
The romance of William of Palerne (otherwise known as The ...

G
I
Msi
NRa
b
c
d
a
ARR2A
KCIMMS
I
rasic
bcf
dT
0d
ARTICLE IN PRESSModel
MBIO-50678; No. of Pages 10
Immunobiology xxx (2011) xxx–xxx
Contents lists available at ScienceDirect
Immunobiology
journa l homepage: www.e lsev ier .de / imbio
acrophages from IBD patients exhibit defective tumour necrosis factor-�ecretion but otherwise normal or augmented pro-inflammatory responses tonfection
air Camposa,1, Fernando Magrob,d,1, Ana Rita Castroa, Joana Cabrala, Pedro Rodriguesa, Ricardo Silvaa,ui Appelbergc, Susana Rodriguesd, Susana Lopesd, Guilherme Macedod, Amélia Sarmentoa,c,∗
CEBIMED – Biomedicine Research Center, Health Sciences Faculty, University Fernando Pessoa, Porto, PortugalInstitute of Pharmacology and Therapeutics, Faculty of Medicine, University of Porto, Porto, PortugalIBMC – Institute for Cellular and Molecular Biology, University of Porto, Porto, PortugalGastroenterology Department, Hospital of São João, Porto, Portugal
r t i c l e i n f o
rticle history:eceived 16 July 2010eceived in revised form9 December 2010ccepted 4 January 2011
eywords:ytokines
BDacrophagesycobacterium avium spp. paratuberculosis
a b s t r a c t
Defects in macrophage function have been implicated in the establishment of Crohn’s disease (CD). How-ever, the response of macrophages from CD patients to live bacteria, particularly Mycobacterium aviumsubsp. paratuberculosis (MAP), has not been addressed. Considering MAP has long been associated toCD, our objective was to assess whether macrophages from CD patients showed impaired inflammatoryresponse to infection by MAP comparing to M. avium subsp. avium (MA) and other live intestinal commen-sal bacteria. Human peripheral blood monocyte-derived macrophages were obtained from CD patients,ulcerative colitis (UC) patients and controls. Following in vitro infection with MAP, MA, Escherichia coli orEnterococcus faecalis, cytokine levels and cell surface receptor expression were evaluated at different timepoints. Macrophages from CD patients showed impaired TNF-� secretion in response to bacterial chal-lenge, but augmented IL-23 secretion and preserved IL-12 secretion and CD-40 expression. In addition,CD macrophages showed low IL-10 secretion. Macrophages from IBD patients showed increased expres-
urface receptorssion of TLR-2 and -4, unaffected by infection. Differences in cytokine secretion observed after bacterialchallenge were not MAP-specific, as other bacteria (E. coli and MA) showed similar effects. Macrophagesfrom UC patients showed a less compromised TNF-� synthesis in response to mycobacterial infectionthan CD macrophages, with increased constitutive IL-12 secretion, and preserved IL-10 secretion. Theincreased IL-23 levels in response to infection and decreased IL-10 production observed in macrophagestribu
from CD patients may conntroduction
Although the aetiology of inflammatory bowel disease (IBD)emains unknown, current knowledge supports that inappropri-
Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
te chronic activation of the innate and adaptive mucosal immuneystems in a genetically susceptible host can lead to mucosalnflammation. Recently, over 30 genomic loci have been impli-ated in the genetic susceptibility to Crohn’s disease (CD) (Van
Abbreviations: CD, Crohn’s disease; UC, ulcerative colitis; IBD, inflammatoryowel disease; HCs, healthy controls; MAP, Mycobacterium avium subsp. paratuber-ulosis; MA, Mycobacterium avium subsp. avium; EC, Escherichia coli; EF, Enterococcusaecalis; TNF, tumour necrosis factor; IL, interleukin.∗ Corresponding author at: CEBIMED, Faculdade de Ciências da Saúde – Universi-ade Fernando Pessoa, Rua Carlos da Maia, 296, 4200-150 Porto, Portugal.el.: +351 225074630; fax: +351 225074637.
E-mail address: [email protected] (A. Sarmento).1 Authors contributed equally to this work.
171-2985/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.imbio.2011.01.002
te to the inflammatory exacerbation observed in those patients.© 2011 Elsevier GmbH. All rights reserved.
Limbergen et al. 2009), and several risk loci are shared with ulcera-tive colitis (UC) (Anderson et al. 2009). CD susceptibility loci includegenes involved in innate immunity mechanisms (innate patternrecognition receptors, autophagy), adaptive immunity mecha-nisms (lymphocyte subset differentiation, antigen presentation)and maintenance of epithelial barrier integrity (Van Limbergenet al. 2009).
Enteric microflora also seem to play a pivotal role in the ini-tiation and maintenance of disease. In fact, mice with abnormaleffector T cell or Treg responses develop colitis only in the pres-ence of intestinal microbiota, but fail to do so when housed undergermfree conditions (Strober et al. 2007). Whether a particularmicroorganism, or a set of microorganisms may cause the disease,
D patients exhibit defective tumour necrosis factor-� secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
remains to be proven.Numerous studies have shown an association between coloniza-
tion by an atypical mycobacterium, Mycobacterium avium subsp.paratuberculosis (MAP), and CD (Abubakar et al. 2008; Feller et al.2007). MAP is the agent of Johne’s disease, which affects rumi-

ING
I
2 nobiol
nCNrMigtt
(rafr(durFni
iarmdv
sbUwmn
op(TrwdopcitftRnecpa1s
CaePti
ARTICLEModel
MBIO-50678; No. of Pages 10
N. Campos et al. / Immu
ants and other mammals and shares some characteristics withD (Glawischnig et al. 2006; Judge et al. 2005; Mcclure et al. 1987).evertheless, due to the complexity in MAP detection and to the
esults of clinical antimicrobial trials (Selby et al. 2007) the role ofAP in CD has not been clarified. Despite these findings, it is legit-
mate to assume that MAP may have the capacity to survive androw in macrophages from genetically susceptible human hosts,hereby playing a role in the exacerbated inflammatory responseo intestinal microflora observed in CD.
Several studies, including a recent research by Smith et al.2009), suggest that a defective macrophage function may beesponsible for the establishment of CD (Marks et al. 2006; Xaviernd Podolsky 2007). Their findings agree with the associationound between innate immunity gene polymorphisms and higherisk for developing CD (Van Limbergen et al. 2009). Smith et al.2009) found that macrophages isolated from CD patients showedecreased pro-inflammatory cytokine production following stim-lation with heat-killed Escherichia coli. However, macrophageesponses to live bacteria, particularly MAP, have not been studied.urthermore, putative defects in cell surface receptor expression,amely pattern-recognition receptors or other receptors involved
n the triggering of inflammation, have not yet been investigated.This study intends to shed some light on a number of pend-
ng questions: (1) assuming a possible link between MAP and CD,re macrophage responses to MAP impaired? (2) Are macrophageesponses to commensal live bacteria affected? (3) Are otheracrophage features, namely cell surface receptor expression,
efective in CD? Is macrophage secretion of IL-23, not tested pre-iously by others, impaired in CD?
In an attempt to explore these questions, we studied cytokineecretion and cell surface receptor expression by human peripherallood monocyte-derived macrophages obtained from CD patients,C patients and healthy controls (HCs), after in vitro infectionith MAP, M. avium subsp. avium (MA) (a MAP-related species ofycobacteria, not previously associated with CD), and two intesti-
al commensal bacteria (Enterococcus faecalis (EF) and E. coli (EC)).For cytokine profile evaluation, we studied the secretion
f TNF-�, IL-12, IL-23 and IL-10. TNF-�, IL-12 and IL-23 areroinflammatory cytokines implicated in IBD immunopathologyMonteleone et al. 2009; Van Deventer 1997; Peluso et al. 2006).NF-� has potent proinflammatory effects, being involved in theecruitment of inflammatory cells to the site of inflammation, asell as in granuloma formation (Van Deventer 1997). TNF-� pro-uction was reported to be increased in gut culture supernatantsf MAP-positive CD patients, as compared to MAP-negative CDatients (Clancy et al. 2007). IL-12 and IL-23, two heterodimericytokines sharing one subunit (p40), are important mediators ofntestinal inflammation. IL-12 plays a role in T cell commitment tohe Th1 lineage (Peluso et al. 2006; Li et al. 2009). IL-23 is importantor the maintenance of a Th17 response, leading to IL-17A secre-ion and tissue injury (Monteleone et al. 2009; Liu et al. 2009).ecently IL-23 was implicated as a key cytokine both in mice intesti-al inflammation (Hue et al. 2006) and in human CD (Monteleonet al. 2009). IL-10 has long been recognized as an anti-inflammatoryytokine. Defects in IL-10 production have been associated with theediatric onset of IBD (Ebert et al. 2009) and IL10-deficient micere known to develop spontaneous colitis (Kuhn et al. 1993). IL-0 was implicated in maintaining Treg cells’ Foxp3 expression anduppressive function in mice with colitis (Murai et al. 2009).
For surface receptor evaluation, we studied the expression ofD-40, TLR-2, TLR-4 and HLA-DR. CD-40 is a key molecule in the
Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
ctivation of immunity and inflammation and is known to be over-xpressed in IBD patients (Danese et al. 2006; Battaglia et al. 1999;olese et al. 2002). TLR-2 and TLR-4 are involved in the recogni-ion of MAP and other mycobacteria (Ferwerda et al. 2007). Anncrease in both TLR receptors was reported in gut biopsies from
PRESSogy xxx (2011) xxx–xxx
IBD patients (Frolova et al. 2008). HLA-DR receptor is involved inantigen-presentation to T cells. Several HLA-DR polymorphisms(Muro et al. 2009) and TLR-associated polymorphisms (includingTLR-2 and -4) were found to be involved in IBD (Browning et al.2007; De Jager et al. 2007; Pierik et al. 2006; Van Limbergen et al.2009).
Materials and methods
Patients and samples
In this study, 69 individuals were included: 25 HCs (mean age,36.16 years; range, 19–71 years; 8 male, 17 female), 22 with CD(mean age, 38.29 years; range, 21–73 years; 11 male, 11 female)and 22 with UC (mean age, 46.09 years; range, 29–79 years; 11male, 11 female). All IBD patients were on 5-aminosalicylate (5-ASA) therapy. 17 CD patients and 6 UC patients were also receivingAzathioprine therapy. None of the patients were receiving anti-TNFtherapy within 3 months of enrolment.
Informed consent was obtained in accordance with the insti-tutional review board regulations at IBMC, University FernandoPessoa and Hospital de São João. Patients with CD were recruitedfrom the Gastroenterology Department at Hospital de São João. CDdiagnosis was based on standard clinical, endoscopic, histologic andradiographic criteria (Sands 2004). HCs were recruited from theAcademic Community of the Health Sciences Faculty, UniversityFernando Pessoa.
Whole blood samples (13.5 ml) were obtained from everyindividual, and peripheral blood mononuclear cell isolation wasperformed as described below.
Cell culture
The blood collected from each individual was diluted 1:2 inphosphate buffered saline (PBS) and placed on Histopaque 1077(Sigma, St. Louis, MO, USA) in 15 ml sterile tubes. Mononuclearcells were separated according to instructions by the manufacturer.The mononuclear cells present in the monolayer were collected,washed 3× in Hank’s Balanced Salt Solution (HBSS, Sigma), and sus-pended in RPMI 1640 with GlutaMAXTM (Invitrogen, Carlsbad, CA,USA) supplemented with antibiotic/antimycotic solution (Invitro-gen), and 10% heat inactivated foetal bovine serum (FBS, Sigma).Cells were counted in a Neubauer chamber. One ml of the cell sus-pension (3 × 106 cells/ml) was seeded in each well of a 24-wellmultidish culture plate. For phagocytosis assessment, cells wereplaced over glass coverslips. Cultures were incubated for 7 days at37 ◦C and 5% CO2, with the medium changed at day 3 after cultureestablishment. At culture day 7, cultures were infected.
In vitro infection of monocyte-derived macrophages
Macrophage cultures were either left uninfected or infectedwith MAP (ATCC 43015, an isolate from a Crohn’s patient), MA(strain 101, an isolate from an AIDS patient), EC (ATCC 25922)or EF (ATCC 29212) at infection ratio of 10 bacteria: 1 cell. Cul-tures were incubated for 3 h to allow phagocytosis of the bacteria.After 3 h, phagocytosis was assessed by Ziehl–Neelsen staining (formycobacteria visualization) or by simple methylene blue staining(for EC and EF visualization) of the coverslips added to culturewells. Bacteria were engulfed by over 35% of cells present in themonolayer, regardless the group of subjects tested, or the bacterial
D patients exhibit defective tumour necrosis factor-� secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
species added (results not shown). Monolayers infected with EC orEF were only assessed at 3 h after infection (T3h) because intra-cellular bacteria are easily eliminated by macrophages but the fewremaining on the extracellular side grow extensively in the culturewells. Cells infected with MAP or MA were assessed at T3h, 3 days
ARTICLE IN PRESSG Model
IMBIO-50678; No. of Pages 10
N. Campos et al. / Immunobiology xxx (2011) xxx–xxx 3
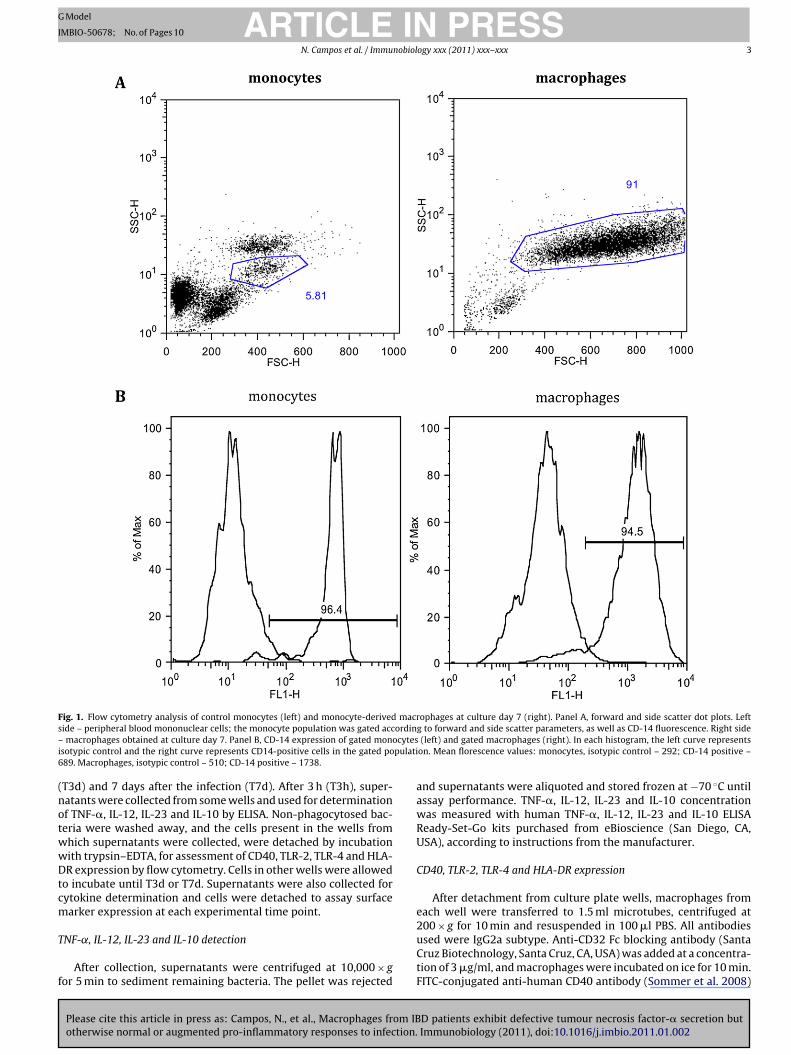
F macrs ordin– ocytesi pulati6
(notwwDtcm
T
f
ig. 1. Flow cytometry analysis of control monocytes (left) and monocyte-derivedide – peripheral blood mononuclear cells; the monocyte population was gated accmacrophages obtained at culture day 7. Panel B, CD-14 expression of gated mon
sotypic control and the right curve represents CD14-positive cells in the gated po89. Macrophages, isotypic control – 510; CD-14 positive – 1738.
T3d) and 7 days after the infection (T7d). After 3 h (T3h), super-atants were collected from some wells and used for determinationf TNF-�, IL-12, IL-23 and IL-10 by ELISA. Non-phagocytosed bac-eria were washed away, and the cells present in the wells fromhich supernatants were collected, were detached by incubationith trypsin–EDTA, for assessment of CD40, TLR-2, TLR-4 and HLA-R expression by flow cytometry. Cells in other wells were allowed
o incubate until T3d or T7d. Supernatants were also collected forytokine determination and cells were detached to assay surfacearker expression at each experimental time point.
Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
NF-˛, IL-12, IL-23 and IL-10 detection
After collection, supernatants were centrifuged at 10,000 × gor 5 min to sediment remaining bacteria. The pellet was rejected
ophages at culture day 7 (right). Panel A, forward and side scatter dot plots. Leftg to forward and side scatter parameters, as well as CD-14 fluorescence. Right side(left) and gated macrophages (right). In each histogram, the left curve represents
on. Mean florescence values: monocytes, isotypic control – 292; CD-14 positive –
and supernatants were aliquoted and stored frozen at −70 ◦C untilassay performance. TNF-�, IL-12, IL-23 and IL-10 concentrationwas measured with human TNF-�, IL-12, IL-23 and IL-10 ELISAReady-Set-Go kits purchased from eBioscience (San Diego, CA,USA), according to instructions from the manufacturer.
CD40, TLR-2, TLR-4 and HLA-DR expression
After detachment from culture plate wells, macrophages fromeach well were transferred to 1.5 ml microtubes, centrifuged at
D patients exhibit defective tumour necrosis factor-� secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
200 × g for 10 min and resuspended in 100 �l PBS. All antibodiesused were IgG2a subtype. Anti-CD32 Fc blocking antibody (SantaCruz Biotechnology, Santa Cruz, CA, USA) was added at a concentra-tion of 3 �g/ml, and macrophages were incubated on ice for 10 min.FITC-conjugated anti-human CD40 antibody (Sommer et al. 2008)
ARTICLE IN PRESSG Model
IMBIO-50678; No. of Pages 10
4 N. Campos et al. / Immunobiology xxx (2011) xxx–xxx
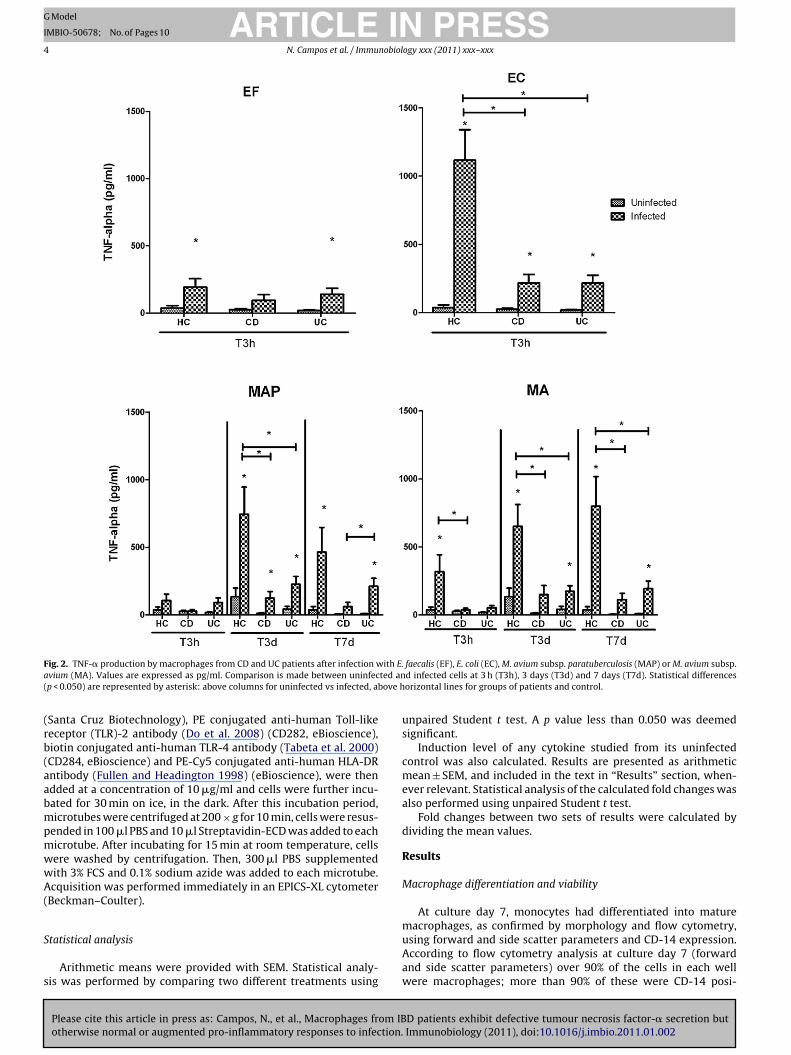
F ith E.a ted an( bove h
(rb(aabmpmwwA(
S
s
ig. 2. TNF-� production by macrophages from CD and UC patients after infection wvium (MA). Values are expressed as pg/ml. Comparison is made between uninfecp < 0.050) are represented by asterisk: above columns for uninfected vs infected, a
Santa Cruz Biotechnology), PE conjugated anti-human Toll-likeeceptor (TLR)-2 antibody (Do et al. 2008) (CD282, eBioscience),iotin conjugated anti-human TLR-4 antibody (Tabeta et al. 2000)CD284, eBioscience) and PE-Cy5 conjugated anti-human HLA-DRntibody (Fullen and Headington 1998) (eBioscience), were thendded at a concentration of 10 �g/ml and cells were further incu-ated for 30 min on ice, in the dark. After this incubation period,icrotubes were centrifuged at 200 × g for 10 min, cells were resus-
ended in 100 �l PBS and 10 �l Streptavidin-ECD was added to eachicrotube. After incubating for 15 min at room temperature, cellsere washed by centrifugation. Then, 300 �l PBS supplementedith 3% FCS and 0.1% sodium azide was added to each microtube.cquisition was performed immediately in an EPICS-XL cytometer
Beckman–Coulter).
Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
tatistical analysis
Arithmetic means were provided with SEM. Statistical analy-is was performed by comparing two different treatments using
faecalis (EF), E. coli (EC), M. avium subsp. paratuberculosis (MAP) or M. avium subsp.d infected cells at 3 h (T3h), 3 days (T3d) and 7 days (T7d). Statistical differencesorizontal lines for groups of patients and control.
unpaired Student t test. A p value less than 0.050 was deemedsignificant.
Induction level of any cytokine studied from its uninfectedcontrol was also calculated. Results are presented as arithmeticmean ± SEM, and included in the text in “Results” section, when-ever relevant. Statistical analysis of the calculated fold changes wasalso performed using unpaired Student t test.
Fold changes between two sets of results were calculated bydividing the mean values.
Results
Macrophage differentiation and viability
At culture day 7, monocytes had differentiated into mature
D patients exhibit defective tumour necrosis factor-� secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
macrophages, as confirmed by morphology and flow cytometry,using forward and side scatter parameters and CD-14 expression.According to flow cytometry analysis at culture day 7 (forwardand side scatter parameters) over 90% of the cells in each wellwere macrophages; more than 90% of these were CD-14 posi-
ARTICLE IN PRESSG Model
IMBIO-50678; No. of Pages 10
N. Campos et al. / Immunobiology xxx (2011) xxx–xxx 5
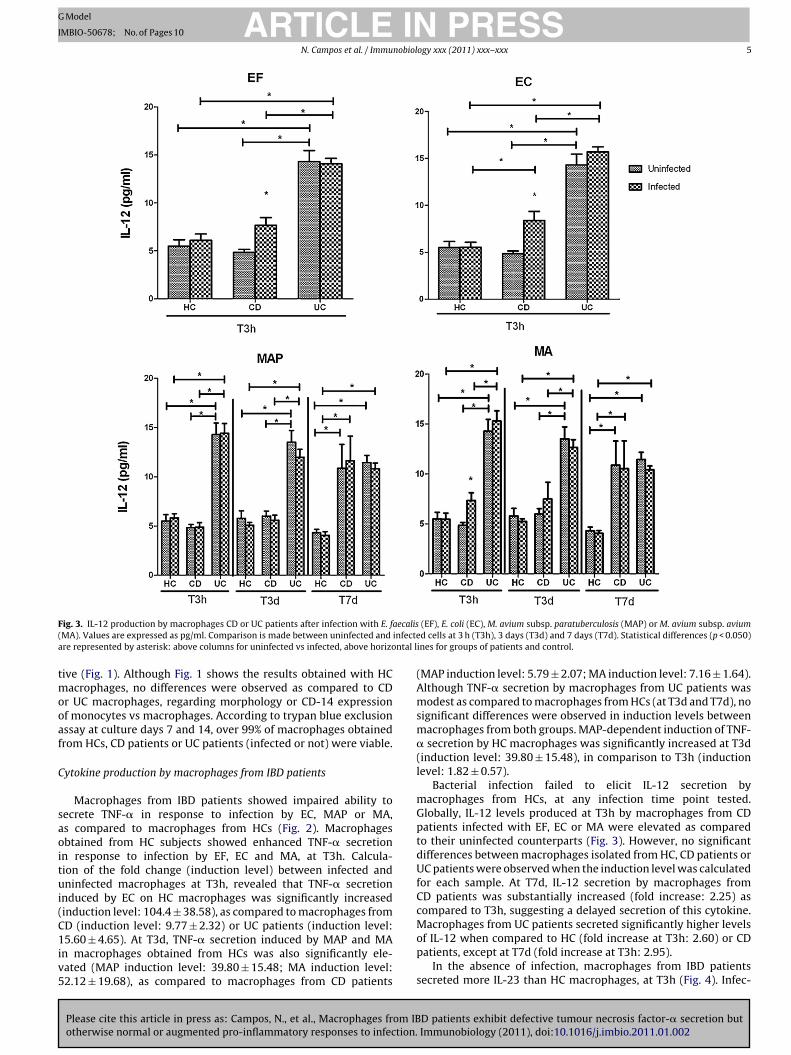
F aecalis( infectea ntal l
tmooaf
C
saoitui(C1iv5
ig. 3. IL-12 production by macrophages CD or UC patients after infection with E. fMA). Values are expressed as pg/ml. Comparison is made between uninfected andre represented by asterisk: above columns for uninfected vs infected, above horizo
ive (Fig. 1). Although Fig. 1 shows the results obtained with HCacrophages, no differences were observed as compared to CD
r UC macrophages, regarding morphology or CD-14 expressionf monocytes vs macrophages. According to trypan blue exclusionssay at culture days 7 and 14, over 99% of macrophages obtainedrom HCs, CD patients or UC patients (infected or not) were viable.
ytokine production by macrophages from IBD patients
Macrophages from IBD patients showed impaired ability toecrete TNF-� in response to infection by EC, MAP or MA,s compared to macrophages from HCs (Fig. 2). Macrophagesbtained from HC subjects showed enhanced TNF-� secretionn response to infection by EF, EC and MA, at T3h. Calcula-ion of the fold change (induction level) between infected andninfected macrophages at T3h, revealed that TNF-� secretion
nduced by EC on HC macrophages was significantly increasedinduction level: 104.4 ± 38.58), as compared to macrophages from
Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
D (induction level: 9.77 ± 2.32) or UC patients (induction level:5.60 ± 4.65). At T3d, TNF-� secretion induced by MAP and MA
n macrophages obtained from HCs was also significantly ele-ated (MAP induction level: 39.80 ± 15.48; MA induction level:2.12 ± 19.68), as compared to macrophages from CD patients
(EF), E. coli (EC), M. avium subsp. paratuberculosis (MAP) or M. avium subsp. aviumd cells at 3 h (T3h), 3 days (T3d) and 7 days (T7d). Statistical differences (p < 0.050)
ines for groups of patients and control.
(MAP induction level: 5.79 ± 2.07; MA induction level: 7.16 ± 1.64).Although TNF-� secretion by macrophages from UC patients wasmodest as compared to macrophages from HCs (at T3d and T7d), nosignificant differences were observed in induction levels betweenmacrophages from both groups. MAP-dependent induction of TNF-� secretion by HC macrophages was significantly increased at T3d(induction level: 39.80 ± 15.48), in comparison to T3h (inductionlevel: 1.82 ± 0.57).
Bacterial infection failed to elicit IL-12 secretion bymacrophages from HCs, at any infection time point tested.Globally, IL-12 levels produced at T3h by macrophages from CDpatients infected with EF, EC or MA were elevated as comparedto their uninfected counterparts (Fig. 3). However, no significantdifferences between macrophages isolated from HC, CD patients orUC patients were observed when the induction level was calculatedfor each sample. At T7d, IL-12 secretion by macrophages fromCD patients was substantially increased (fold increase: 2.25) ascompared to T3h, suggesting a delayed secretion of this cytokine.
D patients exhibit defective tumour necrosis factor-� secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
Macrophages from UC patients secreted significantly higher levelsof IL-12 when compared to HC (fold increase at T3h: 2.60) or CDpatients, except at T7d (fold increase at T3h: 2.95).
In the absence of infection, macrophages from IBD patientssecreted more IL-23 than HC macrophages, at T3h (Fig. 4). Infec-
ARTICLE IN PRESSG Model
IMBIO-50678; No. of Pages 10
6 N. Campos et al. / Immunobiology xxx (2011) xxx–xxx
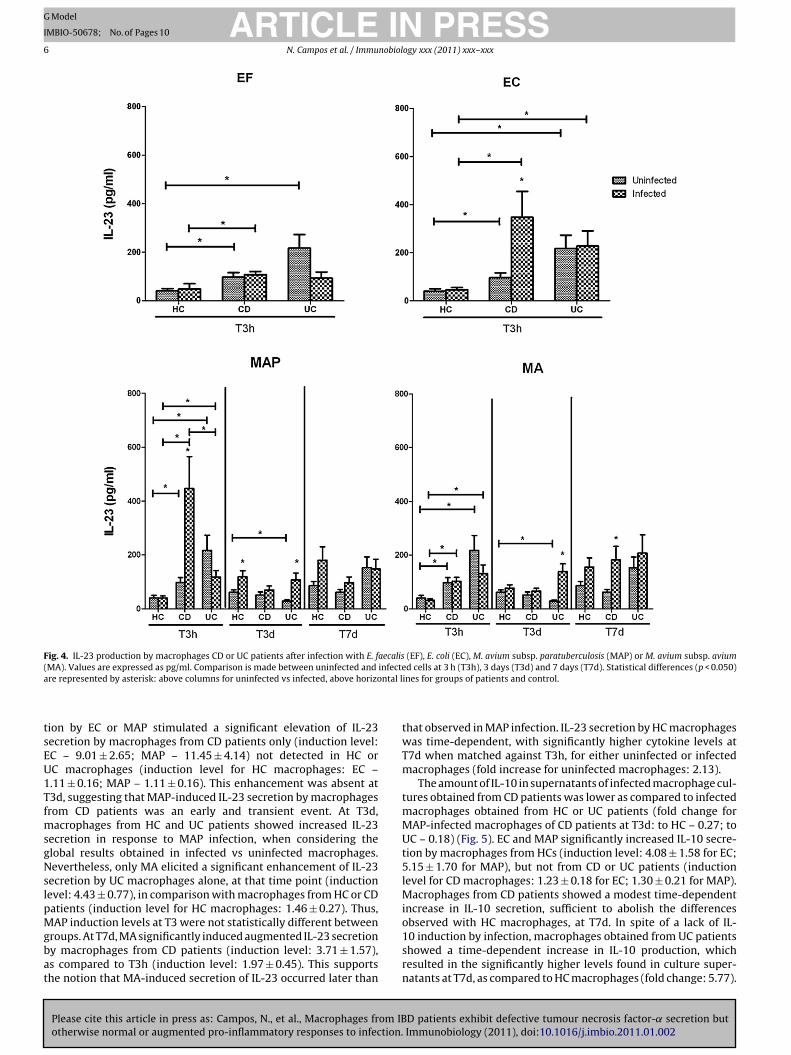
Fig. 4. IL-23 production by macrophages CD or UC patients after infection with E. faecalis (EF), E. coli (EC), M. avium subsp. paratuberculosis (MAP) or M. avium subsp. avium( infectea ntal l
tsEU1TfmsgNslpMgbat
MA). Values are expressed as pg/ml. Comparison is made between uninfected andre represented by asterisk: above columns for uninfected vs infected, above horizo
ion by EC or MAP stimulated a significant elevation of IL-23ecretion by macrophages from CD patients only (induction level:C – 9.01 ± 2.65; MAP – 11.45 ± 4.14) not detected in HC orC macrophages (induction level for HC macrophages: EC –.11 ± 0.16; MAP – 1.11 ± 0.16). This enhancement was absent at3d, suggesting that MAP-induced IL-23 secretion by macrophagesrom CD patients was an early and transient event. At T3d,
acrophages from HC and UC patients showed increased IL-23ecretion in response to MAP infection, when considering thelobal results obtained in infected vs uninfected macrophages.evertheless, only MA elicited a significant enhancement of IL-23
ecretion by UC macrophages alone, at that time point (inductionevel: 4.43 ± 0.77), in comparison with macrophages from HC or CDatients (induction level for HC macrophages: 1.46 ± 0.27). Thus,
Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
AP induction levels at T3 were not statistically different betweenroups. At T7d, MA significantly induced augmented IL-23 secretiony macrophages from CD patients (induction level: 3.71 ± 1.57),s compared to T3h (induction level: 1.97 ± 0.45). This supportshe notion that MA-induced secretion of IL-23 occurred later than
d cells at 3 h (T3h), 3 days (T3d) and 7 days (T7d). Statistical differences (p < 0.050)ines for groups of patients and control.
that observed in MAP infection. IL-23 secretion by HC macrophageswas time-dependent, with significantly higher cytokine levels atT7d when matched against T3h, for either uninfected or infectedmacrophages (fold increase for uninfected macrophages: 2.13).
The amount of IL-10 in supernatants of infected macrophage cul-tures obtained from CD patients was lower as compared to infectedmacrophages obtained from HC or UC patients (fold change forMAP-infected macrophages of CD patients at T3d: to HC – 0.27; toUC – 0.18) (Fig. 5). EC and MAP significantly increased IL-10 secre-tion by macrophages from HCs (induction level: 4.08 ± 1.58 for EC;5.15 ± 1.70 for MAP), but not from CD or UC patients (inductionlevel for CD macrophages: 1.23 ± 0.18 for EC; 1.30 ± 0.21 for MAP).Macrophages from CD patients showed a modest time-dependentincrease in IL-10 secretion, sufficient to abolish the differences
D patients exhibit defective tumour necrosis factor-˛ secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
observed with HC macrophages, at T7d. In spite of a lack of IL-10 induction by infection, macrophages obtained from UC patientsshowed a time-dependent increase in IL-10 production, whichresulted in the significantly higher levels found in culture super-natants at T7d, as compared to HC macrophages (fold change: 5.77).
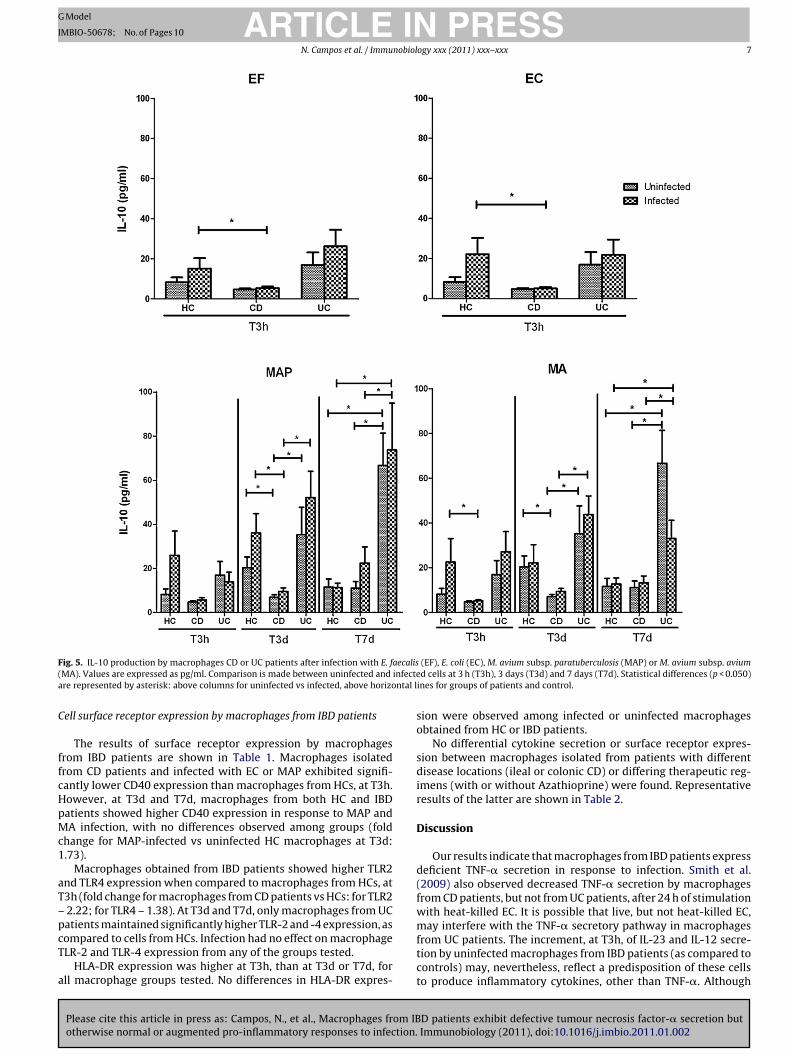
ARTICLE IN PRESSG Model
IMBIO-50678; No. of Pages 10
N. Campos et al. / Immunobiology xxx (2011) xxx–xxx 7
F aecalis( infectea ntal l
C
ffcHpMc1
aT–pcT
a
ig. 5. IL-10 production by macrophages CD or UC patients after infection with E. fMA). Values are expressed as pg/ml. Comparison is made between uninfected andre represented by asterisk: above columns for uninfected vs infected, above horizo
ell surface receptor expression by macrophages from IBD patients
The results of surface receptor expression by macrophagesrom IBD patients are shown in Table 1. Macrophages isolatedrom CD patients and infected with EC or MAP exhibited signifi-antly lower CD40 expression than macrophages from HCs, at T3h.owever, at T3d and T7d, macrophages from both HC and IBDatients showed higher CD40 expression in response to MAP andA infection, with no differences observed among groups (fold
hange for MAP-infected vs uninfected HC macrophages at T3d:.73).
Macrophages obtained from IBD patients showed higher TLR2nd TLR4 expression when compared to macrophages from HCs, at3h (fold change for macrophages from CD patients vs HCs: for TLR22.22; for TLR4 – 1.38). At T3d and T7d, only macrophages from UC
Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
atients maintained significantly higher TLR-2 and -4 expression, asompared to cells from HCs. Infection had no effect on macrophageLR-2 and TLR-4 expression from any of the groups tested.
HLA-DR expression was higher at T3h, than at T3d or T7d, forll macrophage groups tested. No differences in HLA-DR expres-
(EF), E. coli (EC), M. avium subsp. paratuberculosis (MAP) or M. avium subsp. aviumd cells at 3 h (T3h), 3 days (T3d) and 7 days (T7d). Statistical differences (p < 0.050)
ines for groups of patients and control.
sion were observed among infected or uninfected macrophagesobtained from HC or IBD patients.
No differential cytokine secretion or surface receptor expres-sion between macrophages isolated from patients with differentdisease locations (ileal or colonic CD) or differing therapeutic reg-imens (with or without Azathioprine) were found. Representativeresults of the latter are shown in Table 2.
Discussion
Our results indicate that macrophages from IBD patients expressdeficient TNF-� secretion in response to infection. Smith et al.(2009) also observed decreased TNF-� secretion by macrophagesfrom CD patients, but not from UC patients, after 24 h of stimulationwith heat-killed EC. It is possible that live, but not heat-killed EC,
D patients exhibit defective tumour necrosis factor-� secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
may interfere with the TNF-� secretory pathway in macrophagesfrom UC patients. The increment, at T3h, of IL-23 and IL-12 secre-tion by uninfected macrophages from IBD patients (as compared tocontrols) may, nevertheless, reflect a predisposition of these cellsto produce inflammatory cytokines, other than TNF-�. Although

Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
ARTICLE ING Model
IMBIO-50678; No. of Pages 10
8 N. Campos et al. / Immunobiol
Tab
le1
Surf
ace
rece
pto
rex
pre
ssio
nby
mac
rop
hag
esob
tain
edfr
omIB
Dp
atie
nts
,in
resp
onse
toin
fect
ion
.
CD
-40
TLR
-2TL
R-4
HLA
-DR
HC
sC
Dp
atie
nts
UC
pat
ien
tsH
Cs
CD
pat
ien
tsU
Cp
atie
nts
HC
sC
Dp
atie
nts
UC
pat
ien
tsH
Cs
CD
pat
ien
tsU
Cp
atie
nts
T3h U
nin
fect
ed8.
24±
0.72
96.
91±
0.91
46.
28±
1.02
1.95
±0.
262
4.33
±0.
814**
3.73
±0.
677*
1.81
±0.
0956
2.50
±0.
135**
*2.
55±
0.12
3***
226
±21
.421
5±
22.1
223
±24
.3EF
8.39
±0.
995
6.89
±1.
007.
25±
1.66
1.99
±0.
212
4.74
±0.
807**
3.45
4±
0.76
51.
92±
0.10
12.
87±
0.15
1***
2.61
±0.
173**
242
±23
.625
4±
35.4
218
±28
.8EC
8.91
±0.
934
6.09
±0.
870*
5.77
±1.
03*
2.02
±0.
258
4.31
±0.
770**
3.98
±1.
00*
1.87
±0.
112
2.59
±0.
112**
*2.
34±
0.12
7*26
0±
26.4
249
±27
.621
6±
23.9
MA
P8.
33±
0.77
65.
70±
0.81
5*5.
73±
1.11
2.15
±0.
267
4.33
±0.
671**
3.99
±0.
674**
1.91
±0.
0996
2.60
±0.
0785
***
2.68
±0.
132**
*22
8±
24.7
271
±29
.125
3±
24.2
MA
9.30
±0.
865
6.86
±0.
971
5.92
±1.
062.
15±
0.25
24.
97±
0.90
5**3.
634
±0.
720*
1.98
±0.
092
2.79
±0.
134**
*2.
69±
0.08
83**
*21
9±
25.2
235
±40
.821
5±
34.9
T3d U
nin
fect
ed8.
72±
0.87
87.
35±
0.68
77.
50±
1.02
2.77
±0.
239
3.60
±0.
826
5.76
±1.
03**
2.16
±0.
118
2.80
±0.
378
2.96
±0.
295*
92.5
±10
.910
6±
15.5
111
±8.
86M
AP
15.1
±1.
48**
*12
.3±
1.50
*14
.6±
1.52
***
2.46
±0.
246
3.72
±0.
633
6.00
±1.
08**
2.35
±0.
148
2.51
±0.
262
3.07
±0.
314*
98.7
±10
.712
1±
17.9
99.6
±12
.2M
A15
.62
±1.
33**
*15
.0±
1.69
***
15.3
±1.
47**
*3.
02±
0.32
13.
49±
0.59
05.
92±
0.97
5**2.
37±
0.19
12.
74±
0.28
13.
01±
0.29
310
8±
12.8
112
±17
.711
7±
15.3
7T7
d Un
infe
cted
8.42
5±
1.04
10.2
±2.
277.
58±
1.19
1.92
±0.
248
2.02
±0.
446
4.30
±1.
13*
1.91
±0.
101
2.20
±0.
129
2.49
±0.
165**
115
±10
.513
5±
19.3
142
±15
.2M
AP
14.9
±1.
95**
15.6
±2.
0013
.2±
1.78
*2.
12±
0.32
21.
92±
0.26
84.
72±
1.15
*1.
86±
0.14
62.
02±
0.12
12.
17±
0.17
012
9±
13.8
146
±18
.713
6±
11.4
MA
17.0
±1.
75**
*17
.2±
2.13
*16
.2±
1.70
***
1.81
±0.
208
2.42
±0.
545
3.78
±0.
961.
74±
0.12
42.
05±
0.13
71.
83±
0.13
113
1±
11.4
166
±20
.815
2±
18.9
Res
ult
sar
eex
pre
ssed
asm
edia
nfl
uor
esce
nce
inte
nsi
ty.V
alu
esin
bold
ind
icat
esi
gnifi
can
tdif
fere
nce
sas
com
par
edto
un
infe
cted
mac
rop
hag
esfr
omth
esa
me
grou
pof
subj
ects
.Ast
eris
ksin
dic
ate
sign
ifica
ntd
iffe
ren
ces
asco
mp
ared
tom
acro
ph
ages
from
HC
s.*
p<
0.05
0.**
p<
0.01
0.**
*p
<0.
001.
PRESSogy xxx (2011) xxx–xxx
TNF-� is known to play an important role in the pathophysiologyof IBD (confirmed by the effectiveness of treatment with anti-TNF-� antibody), it is possible that in vivo cells, e.g. dendritic cells, maybe activated to secrete TNF-�, as a consequence of the develop-ment of adaptive immunity, overcoming the defect of macrophageTNF-� secretion. Th17 cells are also known to produce TNF-�, andthey play a major role in intestinal inflammation occurring in IBD(Monteleone et al. 2009). Alternatively, TNF-� may be required asan anti-inflammatory cytokine. Using mouse cells, Zakharova andZiegler (2005) have shown that TNF-� inhibits the production ofIL-12 and IL-23 by macrophages and dendritic cells. It is thus con-ceivable that the lowered secretion of TNF-� in IBD patients mayexplain an increased production of cytokines that promote inflam-matory T cell responses.
Interestingly, infection by EC and MAP stimulated elevated IL-23 secretion by macrophages from CD patients alone. It is plausiblethat the EC- and MAP-dependent IL-23 secretion enhancementoccurred as a consequence of phagocytosis, because the high IL-23concentrations obtained at T3h were not observed at T3d and T7d.Recent data suggests that genetic polymorphisms of IL-23R mayconfer protection in CD patients (Tremelling et al. 2007). IL-23 wasfound to be a key cytokine in the development of intestinal inflam-mation in mice (Hue et al. 2006). It has a role in the activation of NKcells as well as in T cell proliferation and stimulation of IFN-� pro-duction (Tan et al. 2009; Monteleone et al. 2009). Induction of Th17cells requires TGF-� and IL-6, but stabilization of the differentiatedTh17 population requires the presence of IL-23. Th17 is producedearly in response to pathogen or antigen stimulation, and it isimportant for the development of early local immune responses.However, increased numbers of Th17 cells were detected in CDand UC patients and are likely to be involved in the pathogene-sis of experimental colitis (Liu et al. 2009; Monteleone et al. 2009;Eastaff-Leung et al. 2009).
Remarkably, a sustained increment in IL-12 secretion by UCmacrophages was detected despite the absence of infection, sug-gesting constitutive activation. This result supports other findings,where mucosal cells of UC patients were found to over-express IL-12 and Stat4 mRNA (Pang et al. 2007). IL-12 is involved in CD4+T cell differentiation into Th1 cells and also in the activation ofNK cells. Induction of IL-12 secretion was observed to result fromligand binding to multiple pattern-recognition receptors (PRR),namely TLRs (Trinchieri and Sher 2007). In fact, we found increasedTLR-2 and TLR-4 expression in UC macrophages. However, expres-sion of these receptors was also augmented in CD macrophagesat T3h, without affecting IL-12 production. It is plausible thatother signals are also required. On the other hand, consider-ing that TLR expression was increased in macrophages from IBDpatients, TNF-� deficiency did not seem to result from lack of thesemolecules.
As noted in our model, HLA-DR expression was unaffected byinfection, but CD40 expression, significant for the activation ofimmunity and inflammation (Danese et al. 2004, 2006) was stim-ulated in response to either MAP or MA. These findings confirmthat both mycobacteria share the ability to elicit an inflamma-tory response. Nevertheless, no differences were observed betweengroups, both for HLA-DR and CD40 expression.
IL-10 secretion by macrophages from CD patients was dimin-ished as compared to HC or UC macrophages, both at T3h andT3d. It was previously observed that CD patients had an impairedability to produce IL-10, and that this impairment might influencethe exacerbation of inflammation (Correa et al. 2009; Marrakchi
D patients exhibit defective tumour necrosis factor-� secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
et al. 2009). Our results fail to support those reported by Smithet al. (2009) who found no significant differences in IL-10 secretionamong macrophages from controls, UC patients or CD patients after24 h of stimulation with heat killed EC. Given that IL-10 has beenimplicated in the control of host inflammatory responses and is pri-

ARTICLE IN PRESSG Model
IMBIO-50678; No. of Pages 10
N. Campos et al. / Immunobiology xxx (2011) xxx–xxx 9
Table 2Cytokine induction fold (at T3h) of macrophages from CD patients undergoing 5-ASA treatment alone or 5-ASA + Azathioprine.
HCs CD patients
5-ASA 5-ASA + Azathioprine p-Value*
TNF-�EC 104.4 ± 38.58 12.26 ± 1.07 7.10 ± 2.01 0.15MAP 1.78 ± 0.53 2.31 ± 0.17 1.25 ± 0.37 0.094MA 29.20 ± 14.90 1.37 ± 0.24 1.59 ± 0.28 0.31
IL-23EC 1.11 ± 0.16 6.44 ± 4.46 9.61 ± 3.14 0.32MAP 1.11 ± 0.16 14.71 ± 8.06 10.97 ± 5.14 0.36MA 0.94 ± 0.18 2.01 ± 0.59 1.85 ± 0.44 0.42
IL-10EC 4.08 ± 1.58 1.20 ± 0.38 1.25 ± 0.22 0.45MAP 5.15 ± 1.70 1.41 ± 0.32 1.26 ± 0.27 0.38
0.17
Eunpair
mri
ieimbatECisswmstii
ciulref
tossatiia
A
(Pa
MA 3.28 ± 1.39 1.72 ±C – E. coli; MAP – M. avium subsp. paratuberculosis; MA – M. avium subsp. avium.
* For CD patients, p values between therapeutic regimens were calculated using
arily produced by monocytes/macrophages, we emphasize theole of defective macrophage IL-10 production in the uncontrollednflammatory status observed in CD patients.
Herein, our results suggest different inflammatory mechanismsnvolved in IBD: (i) macrophages from CD patients first producedlevated IL-23 in response to MAP and EC, and later showedncreased IL-12 but low IL-10 production, (ii) in contrast, UC
acrophages secreted higher IL-12 levels at all time points testedut showed a small time-dependent decrease that correlated withn increase in IL-10 secretion, in spite of lack of inducibility by bac-erial infection at T3h. These results suggest that both MAP andC infection may contribute to the maintenance of Th17 cells byD macrophages through stimulation of IL-23 production, result-
ng in inflammatory tissue damage, uncontrolled by lack of IL-10ecretion. The lack of IL-23 inducibility by UC macrophages, inpite of the observed elevated IL-12 secretion, is in agreementith studies implicating IL23/IL17 in CD (Hölttä et al. 2008). UCacrophages may have decreased ability to contribute for the
tabilization of the Th17 population, but instead may lead tohe generation of Th1 cells through IL-12 secretion. The increasen IL-10 secretion with time may play a part in the control ofnflammation.
Although it was evident that different bacteria elicited diverseytokine responses in CD macrophages, specific effects of MAPnfection alone were not observed. In fact, both EC and MAP stim-lated IL-23 secretion, and both MAP and MA induced similar
evels of TNF-�, although reduced, as compared to control cells. Aseferred above, both MAP and MA also induced comparable CD40xpression. Taken together, one may hypothesise that bacterial sur-ace lipids may be implicated in the activation of CD macrophages.
In conclusion, MAP infection of macrophages did not seemo have any specific effects discriminating it from either ECr MA. Macrophages from CD patients showed impaired TNF-�ecretion in response to bacterial infection, but elevated IL-23ecretion (thereby suggesting maintenance of Th17 cells in vivo)nd low IL-10 secretion, which may contribute to inefficient con-rol of inflammation. On the other hand, UC macrophages showedncreased IL-12 secretion (more suggestive of Th1 differentiationn vivo) and time-dependent increased IL-10 production, therebyllowing the control of inflammation.
cknowledgments
Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
This work was funded by the Portuguese Science FoundationFCT) and COMPETE Program subsidized by FEDER, through theIC/IC/82802/2007 grant. The authors wish to thank all patientsnd control subjects for their participation this study.
1.45 ± 0.39 0.34
ed Student t test.
References
Abubakar, I., Myhill, D., Aliyu, S.H., Hunter, P.R., 2008. Detection of Mycobacteriumavium subspecies paratuberculosis from patients with Crohn’s disease usingnucleic acid-based techniques: a systematic review and meta-analysis. Inflamm.Bowel Dis. 14, 401–410.
Anderson, C.A., Massey, D.C.O., Barrett, J.C., Prescott, N.J., Tremelling, M., Fisher,S.A., Gwilliam, R., Jacob, J., Nimmo, E.R., Drummond, H., Lees, C.W., Onnie, C.M.,Hanson, C., Blaszczyk, K., Ravindrarajah, R., Hunt, S., Varma, D., Hammond, N.,Lewis, G., Attlesey, H., Watkins, N., Ouwehand, W., Strachan, D., Mcardle, W.,Lewis, C.M., Consortium, W.T.C.C., Lobo, A., Sanderson, J., Jewell, D.P., Deloukas,P., Mansfield, J.C., Mathew, C.G., Satsangi, J., Parkes, M., 2009. Investigation ofCrohn’s disease risk loci in ulcerative colitis further defines their molecularrelationship. Gastroenterology 136, 523–529, e3.
Battaglia, E., Biancone, L., Resegotti, A., Emanuelli, G., Fronda, G.R., Camussi, G., 1999.Expression of CD40 and its ligand, CD40L, in intestinal lesions of Crohn’s disease.Am. J. Gastroenterol. 94, 3279–3284.
Browning, B.L., Huebner, C., Petermann, I., Gearry, R.B., Barclay, M.L., Shelling,A.N., Ferguson, L.R., 2007. Has toll-like receptor 4 been prematurely dismissedas an inflammatory bowel disease gene? Association study combined withmeta-analysis shows strong evidence for association. Am. J. Gastroenterol. 102,2504–2512.
Clancy, R., Ren, Z., Turton, J., Pang, G., Wettstein, A., 2007. Molecular evidence forMycobacterium avium subspecies paratuberculosis (MAP) in Crohn’s disease cor-relates with enhanced TNF-alpha secretion. Dig. Liver Dis. 39, 445–451.
Correa, I., Veny, M., Esteller, M., Pique, J.M., Yague, J., Panes, J., Salas, A., 2009. Defec-tive IL-10 production in severe phenotypes of Crohn’s disease. J. Leukocyte Biol.85, 896–903.
Danese, S., Sans, M., Fiocchi, C., 2004. The CD40/CD40L costimulatory pathway ininflammatory bowel disease. Gut 53, 1035–1043.
Danese, S., Sans, M., Scaldaferri, F., Sgambato, A., Rutella, S., Cittadini, A., Pique, J.M.,Panes, J., Katz, J.A., Gasbarrini, A., Fiocchi, C., 2006. TNF-alpha blockade down-regulates the CD40/CD40L pathway in the mucosal microcirculation: a novelanti-inflammatory mechanism of infliximab in Crohn’s disease. J. Immunol. 176,2617–2624.
De Jager, P.L., Franchimont, D., Waliszewska, A., Bitton, A., Cohen, A., Langelier, D.,Belaiche, J., Vermeire, S., Farwell, L., Goris, A., Libioulle, C., Jani, N., Dassopoulos,T., Bromfield, G.P., Dubois, B., Cho, J.H., Brant, S.R., Duerr, R.H., Yang, H., Rotter,J.I., Silverberg, M.S., Steinhart, A.H., Daly, M.J., Podolsky, D.K., Louis, E., Hafler,D.A., Rioux, J.D., Consortium, Q.I.G., Consortium, N.I.G., 2007. The role of theToll receptor pathway in susceptibility to inflammatory bowel diseases. GenesImmun. 8, 387–397.
Do, J.E., Kwon, S.Y., Park, S., Lee, E.-S., 2008. Effects of vitamin D on expression of Toll-like receptors of monocytes from patients with Behcet’s disease. Rheumatology(Oxford) 47, 840–848.
Eastaff-Leung, N., Mabarrack, N., Barbour, A., Cummins, A., Barry, S., 2009. Foxp3(+)regulatory T cells, Th17 effector cells, and cytokine environment in inflammatorybowel disease. J. Clin. Immunol. 30, 80–89.
Ebert, E.C., Panja, A., Das, K.M., Praveen, R., Geng, X., Rezac, C., Bajpai, M., 2009.Patients with inflammatory bowel disease may have a transforming growthfactor-beta-, interleukin (IL)-2- or IL-10-deficient state induced by intrinsic neu-tralizing antibodies. Clin. Exp. Immunol. 155, 65–71.
Feller, M., Huwiler, K., Stephan, R., Altpeter, E., Shang, A., Furrer, H., Pfyffer, G., Jemmi,T., Baumgartner, A., Egger, M., 2007. Mycobacterium avium subspecies paratu-berculosis and Crohn’s disease: a systematic review and meta-analysis. LancetInfect. Dis. 7, 607–613.
D patients exhibit defective tumour necrosis factor-� secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
Ferwerda, G., Kullberg, B.J., De Jong, D.J., Girardin, S.E., Langenberg, D.M., Van Crevel,R., Ottenhoff, T.H., Van Der Meer, J.W., Netea, M.G., 2007. Mycobacterium paratu-berculosis is recognized by Toll-like receptors and NOD2. J. Leukocyte Biol. 82,1011–1018.
Frolova, L., Drastich, P., Rossmann, P., Klimesova, K., Tlaskalova-Hogenova, H., 2008.Expression of Toll-like receptor 2 (TLR2), TLR4, and CD14 in biopsy samples of

ING
I
1 nobiol
F
G
H
H
J
K
L
L
M
M
M
M
M
M
P
Annu. Rev. Genomics Hum. Genet. 10, 89–116.
ARTICLEModel
MBIO-50678; No. of Pages 10
0 N. Campos et al. / Immu
patients with inflammatory bowel diseases: upregulated expression of TLR2 interminal ileum of patients with ulcerative colitis. J. Histochem. Cytochem. 56,267–274.
ullen, D.R., Headington, J.T., 1998. Factor XIIIa-positive dermal dendritic cells andHLA-DR expression in radial versus vertical growth-phase melanomas. J. Cutan.Pathol. 25, 553–558.
lawischnig, W., Steineck, T., Spergser, J., 2006. Infections caused by Mycobacteriumavium subspecies avium, hominissuis, and paratuberculosis in free-rangingred deer (Cervus elaphus hippelaphus) in Austria, 2001–2004. J. Wildl. Dis. 42,724–731.
ölttä, V., Klemmetti, P., Sipponen, T., Westerholm-Ormio, M., Kociubinski, G., Salo,H., Räsänen, L., Kolho, K.-L., Färkkilä, M., Savilahti, E., Vaarala, O., 2008. IL-23/IL-17 immunity as a hallmark of Crohn’s disease. Inflamm Bowel Dis. 14,1175–1184.
ue, S., Ahern, P., Buoncore, S., Kullberg, M.C., Cua, D.J., Mckenzie, B.S., Powrie, F.,Maloy, K.J., 2006. Interleukin-23 drives innate and T cell-mediated intestinalinflammation. J Exp Med. 203, 2473–2483.
udge, J., Kyriazakis, I., Greig, A., Allcroft, D.J., Hutchings, M.R., 2005. Clustering ofMycobacterium avium subsp. paratuberculosis in rabbits and the environment:how hot is a hot spot? Appl. Environ. Microbiol. 71, 6033–6038.
uhn, R., Lohler, J., Rennick, D., Rajewsky, K., Muller, W., 1993. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 75, 263–274.
i, C.-S., Zhang, Q., Lee, K.-J., Cho, S.-W., Lee, K.-M., Hahm, K.-B., Choi, S.-C., Yun,K.-J., Chung, H.-T., Chae, S.-C., 2009. Interleukin-27 polymorphisms are associ-ated with inflammatory bowel diseases in a Korean population. J. Gastroenterol.Hepatol. 24, 1692–1696.
iu, Z.-J., Yadav, P.-K., Su, J.-L., Wang, J.-S., Fei, K., 2009. Potential role of Th17 cellsin the pathogenesis of inflammatory bowel disease. World J. Gastroenterol. 15,5784–5788.
arks, D.J.B., Harbord, M.W.N., Macallister, R., Rahman, F.Z., Young, J., Al-Lazikani, B.,Lees, W., Novelli, M., Bloom, S., Segal, A.W., 2006. Defective acute inflammationin Crohn’s disease: a clinical investigation. Lancet 367, 668–678.
arrakchi, R., Moussa, A., Ouerhani, S., Bougatef, K., Bouhaha, R., Messai, Y., Rouissi,K., Khadimallah, I., Khodjet-El-Khil, H., Najar, T., Benammar-Elgaaeid, A., 2009.Interleukin 10 promoter region polymorphisms in inflammatory bowel diseasein Tunisian population. Inflamm. Res. 58, 155–160.
cclure, H.M., Chiodini, R.J., Anderson, D.C., Swenson, R.B., Thayer, W.R., Coutu,J.A., 1987. Mycobacterium paratuberculosis infection in a colony of stumptailmacaques (Macaca arctoides). J. Infect. Dis. 155, 1011–1019.
onteleone, I., Pallone, F., Monteleone, G., 2009. Interleukin-23 and Th17 cells inthe control of gut inflammation. Mediat. Inflamm. (Article ID 297645).
urai, M., Turovskaya, O., Kim, G., Madan, R., Karp, C.L., Cheroutre, H., Kronenberg,M., 2009. Interleukin 10 acts on regulatory T cells to maintain expression of thetranscription factor Foxp3 and suppressive function in mice with colitis. Nat.Immunol. 10, 1178–1184.
Please cite this article in press as: Campos, N., et al., Macrophages from IBotherwise normal or augmented pro-inflammatory responses to infection.
uro, Lopez, Campilo, Salama, Martinez, Lucas, Valdes, Botella, Minguela, Miras, Car-ballo, Alvarez-Lopez, 2009. The role of human leukocyte antigen (HLA) complexin IBD: Crohn’s disease and ulcerative colitis. Adv. Stud. Biol. 1, 37–41.
ang, Y.H., Zheng, C.Q., Yang, X.Z., Zhang, W.J., 2007. Increased expression and activa-tion of IL-12-induced Stat4 signaling in the mucosa of ulcerative colitis patients.Cell. Immunol. 248, 115–120.
PRESSogy xxx (2011) xxx–xxx
Peluso, I., Pallone, F., Monteleone, G., 2006. Interleukin-12 and Th1 immune responsein Crohn’s disease: pathogenetic relevance and therapeutic implication. WorldJ. Gastroenterol. 12, 5606–5610.
Pierik, M., Joossens, S., Van Steen, K., Van Schuerbeek, N., Vlietinck, R., Rutgeerts,P., Vermeire, S., 2006. Toll-like receptor-1, -2, and -6 polymorphisms influencedisease extension in inflammatory bowel diseases. Inflamm. Bowel Dis. 12, 1–8.
Polese, L., Angriman, I., Cecchetto, A., Norberto, L., Scarpa, M., Ruffolo, C., Barollo,M., Sommariva, A., D’amico, D.F., 2002. The role of CD40 in ulcerative colitis:histochemical analysis and clinical correlation. Eur. J. Gastroenterol. Hepatol.14, 237–241.
Sands, B.E., 2004. From symptom to diagnosis: clinical distinctions among variousforms of intestinal inflammation. Gastroenterology 126, 1518–1532.
Selby, W., Pavli, P., Crotty, B., Florin, T., Radford-Smith, G., Gibson, P., Mitchell, B.,Connell, W., Read, R., Merrett, M., Ee, H., Hetzel, D., 2007. Two-year combinationantibiotic therapy with clarithromycin, rifabutin, and clofazimine for Crohn’sdisease. Gastroenterology 132, 2313–2319.
Smith, A.M., Rahman, F.Z., Hayee, B.H., Graham, S.J., Marks, D.J.B., Sewell, G.W.,Palmer, C.D., Wilde, J., Foxwell, B.M.J., Gloger, I.S., Sweeting, T., Marsh, M., Walker,A.P., Bloom, S.L., Segal, A.W., 2009. Disordered macrophage cytokine secretionunderlies impaired acute inflammation and bacterial clearance in Crohn’s dis-ease. J. Exp. Med. 206, 1883–1897.
Sommer, S., Pudrith, C., Colvin, C., Coussens, P., 2008. Mycobacterium avium sub-species paratuberculosis suppresses expression of IL-12p40 and iNOS genesinduced by signalling through CD40 in bovine monocyte-derived macrophages.Vet. Immunol. Immunopathol. 128, 44–52.
Strober, W., Fuss, I., Mannon, P., 2007. The fundamental basis of inflammatory boweldisease. J. Clin. Invest. 117, 514–521.
Tabeta, K., Yamazaki, K., Akashi, S., Miyake, K., Kumada, H., Umemoto, T., Yoshie,H., 2000. Toll-like receptors confer responsiveness to lipopolysaccharide fromPorphyromonas gingivalis in human gingival fibroblasts. Infect. Immun. 68,3731–3735.
Tan, Z.Y., Bealgey, K.W., Fang, Y., Gong, Y.M., Bao, S., 2009. Interleukin-23: immuno-logical roles and clinical implications. Int. J. Biochem. Cell Biol. 41, 733–735.
Tremelling, M., Cummings, F., Fisher, S.A., Mansfield, J., Gwilliam, R., Keniry, A.,Nimmo, E.R., Drummond, H., Onnie, C.M., Prescott, N.J., Sanderson, J., Bredin, F.,Berzuini, C., Forbes, A., Lewis, C.M., Cardon, L., Deloukas, P., Jewell, D., Mathew,C.G., Parkes, M., Satsangi, J., 2007. IL23R variation determines susceptibility butnot disease phenotype in inflammatory bowel disease. Gastroenterology 132,1657–1664.
Trinchieri, G., Sher, A., 2007. Cooperation of Toll-like receptor signals in innateimmune defence. Nat. Rev. Immunol. 7, 179–190.
Van Deventer, S.J., 1997. Tumour necrosis factor and Crohn’s disease. Gut 40,443–448.
Van Limbergen, J., Wilson, D.C., Satsangi, J., 2009. The genetics of Crohn’s disease.
D patients exhibit defective tumour necrosis factor-� secretion butImmunobiology (2011), doi:10.1016/j.imbio.2011.01.002
Xavier, R.J., Podolsky, D.K., 2007. Unravelling the pathogenesis of inflammatorybowel disease. Nature 448, 427–434.
Zakharova, M., Ziegler, H.K., 2005. Paradoxical anti-inflammatory actions of TNF-alpha: inhibition of IL-12 and IL-23 via TNF receptor 1 in macrophages anddendritic cells. J. Immunol. 175, 5024–5033.
Copyright © 2022 FDOKUMEN




















