
Local Structural Preferences and Dynamics Restrictions in the Urea-Denatured State of SUMO1: NMR...
12
Local Structural Preferences and Dynamics Restrictions in the Urea-Denatured State of SUMO-1: NMR Characterization Ashutosh Kumar,* Sudha Srivastava,* Ram Kumar Mishra, y Rohit Mittal, y and Ramakrishna V. Hosur* *Department of Chemical Sciences and y Department of Biological Sciences, Tata Institute of Fundamental Research, Mumbai, India ABSTRACT We have investigated by multidimensional NMR the structural and dynamic characteristics of the urea-denatured state of activated SUMO-1, a 97-residue protein belonging to the growing family of ubiquitin-like proteins involved in post- translational modifications. Complete backbone amide and 15 N resonance assignments were obtained in the denatured state by using HNN and HN(C)N experiments. These enabled other proton assignments from TOCSY-HSQC spectra. Secondary H a chemical shifts and 1 H- 1 H NOE indicate that the protein chain in the denatured state has structural preferences in the broad b-domain for many residues. Several of these are seen to populate the (f,c) space belonging to polyproline II structure. Although there is no evidence for any persistent structures, many contiguous stretches of three or more residues exhibit structural propensities suggesting possibilities of short-range transient structure formation. The hetero-nuclear 1 H- 15 N NOEs are extremely weak for most residues, except for a few at the C-terminal, and the 15 N relaxation rates show sequence-wise variation. Some of the regions of slow motions coincide with those of structural preferences and these are interspersed by highly flexible residues. The implications of these observations for the early folding events starting from the urea-denatured state of activated SUMO-1 have been discussed. INTRODUCTION Characterization of the unfolded/denatured states of proteins is important for many reasons; here, the term ‘‘unfolded’’ refers to a state completely devoid of any structural prefer- ences and the term ‘‘denatured’’ refers to the lowest energy nonnative state under a given set of conditions (1). The un- folded and the denatured states may coincide under extreme denaturing conditions. Firstly, the denatured state represents the reference state for estimating the stability of a protein. Next, the denatured state is the starting point for the folding process in vivo. In this context, it has been a major area of research to understand how the unfolded chain finds its final folded state destination (2–4). There is now ample evidence to believe that the different members of the unfolded en- semble fold along different pathways on a so-called folding funnel, whose broad end represents the unfolded state, and the narrow end, the folded state (5–7). Although it is almost impossible to know at this point of time, what the exact nature of the denatured state in vivo is, it is believed that the chain is not entirely random but has at least some local preferences on the average that restrict the conformational search by the folding chain. The local preferences could vary depending upon the environmental conditions. In view of this, there is vigorous effort in the literature to characterize the denatured states created by a variety of means, which include use of chemical denaturants (2,8–14), change of pH conditions (15–17), or thermal denaturation (18,19). It has been observed that different conditions populate different conformational states in the ensemble (20,21). Among the various means used for creating denatured states, guanidine denaturation is known to cause the maximum degree of unfolding. This is followed by urea and other denaturing agents. pH denaturation appears to be more tolerant and so is temperature. Sodium dodecyl sulphate micelles cause mostly removal of tertiary interactions (22). Consequently, as must be expected, the equilibrium unfolding pathways in many of these cases have also been found to be different (23–26); some follow a two-state process, whereas others follow mul- tistate processes which have implications for protein stability studies. Thus, study of all such denatured states is expected to provide insights into the different pathways of folding, starting from the different denatured states in the ensemble. The unfolded state and the partially unfolded states have also been thought to be playing important roles in protein function. Recently, the structure-function paradigm has been revisited and a model called the ‘‘protein-quartet model’’ has been put forward, according to which a protein function arises as a consequence of an interplay between folded states, unfolded states, molten globules, and premolten globules (27,28). A direct role of protein unfolding has been seen in transport of proteins across the membranes (29). Lastly, the denatured states of proteins have often been seen to cause protein aggregation (30), and such aggregation is known to be one of the reasons for many diseases in vivo (31,32). One type of structure reported by NMR in denatured proteins is a hydrophobic cluster, usually formed by local side-chain interactions (33–36). Another type of structure reported is a fluctuating secondary structure. In the case of Submitted July 30, 2005, and accepted for publication December 29, 2005. Address reprint requests to Prof. Ramakrishna V. Hosur, Dept. of Chemical Sciences, Tata Institute of Fundamental Research, Homi Bhabha Road, Mumbai 400 005, India. Tel.: 91-22-2280-4545; Fax: 91-22-2280-4610; E-mail: [email protected]. Abbreviations used: HSQC, hetero-nuclear single quantum coherence; NMR, nuclear magnetic resonance; NOESY, nuclear Overhauser effect spectroscopy; SUMO, small ubiquitin related modifier. Ó 2006 by the Biophysical Society 0006-3495/06/04/2498/12 $2.00 doi: 10.1529/biophysj.105.071746 2498 Biophysical Journal Volume 90 April 2006 2498–2509
-
Upload
pioneerurban -
Category
Documents
-
view
1 -
download
0
Transcript of Local Structural Preferences and Dynamics Restrictions in the Urea-Denatured State of SUMO1: NMR...

Local Structural Preferences and Dynamics Restrictions in theUrea-Denatured State of SUMO-1: NMR Characterization
Ashutosh Kumar,* Sudha Srivastava,* Ram Kumar Mishra,y Rohit Mittal,y and Ramakrishna V. Hosur**Department of Chemical Sciences and yDepartment of Biological Sciences, Tata Institute of Fundamental Research, Mumbai, India
ABSTRACT We have investigated by multidimensional NMR the structural and dynamic characteristics of the urea-denaturedstate of activated SUMO-1, a 97-residue protein belonging to the growing family of ubiquitin-like proteins involved in post-translational modifications. Complete backbone amide and 15N resonance assignments were obtained in the denatured state byusing HNN and HN(C)N experiments. These enabled other proton assignments from TOCSY-HSQC spectra. Secondary Ha
chemical shifts and 1H-1H NOE indicate that the protein chain in the denatured state has structural preferences in the broadb-domain for many residues. Several of these are seen to populate the (f,c) space belonging to polyproline II structure. Althoughthere is no evidence for any persistent structures, many contiguous stretches of three or more residues exhibit structuralpropensities suggesting possibilities of short-range transient structure formation. The hetero-nuclear 1H-15N NOEs are extremelyweak formost residues, except for a few at theC-terminal, and the 15N relaxation rates show sequence-wise variation. Some of theregions of slow motions coincide with those of structural preferences and these are interspersed by highly flexible residues. Theimplications of these observations for the early folding events starting from the urea-denatured state of activated SUMO-1 havebeen discussed.
INTRODUCTION
Characterization of the unfolded/denatured states of proteins
is important for many reasons; here, the term ‘‘unfolded’’
refers to a state completely devoid of any structural prefer-
ences and the term ‘‘denatured’’ refers to the lowest energy
nonnative state under a given set of conditions (1). The un-
folded and the denatured states may coincide under extreme
denaturing conditions. Firstly, the denatured state represents
the reference state for estimating the stability of a protein.
Next, the denatured state is the starting point for the folding
process in vivo. In this context, it has been a major area of
research to understand how the unfolded chain finds its final
folded state destination (2–4). There is now ample evidence
to believe that the different members of the unfolded en-
semble fold along different pathways on a so-called folding
funnel, whose broad end represents the unfolded state, and
the narrow end, the folded state (5–7). Although it is almost
impossible to know at this point of time, what the exact
nature of the denatured state in vivo is, it is believed that the
chain is not entirely random but has at least some local
preferences on the average that restrict the conformational
search by the folding chain. The local preferences could vary
depending upon the environmental conditions. In view of
this, there is vigorous effort in the literature to characterize
the denatured states created by a variety of means, which
include use of chemical denaturants (2,8–14), change of pH
conditions (15–17), or thermal denaturation (18,19). It has
been observed that different conditions populate different
conformational states in the ensemble (20,21). Among the
various means used for creating denatured states, guanidine
denaturation is known to cause the maximum degree of
unfolding. This is followed by urea and other denaturing
agents. pH denaturation appears to be more tolerant and so is
temperature. Sodium dodecyl sulphate micelles cause mostly
removal of tertiary interactions (22). Consequently, as must
be expected, the equilibrium unfolding pathways in many
of these cases have also been found to be different (23–26);
some follow a two-state process, whereas others follow mul-
tistate processes which have implications for protein stability
studies. Thus, study of all such denatured states is expected
to provide insights into the different pathways of folding,
starting from the different denatured states in the ensemble.
The unfolded state and the partially unfolded states have
also been thought to be playing important roles in protein
function. Recently, the structure-function paradigm has been
revisited and a model called the ‘‘protein-quartet model’’ has
been put forward, according to which a protein function
arises as a consequence of an interplay between folded states,
unfolded states, molten globules, and premolten globules
(27,28). A direct role of protein unfolding has been seen in
transport of proteins across the membranes (29). Lastly, the
denatured states of proteins have often been seen to cause
protein aggregation (30), and such aggregation is known to
be one of the reasons for many diseases in vivo (31,32).
One type of structure reported by NMR in denatured
proteins is a hydrophobic cluster, usually formed by local
side-chain interactions (33–36). Another type of structure
reported is a fluctuating secondary structure. In the case of
Submitted July 30, 2005, and accepted for publication December 29, 2005.
Address reprint requests to Prof. Ramakrishna V. Hosur, Dept. of Chemical
Sciences, Tata Institute of Fundamental Research, Homi Bhabha Road,
Mumbai 400 005, India. Tel.: 91-22-2280-4545; Fax: 91-22-2280-4610;
E-mail: [email protected].
Abbreviations used: HSQC, hetero-nuclear single quantum coherence;
NMR, nuclear magnetic resonance; NOESY, nuclear Overhauser effect
spectroscopy; SUMO, small ubiquitin related modifier.
� 2006 by the Biophysical Society
0006-3495/06/04/2498/12 $2.00 doi: 10.1529/biophysj.105.071746
2498 Biophysical Journal Volume 90 April 2006 2498–2509

barnase (37,38), the urea-denatured protein shows regions of
fluctuating secondary structure, which correspond fairly well
to the moderately and highly structured regions of the inter-
mediates and the transition states of folding (39). Native- and
nonnative-type secondary structural preferences were seen in
the urea-denatured state of barstar (9), guanidine-denatured
state of HIV-1 protease (2,8,10), and very recently in the
acid-denatured state of hUBF HMG Box 1 (15). A nonnative
a-helical structure was observed in the guanidine denatured
state of b-lactoglobulin, an all-b-protein in the native state
(40). Similarly, b-type preferences were seen in denatured
apomyoglobin, a largely helical protein (41). In some cases,
multiple types of structures have been reported in the dena-
tured state. For example, for the drkN SH3 domain, detailed
NMR studies (42–44) indicate the presence of multiple
structures ranging from conformers with non-native structure
possessing long-range contacts to those with more compact
structures maintaining native-like secondary structure.
There is growing evidence in the literature, mostly based on
circular dichroism (CD) spectroscopy, that chemically dena-
tured proteins populate polyproline (PPII) structure which lies
in the b-domain of (f,c) space of the Ramachandran map.
This is a left-handed helical structure, generally observed in
proline-rich peptides (45–48) and has a characteristic positive
CD band at 225 nm. Further, where residual structures in the
b-domain have been detected by NMR in denatured proteins,
it is suggested that PPII structure may be populated to a sig-
nificant extent (49,50).
In view of the above, we have initiated investigation of
the structural and dynamic characteristics of the variety of
denatured states of the protein, SUMO-1, a 101-residue
protein belonging to the growing family of ubiquitin-like
proteins involved in post-translational modifications (51). It
attaches itself to many target proteins, by a process called
‘‘sumoylation’’. Before sumoylation the protein gets acti-
vated by cleaving off four residues from the C-terminal end.
There have been two reports in the literature on the structure
of the 101-residue SUMO-1 (52,53). We reported the NMR
structure of the activated SUMO-1 at pH 7.4 and 27�C (54).
All three structures were essentially similar and the average
structure of the molecule contains the following secondary
structural elements: b1: 22–28; b2: 33–39; b3: 62–66; b4:
69–72; b5: 87–93; a1: 45–55, and a2: 77–82. The b-strands
form a bent sheet. The regions intervening between second-
ary structures form flexible loops. The amino-terminal 20
residues are highly flexible. In this article, we report NMR
investigations on the characteristics of the denatured state of
SUMO-1 (1–97) created by 8 M urea at pH 5.6 and 27�C.
Hereafter, we refer to this protein as SUMO-1 only for sake
of brevity. Our results indicate that in the urea-denatured
state, a large number of residues have slight (f,c) prefer-
ences in the broad b-domain, which encompasses the ex-
tended structures as well. The dynamic motions at the high
frequency scale appear to be fairly restricted. There are
sequence-wise small variations in the milli- to microsecond
timescale motions. These results provide a description of the
denatured state on the one hand, and also provide clues to the
possible nucleation sites for the folding reaction of the pro-
tein, on the other.
MATERIALS AND METHODS
Protein preparation
SUMO-1 was prepared and purified as described elsewhere (54).
NMR spectroscopy
Isotopically labeled protein samples for NMR experiments were prepared as
described in Mishra et al. (54). The proteins were concentrated to ;1 mM
and exchanged with 0.1 M phosphate buffer (pH 5.6) containing 150 mM
NaCl, 5 mM EDTA, 1 mM DTT, and 8 M urea. The NMR experiments were
started after keeping the solutions for ;3 h so as to reach equilibrium.
All the NMR experiments were performed on a Varian Inova 600 MHz
spectrometer at 27�C (Varian, Cary, NC). A series of two-dimensional and
three-dimensional experiments (see below) were carried out which lasted for
six days. At the end of these the HSQC spectrum was again recorded to check
for the stability of the protein. We observed no change in the HSQC spectra
indicating that the protein was very stable and also had reached equilibrium
state at the beginning of the experiments. HN and 15N resonance assignments
were obtained using HNN and HN(C)N triple resonance experiments (55–
57). HNCA, HN(CO)CA, TOCSY-HSQC, and NOESY-HSQC experiments
(reviewed in (58,59)) facilitated the assignments by providing additional
checks. Standard experimental parameters were used: 24–30 ms for N-Ca and
N-C9 transfers in HNN, HN(C)N, HNCA, and HN(CO)CA; and 60–80 ms for
TOCSY transfer and 150–200 ms for NOESY transfers. The 4–16 scans were
used for each fid in the different experiments. The 40–50 and 80 complex
increments were used along the 15N and C9 dimensions respectively; 60–80
complex increments were used along the aliphatic carbon dimensions; 80–96
complex increments were used along the indirect 1H dimension. Temperature
coefficients of amide protons were measured by recording HSQC spectra at
3�C temperature increments in the range 15–36�C. 15N transverse relaxation
rates (R2) were measured using CPMG delays, 10, 50, 70*, 90, 130, 170*,
210, and 250 ms, where asterisks indicate duplicate measurement. 15N longi-
tudinal relaxation rates (R1) were measured using the inversion recovery
delays 10, 50, 120*, 220, 350, 700*, and 950 ms. Steady-state 1H-15N hetero-
nuclear NOE measurements were carried out with a proton saturation time of
3 s and a relaxation delay of 2 s. For the experiment without proton saturation
the relaxation delay was 5 s. All the relaxation experiments were carried out
using the pulse sequences described by Farrow et al. (60). HN-Ha coupling
constants were measured from a high resolution HSQC spectrum recorded
with 8192 complex t2 points and 512 complex t1 points.
Circular dichroism
Far-UV circular dichroism (CD) spectra of the protein at 27�C, were
recorded on a JASCO–J810 spectropolarimeter (Jasco, Hachioji, Japan), at 0
and 8 M urea concentrations using 0.2 cm cell. The protein concentration
was 20.7 mM. The samples at the appropriate conditions were equilibrated
for at least 10–12 h before CD measurements. Each spectrum was an average
of eight scans (slit width of 2 nm).
RESULTS AND DISCUSSION
Resonance assignments
Resonance assignment in denatured proteins by the standard
triple resonance experiments such as HNCA, HN(CO)CA,
NMR Characterization of Denatured SUMO-1 2499
Biophysical Journal 90(7) 2498–2509

HNCACB, CBCANH, CBCA(CO)NH, etc., which have
proved extremely successful for folded proteins, is severely
hampered because of the high degeneracy of the amide and
carbon chemical shifts (61). Consequently there have been
relatively few proteins on which detailed studies at residue
level have been carried out (61–65). In such situations, recent
experiments that exploit the 15N or CO chemical shifts,
which have good dispersions in both folded and unfolded
proteins, would be extremely valuable (65,66). Presently, we
were able to obtain complete amide proton and backbone15N assignments for SUMO-1, at pH 5.6 and 27�C, in 8 M
urea using the HNN and HN(C)N experiments described
by us a few years ago (55–57). These exploit the good
dispersion of the 15N chemical shifts along two of the three
dimensions, and also provide many checkpoints along the
sequential walk by way of special patterns of peaks around
Gly and Pro residues. Thus side-chain identification does not
become very essential during the sequential walk, and this
enhances the speed of assignment. However, under rare occa-
sions of overlaps, TOCSY-HSQC spectrum, which provides
the spin system information, helps resolve the ambiguities.
An illustrative sequential walk through the HNN spectrum of
SUMO-1 at 27�C, pH 5.6 is shown in Fig. 1 A and the
summary of all the sequential connectivities is shown in Fig.
1 B. Fig. 2 shows the 1H-15N HSQC spectrum with all the
peak assignments. During the course of these sequential as-
signments, many of the side-chain proton assignments were
obtained from the TOCSY-HSQC spectrum.
Structural propensities in the denatured state
Deviations of observed chemical shifts from the random coil
values (secondary shifts) are very good indicators of sec-
ondary structures in proteins. The random coil values are
generally measured using short 5–6 residue peptides, as they
do not possess any structural preferences in solution. These
are particularly useful in denatured proteins where NOEs are
rather scarce. However, there have been more than one set of
random coil values reported in the literature and these differ
due to slightly different experimental conditions. We have
calculated these shifts using two sets, one by Wishart and
Sykes (67,68) and the other by Schwarzinger et al. (69). The
former uses pH 5 and 1 M urea, while the latter uses 8 M urea
and pH 2.3 for the measurements on peptides, GGXGG,
where X is the residue of interest for random coil chemical
shift and G is glycine. In our case the two calculations made
only a marginal difference and we decided to use the data
given by Schwarzinger et al. (70), since these authors have
also described a set of sequence corrections to the random
coil values for all the nuclei.
Among the various secondary chemical shifts (Dd), those
of Ha, 13Ca, and 13CO are the most diagnostic of structural
information in the polypeptide chain (63,64). Thus if some
residues show positive (downfield) Ha and negative (upfield)I3Ca and I3CO structure-dependent chemical shift deviations
(secondary shifts, Dd), then those residues are taken to have
(f,c) preferences in the b-domain of the Ramachandran map
FIGURE 1 (A) Sequential walk through the F1–F3
planes of HNN spectrum of SUMO-1 in 8 M urea at pH
5.6 and 27�C. Sequential connectivities are shown for
Gly-19 to Leu-24 stretch. The F2 (15N) values are
shown at the top for each strip. The black and red con-
tours indicate positive and negative peaks. The distinct
Gly plane serves as the start point in the sequential
assignment. (B) Summary of sequential assignment
obtained from HNN walk along the primary sequence
of SUMO-1. Gly shown in green worked as start/check
point; Pro shown in red worked as break point.
2500 Kumar et al.
Biophysical Journal 90(7) 2498–2509

((f,c) ¼ (�70 to �150�, 90–180�)). Residues with a-helical
propensities ((f,c) ¼ (�40 to �80�, �30 to �60�)) show
the opposite pattern.
We have used Ha secondary shifts to characterize the
residual structure in the protein. Fig. 3 A shows a plot of the
secondary shifts against the sequence for SUMO-1 at pH 5.6
and 27�C. Clearly, from the pattern of the deviations which
show a positive bias, the chain does not appear to be a
random coil. Though the shifts are small, overall there seems
to be some tendency for a large number of residues across the
chain to populate the broad b-region of the (f,c) space; this
includes the extended conformations and PPII structures
((f,c) ¼ (�70 to �80�, 140 to 150�)) as well. There are a
few contiguous stretches of three or more residues showing
positive deviations of .0.08 ppm and these may be taken to
indicate formation of b-structures, perhaps transiently, along
the chain. They have been indicated by solid horizontal bars
inside the figure.
Next, we measured 3J (HN- Ha) coupling constants for all
the residues. Three bond HN-Ha coupling constants are
useful indicators of f-torsion angles. This coupling constant
is of the order of 3–5 Hz for a helical structure (including
PPII) and 8–11 Hz for a b-structure (64). For a random coil, a
weighted average of these values would be observed, which
is typically between 6.0 and 8.0 Hz for most residues. It is
also observed that the random coil value for any residue is
influenced by its N-terminal neighbor and thus two sets of
values have been reported for each residue, depending upon
whether the N-terminal neighbor belongs to one of the two
groups of residues (group I: F, W, H, Y, I, T, V, and group II:
remaining residues except glycine) (71). Thus, under any
given experimental conditions, one can calculate deviation
of the observed coupling constant from the sequence-
dependent random coil value, (Jobs–Jrc), for every residue.
These ‘‘secondary coupling constants,’’ as we call them,
throw valuable light on the secondary structural propensities
along the polypeptide chain. Negative secondary coupling
constants would indicate helical propensities (including PPII)
and positive secondary coupling constants would indicate
b-propensities (71). This led us to the idea that a combined
use of secondary chemical shifts and secondary coupling con-
stants would enable distinction between b and PPII struc-
tures. For a truly b-structure, both the secondary shifts and
the secondary coupling constants would be positive, whereas
for the PPII helix the secondary coupling constant would be
negative, as in the common right-handed a-helices, and the
secondary shifts would be positive, like in the b-structures.
The measured values of the secondary coupling constants
from a high resolution HSQC spectrum of the protein (an
illustrative region to show the quality of the spectral reso-
lution is shown in Fig. 3 B), are shown in Fig. 3 C. The
deviations are mostly ,1.0 Hz for most residues. We may
mention here that the accuracy of secondary coupling con-
stant estimation is ;0.5 Hz for the positive values, and
FIGURE 2 (1H-15N) HSQC spec-
trum of SUMO-1 in 8 M urea at pH
5.6 and 27�C. Residue-specific assign-
ment for each peak is marked on the
spectrum.
NMR Characterization of Denatured SUMO-1 2501
Biophysical Journal 90(7) 2498–2509
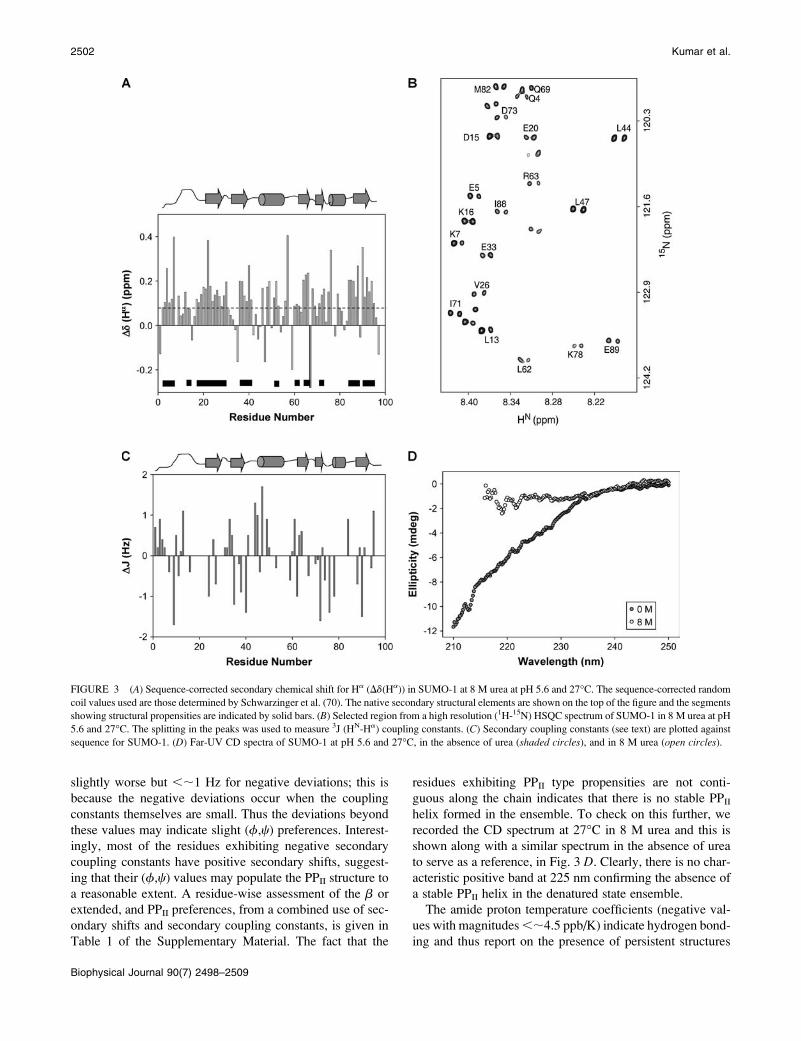
slightly worse but ,;1 Hz for negative deviations; this is
because the negative deviations occur when the coupling
constants themselves are small. Thus the deviations beyond
these values may indicate slight (f,c) preferences. Interest-
ingly, most of the residues exhibiting negative secondary
coupling constants have positive secondary shifts, suggest-
ing that their (f,c) values may populate the PPII structure to
a reasonable extent. A residue-wise assessment of the b or
extended, and PPII preferences, from a combined use of sec-
ondary shifts and secondary coupling constants, is given in
Table 1 of the Supplementary Material. The fact that the
residues exhibiting PPII type propensities are not conti-
guous along the chain indicates that there is no stable PPII
helix formed in the ensemble. To check on this further, we
recorded the CD spectrum at 27�C in 8 M urea and this is
shown along with a similar spectrum in the absence of urea
to serve as a reference, in Fig. 3 D. Clearly, there is no char-
acteristic positive band at 225 nm confirming the absence of
a stable PPII helix in the denatured state ensemble.
The amide proton temperature coefficients (negative val-
ues with magnitudes ,;4.5 ppb/K) indicate hydrogen bond-
ing and thus report on the presence of persistent structures
FIGURE 3 (A) Sequence-corrected secondary chemical shift for Ha (Dd(Ha)) in SUMO-1 at 8 M urea at pH 5.6 and 27�C. The sequence-corrected random
coil values used are those determined by Schwarzinger et al. (70). The native secondary structural elements are shown on the top of the figure and the segments
showing structural propensities are indicated by solid bars. (B) Selected region from a high resolution (1H-15N) HSQC spectrum of SUMO-1 in 8 M urea at pH
5.6 and 27�C. The splitting in the peaks was used to measure 3J (HN-Ha) coupling constants. (C) Secondary coupling constants (see text) are plotted against
sequence for SUMO-1. (D) Far-UV CD spectra of SUMO-1 at pH 5.6 and 27�C, in the absence of urea (shaded circles), and in 8 M urea (open circles).
2502 Kumar et al.
Biophysical Journal 90(7) 2498–2509

(72). For random coils the temperature coefficients are
roughly in the range �6 to �9.5 ppb/K for the different
residues (73). We measured the residue-wise amide proton
temperature coefficients of the protein from HSQC spectra
recorded in the temperature range 15–36�C. Since all the
measured values were negative we calculated their devia-
tions from the random coil values by taking magnitudes
only, and these results are shown in Fig. 4. We see that the
observed deviations D(DdNH/DT) are largely positive. From
the absence of significant negative deviations, it is clear that
there are no stable intramolecular H-bonds formed in most
regions of the polypeptide chain. This rules out the possi-
bility of persistent well-defined structural elements like
a-helix, b-sheet, b-turn, etc., in the denatured state.1H-1H NOEs in folded proteins exhibit secondary struc-
ture specific patterns. b-structures including PPII and type II
turns are characterized by strong Ha(i) – HN(i11) NOEs,
whereas a-helices are characterized by strong HN(i) –
HN(i11) NOEs (74). The a-helices also produce medium
range NOEs, from HN (i) to Ha (i�3) and Ha(i�4). In ad-
dition, several other NOEs will also be seen from the amide
protons to other intraresidue protons. In unfolded proteins,
however, the NOEs are generally weak in the first place due
to greater flexibility of the chain, and the above selectivity
with regard to secondary structure may be lost due to mea-
surably good population of the different secondary structure
types in the ensemble. However, if one does find any kind
of preference in the NOE patterns, then that can be taken to
indicate a higher population of those specific structures. In
SUMO-1 denatured by 8 M urea, the 1H-1H NOEs in the
NOESY-HSQC spectra recorded with a mixing time of
150 ms, are rather sparse, but, interestingly, do show some
pattern. These are shown in Fig. 5, which seem to support the
conclusions derived above from the secondary shifts. A ma-
jor portion of the chain shows fairly intense sequential Ha(i)– HN(i11) NOEs, and only a few of them also show self-
NOEs. In a dipeptide, the sequential distance, daN(i, i11) is
sensitive to torsion angle ci and varies from 2.2 to 3.5 A�, the
shortest distance occurring for ci in the b-domain. Likewise,
the distance between Ha and HN within the same residue,
daN(i, i), which is sensitive to the torsion angle fi, varies
within a narrow range from 2.2 to 2.9 A� (74). Thus, for
those residues where only the sequential peaks are seen, the
(f,c) propensity has a preponderance of theb-structure, which
has the shortest sequential distance and the longest intra-
residue distance. For those residues where both the peaks are
observed, the fi may show slight preference toward a shorter
intraresidue distance, still within the b-domain. From the NOE
data, we conclude that the regions, 9–15, 17–33, 43–48, 61–
68, 71–74, 82–87, and 90–95 have a higher population of
b-structures in the ensemble.
All the above observations taken together indicate that the
SUMO-1 polypeptide chain in 8 M urea does not possess any
stable secondary structures, but has some propensity for for-
mation of transient structures with (f,c) preferences in the
broad b-domain, in 4–5 regions. Several residues in these
regions seem to populate the (f,c) space belonging to the
PPII structure. The structural propensities presently observed
in the 8 M urea-denatured state encompass both native and
nonnative type preferences for the individual residues, and
this would have implications for the folding mechanisms of
the protein.
Dynamics restrictions
Since the protein chain exhibited some local structural pref-
erences in the denatured ensemble, it is intuitively natural to
expect some variations in the motional characteristics along
the length of the chain as well. To probe this explicitly, we
carried out 15N R1, R2 and heteronuclear 1H-15N NOE
measurements, which throw light on the motions over a wide
range of timescales (75–77). Among these, the NOEs are
very sensitive to picosecond timescale motions, R2 values are
sensitive to milli- to microsecond timescale motions (con-
formational transitions) and the R1 values are sensitive to both
low and high frequency motions (nanosecond-to-picosecond
timescale motions). The heteronuclear NOE intensity varies
from ;�4 to 0.9 depending upon the motional correlation
times (75,78). Negative values indicate high frequency
(picosecond timescale) large amplitude motions, whereas
positive values indicate nanosecond timescale motions. In
the denatured states, where there is heterogeneity of the con-
formational states and of the correlation times, these relax-
ation and NOE data can only be interpreted qualitatively to
derive relative motional trends along the chain. Reduced
spectral density analysis based on 15N R1, R2 and heteronu-
clear 1H-15N NOE is the best suited method for studying
motions at residue level in denatured proteins (15,64). How-
ever, in the present case this was hampered by the absence of
measurable 1H-15N NOEs due to their low intensities (see
below); this of course must have a bearing on the motional
characteristics themselves. Nevertheless, we have been able
FIGURE 4 Secondary temperature coefficients. Deviation of amide pro-
ton temperature coefficients from the random coil temperature coefficient
(magnitude mode) at pH 5.6 plotted against the sequence. The native
secondary structure elements are indicated at the top.
NMR Characterization of Denatured SUMO-1 2503
Biophysical Journal 90(7) 2498–2509

to obtain some useful insights into these motions, especially
for the nanosecond timescale components and the confor-
mational transitions, from a qualitative analysis of the R1, R2
relaxation rates.
Fig. 6 shows 1H-15N correlation spectra recorded during
the 1H-15N steady-state NOE measurements. The spectrum
in Fig. 6 A is obtained without irradiation of the protons
and the spectrum in Fig. 6 B is the one recorded with pro-
ton irradiation of 3 s to reach the steady state. Although the
normal spectrum is similar to the typical HSQC spectrum,
the spectrum in Fig. 6 B shows only four peaks; the peaks
have also good intensities, as can be seen from the cross sec-
tion for the weakest of the peaks. This is somewhat unusual
compared to similar studies on other denatured proteins
where the HSQC spectra recorded in the presence of proton
saturation have good peak intensities and a distribution of
positive and negative NOEs has been seen. As a reference,
we show in Fig. 6 C the spectrum on folded SUMO-1 (pH
7.4, 27�C), which was recorded with proton irradiation and
the same protein concentration as in A and B. This spectrum
has clearly good intensities. Thus the present observation of
only four discernible peaks in spectrum B in Fig. 6 must have
dynamic implications. Firstly, two of the four peaks have
greater intensity than the corresponding ones in spectrum A,
and this can happen only if the 1H-15N NOEs are negative.
All the peaks in spectrum B have the same phase and there-
fore the other two peaks also must be negative in sign. As
marked, these peaks belong to the C-terminal of the protein,
and the negative sign indicates that this segment is highly
mobile, exhibiting high amplitude picosecond timescale
motions. This, however, is not surprising, since the terminal
residues generally have been seen to exhibit a greater
flexibility than the other residues. More importantly, the fact
that no other peak is seen in spectrum B, suggests that the
NOEs are very small, perhaps below the noise level and a
simple explanation for this could be that the rest of the entire
FIGURE 5 Summary of sequence-specific1H-1H connectivities obtained from the NOESY-15N-1H HSQC spectrum.
FIGURE 6 Spectra showing 15N-1H heteronu-
clear NOE of SUMO-1 in 8 M urea at pH 5.6 and
27�C. (A) Reference spectrum without irradia-
tion of protons; (B) spectrum with irradiation of
protons for 3 s; and (C) spectrum of the folded
protein (pH 7.4, 27�C) with proton irradiation
as in B. In B, a cross section through the peak
for Q94 (Q94 is enclosed in a box) is shown to
demonstrate the good sensitivity in the spectrum.
The red and black contours indicate negative and
positive peaks.
2504 Kumar et al.
Biophysical Journal 90(7) 2498–2509

chain is relatively more restricted, i.e., the amplitudes of the
high frequency motions are small. And, the correlation times
for the individual N-H vectors (in the nanosecond regime)
may also be such that the steady-state NOEs are nearly zero.
Fig. 7 A shows residue-wise R1 values in the SUMO-
1 protein at 27�C. The average relaxation rate is ;1.69 s�1,
which is typically the value one sees for this size protein.
Interestingly, however, we see some sequence-dependent
variations along the chain, some of which seem to coincide
with the expected motional behaviors in the native state.
For example, the segments/residues (residues 29–31, 40, 60)
belonging to the loops between the native secondary struc-
tures (b1: 22–28; b2: 33–39; b3: 62–66; b4: 69–72; b5: 87–
93; a1: 45–55; and a2: 77–82), which should be more
flexible than other regions, have indeed significantly larger
than average R1 values, indicating larger nanosecond-
timescale motions; these residues are identified by solid
circles in the figure. However, this is not true for other
flexible regions of the native protein, especially at the long
N-terminal segment. These seem to suggest that just as one
can have native- and nonnative-type structural preferences,
there can also be native and nonnative type motional char-
acteristics in the denatured state.
The R2 data shown in Fig. 7 B also displays interesting
sequence-wise variations. In the denatured states, almost
every residue would have some contribution from conforma-
tional exchange (Rex) to its transverse relaxation rate,
R2 ¼ R2;int 1Rex;
where R2,int is the intrinsic transverse relaxation rate. How-
ever, there can be subtle variations in these due to possi-
bilities of differences in short-range interactions because of
sequence effects and consequent different local structural pref-
erences. Thus deviations from the average values, though small
as they would be, can be interpreted to suggest different de-
grees of conformational exchange contributions to the trans-
verse relaxation rates. In Fig. 7 B, among the many residues/
short stretches that have higher than average R2 (3.66 s�1)
values, we see that the stretches, 19–26 (average R2 is 4.17
s�1), and 89–92 (average R2 is 4.12 s�1), are at least four
residues long and coincide with the regions of structural
propensity derived from the secondary chemical shifts (this
data from Fig. 3 A is included in the figure here for ready
comparison). Hence, these regions may be considered to
execute slow segmental motions. The segment 19–26 over-
laps with the native b1 strand and the segment 89–92 over-
laps with the native b5 strand. Furthermore, the loop 29–31
between the b1 and b2 strands that exhibited high R1 value
also shows high R2 values. This indicates, firstly, that the
slow motions representing conformational exchange and the
nanosecond-scale motions of the N-H vectors are indepen-
dent. Secondly, since the b1 and b2 strands interact in the
native structure to form the antiparallel b-sheet, we are
tempted to think that the slow motions in this region help to
facilitate such an interaction in the denatured state as well.
However, as both the strands have some residues with PPII
propensities, the transient structure formation may not be
entirely native-like.
Among the residues that have lower than average R2
values signifying greater flexibilities, special mention must
be made of K37 and S61, which are in the interior of the
chain and have conspicuously low R2 (;2 s�1) values. Such
observations have been made in the past in at least two cases,
namely, apomyoglobin (36) and HIV-1 protease (2) and they
have been taken to indicate molecular hinges along the poly-
peptide chain. Interestingly, in SUMO-1 these residues are
seen to be near the edges of those areas that have exhibited
higher b-propensities along the chain, consolidating their
role as molecular hinges. Another interesting feature notice-
able in the R2 data in Fig. 7 B is that the N-terminal and the
C-terminal residues have distinctly different motional be-
haviors, the C-terminal residues having relatively greater
flexibility.
Implications to protein folding
As pointed out by several authors, the characteristics of the
denatured state have a significant influence in directing the
folding process of a protein (79–83). A denatured state under
some conditions may correspond to an intermediate along
the refolding pathway of a protein (1). The residual structural
propensities in the denatured state are suggestive of initial
FIGURE 7 15N relaxation rates plotted against sequence for SUMO-1 in
8 M urea at pH 5.6 and 27�C. (A) Longitudinal (R1) and (B) transverse (R2)
relaxation rates. The structural propensities seen in Fig. 3 A are also indi-
cated in B with solid bars for ready reference. Solid circles in A and B
identify residues with high flexibility.
NMR Characterization of Denatured SUMO-1 2505
Biophysical Journal 90(7) 2498–2509

folding events along the chain and its dynamic characteris-
tics throw further light on these events. There may be local
and nonlocal interactions and therefore, native and nonnative
propensities as well. Although the local interactions essen-
tially define intrinsic preferences, the nonlocal interactions
would define the topologies. The existence and significance
of nonlocal preferences under strong denaturing conditions,
however, remains a controversy (41,50). For those proteins,
where there is an initial collapse of the polypeptide chain
when refolding is initiated by dilution of denaturant, the
collapse is likely to be a consequence of transient interac-
tions between hydrophobic clusters that persist in the dena-
tured state (33,34,36). In the case of barnase, which exhibits
a fluctuating secondary structure in the denatured state, it is
suggested that folding is initiated around the native-like local
structures (38). In b-lactoglobulin, it was observed that the
early folding intermediate contained a nonnative helix and
this was attributed to the intrinsic high helical preference
of the amino acid residues involved in the denatured state
(40,84).
In SUMO-1 we observe that the protein chain in the urea-
denatured state has (f,c) preferences in the b-domain of the
Ramachandran map in many segments. Several of these are
native-type preferences. Many residues exhibit (f,c) pref-
erences corresponding to the PPII helix. However, there is no
indication of stable secondary structure in the denatured
ensemble. From the dynamics data it appears that in the urea-
denatured state the protein chain is not randomly fluctuating,
but exhibits sequence dependent motional variations. Inter-
estingly, the segment (residues 29–31) exhibiting larger nano-
second timescale motions coincides with the flexible loop
between b1 and b2 strands in the native structure. Although
this is not true for all the flexible regions of the native
structure, it seems to suggest the existence of some native-
type motional signature in the denatured state. The structural
and dynamics results provide extremely valuable clues to the
initial folding events in the protein. Although the structural
preferences indicate possible nucleation sites for the folding
process, the relaxation data indicate how motions can facil-
itate interactions between segments in directing the folding
process. Specifically, the slow conformational exchange ex-
hibited by the loop between b1 and b2 strands may promote
an interaction between the two strands; these are involved in
the formation of an antiparallel b-sheet in the native state.
However, few residues in each of these strands have PPII
propensities and thus the resultant interaction may not be
entirely native-like in the denatured state. Further, residues
K37 and S61, which exhibit greater flexibilities, are seen to
flank regions of higher structural propensities. Thus, they
may serve as molecular hinges for motions of the neighbor-
ing segments that may be transiently ordered in the en-
semble. All these lead to an implication that, possibly, the
conformational transitions and the hinged motions facilitate
short range structure formations, though transiently, in the
denatured state, and thus may serve as early folding events.
Parallel folding events from other regions such as b3, b5,
which also exhibit structural propensities and slow confor-
mational transitions, are also conceivable.
SUMO-1 belongs to the ubiquitin family of proteins, and
shares the same fold topology and the secondary structural
architecture, bbabbab. Thus, it would be interesting to
compare the above results with the folding studies on frag-
ments of ubiquitin reported in the recent past (85–87). It was
observed that the peptide, 1–17, representing the b1-loop-b2
hairpin in ubiquitin, formed a stable nativelike b-hairpin,
which suggested that the peptide segment has an intrinsic
tendency to form such a structure (86). On this basis, the
authors proposed that b1-loop-b2 hairpin formation may
be an initial folding event in the folding mechanism of
ubiquitin. Interestingly, the structural and dynamic prefer-
ences in the denatured state of full-length SUMO-1 protein
described in this work also indicate the above segment as the
folding initiation site. On the other hand, the experiments
with the C-terminal peptide (residues 36–76) of ubiquitin
exhibited nonnative a-helical preferences (87). In SUMO-1
we found some nonnative structural preferences in the broad
b-domain in the N-terminal (residues 2–6, 17–21), and in the
center (residues 48–53) along the protein chain. Thus it
appears that the initial folding events in both the proteins
have a combination of native- and nonnative-like events, but
the finer details have some events in common as well as
some differences. This may not be too surprising considering
that the two proteins have only 18% sequence homology and
consequently the interactions directing the folding process
could have differences.
CONCLUSIONS
In conclusion, we have successfully investigated here, using
a variety of NMR probes, the nature of the urea-denatured
state of SUMO-1, an important protein involved in post-
translational modifications. This became possible because
of the availability of new experimental protocols recently de-
veloped by us for obtaining resonance assignments in un-
folded proteins where the chemical shift dispersion of amide
protons and carbons is poor. Our data indicates that the pro-
tein in the urea-denatured state exhibits structural prefer-
ences in the broad b-domain of the Ramachandran map, over
the major length of the chain. Several residues populate the
(f,c) space corresponding to the PPII helix. The polypeptide
chain has significant motional restrictions and these show
sequence-dependent variations. As the structural propensi-
ties and the motional characteristics in the denatured state
have a major influence on the folding pathways of proteins,
the present observations suggest that, in SUMO-1, folding
from the urea-denatured state may get initiated around the
b1-loop–b2-region along the polypeptide chain. This region
has not only a contiguous stretch of b-propensities, but also
contains many residues exhibiting significant conformational
transitions. A few residues at the edges of these segments
2506 Kumar et al.
Biophysical Journal 90(7) 2498–2509

with structural propensities exhibit high flexibility and thus
may contribute to what one may call hinge motions facili-
tating segment movements and transient structure forma-
tions. Early folding events in a few other regions such as
b3, b5 strands are also conceivable, which would represent
parallel folding processes.
SUPPLEMENTARY MATERIAL
An online supplement to this article can be found by visiting
BJ Online at http://www.biophysj.org.
We thank the Government of India for providing financial support to the
National Facility for High Field NMR at the Tata Institute of Fundamental
Research.
REFERENCES
1. Religa, T. L., J. S. Markson, U. Mayor, S. M. Freund, and A. R. Fersht.2005. Solution structure of a protein denatured state and folding inter-mediate. Nature. 437:1053–1056.
2. Bhavesh, N. S., R. Sinha, P. M. Mohan, and R. V. Hosur. 2003. NMRelucidation of early folding hierarchy in HIV-1 protease. J. Biol. Chem.278:19980–19985.
3. Plotkin, S. S., and J. N. Onuchic. 2002. Understanding protein fold-ing with energy landscape theory. Part I: Basic concepts. Q. Rev.Biophys. 35:111–167.
4. Plotkin, S. S., and J. N. Onuchic. 2002. Understanding protein foldingwith energy landscape theory. Part II: Quantitative aspects. Q. Rev.Biophys. 35:205–286.
5. Chan, H. S., and K. A. Dill. 1998. Protein folding in the landscapeperspective: chevron plots and non-Arrhenius kinetics. Proteins. 30:2–33.
6. Dill, K. A., and H. S. Chan. 1997. From Levinthal to pathways to fun-nels. Nat. Struct. Biol. 4:10–19.
7. Onuchic, J. N., H. Nymeyer, A. E. Garcia, J. Chahine, and N. D. Socci.2000. The energy landscape theory of protein folding: insights intofolding mechanisms and scenarios. Adv. Protein Chem. 53:87–152.
8. Bhavesh, N. S., S. C. Panchal, R. Mittal, and R. V. Hosur. 2001. NMRidentification of local structural preferences in HIV-1 protease tetheredheterodimer in 6 M guanidine hydrochloride. FEBS Lett. 509:218–224.
9. Bhavesh, N. S., J. Juneja, J. B. Udgaonkar, and R. V. Hosur. 2004.Native and nonnative conformational preferences in the urea-unfoldedstate of barstar. Protein Sci. 13:3085–3091.
10. Chatterjee, A., P. Mridula, R. K. Mishra, R. Mittal, and R. V. Hosur.2005. Folding regulates autoprocessing of HIV-1 protease precursor.J. Biol. Chem. 280:11369–11378.
11. Schwalbe, H., K. M. Fiebig, M. Buck, J. A. Jones, S. B. Grimshaw, A.Spencer, S. J. Glaser, L. J. Smith, and C. M. Dobson. 1997. Structuraland dynamical properties of a denatured protein. Heteronuclear 3DNMR experiments and theoretical simulations of lysozyme in 8 M urea.Biochemistry. 36:8977–8991.
12. Shortle, D., and M. S. Ackerman. 2001. Persistence of native-liketopology in a denatured protein in 8 M urea. Science. 293:487–489.
13. Griko, Y., N. Sreerama, P. Osumi-Davis, R. W. Woody, and A. Y.Woody. 2001. Thermal and urea-induced unfolding in T7 RNApolymerase: calorimetry, circular dichroism and fluorescence study.Protein Sci. 10:845–853.
14. Li, Y., F. Picart, and D. P. Raleigh. 2005. Direct characterization of thefolded, unfolded and urea-denatured states of the C-terminal domain ofthe ribosomal protein L9. J. Mol. Biol. 349:839–846.
15. Zhang, X., Y. Xu, J. Zhang, J. Wu, and Y. Shi. 2005. Structural anddynamic characterization of the acid-unfolded state of hUBF HMG
Box 1 provides clues for the early events in protein folding.Biochemistry. 44:8117–8125.
16. Baum, J., C. M. Dobson, P. A. Evans, and C. Hanley. 1989. Char-acterization of a partly folded protein by NMR methods: studies onthe molten globule state of guinea pig a-lactalbumin. Biochemistry.28:7–13.
17. Eliezer, D., J. Yao, H. J. Dyson, and P. E. Wright. 1998. Structural anddynamic characterization of partially folded states of apomyoglobinand implications for protein folding. Nat. Struct. Biol. 5:148–155.
18. Matsuura, H., S. Shimotakahara, C. Sakuma, M. Tashiro, H. Shindo, K.Mochizuki, A. Yamagishi, M. Kojima, and K. Takahashi. 2004.Thermal unfolding of ribonuclease T1 studied by multi-dimensionalNMR spectroscopy. Biol. Chem. 385:1157–1164.
19. Feio, M. J., J. A. Navarro, M. S. Teixeira, D. Harrison, B. G. Karlsson,and M. A. De la Rosa. 2004. A thermal unfolding study of plastocyaninfrom the thermophilic cyanobacterium Phormidium laminosum. Bio-chemistry. 43:14784–14791.
20. Englander, S. W. 2000. Protein folding intermediates and pathwaysstudied by hydrogen exchange. Annu. Rev. Biophys. Biomol. Struct.29:213–238.
21. Onuchic, J. N., and P. G. Wolynes. 2004. Theory of protein folding.Curr. Opin. Struct. Biol. 14:70–75.
22. Otzen, D. E. 2002. Protein unfolding in detergents: effect of micellestructure, ionic strength, pH, and temperature. Biophys. J. 83:2219–2230.
23. Kamal, J. K., M. Nazeerunnisa, and D. V. Behere. 2002. Thermalunfolding of soybean peroxidase. Appropriate high denaturant con-centrations induce cooperativity allowing the correct measurement ofthermodynamic parameters. J. Biol. Chem. 277:40717–40721.
24. Kamal, J. K., and D. V. Behere. 2002. Thermal and conformationalstability of seed coat soybean peroxidase. Biochemistry. 41:9034–9042.
25. Baldwin, R. L. 1995. On-pathway versus off-pathway folding inter-mediates. Fold. Des. 1:R1–R8.
26. Bhuyan, A. K., and J. B. Udgaonkar. 1999. Observation of multistatekinetics during the slow folding and unfolding of barstar. Biochemistry.38:9158–9168.
27. Dunker, A. K., C. J. Brown, J. D. Lawson, L. M. Iakoucheva, and Z.Obradovic. 2002. Intrinsic disorder and protein function. Biochemistry.41:6573–6582.
28. Uversky, V. N. 2002. Natively unfolded proteins: a point wherebiology waits for physics. Protein Sci. 11:739–756.
29. Pohlschroder, M., K. Dilks, N. J. Hand, and R. W. Rose. 2004.Translocation of proteins across archaeal cytoplasmic membranes.FEMS Microbiol. Rev. 28:3.
30. Stefani, M. 2004. Protein misfolding and aggregation: new examples inmedicine and biology of the dark side of the protein world. Biochim.Biophys. Acta. 1739:5–25.
31. Ross, C. A., and M. A. Poirier. 2004. Protein aggregation and neuro-degenerative disease. Nat. Med. 10(Suppl):S10–S17.
32. Dobson, C. M. 2001. Protein folding and its links with human disease.Biochem. Soc. Symp. 1–26.
33. Hodsdon, M. E., and C. Frieden. 2001. Intestinal fatty acid bindingprotein: the folding mechanism as determined by NMR studies. Bio-chemistry. 40:732–742.
34. Lietzow, M. A., M. Jamin, H. J. Jane Dyson, and P. E. Wright. 2002.Mapping long-range contacts in a highly unfolded protein. J. Mol. Biol.322:655–662.
35. Neri, D., M. Billeter, G. Wider, and K. Wuthrich. 1992. NMRdetermination of residual structure in a urea-denatured protein, the 434-repressor. Science. 257:1559–1563.
36. Schwarzinger, S., P. E. Wright, and H. J. Dyson. 2002. Molecularhinges in protein folding: the urea-denatured state of apomyoglobin.Biochemistry. 41:12681–12686.
37. Arcus, V. L., S. Vuilleumier, S. M. Freund, M. Bycroft, and A. R.Fersht. 1995. A comparison of the pH, urea, and temperature-denatured
NMR Characterization of Denatured SUMO-1 2507
Biophysical Journal 90(7) 2498–2509

states of barnase by heteronuclear NMR: implications for the initiationof protein folding. J. Mol. Biol. 254:305–321.
38. Wong, K. B., J. Clarke, C. J. Bond, J. L. Neira, S. M. Freund, A. R.Fersht, and V. Daggett. 2000. Towards a complete description of thestructural and dynamic properties of the denatured state of barnase andthe role of residual structure in folding. J. Mol. Biol. 296:1257–1282.
39. Serrano, L., A. Matouschek, and A. R. Fersht. 1992. The folding of anenzyme. VI. The folding pathway of barnase: comparison with theo-retical models. J. Mol. Biol. 224:847–859.
40. Hamada, D., and Y. Goto. 1997. The equilibrium intermediate ofb-lactoglobulin with non-native a-helical structure. J. Mol. Biol. 269:479–487.
41. Mohana-Borges, R., N. K. Goto, G. J. Kroon, H. J. Dyson, and P. E.Wright. 2004. Structural characterization of unfolded states of apomyo-globin using residual dipolar couplings. J. Mol. Biol. 340:1131–1142.
42. Crowhurst, K. A., M. Tollinger, and J. D. Forman-Kay. 2002.Cooperative interactions and a non-native buried Trp in the unfoldedstate of an SH3 domain. J. Mol. Biol. 322:163–178.
43. Crowhurst, K. A., and J. D. Forman-Kay. 2003. Aromatic and methylNOEs highlight hydrophobic clustering in the unfolded state of an SH3domain. Biochemistry. 42:8687–8695.
44. Mok, K. H., and K. H. Han. 1999. NMR solution conformation of anantitoxic analogue of a-conotoxin GI: identification of a commonnicotinic acetylcholine receptor a1-subunit binding surface for smallligands and a-conotoxins. Biochemistry. 38:11895–11904.
45. Parrot, I., P. C. Huang, and C. Khosla. 2002. Circular dichroism andnuclear magnetic resonance spectroscopic analysis of immunogenicgluten peptides and their analogs. J. Biol. Chem. 277:45572–45578.
46. Tremmel, P., and A. Geyer. 2002. An oligomeric Ser-Pro dipeptidemimetic assuming the polyproline II helix conformation. J. Am. Chem.Soc. 124:8548–8549.
47. Kelly, M. A., B. W. Chellgren, A. L. Rucker, J. M. Troutman, M. G.Fried, A. F. Miller, and T. P. Creamer. 2001. Host-guest study of left-handed polyproline II helix formation. Biochemistry. 40:14376–14383.
48. Moyna, G., H. J. Williams, R. J. Nachman, and A. I. Scott. 1999.Detection of nascent polyproline II helices in solution by NMR insynthetic insect kinin neuropeptide mimics containing the X-Pro-Pro-Xmotif. J. Pept. Res. 53:294–301.
49. Shi, Z., R. W. Woody, and N. R. Kallenbach. 2002. Is polyproline II amajor backbone conformation in unfolded proteins? Adv. ProteinChem. 62:163–240.
50. Jha, A. K., A. Colubri, K. F. Freed, and T. R. Sosnick. 2005. Statisticalcoil model of the unfolded state: resolving the reconciliation problem.Proc. Natl. Acad. Sci. USA. 102:13099–13104.
51. Mahajan, R., C. Delphin, T. Guan, L. Gerace, and F. Melchior. 1997. Asmall ubiquitin-related polypeptide involved in targeting RanGAP1 tonuclear pore complex protein RanBP2. Cell. 88:97–107.
52. Bayer, P., A. Arndt, S. Metzger, R. Mahajan, F. Melchior, R. Jaenicke,and J. Becker. 1998. Structure determination of the small ubiquitin-related modifier SUMO-1. J. Mol. Biol. 280:275–286.
53. Jin, C., T. Shiyanova, Z. Shen, and X. Liao. 2001. Heteronuclearnuclear magnetic resonance assignments, structure and dynamics ofSUMO-1, a human ubiquitin-like protein. Int. J. Biol. Macromol. 28:227–234.
54. Mishra, R. K., S. S. Jatiani, A. Kumar, V. R. Simhadri, R. V. Hosur,and R. Mittal. 2004. Dynamin interacts with members of the sumoy-lation machinery. J. Biol. Chem. 279:31445–31454.
55. Bhavesh, N. S., S. C. Panchal, and R. V. Hosur. 2001. An efficienthigh-throughput resonance assignment procedure for structuralgenomics and protein folding research by NMR. Biochemistry. 40:14727–14735.
56. Chatterjee, A., N. S. Bhavesh, S. C. Panchal, and R. V. Hosur. 2002.A novel protocol based on HN(C)N for rapid resonance assignmentin (15N, 13C) labeled proteins: implications to structural genomics.Biochem. Biophys. Res. Commun. 293:427–432.
57. Panchal, S. C., N. S. Bhavesh, and R. V. Hosur. 2001. Improved 3Dtriple resonance experiments, HNN and HN(C)N, for HN and 15Nsequential correlations in (13C, 15N) labeled proteins: application tounfolded proteins. J. Biomol. NMR. 20:135–147.
58. Permi, P., and A. Annila. 2004. Coherence transfer in proteins. Prog.Nucl. Magn. Reson. Spectrosc. 44:97–137.
59. Tugarinov, V., P. M. Hwang, and L. E. Kay. 2004. Nuclear magneticresonance spectroscopy of high-molecular-weight proteins. Annu. Rev.Biochem. 73:107–146.
60. Farrow, N. A., R. Muhandiram, A. U. Singer, S. M. Pascal, C. M. Kay,G. Gish, S. E. Shoelson, T. Pawson, J. D. Forman-Kay, and L. E. Kay.1994. Backbone dynamics of a free and phosphopeptide-complexedSrc homology 2 domain studied by 15N NMR relaxation. Biochemistry.33:5984–6003.
61. Dyson, H. J., and P. E. Wright. 2004. Unfolded proteins and proteinfolding studied by NMR. Chem. Rev. 104:3607–3622.
62. Barbar, E. 1999. NMR characterization of partially folded and unfoldedconformational ensembles of proteins. Biopolymers. 51:191–207.
63. Dyson, H. J., and P. E. Wright. 2001. Nuclear magnetic resonancemethods for elucidation of structure and dynamics in disordered states.Methods Enzymol. 339:258–270.
64. Dyson, H. J., and P. E. Wright. 2002. Insights into the structure anddynamics of unfolded proteins from nuclear magnetic resonance. Adv.Protein Chem. 62:311–340.
65. Bhavesh, N. S., and R. V. Hosur. 2004. Exploring unstructuredproteins. Proc. Indian Natl. Sci. Acad. 70A:579–596.
66. Chatterjee, A., A. Kumar, J. Chugh, S. Srivastava, N. S. Bhavesh, andR. V. Hosur. 2005. NMR of unfolded proteins. J. Chem. Sci. (IndianAcad. Sci.). 117:3–21.
67. Wishart, D. S., C. G. Bigam, A. Holm, R. S. Hodges, and B. D. Sykes.1995. 1H, 13C and 15N random coil NMR chemical shifts of thecommon amino acids. I. Investigations of nearest-neighbor effects.J. Biomol. NMR. 5:67–81.
68. Wishart, D. S., and B. D. Sykes. 1994. Chemical shifts as a tool forstructure determination. Methods Enzymol. 239:363–392.
69. Schwarzinger, S., G. J. Kroon, T. R. Foss, P. E. Wright, and H. J.Dyson. 2000. Random coil chemical shifts in acidic 8 M urea:implementation of random coil shift data in NMRView. J. Biomol.NMR. 18:43–48.
70. Schwarzinger, S., G. J. Kroon, T. R. Foss, J. Chung, P. E. Wright, andH. J. Dyson. 2001. Sequence-dependent correction of random coilNMR chemical shifts. J. Am. Chem. Soc. 123:2970–2978.
71. Penkett, C. J., C. Redfield, I. Dodd, J. Hubbard, D. L. McBay, D. E.Mossakowska, R. A. Smith, C. M. Dobson, and L. J. Smith. 1997. NMRanalysis of main-chain conformational preferences in an unfoldedfibronectin-binding protein. J. Mol. Biol. 274:152–159.
72. Baxter, N. J., and M. P. Williamson. 1997. Temperature dependence of1H chemical shifts in proteins. J. Biomol. NMR. 9:359–369.
73. Merutka, G., H. J. Dyson, and P. E. Wright. 1995. ‘‘Random coil’’ 1Hchemical shifts obtained as a function of temperature and trifluoroethanolconcentration for the peptide series GGXGG. J. Biomol. NMR. 5:14–24.
74. Wuthrich, K. 1986. NMR of Proteins and Nucleic Acids. John Wiley &Sons.
75. Kay, L. E. 2005. NMR studies of protein structure and dynamics.J. Magn. Reson. 173:193–207.
76. Palmer III, A. G. 1993. Dynamic properties of proteins from NMRspectroscopy. Curr. Opin. Biotechnol. 4:385–391.
77. Palmer III, A. G. 1997. Probing molecular motion by NMR. Curr.Opin. Struct. Biol. 7:732–737.
78. Palmer III, A. G. 2001. NMR probes of molecular dynamics: overviewand comparison with other techniques. Annu. Rev. Biophys. Biomol.Struct. 30:129–155.
79. van Gunsteren, W. F., R. Burgi, C. Peter, and X. Daura. 2001. The keyto solving the protein-folding problem lies in an accurate description ofthe denatured state. Angew. Chem. Int. Ed. Engl. 40:351–355.
2508 Kumar et al.
Biophysical Journal 90(7) 2498–2509

80. Wintrode, P. L., T. Rojsajjakul, R. Vadrevu, C. R. Matthews, and D. L.Smith. 2005. An obligatory intermediate controls the folding of thea-subunit of tryptophan synthase, a TIM barrel protein. J. Mol. Biol.347:911–919.
81. Daggett, V., and A. R. Fersht. 2003. Is there a unifying mechanism forprotein folding? Trends Biochem. Sci. 28:18–25.
82. Daggett, V., A. Li, L. S. Itzhaki, D. E. Otzen, and A. R. Fersht. 1996.Structure of the transition state for folding of a protein derived fromexperiment and simulation. J. Mol. Biol. 257:430–440.
83. Gianni, S., N. R. Guydosh, F. Khan, T. D. Caldas, U. Mayor, G. W.White, M. L. DeMarco, V. Daggett, and A. R. Fersht. 2003. Unifyingfeatures in protein-folding mechanisms. Proc. Natl. Acad. Sci. USA.100:13286–13291.
84. Kuwata, K., R. Shastry, H. Cheng, M. Hoshino, C. A. Batt, Y. Goto,and H. Roder. 2001. Structural and kinetic characterization of earlyfolding events in b-lactoglobulin. Nat. Struct. Biol. 8:151–155.
85. Zerella, R., P. A. Evans, J. M. Ionides, L. C. Packman, B. W. Trotter,J. P. Mackay, and D. H. Williams. 1999. Autonomous folding of apeptide corresponding to the N-terminal b-hairpin from ubiquitin.Protein Sci. 8:1320–1331.
86. Zerella, R., P. Y. Chen, P. A. Evans, A. Raine, and D. H. Williams.2000. Structural characterization of a mutant peptide derived fromubiquitin: implications for protein folding. Protein Sci. 9:2142–2150.
87. Jourdan, M., and M. S. Searle. 2000. Cooperative assembly of a nativelikeubiquitin structure through peptide fragment complexation: energetics ofpeptide association and folding. Biochemistry. 39:12355–12364.
NMR Characterization of Denatured SUMO-1 2509
Biophysical Journal 90(7) 2498–2509




















