
Mass loss and macroinvertebrate colonisation of fish carcasses in riffles and pools of a NW Italian...
11
Mass loss and macroinvertebrate colonisation of Pacific salmon carcasses in south-eastern Alaskan streams DOMINIC T. CHALONER,* MARK S. WIPFLI and JOHN P. CAOUETTE *Department of Entomology, Michigan State University, East Lansing, MI, U.S.A. Pacific Northwest Research Station, USDA Forest Service, Juneau, AK, U.S.A. SUMMARY 1. We examined the spatial and temporal dynamics of pink salmon (Oncorhynchus gorbuscha) carcass decomposition (mass loss and macroinvertebrate colonisation) in south- eastern Alaskan streams. Dry mass and macroinvertebrate fauna of carcasses placed in streams were measured every two weeks over two months in six artificial streams and once after six weeks in four natural streams. We also surveyed the macroinvertebrate fauna and wet mass of naturally occurring salmon carcasses. 2. Carcass mass loss in artificial streams was initially rapid and then declined over time (k ¼ –0.033 day –1 ), and no significant differences were found among natural streams. 3. Several macroinvertebrate taxa colonised carcasses, but chironomid midge (Diptera: Chironomidae) and Zapada (Plecoptera: Nemouridae) larvae were found consistently and were the most abundant (on average 95 and 2%, respectively, of the invertebrates found). Chironomid abundance and biomass increased over time, whereas Zapada abundance and biomass did not. Significant differences in abundance were found among natural streams for Baetis (Ephemeroptera: Baetidae) and Sweltsa (Plecoptera: Chloroperlidae) larvae, while no significant differences were found for chironomid and Zapada abundance or biomass. 4. Our results suggest that salmon carcasses initially undergo a high rate of mass loss that tapers off with time. Chironomid and Zapada larvae are likely to be important in mediating nutrient and energy transfer between salmon carcasses and other components of the freshwater-riparian food web in south-eastern Alaskan streams. Keywords: carcasses, decomposition, invertebrate colonisation, mass loss, Pacific salmon Introduction The annual migration of Pacific salmon (Oncorhynchus spp.) results in the movement of large amounts of organic material into north Pacific rim catchments (Cederholm et al., 1999). These salmon runs are especially remarkable in Alaska and western Canada where salmon populations have, until quite recently, been stable (Baker et al., 1996). Marine-derived nitro- gen and carbon delivered by salmon can be traced into freshwater and terrestrial organisms (Schuldt & Hershey, 1995; Bilby, Fransen & Bisson, 1996; Kline, Goering & Piorkowski, 1997) and may be a resource subsidy critical for maintaining freshwater and ripar- ian productivity (Polis, Anderson & Holt, 1997). However, the ecological processes by which salmon influence ecosystems are not well understood (Cederholm et al., 1999) and the overall influence of marine resource subsidies probably underestimated (Willson, Gende & Marston, 1998). Little is known about carrion decomposition in freshwater (Minshall, Hitchcock & Barnes, 1991; Parmenter & Lamarra, 1991; Merritt & Wallace, 2000), including mass loss and invertebrate colonisa- tion. Much more is known about the decomposition of terrestrial carrion (Hanski, 1987) and other organic inputs to freshwater, such as leaf litter (Webster & Benfield, 1986). Terrestrial carrion, for example, Correspondence: Dominic Chaloner, Department of Biological Sciences, University of Notre Dame, Notre Dame, IN 46556-0369, U.S.A. E-mail: [email protected] Freshwater Biology (2002) 47, 263–273 Ó 2002 Blackwell Science Ltd 263
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Mass loss and macroinvertebrate colonisation of fish carcasses in riffles and pools of a NW Italian...

Mass loss and macroinvertebrate colonisation of Pacificsalmon carcasses in south-eastern Alaskan streams
DOMINIC T. CHALONER,* MARK S. WIPFLI† and JOHN P. CAOUETTE†
*Department of Entomology, Michigan State University, East Lansing, MI, U.S.A.
†Pacific Northwest Research Station, USDA Forest Service, Juneau, AK, U.S.A.
SUMMARY
1. We examined the spatial and temporal dynamics of pink salmon (Oncorhynchusgorbuscha) carcass decomposition (mass loss and macroinvertebrate colonisation) in south-
eastern Alaskan streams. Dry mass and macroinvertebrate fauna of carcasses placed in
streams were measured every two weeks over two months in six artificial streams and
once after six weeks in four natural streams. We also surveyed the macroinvertebrate
fauna and wet mass of naturally occurring salmon carcasses.
2. Carcass mass loss in artificial streams was initially rapid and then declined over time
(k ¼ –0.033 day–1), and no significant differences were found among natural streams.
3. Several macroinvertebrate taxa colonised carcasses, but chironomid midge (Diptera:
Chironomidae) and Zapada (Plecoptera: Nemouridae) larvae were found consistently and
were the most abundant (on average 95 and 2%, respectively, of the invertebrates found).
Chironomid abundance and biomass increased over time, whereas Zapada abundance and
biomass did not. Significant differences in abundance were found among natural streams
for Baetis (Ephemeroptera: Baetidae) and Sweltsa (Plecoptera: Chloroperlidae) larvae, while
no significant differences were found for chironomid and Zapada abundance or biomass.
4. Our results suggest that salmon carcasses initially undergo a high rate of mass loss that
tapers off with time. Chironomid and Zapada larvae are likely to be important in mediating
nutrient and energy transfer between salmon carcasses and other components of the
freshwater-riparian food web in south-eastern Alaskan streams.
Keywords: carcasses, decomposition, invertebrate colonisation, mass loss, Pacific salmon
Introduction
The annual migration of Pacific salmon (Oncorhynchus
spp.) results in the movement of large amounts of
organic material into north Pacific rim catchments
(Cederholm et al., 1999). These salmon runs are
especially remarkable in Alaska and western Canada
where salmon populations have, until quite recently,
been stable (Baker et al., 1996). Marine-derived nitro-
gen and carbon delivered by salmon can be traced
into freshwater and terrestrial organisms (Schuldt &
Hershey, 1995; Bilby, Fransen & Bisson, 1996; Kline,
Goering & Piorkowski, 1997) and may be a resource
subsidy critical for maintaining freshwater and ripar-
ian productivity (Polis, Anderson & Holt, 1997).
However, the ecological processes by which salmon
influence ecosystems are not well understood
(Cederholm et al., 1999) and the overall influence of
marine resource subsidies probably underestimated
(Willson, Gende & Marston, 1998).
Little is known about carrion decomposition in
freshwater (Minshall, Hitchcock & Barnes, 1991;
Parmenter & Lamarra, 1991; Merritt & Wallace,
2000), including mass loss and invertebrate colonisa-
tion. Much more is known about the decomposition
of terrestrial carrion (Hanski, 1987) and other organic
inputs to freshwater, such as leaf litter (Webster &
Benfield, 1986). Terrestrial carrion, for example,
Correspondence: Dominic Chaloner, Department of Biological
Sciences, University of Notre Dame, Notre Dame, IN 46556-0369,
U.S.A. E-mail: [email protected]
Freshwater Biology (2002) 47, 263–273
Ó 2002 Blackwell Science Ltd 263

undergoes a predictable sequence of mass loss and
invertebrate colonisation over time (Hanski, 1987;
Haskell et al., 1989), which can be influenced by
conditions associated with different biomes, such as
grassland versus woodland (Hanski, 1987). Although
obligate necrophagous invertebrate taxa have
evolved only on land where carrion supply is
frequent and predictable (Hanski, 1987; Haskell et al.,
1989), invertebrates are important processors of
marine as well as terrestrial carrion (Britton &
Morton, 1994). Freshwater invertebrates are generally
important agents in the breakdown of organic
material, nutrient cycling and energy flow within
food webs (Cummins, 1973), and can be abundant on
carrion in freshwaters (Vance, Van Dyk & Rowley,
1995; Keiper, Chapman & Foote, 1997; Kline et al.,
1997; Minakawa & Gara, 1999). Invertebrate coloni-
sation has been observed on adult salmon carcasses
throughout the Pacific north-west (Kline et al., 1997;
Minakawa & Gara, 1999), including south-eastern
Alaska where invertebrates have been found scaven-
ging on salmon carcasses (Wipfli, Hudson & Caou-
ette, 1998) and dead salmon alevins and eggs (Ellis,
1970; Elliott & Bartoo, 1981).
Our objectives in this study were to measure the
rates of salmon carcass decomposition, in terms
of mass loss and the abundance and biomass
of invertebrate colonists, and to compare rates of
decomposition among several streams. The results
of this study will contribute to a growing body of
knowledge concerning the influence of decom-
posing salmon on freshwater habitats. A better
understanding of the dynamics of Pacific salmon
decomposition should aid fisheries and ecosystem
management.
Methods
Study location
This study took place in the Margaret Creek catch-
ment on Revillagigedo Island in south-eastern Alaska,
U.S.A. Several anadromous salmonid species, inclu-
ding pink (Oncorhynchus gorbuscha), sockeye (O. nerka),
chum (O. keta) and coho salmon (O. kisutch), regularly
spawn in Margaret and Cobble Creeks, and since
installation of a fish ladder in 1989, in Sprout Fork and
Spike Creek (Bryant, Frenette & McCurdy, 1999).
Spawning takes place from July until late November,
but mostly during August and early September.
Our study combined artificial stream experiments
with natural stream experiments and surveys (cf.
Wipfli et al., 1998, 1999). This enabled us to study
temporal changes in artificial streams and spatial
differences among natural streams, as well as com-
pare patterns under artificial and more realistic
natural conditions. Artificial stream experiments were
conducted in six channels of a larger ‘mesocosm’
(Wipfli et al., 1998 for full description) supplied with
water from a portion of Spike Creek inaccessible to
salmon. Natural stream experiments took place in
Margaret Creek, Cobble Creek, Spike Creek and
Sprout Fork, which represented contrasts in dis-
charge, riparian management, stream habitat, and
history of salmon spawning (Table 1). Surveys of
naturally occurring salmon carcasses were limited to
Cobble Creek and Margaret Creek as only a few
carcasses were found in Sprout Fork and Spike Creek.
Surveys provided information about the invertebrate
abundance on carcasses and the amount of carcass
material present in streams during a natural salmon
run.
Discharge
Dissolved
oxygen Temperature Temperature Riparian
Stream (m3 s–1) (mg L–1) (°C, periodic) (°C, continuous) forest type
Margaret 3.83 10.6 10.4 9.2 Mainstem – mixed
(1.26) (9.9–11.6) (8.6–11.6) (6.2–11.6) old/young growth.
Cobble 0.58 11.1 8.5 n/a Tributary – mixed
(0.17) (10.5–11.7) (7.3–10.1) old/young growth.
Spike 0.19 10.7 9.4 8.8 Tributary – old
(0.12) (10.3–11.4) (7.9–11.0) (5.5–11.4) growth.
Sprout 0.06 10.5 9.1 n/a Tributary – young
(0.01) (10.3–10.8) (7.4–9.8) growth.
Table 1 Baseflow discharge [average
(±1 SEM)], dissolved oxygen [average
(range)], water temperature [periodic
and continuous: average (range)] and
riparian forest type of the four streams
used in this study
264 D.T. Chaloner et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273

Carcasses placed in artificial streams
On 1 September 1997, 24 female pink salmon (48–
52 cm fork length, 1.25–1.49 kg wet mass, 0.35–0.42 kg
dry mass) were taken live from Margaret Creek and
killed. Initial dry mass of each fish was estimated by
multiplying the original wet mass by 0.28 (Wipfli
et al., 1998). Fresh, egg-bearing rather than spawned-
out salmon were used to ensure uniformity among
and between experiments. Salmon carcasses in similar
condition are commonly found in and around
streams. Four carcasses were tethered to the floor of
each artificial stream channel, on top of fibreglass
mesh (mesh size ¼ 1.6 mm) to facilitate their removal.
Channels were dammed to create pools 250 cm long,
18 cm wide and 13 cm deep. During the experiment,
discharge through the channels averaged 0.488 ±
0.001 L s–1 and water temperature, recorded continu-
ously at 1-h intervals with Optic StowawayÒ1 datal-
oggers, averaged 8.9 °C (range 2.4–12.9 °C). Dissolved
oxygen was measured semiweekly using a YSIÒ meter
(model 50B, Yellow Springs Instruments Inc., Yellow-
Springs, OH, USA), and averaged 11.74 mg L–1 (range
10.86–12.89 mg L–1).
After days 17, 28, 42 and 56, a single carcass was
randomly selected and removed from each channel.
Carcasses were rinsed several times with water, and
the washings put through 2-mm and 250-lm nested
sieves. The coarser sieve separated out large pieces of
salmon tissue, from which invertebrates were extrac-
ted without magnification and preserved in 95%
EtOH with material from the finer sieve. Tissue pieces
were frozen and later dried to a constant mass at
80 °C to determine the carcass mass remaining. The
material from the finer sieve consisted primarily of
invertebrates and silt, with insufficient salmon tissue
to justify determining its dry mass.
All invertebrates were extracted from each sample,
except large numbers (>1000) of chironomid midge
(Diptera: Chironomidae) larvae that were subsam-
pled (one-quarter). Invertebrates were preserved
with 70% EtOH and identified to family or genus
using Merritt & Cummins (1996). The lengths of
chironomid midge and Zapada (Plecoptera: Nemouri-
dae) larvae, the dominant carcass invertebrates, were
used in conjunction with established mass-length
equations (Smock, 1980; M.S. Wipfli, unpublished
data) to estimate total invertebrate biomass for each
carcass.
Carcasses placed in natural streams
Salmon carcasses were placed into four streams
(Sprout Fork, Spike, Cobble and Margaret Creeks),
between 2 and 10 September 1998. The pink female
salmon used were similar in condition and size
(48–52 cm fork length, 1.25–2.02 kg wet mass and
0.35–0.56 kg dry mass) to those used in artificial
streams. Carcasses were placed in wire mesh cages
(55 · 25 cm, galvanised fencing wire, mesh
size ¼ 5 cm) with a base of fibreglass mesh (mesh
size ¼ 1.6 mm) to facilitate collection and reduce loss
of material during retrieval. In each stream, four cages
were placed within two log jams where carcasses
were already present (D.T. Chaloner, personal obser-
vation). Cages were secured with parachute cord and
positioned so that they remained submerged and
were reasonably protected from vertebrate scaven-
gers. Temperature dataloggers recorded water tem-
perature hourly, and dissolved oxygen and stream
discharge were measured periodically (Table 1). Data-
loggers in Sprout Fork and Cobble Creek recorded
temperature for only part of the time, but data suggest
that all streams were similar.
Carcass cages were left in place for six weeks, long
enough to allow invertebrates to reach their maximum
abundance, based on artificial stream experiments,
and short enough to avoid autumn floods. At the end
of the experiment, carcasses were removed from cages
and processed as described above. Of the original 32
cages, 25 were recovered: eight from Sprout Fork, five
from Spike Creek, five from Margaret Creek, and
seven from Cobble Creek. Bears had tampered with
the others. One cage from Sprout Fork was completely
buried by sand, and therefore was not included in
statistical analyses.
Carcasses occurring naturally in streams
Surveys of salmon carcasses were undertaken three
times between the first sighting of chum salmon (26
July 1998) and two weeks after live pink salmon were
last seen in the streams (28 September 1998). Surveys
took place along a single 200-m reach of Cobble Creek
1The use of trade or firm names in this publication is for reader
information and does not imply endorsement by the U.S.
Department of Agriculture of any product or service.
Mass loss and macroinvertebrate colonisation of salmon carcasses 265
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273

and Margaret Creek, moving in an upstream direction
to avoid encountering the same carcass twice. During
surveys, relative abundance (0–9, 10–99, 100–999, or
>1000 individuals per carcass) was estimated for
invertebrate taxa found on 30 carcasses. Each taxa
was assigned to a particular category (absent, trace,
present, or common) based upon their overall relative
abundance scores and the number of carcasses on
which they were found. The wet mass of 100 carcasses
was also measured and the carcasses returned to the
stream after weighing.
Experimental design and statistical analyses
A randomised complete block design was used for the
artificial stream experiment, with six channels as
blocks and four time periods as treatments. Each time
period was randomly assigned to one of four carcasses
in each block. Response variables were macroinverte-
brate abundance and biomass. Data were natural
logarithm (ln) transformed to meet the assumptions of
analysis of variance (ANOVA) and analysed with
PROC GLM (SAS Institute, 1989) at a ¼ 0.05. Carcass
mass loss in artificial streams was quantitatively
modelled using percent carcass mass remaining and
with best-fit exponential regression curves, as used in
other studies (Minshall et al., 1991; Parmenter &
Lamarra, 1991).
For the natural stream experiment, we analysed the
data using a nested ANOVA, with carcasses nested
within log jams and log jams nested within streams.
Response variables were percent carcass mass
remaining, macroinvertebrate abundance, and macr-
oinvertebrate biomass. Data were ln-transformed to
meet ANOVA assumptions and analysed with PROC
GLM (SAS Institute, 1989) at a ¼ 0.05. Data from the
surveys of naturally occurring carcasses were not
analysed statistically because of the structure and
qualitative nature of the data.
Results
Carcasses placed in artificial streams
Salmon carcasses changed in appearance over time,
although distinct stages were not evident. A thick,
furry mould-like coating developed within two days
of carcasses being placed in channels, which gradu-
ally turned into a dark slime layer as organic material
and silt accumulated on carcasses. The muscle tissue
became paste-like within a month and more fluid with
time, while skin, eggs and internal organs persisted
throughout the experiment. The percent carcass mass
remaining reflected these observations and fitted a
single exponential decay curve (Fig. 1). A double
exponential regression curve was unsuitable given the
sampling intervals and the experimental duration (cf.
Parmenter & Lamarra, 1991).
Of the 23 invertebrate taxa found on carcasses,
chironomid midge and Zapada larvae were the most
abundant taxa, constituting 97% (range 83–99%) and
3% (range <1–13%) of invertebrates found, respect-
ively (Table 2). Invertebrates were found predomin-
antly on the carcass surface but were also found
among the eggs and in the gill and mouth cavities
when carcasses were removed after six to eight weeks.
Chironomid abundance and biomass increased sig-
nificantly over time (P < 0.001, Table 2), whereas
Zapada biomass and abundance did not (P > 0.05,
Table 2) (Fig. 2).
Carcasses placed in natural streams
Salmon carcasses recovered from a single log jam
exhibited a range of percent remaining values (e.g.
8–29%). Some carcasses had missing parts, while
others were fully intact, which partly reflected how
exposed carcasses were within the log jams. How-
ever, the average carcass mass percent remaining for
Fig. 1 Mean carcass mass remaining (percentage of original dry
mass) after sampling salmon carcasses placed in artificial
streams. Error bars represent ±1 SEM.
266 D.T. Chaloner et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273

natural streams (26 ± 2%, mean ± SEM) was similar
to that found in artificial streams after six weeks
(20 ± 2%, mean ± SEM). As in artificial streams,
skin, eggs and internal organs had not decomposed
appreciably, while the muscle tissue was paste-like.
Carcasses were covered with a dense, furry, blue-
gray mould-like layer, firmly attached to the skin,
unlike the slime layer found in artificial streams.
Overall, there were no obvious differences in carcass
appearance among natural streams and no signi-
ficant differences amongst streams in percent
mass remaining (P > 0.05), suggesting that carcass
decomposition was similar amongst streams in
terms of mass loss.
Table 2 Functional feeding group (c-g, collector-gatherer; om, omnivore; p, predator; sc, scraper; sh, shredder; after Merritt &
Cummins, 1996), mean percent relative abundance (calculated by pooling across all time periods or streams), mean actual abundance
(number per carcass), mean biomass (mg per carcass), and A N O V A results (with time or stream as the treatment, n = 24, d.f. = 3, 15) for
benthic macroinvertebrates collected from carcasses placed in artificial or natural streams. Values for biomass given in parentheses
below those for abundance
Artificial streams Natural streams
Functional
feeding Relative AbundanceA N O V AA N O V A
Relative AbundanceA N O V AA N O V A
Taxon group abundance (biomass) F P abundance (biomass) F P
Arthropoda
Insecta
Ephemeroptera* c-g, sc 0.2 1.9 nt†
Baetidae
Baetis c-g, sc 0.8 18.4 10.22 0.024
Other Ephemeroptera‡ c-g, sc 0.4 6.0 nt
Plecoptera
Nemouridae
Zapada sh 2.5 33.5 0.67 0.581 2.2 32.2 0.42 0.749
(15.1) (2.86) (0.072) (31.5) (0.51) (0.696)
Chloroperlidae
Sweltsa p 0.6 8.0 18.26 0.009
Capniidae sh 1.7 19.0 2.56 0.193
Other Plecoptera§,– sh, p 0.1 1.5 nt <0.1 0.3 nt
Coleoptera** several <0.1 0.1 nt 0.1 0.8 nt
Trichoptera†† several 0.4 4.4 nt
Onocosmoecus sh 0.4 5.0 1.07 0.457
Other Trichoptera‡‡ several 0.4 6.2 1.89 0.273
Diptera
Chironomidae several 96.6 2155.8 22.11 <0.001 92.6 1526.5 2.91 0.164
(783.7) (75.85) (<0.001) (251.7) (1.49) (0.346)
Simuliidae cf 0.1 3.8 nt
Other Diptera§§,–– several 0.2 2.2 nt 0.4 5.2 nt
Arachnida
Acari om <0.1 0.3 nt <0.1 0.1 nt
Annelida
Oligochaeta c-g <0.1 0.3 nt 0.2 2.9 nt
All taxa 100.0 2200.2 19.63 <0.001 100.0 1635.6 2.66 0.184
*Baetis, Cinygmula, Paraleptophlebia.
†Not tested.
‡Cinygmula, Drunella doddsi, Ephemerella, Paraleptophlebia, Serratella.
§Artificial streams: Capniidae, Leuctridae, Sweltsa.
–Natural streams: Hesperoperla, Leuctridae.
**Dytiscidae, Elmidae.
††Ecclisomyia, Micrasema, Onocosmoecus, Polycentropus, Rhyacophila.
‡‡Ecclisomyia, Hydropsychidae, Micrasema, Polycentropus, Rhyacophila.
§§Artificial streams: Ceratopogonidae, Empididae, Simuliidae, Tipulidae.
––Natural streams: Ceratopogonidae, Empididae, Tipulidae.
Mass loss and macroinvertebrate colonisation of salmon carcasses 267
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273

Of the 27 invertebrate taxa found on carcasses
(Table 2), chironomid midges, Zapada, Capniidae
(Plecoptera), Baetis (Ephemeroptera: Baetidae), Sweltsa
(Plecoptera: Chloroperlidae), and Onocosmoecus (Tri-
choptera: Limnephilidae) larvae were the most com-
mon. Some invertebrates, especially chironomids,
were found in the gills and mouth and buried within
carcass tissue. As in artificial streams, chironomids
and Zapada were the only taxa found on all carcasses
and constituted 93% (range 83–99%) and 2% (range
<1–5%) of the invertebrates found, respectively. The
low statistical power of this study design meant only
large differences among streams, relative to within-
stream variation (between log jams), would be detect-
able. So although there appeared to be considerable
variation among streams in the abundance of several
taxa (Fig. 3), these differences were only significant
for the abundance of Sweltsa (P ¼ 0.0085) and Baetis
(P ¼ 0.024) (Table 2, Fig. 3). For the abundance of all
other taxa and biomass of chironomids and Zapada, no
significant differences were found among streams
(P > 0.05) (Table 2, Figs 3 and 4).
Carcasses occurring naturally in streams
Naturally occurring carcass material was found in a
variety of forms, reflecting the many ways that fish-
died (e.g. natural death, predator kills) and carcasses
Fig. 2 Mean abundance and biomass estimates of chironomid midge and Zapada larvae found on salmon carcasses placed in
artificial streams. Error bars represent ±1 SEM.
268 D.T. Chaloner et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273

were moved around streams (e.g. scavenging, spates).
Carcass material ranged from complete fish (both
fresh and well decomposed) to tissue fragments. The
more decomposed material was often covered with a
layer of dense, mould-like biofilm, similar to that
found on carcasses artificially placed in natural
streams. The average mass of carcass material found
during the first survey of Margaret Creek (Fig. 5)
reflected partial consumption of chum salmon by
predators. By the first survey of Cobble Creek,
predators left chum carcasses virtually untouched, in
contrast to the smaller pink salmon that began to
appear in streams at this time. Towards the end of the
salmon run, whole carcasses were seen less fre-
quently, while small to medium-sized (5–20-cm
diameter) tissue fragments were more common. The
average mass of carcass material found over time
(Fig. 5) reflected these observations.
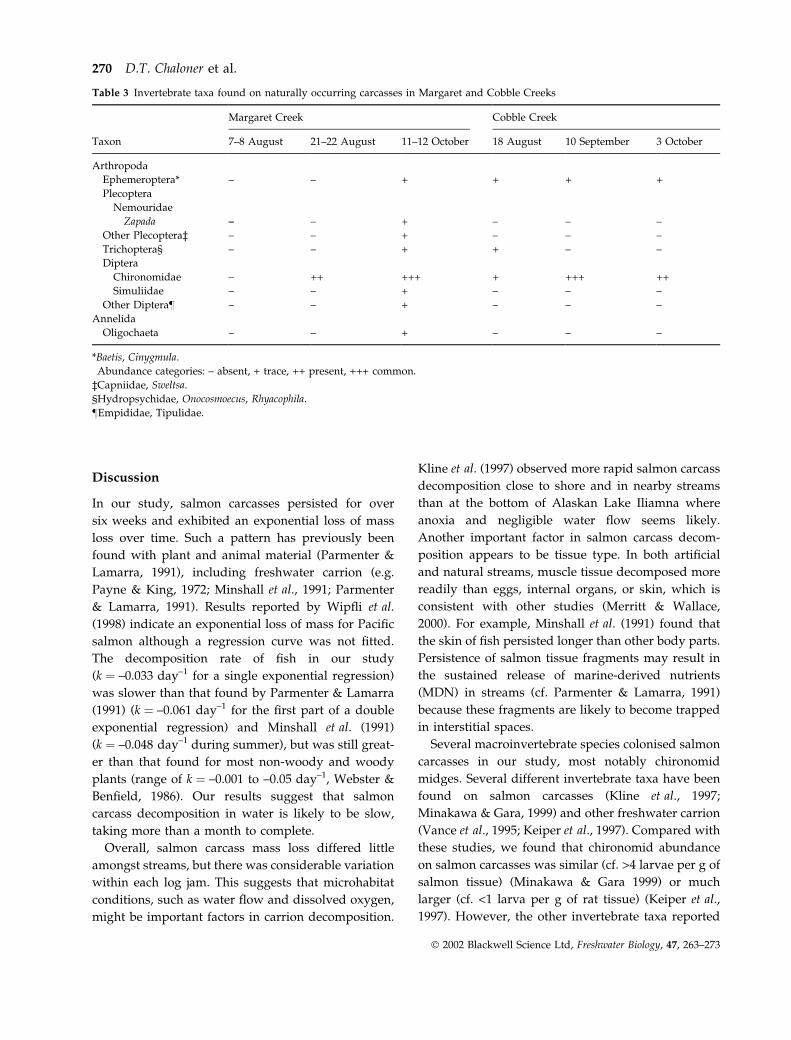
Several invertebrate taxa were found on naturally
occurring salmon carcasses (Table 3), with the most
abundant and common being chironomid larvae,
consistent with both artificial and natural stream
experiments. Zapada larvae were also found, but not
as often as mayflies and caddisflies. Fewer inverte-
brates were observed on carcasses in Cobble Creek
than Margaret Creek, consistent with the invertebrate
abundance found on artificially placed carcasses. In
Margaret Creek, invertebrates were not found on
carcasses in early August, but were present in Octo-
ber. In Cobble Creek, invertebrates were observed on
carcasses on all surveying dates. In both streams,
invertebrates were found in the mouth and on gills,
fin rays, and the surface of carcasses.
Fig. 3 Mean abundance of the most abundant invertebrate
taxa found on salmon carcasses artificially placed in natural
streams for six weeks. Error bars represent ±1 SEM.
Fig. 4 Mean biomass estimates of chironomid midge (j) and
Zapada (h) larvae found on salmon carcasses artificially placed
in natural streams for 6 weeks. Error bars represent ±1 SEM.
Fig. 5 Mean wet mass of carcass material found naturally in
Cobble (–j–) and Margaret (–r–) Creeks. Error bars represent
±1 SEM.
Mass loss and macroinvertebrate colonisation of salmon carcasses 269
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273
Discussion
In our study, salmon carcasses persisted for over
six weeks and exhibited an exponential loss of mass
loss over time. Such a pattern has previously been
found with plant and animal material (Parmenter &
Lamarra, 1991), including freshwater carrion (e.g.
Payne & King, 1972; Minshall et al., 1991; Parmenter
& Lamarra, 1991). Results reported by Wipfli et al.
(1998) indicate an exponential loss of mass for Pacific
salmon although a regression curve was not fitted.
The decomposition rate of fish in our study
(k ¼ –0.033 day–1 for a single exponential regression)
was slower than that found by Parmenter & Lamarra
(1991) (k ¼ –0.061 day–1 for the first part of a double
exponential regression) and Minshall et al. (1991)
(k ¼ –0.048 day–1 during summer), but was still great-
er than that found for most non-woody and woody
plants (range of k ¼ –0.001 to –0.05 day–1, Webster &
Benfield, 1986). Our results suggest that salmon
carcass decomposition in water is likely to be slow,
taking more than a month to complete.
Overall, salmon carcass mass loss differed little
amongst streams, but there was considerable variation
within each log jam. This suggests that microhabitat
conditions, such as water flow and dissolved oxygen,
might be important factors in carrion decomposition.
Kline et al. (1997) observed more rapid salmon carcass
decomposition close to shore and in nearby streams
than at the bottom of Alaskan Lake Iliamna where
anoxia and negligible water flow seems likely.
Another important factor in salmon carcass decom-
position appears to be tissue type. In both artificial
and natural streams, muscle tissue decomposed more
readily than eggs, internal organs, or skin, which is
consistent with other studies (Merritt & Wallace,
2000). For example, Minshall et al. (1991) found that
the skin of fish persisted longer than other body parts.
Persistence of salmon tissue fragments may result in
the sustained release of marine-derived nutrients
(MDN) in streams (cf. Parmenter & Lamarra, 1991)
because these fragments are likely to become trapped
in interstitial spaces.
Several macroinvertebrate species colonised salmon
carcasses in our study, most notably chironomid
midges. Several different invertebrate taxa have been
found on salmon carcasses (Kline et al., 1997;
Minakawa & Gara, 1999) and other freshwater carrion
(Vance et al., 1995; Keiper et al., 1997). Compared with
these studies, we found that chironomid abundance
on salmon carcasses was similar (cf. >4 larvae per g of
salmon tissue) (Minakawa & Gara 1999) or much
larger (cf. <1 larva per g of rat tissue) (Keiper et al.,
1997). However, the other invertebrate taxa reported
Table 3 Invertebrate taxa found on naturally occurring carcasses in Margaret and Cobble Creeks
Margaret Creek Cobble Creek
Taxon 7–8 August 21–22 August 11–12 October 18 August 10 September 3 October
Arthropoda
Ephemeroptera* –† – + + + +
Plecoptera
Nemouridae
Zapada – – + – – –
Other Plecoptera‡ – – + – – –
Trichoptera§ – – + + – –
Diptera
Chironomidae – ++ +++ + +++ ++
Simuliidae – – + – – –
Other Diptera– – – + – – –
Annelida
Oligochaeta – – + – – –
*Baetis, Cinygmula.
†Abundance categories: – absent, + trace, ++ present, +++ common.
‡Capniidae, Sweltsa.
§Hydropsychidae, Onocosmoecus, Rhyacophila.
–Empididae, Tipulidae.
270 D.T. Chaloner et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273

by previous studies (e.g. Kline et al., 1997; Minakawa
& Gara, 1999), such as stoneflies, notably Zapada and
Sweltsa, and caddisflies, notably Ecclisomyia and
Onocosmoecus, we found at much smaller abundances.
Invertebrates were also much less abundant on
naturally occurring carcasses than on those artificially
placed in natural or artificial streams. Focusing
surveys on carcasses trapped in log jams for several
weeks, may have produced results more similar to
those for other parts of the study because inverte-
brates are likely to be dislodged from carcasses that
are washed downstream.
Salmon carcasses could be important microhabitats
for chironomid midges and Zapada, representing
potential ‘ecological hotspots’ of invertebrate abun-
dance in streams. Wipfli et al. (1998) found that
chironomid abundance in the mineral substrate of
mesocosm pools declined after an early peak to about
12 000 larvae m–2. In our study, using the same
mesocosm, chironomid abundance increased steadily
to about 84 000 larvae m–2 on carcasses. In contrast,
Zapada abundance progressively increased through-
out the study by Wipfli et al. (1998), reaching about
700 larvae m–2 in mineral substrate as compared with
about 3700 larvae m–2 on carcasses in our study.
Chironomid larvae can reach larger population den-
sities in organic substrates (e.g. 250 000 larvae m–2,
Armitage, Cranston & Pinder, 1995), even in the
presence of the low dissolved oxygen levels often
associated with submerged carrion (Merritt &
Wallace, 2000).
Organisms are believed to use carrion as a source of
food and/or shelter (Merritt & Wallace, 2000). Inver-
tebrate consumption of salmon carcass material has
been indicated by anecdotal observations (Elliott &
Bartoo, 1981; Minakawa & Gara, 1999) and stable
isotope analyses (Bilby et al., 1996; Kline et al., 1997),
and seems likely for invertebrates belonging to the
‘shredder’ functional feeding group, such as Zapada,
that chew, burrow, or mine into coarse particulate
organic material (Merritt & Cummins, 1996). Inver-
tebrate colonists could help fragment carcass material,
promote nutrient leaching, and oxygenate anoxic
subsurface areas, as has been shown with other
organic substrates (Parmenter & Lamarra, 1991).
Minshall et al. (1991) suggested that invertebrates
could also transfer decomposing microbes between
carcasses, and from the surface to the carcass interior.
The thick mould-like microbial development
observed on carrion (e.g. Minshall et al., 1991; Vance
et al., 1995; Kline et al., 1997; Merritt & Wallace, 2000;
this study) is probably involved in decomposition and
could sequester nutrients leaching from carcasses. The
biofilm associated with salmon tissue and its interac-
tion with carcass invertebrates warrants study, given
what little is known about biofilm associated with
animal tissue (Stevenson, Bothwell & Lowe, 1996).
Salmon carcass invertebrates may facilitate the
transfer of MDN to higher trophic levels, such as
predatory fishes and insectivorous birds. Carcass
tissue is an important food source for fish (Bilby et al.,
1996), mammals and birds (Willson et al., 1998) but
carcasses become fragmented, buried, or washed away
(Cederholm et al., 1989). In these situations, carcass
invertebrates could convert tissue and associated
biofilm into food for other invertebrates and higher
trophic levels, as invertebrate tissue and faecal
material. Chironomids can produce prodigious
amounts of faecal material and are important food
item for many predators (Armitage et al., 1995). Car-
cass invertebrates, because of their proximity to the
MDN source, may also assimilate more MDN than
invertebrates in the same stream but on different
substrate types. Early invertebrate colonists, although
small in biomass, might be disproportionately
important in this respect because of their presence
during the large initial loss of mass. Cederholm et al.
(1989) emphasised the importance of physical MDN
retention devices (e.g. large wood structures) in
streams. Carcass biofilm and invertebrates may repre-
sent biological MDN retention devices also important
in streams but at a smaller scale, similar to the chemical
sorption processes discussed by Bilby et al. (1996).
The processes and mechanisms by which Pacific
salmon influence ecosystems are complex and involve
many factors, including the rate at which carcasses
lose mass and become colonised by invertebrates. Our
study indicates that salmon carcasses lose mass in a
way similar to other types of organic material and are
colonised by invertebrates in high densities. Better
understanding of salmon decomposition in freshwa-
ter-riparian ecosystems will contribute to the devel-
opment of more holistic Pacific salmon management
and restoration strategies, which can include placing
hatchery carcasses in streams (Ashley & Slaney, 1997).
The design of such projects would ideally reflect the
influence that salmon have upon all trophic levels in
freshwater-riparian food webs.
Mass loss and macroinvertebrate colonisation of salmon carcasses 271
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273

Acknowledgments
We thank the following: Ketchikan Ranger District
(USDA Forest Service) personnel for logistical support
and use of Margarita Bay camp facilities; Rachel
Baker, Derek Busch, Kim Frangos, Adam Herron,
John Hudson, Gary Lamberti, Kristine Martin, Rich-
ard Merritt, Warren Mitchell, Jeff Nichols, David
Sperry, and Brenda Wright for their enthusiastic help,
advice, and support; Timothy Max for helpful statis-
tical advice; Patrick Armitage, Mason Bryant, John
Hudson, Gary Lamberti, Richard Merritt, William
Perry, Deanna Stouder, and Roger Wotton for their
many helpful suggestions on the manuscript and
stimulating discussions about dead fish; Linda Daniel
for important editorial guidance; two anonymous
reviewers for their useful comments. This research
was supported by the Pacific Northwest Research
Station (USDA Forest Service) (Cooperative Research
Agreement: PNW 96-3012-2-CA) and by a grant from
the USDA-CSREES National Research Initiative Com-
petitive Grants Program (99-35101-8592).
References
Armitage P.D., Cranston P.S. & Pinder L.C.V. (Eds)
(1995) The Chironomidae: Biology and Ecology of Non-
Biting Midges. Chapman & Hall, New York.
Ashley K.I. & Slaney P.A. (1997) Accelerating recovery of
stream and pond productivity by low-level nutrient
enrichment. In: Fish Habitat Rehabilitation Procedures
(Eds P.A. Slaney & D. Zaldokas), pp. 239–262. Water-
shed Restoration Technical Circ., no. 9, British Colum-
bia Ministry of Environment, Lands and Parks and
Ministry of Forests, Vancouver.
Baker T.T., Wertheimer A.C., Burkett R.D., Dunlap R.,
Eggers D.M., Fritts E.I., Gharrett A.J., Holmes R.A. &
Wilmot R.L. (1996) Status of Pacific salmon and
steelhead escapments in southeastern Alaska. Fisheries,
21, 6–18.
Bilby R.E., Fransen B.R. & Bisson P.A. (1996) Incorpor-
ation of nitrogen and carbon from spawning coho
salmon into the trophic system of small streams:
evidence from stable isotopes. Canadian Journal of
Fisheries and Aquatic Science, 53, 164–173.
Britton J.C. & Morton B. (1994) Marine carrion and
scavengers. Oceanography and Marine Biology: An
Annual Review, 32, 369–434.
Bryant M.D., Frenette B.J. & McCurdy S.J. (1999) Colon-
ization of a watershed by anadromous salmonids
following the installation of a fish ladder in Margaret
Creek, Southeast Alaska. North American Journal of
Fisheries Management, 19, 1129–1136.
Cederholm C.J., Houston D.B., Cole D.L. & Scarlett W.J.
(1989) Fate of coho salmon (Oncorhynchus kisutch)
carcasses in spawning streams. Canadian Journal of
Fisheries Aquatic Science, 46, 1347–1355.
Cederholm C.J., Kunze M.D., Murota T. & Sibatani A.
(1999) Pacific salmon carcasses: essential contributions
of nutrients and energy for aquatic and terrestrial
ecosystems. Fisheries, 24, 6–15.
Cummins K.W. (1973) Trophic relations of aquatic
insects. Annual Review of Entomology, 18, 183–206.
Elliott S.T. & Bartoo R. (1981) Relation of larval Polyped-
ilum (Diptera: Chironomidae) to pink salmon eggs and
alevins in an Alaskan stream. Progressive Fish Culturist,
43, 220–221.
Ellis R.J. (1970) Alloperla stonefly nymphs: predators or
scavengers on salmon eggs and alevins? Transactions of
the American Fisheries Society, 99, 677–683.
Hanski I. (1987) Nutritional ecology of dung- and carrion-
feeding insects. In: Nutritional Ecology of Insects, Mites,
Spiders and Related Invertebrates (Eds F. Slansky & J.G.
Rodriguez), pp. 837–884. Wiley, New York.
Haskell N.H., McShaffrey D.G., Hawley D.A., Williams
R.E. & Pless J.E. (1989) Use of aquatic insects in
determining submersion interval. Journal of Forensic
Science, 34, 622–632.
Keiper J.B., Chapman E.G. & Foote B.A. (1997) Midge
larvae (Diptera: Chironomidae) as indicators of post-
mortem submersion interval of carcasses in a wood-
land stream: a preliminary report. Journal of Forensic
Science, 42, 1074–1079.
Kline T.C., Goering J.J. & Piorkowski R.J. (1997) The
effect of salmon carcasses on Alaskan freshwaters. In:
Freshwaters of Alaska: Ecological Syntheses (Eds A.M.
Milner & M.W. Oswood), pp. 179–204. Springer-Ver-
lag, New York.
Merritt R.W. & Cummins K.W. (Eds) (1996) An Introduc-
tion to the Aquatic Insects of North America. Kendall/
Hunt, Dubuque.
Merritt R.W. & Wallace J.R. (2000) The role of aquatic
insects in forensic investigations. In: Forensic Entomol-
ogy: The Utility of Arthropods in Legal Investigations (Eds
J.H. Byrd & J.L. Castner), pp. 177–222. CRC Press, Boca
Raton.
Minakawa N. & Gara R.I. (1999) Ecological effects of a
chum salmon (Oncorhynchus keta) spawning run in a
small stream of the Pacific Northwest. Journal of
Freshwater Ecology, 14, 327–335.
Minshall G.W., Hitchcock E. & Barnes J.R. (1991)
Decomposition of rainbow trout (Oncorhynchus mykiss)
carcasses in a forest stream ecosystem inhabited only
272 D.T. Chaloner et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273

by non-anadromous fish populations. Canadian Journal
of Fisheries and Aquatic Science, 48, 191–195.
Parmenter R.R. & Lamarra V.A. (1991) Nutrient cycling
in a fresh-water marsh: the decomposition of fish and
waterfowl carrion. Limnology and Oceanography, 36,
976–987.
Payne J.A. & King E.W. (1972) Insect succession and
decomposition of pig carcasses in water. Journal of the
Georgia Entomological Society, 7, 153–162.
Polis G.A., Anderson W.B. & Holt R.D. (1997) Toward an
integration of landscape and food web ecology: the
dynamics of spatially subsidized food webs. Annual
Review of Ecology and Systematics, 28, 289–316.
SAS Institute Inc. (1989) SAS/STAT User’s Guide, Version
6, 4th edn., Vol. 2. SAS Institute, Cary.
Schuldt J.A. & Hershey A.E. (1995) Effect of salmon
carcass decomposition on Lake Superior tributary
streams. Journal of the North American Benthological
Society, 14, 259–268.
Smock L.A. (1980) Relationships between body size and
biomass of aquatic insects. Freshwater Biology, 10, 375–
383.
Stevenson R.J., Bothwell M.L. & Lowe R.L. (Eds) (1996)
Algal Ecology. Academic Press, San Diego, CA.
Vance G.M., Van Dyk J.K. & Rowley W.A. (1995) A
device for sampling aquatic insects associated with
carrion in water. Journal of Forensic Science, 40, 479–482.
Webster J.R. & Benfield E.F. (1986) Vascular plant
breakdown in freshwater ecosystems. Annual Review
of Ecology and Systematics, 17, 567–94.
Willson M.F., Gende S.M. & Marston B. (1998) Fishes and
the forest: expanding perspectives on fish–wildlife
interactions. Bioscience, 48, 455–462.
Wipfli M.S., Hudson J.P. & Caouette J.P. (1998) Influence
of salmon carcasses on stream productivity: response
of biofilm and benthic macroinvertebrates in south-
eastern Alaska, USA. Canadian Journal of Fisheries and
Aquatic Science, 55, 1503–1511.
Wipfli M.S., Hudson J.P., Chaloner D.T. & Caouette J.P.
(1999) The influence of salmon spawner densities on
stream productivity in Southeast Alaska. Canadian Jour-
nal of Fisheries and Aquatic Science, 56, 1600–1611.
(Manuscript accepted 28 May 2001)
Mass loss and macroinvertebrate colonisation of salmon carcasses 273
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 263–273




















