
Loss of High-Energy Phosphate following Hyperthermia Demonstrated by in Vivo 31P-Nuclear Magnetic...
7
[CANCER RESEARCH 44, 633-638, February 1984] Loss of High-Energy Phosphate following Hyperthermia Demonstrated by in Vivo 31P-Nuclear Magnetic Resonance Spectroscopy1 Michael B. Lilly,2 Thian C. Ng, William T. Evanochko, Charles R. Katholi, Narinder G. Kumar, Gabriel A. Elgavish, John R. Durant, Raymond Hiramoto, Vithal Ghanta, and Jerry D. Glickson Departments of Medicine [M. B. L, J. R. D.], Biochemistry [T. C. N., W. T.E., N. G. K., G. A. £.,J. D. G.¡,Microbiology [R. H., V. G.], and Biomathematics [C. R. K.J, and the Comprehensive Cancer Center, University of Alabama in Birmingham, Birmingham, Alabama 35294 ABSTRACT We have used in vivo 31P-nuclear magnetic resonance spec- troscopy to study the changes in high-energy phosphates follow ing hyperthermia. Immediately after heating, there is a fall in adenosine triphosphate and apparent intracellular pH and an increase in inorganic phosphate. Following sublethal heating (40° for 15 min), these changes were partial, and they resolved over the subsequent 45 hr. With tumors given severe hyperthermia (47°for 15 min), there was complete disappearance of adenosine triphosphate, with no recovery by 24 hr posttreatment. Qualita tively similar effects were seen after heating of normal leg muscle. The degree of fall of the adenosine triphosphate/inorganic phos phate concentration ratio was directly proportional to the heat dose and to thermal cell kill. 31P-Nuclear magnetic resonance spectroscopy may be useful in thermal dosimetry and treatment evaluation following hyperthermia. INTRODUCTION Hyperthermia is a potential antitumor modality which is cur rently the subject of extensive investigation. Following exposure to heat, virtually every phase of metabolism becomes disrupted. No clear concensus exists, however, as to the primary metabolic events associated with thermal cell kill. Studies have demon strated that respiration, anaerobic glycolysis, and DNA, RNA, and protein synthesis are inhibited at temperatures above 42°. These studies have been summarized in recent reviews (5, 11). The majority of the available data are derived from in vitro systems. Cellular metabolism depends greatly on environmental factors. Because of the variable metabolic states of cells within a solid tumor, as well as modifying host factors, it is sometimes difficult to extrapolate from in vitro data to the in vivo situation. Thus, the ability to study the metabolism of tumors in situ could help clarify which events are most important in thermal cell kill. NMR3 is a noninvasive spectroscopic technique that can be used to study the metabolism of tissues both in vitro and in vivo. Isotopes that have been detected in living cells include 1H (6), 13C(27), 15N(21), 19F(13), 23Na(10), and 31P(8). 31Pis the most extensively studied isotope in biological samples. It offers the advantages of moderately high sensitivity (detectable at concen trations >1 ITIM), 100% natural abundance, relative spectral simplicity, and presence in a number of detectable key respiratory intermediates. The apparent intracellular pH can also be deter- 1Supported in part by Grants CA 13148 and CA 16699 from NIH. 2 To whom requests for reprints should be addressed, at Comprehensive Cancer Center, University of Alabama in Birmingham, Birmingham, Ala. 35294. 'The abbreviations used are: NMR, nuctear magnetic resonance; PME, phos- phomonoesters; PCr, phosphocreatinine; PCA, perchloric acid. Received September 8, 1982; accepted September 28,1983. mined from the chemical shift of the P¡peak in the 31P-NMR spectra (24). The development of surface coils (1) and field focusing tech niques (18) has led to the noninvasive measurement of NMR spectra of specific tissue and organs. These methods have been used to monitor the in vivo 31P-NMR spectra of muscle and brain (1), kidney (2), and liver (18) in experimental animals, as well as muscle in humans (26). We have recently used a surface coil probe to obtain in vivo 31P-NMR spectra from s.c. murine tumors and human tumors in nude mice and to monitor the response of some of these tumors to radiation and chemotherapy (15, 23). In this paper, we report our use of 31P-NMR spectroscopy to record the changes in high- energy phosphates following hyperthermic treatment of a murine osteosarcoma and murine leg muscle and relate these spectral changes to heat dose and to an estimate of cell kill. MATERIALS AND METHODS Murine Tumor System. Female C3H mice (8 to 12 weeks old) were given s.c. inoculations of s.c. 1 to 3 x 106 trypan blue-negative Dunn osteosarcoma cells. The single-cell suspension was prepared from intact tumors by a Pronase digestion technique (17). Tumor damage from hyperthermia was quantitated by measuring the change in serum alkaline phosphatase, a tumor-associated marker, by a colorimetrie procedure (17). This marker has previously been related quantitatively to the total number of viable tumor cells in the mouse (20). We have seen, after hyperthermia, a reproducible fall in serum alkaline phosphatase that is proportional to the heat dose and that parallels gross tumor regression (data not shown). Alkaline phosphatase measurements were made on Days -1 and 6 after treatment. Mice were anesthetized with Nembutal for all treatments and spectroscopic measurements. The skin overlying the tumor was shaved before spectroscopy. Hyperthermia Treatment. Three weeks after inoculation, the tumors measured 1.5 to 2.0 cm in diameter and contained 1to5x 10B cells. Hyperthermia was then administered via a capacitive radiofrequency device of our own design. A signal generator (Model 3002; Wavetek, Inc., San Diego, Calif.) produced a 1.5-MHz signal, which was then fed to an amplifier (Model 31OL; ENI, Inc., Rochester, N.Y.). The power was transmitted to the mouse via a coaxial cable, which terminated in 2 brass electrodes. These electrodes bracketed the tumor and were coupled to it by standard electrode jelly. Temperature measurements were made with an electronic thermometer (BAT-8; Bailey Instruments, Saddle Brook, N. J.) and Teflon-sheathed copper-constantan thermocouples (Bailey IT-21 ). The thermocouples were inserted along a 25-gauge needle track along the longest axis of the tumor. A small amount of radiofre quency power (0.5 watt) was applied, and the thermocouple was moved back and forth until it was at the point of maximum temperature. Power was then increased until the thermocouple registered the target temper ature. Treatment duration was timed from the moment target tempera ture was reached. Power was adjusted manually to keep the tumor within ±0.1°of target temperature. Similar procedures were used for heating of normal murine thigh muscle. FEBRUARY 1984 633 on April 5, 2015. © 1984 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Loss of High-Energy Phosphate following Hyperthermia Demonstrated by in Vivo 31P-Nuclear Magnetic...

[CANCER RESEARCH 44, 633-638, February 1984]
Loss of High-Energy Phosphate following Hyperthermia Demonstrated byin Vivo 31P-Nuclear Magnetic Resonance Spectroscopy1
Michael B. Lilly,2 Thian C. Ng, William T. Evanochko, Charles R. Katholi, Narinder G. Kumar, Gabriel A. Elgavish,
John R. Durant, Raymond Hiramoto, Vithal Ghanta, and Jerry D. Glickson
Departments of Medicine [M. B. L, J. R. D.], Biochemistry [T. C. N., W. T. E., N. G. K., G. A. £.,J. D. G.¡,Microbiology [R. H., V. G.], and Biomathematics [C. R. K.J,and the Comprehensive Cancer Center, University of Alabama in Birmingham, Birmingham, Alabama 35294
ABSTRACT
We have used in vivo 31P-nuclear magnetic resonance spec-
troscopy to study the changes in high-energy phosphates follow
ing hyperthermia. Immediately after heating, there is a fall inadenosine triphosphate and apparent intracellular pH and anincrease in inorganic phosphate. Following sublethal heating (40°
for 15 min), these changes were partial, and they resolved overthe subsequent 45 hr. With tumors given severe hyperthermia(47°for 15 min), there was complete disappearance of adenosine
triphosphate, with no recovery by 24 hr posttreatment. Qualitatively similar effects were seen after heating of normal leg muscle.The degree of fall of the adenosine triphosphate/inorganic phosphate concentration ratio was directly proportional to the heatdose and to thermal cell kill. 31P-Nuclear magnetic resonance
spectroscopy may be useful in thermal dosimetry and treatmentevaluation following hyperthermia.
INTRODUCTION
Hyperthermia is a potential antitumor modality which is currently the subject of extensive investigation. Following exposureto heat, virtually every phase of metabolism becomes disrupted.No clear concensus exists, however, as to the primary metabolicevents associated with thermal cell kill. Studies have demonstrated that respiration, anaerobic glycolysis, and DNA, RNA,and protein synthesis are inhibited at temperatures above 42°.
These studies have been summarized in recent reviews (5, 11).The majority of the available data are derived from in vitrosystems. Cellular metabolism depends greatly on environmentalfactors. Because of the variable metabolic states of cells withina solid tumor, as well as modifying host factors, it is sometimesdifficult to extrapolate from in vitro data to the in vivo situation.Thus, the ability to study the metabolism of tumors in situ couldhelp clarify which events are most important in thermal cell kill.
NMR3 is a noninvasive spectroscopic technique that can be
used to study the metabolism of tissues both in vitro and in vivo.Isotopes that have been detected in living cells include 1H (6),13C(27), 15N(21), 19F(13), 23Na(10), and 31P(8). 31Pis the most
extensively studied isotope in biological samples. It offers theadvantages of moderately high sensitivity (detectable at concentrations >1 ITIM), 100% natural abundance, relative spectralsimplicity, and presence in a number of detectable key respiratoryintermediates. The apparent intracellular pH can also be deter-
1Supported in part by Grants CA 13148 and CA 16699 from NIH.2To whom requests for reprints should be addressed, at Comprehensive Cancer
Center, University of Alabama in Birmingham, Birmingham, Ala. 35294.'The abbreviations used are: NMR, nuctear magnetic resonance; PME, phos-
phomonoesters; PCr, phosphocreatinine; PCA, perchloric acid.Received September 8, 1982; accepted September 28,1983.
mined from the chemical shift of the P¡peak in the 31P-NMR
spectra (24).The development of surface coils (1) and field focusing tech
niques (18) has led to the noninvasive measurement of NMRspectra of specific tissue and organs. These methods have beenused to monitor the in vivo 31P-NMR spectra of muscle and brain
(1), kidney (2), and liver (18) in experimental animals, as well asmuscle in humans (26).
We have recently used a surface coil probe to obtain in vivo31P-NMR spectra from s.c. murine tumors and human tumors in
nude mice and to monitor the response of some of these tumorsto radiation and chemotherapy (15, 23). In this paper, we reportour use of 31P-NMR spectroscopy to record the changes in high-
energy phosphates following hyperthermic treatment of a murineosteosarcoma and murine leg muscle and relate these spectralchanges to heat dose and to an estimate of cell kill.
MATERIALS AND METHODS
Murine Tumor System. Female C3H mice (8 to 12 weeks old) weregiven s.c. inoculations of s.c. 1 to 3 x 106 trypan blue-negative Dunn
osteosarcoma cells. The single-cell suspension was prepared from intact
tumors by a Pronase digestion technique (17). Tumor damage fromhyperthermia was quantitated by measuring the change in serum alkalinephosphatase, a tumor-associated marker, by a colorimetrie procedure
(17). This marker has previously been related quantitatively to the totalnumber of viable tumor cells in the mouse (20). We have seen, afterhyperthermia, a reproducible fall in serum alkaline phosphatase that isproportional to the heat dose and that parallels gross tumor regression(data not shown). Alkaline phosphatase measurements were made onDays -1 and 6 after treatment. Mice were anesthetized with Nembutal
for all treatments and spectroscopic measurements. The skin overlyingthe tumor was shaved before spectroscopy.
Hyperthermia Treatment. Three weeks after inoculation, the tumorsmeasured 1.5 to 2.0 cm in diameter and contained 1 to 5 x 10B cells.
Hyperthermia was then administered via a capacitive radiofrequencydevice of our own design. A signal generator (Model 3002; Wavetek,Inc., San Diego, Calif.) produced a 1.5-MHz signal, which was then fed
to an amplifier (Model 31OL; ENI, Inc., Rochester, N.Y.). The power wastransmitted to the mouse via a coaxial cable, which terminated in 2 brasselectrodes. These electrodes bracketed the tumor and were coupled toit by standard electrode jelly. Temperature measurements were madewith an electronic thermometer (BAT-8; Bailey Instruments, SaddleBrook, N. J.) and Teflon-sheathed copper-constantan thermocouples(Bailey IT-21 ). The thermocouples were inserted along a 25-gauge needle
track along the longest axis of the tumor. A small amount of radiofrequency power (0.5 watt) was applied, and the thermocouple was movedback and forth until it was at the point of maximum temperature. Powerwas then increased until the thermocouple registered the target temperature. Treatment duration was timed from the moment target temperature was reached. Power was adjusted manually to keep the tumorwithin ±0.1°of target temperature. Similar procedures were used for
heating of normal murine thigh muscle.
FEBRUARY 1984 633
on April 5, 2015. © 1984 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

M.B. Lilly et al.
NMR Spectroscopy. All in vivo 31P-NMR spectra were measuredwithout 'H decoupling on a Bruker CXP-200/300 spectrometer (89-mm
bore) operating at 80.96 MHz in the Fourier transform mode. We usedthe following spectral parameters: 25-/isec pulse (65°flip angle at the
center of the coil); 4K data points with quadrature detection; and 3.2-
sec recycle time per scan. Resolution was enhanced by the convolutiondifference method (line broadenings, 15 and 400 Hz). We confirmedabsence of saturation by varying the recycle time. Tumor spectra werethe sum of at least 512 scans, while those from muscle were the sum ofat least 256 scans. Spectra of tumor extracts were measured by Fouriertransform NMR on a Bruker WH-400 spectrometer.
The surface coil consisted of a 3-turn, flat, circular coil. It was woundfrom a 16-gauge, plastic-coated copper wire and had a diameter of 19mm. The sensitivity (mapped with a sample of phosphoric acid in a 2-
mm capillary) decreased monotonically from the center of the surfacecoil and was negligible beyond about 7 mm along an axis perpendicularto the coil and beyond about 8 mm along an axis in the plane of the coil.The anesthetized mouse was placed in a polyethylene cage whichcontained a hole through which the tumor or leg protruded. The cagewas placed 1.2 cm from the surface coil, with the tissue positioned atthe center of the coil. That only tumor resonances were detected wasconfirmed by the absence of signals when a normal mouse was placedin the probe.
The assignments of resonances to PME, PI, PCr, ADP, ATP, andreduced and oxidized nicotinamide adenine phosphate (collectively referred to as NAD) were made from published values (7). The apparent4
intracellular pH was estimated from the chemical shift of the P, peak.This shift was calibrated with a solution of 12.5 ITIM phosphoric acid,12.5 mM phosphocreatine, and 125 mw NaCI.5 PCr (0.0 ppm) or the ATP
«-phosphorus peak (-7.5 ppm) served as references. Relative concen
trations were estimated from the integrated areas of the corresponding3IP-NMR resonances.
PCA Extraction of Solid Tumors. Immediately after recording of invivo 31P-NMR spectra, an osteosarcoma (0.9 g) was freeze-clamped by
compression between 2 blocks of lead which had been precooled to thetemperature of liquid nitrogen. The mouse was then sacrificed by cervicaldislocation, and the tumor was excised and weighed while still cold.
The frozen tumor (submerged in liquid nitrogen) was pulverized witha precooled mortar and pestle, and cold (4°)0.5 N PCA was added (1
mliter PCA per g of tumor weight). Following evaporation of the cryogen,the PCA-tumor suspension was allowed to stand for 5 to 10 min at 4°
and then centrifugea to remove cellular debris. The supernatant wasremoved and neutralized with 2.5 N potassium bicarbonate. Followinganother centrifugation to remove precipitated potassium perchlorate, thesupernatant was lyophilized to dryness. The residue was dissolved in aminimal amount of water and centrifugea again, and the supernatantwas passed through a column (1x8 cm) of Chelex 100 (sodium form;Bio-Rad Laboratories, Richmond, Calif.). The eluate was again lyophilized
and dissolved in D2O, and the pH of the solution was adjusted to that ofthe in vivo tumor.
RESULTS
Chart 1 presents 3'P-NMR spectra from a murine osteosar
coma. The upper spectrum was obtained from the tumor in vivo.The major peaks are derived from PME (primarily phosphocholineand phosphoethanolamine), Ph and ATP. Several other nucleo-
side triphosphates may contribute to the resonances labeledATP, but their concentrations are much lower. For this reason,the nucleoside triphosphate peaks are labeled only as ATP.Several minor peaks are seen between -2 and 4 ppm. The lower
•rr i i—i—r—
75 01 1 1 1
-7.5 -15
4Roberts et al. (25) have noted that changes in divalent cation binding to P,canlead to changes in the P, chemical shift comparable to those produced by H*binding. For this reason, we refer to "apparent" intracellularpH.
5S. Digemess and E. Blackstone, personalcommunication.
Chart 1. 31P-NMRspectra from untreated Dunn osteosarcoma. Top spectrum
is from in vivo tumor. Bottom spectrum is from PCA extract of same tumor. Identityof the major peaks: (I) ATP9;(//) undinediphosphoglucose(tentative);(///)ATP„;(IV)ATP,; (V) PCr; (VI) glycerophosphocholineand glycerophosphoethanolamine;(VII)P.;and (VIII)phosphocholineand phosphoethanolamine.Additionalsmall peaks canbe seen on the extract spectrum. ADP„is the "shoulder" on the right side of peakIV, ADP„is the "shoulder" on the left side of peak III, and NAD is the "shoulder"onthe right side of peak III. These peaks would be more obvious on an expanded-scale spectrum.
spectrum is from a PCA extract of the same tumor. The freeze-
clamp process appears to have prevented loss of ATP afterdeath, as evidenced by the lack of significant amounts of ADP.While the 2 spectra are basically similar, certain differences areapparent. There is an obvious difference in resolution betweenthe extract and in vivo spectra, with quite a bit of broadeningseen in the latter. This difference is because the phosphorusmetabolites in vivo are largely bound to paramagnetic ions orlarge molecules. Preparation of the PCA extract removes mostof these paramagnetic ions and produces sharper peaks. Thesechanges impair the ability of in vivo NMR spectroscopy to detectand quantitate changes in minor phosphorus-containing species.
For instance, the small peak labeled II (possibly uridine diphosphoglucose) on the extract spectrum is not distinguished fromthe much larger peak III (primarily the «-phosphorus of ATP) on
the in vivo tracing. Peak V (PCr) on the in vivo spectrum appearsto have 2 components, whereas only one is present in theextract. This difference is likely due to the poorer signal/noiseratio seen with in vivo tracing, though we cannot exclude theoccurrence of a labile phosphorus-containing species which was
destroyed during preparation of the extract. We have rarely seen2 peaks in this area on spectra of PCA extracts from other tumorlines. The relative heights of the peaks differ between the 2spectra. For instance, the in vivo spectrum shows much lessPCr (peak V) than does ATP (peak I). In the extract spectrum,however, peak V is taller than peak I. These results are notdiscordant, however, when one examines the area under thepeaks. The integral curve clearly shows that the relative amounts
634 CANCER RESEARCH VOL. 44
on April 5, 2015. © 1984 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Hyperthermia and NMR
of PCr and ATP are similar to those measured in vivo. A finaldifference is in the chemical shift of the ATP peaks. The relativeup-field (more negative) shift of the ATP peaks in the extract isdue to the removal of Mg2+ during preparation.
Charts 2 and 3 show serial 31P-NMR spectra of tumors in 2
mice. The bottom spectrum in each chart presents the pretreatment condition of the tumor. Peaks I, II, and III are derivedprimarily from adenine nucleotides, while Peak V represents P¡.The tumor studied in Chart 1 was heated to 40°for 15 min. This
heat dose produced no apparent cell kill, as demonstrated bysteady tumor growth and rise in serum alkaline phosphatase.There was an immediate decrease in peaks representing ATPand an increase in the P¡peak, reflecting a 50% fall in the ATP/P¡ratio. Five hr later, there was slight recovery of the ATP/P¡ratio. Forty-five hr after hyperthermia, the ATP/P¡ratio had
returned to pretreatment values, with little change thereafter.The apparent intracellular pH, as reflected in the shift of the P¡peak, changed synchronously with the ATP/P( ratio. Immediatelyafter treatment, there was a drop in intracellular pH from 7.2 to6.7. Subsequently, the pH also increased to a maximum at 45hr after treatment.
Chart 3 shows spectra from an osteosarcoma exposed to alarger thermal dose, 47°for 15 min. This heat dose produced at
least 2 logs of cell kill. This was apparent from the shrinkage ofthe tumor (~3 x 108 cells) to below the limits of palpation (~106
cells) and from the fall in the serum alkaline phosphatase tobelow background levels. Immediately after treatment, there wascomplete conversion of ATP to P¡.No recovery of ATP occurredby 24 hr, at which time the tumor grossly necrosed and sloughedoff, preventing further spectroscopic measurements. Smallamounts of PME and NAD (peak at -8 ppm on the 0-hr spectrum)
may have persisted for a few hr after treatment but eventuallydisappeared.
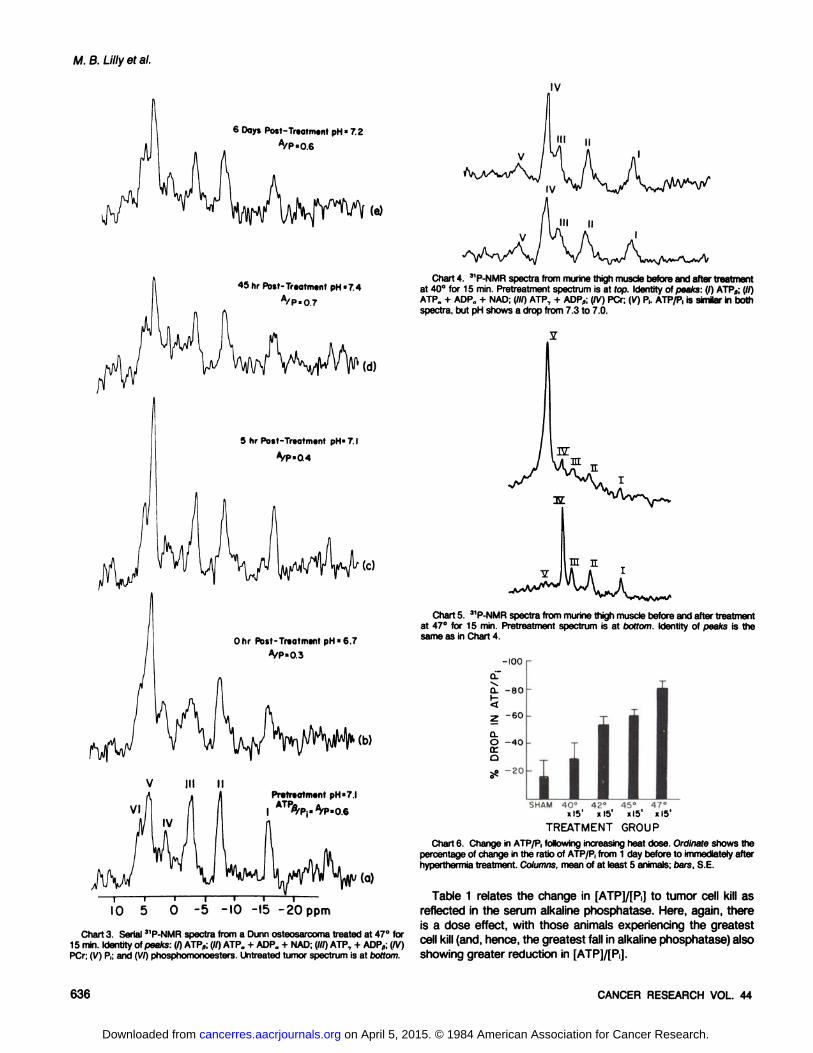
Charts 4 and 5 show pre- and posttreatment 31P-NMR spectra
from the thigh muscle of 2 mice. The pretreatment spectrum ineach case shows the expected large PCr peak, with smallerpeaks from ATP and little P¡.Following mild hyperthermia of 40°
for 15 min (Chart 4), there was some deterioration in the signal/noise ratio. There was no fall in the ATP/P, ratio. The P¡peak didshow an upfield shift, suggesting a fall in the muscle pH from 7.3to 7.0. No functional impairment ensued. After heating to 47°for
15 min (Chart 5), the leg muscle shows nearly complete conversion of high-energy phosphates (both PCr and ATP) to P,, with a
corresponding fall in intracellular pH. No further spectra wererecorded and, several hr later, the leg had become an eschar,without functional ability. These changes in phosphorus metabolites following heating of muscle are qualitatively similar to thoseseen after tumor hyperthermia. The tumor is clearly more sensitive at lower heat doses but, at higher temperatures, the magnitude of P¡loss is similar.
The changes seen by NMR spectroscopy in ATP and P¡maybe useful in the study of thermal dosimetry. Chart 6 presents themean change in the ATP/P¡ratio over the period of a singlehyperthermia treatment, at various heat doses. There is anobvious dose effect, with the higher heat doses producinggreater reduction in ATP and increase in P¡.It can be seen thateven sham-heated animals had a slight decrease in Ph This likely
results from a decrease in tumor tissue perfusion during spectroscopy as a result of the combined effects of pentobarbitalanesthesia and bleeding into the tumor from thermocouple placement.
24 hr Post-Treatment pH-7.3
A/p «0.0
I2 hr Post-Treatment pH=70
A/P=0.0
6 hr Post-Treatment pH=6.8
A/P =00
(0
0 hr Post-Treatment pH=5.9
A/P =O.I
Pretreatment pH »7.4
IO -5 -IO -15 -20 ppm
Chart 2. Serial 3'P-NMR spectra from a Dunn osteosarcoma tumor treated at40°for 15 min. Identity of peaks: (I) ATPfl; (//) ATP. + ADP„+ NAD; (///) ATP, +
ADPfl; (IV) glycerophosphocholine and glycerophosphoethanolamine; (V) P,; and (VI)phosphomonoesters. Untreated tumor spectrum is at bottom.
FEBRUARY 1984 635
on April 5, 2015. © 1984 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
M.B. Lilly et al.
6 Days Post-Treoiment pH«7.2
A/P»0.6
45hr Post-Treatment pH«7.4
A/P*0.7
5 hr Pott-Treatment pH'7.1
A/P-0.4
Ohr Post-Treatment pH«6.7
A/P-0.3
Chart 4. 31P-NMR spectra from murine thigh muscle before and after treatmentat 40°for 15 min. Pretreatment spectrum is at top. Identity of peaks: (I) ATP„;(//)
ATP„+ ADP„+ NAD; (III) ATP, + ADP„;(IV) PCr; (V) P,. ATP/P, is similar in bothspectra, but pH shows a drop from 7.3 to 7.0.
Chart 5. 31P-NMR spectra from murine thigh muscle before and after treatmentat 47°for 15 min. Pretreatment spectrum is at bottom. Identity of peaks is the
same as in Chart 4.
-IOO
oTCL -80
5z -60
CLO -40tro^o -20 ii
SHAM 40°«I5'
42°Xl5'
45°xl5'
47°Xl5'
IO 5 0-5 -IO -15 -20ppm
Chart 3. Serial 3'P-NMR spectra from a Dunn osteosarcoma treated at 47°for
15 min. Identity of peaks: (I) ATP„;(//) ATP0 + ADP„+ NAD; (///) ATP, + ADP«;(IV)PCr; (V) P,; and (VI) phosphomonoesters. Untreated tumor spectrum is at bottom.
TREATMENT GROUP
Chart 6. Change in ATP/P, following increasing heat dose. Ordinate shows thepercentage of change in the ratio of ATP/P, from 1 day before to immediately afterhyperthermia treatment. Columns, mean of at least 5 animals; oars, S.E.
Table 1 relates the change in [ATP]/[P¡]to tumor cell kill asreflected in the serum alkaline phosphatase. Here, again, thereis a dose effect, with those animals experiencing the greatestcell kill (and, hence, the greatest fall in alkaline phosphatase)alsoshowing greater reduction in [ATP]/[PJ.
636 CANCER RESEARCH VOL. 44
on April 5, 2015. © 1984 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Hyperthermia and NMR
Table 1Relationship of ATP/Pi concentration to alkaline phosphatase
% of decrease in alkaline phosphatase
<20 (n = 5)20-80 (n = 4)
>80 (n = 7)
Median % of change in [ATP]/[PJ-11 (60to-73)a-56 (-28 to -78)-80 (-58 to -96)
" Numbers in parentheses, range.
DISCUSSION
Several workers have pointed out differences in the responseof tumors heated in vivo compared to that of those treated invitro (14). These differences include a more rapid cell death ratein excess of that expected from direct thermal injury (22). Somehave speculated that vascular damage may be one of the primarymechanisms of cell death following in vivo hyperthermia (28). Wehave demonstrated a prompt, dose-related loss of high-energy
phosphate, increase in P¡,and fall in intracellular pH in murineosteosarcomas treated with hyperthermia. Such changes areknown to occur in devascularized tissues (4, 23) and are consistent with previous reports of impaired blood flow and oxygenand glucose utilization (3, 29) following hyperthermia.
Our data on ATP fluxes after hyperthermia are at variancewith those of Herman ef al. (19). Using Chinese hamster ovarycells in culture, they found no significant change in ATP concentration, even after substantial heat exposure. An obvious explanation of this discrepancy is that the loss of high-energy phos
phate results primarily from vascular impairment, a parameternot present in tissue culture systems. Our findings of a directcorrelation between loss of ATP, cell kill, and heat dose, whencompared with the previous in vitro data (19), are consistentwith the view that vascular factors may be of extreme importancein in vivo thermal cell kill.
These observations also reinforce the importance of selectingclinically appropriate models for combined modality trials involving hyperthermia. In vivo and in vitro studies may differ markedlyin the effects of treatment on levels of ATP, a substance ofimportance to most cellular processes. Hypoxia and impairedblood flow may greatly modify tumor responses to radiation andchemotherapy. The application of in vivo NMR spectroscopy tohumans is at hand (26). Such technology may therefore helpelucidate the metabolic events occurring during clinical hyperthermia treatment and may help clarify which tumor models mostclosely resemble their human counterparts metabolically.
The clinical development of hyperthermia has been hinderedby imprecise dosimetry. Inhomogenous heat distribution, variablemetabolic states of the cells, and electrical interference withthermocouples have impeded attempts to define a heat "dose"
in physical terms. Much of the recent progress in chemotherapyhas resulted from the use of biological end points (of eitherantitumor effect or toxicity) to define drug dose. The practice oftreating embryonal carcinomas until disappearance of humanchorionic gonadotropin from the blood is a well-known example.
More recently, it has been shown that patients undergoingadjuvant chemotherapy for breast cancer have fewer relapses ifenough drug is given to produce a constant level of myelo-
suppression (9). Changes in the levels of several glycolytic andrespiratory intermediates have been used to predict tissue viability in organ transplant (4) and tumor therapy (12) studies. Theability to express thermal effects in terms of cell damage, rather
than time at temperature, may improve dosimetry. 31P-NMR
spectroscopy offers that potential.In spite of its considerable promise for monitoring phosphorus-
containing species noninvasively, in vivo 31P-NMR spectroscopy
does suffer from some technical limitations. This analytical toolis relatively insensitive. Quantitative information is limited to onlya few of the most abundant phosphorus species. Peaks derivedfrom ATP and P¡are large and present in most spectra. Previousdata6 suggest that the S.D. from measurement of these peaks
is around ±10%. Minor peaks, such as PCr, may be presentonly in very small tumors that are rapidly growing. The variancein measurement of those peak areas may be as high as ±50%.7
We have also noted deterioration of the signal/noise ratio following interventions that disturb tissue homogeneity. This is aparticular problem after hyperthermia treatment, which is associated with hemorrhage from thermocouple insertion, edema,and necrosis of tissues. This variation in signal/noise ratio amongspectra explains our use of a ratio, ATP/P¡,to express quantitative information. Such a ratio is less likely to be affected byvariations in noise than is the absolute area under any particularpeak. At times, it is also necessary to record many more scansper spectrum after treatment than before. We are currentlyexploring several modifications of the surface coil probe toimprove the signal/noise ratio. Our preliminary data suggest thata shielded solenoidal probe may improve the signal/noise ratioby a factor of 3.
The precise biological events leading to thermally induced lossof ATP remain to be defined. While some of the ATP changesare transient, we have still seen a clear-cut dose effect when
NMR spectra are compared with either heat dose or cell kill.Thus, we feel that the changes in 31P-NMR spectra after hyper
thermia reflect metabolically important events. NMR spectroscopy has the advantages of being rapid, noninvasive, and non-perturbing. We feel that in vivo 31P-NMR spectroscopy may be
able to provide prompt feedback on the effectiveness of hyperthermia treatment. Studies to investigate this possibility are nowin progress.
ACKNOWLEDGMENTS
The authors express their gratitude to Dr. Ted Sakai for preparation of the PCAextracts and to Frances Smith, Sharon Garrison, and Maxine Aycock for theirassistance in preparing the manuscript.
ADDENDUMA recent publication by Findly ef al. (16) has shown that in vivo 31P-NMR
spectroscopy can show loss of ATP following heat shock of Tetrahymena.
REFERENCES
1. Ackerman, J. J. H., Grove, T. H., Wong, G. G., Gadian, D. G., and Radda, G.K. Mapping of metabolites in whole animals by :)'P-NMR using surface coils.
Nature (Lond.), 283; 167-170,1980.2. Balaban, R. S., Gadian, D. G., and Radda, G. K. A 31P-NMR study of the rat
kidney in vivo. Kidney Int., 20: 575-579, 1981.3. Sicher, H. l., Hetzel, P. W., Sandhu, T. S., Frinak, S., Vaupel, P., O'hara, M.
D., and O'Brien, T. Effects of hyperthermia on normal and tumor microenviron-
ment. Radiology, 737: 523-530,1980.4. Bore, P. J., Papatheofanis, l., and Seils, R. A. Adenosine triphosphate regen
eration and function in the rat kidney following warm ischemia. Transplantation(Baltimore), 27: 235-237,1979.
6 Unpublished observations.7 Unpublished observation.
FEBRUARY 1984 637
on April 5, 2015. © 1984 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

M.B. Lilly et al.
5. Bronk, B. W. Thermal potentiation of mammalian cell killing: clues for understanding and potential for tumor therapy. Adv. Radiât.Biol., 6:267-324,1976.
6. Brown, F. F., Campbell, I. D., Kuchel, P. W., and Rabenstein, D. C. Humanerylhrocyte metabolism studies by 'H spin echo NMR. FEBS Lett., 82:12-16,
1977.7. Burt, C. T., Cohen, S. M., and Barany, M. A. Analysis of intact tissues with
31P-NMR. Annu. Rev. Biophys. Bioeng., 8: 1-26, 1970.
8. Burt, C. T., Glonek, T., and Barany, M. Analysis of phosphate metabolities,the intracellular pM, and the state of adenosine triphosphate in intact muscleby phosphorus NMR. J. Biol. Chem., 257: 2584-2591,1976.
9. Carpenter, J. T., Maddox, W. A., Laws, H. L, Wirtschafter, D. D., and Soong,S. J. Favorable factors in the adjuvant therapy of breast cancer. Cancer(Phila.), 50:18-23, 1982.
10. DeLayre, J. L., Ingwall, J. S., Malloy, C., and Fossel, E. T. Gated sodium-23nuclear magnetic resonance images of an isolated perfused working rat heart.Science (Wash. D. C.), 212: 935-936,1981.
11. Dewey, W. C., Hopwood, L. E., Sapareto, S. A., and Gerweck, L. E. Cellularresponses to combinations of hyperthermia and radiation. Radiology, 123:463-474,1977.
12. Dickson, J. A., and Shah, D. M. Effects of hyperthermia (42"C) on thebiochemistry and growth of a malignant cell line. Eur. J. Cancer, 8: 561-571,
1972.13. Dobson, C. M., Kirk, K. L., Poulsen, F. M.. Vareri, C. R., and Vecchinoe, J. J.
Studies of human platelets by 19Fand 31PNMR. FEBS Lett., 99: 141, 1979.
14. Eddy, H. A. Alterations in tumor microvasculature during hyperthermia. Radiology, 737:515-521,1980.
15. Evanochko, W. T., Ng, T. C., Lilly, M. B., Lawson, A. J., Corbe«,T. H., Durant,J. R., and Glickson, J. D. In vivo 3'P-NMR study of the metabolism of murinemammary 16/C adenocarcinoma and its response to chemotherapy, X-radia-tion, and hyperthermia. Proc. Nati. Acad. Sei. U. S. A., 80: 334-338,1983.
16. Findly, R. C., Gillies, R. J., and Shulman, R. G. In vivo phosphorus-31NMR
reveals lowered ATP during heat shock of Tetrahymena. Science (Wash. D.C.), 279. 1223-1225, 1983.
17. Ghanta, V. K., Hiramoto, R. N., Weiss, A. B., and Caudill, L. Monitoring ofmurine osteosarcoma by serial alkaline phosphatase determinations. J. Nati.Cancer Inst., 57: 837-839,1976.
18. Gordon, R. E., Hanley, D. E., Shaw, D., Gadian, D. G., Radda, G. K., Styles,P., Bore, P. J., and Chan, L. Localization of metabolites in animals using 3'P
topical magnetic resonance. Nature (Lond.) 287: 736-738, 1980.19. Herman, T. S., Henle, K. J., and Nagle, W. A. Effects of precooling on the
development of thermotolerance and sensitization by step-down heating (Abstract). Radiation Res., 91: 285, 1982.
20. Hiramoto, R. N., Ghanta, V. K., Soong, S., and Hurst, D. C. An experimentalapproach to relate a tumor-associated enzyme marker to tumor cell number.Cancer Res., 37: 365-368,1977.
21. Lapidot, A., and Irving, C. S. Comparative in vivo 15N-NMR study of the cell
wall compartments of five gram-positive bacteria. Biochemistry, 78: 1788-1796,1979.
22. Marmour, J. B., Hahn, N., and Hahn, G. M. Tumor cure and cell survival afterlocalized radiofrequency heating. Cancer Res., 37: 879-883, 1977.
23. Ng, T. C., Evanochko, W. T., Hiramoto, R. N., Ghanta, V. K., Lilly, M. B.,Lawson, A. J., Corbett, T. H., Durant, J. R., and Glickson, J. D. 3'P-NMRspectroscopy of in vivo tumors. J. Magn. Reson., 49: 271-286, 1982.
24. Radda, G. K., Ackerman, T. T. H., Bore, P., Sehr, T., Wong, G., Ross, B. D.,Green, Y., Bartlett, S., and Lowry, M. 31P-NMR studies on kidney intracellularpH in acute renal acidosis. Int. J. Biochem., 72: 277-281, 1980.
25. Roberts, J. K. M., Wade-Jardetzsky, N., and Jardetzsky, 0. Intracellular pHmeasurements by 3'P-NMR. Influence of factors other than pH on 31Pchemical
shifts. Biochemistry, 20: 5389-5392, 1981.26. Ross, B. D., Radda, G. K., Gadian, D. G., Rocker, G., Esiri, M., and Smith, J.
F. Examination of a case of suspected McArdle's Syndrome by 31P-NMR. New
Engl. J. Med., 304: 1338-1343, 1981.27. Schulman, R. G., Ugurbil, K., Brown, T. R., Rottenberg, H., Ogawa, S.,
Yamane, T., Navon, G., Der Hollander, J. A., and Cohen, S. M. 31P- and '3C-
NMR studies of metabolism and bioenergetics in suspensions of living cells.Biophys. J., 27: 154a, 1978.
28. Song, C. W., Kang, M. S., Rhee, J. G., and Levitt, S. H. Effect of hyperthermiaon vascular function, pH, and cell survival. Radiology, 737: 795-803,1980.
29. Vaupel, P., Ostoheimer, K., and Muller-Kleiser, W. Circulatory and metabolicresponses of malignant tumors during localized hyperthermia. J. Cancer Res.Clin. Oncol., 98. 15-29, 1980.
638 CANCER RESEARCH VOL. 44
on April 5, 2015. © 1984 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1984;44:633-638. Cancer Res Michael B. Lilly, Thian C. Ng, William T. Evanochko, et al. Spectroscopy
P-Nuclear Magnetic Resonance31 in VivoDemonstrated by Loss of High-Energy Phosphate following Hyperthermia
Updated version
http://cancerres.aacrjournals.org/content/44/2/633
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on April 5, 2015. © 1984 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from




















