
Mechanical stress induces tumor necrosis factor- production through Ca2+ release-dependent TLR2...
8
Mechanical stress induces tumor necrosis factor- production through Ca 2 release-dependent TLR2 signaling Han Geun Kim, 1,2 * Joo Yun Kim, 1,3 * Min Geun Gim, 1 Jung Min Lee, 1,2 and Dae Kyun Chung 1,2 1 Graduate School of Biotechnology and Institute of Life Science and Resources and 2 Skin Biotechnology Center, Kyung Hee University, Yongin; and 3 Division of Cardiology, Samsung Medical Center and Samsung Biomedical Research Institute, Sungkyunkwan University School of Medicine, Seoul, Korea Submitted 14 February 2008; accepted in final form 4 June 2008 Kim HG, Kim JY, Gim MG, Lee JM, Chung DK. Mechanical stress induces tumor necrosis factor- production through Ca 2 release-dependent TLR2 signaling. Am J Physiol Cell Phys- iol 295: C432–C439, 2008. First published June 11, 2008; doi:10.1152/ajpcell.00085.2008.—We studied centrifugation-medi- ated mechanical stress-induced tumor necrosis factor- (TNF-) pro- duction in the monocyte-like cell line THP-1. The induction of TNF- by mechanical stress was dependent on the centrifugation speed and produced the highest level of TNF- after 1 h of stimulation. TNF- production returned to normal levels after 24 h of stimulation. Me- chanical stress also induced Toll-like receptor-2 (TLR2) mRNA in proportion to the expression of TNF-. The inhibition of TLR2 signaling by dominant negative myeloid differentiation factor 88 (MyD88) blocked TNF- expression response to mechanical stress. After transient overexpression of TLR2 in HEK-293 cells, mechanical stress induced TNF- mRNA production. Interestingly, mechanical stress activated the c-Src-dependent TLR2 phosphorylation, which is necessary to induce Ca 2 fluxes. When THP-1 cells were pretreated with BAPTA-AM, thapsigargin, and NiCl 2 6H 2 O, followed by me- chanical stimulation, both TLR2 and TNF- production were inhib- ited, indicating that centrifugation-mediated mechanical stress induces both TLR2 and TNF- production through Ca 2 releases from intra- cellular Ca 2 stores following TLR2 phosphorylation. In addition, TNF- treatment in THP-1 cells induced TLR2 production in re- sponse to mechanical stress, whereas the preincubation of anti-TNF- antibody scarcely induced the mechanical stress-mediated production of TLR2, indicating that TNF- produced by mechanically stimulated THP-1 cells affected TLR2 production. We concluded that TNF- production induced by centrifugation-mediated mechanical stress is dependent on MyD88-dependent TLR2 signaling that is associated with Ca 2 release and that TNF- production induced by mechanical stress affects TLR2 production. centrifugation; Toll-like receptor; THP-1 cells MECHANICAL STRESS INDUCES the production of growth-related proto-oncogene such as c-fos or c-myc in osteoblasts (20). c-fos and c-myc are considered significant genes because they play an important role in promoter regions of several genes related to the growth and mineralization of bone, including osteocal- cin, alkaline phosphatase, and collagen type 1 (9, 10). Vortex- or shear stress-mediated mechanical stress induces cell adhe- sion of THP-1 cells. In addition, THP-1 cells stimulated by platelet-derived microparticles produced by high shear stress induce cytokine expression such as tumor necrosis factor- (TNF-) and IL-1. These alterations in monocytes can induce the progression of atherosclerosis on the vessel wall (11, 17, 23). Atherosclerosis is regarded as a chronic inflammatory disease of the vessel wall and is characterized by the accumu- lation of lipid-laden macrophages and foam cells in the large arteries (15, 21). In general, monocytes respond to endogenous or exogenous ligands through pattern-recognition receptors, leading to the expression of inflammatory cytokines such as TNF-, IL-6, fibrinogen, and soluble vascular cell adhesion molecule 1. These molecules have well-described functions in inflammation that have been associated with atherosclerotic progression (19, 22). Furthermore, TNF- is a major mediator in the induction of the acute phase response (13) and affects the leukocyte-endothelial cell adhesion (2). Recently, research has focused on the mechanisms by which cytokine and chemokine expressions, as well as biofunctional changes, are induced by mechanical stress. The early cellular response to mechanical stress is the influx of Ca 2 , which leads to increased cytosolic Ca 2 and then induces changes in the intracellular activation of numerous molecules and NO pro- duction (11). Although more studies on the mechanical stress- mediated signaling pathway are necessary, mechanical stress may induce cytokine expression through NF-B activation (25). In addition, mechanical stress induces Toll-like receptor (TLR) expression as well as cytokine expression. Liang et al. (8) reported that TLR4 mRNA was upregulated after shear stress. Previous studies have shown that TLR signaling has an important function in the link between atherosclerosis and defense against both foreign pathogens and endogenously gen- erated inflammatory ligands (6, 16). These results suggest that TLR may be linked to mechanical stress-mediated cytokine production and thus may induce inflammation, atherosclerosis, and arthritis. In blood vessels, monocytes may be affected by blood flow as well as the size, shape, branching, and partial obstructions of the vessel. However, there is little evidence that monocytes induce the inflammatory cytokines in response to the mechan- ical stress caused by such an environment. The regulatory mechanism of proinflammatory cytokine expression by me- chanical stress-mediated TLR signaling is also insufficient. In this study, therefore, we examined the expression pattern of cytokines, such as TNF-, that are induced in THP-1 mono- cyte-like cells by mechanical stress. We also studied that TLR signaling and Ca 2 release are required for mechanical stress- mediated TNF- production. * H. G. Kim and J. Y. Kim contributed equally to this work. Address for reprint requests and other correspondence: D. K. Chung, Graduate School of Biotechnology and Institute of Life Science and Resources, Kyung Hee Univ., Yongin 449-701, Korea (e-mail: [email protected]). The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Am J Physiol Cell Physiol 295: C432–C439, 2008. First published June 11, 2008; doi:10.1152/ajpcell.00085.2008. 0363-6143/08 $8.00 Copyright © 2008 the American Physiological Society http://www.ajpcell.org C432
Transcript of Mechanical stress induces tumor necrosis factor- production through Ca2+ release-dependent TLR2...

Mechanical stress induces tumor necrosis factor-� production through Ca2�
release-dependent TLR2 signaling
Han Geun Kim,1,2* Joo Yun Kim,1,3* Min Geun Gim,1 Jung Min Lee,1,2 and Dae Kyun Chung1,2
1Graduate School of Biotechnology and Institute of Life Science and Resources and 2Skin Biotechnology Center, Kyung HeeUniversity, Yongin; and 3Division of Cardiology, Samsung Medical Center and Samsung Biomedical Research Institute,Sungkyunkwan University School of Medicine, Seoul, Korea
Submitted 14 February 2008; accepted in final form 4 June 2008
Kim HG, Kim JY, Gim MG, Lee JM, Chung DK. Mechanicalstress induces tumor necrosis factor-� production through Ca2�
release-dependent TLR2 signaling. Am J Physiol Cell Phys-iol 295: C432–C439, 2008. First published June 11, 2008;doi:10.1152/ajpcell.00085.2008.—We studied centrifugation-medi-ated mechanical stress-induced tumor necrosis factor-� (TNF-�) pro-duction in the monocyte-like cell line THP-1. The induction of TNF-�by mechanical stress was dependent on the centrifugation speed andproduced the highest level of TNF-� after 1 h of stimulation. TNF-�production returned to normal levels after 24 h of stimulation. Me-chanical stress also induced Toll-like receptor-2 (TLR2) mRNA inproportion to the expression of TNF-�. The inhibition of TLR2signaling by dominant negative myeloid differentiation factor 88(MyD88) blocked TNF-� expression response to mechanical stress.After transient overexpression of TLR2 in HEK-293 cells, mechanicalstress induced TNF-� mRNA production. Interestingly, mechanicalstress activated the c-Src-dependent TLR2 phosphorylation, which isnecessary to induce Ca2� fluxes. When THP-1 cells were pretreatedwith BAPTA-AM, thapsigargin, and NiCl2 �6H2O, followed by me-chanical stimulation, both TLR2 and TNF-� production were inhib-ited, indicating that centrifugation-mediated mechanical stress inducesboth TLR2 and TNF-� production through Ca2� releases from intra-cellular Ca2� stores following TLR2 phosphorylation. In addition,TNF-� treatment in THP-1 cells induced TLR2 production in re-sponse to mechanical stress, whereas the preincubation of anti-TNF-�antibody scarcely induced the mechanical stress-mediated productionof TLR2, indicating that TNF-� produced by mechanically stimulatedTHP-1 cells affected TLR2 production. We concluded that TNF-�production induced by centrifugation-mediated mechanical stress isdependent on MyD88-dependent TLR2 signaling that is associatedwith Ca2� release and that TNF-� production induced by mechanicalstress affects TLR2 production.
centrifugation; Toll-like receptor; THP-1 cells
MECHANICAL STRESS INDUCES the production of growth-relatedproto-oncogene such as c-fos or c-myc in osteoblasts (20). c-fosand c-myc are considered significant genes because they playan important role in promoter regions of several genes relatedto the growth and mineralization of bone, including osteocal-cin, alkaline phosphatase, and collagen type 1 (9, 10). Vortex-or shear stress-mediated mechanical stress induces cell adhe-sion of THP-1 cells. In addition, THP-1 cells stimulated byplatelet-derived microparticles produced by high shear stressinduce cytokine expression such as tumor necrosis factor-�(TNF-�) and IL-1�. These alterations in monocytes can inducethe progression of atherosclerosis on the vessel wall (11, 17,
23). Atherosclerosis is regarded as a chronic inflammatorydisease of the vessel wall and is characterized by the accumu-lation of lipid-laden macrophages and foam cells in the largearteries (15, 21). In general, monocytes respond to endogenousor exogenous ligands through pattern-recognition receptors,leading to the expression of inflammatory cytokines such asTNF-�, IL-6, fibrinogen, and soluble vascular cell adhesionmolecule 1. These molecules have well-described functions ininflammation that have been associated with atheroscleroticprogression (19, 22). Furthermore, TNF-� is a major mediatorin the induction of the acute phase response (13) and affects theleukocyte-endothelial cell adhesion (2).
Recently, research has focused on the mechanisms by whichcytokine and chemokine expressions, as well as biofunctionalchanges, are induced by mechanical stress. The early cellularresponse to mechanical stress is the influx of Ca2�, which leadsto increased cytosolic Ca2� and then induces changes in theintracellular activation of numerous molecules and NO pro-duction (11). Although more studies on the mechanical stress-mediated signaling pathway are necessary, mechanical stressmay induce cytokine expression through NF-�B activation(25). In addition, mechanical stress induces Toll-like receptor(TLR) expression as well as cytokine expression. Liang et al.(8) reported that TLR4 mRNA was upregulated after shearstress. Previous studies have shown that TLR signaling has animportant function in the link between atherosclerosis anddefense against both foreign pathogens and endogenously gen-erated inflammatory ligands (6, 16). These results suggest thatTLR may be linked to mechanical stress-mediated cytokineproduction and thus may induce inflammation, atherosclerosis,and arthritis.
In blood vessels, monocytes may be affected by blood flowas well as the size, shape, branching, and partial obstructions ofthe vessel. However, there is little evidence that monocytesinduce the inflammatory cytokines in response to the mechan-ical stress caused by such an environment. The regulatorymechanism of proinflammatory cytokine expression by me-chanical stress-mediated TLR signaling is also insufficient. Inthis study, therefore, we examined the expression pattern ofcytokines, such as TNF-�, that are induced in THP-1 mono-cyte-like cells by mechanical stress. We also studied that TLRsignaling and Ca2� release are required for mechanical stress-mediated TNF-� production.
* H. G. Kim and J. Y. Kim contributed equally to this work.Address for reprint requests and other correspondence: D. K. Chung,
Graduate School of Biotechnology and Institute of Life Science and Resources,Kyung Hee Univ., Yongin 449-701, Korea (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Cell Physiol 295: C432–C439, 2008.First published June 11, 2008; doi:10.1152/ajpcell.00085.2008.
0363-6143/08 $8.00 Copyright © 2008 the American Physiological Society http://www.ajpcell.orgC432
MATERIALS AND METHODS
Cell culture and cell stimulation experiments. The human mono-cyte-like cell line THP-1 and human embryonic kidney (HEK)-293cell lines were obtained from Korean Cell Line Bank (KCLB, Seoul,Korea) and maintained in RPMI 1640 and MEM, respectively. Allmedia were supplemented with 10% FBS, antibiotics (100 units ofpenicillin and 100 �g/ml streptomycin) in a 37°C incubator with5% CO2.
A 5804R centrifuge (Eppendorf, Germany) with a microtiter platerotor was used to apply mechanical stress on the THP-1 and HEK-293cells. The average shear stress on the cell surface during centrifuga-tion was calculated by Dr. Michael R. King (University of Rochester,Rochester, NY; personal communication) according to the followingformulation (14): � � 0.75 � � � U/a, where � is the bufferviscosity, U is the sedimentation velocity of the cell, and a is the cellradius. For a cell that is approximately the size of a neutrophil (a �4 �m), a centrifugation speed of 280 g corresponds to an averageshear stress on the cell surface of 2.06 dyn/cm2. In this regime of flow,the shear stress will scale linearly with the centrifugation speed. In thisstudy, the centrifugation speeds of 10, 180, and 461 g correspond toaverage shear stress on the cell surface of 0.074, 1.3, and 3.4 dyn/cm2,respectively.
RNA preparation and real-time PCR. Total RNA was extractedfrom THP-1 or HEK-293 cells by using the guanidinium thiocyanate-
acid phenol-chloroform method. cDNA was synthesized with theImprom-II reverse transcription system (Promega) according to themanufacturer’s instructions. To quantify the TNF-� and TLR2mRNA, we carried out real-time PCR amplification with the ABIPrism 7000 sequence detection system (Applied BioSystems) anddetected the PCR products with SYBR green. The following primerswere used: TNF-� forward, 5-CTCTTCTGGCTCAAAAAGAG-AATT-3; TNF-� reverse, 5-AGGCCCCAGTTTGAATTCTT-3;TLR2 forward, 5-ACCTAGGGGAAACATCTCT-3; and TLR2 re-verse, 5-AGCTCTGTAGATCTGAAGCATC-3.
Establishment of dominant negative MyD88 and transient trans-fection. Dominant negative myeloid differentiation factor 88(dnMyD88) was manufactured using the method described byWang et al. (27) with minor modifications. dnMyD88 (amino acids156 –296) that contains the Toll/IL-1 receptor domain was ampli-fied from total RNA extracted from THP-1 cells by RT-PCR andcloned into pCMV-Tag 2A (Stratagene). THP-1 and HEK-293cells were transfected with the expression vector using theWelFec-Q transfection reagent (JBI, Daegu, Korea) according tothe manufacturer’s instructions. Thirty-six hours after transfection,the cells were used for further analysis. The PCR primers used forRT-PCR were as follows: dnMyD88 forward, 5- GCTAGCAT-GCCTGAGCGTTTCG-3; and dnMyD88 reverse, 5-GGATCCT-CAGGGCAGGGACAA-3.
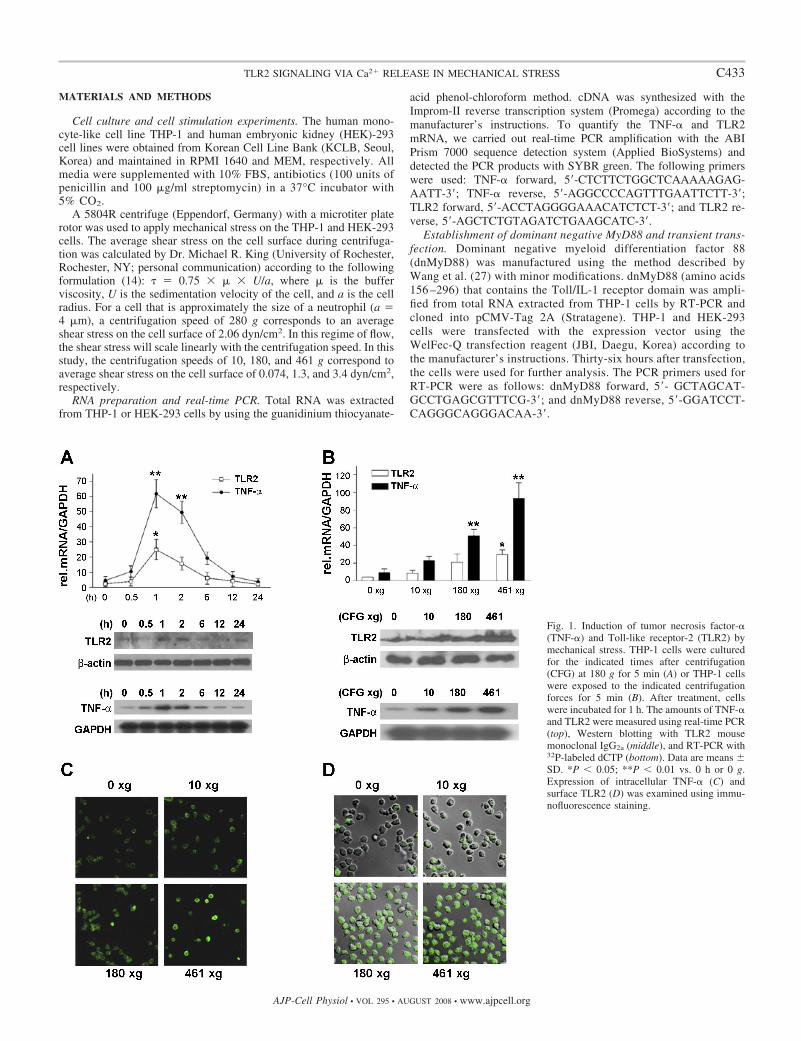
Fig. 1. Induction of tumor necrosis factor-�(TNF-�) and Toll-like receptor-2 (TLR2) bymechanical stress. THP-1 cells were culturedfor the indicated times after centrifugation(CFG) at 180 g for 5 min (A) or THP-1 cellswere exposed to the indicated centrifugationforces for 5 min (B). After treatment, cellswere incubated for 1 h. The amounts of TNF-�and TLR2 were measured using real-time PCR(top), Western blotting with TLR2 mousemonoclonal IgG2a (middle), and RT-PCR with32P-labeled dCTP (bottom). Data are means SD. *P � 0.05; **P � 0.01 vs. 0 h or 0 g.Expression of intracellular TNF-� (C) andsurface TLR2 (D) was examined using immu-nofluorescence staining.
C433TLR2 SIGNALING VIA Ca2� RELEASE IN MECHANICAL STRESS
AJP-Cell Physiol • VOL 295 • AUGUST 2008 • www.ajpcell.org
NF-�B luciferase reporter assay. THP-1 cells (5 � 105 cells/well)were seeded onto 12-well plates. After 24 h, the cells were transientlycotransfected with the pNF-�B-Luc and pRL-SV40 vectors. Thirty-six hours after transfection, the cells were stimulated by centrifugationat 180 g for 5 min. The cells were then incubated for another 18 h.Total cell lysates were prepared, and the luciferase activity of theselysates were measured using a reporter assay system (Promega). TheRenilla luciferase reporter gene (10 ng/well) was used as an internalcontrol.
Immunofluorescence staining. After stimulation, THP-1 cells werefixed with 4% paraformaldehyde. Cells were permeabilized with 0.5%Triton X-100, blocked with phosphate-buffered saline containing 1%bovine serum albumin, and incubated with polyclonal anti-TLR2(Santa Cruz Biotechnology) or anti-TNF-� (Sigma) antibody inblocking buffer at room temperature. Donkey anti-mouse IgG-FITC(Santa Cruz Biotechnology) in blocking buffer was added as asecondary antibody. Fluorescence was examined using a confocalmicroscope.
Statistical analysis. All experiments were performed at least threetimes. The data shown are representative results of means SD oftriplicate cultures. A paired t-test was used to determine the signifi-cance of the data. P values �0.05 were considered significant.
RESULTS
TLR2 mRNA was increased in response to mechanicalstress. To date, 10 members of the TLR family have beenidentified in humans (26). Different types of ligands inducedistinct types of immune responses based on the activation ofimmune cell subsets that express corresponding TLR profiles.In our experiment, THP-1 cells that were stimulated by me-chanical stress induced TLR2 and TLR4 mRNA. In particular,TLR2 mRNA was significantly induced (data not shown).Therefore, we investigated the effect of TLR2 signaling onTNF-� expression after mechanical stress.
Mechanical stress induced TNF-� and TLR2 productionfrom THP-1 cells. To examine the effect of mechanical stresson the cultured cells, we studied centrifugation-mediatedTNF-� and TLR2 production. Both TLR2 and TNF-� mRNAproduction peaked at 1 h after mechanical stimulation and thendecreased slowly (Fig. 1A, top). The TLR2 and TNF-� mRNAlevels recovered to their normal levels after 24 h. The levels ofTLR2 mRNA were confirmed by measuring the levels of TLR2protein using Western blotting (Fig. 1A, middle). RT-PCR ofTNF-� expression using 32P-labeled dCTP showed clearly thatthe production of TNF-� mRNA peaked 1 h after the cells werestimulated by centrifugation at 180 g for 5 min (Fig. 1A,bottom).
To examine the effect of mechanical stress on TNF-� andTLR2 expression, we exposed THP-1 cells to various ranges ofcentrifugation speeds. Previous study demonstrated that cen-trifugation at 3 g altered c-fos and osteocalcin gene expressionin MC3T3-E1 osteoblasts (9). Because a centrifugation forceof 3 g was not available in our experimental system, we useda force of 10 g as the lowest mechanical stress source. We alsoused a centrifugal force of 180 g (1,000 rpm) in our routine celltransfer experiments. To define our observation of the changein gene expression, we used a centrifugal force of 461 g (1,800rpm) in this experiment. The centrifugation speeds of 10, 180,and 461 g correspond to an average shear stress on the cellsurface of 0.074, 1.3, and 3.4 dyn/cm2, respectively.
The productions of TNF-� and TLR2 mRNA were increased ina speed-dependent manner (Fig. 1B, top). In addition, we used
Western blotting to examine the expression of TLR2 (Fig. 1B,middle) and 32P-labeled RT-PCR for the detection of TNF-� (Fig.1B, bottom). These alternative experimental methods showedresults similar to those obtained using real-time PCR.
To confirm the TNF-� and TLR2 expression induced bymechanical stress, we performed immunofluorescence stainingwith antibodies for TNF-� and TLR2. Confocal microscopyshowed that both TLR2 (Fig. 1C) and TNF-� (Fig. 1D) wereinduced in a speed-dependent manner. These results suggestedthat mechanical stress upregulated TLR2 expression and thatTNF-� production was proportional to TLR2 expression.
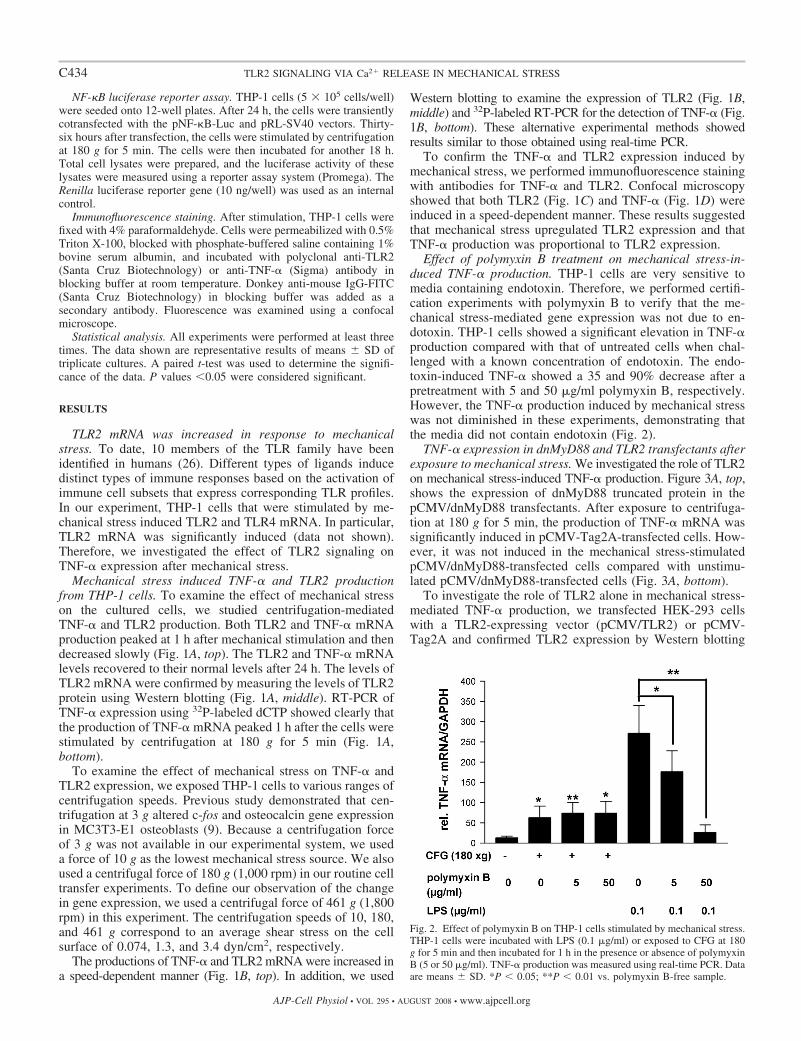
Effect of polymyxin B treatment on mechanical stress-in-duced TNF-� production. THP-1 cells are very sensitive tomedia containing endotoxin. Therefore, we performed certifi-cation experiments with polymyxin B to verify that the me-chanical stress-mediated gene expression was not due to en-dotoxin. THP-1 cells showed a significant elevation in TNF-�production compared with that of untreated cells when chal-lenged with a known concentration of endotoxin. The endo-toxin-induced TNF-� showed a 35 and 90% decrease after apretreatment with 5 and 50 �g/ml polymyxin B, respectively.However, the TNF-� production induced by mechanical stresswas not diminished in these experiments, demonstrating thatthe media did not contain endotoxin (Fig. 2).
TNF-� expression in dnMyD88 and TLR2 transfectants afterexposure to mechanical stress. We investigated the role of TLR2on mechanical stress-induced TNF-� production. Figure 3A, top,shows the expression of dnMyD88 truncated protein in thepCMV/dnMyD88 transfectants. After exposure to centrifuga-tion at 180 g for 5 min, the production of TNF-� mRNA wassignificantly induced in pCMV-Tag2A-transfected cells. How-ever, it was not induced in the mechanical stress-stimulatedpCMV/dnMyD88-transfected cells compared with unstimu-lated pCMV/dnMyD88-transfected cells (Fig. 3A, bottom).
To investigate the role of TLR2 alone in mechanical stress-mediated TNF-� production, we transfected HEK-293 cellswith a TLR2-expressing vector (pCMV/TLR2) or pCMV-Tag2A and confirmed TLR2 expression by Western blotting
Fig. 2. Effect of polymyxin B on THP-1 cells stimulated by mechanical stress.THP-1 cells were incubated with LPS (0.1 �g/ml) or exposed to CFG at 180g for 5 min and then incubated for 1 h in the presence or absence of polymyxinB (5 or 50 �g/ml). TNF-� production was measured using real-time PCR. Dataare means SD. *P � 0.05; **P � 0.01 vs. polymyxin B-free sample.
C434 TLR2 SIGNALING VIA Ca2� RELEASE IN MECHANICAL STRESS
AJP-Cell Physiol • VOL 295 • AUGUST 2008 • www.ajpcell.org
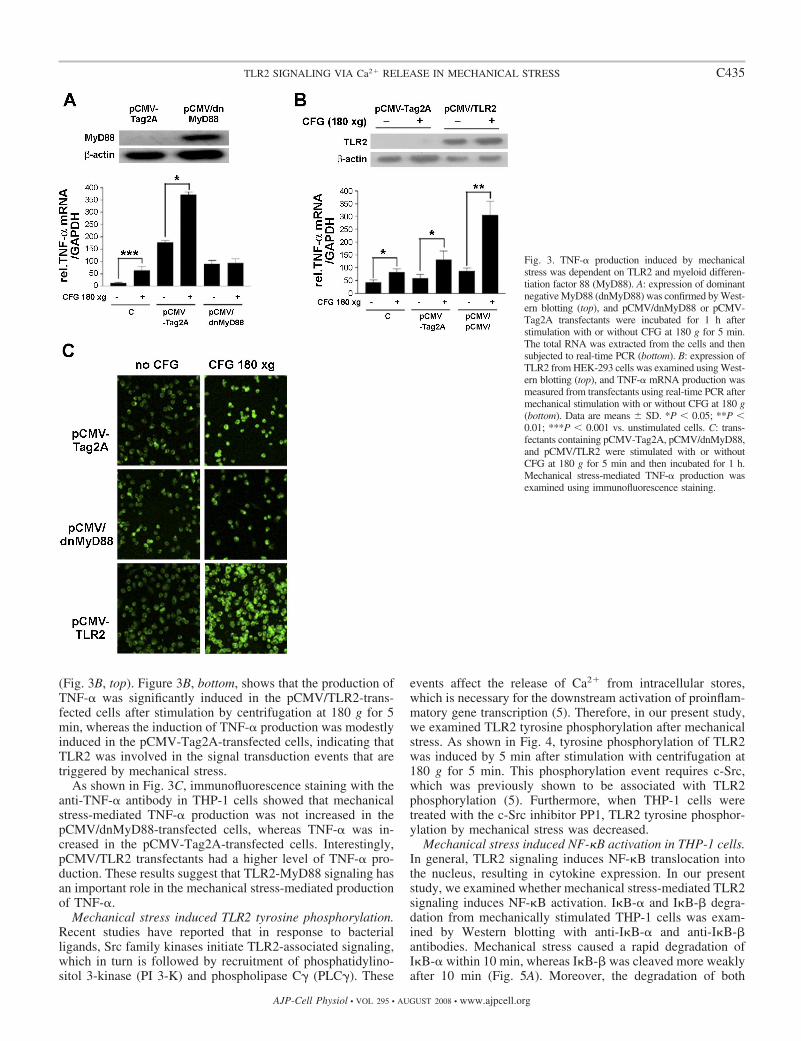
(Fig. 3B, top). Figure 3B, bottom, shows that the production ofTNF-� was significantly induced in the pCMV/TLR2-trans-fected cells after stimulation by centrifugation at 180 g for 5min, whereas the induction of TNF-� production was modestlyinduced in the pCMV-Tag2A-transfected cells, indicating thatTLR2 was involved in the signal transduction events that aretriggered by mechanical stress.
As shown in Fig. 3C, immunofluorescence staining with theanti-TNF-� antibody in THP-1 cells showed that mechanicalstress-mediated TNF-� production was not increased in thepCMV/dnMyD88-transfected cells, whereas TNF-� was in-creased in the pCMV-Tag2A-transfected cells. Interestingly,pCMV/TLR2 transfectants had a higher level of TNF-� pro-duction. These results suggest that TLR2-MyD88 signaling hasan important role in the mechanical stress-mediated productionof TNF-�.
Mechanical stress induced TLR2 tyrosine phosphorylation.Recent studies have reported that in response to bacterialligands, Src family kinases initiate TLR2-associated signaling,which in turn is followed by recruitment of phosphatidylino-sitol 3-kinase (PI 3-K) and phospholipase C� (PLC�). These
events affect the release of Ca2� from intracellular stores,which is necessary for the downstream activation of proinflam-matory gene transcription (5). Therefore, in our present study,we examined TLR2 tyrosine phosphorylation after mechanicalstress. As shown in Fig. 4, tyrosine phosphorylation of TLR2was induced by 5 min after stimulation with centrifugation at180 g for 5 min. This phosphorylation event requires c-Src,which was previously shown to be associated with TLR2phosphorylation (5). Furthermore, when THP-1 cells weretreated with the c-Src inhibitor PP1, TLR2 tyrosine phosphor-ylation by mechanical stress was decreased.
Mechanical stress induced NF-�B activation in THP-1 cells.In general, TLR2 signaling induces NF-�B translocation intothe nucleus, resulting in cytokine expression. In our presentstudy, we examined whether mechanical stress-mediated TLR2signaling induces NF-�B activation. I�B-� and I�B-� degra-dation from mechanically stimulated THP-1 cells was exam-ined by Western blotting with anti-I�B-� and anti-I�B-�antibodies. Mechanical stress caused a rapid degradation ofI�B-� within 10 min, whereas I�B-� was cleaved more weaklyafter 10 min (Fig. 5A). Moreover, the degradation of both
Fig. 3. TNF-� production induced by mechanicalstress was dependent on TLR2 and myeloid differen-tiation factor 88 (MyD88). A: expression of dominantnegative MyD88 (dnMyD88) was confirmed by West-ern blotting (top), and pCMV/dnMyD88 or pCMV-Tag2A transfectants were incubated for 1 h afterstimulation with or without CFG at 180 g for 5 min.The total RNA was extracted from the cells and thensubjected to real-time PCR (bottom). B: expression ofTLR2 from HEK-293 cells was examined using West-ern blotting (top), and TNF-� mRNA production wasmeasured from transfectants using real-time PCR aftermechanical stimulation with or without CFG at 180 g(bottom). Data are means SD. *P � 0.05; **P �0.01; ***P � 0.001 vs. unstimulated cells. C: trans-fectants containing pCMV-Tag2A, pCMV/dnMyD88,and pCMV/TLR2 were stimulated with or withoutCFG at 180 g for 5 min and then incubated for 1 h.Mechanical stress-mediated TNF-� production wasexamined using immunofluorescence staining.
C435TLR2 SIGNALING VIA Ca2� RELEASE IN MECHANICAL STRESS
AJP-Cell Physiol • VOL 295 • AUGUST 2008 • www.ajpcell.org
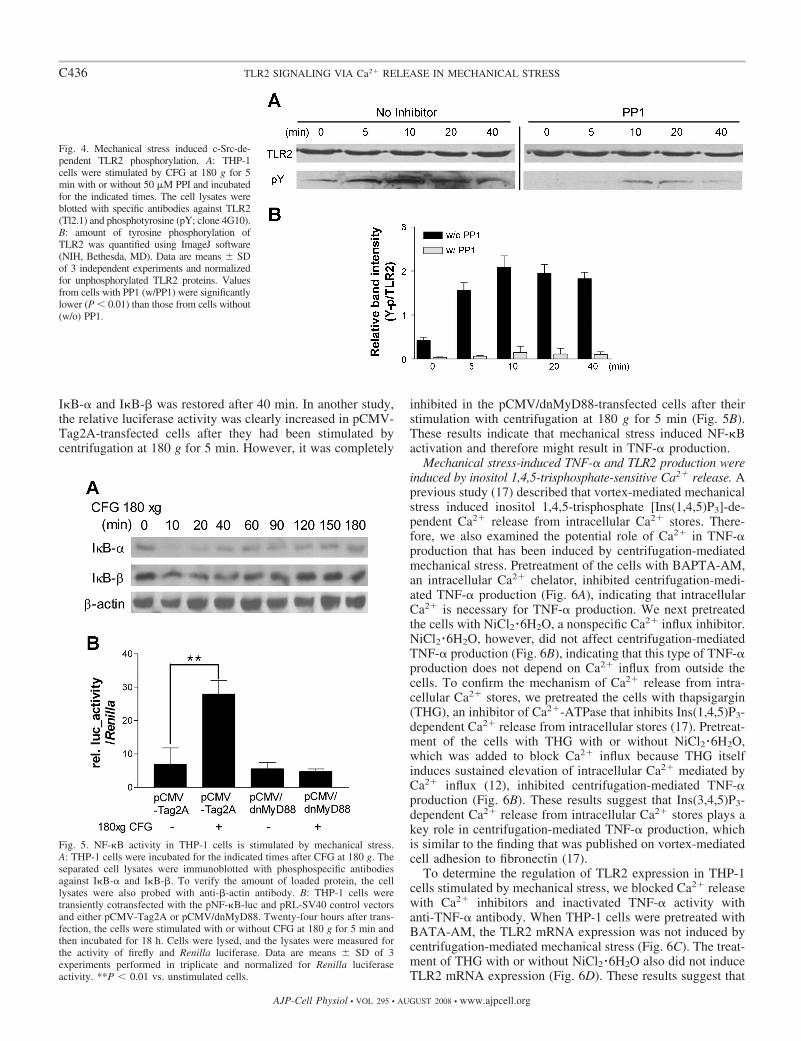
I�B-� and I�B-� was restored after 40 min. In another study,the relative luciferase activity was clearly increased in pCMV-Tag2A-transfected cells after they had been stimulated bycentrifugation at 180 g for 5 min. However, it was completely
inhibited in the pCMV/dnMyD88-transfected cells after theirstimulation with centrifugation at 180 g for 5 min (Fig. 5B).These results indicate that mechanical stress induced NF-�Bactivation and therefore might result in TNF-� production.
Mechanical stress-induced TNF-� and TLR2 production wereinduced by inositol 1,4,5-trisphosphate-sensitive Ca2� release. Aprevious study (17) described that vortex-mediated mechanicalstress induced inositol 1,4,5-trisphosphate [Ins(1,4,5)P3]-de-pendent Ca2� release from intracellular Ca2� stores. There-fore, we also examined the potential role of Ca2� in TNF-�production that has been induced by centrifugation-mediatedmechanical stress. Pretreatment of the cells with BAPTA-AM,an intracellular Ca2� chelator, inhibited centrifugation-medi-ated TNF-� production (Fig. 6A), indicating that intracellularCa2� is necessary for TNF-� production. We next pretreatedthe cells with NiCl2 �6H2O, a nonspecific Ca2� influx inhibitor.NiCl2 �6H2O, however, did not affect centrifugation-mediatedTNF-� production (Fig. 6B), indicating that this type of TNF-�production does not depend on Ca2� influx from outside thecells. To confirm the mechanism of Ca2� release from intra-cellular Ca2� stores, we pretreated the cells with thapsigargin(THG), an inhibitor of Ca2�-ATPase that inhibits Ins(1,4,5)P3-dependent Ca2� release from intracellular stores (17). Pretreat-ment of the cells with THG with or without NiCl2 �6H2O,which was added to block Ca2� influx because THG itselfinduces sustained elevation of intracellular Ca2� mediated byCa2� influx (12), inhibited centrifugation-mediated TNF-�production (Fig. 6B). These results suggest that Ins(3,4,5)P3-dependent Ca2� release from intracellular Ca2� stores plays akey role in centrifugation-mediated TNF-� production, whichis similar to the finding that was published on vortex-mediatedcell adhesion to fibronectin (17).
To determine the regulation of TLR2 expression in THP-1cells stimulated by mechanical stress, we blocked Ca2� releasewith Ca2� inhibitors and inactivated TNF-� activity withanti-TNF-� antibody. When THP-1 cells were pretreated withBATA-AM, the TLR2 mRNA expression was not induced bycentrifugation-mediated mechanical stress (Fig. 6C). The treat-ment of THG with or without NiCl2 �6H2O also did not induceTLR2 mRNA expression (Fig. 6D). These results suggest that
Fig. 4. Mechanical stress induced c-Src-de-pendent TLR2 phosphorylation. A: THP-1cells were stimulated by CFG at 180 g for 5min with or without 50 �M PPI and incubatedfor the indicated times. The cell lysates wereblotted with specific antibodies against TLR2(Tl2.1) and phosphotyrosine (pY; clone 4G10).B: amount of tyrosine phosphorylation ofTLR2 was quantified using ImageJ software(NIH, Bethesda, MD). Data are means SDof 3 independent experiments and normalizedfor unphosphorylated TLR2 proteins. Valuesfrom cells with PP1 (w/PP1) were significantlylower (P � 0.01) than those from cells without(w/o) PP1.
Fig. 5. NF-�B activity in THP-1 cells is stimulated by mechanical stress.A: THP-1 cells were incubated for the indicated times after CFG at 180 g. Theseparated cell lysates were immunoblotted with phosphospecific antibodiesagainst I�B-� and I�B-�. To verify the amount of loaded protein, the celllysates were also probed with anti-�-actin antibody. B: THP-1 cells weretransiently cotransfected with the pNF-�B-luc and pRL-SV40 control vectorsand either pCMV-Tag2A or pCMV/dnMyD88. Twenty-four hours after trans-fection, the cells were stimulated with or without CFG at 180 g for 5 min andthen incubated for 18 h. Cells were lysed, and the lysates were measured forthe activity of firefly and Renilla luciferase. Data are means SD of 3experiments performed in triplicate and normalized for Renilla luciferaseactivity. **P � 0.01 vs. unstimulated cells.
C436 TLR2 SIGNALING VIA Ca2� RELEASE IN MECHANICAL STRESS
AJP-Cell Physiol • VOL 295 • AUGUST 2008 • www.ajpcell.org
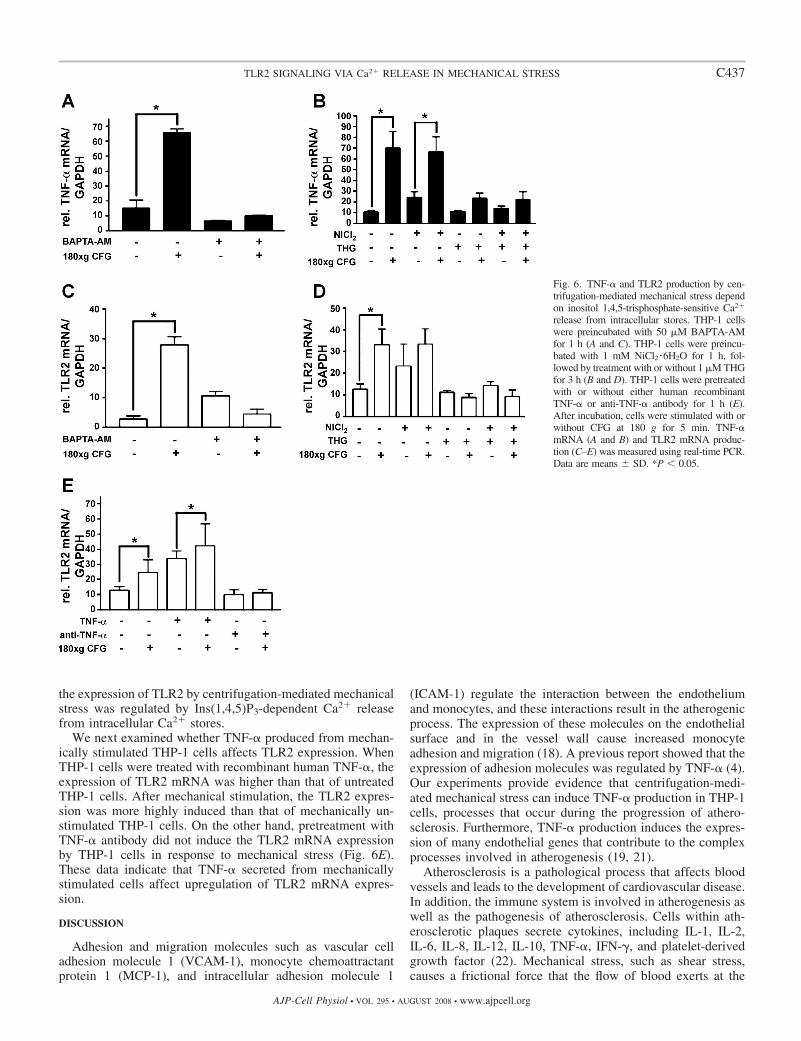
the expression of TLR2 by centrifugation-mediated mechanicalstress was regulated by Ins(1,4,5)P3-dependent Ca2� releasefrom intracellular Ca2� stores.
We next examined whether TNF-� produced from mechan-ically stimulated THP-1 cells affects TLR2 expression. WhenTHP-1 cells were treated with recombinant human TNF-�, theexpression of TLR2 mRNA was higher than that of untreatedTHP-1 cells. After mechanical stimulation, the TLR2 expres-sion was more highly induced than that of mechanically un-stimulated THP-1 cells. On the other hand, pretreatment withTNF-� antibody did not induce the TLR2 mRNA expressionby THP-1 cells in response to mechanical stress (Fig. 6E).These data indicate that TNF-� secreted from mechanicallystimulated cells affect upregulation of TLR2 mRNA expres-sion.
DISCUSSION
Adhesion and migration molecules such as vascular celladhesion molecule 1 (VCAM-1), monocyte chemoattractantprotein 1 (MCP-1), and intracellular adhesion molecule 1
(ICAM-1) regulate the interaction between the endotheliumand monocytes, and these interactions result in the atherogenicprocess. The expression of these molecules on the endothelialsurface and in the vessel wall cause increased monocyteadhesion and migration (18). A previous report showed that theexpression of adhesion molecules was regulated by TNF-� (4).Our experiments provide evidence that centrifugation-medi-ated mechanical stress can induce TNF-� production in THP-1cells, processes that occur during the progression of athero-sclerosis. Furthermore, TNF-� production induces the expres-sion of many endothelial genes that contribute to the complexprocesses involved in atherogenesis (19, 21).
Atherosclerosis is a pathological process that affects bloodvessels and leads to the development of cardiovascular disease.In addition, the immune system is involved in atherogenesis aswell as the pathogenesis of atherosclerosis. Cells within ath-erosclerotic plaques secrete cytokines, including IL-1, IL-2,IL-6, IL-8, IL-12, IL-10, TNF-�, IFN-�, and platelet-derivedgrowth factor (22). Mechanical stress, such as shear stress,causes a frictional force that the flow of blood exerts at the
Fig. 6. TNF-� and TLR2 production by cen-trifugation-mediated mechanical stress dependon inositol 1,4,5-trisphosphate-sensitive Ca2�
release from intracellular stores. THP-1 cellswere preincubated with 50 �M BAPTA-AMfor 1 h (A and C). THP-1 cells were preincu-bated with 1 mM NiCl2 �6H2O for 1 h, fol-lowed by treatment with or without 1 �M THGfor 3 h (B and D). THP-1 cells were pretreatedwith or without either human recombinantTNF-� or anti-TNF-� antibody for 1 h (E).After incubation, cells were stimulated with orwithout CFG at 180 g for 5 min. TNF-�mRNA (A and B) and TLR2 mRNA produc-tion (C–E) was measured using real-time PCR.Data are means SD. *P � 0.05.
C437TLR2 SIGNALING VIA Ca2� RELEASE IN MECHANICAL STRESS
AJP-Cell Physiol • VOL 295 • AUGUST 2008 • www.ajpcell.org

endothelial surface of the vessel wall and results in the pro-gression of atherosclerosis (24). Shear stress is defined as theviscous drag of blood over the endothelium. Both animal andhuman models have shown that atherosclerotic lesions occurpredominantly at specific arterial regions with low or disturbedflow, where endothelial shear stress (ESS) occurs and wherethe atheroprotective genes are suppressed, whereas theproatherogenic genes are upregulated, thereby promoting theatherosclerotic process (3). These results suggest that mechan-ical stress plays an important role in the atherosclerotic pro-cess.
The response of cells to mechanical stress is similar to theresponse of ligand-mediated TLR signaling in the gene expres-sion of cytokines, chemokines, growth factors, and transcrip-tion factors, suggesting an interrelation between mechanicalstress and TLR. Recently, Liang et al. (8) showed that over-expression of TLR4 could be induced by mechanical stress. Inour present study, TLR2 was significantly induced by mechan-ical stress, whereas other TLRs were only moderately induced.Therefore, we examined whether TLR2 signaling participatedin mechanical stress-mediated TNF-� production. WhenTHP-1 cells were transfected with dnMyD88, the expression ofTNF-� after the exposure to mechanical stress was not in-duced, whereas TLR2-transfected cells, after their exposure tomechanical stress, had higher TNF-� production comparedwith unstimulated TLR2-transfected cells. These results indi-cate that mechanical stress-mediated TNF-� expression wasinduced through MyD88-dependent TLR2 signaling. By usinga genetic loss-of-function approach, a recent study (16) showedthat MyD88 has an important role in the development ofatherosclerosis in murine models of atherogenesis. This studyshowed that a deficiency in MyD88 led to a significant reduc-tion in plaque size, lipid content, and expression of proinflam-matory genes (16). Another study (7) has suggested thatlaminar flow induced SP1 serine phosphorylation and therebyblocked SP1 binding to the TLR2 promoter, which is requiredfor TLR2 expression. This regulatory mechanism of TLR2production may contribute to an atheroprotective role in ath-erosclerotic lesion formation. Therefore, our study, whichshows that TLR2 and MyD88 signaling mediated mechanicalstress-mediated TNF-� production, may provide further insightinto the mechanisms of mechanical stress in atheroscleroticformation.
Signal transduction through TLR2 is initiated by c-Src-depen-dent phosphorylation of TLR2, which is necessary to induce theassociation of PI3K and TLR2 signaling. The generation ofphosphatidylinositol 3,4,5-trisphosphate [PI(3,4,5)P3] by PI 3-Kactivates PLC� and is involved in intracellular Ca2� release bygenerating Ins(1,4,5)P3 (1). PLC� activity is regulated by plasmamembrane localization and tyrosine phosphorylation. Further-more, PLC� activity is associated with TLR2 in response to theTLR2 agonist (5). In the present study, we showed that centrifu-gation-mediated mechanical stress induced TNF-� and TLR2production through Ins(1,4,5)P3-dependent Ca2� release fromintracellular Ca2� stores. These results were confirmed by thepretreatment of cells with Ca2� inhibitors. Moreover, the TNF-�produced from the mechanically stimulated-THP-1 cells affectedTLR2 mRNA production. In addition, TLR2 signal transductionby mechanical stress induced TNF-� production through aMyD88-dependent pathway that included NF-�B activation.These results suggest that mechanical stress on THP-1 cells leads
to mutual action between TLR2 and TNF-�, which may occur viaintracellular Ca2� release and a MyD88-dependent NF-�B signalpathway.
In conclusion, we showed that centrifugation-mediated me-chanical stress upregulated TNF-� and TLR2 production inTHP-1 cells and that c-Src-dependent TLR2 phosphorylationfollowed by MyD88-dependent NF-�B activation and Ca2�
release-dependent TNF-� production were involved in thismechanism. These findings might elucidate another aspect ofthe signaling of mechanical stress-mediated atherosclerosis.
ACKNOWLEDGMENTS
We thank Michael R. King for calculation of the amount of shear stress onthe cell surface during centrifugation.
GRANTS
This work was supported by Regional Innovation Center Grant RIC07-06-04 (Skin Biotechnology Center), funded by the Ministry of KnowledgeEconomy, Republic of Korea.
REFERENCES
1. Bae YS, Cantley LG, Chen CS, Kim SR, Kwon KS, Rhee SG.Activation of phospholipase C-� by phosphatidylinositol 3,4,5-trisphos-phate. J Biol Chem 273: 4465–4469, 1998.
2. Bevilacqua MP, Pober JS, Mendrick DL, Cotran RS, Gimbrone MA.Identification of an inducible endothelial-leukocyte adhesion molecule.Proc Natl Acad Sci USA 84: 9238–9242, 1987.
3. Chatzizisis YS, Coskun AU, Jonas M, Edelman ER, Feldman CL,Stone PH. Role of endothelial shear stress in the natural history ofcoronary atherosclerosis and vascular remodeling: molecular, cellular, andvascular behavior. J Am Coll Cardiol 49: 2379–2393, 2007.
4. Chiu JJ, Lee PL, Chen CN, Lee CI, Chang SF, Chen LJ, Lien SC, KoYC, Usami S, Chien S. Shear stress increases ICAM-1 and decreasesCAM-1 and E-selectin expressions induced by tumor necrosis factor-� inendothelial cells. Arterioscler Thromb Vasc Biol 24: 73–79, 2004.
5. Chun J, Prince A. Activation of Ca2�-dependent signaling by TLR2.J Immunol 177: 1330–1337, 2006.
6. Cook DN, Pisetsky DS, Schwartz DA. Toll-like receptors in the patho-genesis of human disease. Nat Immunol 5: 975–979, 2004.
7. Dunzendorfer S, Lee HK, Tobias PS. Flow-dependent regulation ofendothelial Toll-like receptor 2 expression through inhibition of SP1activity. Circ Res 95: 684–691, 2004.
8. Liang F, Huang N, Wang B, Chen H, Wu L. Shear stress inducesinterleukin-8 mRNA expression and transcriptional activation in humanvascular endothelial cells. Chin Med J (Engl) 115: 1838–1842, 2002.
9. Fitzgerald J, Hughes-Fulford M. Gravitational loading of a simulatedlaunch alters mRNA expression in osteoblasts. Exp Cell Res 228: 168–171, 1996.
10. Fitzgerald J, Hughes-Fulford M. Mechanically induced c-fos expressionis mediated by cAMP in MC3T3-E1 osteoblast. FASEB J 13: 553–557,1999.
11. Kristopher SC, Avrum IG. The role of shear stress in the pathogenesisof atherosclerosis. Lab Invest 85: 9–23, 2005.
12. Kuroiwa-Matsumoto M, Hirano K, Ahmed A, Kawasaki J, NishimuraJ, Kanaide H. Mechanisms of the thapsigargin-induced Ca2� entry in insitu endothelial cells of the porcine aortic valve and the endothelium-dependent relaxation in the porcine coronary artery. Br J Pharmacol 131:115–123, 2000.
13. Kushner I. Regulation of the acute phase response by cytokines. PerspectBiol Med 36: 611–622, 1993.
14. Lee D, King MR. Shear-induced capping of L-selectin on the neutrophilsurface during centrifugation. J Immunol Methods 328: 97–105, 2007.
15. Lusis AJ. Atherosclerosis. Nature 407: 233–241, 2000.16. Michelsen KS, Doherty TM, Shah PK, Arditi M. TLR signaling: an
emerging bridge from innate immunity to atherogenesis. J Immunol 173:5901–5907, 2004.
17. Noboru A, Hajime T, Toru K, Hidenori A. Vortex-mediated mechanicalstress induces integrin-dependent cell adhesion mediated by inositol 1,4,5-triphosphate-sensitive Ca2� release in THP-1 cells. J Biol Chem 278:9327–9331, 2003.
C438 TLR2 SIGNALING VIA Ca2� RELEASE IN MECHANICAL STRESS
AJP-Cell Physiol • VOL 295 • AUGUST 2008 • www.ajpcell.org

18. Piga R, Naito Y, Kokura S, Handa O, Yoshikawa T. Short-term highglucose exposure induces monocyte-endothelial cells adhesion and trans-migration by increasing VCAM-1 and MCP-1 expression in human aorticendothelial cells. Atherosclerosis 193: 328–334, 2007.
19. Popa C, Netea MG, van Riel PL, van der Meer JW, Stalenhoef AF.The role of TNF-� in chronic inflammatory conditions, intermediarymetabolism, and cardiovascular risk. J Lipid Res 48: 751–762, 2007.
20. Raymond RT, Vicki LV, Millie H. Vibrational force alters mRNAexpression in osteoblasts. FASEB J 11: 493–497, 1997.
21. Ross R. The pathogenesis of atherosclerosis: a perspective for the 1990s.Nature 362: 801–809, 1993.
22. Sherer Y, Shoenfeld Y. Mechanisms of disease: atherosclerosis in auto-immune diseases. Nat Clin Pract Rheumatol 2: 99–106, 2006.
23. Shosaku N, Narendra NT, Takashi N, James C, Shirou F, Junichi K.High-shear-stress-induced activation of platelets and microparticles en-
hances expression of cell adhesion molecules in THP-1 and endothelialcells. Atherosclerosis 158: 277–287, 2001.
24. Siasos G, Tousoulis D, Siasou Z, Stefanadis C, Papavassiliou AG.Shear stress, protein kinases and atherosclerosis. Curr Med Chem 14:1567–1572, 2007.
25. Sudha A, James D, Ping L, Anupam V, Cynthia H, Christopher HE,Nicholas P. Role of NF-�B transcription factors in anti-inflammatory andproinflammatory actions of mechanical signals. Arthritis Rheum 50: 3541–3548, 2004.
26. Takeda K, Akira S. Toll-like receptors in innate immunity. Int Immunol17: 1–14, 2005.
27. Wang J, Kobayashi M, Han M, Choi S, Takano M, Hashino S, TanakaJ, Kondoh T, Kawamura KI, Hosokawa M. MyD88 is involved in thesignaling pathway for taxtol-induced apoptosis and TNF-� expression inhuman myelomonocytic cells. Br J Haematol 118: 638–645, 2002.
C439TLR2 SIGNALING VIA Ca2� RELEASE IN MECHANICAL STRESS
AJP-Cell Physiol • VOL 295 • AUGUST 2008 • www.ajpcell.org




















