
Eutrophication and the dietary promotion of sea turtle tumors
Upload
independentCategory
view
0download
0
Estuarine, Coastal and Shelf Science 65 (2005) 697e707
www.elsevier.com/locate/ecss
Management of a shallow temperate estuary to controleutrophication: The effect of hydrodynamics on the
system’s nutrient loading
A.I. Lillebø*, J.M. Neto, I. Martins, T. Verdelhos, S. Leston, P.G. Cardoso,S.M. Ferreira, J.C. Marques, M.A. Pardal
IMAR e Institute of Marine Research, Department of Zoology, University of Coimbra, 3004-517 Coimbra, Portugal
Received 13 January 2005; accepted 15 July 2005
Available online 26 August 2005
Abstract
The Mondego estuary, a shallow warm-temperate intertidal system located on the west coast of Portugal, has for some decadesbeen under severe ecological stress, mainly caused by eutrophication. Water circulation in this system was, until 1998, mainlydependent on tides and on the freshwater input of a small tributary artificially controlled by a sluice. After 1998, the sluice opening
was effectively minimised to reduce the nutrient loading, and the system hydrodynamics improved due to engineering work in theupstream areas. The objective of the present study was to evaluate the effect of the mitigation measures implemented in 1998.Changes to the hydrodynamics of the system were assessed using precipitation and salinity data in relation to the concentrations of
dissolved inorganic nutrients, as well as the linkage between dissolved N:P ratios and the biological parameters (phytoplanktonchlorophyll a concentrations, green macroalgal biomass and seagrass biomass). Two distinctive periods were compared, over a tenyear period: from January 1993 to January 1997 and from January 1999 until January 2003. The effective reduction in the dissolvedN:P atomic ratio from 37.7 to 13.2 after 1998 is a result of lowered ammonia, but not the oxidised forms of nitrogen (nitrate plus
nitrite), or increased concentrations of dissolved inorganic phosphorus. Results suggest that the phytoplankton is not nutrientlimited, yet maximum and mean biomass of green macroalgae was reduced by one order of magnitude after the mitigation measures.This suggests that besides lowering the water residence time of the system, macroalgal growth became nitrogen limited. In parallel to
these changes the seagrass-covered area and biomass of Zostera noltii showed signs of recovery.� 2005 Elsevier Ltd. All rights reserved.
Keywords: hydrodynamics; river management; phosphorus; nitrogen; chlorophyll a; Ulva, Zostera, Hysteresis
1. Introduction
Worldwide, the nutrient enrichment of shallowcoastal areas has led to a decline in environmentalquality, especially close to densely populated areasaffected by eutrophication. As a result of nutrient-induced changes, particularly increased levels of growth
* Corresponding author.
E-mail address: [email protected] (A.I. Lillebø).
0272-7714/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.ecss.2005.07.009
limiting nutrients, perennial benthic macrophytes arecommonly replaced by fast-growing opportunistic algae(epiphytic green macroalgae and/or phytoplankton),which in many systems has affected the entire trophicstructure (e.g. Valiela et al., 1997; Raffaelli et al., 1998;Cardoso et al., 2004). Thus, conservation and restora-tion of coastal areas has become a priority during thelast decades (de Jonge, 2000; van Katwijk et al., 2000; deJonge and de Jong, 2002; Kendrick et al., 2002). Thegeneral aim of restoration programmes is to returna system from an altered or disturbed condition to

698 A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
a previously existing stable state condition (Zhang et al.,2003; Webster and Harris, 2004). Yet, when theresponse trajectory of the system to reduced drivingvariable (e.g. nutrient loads reduction) is very differentfrom the trajectory as the driving variable intensifies(e.g. rising nutrient loads), the system shows pro-nounced hysteresis (Scheffer et al., 2001; Beisner et al.,2003; Webster and Harris, 2004). Eutrophication maypush the system into an alternate stable state by positivefeedback processes from which recovery is very difficultto achieve (Scheffer et al., 2001). Thus, to assess thesuccess of restoration programmes it is necessary tounderstand the processes which have driven the ob-served ecological changes and to monitor the restorationprocess (Zhang et al., 2003; Cardoso et al., 2004, Pardalet al., 2004; Webster and Harris, 2004).
The main inputs contributing to the nutrient balanceand status of estuarine systems are the external pointand diffuse sources plus the internal biogeochemicalmineralization processes (Zwolsman, 1994; Lillebøet al., 2004; Coelho et al., 2004). Additionally, thephysical and chemical characteristics and the ecology ofthese shallow, yet highly complex and dynamic ecosys-tems are strongly influenced by the hydrology of thesystems (Martins et al., 2001).
Like many other estuaries worldwide (McMahon andWalker, 1998; Beukema et al., 1999; Norkko andBonsdorff, 2000; Sfriso et al., 2001), the Mondegoestuary has been under environmental stress by eutro-phication processes. As a consequence, the seagrass(Zostera noltii) bed has almost disappeared, reducing inextent from 15 ha in the early 1980’s to 0.02 ha in themid-1990’s (e.g. Dolbeth et al., 2003; Marques et al.,2003; Pardal et al., 2004). To evaluate the effect of theimplemented mitigation measures, in 1998, the objectiveof the present study was to assess the hydrodynamics,using precipitation and salinity data, in relation to theconcentrations of dissolved inorganic nutrients, as wellas the linkage between dissolved N:P ratios with thebiological parameters (phytoplankton chlorophyll a con-centrations, green macroalgal biomass and seagrassbiomass) before and after 1998, over a 10-year period.
2. Material and methods
2.1. Description of the study site
The Mondego estuary is a warm-temperate intertidalsystem located on the west coast of Portugal (Fig. 1).The construction of harbour facilities has imposedsevere changes of this system since the 1930s, particu-larly the construction of stone walls to regulate the mainnavigation channels, and the construction of small waterreservoirs for agriculture and aquaculture purposes. The
Mondego estuary is relatively small (1600 ha) andreceives agricultural runoff from 15 000 ha of upstreamcultivated land, mainly dominated by rice fields. Themain changes that have affected the system includemodified riverbed topography, changed hydrodynamics,increased water turbidity and increased concentration ofgrowth limiting nutrients (Martins et al., 2001).
The Mondego estuary is about 7 km long and is 2e3 km across at its widest part, and comprises a northernand southern arm, separated by the Murraceira island.The northern arm is the deeper (G10 m during high tide,tidal range 0.5e3.5 m), constituting the main navigationchannel and the location of the Figueira da Foz harbour.The southern arm is shallower (2e4 m during high tide),and is characterised by large areas of intertidal flatsexposed during low tide. Until 1998, the southern armwas almost silted up in the innermost areas, and the riveroutflow occurred mainly via the northern arm (Fig. 1).Water circulation in the southern arm was mostlydependent on the tides and on the freshwater input fromthe Pranto River, a small tributary with a flow controlledby a sluice, which was regulated according to the waterlevel of rice fields in the Mondego Valley (Fig. 1). Thisfreshwater input represented an important source ofnutrients into the southern arm. Thus, the system hasbeen under environmental stress by eutrophication pro-cesses. A previous study, conducted between January1993 and January 1997, concluded that the inter-annualvariation of macroalgal biomass in the Mondego estuarywas controlled by hydrodynamics, which in turndepended on precipitation and river management,according to the water requirements of the upstream ricecrop (Martins et al., 2001).
The Pranto river sluice opening was effectively reducedin 1998 to lower the nutrient loading of the south arm.Most of the nutrient enriched Pranto freshwater isdiverted to the northern arm by another sluice locatedfurther upstream. Furthermore, to improve the hydraulicregime, the upstream connection between the two armswas enlarged, allowing water to flow from the north armat high tide situations (Fig. 1). In addition, the remainingseagrass patches were protected with wooden stakes toprevent further disturbance of that area (by fishermenwho were digging in the sediment looking for bait), andseveral forums were convened to apprise local people ofthe ecological and economic importance of the seagrassbed.
2.2. Sampling procedure and analysis
The long-term monitoring program in the Mondegoestuary has been carried out since the early 1990s by anIMAR-Institute research team, within the scope ofEuropean and national projects. In order to assess theeutrophication process in the south arm of the estuary,three distinct areas were monitored (A, B and C in

699A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
Figueira da Foz Harbour
AtlanticOcean
Intertidal areas
(40º 08’ N, 8º 50’W)
POR
TU
GA
L
N
0 1km
South arm
North arm
Managementareas
AB
Long termmonitoring area
Sluice
PrantoRiver
C
Pranto river
South arm
SeaNorth arm
January 1993/1997
Pranto river
South armNorth arm
SeaNorth arm
1 Km
January 1999/2003
Fig. 1. The Mondego estuary, with the location of the long term monitoring area (A, B and C, where the macroalgal coverage gradient increases from
downstream to upstream); the alterations implemented in 1998 and the freshwater main point sources before and after the management.
Fig. 1). The choice of these areas was dictated byobservations of a macroalgal coverage gradient in thesouth arm, increasing from downstream to upstream.Originally, in the early 1980’s, the three areas werecovered by rooted macrophytes; however, as theeutrophication increased, together with human distur-bance caused by an intensive baitdigging, Zostera noltiideclined progressively. Currently, the seagrass bed isrestricted to the marine end of the estuary, having beenreplaced elsewhere by green macroalgae.
At each sampling site, primary producers andassociated macrofauna were sampled monthly (mini-mum 6 cores, with 141 cm2 section) between January
1993 and January 2003. Sampling was conducted duringthe day period just after ebbing. At the same time, watertemperature and salinity were measured, while watersamples (approximately 250 ml) were collected foranalysis of dissolved inorganic nitrogen and phospho-rus. Samples were immediately filtered (Whatman GF/Fglass-fibre filter) and stored frozen at �18 �C untilanalysis following standard methods described inLimnologisk Metodik (1992) for ammonia (NH3-N)and phosphate (PO4-P) and in Strickland and Parsons(1972) for nitrate (NO3-N), and nitrite (NO2-N). It ispossible that underestimation of ammonia occurred dueto temporary freezing of samples. The phytoplankton

700 A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
chlorophyll a (Chl a) determinations were performed byfiltering 0.5e1.0 L of water through Whatman GF/Cglass-fibre filters followed by extraction according toParsons et al. (1985). In the field and during trans-portation to the lab, samples were stored on ice and lightprotected. Samples from the three monitoring areas(A, B and C) were collected and analysed separately.Due to the small distance between the sites, data wereput together and presented/related as mean values(GSE) for the south arm of the estuary. Greenmacroalgae (mostlyUlva intestinalis andUlva compressa,according to the recent revision of Hayden et al., 2003)and the seagrass Zostera noltii, were dried (for 48 h at60 �C) and ash free dry weight (AFDW) was assessedafter combustion of samples for 8 h at 450 �C.
3. Results
The seasonal and inter-annual variation of salinityin the south arm of the Mondego estuary is strongly
influenced by precipitation (Fig. 2). These two param-eters showed a highly significant (P ! 0.001) negativelinear relationship for both periods of study, althoughwith higher correlation for the 1999e2003 period.
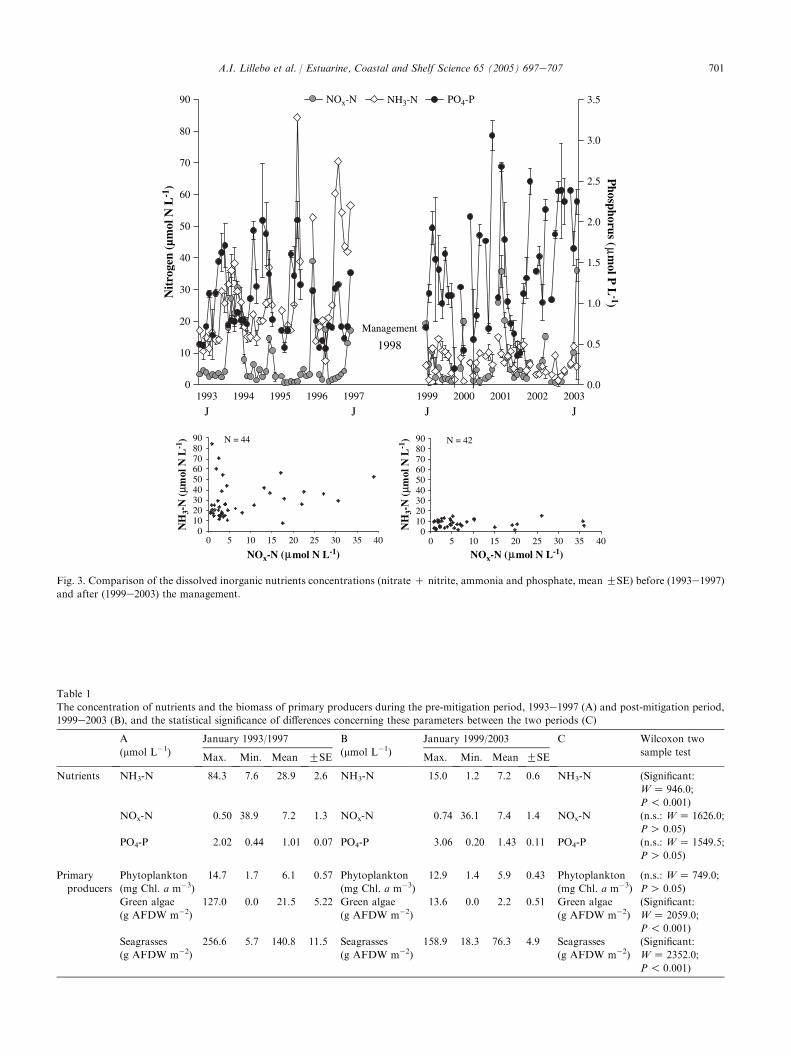
The changes implemented in the hydrodynamics ofthe system promoted a noteworthy reduction in theconcentration of the ammonia, but not in the oxidisedforms of nitrogen (nitrate plus nitrite) or in theconcentrations of the dissolved inorganic phosphorus(Fig. 3). The concentrations of ammonia rangedbetween 7.6 mmol L�1 and 84.3 mmol L�1 during the1993e1997 period, and after the Pranto river sluiceaperture was effectively reduced, the ammonia concen-trations dropped to values lower than 15 mmol L�1,(Table 1). The concentrations of oxidised nitrogen formswere not significantly different with a mean value of7.2 mmol N L�1 in the first period, and 7.4 mmol N L�1
after 1998 (Table 1). Similarly, there were no significantdifferences between phosphate concentrations beforeand after 1998 (Table 1). For both periods, before andafter 1998, the oxidised forms of nitrogen exhibited
J1993 1994 1995 1996 1997 1999 2000 2001 2002
J2003
J J
1998
Management
0
50
100
150
200
250
300
350
400
Pre
cipi
tati
on (
mm
)
0
5
10
15
20
25
30
35
Salinity
SalinityPrecipitation
0
5
10
15
20
25
30
35
50 100 200 300150 250 350
Precipitation (mm)
N = 46
0
5
10
15
20
25
30
35
050 100 200 300150 250 350
Precipitation (mm)
Salin
ity
R2 = 0.39 N = 40R2 = 0.67
0
Fig. 2. The variation of precipitation (mm) and salinity (mean GSE) between January 1993 and January 2003, and the relation between precipitation
(mm) and salinity before (1993e1997) and after (1999e2003) the management.
701A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
0
10
20
30
40
50
60
70
80
90
Nit
roge
n (µ
mol
N L
-1)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Phosphorus ( m
ol P L
-1)
NOx-N NH3-N PO4-P
J1993 1994 1995 1996 1997 1999 2000 2001 2002
J2003
J J
1998
Management
0102030405060708090
0 5 10 15 20 25 30 35 40
NOx-N ( mol N L-1)
NH
3-N
( m
ol N
L-1
) N = 44
0102030405060708090
0 5 10 15 20 25 30 35 40
N = 42
NH
3-N
( m
ol N
L-1
)
NOx-N ( mol N L-1)
Fig. 3. Comparison of the dissolved inorganic nutrients concentrations (nitrate C nitrite, ammonia and phosphate, mean GSE) before (1993e1997)
and after (1999e2003) the management.
Table 1
The concentration of nutrients and the biomass of primary producers during the pre-mitigation period, 1993e1997 (A) and post-mitigation period,
1999e2003 (B), and the statistical significance of differences concerning these parameters between the two periods (C)
A
(mmol L�1)
January 1993/1997 B
(mmol L�1)
January 1999/2003 C Wilcoxon two
sample testMax. Min. Mean GSE Max. Min. Mean GSE
Nutrients NH3-N 84.3 7.6 28.9 2.6 NH3-N 15.0 1.2 7.2 0.6 NH3-N (Significant:
WZ 946.0;
P ! 0.001)
NOx-N 0.50 38.9 7.2 1.3 NOx-N 0.74 36.1 7.4 1.4 NOx-N (n.s.: WZ 1626.0;
P O 0.05)
PO4-P 2.02 0.44 1.01 0.07 PO4-P 3.06 0.20 1.43 0.11 PO4-P (n.s.: WZ 1549.5;
P O 0.05)
Primary
producers
Phytoplankton
(mg Chl. a m�3)
14.7 1.7 6.1 0.57 Phytoplankton
(mg Chl. a m�3)
12.9 1.4 5.9 0.43 Phytoplankton
(mg Chl. a m�3)
(n.s.: WZ 749.0;
P O 0.05)
Green algae
(g AFDW m�2)
127.0 0.0 21.5 5.22 Green algae
(g AFDW m�2)
13.6 0.0 2.2 0.51 Green algae
(g AFDW m�2)
(Significant:
WZ 2059.0;
P ! 0.001)
Seagrasses
(g AFDW m�2)
256.6 5.7 140.8 11.5 Seagrasses
(g AFDW m�2)
158.9 18.3 76.3 4.9 Seagrasses
(g AFDW m�2)
(Significant:
WZ 2352.0;
P ! 0.001)

702 A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
a highly significant (P! 0.001) negative linear relationwith salinity (1993e97 R2Z 0.67, NZ 46 and 1999e03R2Z 0.67, NZ 42) and a highly significant (P! 0.001)positive linear relationship with precipitation (1993e97R2Z 0.40, NZ 47 and 1999e03 R2Z 0.43, NZ 41)(Fig. 4), suggesting that the Mondego system mainsource for these nutrients are the freshwater inputs.
The mean water temperature showed a clear seasonalvariation, closely following the air temperature (Fig. 5).Phosphate concentrations showed a highly significant(P ! 0.001) positive linear relationship with watertemperature (R2 Z 0.44, N Z 40) for the 1993e1997
period, suggesting a clear dependency on mineralizationprocesses. After 1998, this relationship disappeared andhigh concentrations of phosphate were present even atlow temperatures.
Ammonia did not show any relationship with pre-cipitation, salinity, temperature, nor with the otherdissolved inorganic nutrients (NOx-N and PO4-P). Thedissolved N:P atomic ratio showed a significant decrease(Wilcoxon two sample test: W Z 2882.5; P ! 0.001)from the pre- (mean N:P Z 37.7) to post-management(mean N:P Z 13.2) period (Fig. 6), mostly due to thereduction in the concentration of ammonia. This
J1993 1994 1995 1996 1997 1999 2000 2001 2002
J2003
J J
1998
Management
0
5
10
15
20
25
30
35
Salin
ity
0
5
10
15
20
25
30
35
40
45NOx-NSalinity
NO
x ( mol N
L-1)
05
1015202530354045
0 5 10 15 20 25 30 35
Salinity
05
1015202530354045
0 5 10 15 20 25 30 35
Salinity
05
1015202530354045
0 50 100 150 200 250 350
Precipitation (mm)
05
1015202530354045
100 150 250300 0 50 200 300 350
Precipitation (mm)
NO
x (
mol
N L
-1)
N = 47R2 = 0.40
N = 46R2 = 0.67
N = 42R2 = 0.67
N = 41R2 = 0.43
NO
x (
mol
N L
-1)
Fig. 4. The variation of salinity (mean G SE) and the concentration of the oxidised forms of nitrogen (NOx Z NO3-N C NO2-N, mean GSE)
between January 1993 and January 2003, and the relation between salinity and the oxidised forms of nitrogen, and precipitation and the oxidised
forms of nitrogen before (1993e1997) and after (1999e2003) the management.

703A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
R2 = 0.44N=40
0 5 10 15 20 25 30 35
Water temperature (ºC) Water temperature (ºC)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
0 5 10 15 20 25 30 35
N=42
Tem
pera
ture
(ºC
)
1998
Management
J1993 1994 1995 1996 1997 1999 2000 2001 2002
J2003
J J
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5PO4-PTemp. air Temp. water
0
5
10
15
20
25
30
35
Phosphorus ( m
ol P L
-1)P
O4-
P (
m
ol P
L-1
)
Fig. 5. The variation of mean temperature ( �C) in the air and in the water, and the concentration of phosphate (PO4-P, mean GSE) between January
1993 and January 2003, and the relation between mean water temperature and the concentration of phosphate before (1993e1997) and after (1999e2003) the management.
difference is evidenced by analysing the dissolved N:Patomic ratio, separating the nitrogen oxidize forms(NOx-N:P) from ammonia (NH3-N:P) (Fig. 6). Forboth periods, the dissolved N:P atomic ratio exhibiteda highly significant (P ! 0.001) negative linear relationwith salinity (1993e97 R2 Z 0.55, N Z 35 and 1999e03R2 Z 0.67, N Z 40) (Fig. 6).
The biological parameters for opportunistic primaryproducers showed that green macroalgal biomass,mostly Ulva intestinalis and Ulva compressa, wassignificantly reduced (Table 1) after the implementationof the mitigation measures (Fig. 7A): maximum biomass127.0 g AFDW m�2 before and 13.6 g AFDW m�2 after1998. Yet, the phytoplankton chlorophyll a concentra-tions did not differ (Fig. 7): mean concentration:6.1 mg m�3 G 0.6 SE before and 5.9 mg m�3 G 0.4 SEafter 1998 (Table 1).
The biomass of Zostera noltii sharply decreased fromJanuary 1993 to January 1997, (Fig. 7B). After themitigation measures, the seagrass partly recovered:maximum and minimum biomass was respectively256.6 and 5.7 g AFDW m�2 before and 158.9 and18.3 g AFDWm�2 after 1988. However, the biomass wasstill significantly lesser than in the first period (Table 1).
4. Discussion
Water circulation in the south arm of the Mondegoestuary was mainly dependent on tides and freshwaterinput by the tributary Pranto river during the period of1993 to 1997. This is clear from the highly significantnegative linear relation between salinity and precipita-tion. Comparatively, in the post-management period,the explained variance was lower, which is supported bya time lag between the occurrence of precipitation andthe freshwater discharge from the River sluice (Martinset al., 2001). After the enlargement of the upstreamconnection between the two arms, higher variance in thenegative linear relation between salinity and precipita-tion clearly suggests a higher freshwater input from thenorth arm through overflow episodes. Despite thechanges in the hydrodynamics of the system, the highestsalinities were recorded during the warmer/dry periods,when the freshwater inputs are lower and the runoffepisodes negligible. This indicates a major penetrationof seawater during spring and summer. On the otherhand, during autumn and winter the considerableincrease and predominance of freshwater inputs lowerthe salinity.

704 A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
R2 = 0.55 R2 = 0.67N = 35
0102030405060708090
0 5 10 15 20 25 30 35
Salinity Salinity
N:P
ato
mic
rat
io
N = 40
0102030405060708090
0 5 10 15 20 25 30 35
N:P
ato
mic
rat
io
N:P
atomic ratio
0
10
20
30
40
50
60
70
80
90
0
10
20
30
40
50
60
70
80
90NH3-N:P NOx-N:P N:P
J1993 1994 1995 1996 1997 1999 2000 2001 2002
J2003
J J
1998
Management
N:P
ato
mic
rat
io
Fig. 6. Comparison of the N:P atomic ratio in the water, between January 1993 and January 2003, considering the total dissolved inorganic nitrogen,
just the ammonia formof nitrogen and just the oxidised forms of nitrogen, and the relation between theN:P atomic ratio and salinity before (1993e1997)
and after (1999e2003) the management.
For both periods, before and after 1998, there isa highly significant negative linear relation betweensalinity and the oxidised forms of dissolved nitrogenand a highly significant positive linear relation betweenprecipitation and the oxidised forms of dissolved nitro-gen, suggesting a strong dependency on the freshwaterinputs from diffuse and/or point sources The oxidizednitrogen forms were not significantly reduced after 1998.Probably due to the fact that concentrations of nitrogenare often several times higher in rivers than in receivingcoastal seawater (Yin et al., 2001), and this estuarinesystem receives an agricultural runoff from an upstreamcultivated area ten times larger than the estuary itself.Moreover, rain water can also constitute a source ofnitrate to aquatic systems (Berner and Berner, 1996).
Rivers may supply much of the nitrogen andphosphorus to estuaries, but most of it may alsointernally recycle within the estuary through biogeo-chemical processes. Thus, external sources for nutrientscan also include the regeneration of nutrients fromorganic and inorganic matter carried by rivers (Bernerand Berner, 1996). So, the Pranto river could alsoconstitute a source of nutrients in the form of suspendedparticulate organic and/or inorganic matter.
The absence of a linear relationship between ammoniaand salinity or precipitation, and between phosphateand salinity or precipitation, before and after 1998,suggests that the main source of these nutrients, in thedissolved form, was not dependent on the freshwaterinputs. Both nutrients can be directly related to thebiological activity and mineralization of organic matterwithin the system (e.g. Berner and Berner, 1996; Lillebøet al., 2002). In fact, most of the phosphorus enters thesystem in the particulate form (Lillebø et al., 2002). Forthe period 1993 to 1997, the highly significant positivelinear relation of phosphate with temperature impliesa dependency on mineralization processes (Asmus et al.,2000; Lillebø et al., 2002, 2004). This means thatecological stress may still persist due to internal P-loading (Coelho et al., 2004) in a similar way as observedin other temperate estuaries (Holmboe et al., 1999) andshallow lakes (e.g. Jensen and Andersen, 1992; Zhanget al., 2003). After 1998, this relation is not significant,since high phosphate concentrations also persist intoautumn and winter, with low temperatures (between6 �C and 12 �C), suggesting the existence of additionalphosphate sources, possibly aquaculture farms or smallindustries.

705A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
Ammonia did not show any relationship withtemperature, which is in agreement with previous studiesshowing that primary producers may control thesediment/water flux of ammonia (Lillebø et al., 2002).Higher ammonia efflux rates occur seasonally, whenprimary producers are less active (autumn and winter),
J
1993 1994 1995 1996 1997 1999 2000 2001 2002
J
2003
J J
1998Management
0
5
10
15
20
25
30
35
Chl
orop
hyll
a (m
g m
-3)
0
20
40
60
80
100
120
140
160
180
200
Green m
acroalgal (g AF
DW
m-2)
Chl. a concentration Green macroalgal biomass
J
1993 1994 1995 1996 1997 1999 2000 2001 2002
J2003
J J
A
B
C
R2 = 0.85
R2 = 0.54N = 42
N = 45
0
50
100
150
200
250
300
Zos
tera
nol
tii (
g A
FD
W m
-2)
1998
Management
0
25
50
75
100
200 4000 100 300 500
Nitrogen loading rates (kg ha-1 yr-1)
Per
cent
pri
mar
y pr
oduc
tion
phytoplankton
macroalgae
eelgrass
Fig. 7. A) Comparison of the mean concentration of chlorophyll a
(mg m�3, mean GSE) and the mean biomass of green macroalgae
(mg AFDW m�2, mean GSE) before the management (1993e1997)
and after (1999e2003); B) Comparison of the mean biomass of Zostera
noltii at the long monitoring field station A (mg AFDW m�2) before
the management (1993e1997) and after (1999e2003); C) Hypothetical
pattern of change in the relative contribution of these three major
groups of producers in shallow temperate estuaries with shorter water
residence time, redrawn after Valiela et al. (1997).
and daily when heterotrophic respiration is not com-pensated by primary production (night period) (Lillebøet al., 2002).
After the diversion of most of the freshwater from thePranto river directly into the northern arm, throughanother sluice located further upstream, the concen-trations of ammonia in the south arm were effectivelyreduced. This suggests that the river Pranto waspreviously the major source for the ammonia form ofnitrogen. In addition, a linear correlation often existsbetween ammonia concentrations and the suspendedparticulate matter in the water column of temperateintertidal shallow systems (Rocha et al., 1995).
For both periods (pre- and post-mitigation), thedissolved N:P atomic ratio exhibited a highly significantnegative linear relation with salinity, suggesting a closerelation with the freshwater inputs. Since no relevantchanges occurred in the concentrations of dissolvedinorganic phosphorus between pre- and post-mitigation,lack of ammonia derived from the Pranto river causedthe effective reduction in the N:P atomic ratio after theregulation. The opportunistic primary producer param-eters suggest that phytoplankton is not nutrient limited,since chlorophyll a concentrations did not show anysignificant changes throughout the period between 1993and 2003. Due to a comparatively higher surface area/volume relation, phytoplankton have higher affinity fornitrogen forms, with higher uptake rates than macro-algae for both ammonia and nitrate per unit of biomass(Hein et al., 1995; Collos, 1998). However, this onlymakes phytoplankton competitively superior under lownitrogen availability, which is closely related to nitrogenloading and water residence time (Hein et al., 1995;Valiela et al., 1997). On the other hand, the biomassof green macroalgae was reduced about one order ofmagnitude after the mitigation measures. This showsthat the improvement of the hydrodynamics of thesystem lowered dissolved inorganic nitrogen concen-trations, which caused macroalgae with low nitrogenaffinity to be nitrogen limited. In fact, nitrogen oftenappears to be the limiting nutrient in estuarine systems(Hein et al., 1995; Yin et al., 2001), namely in the formof ammonia (Rocha et al., 1995). Ultimately, theresponse of phytoplankton, green macroalgae and theseagrass Zostera noltii to the changes of nitrogenloading in the Mondego estuary clearly confirms thepattern of relative contribution of these three majorgroups of producers, in shallow temperate estuaries withshorter water residence time, proposed by Valiela et al.(1997) (Fig. 7C).
Aquatic ecosystems may shift between an oligotro-phic/benthic state (with clear water and macrophytedominated) and an euthrophic/pelagic state (with turbidwater and plankton dominated) (Valiela et al., 1997;Scheffer et al., 2001; Zhang et al., 2003; Webster andHarris, 2004). The switch between the two states is often

706 A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
abrupt and the ecosystem response to perturbation ishighly non-linear and complex (Scheffer et al., 2001;Beisner et al., 2003; Marques et al., 2003; Zhang et al.,2003; Cardoso et al., 2004). The non-linearity arisesfrom the strong interactions between the benthosand the pelagic compartments and from competitionbetween primary producers for light and nutrients(Scheffer et al., 2001; Zhang et al., 2003; Webster andHarris, 2004). When the mean dissolved N:P ratio wasreduced in the Mondego system, the large greenmacroalgae biomass was reduced, followed by anincrease of Zostera noltii biomass and covered area.Yet, seagrass biomass and aerial cover remained muchlower than during the initial state. These primaryproducers are bio-stabilisers due to the immediatechanges in the physical environment (e.g. decreasingtidal currents, wave action and sediment resuspensionand enhancing sediment cohesiveness and settling ofsuspended matter (Widdows and Brinsley, 2002). Thehysteresis response may be related to the sedimentarychanges that followed the loss of seagrass (Cardosoet al., 2004).
The state of a system depends on the history ofdisturbances, since an ecosystem is set to provideresilience to buffer changes for certain periods of time(Beisner et al., 2003; Zhang et al., 2003). The restorationof ecosystems into a natural stable state should befocused on monitoring events after the cessation ofa perturbation and the responses to the reversal ofa perturbation. Thus, integrated catchment managementand the reduction of nutrient loads from all sources,together with additional measures (e.g. directionalhabitat reconstruction) are the way for long-termmanagement solution for estuarine systems.
Acknowledgments
This study was supported by the POCTI e Formar eQualificar e Medida 1.1 program (Portuguese FCT)through a Post-Doc grant (SFRH/BPD/5650/2001) (A.I. Lillebø), and a PhD grant (Praxis XXI/BD/18422/98)to J.M. Neto.
References
Asmus, R.M., Sprung, M., Asmus, H., 2000. Nutrient fluxes in
intertidal communities of a southEuropean lagoon (RiaFormosa)e
similarities and differences with a northern wadden sea bay (Syl-
Rømø bay). Hydrobiologia 436, 217e235.
Beisner, B.E., Haydon, D.T., Cuddington, K., 2003. Alternative stable
stages in ecology. Frontiers in Ecology and the Environment 1,
376e382. www.frontiersinecoly.org.
Berner, E.K., Berner, R.A., 1996. Marginal marine environments:
estuaries. In: The Global Environment: Water, Air and Geo-
chemical Cycles. Prentice-Hall, Upper Saddle River, NJ, pp.
284e311.
Beukema, J.J., Flach, G.C., Dekker, R., Starink, M., 1999. A long-
term study of the recovery of the macrozoobenthos on large
defaunated plots on a tidal flat in the Wadden Sea. Journal of Sea
Research 42, 235e254.Cardoso, P., Pardal, M.A., Lillebø, A.I., Ferreira, S.M., Raffaelli, D.,
Marques, J.C., 2004. Dynamic changes in seagrass assemblages
under eutrophication and implications for recovery. Journal of
Experimental Marine Biology and Ecology 302, 233e248.Coelho, J.P., Flindt, M.R., Jensen, H.S., Lillebø, A.I., Pardal, M.A.,
2004. Phosphorus speciation and availability in intertidal sediments
of a temperate estuary: relation to eutrophication and annual
P-fluxes. Estuarine, Coastal and Shelf Science 61, 583e590.Collos, Y., 1998. Nitrate uptake, nitrite release and uptake, and new
production estimates.MarineEcologyProgress Series 171, 293e301.
Dolbeth, M., Pardal, M.A., Lillebø, A.I., Azeiteiro, U., Marques, J.C.,
2003. Short term and long term effects of eutrophication on the
secondary production of an intertidal macrobenthic community.
Marine Biology 143, 1229e1238.
Hayden, H.S., Blomster, J., Maggs, C.A., Silva, P.C., Stanhope, M.J.,
Waalland, J.R., 2003. Linnaeus was right all along: Ulva and
Enteromorpha are not distinct genera. European Journal of
Phycology 38, 277e294.
Hein, M., Pedersen, M.F., Sand-Jensen, M., 1995. Size dependent
nitrogen uptake in micro-and macroalgae. Marine Ecology
Progress Series 118, 247e253.
Holmboe, N., Jensen, H.S., Andersen, F.Ø., 1999. Nutrient addition
bioassays as indicators of nutrient limitation of phytoplankton in
an eutrophic estuary. Marine Ecology Progress Series 186, 95e104.
Jensen, H.S., Andersen, F.Ø., 1992. Importance of temperature,
nitrate and pH for phosphate release from aerobic sediments of
four shallow, eutrophic lakes. Limnology and Oceanography 37,
577e589.
de Jonge, V.N., 2000. Importance of temporal and spatial scales in
applying biological and physical process knowledge in coastal
management, an example for the Ems estuary. Continental Shelf
Research 20, 1655e1686.
de Jonge, V.N., de Jong, D.J., 2002. Ecological restoration in coastal
areas in the Netherlands: concepts, dilemmas and some examples.
Hydrobiologia 478, 7e28.
van Katwijk, M.M., Hermus, D.C.R., de Jong, D.J., Asmus, R.M., de
Jonge, V.N., 2000. Habitat suitability of the Wadden Sea for
restoration of Zostera marina beds. Helgoland Marine Research
54, 117e128.
Kendrick, G.A., Aylward, M.J., Hegge, M.L., Cambridge, K.,
Hillman, K., Wyllie, A., Lord, D.A., 2002. Changes in seagrass
coverage in Cockburn Sound, Western Australia between 1967 and
1999. Aquatic Botany 73, 75e87.
Lillebø, A.I., Flindt, M.R., Pardal, M.A., Martins, I., Neto, J.M.,
Marques, J.C., 2002. Nutrient dynamics in the intertidal pools of
the Mondego estuary. II e Seasonal efflux of PO4-P and NH4-N in
bare bottom and vegetated pools. In: Pardal, M.A., Marques, J.C.,
Graca, M.A. (Eds.), Aquatic Ecology of the Mondego River Basin.
Global Importance of Local Experience. Imprensa da Universidade
de Coimbra, Coimbra, pp. 257e272.
Lillebø, A.I., Neto, J.M., Flindt, M.R., Marques, J.C., Pardal, M.A.,
2004. Phosphorous dynamics in a temperate intertidal estuary.
Estuarine, Coastal and Shelf Science 61, 101e109.
Limnologisk Metodik, 1992. Ferskvandsbiologisk Laboratorium. Kø-
benhavnsUniversitet (Ed.), AkademiskForlag.København, 172 pp.
Marques, J.C., Nielsen, S.N., Pardal, M.A., Jørgensen, S.E., 2003.
Impact of eutrophication and river management within a frame-
work of ecosystem theories. Ecological Modelling 166, 147e168.
Martins, I., Pardal, M.A., Lillebø, A.I., Flindt, M.R., Marques, J.C.,
2001. Hydrodynamics as a major factor controlling the occurrence
of green macroalgal blooms in a eutrophic estuary: a case study on
the influence of precipitation and river management. Estuarine,
Coastal and Shelf Science 52, 165e177.

707A.I. Lillebø et al. / Estuarine, Coastal and Shelf Science 65 (2005) 697e707
McMahon, K., Walker, D.I., 1998. Fate of seasonal, terrestrial
nutrient inputs to a shallow seagrass dominated embayment.
Estuarine, Coastal and Shelf Science 46, 15e25.
Norkko, A., Bonsdorff, E., Norkko, A., 2000. Drifting algal mats as an
alternative habitat for benthic invertebrates: species specific
responses to a transient resource. Journal of Experimental Marine
Biology and Ecology 248, 79e104.
Pardal, M.A., Cardoso, P.G., Sousa, J.P., Marques, J.C., Raffaelli, D.,
2004. Assessing environmental quality: a novel approach. Marine
Ecology Progress Series 267, 1e8.
Parsons, T.R., Maita, Y., Lally, C.M., 1985. Pigments. In: A Manual
of Chemical and Biological Methods for Seawater Analysis.
Pergamon Press, pp. 101e104.
Raffaelli, D.G., Raven, J.A., Poole, L.J., 1998. Ecological impact of
green macroalgal blooms. Oceanography and Marine Biology.
Annual Review 36, 97e125.Rocha, C., Cabecadas, G., Brogueira, M.J., 1995. On the importance
of sediment-water exchange processes of ammonia to primary
production in shallow areas of the Sado estuary (Portugal).
Netherlands Journal of Aquatic Ecology 29, 265e273.
Scheffer, M., Carpenter, S., Foley, J.A., Folke, C., Walker, B., 2001.
Catastrophic shifts in ecosystems. Nature 413, 591e596.
Sfriso, A., Birkmeyer, T., Ghetti, P.F., 2001. Benthic macrofauna
changes in areas of Venice lagoon populated by seagrasses or
seaweeds. Marine Environmental Research 52, 323e349.
Strickland, J.D.M., Parsons, T.R., 1972. A Practical Handbook of
Seawater Analysis (Bulletin 167, second ed.). In: Fisheries Research
Board of Canada, Ottawa, pp. 71e80.
Valiela, I., McClelland, J., Hauxwell, J., Behr, P.J., Hersh, D.,
Foreman, K., 1997. Macroalgal blooms in shallow estuaries:
Controls and ecophysiological and ecosystem consequences.
Limnology and Oceanography 4215, 1105e1118.
Webster, I.T., Harris, G.P., 2004. Anthropogenic impacts on the
ecosystems of coastal lagoons: modelling fundamental biogeo-
chemistry process and management implications. Marine and
Freshwater Research 55, 67e78.
Widdows, J., Brinsley, M., 2002. Impact of biotic and abiotic processes
on sediment dynamics and the consequences to the structure and
functioning of the intertidal zone. Journal of Sea Research 48,
143e156.
Yin, K., Qiam, P., Wu, M.C.S., Chen, J.C., Huang, L., Song, X.,
Jian, W., 2001. Shift from P to N limitation of phytoplankton
growth across the Pearl River estuarine plume during summer.
Marine Ecology Progress Series 221, 17e28.Zhang, J., Jørgensen, S.E., Beklioglu, M., Ince, O., 2003. Hysteresis in
vegetation shift e Lake Mogan prognoses. Ecological Modelling
164, 227e238.
Zwolsman, J.J.G., 1994. Seasonal variability and biogeochemistry of
phosphorus in the Scheldt Estuary, South-west Netherlands.
Estuarine, Coastal and Shelf Science 39, 227e248.
Copyright © 2022 FDOKUMEN




















