
MASKING INTERFERENCE AND THE EVOLUTION OF THE ACOUSTIC COMMUNICATION SYSTEM IN THE AMAZONIAN...
14
1874 2006 The Society for the Study of Evolution. All rights reserved. Evolution, 60(9), 2006, pp. 1874–1887 MASKING INTERFERENCE AND THE EVOLUTION OF THE ACOUSTIC COMMUNICATION SYSTEM IN THE AMAZONIAN DENDROBATID FROG ALLOBATES FEMORALIS ADOLFO AME ´ ZQUITA, 1,2,3 WALTER HO ¨ DL, 3,4 ALBERTINA PIMENTEL LIMA, 5 LINA CASTELLANOS, 1,6 LUCIANA ERDTMANN, 1,5,7 AND MARIA CARMOZINA DE ARAU ´ JO 5,8 1 Department of Biological Sciences, University of Los Andes, AA 4976, Bogota ´, Colombia 2 E-mail: [email protected] 3 Department of Evolutionary Biology, University of Vienna, Althanstrasse 14, A 1090, Vienna, Austria 4 E-mail: [email protected] 5 Instituto Nacional de Pesquisas da Amazonia, Ecologia, Manaus, Brazil E-mail: [email protected] 6 E-mail: [email protected] 7 E-mail: [email protected] 8 E-mail: [email protected] Abstract. The efficacy of communication relies on detection of species-specific signals against the background noise. Features affecting signal detection are thus expected to evolve under selective pressures represented by masking noise. Spectral partitioning between the auditory signals of co-occurring species has been interpreted as the outcome of the selective effects of masking interference. However, masking interference depends not only on signal’s frequency but on receiver’s range of frequency sensitivity; moreover, selection on signal frequency can be confounded by selection on body size, because these traits are often correlated. To know whether geographic variation in communication traits agrees with predictions about masking interference effects, we tested the hypothesis that variation in the male-male communication system of the Amazonian frog, Allobates femoralis, is correlated with the occurrence of a single species calling within an overlapping frequency range, Epipedobates trivittatus. We studied frogs at eight sites, four where both species co-occur and four where A. femoralis occurs but E. trivittatus does not. To study the sender component of the communication system of A. femoralis and to describe the use of the spectral range, we analyzed the signal’s spectral features of all coactive species at each site. To study the receiver component, we derived frequency-response curves from playback experiments conducted on territorial males of A. femoralis under natural conditions. Most geographic variation in studied traits was correlated with either call frequency or with response frequency range. The occurrence of E. trivittatus significantly predicted narrower and asymmetric frequency-response curves in A. femoralis, without concomitant differences in the call or in body size. The number of acoustically coactive species did not significantly predict variation in any of the studied traits. Our results strongly support that the receiver but not the sender component of the communication system changed due to masking interference by a single species. Key words. Acoustic communication, Dendrobatidae, geographic variation, masking interference, playback experi- ments, spectral partitioning. Received February 10, 2006. Accepted June 26, 2006. Communication in sexually reproducing animals mediates species recognition and mating. Therefore, the study of the selective forces that act on communication traits improves our understanding of the mechanisms behind evolutionary processes such as adaptation, speciation, and sexual selection (Blair 1964; Jones 1997; Ryan and Rand 1993; Boughman 2002). Research on the evolution of mating signals has his- torically focused on the correlates of geographic variation in signal features (e.g., Hunter and Krebs 1979; Nelson 1989; Loftus-Hill and Littlejohn 1992; Irwin 2000), and on the relationship between signal features, receivers’ (usually fe- male) preference, and breeding success of senders and re- ceivers (e.g., Houde and Endler 1990; Welch et al. 1998; Widemo and Sæther 1999; Grether 2000). However, before receivers decode and react to signals, they must detect them against the background noise (Forrest 1994; Bradbury and Vehrencamp 1998). Signal detection may impose selective constraints on both signal production and signal perception mechanisms (Ryan and Brenowitz 1985; Narins and Zelick 1988; Anderson and McGregor 1999), and also in commu- nication systems where receivers are not females, for ex- ample, males (agonistic interactions), juveniles (parent-off- spring communication), and predators (aposematism). Among acoustically communicating species that breed to- gether, heterospecific signals often represent the noise source interfering with signal detection (Otte 1974; Ryan and Bren- owitz 1985; Gerhardt and Klump 1988; Narins and Zelick 1988). Therefore, calling within species-specific frequency ranges (spectral partitioning) has been interpreted as the evo- lutionary outcome of masking interference (Ho ¨dl 1977; Dre- wry and Rand 1983; Duellman and Pyles 1983, Wilczynski et al. 1993; Narins 1995; Lu ¨ddecke et al. 2000; Chek et al. 2003; for discussions on partitioning beyond the spectral di- mension of the signals, see Nelson 1989; Wollerman and Wiley 2002). However, the probability of masking interfer- ence depends not only on the signal’s frequency; the degree of tuning in receivers’ ears determines how much of the het- erospecific noise is filtered out (Wilczynski et al. 1993; Hall 1994; Ro ¨mer 1998). Moreover, because body size constrains signal’s frequency within and across several animal taxa (Martin 1972; Ryan and Brenowitz 1985; Bennet-Clark 1998), selection on signal frequency can be confounded by selection on body size. One way to evidence masking interference effects on com- munication is to compare populations of the same species
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of MASKING INTERFERENCE AND THE EVOLUTION OF THE ACOUSTIC COMMUNICATION SYSTEM IN THE AMAZONIAN...

1874
� 2006 The Society for the Study of Evolution. All rights reserved.
Evolution, 60(9), 2006, pp. 1874–1887
MASKING INTERFERENCE AND THE EVOLUTION OF THE ACOUSTICCOMMUNICATION SYSTEM IN THE AMAZONIAN DENDROBATID FROG
ALLOBATES FEMORALIS
ADOLFO AMEZQUITA,1,2,3 WALTER HODL,3,4 ALBERTINA PIMENTEL LIMA,5 LINA CASTELLANOS,1,6
LUCIANA ERDTMANN,1,5,7 AND MARIA CARMOZINA DE ARAUJO5,8
1Department of Biological Sciences, University of Los Andes, AA 4976, Bogota, Colombia2E-mail: [email protected]
3Department of Evolutionary Biology, University of Vienna, Althanstrasse 14, A 1090, Vienna, Austria4E-mail: [email protected]
5Instituto Nacional de Pesquisas da Amazonia, Ecologia, Manaus, BrazilE-mail: [email protected]
6E-mail: [email protected]: [email protected]: [email protected]
Abstract. The efficacy of communication relies on detection of species-specific signals against the background noise.Features affecting signal detection are thus expected to evolve under selective pressures represented by masking noise.Spectral partitioning between the auditory signals of co-occurring species has been interpreted as the outcome of theselective effects of masking interference. However, masking interference depends not only on signal’s frequency buton receiver’s range of frequency sensitivity; moreover, selection on signal frequency can be confounded by selectionon body size, because these traits are often correlated. To know whether geographic variation in communication traitsagrees with predictions about masking interference effects, we tested the hypothesis that variation in the male-malecommunication system of the Amazonian frog, Allobates femoralis, is correlated with the occurrence of a single speciescalling within an overlapping frequency range, Epipedobates trivittatus. We studied frogs at eight sites, four whereboth species co-occur and four where A. femoralis occurs but E. trivittatus does not. To study the sender componentof the communication system of A. femoralis and to describe the use of the spectral range, we analyzed the signal’sspectral features of all coactive species at each site. To study the receiver component, we derived frequency-responsecurves from playback experiments conducted on territorial males of A. femoralis under natural conditions. Mostgeographic variation in studied traits was correlated with either call frequency or with response frequency range. Theoccurrence of E. trivittatus significantly predicted narrower and asymmetric frequency-response curves in A. femoralis,without concomitant differences in the call or in body size. The number of acoustically coactive species did notsignificantly predict variation in any of the studied traits. Our results strongly support that the receiver but not thesender component of the communication system changed due to masking interference by a single species.
Key words. Acoustic communication, Dendrobatidae, geographic variation, masking interference, playback experi-ments, spectral partitioning.
Received February 10, 2006. Accepted June 26, 2006.
Communication in sexually reproducing animals mediatesspecies recognition and mating. Therefore, the study of theselective forces that act on communication traits improvesour understanding of the mechanisms behind evolutionaryprocesses such as adaptation, speciation, and sexual selection(Blair 1964; Jones 1997; Ryan and Rand 1993; Boughman2002). Research on the evolution of mating signals has his-torically focused on the correlates of geographic variation insignal features (e.g., Hunter and Krebs 1979; Nelson 1989;Loftus-Hill and Littlejohn 1992; Irwin 2000), and on therelationship between signal features, receivers’ (usually fe-male) preference, and breeding success of senders and re-ceivers (e.g., Houde and Endler 1990; Welch et al. 1998;Widemo and Sæther 1999; Grether 2000). However, beforereceivers decode and react to signals, they must detect themagainst the background noise (Forrest 1994; Bradbury andVehrencamp 1998). Signal detection may impose selectiveconstraints on both signal production and signal perceptionmechanisms (Ryan and Brenowitz 1985; Narins and Zelick1988; Anderson and McGregor 1999), and also in commu-nication systems where receivers are not females, for ex-ample, males (agonistic interactions), juveniles (parent-off-spring communication), and predators (aposematism).
Among acoustically communicating species that breed to-gether, heterospecific signals often represent the noise sourceinterfering with signal detection (Otte 1974; Ryan and Bren-owitz 1985; Gerhardt and Klump 1988; Narins and Zelick1988). Therefore, calling within species-specific frequencyranges (spectral partitioning) has been interpreted as the evo-lutionary outcome of masking interference (Hodl 1977; Dre-wry and Rand 1983; Duellman and Pyles 1983, Wilczynskiet al. 1993; Narins 1995; Luddecke et al. 2000; Chek et al.2003; for discussions on partitioning beyond the spectral di-mension of the signals, see Nelson 1989; Wollerman andWiley 2002). However, the probability of masking interfer-ence depends not only on the signal’s frequency; the degreeof tuning in receivers’ ears determines how much of the het-erospecific noise is filtered out (Wilczynski et al. 1993; Hall1994; Romer 1998). Moreover, because body size constrainssignal’s frequency within and across several animal taxa(Martin 1972; Ryan and Brenowitz 1985; Bennet-Clark1998), selection on signal frequency can be confounded byselection on body size.
One way to evidence masking interference effects on com-munication is to compare populations of the same species

1875MASKING INTERFERENCE IN FROG COMMUNICATION
across different acoustic environments. Although the geo-graphic variation approach has been strongly recommended(Romer 1998; Wilczynski and Ryan 1999; Gerhardt and Hub-er 2002), the empirical evidence is still lacking. Such evi-dence should indicate that both signal production and signalperception follow rules that maximize signal contrast againstlocal noise sources. The breeding aggregations of acousti-cally communicating anurans provide valuable research mod-els to test masking interference effects (Narins and Zelick1988; Greenfield 1994; Wiley 1994; Gerhardt and Huber2002, Wollerman and Wiley 2002). Heterospecific calls canseverely impair signal detection in frogs (Gerhardt andKlump 1988; Schwartz and Gerhardt 1989; Narins 1992;Schwartz 1993; Wiley 1994; Wollerman 1999). Also, malesof several phylogenetically distant species breeding nearstreams and waterfalls, both sources of low-frequency noise,converge in producing unusually high-frequency calls andamazingly similar visual displays (Hodl and Amezquita2001). Nonetheless, the question of whether the communi-cation system of a single species can geographically divergebecause of masking interference is not yet resolved. We com-pared here the communication system of a wide-ranging frogspecies in localities with and without another species whosesignals are potential sources of masking interference.
The dendrobatid frog Allobates (Phyllobates in Silverstone1976; Epipedobates in Myers 1987; Allobates in Zimmermannand Zimmermann 1988) femoralis (Anura: Dendrobatidae) iswidely distributed throughout the Amazon basin, includingareas among which the number of acoustically coactive spe-cies varies from two to eight (Toft and Duellman 1979; Rod-rıguez and Cadle 1990; Zimmerman and Rodrigues 1990; W.Hodl and A. Amezquita, pers. obs.). The possession of aterritory is the usual precondition for female attraction(Roithmair 1992) and males signal territory ownership toother males by uttering advertisement calls from slightly el-evated positions on the forest floor. Vocally active intrudersare approached and attacked by territorial males (Hodl 1982,1987). The phonotactic approach can be elicited experimen-tally by playing back synthetic advertisement calls (Hodl1987; Narins et al. 2003, Hodl et al. 2004). Based on thedistribution of male responses towards synthetic calls of var-ious frequencies, a frequency-response curve can be derived(Amezquita et al. 2005; Rojas et al. 2006).
The present study aimed at testing whether geographic var-iation in the male-male communication system of A. femoralisagrees with predictions of masking interference effects. Wechose Epipedobates trivittatus as the potentially masking spe-cies because a previous study at a single site (Amezquita etal. 2005) had shown that A. femoralis and E. trivittatus callsoverlap in frequency, that calling activity of both speciesconcide, and that males of A. femoralis react less towards‘‘homospecific’’ synthetic calls within the frequency rangeof E. trivittatus calls. We specifically predict that commu-nication traits of A. femoralis should be correlated with theoccurrence of E. trivittatus, in a way that minimizes between-species spectral overlap (Fig. 1A). To test this prediction: (1)We compared male calls and male phonotactic reactionsamong eight sites, four in which both species co-occur andfour where A. femoralis occurs but E. trivittatus does not. (2)Because the spectral range available for acoustic communi-
cation may also depend on how many species are sharing it(Hodl 1977; Drewry and Rand 1983; Duellman and Pyles1983, Nelson 1989; Chek et al. 2003), we also tested whetherthe number of acoustically coactive species is correlated withvariation in the communication traits of A. femoralis. (3)Because signal production and signal perception mechanismsneed not evolve in tandem (i.e., as a unit; Ryan et al. 1992;Arak and Enquist 1993; Wiley 1994; Wilczynski and Ryan1999), we tested for geographic covariation between call fre-quency traits, response frequency traits, and body size. (4)Moreover, because geographic differentiation may reflect theeffect of distance on environmental similarity and gene flowbetween populations (Sokal 1979; Endler 1983; Legendre etal. 2002), we tested for distance effects on the subsets ofcovarying traits.
METHODS
We studied the main auditory signal of A. femoralis andthe acoustically coactive species, as well as the phonotacticresponse of male A. femoralis, at eight sites distributedthroughout the Amazon basin (Fig. 2, Table 1). Every sitewas visited at the beginning of the rainy season when thebreeding activity of most diurnally calling species is high(Schluter 1980; Aichinger 1987; Gottsberger and Gruber2004). Between January 2002 and February 2004, we con-ducted field journeys lasting between 12 and 27 days each.Most calling males of A. femoralis were found at the edgeof terra firma forest or associated with forest gaps; we didnot see differences among study populations in microhabitatstructure that may have covaried with our experimental fac-tors. To minimize our impact on frogs’ motivational stateand acoustic environment, we followed at each site the sameprotocol: (1) we characterized the advertisement calls of thepopulation of A. femoralis by recording a preliminary sampleof males without capturing them; (2) we analyzed the dataon call characteristics to synthesize stimulus calls; (3) weconducted playback experiments to test the effect of call fre-quency on the probability of male response; (4) we identifiedthe acoustically coactive species by recording the acousticenvironment at the sites and times of day when males of A.femoralis called; (5) we recorded calls of each acousticallycoactive species; (6) we recorded a new sample of A. fe-moralis calls, captured the males, and measured their bodysize.
Call Recording and Analysis
To characterize signals, we tape recorded (Sony WM D6Crecorder) between three and 34 spontaneous and consecutivecalls with a microphone (AKG D-190-E, AKG AcousticsGmbH, Vienna Austria; Shure BG4.1 or ME-62/K6, Senn-heiser Electronic GmbH, Wedemark, Germany) positioned ata distance of 0.5 m to 1.5 m in front of each male. Recordingswere digitized at 22 kHz and spectral parameters of the callswere analyzed calculating power spectra (Window: Black-man, DFT: 2048 samples, 3 dB filter bandwidth: 18.5 Hz)on Canary 1.2.4 software (Charif et al. 1995). A subset ofthree calls per male was analyzed and each measured param-eter was then averaged to represent the smallest statisticalunit of analysis. According to our preliminary analyses and

1876 ADOLFO AMEZQUITA ET AL.
FIG. 1. (A) Schematic representation of two predictions about masking interference effects (arrows) of the advertisement call ofEpipedobates trivittatus (grey rectangle) on the communication system (bell-shaped curves) of Allobates femoralis. Due to maskinginterference, the communication system of A. femoralis might have experienced a shift upwards along the frequency axis (above) or anasymmetric decrease in the spectral bandwidth (Amezquita et al. 2005). (B) Actual frequency traits derived from bell-shaped curves(either the call power spectrum or the frequency-response curve) and used to test the predictions. CLF, call low frequency; CPF, callpeak frequency; CFR, call frequency range; CHF, call high frequency; RLF, response low frequency; RPF, response peak frequency;RFR, response frequency range; RHF, response high frequency.
a parallel study (H. Gasser, A. Amezquita, W. Hodl, unpubl.data), the within-male coefficient of variation ranged from0.7% (call duration and spectral properties of the call) to 3%(note duration).
We analyzed calls of A. femoralis to prepare the syntheticstimuli necessary for playback experiments and to charac-terize the populations. Each call was described by its temporal(call duration, intercall interval, note duration, internote in-tervals, Fig. 3, inset) and spectral (peak, low and high fre-quency, frequency range, Fig. 1B) parameters. Low and highfrequency were measured at 20 dB (re 20 �Pa) below thepeak intensity, the value at which the signal energy could
still be clearly distinguished from background noise in ourrecordings. With the exceptions explained in some detail, theterminology and procedures for measuring call parametersfollow Cocroft and Ryan (1995).
We also measured the signal’s frequency traits of theacoustically coactive species to describe their potential formasking interference. If the signal consisted of multiple unitsproduced within a similar frequency range, we analyzed theunit with the highest energy content (e.g., Colostethus trili-neatus, Leticia, Fig. 3). If the signal contained two or moreappreciable harmonics (e.g., Adenomera andreae, Fig. 3), weanalyzed the harmonic with the highest energy content.

1877MASKING INTERFERENCE IN FROG COMMUNICATION
FIG. 2. Distribution of the eight study sites throughout the Amazon basin. Shaded area denotes elevation above 700 m.
TABLE 1. Location of study sites (latitude, longitude), number of species (nSPP), and occurrence of Epipedobates trivittatus (0 � no,1 � yes); frequency traits of Allobates femoralis calls (n, number of recorded males; CLF, call low frequency; CPF, call peak frequency;CHF, call high frequency; mean � SD is given) and characteristics of the playback experiments on A. femoralis (Males, number of testedmales; stimuli, number of different stimuli; MF, minimum–maximum values of midfrequency of the synthetic stimuli).
Site Coordinates (deg) nSPP(E.t.) n CLF (kHz) CPF (kHz) CHF (kHz) Males Stimuli MF (kHz)
Hileia �3.1977, �60.4425 3 (0) 15 2.50 � 0.12 3.18 � 0.15 3.83 � 0.15 27 30 1.00–5.52Arataı 3.9907, �52.5901 3 (0) 15 2.86 � 0.14 3.40 � 0.18 3.97 � 0.17 23 23 1.36–5.18Ducke �2.9333, �59.9744 4 (0) 16 2.32 � 0.10 2.93 � 0.18 3.43 � 0.14 23 25 0.97–4.84Treviso �3.1491, �54.8403 7 (0) 16 2.88 � 0.09 3.39 � 0.12 4.04 � 0.07 27 27 1.11–5.35Leticia �4.1233, �69.9491 4 (1) 35 2.64 � 0.12 3.16 � 0.20 3.68 � 0.18 18 24 1.20–5.08Careiro �3.3547, �59.8605 5 (1) 17 2.63 � 0.13 3.15 � 0.19 3.85 � 0.18 19 23 1.17–5.02Catuaba �10.0742, �67.6249 6 (1) 12 2.80 � 0.09 3.38 � 0.13 3.93 � 0.14 23 25 1.53–5.39Panguana �9.6137, �74.9355 8 (1) 14 2.58 � 0.08 3.15 � 0.13 3.87 � 0.09 21 21 1.18–5.05
Synthesis of Experimental Stimuli
The advertisement calls employed as stimuli were assem-bled using SoundEdit software 2.0.3 (Weary and Weisman1993). First, one ‘‘average’’ call was synthesized accordingto the average temporal and spectral parameters of the pop-ulation. To mimic the note envelope of natural calls, weapplied to each synthetic note the fast option of the fade-ineffect (the sound’s amplitude is increased quickly at first,then levels off toward the end of the sound) and the slowoption of the fade-out effect (the sound’s amplitude decreasesslowly at first, then it decreases rapidly toward the end ofthe sound). Because the number of notes per call variesamong the studied populations of A. femoralis, we prepared
synthetic calls consisting of two (Catuaba), three (Panguana),or four (all other populations) notes. Here, we report malereactions only towards calls with the same number of notesas the calls they produce.
To test the effect of call frequency on male phonotacticresponse, we first synthesized 12–15 stimulus calls per pop-ulation by simultaneously varying the frequency of each notebut keeping the temporal properties of the average call con-stant. The frequency range encompassed by stimulus callsextended beyond variation of natural calls at each population;a wider frequency range was necessary at the Hileia site toobtain a sufficient number of negative phonotactic reactions.
To replicate each stimulus call, we randomly modified its

1878 ADOLFO AMEZQUITA ET AL.
FIG
.3.
Sou
ndsp
ectr
ogra
ms
ofth
ree-
seco
nds
reco
rdin
gsof
the
mai
nau
dito
rysi
gnal
of20
anur
anan
don
eor
thop
tera
nsp
ecie
sre
gist
ered
atth
est
udy
site
s.O
scil
logr
ams
are
pres
ente
dbe
low
each
spec
trog
ram
toem
phas
ize
betw
een-
spec
ies
diff
eren
ces
inth
ete
mpo
ral
stru
ctur
eof
the
sign
al.
Six
seco
nds
ofth
eP
hyze
laph
ryne
sp.’
sca
ll(T
revi
sosi
te)
are
pres
ente
dto
outl
ine
sile
ntin
terv
als
betw
een
call
bout
s.T
hein
set
pres
ents
inm
ore
deta
ilon
eof
the
adve
rtis
emen
tca
lls
ofA
llob
ates
fem
oral
isat
Let
icia
.

1879MASKING INTERFERENCE IN FROG COMMUNICATION
TABLE 2. Logistic regression analysis of the effect of stimulus call frequency and population on the probability of eliciting a phonotacticresponse during playback experiments on Allobates femoralis males. See Methods for a detailed explanation of variables and criteria.
Variables B SE Wald df P
Model if term removed
LL �2 Log LR P of change
In the equationStimulus frequency � Side 3.568 0.497 51.523 1 �0.001 �127.702 105.668 �0.001Population 20.645 7 0.004 �88.636 27.536 �0.001Stimulus frequency � Population � Side 11.614 7 0.114 �81.291 12.845 0.076Side �11.577 1.621 50.985 1 �0.001 �127.416 105.096 �0.001Constant 4.158 0.603 47.572 1 �0.001
Not in the equation ScoreStimulus frequency � Population 3.733 7 0.810Sound pressure level 0.711 1 0.399Stimulus frequency 1.124 1 0.289Overall statistics 5.401 9 0.798
temporal features within the population range of variation,without altering the experimental factor (call frequency). Arandom number (between 0 and 1) was multiplied by thestandard deviation of each temporal feature and the resultingvalue was either added or subtracted (according to a secondrandom number) to the population’s average (Amezquita etal. 2005). Hence, we generated for each population between21–30 replicates of the stimulus calls that varied less thanone standard deviation in the noninvestigated call features.
Playback Experiments
We report the result of 197 playback experiments with 197different synthetic signals on 159 males (Table 1). Becausethe relevant unit of analysis is the signal used in a playback,we avoided pseudoreplication by using a different signal inevery trial (Kroodsma 1989, 1990; Hopp and Morton 1998;Amezquita et al. 2005; Rojas et al. 2006). Because nonin-dependence of some test subjects (caused by reusing somemales) may influence the generality of the conclusions weexpected to draw, a maximum of two experiments separatedby a period of no calling activity (one night or one noon forA. femoralis) were performed with a single male. Addition-ally, we never tested two calls of similar frequency on thesame individual. Stimulus calls were broadcast by a battery-powered loudspeaker (Sony SRS-A27, Sony SRS-A47, orSony SRS-57, Sony Corp., Tokyo) connected to a CD player(Panasonic Sl-SX320, Panasonic Corp., Secaucus, NJ, or Sil-va Schneider, Anif Anif, Austria CDP 286 AS). To avoidimmediate effects of E. trivittatus calls on A. femoralis re-sponse, we stimulated males at sound pressure levels of 72–80 dB (peak intensity, fast time, measured with a Sound-Level Meter RO-1350 Roline, Rotronic AG, Bassersdorf,Switzerland or RION Co., Toyko, NC-73/M91), well abovethe sound pressure level (SPL) of both conspecific (Hodl1987) and heterospecific calls (54–62 dB, this study and re-cordings of the acoustic environment) measured at males’calling positions. Because unavoidable differences in micro-habitat structure affect the SPL at which tested males perceivethe stimulus calls, we positioned the loudspeaker between100–230 cm away from the male, according to the probabilityof excessive sound attenuation by vegetation. Moreover, wemeasured the stimulus’ SPL at the focal-male position im-mediately after the experiment, and tested its potential effect
as a covariate in the corresponding statistical model (Table2).
The playback experiments were conducted following theprotocols published elsewhere (Narins et al. 2003; Hodl etal. 2004; Amezquita et al. 2005). In brief, we broadcast seriesof calls according to the average pattern of spontaneous call-ing among several A. femoralis populations (Hodl 1987; Na-rins et al. 2003). During a single experiment (lasting up to220 sec) up to 10 consecutive bouts (18–22 sec per bout)were broadcast, each bout consisting of 10 synthetic calls(9–11 sec duration) followed by a silent interval. The outputvariable was binary. If the male under test crossed a perimeterof 30 cm around the loudspeaker, we stopped the experimentand declared a positive phonotactic response. Trials in whichmales failed to approach the loudspeaker within the experi-ment time were counted as negative only if the individualunder test exhibited a positive phonotactic approach in asubsequent control trial performed with the average call.Most tests occurred between 0535–1100 hours and 1430–1825 hours, under rainless conditions, when at least severalindividuals of A. femoralis were calling.
Statistical Analyses
Our statistical approach involves five steps. First, we cal-culated average values to characterize four frequency traitsof A. femoralis calls at each study site: the call peak frequency(CPF), the call frequency range (CFR), and the call low(CLF), and high (CHF) frequency (Fig. 1B). Second, we de-rived frequency-response curves from male reactions to play-back experiments, to interpolate four equivalent response fre-quency traits: the response peak frequency (RPF), the re-sponse frequency range (RFR), and the response low (RLF)and high (RHF) frequency (Fig. 1B). Third, we examined thepatterns of geographic covariation between body size, call-and response-frequency traits. Fourth, we tested for corre-lation between subsets of covarying traits and geographicdistance. Fifth, we tested the main prediction by running aset of regression models on the effect of the occurrence ofE. trivittatus and the number of coactive species (covariate)on the frequency traits of the communication system of A.femoralis.

1880 ADOLFO AMEZQUITA ET AL.
Estimation of frequency-response curves
Because males were tested with synthetic calls encom-passing a wide-frequency range approximately centered onthe population’s average, the probability of male positiveresponse can be expected to be low towards low-frequencycalls, high towards intermediate-frequency (i.e., close to thepopulation average) calls, and again low towards high-fre-quency calls. Such response pattern was tested as the inter-action effect of stimulus frequency and side (a factor withtwo levels: stimulus frequency below or above the populationaverage) on male response (see also Amezquita et al. 2005;Rojas et al. 2006). Because the output variable was binary,the males either approached (1) or not (0) within the exper-imental time to the loudspeaker, a logistic regression modelwas appropriate (Menard 2001). If the null hypothesis is re-jected, then two (one positive and one negative) logisticcurves can be fitted to the original data and fused in a bell-like curve representing the predicted probability of male re-sponse (Fig. 1B). By adding the factor population and thecorresponding interactions with stimulus frequency and sideas potential predictors, the model tested whether populationsdiffer in the form (width and symmetry) and position (alongthe scale of stimulus frequency) of their frequency-responsecurves. Finally, by adding the SPL of the stimulus at males’initial position as a covariate, the model tested the potentialeffect of stimulus intensity on male response.
We tested the resulting logistic model with a backwardstepwise procedure. Although stepwise procedures may beinappropriate for theory testing, they are useful in purelypredictive research and exploratory research (Hosmer andLemeshow 1989; Menard 2001). This part of our analysis ispurely predictive because our main concern is to identify asubset of variables that provides accurate predictions of maleresponse, that is, to build a frequency-response curve. Wechose the backward elimination (instead of forward addition)procedure because it reduces the risk of suppressor effects,that is, that the effect of one variable is not significant becausethe effect of another variable is not controlled or held constant(Agresti and Finlay 1997). Four parameters were interpolatedfrom the frequency-response curves of each population (Fig.1B): the response peak frequency (RPF) represents the mid-point between the two frequency values at which the prob-ability of male positive reaction was 0.9; the response fre-quency range (RFR) represents the range of frequency valuesat which the model predicts that males most probably reactto the stimulus; that is, the probability of reaction is above0.5; the response low (RLF) and high (RHF) frequency valuesrepresent the lower and upper limits of the RFR. Because asingle frequency-response curve is derived for each popu-lation, and because each male’s reaction is coded as a singlebinary variable, we are unable to provide estimates of inter-individual variation in frequency recognition curves.
Geographic pattern of covariation
Populations are inadequate units of analysis for statisticaltests, because their independence is compromised by geo-graphic distance or phylogenetic relationships (Sokal 1979;Manly 1997; Fortin and Gurevitch 1993; Legendre et al.2002). Therefore, we used here comparisons between pairs
of populations as the statistical units, and estimated statisticalsignificance of tests by permutation techniques (Mantel 1967;Sokal 1979). The Mantel approach was first used to examinethe pattern of geographic correlation between the studiedtraits. We calculated between-populations differences (Eu-clidean distance) for each original variable (body size, call-and response-frequency traits) and then calculated Mantelcorrelation coefficients between each pair of the resulting(population � population) distance matrices. In turn, theMantel correlation coefficients were used to perform a prin-cipal coordinates ordination (Gower 1966; Legendre and La-pointe 2004) that represents the degree of congruence be-tween studied traits across the study sites. The Mantel ap-proach was also used to test for correlation between eachsubset of intercorrelated variables and geographic distance,the latter calculated on a GPS.
Model testing by multiple regression
To test the effect of the occurrence of E. trivittatus and thenumber of acoustically coactive species (covariate) on eachfrequency trait we ran multiple permutational regressions onthe software Permute 3.4 (Legendre et al. 1994). Severalauthors have suggested using multiple regression as an ex-tension of the Mantel test for two or more input variables(Smouse et al. 1986; Legendre et al. 1994; Manly 1997); thepermutation of raw data has shown good power and correctrates of type I error (Anderson and Legendre 1999). We rana separate regression for each frequency trait because we hadspecific predictions for them (Fig. 1 above); the significancelevel was correspondingly adjusted (0.05/number of tests)according to the Bonferroni procedure (Legendre and Le-gendre 1998).
As input and output variables, we used the skew-symmetricmatrices containing the absolute difference (value in popu-lation 2 � value in population 1) between all (randomlycontrasted) pairs of populations (Sokal 1979; Fortin and Gur-evitch 1993; Legendre 2000). Only the upper triangular partof the matrices was used because it contains all the infor-mation needed to compare populations (Casgrain et al. 1996).The occurrence of E. trivittatus was coded as �1 (if E. tri-vittatus did not occur in the first population but it did in thesecond), 0 (if E. trivittatus occurred in neither or occurred inboth populations), or 1 (if E. trivittatus occurred in the firstpopulation but did not in the second). By allowing negativephenetic differences in the matrices (Sokal 1979; Manly1997), we were able to specify the direction of significantrelationships, that is, not only whether large differences insome trait predict large differences in other, but whether anincrease in the former predicts either an increase or a decreasein the latter.
RESULTS
At the study sites, between three and eight species wererecorded from A. femoralis calling positions at times of daywhen individuals of this species were calling (Fig. 3). Threeadditional species (Dendrobates ventrimaculatus at Leticiaand Careiro, D. castaneoticus at Treviso, and Dendrophryn-iscus minutus at Careiro) were active and apparently breedingbut their calls were not registered in samples of acoustic

1881MASKING INTERFERENCE IN FROG COMMUNICATION
FIG. 4. Frequency traits of the main auditory signal of 20 anuranand one orthopteran species registered at the study sites. Each circlerepresents the average peak frequency of one male. The low andhigh limits of the frequency range correspond to among-individualsaverages of measurements performed at 20 dB below the peak in-tensity of the call. The study species (Af, Allobates femoralis) andthe species hypothesized to represent a source of masking inter-ference (Et, Epipedobates trivittatus) are outlined. Within each studysite, species are sorted according to the frequency range of theiradvertisement calls. From left to right: Hileia (HIL): A. femoralis,Colostethus sp2, and Adenomera andreae; Arataı (ARA): A. femor-alis, E. hahneli, and A. andreae; Ducke (DUC): A. femoralis, C.stepheni, A. andreae, and Colostethus sp3; Treviso (TRE): Phyze-laphryne sp., Bufo aff. typhonius, A. femoralis, Colostethus sp6, Co-lostethus sp7, A. andreae, and Colostethus sp5; Leticia (LET): E.trivittatus, A. femoralis, Unidentified orthopteran, C. trilineatus; Ca-tuaba (CAT): E. trivittatus, A. femoralis, E. hahneli, Colostethus sp4,and C. aff. trilineatus 2; Careiro (CAR): Vanzolinius discodactylus,E. trivittatus, A. femoralis, Colostethus sp2, A. andreae, and Colos-tethus sp1; Panguana (PAN): E. trivittatus, A. femoralis, E. petersi,A. andreae, E. hahneli, E. aff. pictus, C. aff. trilineatus 1, and Den-drobates ventrimaculatus.
FIG. 5. Geographic variation in the frequency-response curves ofAllobates femoralis males. The probability of male response is in-terpolated from a logistic regression analysis of male binary re-sponses during field playback experiments (see Table 2). Study sitesare organized according to the absence (left) or presence (right) ofEpipedobates trivittatus, and to the increasing (up to down) numberof acoustically coactive species (Table 1).
environment, probably because they are produced too sparse-ly and at a very low intensity. At the four sites where E.trivittatus occurs, its call frequency range overlapped with A.femoralis (Fig. 4). At the Panguana site, the call peak fre-quency and the call frequency range of E. petersi overlappedwith A. femoralis (Fig. 4).
The logistic regression analysis of male A. femoralis re-sponses to playback experiments included three variables assignificant predictors (83.2 % of correct classifications; Na-gelkerke R2 � 0.615): the interaction stimulus frequency �

1882 ADOLFO AMEZQUITA ET AL.
TABLE 3. Matrix of Mantel correlation coefficients (above diagonal) and associated significance levels (below diagonal) between bodysize (SVL) and the spectral features of the advertisement call and the frequency-response curve of Allobates femoralis.
SVL CPF CLF CHF RPF RLF RHF CFR RFR
Snout-vent length (SVL) 0.49 0.44 0.49 0.50 0.21 �0.13 �0.31 �0.22Call peak frequency (CPF) 0.0276 0.91 0.75 0.59 �0.23 0.32 �0.25 �0.11Call low frequency (CLF) 0.0458 �0.0001 0.71 0.45 �0.15 0.21 �0.16 �0.05Call high frequency (CHF) 0.0660 0.0009 0.0018 0.27 �0.35 0.28 �0.21 �0.11Response peak frequency (RPF) 0.0382 0.0040 0.0214 0.1494 0.09 0.15 �0.26 �0.19Response low frequency (RLF) 0.2674 0.1768 0.2740 0.1260 0.3620 0.02 0.15 0.53Response high frequency (RHF) 0.2493 0.0479 0.1438 0.0770 0.2258 0.4683 �0.08 0.67Call frequency range (CFR) 0.1224 0.1275 0.2356 0.2029 0.1274 0.2705 0.3414 0.10Response frequency range (RFR) 0.1622 0.2922 0.4064 0.2983 0.1784 0.0073 0.0070 0.3049
side (a factor with two levels: stimulus frequency below orabove the population average), side, and population (Table2). The three-way interaction stimulus frequency � popu-lation � side was also accepted as predictor but its effectwas very low. Between-experiments variation in perceivedstimulus intensity did not affect male response (Table 2). Theprobability of male reaction predicted by the model was usedto build frequency-response curves for each population,which differed in form (width and symmetry) and position(along the stimulus frequency axis) (Fig. 5).
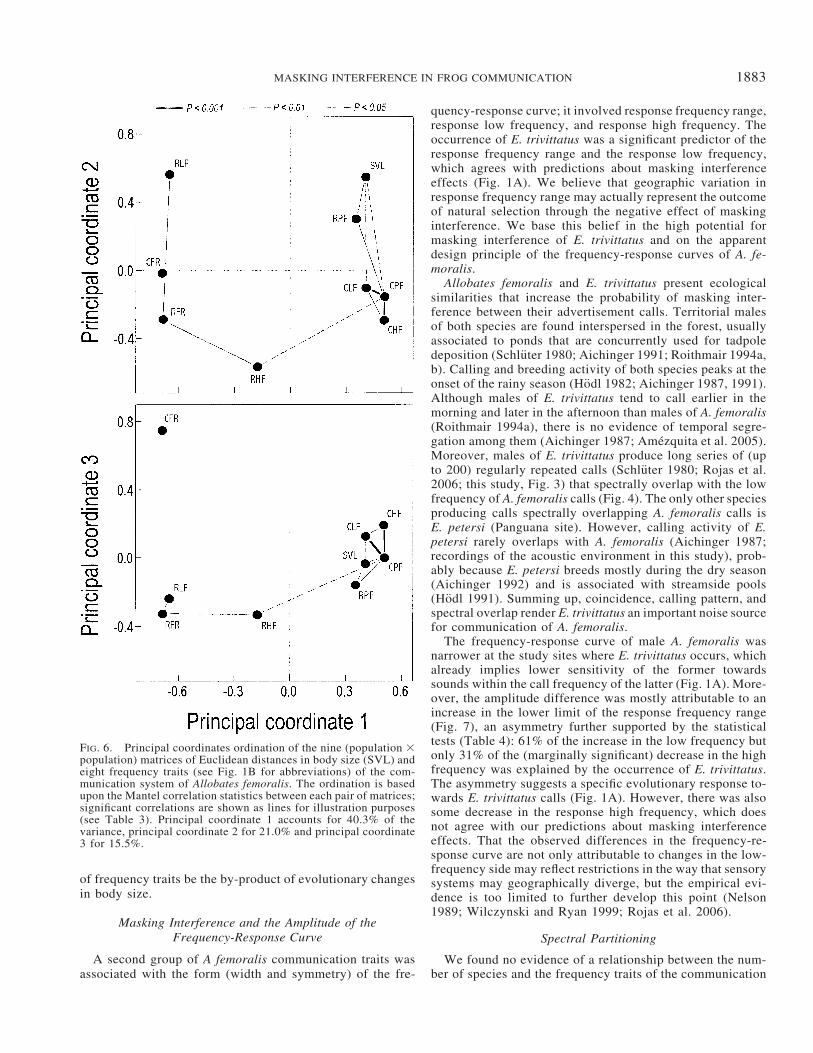
The frequency traits of the communication system werehighly correlated among them and with male body size (Table3). The principal coordinates analysis revealed three groupsof variables that tend to covary across the study sites (Fig.6). First, call peak, low and high frequency were highly cor-related among them as well as with response peak frequencyand body size. Second, response frequency range was sig-nificantly correlated with response low and high frequency.Third, call frequency range, which was not correlated withany other of the studied traits. Geographic distance betweenpopulations was not correlated with multivariate Euclideandistances calculated from the first (Mantel test; Mantel r ��0.17, one-tailed P � 0.358), second (Mantel r � 0.22, one-tailed P � 0.163), or third (Mantel r � �0.09, one-tailed P� 0.347) group of variables.
To remove the confounding effect of body size on callfrequency traits (Table 3), we used raw individual data toregress call peak frequency (linear regression; N � 126 inall cases; R2 � 0.20, F � 31.5, P � 0.001), call low frequency(R2 � 0.38, F � 36.9, P � 0.001), and call high frequency(R2 � 0.18, F � 13.5, P � 0.001) on male size. The residualsof these regressions were averaged per population and usedas size-independent estimates of call peak, low, and highfrequency.
Among populations, the occurrence of E. trivittatus was ahighly significant predictor of response low frequency andresponse frequency range (Table 4, Fig. 7). Where E. trivit-tatus was present, the frequency-response curve of male A.femoralis was narrower due to an increase in the responselow frequency and a marginally significant (Table 4) decreasein the response high frequency (Fig. 7). Neither the occur-rence of E. trivittatus nor the number of species had a sig-nificant effect on the call frequency traits (Table 4).
DISCUSSION
Our results demonstrate that the male-male communicationsystem of A. femoralis did not evolve as a unit. There were
three recognizable groups of intercorrelated frequency traits,the first associated to the position (along the frequency axis)of the call and the response, the second associated to the form(width and symmetry) of the frequency-response curve, andthe third including only the call frequency range. The oc-currence of E. trivittatus was a significant predictor of var-iation in traits associated with the form of the frequency-response curve, but not with variation in any other trait. Al-together, the results agree with predictions about the selectiveeffect of masking interference by a single species; they alsoconstitute the first correlational evidence of a commonly in-voked adaptation for communication in acoustic assem-blages.
Variation in Frequency Traits Correlated with Body Size
The first group of covarying traits described the positionof the communication system along the frequency axis: callpeak frequency, response peak frequency, as well as call lowand high frequency. None of these traits was related to theoccurrence of E. trivittatus or to the number of acousticallycoactive species. Instead, geographic differentiation in call/response peak frequency (and associated traits) was corre-lated with concomitant differences in body size. This resultis not surprising, because body size is well known to influencethe spectral features of the signal (call frequency) and theauditory system (basilar papilla tuning) in anurans (Martin1972; Ryan 1988; Wagner 1989; Keddy-Hector et al. 1992).In turn geographic variation in body size is widespread, andoften correlated with environmental differences in humidity(Calhoon and Jameson 1970; Nevo 1973; Nevo and Capran-ica 1985; Lee 1993) and temperature (Narins and Smith 1986;Asquith et al. 1988).
There is also some evidence that call frequency and basilarpapilla tuning can exhibit size-independent variation amongpopulations (Ryan and Wilczynski 1991; Keddy-Hector et al.1992; McClelland et al. 1996), which can be correlated withenvironmental differences in habitat structure (Wilczynskiand Ryan 1999). In A. femoralis, we retested the effect ofthe occurrence of E. trivittatus and the number of species onthe residual (size independent) estimates of call frequencytraits, instead of the original data, but found no significantrelationships (Table 4). In sum, we have evidence of geo-graphic variation in call/response peak frequency (and as-sociated traits) apparently not correlated with masking in-terference; also, we cannot discard that variation in this group
1883MASKING INTERFERENCE IN FROG COMMUNICATION
FIG. 6. Principal coordinates ordination of the nine (population �population) matrices of Euclidean distances in body size (SVL) andeight frequency traits (see Fig. 1B for abbreviations) of the com-munication system of Allobates femoralis. The ordination is basedupon the Mantel correlation statistics between each pair of matrices;significant correlations are shown as lines for illustration purposes(see Table 3). Principal coordinate 1 accounts for 40.3% of thevariance, principal coordinate 2 for 21.0% and principal coordinate3 for 15.5%.
of frequency traits be the by-product of evolutionary changesin body size.
Masking Interference and the Amplitude of theFrequency-Response Curve
A second group of A femoralis communication traits wasassociated with the form (width and symmetry) of the fre-
quency-response curve; it involved response frequency range,response low frequency, and response high frequency. Theoccurrence of E. trivittatus was a significant predictor of theresponse frequency range and the response low frequency,which agrees with predictions about masking interferenceeffects (Fig. 1A). We believe that geographic variation inresponse frequency range may actually represent the outcomeof natural selection through the negative effect of maskinginterference. We base this belief in the high potential formasking interference of E. trivittatus and on the apparentdesign principle of the frequency-response curves of A. fe-moralis.
Allobates femoralis and E. trivittatus present ecologicalsimilarities that increase the probability of masking inter-ference between their advertisement calls. Territorial malesof both species are found interspersed in the forest, usuallyassociated to ponds that are concurrently used for tadpoledeposition (Schluter 1980; Aichinger 1991; Roithmair 1994a,b). Calling and breeding activity of both species peaks at theonset of the rainy season (Hodl 1982; Aichinger 1987, 1991).Although males of E. trivittatus tend to call earlier in themorning and later in the afternoon than males of A. femoralis(Roithmair 1994a), there is no evidence of temporal segre-gation among them (Aichinger 1987; Amezquita et al. 2005).Moreover, males of E. trivittatus produce long series of (upto 200) regularly repeated calls (Schluter 1980; Rojas et al.2006; this study, Fig. 3) that spectrally overlap with the lowfrequency of A. femoralis calls (Fig. 4). The only other speciesproducing calls spectrally overlapping A. femoralis calls isE. petersi (Panguana site). However, calling activity of E.petersi rarely overlaps with A. femoralis (Aichinger 1987;recordings of the acoustic environment in this study), prob-ably because E. petersi breeds mostly during the dry season(Aichinger 1992) and is associated with streamside pools(Hodl 1991). Summing up, coincidence, calling pattern, andspectral overlap render E. trivittatus an important noise sourcefor communication of A. femoralis.
The frequency-response curve of male A. femoralis wasnarrower at the study sites where E. trivittatus occurs, whichalready implies lower sensitivity of the former towardssounds within the call frequency of the latter (Fig. 1A). More-over, the amplitude difference was mostly attributable to anincrease in the lower limit of the response frequency range(Fig. 7), an asymmetry further supported by the statisticaltests (Table 4): 61% of the increase in the low frequency butonly 31% of the (marginally significant) decrease in the highfrequency was explained by the occurrence of E. trivittatus.The asymmetry suggests a specific evolutionary response to-wards E. trivittatus calls (Fig. 1A). However, there was alsosome decrease in the response high frequency, which doesnot agree with our predictions about masking interferenceeffects. That the observed differences in the frequency-re-sponse curve are not only attributable to changes in the low-frequency side may reflect restrictions in the way that sensorysystems may geographically diverge, but the empirical evi-dence is too limited to further develop this point (Nelson1989; Wilczynski and Ryan 1999; Rojas et al. 2006).
Spectral Partitioning
We found no evidence of a relationship between the num-ber of species and the frequency traits of the communication

1884 ADOLFO AMEZQUITA ET AL.
TABLE 4. Summary of the permutational regression analyses to test the effect of the occurrence of Epipedobates trivittatus and thenumber of acoustically coactive species (covariate) on the frequency traits of the sender (call and residuals of call features on body size)and the receiver (frequency-response curve) portion of the communication system of Allobates femoralis. Significance levels are assessedafter Bonferroni’s correction (0.05/number of tests � 0.0045): **statistically significant, MSmarginally significant.
Dependent Model R2 One-tailed P Source Std. coeff. Two-tailed P
Advertisement callPeak frequency 0.00 0.9927 TRIVI �0.0357 0.8888
SPP 0.0217 0.9315Low frequency 0.02 0.8733 TRIVI 0.0597 0.8188
SPP 0.0891 0.7575High frequency 0.12 0.2933 TRIVI �0.1445 0.5620
SPP 0.3851 0.1382Frequency range 0.31 0.0395 TRIVI �0.4211 0.1026
SPP 0.6132 0.0117Resid. of peak freq. 0.24 0.0553 TRIVI �0.5361 0.0203
SPP 0.1235 0.5619Resid. of low freq. 0.07 0.5396 TRIVI �0.2875 0.2779
SPP 0.1014 0.7121Resid. of high freq. 0.19 0.1462 TRIVI �0.4055 0.0991
SPP 0.4371 0.1051
Frequency-response curvePeak frequency 0.06 0.5011 TRIVI 0.2487 0.2932
SPP �0.2201 0.3494Low frequency 0.61** 0.0030 TRIVI 0.8286** 0.0015
SPP �0.1107 0.5973High frequency 0.31MS 0.0193 TRIVI �0.5023 0.0259
SPP �0.0984 0.6541Frequency range 0.58** 0.0030 TRIVI �0.7635** 0.0030
SPP �0.0015 0.9943
FIG. 7. Relationship between traits of the frequency-response curves (low frequency, frequency range, and high frequency) of Allobatesfemoralis males and the occurrence of Epipedobates trivittatus. Each circle represents the absolute difference between two populations,which represents the unit of analysis in permutational multiple regressions (see Methods); lines are shown for significant relationships(see Table 4). The difference in the occurrence of E. trivittatus was coded in a skew-symmetric matrix as �1 (if it did not occur in thefirst population but occurred in the second one), 0 (if it occurred in neither or occurred in both populations), or 1 (if it occurred in thefirst population but not in the second).
system of A. femoralis, indicating that the potential for mask-ing interference does not increase with the number of acous-tically coactive species. Does this result contradict the ideathat spectral partitioning among species that breed togetheris the evolutionary outcome of masking interference? On the
one hand, if the communication system of each species isinfluenced only by those species calling within adjacent fre-quency ranges (as is strongly suggested by this study), thenthe sum of the evolutionary interactions between species pairsmay lead the system towards spectral partitioning (Otte 1974;

1885MASKING INTERFERENCE IN FROG COMMUNICATION
Nelson 1989). In this sense, the results of this study actuallyprovide some cue about the mechanisms by which maskinginterference may lead to spectral partitioning. On the otherhand, the concept of spectral partitioning has been histori-cally applied to between-species differences in call frequency(Hodl 1977; Drewry and Rand 1983; Duellman and Pyles1983; Narins 1995; Luddecke et al. 2000; Chek et al. 2003)rather than in the sensory system (Wilczynski et al. 1993).Because call frequency and response peak frequency are cor-related with body size in A. femoralis (Table 3, Fig. 6), sup-portive evidence for the role of masking interference in spec-tral partitioning should have involved size-independent var-iation in the position (along the frequency axis) of the com-munication system. In this sense, our study does not provideevidence of a mechanistic relationship between masking in-terference and spectral partitioning.
Why were the changes in the frequency-response curve notparalleled by concomitant changes in the call? One reasoncould be that the auditory system’s sensitivity extends beyondthe signal’s frequency range, and thus has a large potentialto diverge under noisy environments. In A. femoralis, theresponse frequency range is up to 2 kHz wider than the callfrequency range, and there is no correlation among them. The‘‘extended’’ sensory range may reflect multiple roles of sen-sory system (Megela-Simmons et al. 1985; Narins and Zelick1988; Wilczynski et al. 1995), that is, that it is used not onlyfor communication but also for detection of other relevantenvironmental sounds (e.g., sounds originating from preys orpredators). However, an extended sensory range also increas-es the detection of nonrelevant information thus reducing thecontrast between the signal and the background noise. Thus,the response frequency range should reflect a compromiseaccording to the frequency distribution of local noise sources.Our behavioral study supports this idea.
Conclusion and Further Research
In this study we demonstrate for the first time geographicvariation in the response frequency range that agrees withpredictions about the selective effect of masking interference.Males of A. femoralis react less towards low-frequency callsat sites where E. trivittatus calls within this frequency range.We do not know whether the change in the acoustic criteriais due to shifts in the peripheral sensitivity (Gerhardt 1987;Ryan and Wilczynski 1988; Ryan et al. 1992) or to modifi-cations in the brain template (Fuzessery 1988; Hall 1994;Pollack 1998). However, our experiments were conductedunder realistic conditions of male motivational state andbackground noise, which suggests that, regardless of thephysiological mechanism behind it, the frequency-responsepattern should have ecological consequences at the individuallevel. The negative effect of noise masking in anuran socialinteractions is well documented (Ehret and Gerhardt 1980;Schwartz and Wells 1983; Gerhardt and Klump 1988; Woll-erman 1999). However, to claim an adaptation in the case ofA. femoralis, the inter-individual variance in acoustic criteriais still to be assessed, as well as the fitness consequences ofbearing variants of the communication frequency traits.
ACKNOWLEDGMENTS
This study was mainly supported by a grant to W. Hodlfrom the Austrian Science Foundation (FWF-P15345). Ad-ditional support for the study at Leticia was provided asgrants to L. Castellanos and to A. Amezquita from the Re-search Committee of the Faculty of Sciences at the Univ-ersidad de Los Andes, Bogota. For providing field assistancewe are grateful to K. Siu-Ting and N. Ellinger, Peru; G. deLuna, O. Ramos, A. Velez, B. Rojas, S. Flechas, C. Ulloa,C. Guarnizo, Colombia; J. R. D. Souza, V. M. L. Guida, R.S. Costa, L. G. Moura, M. A. Freitas, M. Frimmel, Brazil;H. and D. Dell’mour, C. Proy, and P. Narins, French Guiana.For providing invaluable logistic support we especially thankC. Keller, P. Gaucher, and H. Gasser. For sharing their ex-perience as local inhabitants of the study sites, we thank thecommunity of Cercaviva, Colombia; Moro’s family, Peru;and R. N., and D. Antonia, Brazil. Research permits wereprovided by CorpoAmazonia, Colombia, Permit no. 1128;IBAMA/RAN, Brazil, Permit no. 004/03; and INRENA, Peru,Permit no. 078-2003).
LITERATURE CITED
Agresti, A., and B. Finlay. 1997. Statistical methods for the socialsciences. Prentice Hall, Upper Saddle River, NJ.
Aichinger, M. 1987. Annual activity patterns of anurans in a sea-sonal Neotropical environment. Oecologia 71:583–592.
———. 1991. Tadpole transport in relation to rainfall, fecundity,and body size in five species of poison-dart frogs from Ama-zonian Peru. Amph.-Rept. 12:49–55.
———. 1992. Fecundity and breeding sites of an anuran communityin a seasonal tropical environment. Stud. Neotrop. Fauna En-viron. 27:9–18.
Amezquita, A., L. Castellanos, and W. Hodl. 2005. Auditory match-ing of male Epipedobates femoralis (Anura: Dendrobatidae) un-der field conditions: the role of spectral and termporal call fea-tures. Anim. Behav. 70:1377–1386.
Anderson, M. J. and P. Legendre. 1999. An empirical comparisonof permutation methods for tests of partial regression coefficientsin a linear model. J. Stat. Comput. Simul. 62:271–303.
Anderson, S., and P. K. McGregor. 1999. Animal communication:what is the signal to noise ratio? Trends Ecol. Evol. 14:174–175.
Arak, A., and M. Enquist. 1993. Hidden preferences and the evo-lution of signals. Philos. Trans. R. Soc. Lond. B 340:207–213.
Asquith, A., R. Altig, and P. Zimba. 1988. Geographic variation inthe mating call of the green treefrog Hyla cinerea. Am. Midl.Nat. 119:101–110.
Bennet-Clark, H. C. 1998. Size and scale effects as constraints ininsect sound communication. Philos. Trans. R. Soc. Lond. B353:407–419.
Blair, W. F. 1964. Evolution at populational and interpopulationallevels. Isolating mechanisms and interspecies interactions in an-uran amphibians. Q. Rev. Biol. 39:333–344.
Boughman, J. W. 2002. How sensory drive can promote speciation?Trends Ecol. Evol. 17:571–587.
Bradbury, J. W., and S. L. Vehrencamp. 1998. Principles of animalcommunication. Sinauer, Sunderland, MA.
Calhoon, R. E., and D. L. Jameson. 1970. Canonical correlationbetween variation in weather and variation in size in the Pacifictree frog, Hyla regilla, in southern California. Copeia 1970:124–134.
Casgrain, P., P. Legendre, J.-L. Sixou, and C. Mouton. 1996. Agraph-theory method to establish serological relationships withina bacterial taxon, with example from Porphyromonas gingivalis.J. Microbiol. Meth. 26:225–236.
Charif, R. A., S. Mitchell, and C. W. Clark. 1995. Canary 1.12user’s manual. Cornell Laboratory of Ornithology, Ithaca, NY.
Chek, A. A., J. P. Bogart, and S. C. Lougheed. 2003. Mating signal

1886 ADOLFO AMEZQUITA ET AL.
partitioning in multi-species assemblages: a null model test usingfrogs. Ecol. Lett. 6:235–247.
Cocroft, R. B., and M. J. Ryan. 1995. Patterns of advertisementcall evolution in toads and chorus frogs. Anim. Behav. 49:283–303.
Drewry, G. E., and A. S. Rand. 1983. Characteristics of an acousticcommunity: Puerto Rican frogs of the genus Eleutherodactylus.Copeia 1983:941–953.
Duellman, W. E., and R. E. Pyles. 1983. Acoustic resource parti-tioning in anuran communities. Copeia 1983:639–649.
Ehret, G., and H. C. Gerhardt. 1980. Auditory masking and effectsof noise on responses of of the green treefrog (Hyla cinerea) tosynthetic mating calls. J. Comp. Physiol. A 141:13–18.
Endler, J. A. 1983. Testing causal hypotheses in the study of geo-graphic variation. Pp. 425–443 in J. Felsenstein, ed. Numericaltaxonomy. Springer, Berlin.
Forrest, T. G. 1994. From sender to receiver: propagation and en-vironmental effects on acoustic signals. Am. Zool. 34:644–654.
Fortin, M. J., and J. Gurevitch. 1993. Mantel tests: Spatial structurein field experiments. Pp. 342–359 in S. M. Scheiner and J. Gur-evitch, eds. Design and analysis of ecological experiments.Chapman and Hall, New York.
Fuzessery, Z. M. 1988. Frequency tuning in the anuran centralauditory system. Pp. 253–273 in B. Fritzsch, M. J. Ryan, W.Wilczynski, T. E. Hetherington, and W. Walkowiak, eds. Theevolution of the amphibian auditory system. John Wiley, NewYork.
Gerhardt, H. C. 1987. Evolutionary and neurobiological implica-tions of selective phonotaxis in the green treefrog, Hyla cinerea.Anim. Behav. 35:1479–1489.
Gerhardt, H. C., and F. Huber. 2002. Acoustic communication ininsects and anurans. Common problems and diverse solutions.Univ. of Chicago Press, Chicago, IL.
Gerhardt, H. C., and G. M. Klump. 1988. Masking of acousticsignals by the chorus background noise in the green tree frog:A limitation on mate choice. Anim. Behav. 36:1247–1249.
Gottsberger, B., and E. Gruber. 2004. Temporal partitioning of re-productive activity in a Neotropical anuran community. J. Trop.Ecol. 20:271–280.
Gower, J. C. 1966. Some distance properties of latent root andvector methods used in multivariate analysis. Biometrika 53:325–338.
Greenfield, M. D. 1994. Synchronous and alternating choruses ininsects and anurans: common mechanisms and diverse functions.Am. Zool. 34:605–615.
Grether, G. F. 2000. Carotenoid limitation and mate preferenceevolution: a test of the indicator hypothesis in guppies (Poeciliareticulata). Evolution 54:1712–1724.
Hall, J. C. 1994. Central processing of communication sounds inthe anuran auditory system. Am. Zool. 34:670–684.
Hodl, W. 1977. Call differences and calling site segregation inanuran species from central Amazonian floating meadows. Oec-ologia 28:351–363.
———. 1982. Phyllobates femoralis (Dendrobatidae): Rufverhaltenund akustische Orientierung der Mannchen (Freilandaufnah-men). Wiss. Film Ctf 1788 BHWK Wien. Encyclopaedia Ci-nematographica, E2822.
———. 1987. Dendrobates femoralis (Dentrobatidae): a handly fel-low for frog bioacoustics. Proceedings of the 4th Ordinary Gen-eral Meeting of the Societas Europaea Herpetologica, Nijmegen201–204.
———. 1991. Calling behaviour and spectral stratification in dart-poison frogs (Dendrobatidae). Wiss. Film Ctf 2444 OWF Wien.
Hodl, W., and A. Amezquita. 2001. Visual signaling in anuranamphibians. Pp. 493–530 in M. J. Ryan, ed. Anuran commu-nication. Smithsonian Institution Press, Washington, DC.
Hodl, W., A. Amezquita, and P. M. Narins. 2004. The role of callfrequency and the auditory papillae in phonotactic behavior inmale Dart-poison frogs Epipedobates femoralis (Dendrobatidae).J Comp. Physiol. A 190:823–829.
Hopp, S. L., and E. S. Morton. 1998. Sound playback studies. Pp.323–352 in S.L. Hopp, M. J. Owren, and C. S. Evans, eds.Animal acoustic communication. Springer, Berlin.
Hosmer, D. W., and S. Lemeshow. 1989. Applied logistic regres-sion. John Wiley, New York.
Houde, A. E., and J. A. Endler. 1990. Correlated evolution of femalemating preferences and male color patterns in the guppy Poeciliareticulata. Science 248:1405–1408.
Hunter, M. L., and J. R. Krebs. 1979. Geographical variation in thesong of the great tit (Parus major) in relation to ecological fac-tors. J. Anim. Ecol. 48:759–785.
Irwin, D. E. 2000. Song variation in an avian ring species. Evolution54:998–1010.
Jones, G. 1997. Acoustic signals and speciation: The roles of naturaland sexual selection in the evolution of cryptic species. Adv.Stud. Behav. 26:317–354.
Keddy-Hector, A. C., W. Wilczynski, and M. J. Ryan. 1992. Callpatterns and basilar papilla tuning in cricket frogs. II. Intrapop-ulation variation and allometry. Brain Behav. Evol. 39:238–246.
Kroodsma, D. E. 1989. Suggested experimental designs for songplaybacks. Anim. Behav. 37:600–609.
———. 1990. Using appropriate experimental designs for intendedhypotheses in ‘‘song’’ playbacks, with examples for testing ef-fects of song repertoire sizes. Anim. Behav. 40:1138–1150.
Lee, J. C. 1993. Geographic variation in size and shape of Neo-tropical frogs: a precipitation gradient analysis. Occ. Pap. Mus.Nat. Hist. Univ. Kansas 163:1–20.
Legendre, P. 2000. Comparison of permutation methods for thepartial correlation and partial Mantel tests. J. Statist. Comput.Simul. 67:37–73.
Legendre, P., and F. J. Lapointe. 2004. Assessing the congruenceamong distance matrices: single malt Scotch whiskies revisited.Aust. NZ J. Stat. 46:615–630.
Legendre, P., and L. Legendre. 1998. Numerical ecology. Elsevier,Amsterdam.
Legendre, P., F.-J. Lapointe, and P. Casgrain. 1994. Modeling brainevolution from behavior: a permutational regression approach.Evolution 48:1487–1499.
Legendre, P., M. R. T. Dale, M. J. Fortin, J. Gurevitch, M. Hohn,and D. Myers. 2002. The consequences of spatial structure forthe design and analysis of ecological field surveys. Ecography25:601–615.
Loftus-Hills, J. J., and M. J. Littlejohn. 1992. Reinforcement andreproductive character displacement in Gastrophryne carolinen-sis and G. olivacea (Anura: Microhylidae): a reexamination. Evo-lution 46:896–906.
Luddecke, H., A. Amezquita, X. Bernal, and F. Guzman. 2000.Partitioning of vocal activity in a Neotropical highland-frogcommunity. Stud. Neotrop. Fauna Environ. 35:185–194.
Manly, B. 1997. Randomization, bootstrap, and Monte Carlo meth-ods in biology. Chapman and Hall, London.
Mantel, N. 1967. The detection of disease clustering and a gener-alized regression approach. Cancer Res. 27:209–220.
Martin, W. F. 1972. Evolution of vocalization in the genus Bufo.Pp. 279–309 in W. F. Blair, ed. Evolution in the genus Bufo.Ch. 15. University of Texas Press, Austin, TX.
McClelland, B. E., W. Wilczynski, and M. J. Ryan. 1996. Corre-lations between call characteristics and morphology in malecricket frogs (Acris crepitans). J. Exp. Biol. 199:1907–1919.
Megela-Simmons, A., C. F. Moss, and K. M. Daniel. 1985. Be-havioral audiograms of the bullfrog (Rana catesbeiana) and thegreen tree frog (Hyla cinerea). J. Acoust. Soc. Am. 78:1236–1244.
Menard, S. 2001. Applied logistic regression analysis. Sage Uni-versity papers series on quantitative applications in the socialsciences, 07–106. Sage Publications, Thousand Oaks, CA.
Myers, C. W. 1987. New generic names for some Neotropical poi-son frogs (Dendrobatidae). Pap. Avul. Zool. 36:301–306.
Narins, P. M. 1992. Evolution of anuran chorus behavior: neuraland behavioral constraints. Am. Nat. 139:S90–S104.
———. 1995. Vibration communication in vertebrates. Pp. 127–148 in F. G. Barth, and A. Schmidt, eds. Ecology of sensing.Springer, Berlin.
Narins, P. M., and S. L. Smith. 1986. Clinal variation in anuranadvertisement calls: basis for acoustic isolation? Behav. Ecol.Sociobiol. 19:135–141.

1887MASKING INTERFERENCE IN FROG COMMUNICATION
Narins, P. M., and R. Zelick. 1988. The effects of noise on auditoryprocessing and behavior in amphibians. Pp. 511–536 in B.Fritzsch, M. J. Ryan, W. Wilczynski, T. E. Hetherington, andW. Walkowiak, eds. The evolution of the amphibian auditorysystem. John Wiley, New York.
Narins, P. M., W. Hodl, and D. S. Grabul. 2003. Bimodal signalrequisite for agonistic behavior in a dart-poison frog, Epipe-dobates femoralis. Proc. Natl. Acad. Sci. USA 100:577–580.
Nelson, D. A. 1989. The importance of invariant and distinctivefeatures in species recognition of bird song. Condor 91:120–130.
Nevo, E. 1973. Adaptive variation in size of cricket frogs. Ecology54:1271–1281.
Nevo, E., and R. R. Capranica. 1985. Evolutionary origin of etho-logical reproductive isolation in cricket frogs. Evol. Biol. 19:147–214.
Otte, D. 1974. Effects and functions in the evolution of signalingsystems. Annu. Rev. Ecol. System. 5:385–417.
Pollack, G. S. 1998. Neural processing of acoustic signals. Pp. 139–196 in R. R. Hoy, A. N. Popper, and R. R. Fay, eds. Comparativehearing: insects. Springer, New York.
Rodrıguez, L. B., and J. E. Cadle. 1990. A preliminary overviewof the herpetofauna of Cocha Cashu, Manu National Park, Peru.Pp. 410–425 in A. H. Gentry, ed. Four Neotropical rainforests.Yale Univ. Press, New Haven, CT.
Roithmair, M. 1992. Territoriality and male mating success in thedart-poison frog, Epipedobates femoralis (Dendrobatidae, Anu-ra). Ethology 92:331–343.
———. 1994a. Field studies on reproductive behaviour in two dart-poison frog species (Epipedobates femoralis, Epipedobates tri-vittatus) in Amazonian Peru. Herpetol. J. 4:77–85.
———. 1994b. Male territoriality and female mate selection in thedart-poison frog Epipedobates trivittatus (Dendrobatidae: Anu-ra). Copeia 1994:107–115.
Rojas, B., A. Amezquita, and A. Delgadillo. 2006. Matching andsymmetry in the frequency recognition curve of the poison frogEpipedobates trivittatus. Ethology 112:564–571.
Romer, H. 1998. The sensory ecology of acoustic communicationin insects. Pp. 63–96 in R. R. Hoy, A. N. Popper, and R. R. Fay,eds. Comparative hearing: insects. Springer, New York.
Ryan, M. J. 1988. Energy, calling, and selection. Am. Zool. 28:885–898.
Ryan, M. J., and E. A. Brenowitz. 1985. The role of body size,phylogeny, and ambient noise in the evolution of bird song. Am.Nat. 126:87–100.
Ryan, M. J., and A. S. Rand. 1993. Species recognition and sexualselection as a unitary problem in animal communication. Evo-lution 47:647–657.
Ryan, M. J., and W. Wilczynski. 1988. Coevolution of sender andreceiver: effect on local mate preference in cricket frogs. Science240:1786–1787.
———. 1991. Evolution of intraspecific variation in the advertise-ment call of a cricket frog (Acris crepitans, Hylidae). Biol. J.Linn. Soc. 44:249–271.
Ryan, M. J., S. A. Perrill, and W. Wilczynski. 1992. Auditory tuningand call frequency predict population-based mating preferencesin the cricket frog, Acris crepitans. Am. Nat. 139:1370–1383.
Schluter, A. 1980. Bio-akustische Untersuchungen an Dendroba-tiden in einem begrenzten Gebiet des tropischen Regenwaldesvon Peru (Amphibia: Salientia: Dendrobatidae). Salamandra 16:149–161.
Schwartz, J. J. 1993. Male calling behavior, female discriminationand acoustic interference in the Neotropical treefrog Hyla mi-crocephala under realistic acoustic conditions. Behav. Ecol. So-ciobiol. 32:401–414.
Schwartz, J. J., and H. C. Gerhardt. 1989. Spatially mediated releasefrom auditory masking in an anuran amphibian. J. Comp. Phy-siol. A 166:37–41.
Schwartz, J. J., and K. D. Wells. 1983. The influence of backgroundnoise on the behavior of a Neotropical treefrog, Hyla ebraccata.Herpetologica 39:121–129.
Silverstone, P. A. 1976. A revision of the poison-arrow frogs ofthe genus Phyllobates Bibron in Sagra (family Dendrobatidae).Nat. Hist. Mus. Los Angeles Co., Sci. Bull. 27:1–53.
Smouse, P. E., J. C. Long, and R. R. Sokal. 1986. Multiple re-gressions and correlation extensions of the Mantel tests of matrixcorrespondence. Syst. Zool. 35:627–632.
Sokal, R. R. 1979. Testing statistical significance of geographicvariation patterns. Syst. Zool. 28:227–232.
Toft, C. A., and W. E. Duellman. 1979. Anurans of the lower RıoLlullapichis, Amazonian Peru: a preliminary analysis of com-munity structure. Herpetologica 35:71–77.
Wagner, W. E., Jr. 1989. Social correlates of variation in malecalling behavior in Blanchard’s cricket frog, Acris crepitansblanchardi. Ethology 82:27–45.
Weary, D., and R. Weisman. 1993. SoundEdit. Vers. 2.0.3. Anim.Behav. 45:417–418.
Welch, A. M., R. D. Semlitsch, and H. C. Gerhardt. 1998. Callduration as an indicator of genetic quality in male gray tree frogs.Science 280:1928–1930.
Widemo, F., and S. A. Sæther. 1999. Beauty is in the eye of thebeholder: causes and consequences of variation in mating pref-erences. Trends Ecol. Evol. 14:26–31.
Wilczynski, W., and M. J. Ryan. 1999. Geographic variation inanimal communication systems. Pp. 234–261 in S. A. Foster andJ. A. Endler, eds. Geographic variation in behavior. Oxford Univ,Press, New York.
Wilczynski, W., B. E. McClelland, and A. S. Rand. 1993. Acoustic,auditory, and morphological divergence in three species of neo-tropical frog. J. Comp. Physiol. A 172:425–438.
Wilczynski, W., A. S. Rand, and M. J. Ryan. 1995. The processingof spectral cues by the call analysis system of the tungara frog,Physalaemus pustulosus. Anim. Behav. 49:911–929.
Wiley, R. H. 1994, Errors, exaggeration, and deception in animalcommunication. Pp. 157–189 in L. Real, ed. Behavioral mech-anisms in evolutionary ecology. Univ. of Chicago Press, Chi-cago, IL.
Wollerman, L. 1999. Acoustic interference limits call detection ina neotropical frog Hyla ebraccata. Anim. Behav. 57:529–536.
Wollerman, L., and R. H. Wiley. 2002. Possibilities for error duringcommunication by Neotropical frogs in a complex acoustic en-vironment. Behav. Ecol. Sociobiol. 52:465–473.
Zimmerman, B. L., and M. T. Rodrigues. 1990. Frogs, snakes, andlizards of the INPA-WWF reserves near Manaus, Brazil. Pp.427–453 in A. H. Gentry, ed. Four Neotropical rainforests. YaleUniv. Press, New Haven, CT.
Zimmermann, H., and E. Zimmermann. 1988. Etho-Taxonomie undzoogeographische Artengruppenbildung bei Pfeilgiftfroschen(Anura: Dendrobatidae). Salamandra 24:125–160.
Corresponding Editor: M. Noor




















