about theVEGF-R1 signal transduction pathway in macrophages; however,because other members of the VEGF-R family activate bothERK1/2 and p38 MAKP for chemotaxis,123 it is likely thatVEGF-R1 signals in a similar fashion. Similarly, ET-2–mediatedchemotaxis of macrophages also involves activation of the MAPKpathway, which is inhibited by both pertussis toxin and hypoxia.50
These data indicate that hypoxia may abrogate an ET-2– andVEGF-mediated signal transduction mechanism by up-regulatingMKP-1. Although there is little evidence to suggest that EMAP IIsignals via MAPK, this chemoattractant may also be subjected tothe same inhibitory process.
On first sight, the role of MKP-1 in inhibiting TAM migrationwithin hypoxic areas is appealing. However, several recent studieshave indicated that this process is more complex. Schioppa et alfound that hypoxia significantly increased the expression ofCXCR4 on peripheral blood monocytes, monocyte-derived macro-phages, TAMs, endothelial cells, and tumor cells. Furthermore, incontrast to CCR2/CCL2, the up-regulation of CXCR4 was found toenhance monocyte migration in response to CXCL12 in hypoxicconditions.124 Like many other hypoxia-responsive genes, CXCR4up-regulation is mediated in an HIF-1�–dependent manner.124,125
As with other chemokines, binding of CXCL12 to CXCR4elicits chemotaxis via activation of p38 MAPK and ERK.126 Theparadoxic finding that CCL2-mediated macrophage migration isinhibited by hypoxic induction of MKP-1, whereas CXCL12-mediated migration under hypoxic conditions is enhanced, isintriguing. Peripheral blood lymphocytes, which use similar chemo-tactic signal transduction mechanisms, migrate normally in re-sponse to CCL2 under hypoxic conditions.115 Furthermore, themigration of macrophages toward ET-2 is inhibited by hypoxiawhereas the migration of breast carcinoma cells is not.106 Thus, itappears that hypoxia may selectively and differentially modulatethe expression and function of chemoattractant receptors ondifferent cell types. It would be interesting to determine thechemotactic response of macrophages in response to simultaneousstimulation with CXCL12 and CCL2 in hypoxic conditions.
The relevance of elevated CXCR4 expression on monocytes,macrophages, and TAMs by hypoxia is questionable. To date moststudies have found that most tumor types do not expressCXCL12,71-74 suggesting that this chemokine is unlikely to becentrally involved in attracting TAMs into hypoxic tumor areas(although CXCL12 has been shown to be up-regulated by hypoxiain synovia fibroblasts, so hypoxic up-regulation of CXCR4 bymonocytes in the synovium may be important in rheumatoidarthritis). So why do TAMs up-regulate CXCR4 in hypoxic areas oftumors? One reason may be the important contribution thatCXCR4 appears to have in angiogenesis. For example, CXCR4-deficient mice show defects in the branching and remodeling ofcertain blood vessels,127 and expression of CXCR4 appears to becrucial for endothelial cell morphogenesis and angiogenesis.128 Theup-regulation of CXCR4 on endothelial cells along with othernumerous cell types by hypoxia might be part of an integratedresponse that allows both the generation of new blood vessels andthe remodeling of the existing circulation in order to alleviatehypoxic stress. Because TAMs significantly contribute to tumor
angiogenesis, elevated levels of TAM CXCR4 may have a moreimportant role in remodeling the tumor vasculature than directingTAM migration.
Thus, a common theme concerning the entrapment of TAMsat hypoxic sites in tumors is emerging. It appears that during theearly development of tumors, tumor cells secrete elevated levelsof CC chemokines and/or CSF-1, which then attract monocytesfrom the vasculature into the tumor. As the tumor progresses andareas of hypoxia/necrosis develop further, monocyte/TAM che-moattractants such as VEGF, endothelins, EMAP II, and possi-bly other hitherto undefined factors are liberated in a processthat is likely to require hypoxia inducible transcription factors,in particular HIF-1 and HIF-2, and possibly ATF-4, Ets-1, and/orNF�B. Together these factors act to attract monocytes intotumors and then direct TAMs toward areas of hypoxia/necrosisalong a chemoattractant gradient.
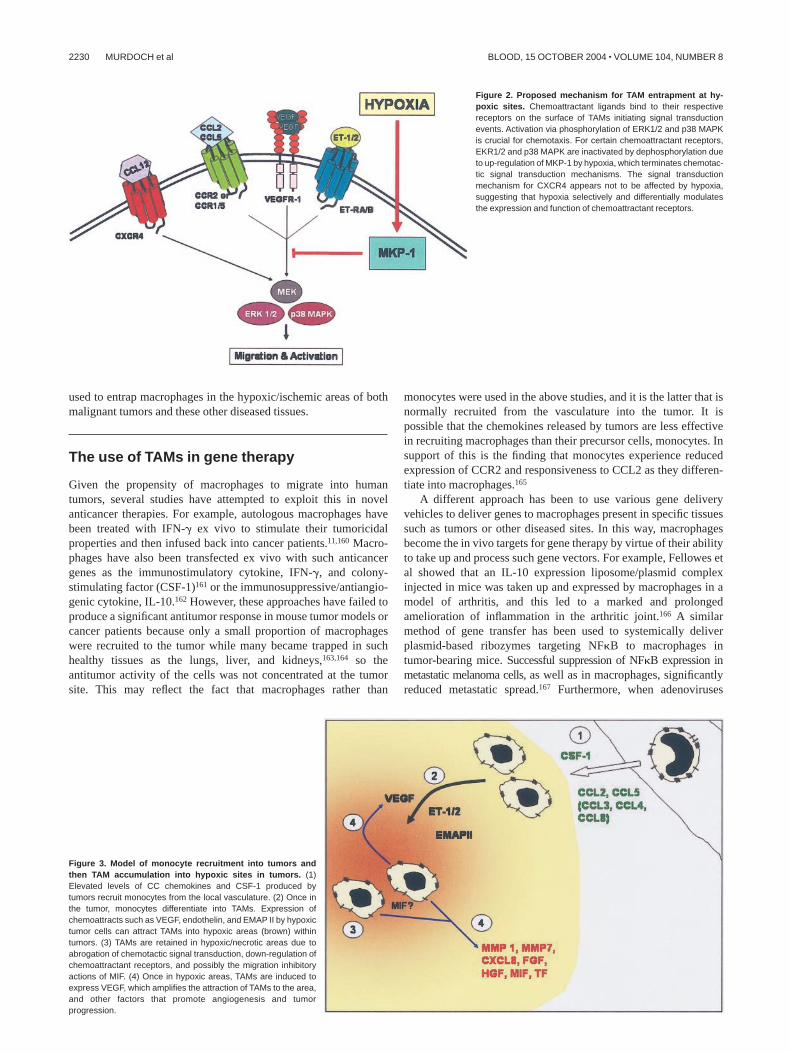
Although much further investigation is needed to understandthe puzzle of CCL2 and CXCL12 in the hypoxia-induced inhibition/stimulation of macrophage chemotaxis, a number of potentialmechanisms appear to be used to retain macrophages in hypoxicareas in tumors. A rapid, initial mechanism whereby hypoxiaand/or TNF-� inactivates MAPK, thus abrogating the intracellularsignaling cascade needed for migration to certain TAM chemoattrac-tant receptors, and a second, slower mechanism involving suchimmunomodulatory molecules as TNF-� and interferon-� (IFN-�),which get to down-regulate the expression of CCR2 and otherchemoattractant receptors113,129 in order to prevent migration ofTAMs away from the area (Figures 2 and 3).
To further add to the complexity, TAMs may also be trapped inhypoxic areas due to the actions of macrophage migration inhibi-tory factor (MIF), a cytokine that was originally found to inhibit therandom migration of macrophages.130,131 This cytokine is ubiqui-tously expressed by both immune and nonimmune cells, includingmacrophages132 and tumor cells,133 and has been shown to have arole in numerous disease states including tumor progression andneoplasia.134 The actions of MIF on macrophage migration aresomewhat controversial at present, because little in vitro or in vivoevidence has since been published to substantiate the originalfinding that MIF inhibits macrophage migration. Even so, Bacheret al showed strong MIF protein accumulation in close associationwith necrotic areas as well as in tumor cells surrounding bloodvessels in glioblastoma multiforme.135 Moreover, using microarraytechnology MIF has been shown to be up-regulated by hypoxia inglioblastoma, breast and squamous carcinoma cell lines, and inhuman macrophages,96-98,118 suggesting that MIF may play a role inTAM functions at these sites even if it is not involved with TAMentrapment per se.
Thus, macrophages may be trapped in hypoxic/necrotic areas oftumors by a combination of down-regulation of chemotactic signaltransduction mechanisms, chemoattractant receptors, and possiblyby the antichemotactic effects of MIF (Figure 3). As hypoxiaup-regulates VEGF release by macrophages,21,23,136 TAMs may beinduced in hypoxic areas to secrete VEGF and thus attract moreTAMs to these areas. The expression of proangiogenic cytokinesand enzymes would finally culminate in the onset of angiogenesis,revascularization, and thus the expansion of tumor cells in the area(Figure 3).
Importantly, elevated levels of the many of these chemoattrac-tants found in tumors have also been seen in other disease tissueslike atherosclerotic plaques and rheumatoid arthritic joints, inwhich macrophage accumulation in hypoxic areas has been ob-served (Table 2). Thus, it is possible that similar mechanisms are
RECRUITMENT OF MACROPHAGES INTO TUMORS 2229BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8
used to entrap macrophages in the hypoxic/ischemic areas of bothmalignant tumors and these other diseased tissues.
The use of TAMs in gene therapy
Given the propensity of macrophages to migrate into humantumors, several studies have attempted to exploit this in novelanticancer therapies. For example, autologous macrophages havebeen treated with IFN-� ex vivo to stimulate their tumoricidalproperties and then infused back into cancer patients.11,160 Macro-phages have also been transfected ex vivo with such anticancergenes as the immunostimulatory cytokine, IFN-�, and colony-stimulating factor (CSF-1)161 or the immunosuppressive/antiangio-genic cytokine, IL-10.162 However, these approaches have failed toproduce a significant antitumor response in mouse tumor models orcancer patients because only a small proportion of macrophageswere recruited to the tumor while many became trapped in suchhealthy tissues as the lungs, liver, and kidneys,163,164 so theantitumor activity of the cells was not concentrated at the tumorsite. This may reflect the fact that macrophages rather than
monocytes were used in the above studies, and it is the latter that isnormally recruited from the vasculature into the tumor. It ispossible that the chemokines released by tumors are less effectivein recruiting macrophages than their precursor cells, monocytes. Insupport of this is the finding that monocytes experience reducedexpression of CCR2 and responsiveness to CCL2 as they differen-tiate into macrophages.165
A different approach has been to use various gene deliveryvehicles to deliver genes to macrophages present in specific tissuessuch as tumors or other diseased sites. In this way, macrophagesbecome the in vivo targets for gene therapy by virtue of their abilityto take up and process such gene vectors. For example, Fellowes etal showed that an IL-10 expression liposome/plasmid complexinjected in mice was taken up and expressed by macrophages in amodel of arthritis, and this led to a marked and prolongedamelioration of inflammation in the arthritic joint.166 A similarmethod of gene transfer has been used to systemically deliverplasmid-based ribozymes targeting NF�B to macrophages intumor-bearing mice. Successful suppression of NF�B expression inmetastatic melanoma cells, as well as in macrophages, significantlyreduced metastatic spread.167 Furthermore, when adenoviruses
Figure 2. Proposed mechanism for TAM entrapment at hy-poxic sites. Chemoattractant ligands bind to their respectivereceptors on the surface of TAMs initiating signal transductionevents. Activation via phosphorylation of ERK1/2 and p38 MAPKis crucial for chemotaxis. For certain chemoattractant receptors,EKR1/2 and p38 MAPK are inactivated by dephosphorylation dueto up-regulation of MKP-1 by hypoxia, which terminates chemotac-tic signal transduction mechanisms. The signal transductionmechanism for CXCR4 appears not to be affected by hypoxia,suggesting that hypoxia selectively and differentially modulatesthe expression and function of chemoattractant receptors.
Figure 3. Model of monocyte recruitment into tumors andthen TAM accumulation into hypoxic sites in tumors. (1)Elevated levels of CC chemokines and CSF-1 produced bytumors recruit monocytes from the local vasculature. (2) Once inthe tumor, monocytes differentiate into TAMs. Expression ofchemoattracts such as VEGF, endothelin, and EMAP II by hypoxictumor cells can attract TAMs into hypoxic areas (brown) withintumors. (3) TAMs are retained in hypoxic/necrotic areas due toabrogation of chemotactic signal transduction, down-regulation ofchemoattractant receptors, and possibly the migration inhibitoryactions of MIF. (4) Once in hypoxic areas, TAMs are induced toexpress VEGF, which amplifies the attraction of TAMs to the area,and other factors that promote angiogenesis and tumorprogression.
2230 MURDOCH et al BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8

encoding the heat shock protein heme oxygenase 1 (HO-1) wereadministered by intratracheal inoculation into a murine model ofacute lung injury caused by influenza virus, HO-1 was found to beexpressed by alveolar macrophages and respiratory epithelial cells,which enhanced survival and decreased inflammatory cells in thelung.168,169 However, the disadvantage with this type of therapy isthe lack of specificity of the liposome/plasmid complex, becausemacrophages in nondiseased tissues and indeed other cell types caninternalize and express the therapeutic gene.
The finding that TAMs accumulate in hypoxic areas in tumorsled us to propose that macrophages might also be used as deliveryvehicles to target gene therapy to these otherwise largely inacces-sible areas of tumors. In this novel approach, autologous mono-cytes would be isolated from a given patient, differentiated intomacrophages ex vivo, transfected with a hypoxia-activated thera-peutic gene, and then reinfused back into the patient.170 Thetransfected macrophages would then be taken up from the blood-stream into the primary tumor (as well as any secondary tumorspresent elsewhere in the body) and accumulate in hypoxic tumorareas, where they would express the therapeutic gene.
So far we have shown that when macrophages transfected witha hypoxia-regulated gene for the prodrug-activating enzyme,cytochrome, P450—cocultured in vitro with breast tumor multicel-lular spheroids (small 3-D tumor masses grown in vitro from breasttumor cell lines)—rapidly migrated into them and expressed theP450 enzyme. When these spheroids were then exposed to theprodrug, cyclophosphamide, the P450 enzyme expressed by hy-poxic macrophages in the multicellular tumor spheroids convertedthe prodrug into its active, cytotoxic metabolite. This then diffusedout of transfected macrophages (which are nondividing cells andthus refractory to its effects) and was intercalated into the DNA ofsurrounding tumor cells, causing cell death during their subsequentmitosis. This novel approach combines 2 levels of tumor targeting—the ability of macrophages to “home” to the hypoxic areas oftumors with the hypoxic activation of the therapeutic gene.170
Although the in vivo efficacy of this novel approach has yet to betested, reintroduction of transfected macrophages back into tumorpatients is also likely to meet with limited success unless ways ofenhancing macrophage uptake specifically by tumors are used. Wehope, now that we have a more comprehensive picture of the
chemoattractants involved in the uptake of monocytes by tumors, itmay be possible to up-regulate the responsiveness of such therapeu-tically “armed” macrophages to tumor-derived signals.
Conclusion
This review has indicated that monocyte recruitment into tumors isa complex process involving a number of molecules. As mentionedpreviously, the recruitment of macrophages to sites of hypoxia isnot restricted to tumors but also occurs in various other diseases.This begs the question of whether it might be possible to exploit ourknowledge of how macrophages are recruited/trapped in thesehypoxic/necrotic areas to enhance the uptake of therapeuticallyarmed macrophages (ie, bearing hypoxically activated therapeuticDNA constructs) to such tissues. For example, it may be sufficientto up-regulate expression of receptors for just one of thesemolecules, even if it is not the central one, in the hypoxicaccumulation of macrophages. The expression of these chemoattrac-tants may be temporally and/or spatially regulated in order torecruit and retain macrophages in these areas. However, the preciserole that each of these chemoattractants plays in this part of theprocess and, more importantly, how they cooperate with oneanother remains largely unknown.
Alternatively, new therapies involving the use of peptides orsmall molecule antagonists to inhibit macrophage receptors forspecific chemoattractants produced by hypoxic areas of tumorsmay help to reduce macrophage accumulation in these sites whileleaving macrophage functions in healthy tissues unaffected. Thiswould then remove their proangiogenic contribution under hypoxiaand reduce the net proangiogenic activity of cells in such areas.
Acknowledgments
The authors thank James Raleigh, University of North CarolinaSchool of Medicine, for donation of pimonidazole-labeled tumorsections, and Elizabeth Macrae, University of Sheffield, UnitedKingdom, for the murine PC3 prostate carcinoma sections.
References
1. Ross JA. The biology of the macrophage. In:Burke B, Lewis CE, eds. The Macrophage. Ox-ford, United Kingdom: Oxford University Press;2002:P1-P72.
2. Kelly PM, Davison RS, Bliss E, McGee JO. Mac-rophages in human breast disease: a quantitativeimmunohistochemical study. Br J Cancer. 1988;57:174-177.
3. Leek RD, Harris AL, Lewis CE. Cytokine net-works in solid human tumors: regulation of angio-genesis. J Leukoc Biol. 1994;56:423-435.
4. Yamashiro S, Takeya M, Nishi T, et al. Tumor-derived monocyte chemoattractant protein-1induces intratumoral infiltration of monocyte-derived macrophage subpopulation in transplantedrat tumors. Am J Pathol. 1994;145:856-867.
5. Pollard JW. Tumour-educated macrophages pro-mote tumour progression and metastasis. NatRev Cancer. 2004;4:71-78.
6. Mantovani A, Sozzani S, Locati M, et al. Infiltra-tion of tumours by macrophages and dendriticcells: tumour-associated macrophages as a para-digm for polarized M2 mononuclear phagocytes.Novartis Found Symp. 2004;256:137-145.
Table 2. Evidence for elevated levels of certain chemoattractants in ischemic areas of diseases where macrophages have been foundto accumulate in vivo
Dermal sounds Atherosclerosis Rheumatoid arthritisProliferativeretinopathy
CC chemokines Y (Heinrich et al137) Y (Burke-Gaffney et al138,
Linton and Fazio139)
Y (DeVries et al140, Bodolay et al141) Y (Capeans et al142,
Abu el-Asrar et al143)
CXCL12 ND ND Y (Pufe et al144) ND
VEGF Y (Dvorak et al145) Y (Inoue et al146) Y (Pufe et al144, Afuwape et al147) Y (Ishida et al148)
CSF-1 Y (Wu et al149) Y (Clinton et al150, Qiao et al151) Y (Seitz et al152) Y (Elner et al153)
EMAP II ND ND ND ND
Endothelin Y (Black et al154, Lipa et al155) Y (Fan et al156, Best et al157) Y (Haq et al158) Y (Oku et al159)
Data in parentheses are references.Y indicates elevated levels observed; and ND, not determined.
RECRUITMENT OF MACROPHAGES INTO TUMORS 2231BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8

7. Leek RD, Harris AL. Tumor-associated macro-phages in breast cancer. J Mammary Gland BiolNeoplasia. 2002;7:177-189.
8. Blankenstein T. The role of inflammation in tu-mour growth and tumour suppression. NovartisFound Symp. 2004;256:205-210.
9. Blankenstein T, Qin ZH, Uberla K, et al. Tumorsuppression after tumor cell-targeted tumor ne-crosis factor alpha gene transfer. J Exp Med.1991;173:1047-1052.
10. Mantovani A, Sozzani S, Locati M, Allavena P,Sica A. Macrophage polarization: tumor-associ-ated macrophages as a paradigm for polarizedM2 mononuclear phagocytes. Trends Immunol.2002;23:549-555.
11. Bingle L, Brown NJ, Lewis CE. The role of tu-mour-associated macrophages in tumour pro-gression: implications for new anticancer thera-pies. J Pathol. 2002;196:254-265.
12. Vaupel P, Kallinowski F, Okunieff P. Blood flow,oxygen and nutrient supply, and metabolic micro-environment of human tumors: a review. CancerRes. 1989;49:6449-6465.
13. Raleigh JA, Chou SC, Bono EL, Thrall DE, VariaMA. Semiquantitative immunohistochemical anal-ysis for hypoxia in human tumors. Int J RadiatOncol Biol Phys. 2001;49:569-574.
14. Brown JM, Giaccia AJ. The unique physiology ofsolid tumors: opportunities (and problems) forcancer therapy. Cancer Res. 1998;58:1408-1416.
15. Brown JM. Exploiting the hypoxic cancer cell:mechanisms and therapeutic strategies. Mol MedToday. 2000;6:157-162.
16. Vaupel P, Kelleher DK, Hockel M. Oxygen statusof malignant tumors: pathogenesis of hypoxiaand significance for tumor therapy. Semin Oncol.2001;28:29-35.
17. Negus RP, Stamp GW, Hadley J, Balkwill FR.Quantitative assessment of the leukocyte infil-trate in ovarian cancer and its relationship to theexpression of C-C chemokines. Am J Pathol.1997;150:1723-1734.
18. Silzle T, Kreutz M, Dobler MA, Brockhoff G,Knuechel R, Kunz S. Tumor-associated fibro-blasts recruit blood monocytes into tumor tissue.Eur J Immunol. 2003;33:1311-1320.
19. Leek RD, Lewis CE, Whitehouse R, Greenall M,Clarke J, Harris AL. Association of macrophageinfiltration with angiogenesis and prognosis ininvasive breast carcinoma. Cancer Res. 1996;56:4625-4629.
20. Leek RD, Landers RJ, Harris AL, Lewis CE. Ne-crosis correlates with high vascular density andfocal macrophage infiltration in invasive carci-noma of the breast. Br J Cancer. 1999;79:991-995.
21. Lewis JS, Landers RJ, Underwood JC, Harris AL,Lewis CE. Expression of vascular endothelialgrowth factor by macrophages is up-regulated inpoorly vascularized areas of breast carcinomas.J Pathol. 2000;192:150-158.
22. Onita T, Ji PG, Xuan JW, et al. Hypoxia-induced,perinecrotic expression of endothelial Per-ARNT-Sim domain protein-1/hypoxia-inducible factor-2alpha correlates with tumor progression, vascu-larization, and focal macrophage infiltration inbladder cancer. Clin Cancer Res. 2002;8:471-480.
23. Burke B, Giannoudis A, Corke KP, et al. Hypoxia-induced gene expression in human macro-phages: implications for ischemic tissues andhypoxia-regulated gene therapy. Am J Pathol.2003;163:1233-1243.
24. Crowther M, Brown NJ, Bishop ET, Lewis CE.Microenvironmental influence on macrophageregulation of angiogenesis in wounds and malig-nant tumors. J Leukoc Biol. 2001;70:478-490.
25. Bjornheden T, Levin M, Evaldsson M, Wiklund O.Evidence of hypoxic areas within the arterial wallin vivo. Arterioscler Thromb Vasc Biol. 1999;19:870-876.
26. Hollander AP, Corke KP, Freemont AJ, Lewis CE.Expression of hypoxia-inducible factor 1alpha bymacrophages in the rheumatoid synovium: impli-cations for targeting of therapeutic genes to theinflamed joint. Arthritis Rheum. 2001;44:1540-1544.
27. Esser P, Heimann K, Wiedemann P. Macro-phages in proliferative vitreoretinopathy and pro-liferative diabetic retinopathy: differentiation ofsubpopulations. Br J Ophthalmol. 1993;77:731-733.
28. Zlotnik A, Yoshie O. Chemokines: a new classifi-cation system and their role in immunity. Immu-nity. 2000;12:121-127.
29. Murdoch C, Finn A. Chemokine receptors andtheir role in inflammation and infectious diseases.Blood. 2000;95:3032-3043.
30. Balkwill F. Chemokine biology in cancer. SeminImmunol. 2003;15:49-55.
31. Vicari AP, Caux C. Chemokines in cancer. Cyto-kine Growth Factor Rev. 2002;13:143-154.
32. Strieter RM, Belperio JA, Phillips RJ, Keane MP.Chemokines: angiogenesis and metastases inlung cancer. Novartis Found Symp. 2004;256:173-184.
33. Ueno T, Toi M, Saji H, et al. Significance of mac-rophage chemoattractant protein-1 in macro-phage recruitment, angiogenesis, and survival inhuman breast cancer. Clin Cancer Res. 2000;6:3282-3289.
34. Negus RP, Stamp GW, Relf MG, et al. The detec-tion and localization of monocyte chemoattractantprotein-1 (MCP-1) in human ovarian cancer.J Clin Invest. 1995;95:2391-2396.
35. Graves DT, Barnhill R, Galanopoulos T, Antonia-des HN. Expression of monocyte chemotacticprotein-1 in human melanoma in vivo. Am JPathol. 1992;140:9-14.
36. Arenberg DA, Keane MP, DiGiovine B, et al. Mac-rophage infiltration in human non-small-cell lungcancer: the role of CC chemokines. Cancer Im-munol Immunother. 2000;49:63-70.
37. Leung SY, Wong MP, Chung LP, Chan AS, YuenST. Monocyte chemoattractant protein-1 expres-sion and macrophage infiltration in gliomas. ActaNeuropathol (Berl). 1997;93:518-527.
38. Sato K, Kuratsu J, Takeshima H, Yoshimura T,Ushio Y. Expression of monocyte chemoattrac-tant protein-1 in meningioma. J Neurosurg. 1995;82:874-878.
39. Ohta M, Kitadai Y, Tanaka S, et al. Monocyte che-moattractant protein-1 expression correlates withmacrophage infiltration and tumor vascularity inhuman esophageal squamous cell carcinomas.Int J Cancer. 2002;102:220-224.
40. Luboshits G, Shina S, Kaplan O, et al. Elevatedexpression of the CC chemokine regulated onactivation, normal T cell expressed and secreted(RANTES) in advanced breast carcinoma. Can-cer Res. 1999;59:4681-4687.
41. Niwa Y, Akamatsu H, Niwa H, Sumi H, Ozaki Y,Abe A. Correlation of tissue and plasma RANTESlevels with disease course in patients with breastor cervical cancer. Clin Cancer Res. 2001;7:285-289.
42. Mrowietz U, Schwenk U, Maune S, et al. The che-mokine RANTES is secreted by human mela-noma cells and is associated with enhanced tu-mour formation in nude mice. Br J Cancer. 1999;79:1025-1031.
43. Tang RP, Kacinski B, Validire P, et al. Oncogeneamplification correlates with dense lymphocyteinfiltration in human breast cancers: a role for he-matopoietic growth factor release by tumor cells?J Cell Biochem. 1990;44:189-198.
44. Lin EY, Nguyen AV, Russell RG, Pollard JW.Colony-stimulating factor 1 promotes progressionof mammary tumors to malignancy. J Exp Med.2001;193:727-740.
45. Kacinski BM. CSF-1 and its receptor in ovarian,
endometrial and breast cancer. Ann Med. 1995;27:79-85.
46. Ferrara N, Davis-Smyth T. The biology of vascu-lar endothelial growth factor. Endocr Rev. 1997;18:4-25.
47. Kao J, Ryan J, Brett G, et al. Endothelial mono-cyte-activating polypeptide II. A novel tumor-derived polypeptide that activates host-responsemechanisms. J Biol Chem. 1992;267:20239-20247.
48. Clarijs R, Schalkwijk L, Ruiter DJ, de W. EMAP-IIexpression is associated with macrophage accu-mulation in primary uveal melanoma. Invest Oph-thalmol Vis Sci. 2003;44:1801-1806.
49. Grant K, Loizidou M, Taylor I. Endothelin-1: amultifunctional molecule in cancer. Br J Cancer.2003;88:163-166.
50. Grimshaw MJ, Wilson JL, Balkwill FR. Endothe-lin-2 is a macrophage chemoattractant: implica-tions for macrophage distribution in tumors. EurJ Immunol. 2002;32:2393-2400.
51. Bottazzi B, Walter S, Govoni D, Colotta F, Man-tovani A. Monocyte chemotactic cytokine genetransfer modulates macrophage infiltration,growth, and susceptibility to IL-2 therapy of a mu-rine melanoma. J Immunol. 1992;148:1280-1285.
52. Nesbit M, Schaider H, Miller TH, Herlyn M. Low-level monocyte chemoattractant protein-1 stimu-lation of monocytes leads to tumor formation innontumorigenic melanoma cells. J Immunol.2001;166:6483-6490.
53. Locati M, Deuschle U, Massardi ML, et al. Analy-sis of the gene expression profile activated by theCC chemokine ligand 5/RANTES and by lipopoly-saccharide in human monocytes. J Immunol.2002;168:3557-3562.
54. Robinson SC, Scott KA, Balkwill FR. Chemokinestimulation of monocyte matrix metalloprotein-ase-9 requires endogenous TNF-alpha. Eur J Im-munol. 2002;32:404-412.
55. Cross AK, Woodroofe MN. Chemokine modula-tion of matrix metalloproteinase and TIMP pro-duction in adult rat brain microglia and a humanmicroglial cell line in vitro. Glia. 1999;28:183-189.
56. Azenshtein E, Luboshits G, Shina S, et al. TheCC chemokine RANTES in breast carcinoma pro-gression: regulation of expression and potentialmechanisms of promalignant activity. CancerRes. 2002;62:1093-1102.
57. Coussens LM, Werb Z. Matrix metalloproteinasesand the development of cancer. Chem Biol. 1996;3:895-904.
58. Mazzucchelli L, Loetscher P, Kappeler A, et al.Monocyte chemoattractant protein-1 gene ex-pression in prostatic hyperplasia and prostateadenocarcinoma. Am J Pathol. 1996;149:501-509.
59. Chetcuti A, Margan S, Mann S, et al. Identificationof differentially expressed genes in organ-con-fined prostate cancer by gene expression array.Prostate. 2001;47:132-140.
60. Scotton C, Milliken D, Wilson J, Raju S, Balkwill F.Analysis of CC chemokine and chemokine recep-tor expression in solid ovarian tumours. Br J Can-cer. 2001;85:891-897.
61. Milliken D, Scotton C, Raju S, Balkwill F, Wilson J.Analysis of chemokines and chemokine receptorexpression in ovarian cancer ascites. Clin CancerRes. 2002;8:1108-1114.
62. Belperio JA, Keane MP, Arenberg DA, et al. CXCchemokines in angiogenesis. J Leukoc Biol.2000;68:1-8.
63. Salcedo R, Oppenheim JJ. Role of chemokines inangiogenesis: CXCL12/SDF-1 and CXCR4 inter-action, a key regulator of endothelial cell re-sponses. Microcirculation. 2003;10:359-370.
64. Patel L, Charlton SJ, Chambers JK, MacpheeCH. Expression and functional analysis of che-mokine receptors in human peripheral blood leu-kocyte populations. Cytokine. 2001;14:27-36.
2232 MURDOCH et al BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8

65. Moser B, Barella L, Mattei S, et al. Expression oftranscripts for two interleukin 8 receptors in hu-man phagocytes, lymphocytes and melanomacells. Biochem J. 1993;294(pt 1):285-292.
66. Gerszten RE, Garcia-Zepeda EA, Lim YC, et al.MCP-1 and IL-8 trigger firm adhesion of mono-cytes to vascular endothelium under flow condi-tions. Nature. 1999;398:718-723.
67. Di Marzio, Tse J, Landau NR. Chemokine recep-tor regulation and HIV type 1 tropism in mono-cyte-macrophages. AIDS Res Hum Retroviruses.1998;14:129-138.
68. Wang J, Guan E, Roderiquez G, Calvert V, Al-varez R, Norcross MA. Role of tyrosine phos-phorylation in ligand-independent sequestrationof CXCR4 in human primary monocytes-macro-phages. J Biol Chem. 2001;276:49236-49243.
69. Muller A, Homey B, Soto H, et al. Involvement ofchemokine receptors in breast cancer metasta-sis. Nature. 2001;410:50-56.
70. Scotton CJ, Wilson JL, Scott K, et al. Multiple ac-tions of the chemokine CXCL12 on epithelial tu-mor cells in human ovarian cancer. Cancer Res.2002;62:5930-5938.
71. Begum NA, Coker A, Shibuta K, et al. Loss ofhIRH mRNA expression from premalignant ad-enomas and malignant cell lines. Biochem Bio-phys Res Commun. 1996;229:864-868.
72. Shibuta K, Begum NA, Mori M, Shimoda K, Akiyo-shi T, Barnard GF. Reduced expression of theCXC chemokine hIRH/SDF-1alpha mRNA inhepatoma and digestive tract cancer. Int J Can-cer. 1997;73:656-662.
73. Schrader AJ, Lechner O, Templin M, et al.CXCR4/CXCL12 expression and signalling in kid-ney cancer. Br J Cancer. 2002;86:1250-1256.
74. Phillips RJ, Burdick MD, Lutz M, Belperio JA,Keane MP, Strieter RM. The stromal derived fac-tor-1/CXCL12-CXC chemokine receptor 4 biologi-cal axis in non-small cell lung cancer metastases.Am J Respir Crit Care Med. 2003;167:1676-1686.
75. Kawasaki ES, Ladner MB. Molecular biology ofmacrophage colony-stimulating factor. ImmunolSer. 1990;49:155-176.
76. Rambaldi A, Young DC, Griffin JD. Expression ofthe M-CSF (CSF-1) gene by human monocytes.Blood. 1987;69:1409-1413.
77. Stanley ER, Guilbert LJ, Tushinski RJ, BartelmezSH. CSF-1—a mononuclear phagocyte lineage-specific hemopoietic growth factor. J Cell Bio-chem. 1983;21:151-159.
78. Csar XF, Wilson NJ, McMahon KA, et al. Pro-teomic analysis of macrophage differentiation.p46/52(Shc) Tyrosine phosphorylation is requiredfor CSF-1-mediated macrophage differentiation.J Biol Chem. 2001;276:26211-26217.
79. Wang JM, Griffin JD, Rambaldi A, Chen ZG, Man-tovani A. Induction of monocyte migration by re-combinant macrophage colony-stimulating factor.J Immunol. 1988;141:575-579.
80. Dorsch M, Hock H, Kunzendorf U, DiamantsteinT, Blankenstein T. Macrophage colony-stimulatingfactor gene transfer into tumor cells induces mac-rophage infiltration but not tumor suppression.Eur J Immunol. 1993;23:186-190.
81. Adams J, Carder PJ, Downey S, et al. Vascularendothelial growth factor (VEGF) in breast can-cer: comparison of plasma, serum, and tissueVEGF and microvessel density and effects of ta-moxifen. Cancer Res. 2000;60:2898-2905.
82. Barleon B, Sozzani S, Zhou D, Weich HA, Man-tovani A, Marme D. Migration of human mono-cytes in response to vascular endothelial growthfactor (VEGF) is mediated via the VEGF receptorflt-1. Blood. 1996;87:3336-3343.
83. Sawano A, Iwai S, Sakurai Y, et al. Flt-1, vascularendothelial growth factor receptor 1, is a novelcell surface marker for the lineage of monocyte-macrophages in humans. Blood. 2001;97:785-791.
84. Hiratsuka S, Minowa O, Kuno J, Noda T, ShibuyaM. Flt-1 lacking the tyrosine kinase domain is suf-ficient for normal development and angiogenesisin mice. Proc Natl Acad Sci U S A. 1998;95:9349-9354.
85. Leek RD, Hunt NC, Landers RJ, Lewis CE,Royds JA, Harris AL. Macrophage infiltration isassociated with VEGF and EGFR expression inbreast cancer. J Pathol. 2000;190:430-436.
86. Kao J, Houck K, Fan Y, et al. Characterization ofa novel tumor-derived cytokine. Endothelial-monocyte activating polypeptide II. J Biol Chem.1994;269:25106-25119.
87. Knies UE, Kroger S, Clauss M. Expression ofEMAP II in the developing and adult mouse. Apo-ptosis. 2000;5:141-151.
88. Knies UE, Behrensdorf HA, Mitchell CA, et al.Regulation of endothelial monocyte-activatingpolypeptide II release by apoptosis. Proc NatlAcad Sci U S A. 1998;95:12322-12327.
89. Masaki T, Ninomiya H, Sakamoto A, Okamoto Y.Structural basis of the function of endothelin re-ceptor. Mol Cell Biochem. 1999;190:153-156.
90. Kusuhara M, Yamaguchi K, Nagasaki K, et al.Production of endothelin in human cancer celllines. Cancer Res. 1990;50:3257-3261.
91. Cui P, Tani K, Kitamura H, et al. A novel bioactive31-amino acid endothelin-1 is a potent chemotac-tic peptide for human neutrophils and monocytes.J Leukoc Biol. 2001;70:306-312.
92. Bessis M, de Boisfleury C. Facts and speculationabout necrotaxis (chemotaxis toward a dyingcell). Blood Cells. 1984;10:5-22.
93. Laskin DL, Soltys RA, Berg RA, Riley DJ. Activa-tion of alveolar macrophages by native and syn-thetic collagen-like polypeptides. Am J RespirCell Mol Biol. 1994;10:58-64.
94. Lee AH, Dublin EA, Bobrow LG, Poulsom R. Inva-sive lobular and invasive ductal carcinoma of thebreast show distinct patterns of vascular endothe-lial growth factor expression and angiogenesis.J Pathol. 1998;185:394-401.
95. Brown LF, Berse B, Jackman RW, et al. Expres-sion of vascular permeability factor (vascular en-dothelial growth factor) and its receptors in breastcancer. Hum Pathol. 1995;26:86-91.
96. Lal A, Peters H, Croix B, et al. Transcriptional re-sponse to hypoxia in human tumors. J Natl Can-cer Inst. 2001;93:1337-1343.
97. Bando H, Toi M, Kitada K, Koike M. Genes com-monly upregulated by hypoxia in human breastcancer cells MCF-7 and MDA-MB-231. BiomedPharmacother. 2003;57:333-340.
98. Koong AC, Denko NC, Hudson KM, et al. Candi-date genes for the hypoxic tumor phenotype.Cancer Res. 2000;60:883-887.
99. Kim MS, Kwon HJ, Lee YM, et al. Histonedeacetylases induce angiogenesis by negativeregulation of tumor suppressor genes. Nat Med.2001;7:437-443.
100. Raleigh JA, Calkins A, Rinker LH, et al. Hypoxiaand vascular endothelial growth factor expressionin human squamous cell carcinomas using pi-monidazole as a hypoxia marker. Cancer Res.1998;58:3765-3768.
101. Janssen HL, Haustermans KM, Sprong D, et al.HIF-1A, pimonidazole, and iododeoxyuridine toestimate hypoxia and perfusion in human head-and-neck tumors. Int J Radiat Oncol Biol Phys.2002;54:1537-1549.
102. Zhang FR, Schwarz MA. Pro-EMAP II is not pri-marily cleaved by caspase-3 and -7. Am J PhysiolLung Cell Mol Physiol. 2002;282:L1239-L1244.
103. Matschurat S, Knies UE, Person V, et al. Regula-tion of EMAP II by hypoxia. Am J Pathol. 2003;162:93-103.
104. Grimshaw MJ, Naylor S, Balkwill FR. Endothe-lin-2 is a hypoxia-induced autocrine survival fac-tor for breast tumor cells. Mol Cancer Ther. 2002;1:1273-1281.
105. Spinella F, Rosano L, Di C, Natali PG, Bagnato A.Endothelin-1 induces vascular endothelial growthfactor by increasing hypoxia-inducible factor-1alpha in ovarian carcinoma cells. J Biol Chem.2002;277:27850-27855.
106. Grimshaw MJ, Hagemann T, Ayhan A, Gillett CE,Binder C, Balkwill FR. A role for endothelin-2 andits receptors in breast tumor cell invasion. CancerRes. 2004;64:2461-2468.
107. Cramer T, Yamanishi Y, Clausen BE, et al. HIF-1alpha is essential for myeloid cell-mediated in-flammation. Cell. 2003;112:645-657.
108. Bosco MC, Puppo M, Pastorino S, et al. Hypoxiaselectively inhibits monocyte chemoattractantprotein-1 production by macrophages. J Immu-nol. 2004;172:1681-1690.
109. Xu L, Xie K, Mukaida N, Matsushima K, FidlerIJ. Hypoxia-induced elevation in interleukin-8 ex-pression by human ovarian carcinoma cells. Can-cer Res. 1999;59:5822-5829.
110. Shi Q, Xiong Q, Le X, Xie K. Regulation of inter-leukin-8 expression by tumor-associated stressfactors. J Interferon Cytokine Res. 2001;21:553-566.
111. Hitchon C, Wong K, Ma G, Reed J, Lyttle D, El-Gabalawy H. Hypoxia-induced production of stro-mal cell-derived factor 1 (CXCL12) and vascularendothelial growth factor by synovial fibroblasts.Arthritis Rheum. 2002;46:2587-2597.
112. Blades MC, Ingegnoli F, Wheller SK, et al. Stro-mal cell-derived factor 1 (CXCL12) inducesmonocyte migration into human synovium trans-planted onto SCID mice. Arthritis Rheum. 2002;46:824-836.
113. Sica A, Saccani A, Bottazzi B, et al. Defective ex-pression of the monocyte chemotactic protein-1receptor CCR2 in macrophages associated withhuman ovarian carcinoma. J Immunol. 2000;164:733-738.
114. Negus RP, Turner L, Burke F, Balkwill FR. Hyp-oxia down-regulates MCP-1 expression: implica-tions for macrophage distribution in tumors.J Leukoc Biol. 1998;63:758-765.
115. Turner L, Scotton C, Negus R, Balkwill F. Hypoxiainhibits macrophage migration. Eur J Immunol.1999;29:2280-2287.
116. Grimshaw MJ, Balkwill FR. Inhibition of monocyteand macrophage chemotaxis by hypoxia and in-flammation—a potential mechanism. Eur J Immu-nol. 2001;31:480-489.
117. Seta KA, Kim R, Kim HW, Millhorn DE, BeitnerJ. Hypoxia-induced regulation of MAPK phospha-tase-1 as identified by subtractive suppressionhybridization and cDNA microarray analysis.J Biol Chem. 2001;276:44405-44412.
118. White JR, Harris RA, Lee SR, et al. Genetic am-plification of the transcriptional response to hyp-oxia as a novel means of identifying regulators ofangiogenesis. Genomics. 2004;83:1-8.
119. Sun H, Charles CH, Lau LF, Tonks NK. MKP-1(3CH134), an immediate early gene product, is adual specificity phosphatase that dephosphory-lates MAP kinase in vivo. Cell. 1993;75:487-493.
120. Franklin CC, Kraft AS. Conditional expression ofthe mitogen-activated protein kinase (MAPK)phosphatase MKP-1 preferentially inhibits p38MAPK and stress-activated protein kinase inU937 cells. J Biol Chem. 1997;272:16917-16923.
121. Ashida N, Arai H, Yamasaki M, Kita T. Differentialsignaling for MCP-1-dependent integrin activationand chemotaxis. Ann N Y Acad Sci. 2001;947:387-389.
122. Wain JH, Kirby JA, Ali S. Leucocyte chemotaxis:examination of mitogen-activated protein kinaseand phosphoinositide 3-kinase activation bymonocyte chemoattractant proteins-1, -2, -3 and-4. Clin Exp Immunol. 2002;127:436-444.
123. Rousseau S, Houle F, Landry J, Huot J. p38 MAPkinase activation by vascular endothelial growth
RECRUITMENT OF MACROPHAGES INTO TUMORS 2233BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8

factor mediates actin reorganization and cell mi-gration in human endothelial cells. Oncogene.1997;15:2169-2177.
124. Schioppa T, Uranchimeg B, Saccani A, et al.Regulation of the chemokine receptor CXCR4 byhypoxia. J Exp Med. 2003;198:1391-1402.
125. Staller P, Sulitkova J, Lisztwan J, Moch H, Oake-ley EJ, Krek W. Chemokine receptor CXCR4downregulated by von Hippel-Lindau tumour sup-pressor pVHL. Nature. 2003;425:307-311.
126. Sun Y, Cheng Z, Ma L, Pei G. Beta-arrestin2 iscritically involved in CXCR4-mediated chemo-taxis, and this is mediated by its enhancement ofp38 MAPK activation. J Biol Chem. 2002;277:49212-49219.
127. Tachibana K, Hirota S, Iizasa H, et al. The che-mokine receptor CXCR4 is essential for vascular-ization of the gastrointestinal tract. Nature. 1998;393:591-594.
128. Salvucci O, Yao L, Villalba S, Sajewicz A, Pitta-luga S, Tosato G. Regulation of endothelial cellbranching morphogenesis by endogenous che-mokine stromal-derived factor-1. Blood. 2002;99:2703-2711.
129. Penton-Rol G, Polentarutti N, Luini W, et al. Se-lective inhibition of expression of the chemokinereceptor CCR2 in human monocytes by IFN-gamma. J Immunol. 1998;160:3869-3873.
130. David JR. Delayed hypersensitivity in vitro: itsmediation by cell-free substances formed by lym-phoid cell-antigen interaction. Proc Natl Acad SciU S A. 1966;56:72-77.
131. Bloom BR, Bennett B. Mechanism of a reaction invitro associated with delayed-type hypersensitiv-ity. Science. 1966;153:80-82.
132. Calandra T, Bernhagen J, Mitchell RA, Bucala R.The macrophage is an important and previouslyunrecognized source of macrophage migrationinhibitory factor. J Exp Med. 1994;179:1895-1902.
133. Meyer-Siegler K, Hudson PB. Enhanced expres-sion of macrophage migration inhibitory factor inprostatic adenocarcinoma metastases. Urology.1996;48:448-452.
134. Mitchell RA, Bucala R. Tumor growth-promotingproperties of macrophage migration inhibitoryfactor (MIF). Semin Cancer Biol. 2000;10:359-366.
135. Bacher M, Schrader J, Thompson N, et al. Up-regulation of macrophage migration inhibitory fac-tor gene and protein expression in glial tumorcells during hypoxic and hypoglycemic stress in-dicates a critical role for angiogenesis in glioblas-toma multiforme. Am J Pathol. 2003;162:11-17.
136. Burke B, Tang N, Corke KP, et al. Expression ofHIF-1alpha by human macrophages: implicationsfor the use of macrophages in hypoxia-regulatedcancer gene therapy. J Pathol. 2002;196:204-212.
137. Heinrich SA, Messingham KA, Gregory MS, et al.Elevated monocyte chemoattractant protein-1levels following thermal injury precede monocyterecruitment to the wound site and are controlled,in part, by tumor necrosis factor-alpha. WoundRepair Regen. 2003;11:110-119.
138. Burke-Gaffney A, Brooks AV, Bogle RG. Regula-tion of chemokine expression in atherosclerosis.Vascul Pharmacol. 2002;38:283-292.
139. Linton MF, Fazio S. Macrophages, inflammation,and atherosclerosis. Int J Obes Relat Metab Dis-ord. 2003;27(suppl 3):S35-S40.
140. DeVries ME, Ran L, Kelvin DJ. On the edge: the
physiological and pathophysiological role of che-mokines during inflammatory and immunologicalresponses. Semin Immunol. 1999;11:95-104.
141. Bodolay E, Koch AE, Kim J, Szegedi G, Szek-anecz Z. Angiogenesis and chemokines in rheu-matoid arthritis and other systemic inflammatoryrheumatic diseases. J Cell Mol Med. 2002;6:357-376.
142. Capeans C, de Rojas MV, Lojo S, Salorio MS.C-C chemokines in the vitreous of patients withproliferative vitreoretinopathy and proliferativediabetic retinopathy. Retina. 1998;18:546-550.
143. Abu el-Asrar AM, Van Damme J, Put W, et al.Monocyte chemotactic protein-1 in proliferativevitreoretinal disorders. Am J Ophthalmol. 1997;123:599-606.
144. Pufe T, Petersen W, Tillmann B, Mentlein R.Splice variants VEGF121 and VEGF165 of theangiogenic peptide vascular endothelial cellgrowth factor are expressed in the synovial tissueof patients with rheumatoid arthritis. J Rheumatol.2001;28:1482-1485.
145. Dvorak HF, Brown LF, Detmar M, Dvorak AM.Vascular permeability factor/vascular endothelialgrowth factor, microvascular hyperpermeability,and angiogenesis. Am J Pathol. 1995;146:1029-1039.
146. Inoue M, Itoh H, Ueda M, et al. Vascular endothe-lial growth factor (VEGF) expression in humancoronary atherosclerotic lesions: possible patho-physiological significance of VEGF in progressionof atherosclerosis. Circulation. 1998;98:2108-2116.
147. Afuwape AO, Kiriakidis S, Paleolog EM. The roleof the angiogenic molecule VEGF in the patho-genesis of rheumatoid arthritis. Histol His-topathol. 2002;17:961-972.
148. Ishida S, Usui T, Yamashiro K, et al. VEGF164-mediated inflammation is required for pathologi-cal, but not physiological, ischemia-induced reti-nal neovascularization. J Exp Med. 2003;198:483-489.
149. Wu L, Yu YL, Galiano RD, Roth SI, Mustoe TA.Macrophage colony-stimulating factor acceler-ates wound healing and upregulates TGF-beta1mRNA levels through tissue macrophages.J Surg Res. 1997;72:162-169.
150. Clinton SK, Underwood R, Hayes L, ShermanML, Kufe DW, Libby P. Macrophage colony-stimu-lating factor gene expression in vascular cellsand in experimental and human atherosclerosis.Am J Pathol. 1992;140:301-316.
151. Qiao JH, Tripathi J, Mishra NK, et al. Role of mac-rophage colony-stimulating factor in atherosclero-sis: studies of osteopetrotic mice. Am J Pathol.1997;150:1687-1699.
152. Seitz M, Loetscher P, Fey MF, Tobler A. Constitu-tive mRNA and protein production of macrophagecolony-stimulating factor but not of other cyto-kines by synovial fibroblasts from rheumatoid ar-thritis and osteoarthritis patients. Br J Rheumatol.1994;33:613-619.
153. Elner SG, Elner VM, Jaffe GJ, Stuart A, KunkelSL, Strieter RM. Cytokines in proliferative diabeticretinopathy and proliferative vitreoretinopathy.Curr Eye Res. 1995;14:1045-1053.
154. Black CE, Huang N, Neligan PC, Forrest CR,Lipa JE, Pang CY. Vasoconstrictor effect andmechanism of action of endothelin-1 in humanradial artery and vein: implication of skin flap va-sospasm. J Cardiovasc Pharmacol. 2003;41:460-467.
155. Lipa JE, Neligan PC, Perreault TM, et al. Vaso-
constrictor effect of endothelin-1 in human skin:role of ETA and ETB receptors. Am J Physiol.1999;276:H359-H367.
156. Fan J, Unoki H, Iwasa S, Watanabe T. Role ofendothelin-1 in atherosclerosis. Ann N Y AcadSci. 2000;902:84-93.
157. Best PJ, Lerman A. Endothelin in cardiovasculardisease: from atherosclerosis to heart failure.J Cardiovasc Pharmacol. 2000;35:S61-S63.
158. Haq A, El-Ramahi K, Al-Dalaan A, Al-Sedairy ST.Serum and synovial fluid concentrations of endo-thelin-1 in patients with rheumatoid arthritis.J Med. 1999;30:51-60.
159. Oku H, Kida T, Sugiyama T, Hamada J, Sato B,Ikeda T. Possible involvement of endothelin-1 andnitric oxide in the pathogenesis of proliferativediabetic retinopathy. Retina. 2001;21:647-651.
160. Hennemann B, Beckmann G, Eichelmann A,Rehm A, Andreesen R. Phase I trial of adoptiveimmunotherapy of cancer patients using mono-cyte-derived macrophages activated with inter-feron gamma and lipopolysaccharide. CancerImmunol Immunother. 1998;45:250-256.
161. Lei H, Ju DW, Yu Y, et al. Induction of potent anti-tumor response by vaccination with tumor lysate-pulsed macrophages engineered to secrete mac-rophage colony-stimulating factor and interferon-gamma. Gene Ther. 2000;7:707-713.
162. Huang S, Xie K, Bucana CD, Ullrich SE, Bar E.Interleukin 10 suppresses tumor growth and me-tastasis of human melanoma cells: potential inhi-bition of angiogenesis. Clin Cancer Res. 1996;2:1969-1979.
163. Bartholeyns J, Lopez M, Andreesen R. Adoptiveimmunotherapy of solid tumors with activatedmacrophages: experimental and clinical results.Anticancer Res. 1991;11:1201-1204.
164. Faradji A, Bohbot A, Schmitt G, et al. Phase I trialof intravenous infusion of ex-vivo-activated au-tologous blood-derived macrophages in patientswith non-small-cell lung cancer: toxicity and im-munomodulatory effects. Cancer Immunol Immu-nother. 1991;33:319-326.
165. Fantuzzi L, Borghi P, Ciolli V, Pavlakis G, Be-lardelli F, Gessani S. Loss of CCR2 expressionand functional response to monocyte chemotacticprotein (MCP-1) during the differentiation of hu-man monocytes: role of secreted MCP-1 in theregulation of the chemotactic response. Blood.1999;94:875-883.
166. Fellowes R, Etheridge CJ, Coade S, et al. Amelio-ration of established collagen induced arthritis bysystemic IL-10 gene delivery. Gene Ther. 2000;7:967-977.
167. Kashani-Sabet M, Liu Y, Fong S, et al. Identifica-tion of gene function and functional pathways bysystemic plasmid-based ribozyme targeting inadult mice. Proc Natl Acad Sci U S A. 2002;99:3878-3883.
168. Inoue S, Suzuki M, Nagashima Y, et al. Transferof heme oxygenase 1 cDNA by a replication-defi-cient adenovirus enhances interleukin 10 produc-tion from alveolar macrophages that attenuateslipopolysaccharide-induced acute lung injury inmice. Hum Gene Ther. 2001;12:967-979.
169. Hashiba T, Suzuki M, Nagashima Y, et al. Adeno-virus-mediated transfer of heme oxygenase-1cDNA attenuates severe lung injury induced bythe influenza virus in mice. Gene Ther. 2001;8:1499-1507.
170. Griffiths L, Binley K, Iqball S, et al. The macro-phage—a novel system to deliver gene therapy topathological hypoxia. Gene Ther. 2000;7:255-262.
2234 MURDOCH et al BLOOD, 15 OCTOBER 2004 � VOLUME 104, NUMBER 8




















