
Livro do curso - Projeto Dinâmica Biológica de Fragmentos ...
345
Ecologia da Floresta Amazônica 2012 Livro do curso Foto: Lucas Navarro Paolucci
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Livro do curso - Projeto Dinâmica Biológica de Fragmentos ...

Ecologia da Floresta Amazônica 2012
Livro do curso
Foto: Lucas Navarro Paolucci

2
Coordenadores e monitores
Logística
Ary Jorge Correa Ferreira
Apoio logístico
Maria Rosely Cavalcante Hipólito
Jéssica Iara Corísco da Silva
Alexandra Maria Ferreira Silveira
Ayezha Oliveira Hidalgo
Cozinheiros
Maria Eduarda da Costa Benício
Raimundo José Maria de Souza
Motoristas
Luiz Raimundo de Queiroz
Antônio José Moraes Pereira
Paulo Estefano D.
Bobrowiec
Thiago G. Kloss
José Luis C.
Camargo
Paulo Enrique
C. Peixoto Marcos C. Vieira

3

4

5
Índice
Floresta fragmentada - Dimona
Projetos orientados
A abundância e riqueza de plântulas e sementes abaixo de abrigos utilizados por
morcegos frugívoros é maior?
Joselândio Corrêa Santos, Ana Carolina Vieira Pires, José Hidasi Neto, Sarah Freitas Magalhães Silva...........................................13
A cobertura foliar por liquens e briófitas produz custos para Duguetia cf. flagellaris
(Annonaceae)?
Daniel Passos, Bianca Silva, Cibele Paiva & Vidal Carrascosa...........................................................................................................22
A riqueza de plantas aumenta a riqueza de herbívoros, mas não dos predadores
Renata Pimentel Rocha, Caroline Marques Dracxler, Leonardo Oliveira, Bruno Travassos de Britto................................................28
Espécies de animais aquáticos e semiaquáticos se distribuem entre poças temporárias
independentemente da qualidade do habitat
Mariana Velasque, Ana Claudia Rorato, Ludmila Rattis, Randolpho Gonçalves Dias Terceiro.........................................................35
O tamanho da domácia foliar determina sua ocupação por ácaros?
Tatiane Gomes Calaça Menezes, Daniela Pinto Coelho, Lorena Costa Pinto e Artur Alcantara Madeira..............,............................43
Projetos livres
Alocação diferencial de recurso em Vismia guianensis (Clusiaceae) associada a
condições de luminosidade
Randolpho Gonçalves Dias Terceiro, José Hidasi Neto, Sarah Freitas, Lorena Costa Pinto...............................................................49

6
Distribuição espacial em Selaginella amazonica (Selaginellaceae) em função da
umidade
Leonardo Queiroz de Oliveira, Bianca Ferreira da Silva, Mariana Velasque & Joselândio Corrêa Santos.........................................51
O investimento em reprodução não diminui o investimento em defesa em Miconia
dispar (Melastomataceae)
Daniela Coelho, Ludmila Rattis, Tatiane Menezes & Cibele Paiva.....................................................................................................52
Por que o hemíptero Jalisus ossae (Reduviidae) não desencadeia resposta defensiva da
formiga Pheidole minutula em Maieta poeppigii (Melastomataceae)?
Ana Carolina Vieira Pires, Ana Claudia Rorato Vitor, Caroline Marques Dracxler e Daniel Passos..................................................53
Recrutamento de formigas na mirmecófita Hirtella sp.: estudo em plantas com
diferentes níveis de estresse
Bruno Travassos, Artur Madeira, Vidal Carrascosa, Renata Rocha.....................................................................................................55
Várzea
Projetos orientados
A seleção de habitats por cupins em áreas alagadas não visa redução dos custos de
deslocamento até o solo
Sarah Freitas Magalhães, Caroline Marques Dracxler, Renata Pimentel Rocha e Randolpho Dias Terceiro......................................56
O crescimento vertical da macrófita Eichhornia crassipes (Pontederiaceae) é maior em
ambientes com menor disponibilidade de água
José Hidasi, Artur Madeira, Cibele Paiva e Daniel Passos...................................................................................................................62

7
Ocorrência de grupos funcionais de peixes em dois rios Amazônicos com diferentes
transparências de água
Lorena Pinto, Ludmila Rattis, Tatiane Calaça e Bruno Travassos.................................................................................................67
Plantas não sincronizam a produção de frutos de acordo com a vazante em uma
comunidade de várzea no Rio Solimões
Ana Claudia Rorato Vitor, Leonardo Oliveira, Vidal Carrascosa, Mariana Velasque.........................................................................75
Respostas comportamentais em presas de ninfas de Odonata (Subordem: Anisoptera)
Bianca Silva, Joselândio Santos, Ana Carolina Vieira & Daniela Coelho...........................................................................................82
Projetos livres
A herbivoria em Eichhornia crassipes (Pontederiaceae) não depende do estresse hídrico
Cibele Paiva, Lorena Pinto, Mariana Velasque, Ana Carolina Vieira Pires, Leonardo Oliveira.........................................................88
A transparência da água como filtro ambiental para a pigmentação de espécies de peixes
em rios amazônicos
Ludmila Rattis, Daniel Passos, Daniela Coelho, Joselândio Santos e Renata Rocha........................................................................89
Complexidade estrutural do habitat influenciando o padrão de frequência relativa de
espécies em comunidades de formigas
Vidal Carrascosa, Randolpho Dias Terceiro, Bruno Travassos, José Hidasi & Bianca Ferreira..........................................................90
Plantas sujeitas ao alagamento sazonal possuem menor variação no tamanho das folhas
Caroline Marques Dracxler, Artur Alcântara Madeira, Tatiane Gomes Calaça Menezes, Sarah Freitas Magalhães e Ana Claúdia
Rorato....................................................................................................................................................................................................92

8
Igapó – Anavilhanas
Projetos orientados
A coloração alar é um indicativo da qualidade dos machos em Diastatops sp. (Odonata:
Libelullidae)
Daniela P. Coelho, Leonardo Oliveira, José Hidasi e Joselândio C. Santos.........................................................................................93
A escolha do local de oviposição de um lepidóptero é influenciada pela disponibilidade
de recurso
Vidal Carrascosa, Sarah Freitas Magalhães, Renata Pimentel Rocha e Tatiane Gomes Calaça Menezes........................................101
Diferentes períodos de inundação influenciam resistência de folhas em plantas no igapó
Bruno Travassos, Ana Claudia Rorato, Mariana Velasque e Ana Carolina Vieira Pires..................................................................107
Formigas que planam, nadam? Relação entre queda direcional e capacidade de natação
em três espécies de formigas de uma área alagável na Amazônia Central
Lorena Pinto, Caroline Marques Dracxler, Randolpho Dias Terceiro & Cibele Paiva.....................................................................114
O tempo de submersão de plântulas de Dalbergia sp. gera um investimento diferencial
entre os sistemas aéreo e radicular?
Artur Madeira, Bianca Silva, Daniel Passos, Ludmila Rattis............................................................................................................122
Projetos livres
A borboleta Glutophrissa drusilla (Pieridae) prefere cruzar rios mais largos entre as
manchas florestais
Mariana Velasque, Ana Carolina Vieira Pires, Tatiane G. C. Menezes, Renata Pimentel Rocha e Ludmila Rattis.........................128

9
O tamanho das pernas de formigas em áreas alagáveis influencia sua capacidade de
deslocamento na água?
Caroline Marques Dracxler, Lorena Costa Pinto, Vidal Carrascosa, Daniela Pinto Coelho e Sarah Freitas Magalhães..................130
Pernas para que te quero? Função das pernas na natação da formiga Camponotus sp.
(Formicidae)
Randolpho Dias-Terceiro, Artur Madeira, Daniel Passos, Cibele Paiva & Leonardo Oliveira.........................................................131
Variação da capacidade de armazenamento de água na planta Psychotria sp. mediada
por filtro ambiental
Ana Claudia Rorato, Joselândio Santos, Bianca Ferreira, José Hidasi Neto & Bruno Travassos.....................................................132
Floresta contínua – Km41
Projetos orientados
A aranha Faiditus subflavus (Theridiidae) selecionam indivíduos de Maieta guianensis
(Melastomataceae) com menor dano foliar?
Leonardo Oliveira, Renata Rocha, José Hidasi e Ana Carolina Vieira Pires.....................................................................................134
Importância relativa do espaço e do ambiente na explicação da diversidade beta: um
teste da Teoria do Nicho x Teoria Neutra em pequena escala
Daniela Coelho, Bruno Travassos, Randolpho Dias-Terceiro e Cibele Paiva...................................................................................140
Interação formiga-planta: a abundância da planta não afeta o número de formigas
mutualistas com que ela interage
Sarah Freitas Magalhães Silva, Ana Claudia Rorato, Artur Alcântara Madeira e Mariana Velasque...............................................147

10
Onde os similares também têm vez: árvores com morfologia foliar semelhante
coexistem em uma floresta secundária
Caroline Marques Dracxler, Joselândio Corrêa Santos, Ludmila Rattis e Bianca Ferreira da Silva.................................................153
Plantas com folhas menores impedem o estabelecimento de minas e galhas
Daniel Passos, Tatiane Calaça, Lorena Pinto & Vidal Carrascosa....................................................................................................159
Projetos individuais
Diferença no comportamento de forrageio de formigas associadas a nectários
extraflorais e exsudato de afídeos
Ana Carolina Vieira Pires..................................................................................................................................................................165
Convergência de características funcionais foliares entre uma floresta em regeneração e
uma floresta primária na Amazônia Central
Ana Claudia Rorato............................................................................................................................................................................177
Distribuição de galhas em folhas de Vismia japurensis (Clusiaceae): existe uma seleção
do local de oviposição?
Artur Alcantara Madeira....................................................................................................................................................................189
Quando uma ajuda é bem-vinda: briófitas facilitam o estabelecimento de plantas sobre
troncos caídos
Bianca Ferreira da Silva.....................................................................................................................................................................197
As formigas não utilizam sinal químico de formigas outros territórios para melhorar sua
eficiência de forrageio
Bruno Travassos.................................................................................................................................................................................205

11
Agregação de palmeiras não aumenta o consumo por herbívoros em plântulas de Attalea
sp.
Caroline Marques Dracxler................................................................................................................................................................213
Interação mutualista entre afídeos e formigas não altera a eficácia de defesa da
mirmecófita Tococa sp. por formigas
Cibele Paiva........................................................................................................................................................................................223
Pequenos camuflados e grandes esquivos? O tamanho corpóreo não atua na seleção de
habitat em peixes de riachos amazônicos
Daniel Passos......................................................................................................................................................................................230
A herbivoria em mirmecófitas afetando a resposta da colônia de formigas
Daniela Pinto Coelho.........................................................................................................................................................................239
Habitats com maior luminosidade não representam melhores territórios de acasalamento
para machos da libélula Chalcopteryx scintilans (Odonata: Polythoridae)
José Hidasi Neto.................................................................................................................................................................................248
Mutualismo de defesa entre plantas mirmecófitas e formigas reduz o investimento em
defesas estruturais em espécies de Melastomataceae
Joselândio Corrêa Santos...................................................................................................................................................................257
A condição alimentar das larvas de formigas-leão influencia no tamanho e no tempo de
construção do funil de areia?
Leonardo Queiroz de Oliveira............................................................................................................................................................267

12
Relação entre tamanho corporal, respostas de defesa e posição na teia de indivíduos de
uma espécie de aranha (Pholcidae) que vive em agregações na Amazônia Central
Lorena Costa Pinto.............................................................................................................................................................................275
A influência da agregação espacial de Maieta guianensis (Melastomataceae) sobre a
intensidade da herbivoria
Ludmila Rattis....................................................................................................................................................................................287
Tricomas foliares auxiliam formigas na detecção de herbívoros em plantas mirmecófita?
Mariana Velasque...............................................................................................................................................................................298
Folha consumida é menos querida? Seleção de habitat de Aranhas Faiditus subflavus
(Theridiidae) por folhas de Maieta guianensis (Melastomataceae)
Randolpho Gonçalves Dias Terceiro..................................................................................................................................................306
Quanto menor o guerreiro, maior a armadura: defesa estrutural em plantas com
diferentes tamanhos foliares
Renata Pimentel Rocha......................................................................................................................................................................313
A resposta de captura em uma aranha orbitela aumenta com a redução do valor da presa
Sarah Freitas Magalhães Silva...........................................................................................................................................................320
Árvores grandes limitam o tamanho de suas vizinhas?
Tatiane Gomes Calaça Menezes........................................................................................................................................................327
Qual é a influência da qualidade do macho da espécie Chalcopteryx scintilans (Odonata:
Polythoridae) no tempo de luta por um território?
Vidal C. Carrascosa............................................................................................................................................................................336

13
Dimona – projetos orientados
A abundância e riqueza de plântulas e sementes abaixo de abrigos utilizados por
morcegos frugívoros é maior?
Joselândio Corrêa Santos, Ana Carolina Vieira Pires, José Hidasi Neto, Sarah Freitas
Magalhães Silva
Introdução
A dispersão de sementes promove o aumento e manutenção da diversidade de plantas,
aumentando a chance de colonização de novos ambientes e a ampliação da distribuição
das espécies. Assim, o processo de dispersão de sementes está relacionado com o
padrão de distribuição das espécies vegetais.
As plantas de ambientes tropicais possuem uma clara associação com grupos de
dispersores animais (Jordano 2000). Em muitos casos, as plantas que alocam nutrientes
de alto valor em seus frutos atraem potenciais animais dispersores. Além disso, o
tamanho do fruto está associado com a quantidade de reservas ou tecidos nutritivos que
ele contém, ou seja, frutos maiores possuem potencialmente mais reservas que frutos
pequenos. Consequentemente, isso estaria determinando a escolha dos frutos por
animais, que escolheriam frutos tão grandes quanto pudessem manipular.
Nos trópicos, os principais dispersores dos propágulos das plantas são os
vertebrados, em especial aves e morcegos (Galindo-Gonzalez 2000; Mendellin & Gaona
1999). Esses animais são conhecidos por manipular e ingerir pequenos frutos e liberar
as sementes em suas fezes (Galindo-Gonzalez 1998). Em geral, o consumo dos frutos é
feito ainda na planta em frutificação, entretanto algumas espécies possuem locais

14
preferenciais para a manipulação do alimento, que podem ser considerados como sítios
de alimentação ou poleiros. Assim, haverá locais onde a deposição de sementes e,
consequentemente, também de plântulas, serão diferenciadas de locais relativamente
próximos, devido a chuvas de sementes adicionais provenientes dos restos da
alimentação e das fezes dos animais (Melo 2009).
Algumas espécies frugívoras de morcegos, como as do gênero Carolia,
constroem tendas em copas de árvores e arbustos do sub-bosque das florestas (Melo
2009). Essas tendas são confeccionadas a partir da modificação da disposição da lâmina
foliar por mordidas dos morcegos no pecíolo ou nas regiões próximas à nervura foliar,
conforme o tipo de folha (Stoner 2000). Pouco se sabe sobre o objetivo da construção
dessas estruturas, mas acredita-se que sejam locais de descanso, acasalamento, paradas
estratégicas para manipulação de frutos durante o forrageio ou mesmo abrigos para
situações adversas, tais como chuvas, risco eminente de predação, entre outros. A única
certeza que se tem é que são bastante recorrentes nas florestas tropicais úmidas e que
são ativamente construídas por morcegos.
Considerando que os morcegos frugívoros constroem abrigos para utilizar como
locais para alimentação, o objetivo desse trabalho foi conhecer qual o papel dos
morcegos construtores de abrigo sobre a dispersão de sementes e plântulas em uma
floresta tropical úmida. Nossa previsão é que existe um aumento no número de
indivíduos e morfoespécies de sementes e plântulas em locais abaixo dos abrigos,
quando comparado aos locais sem o abrigo, pois os morcegos que utilizam o abrigo
devem promover um incremento na chuva de sementes.

15
Material e Métodos
Área de estudo
Nosso estudo foi realizado na Fazenda Dimona (2º 20’S; 60º 06’O), uma das áreas do
Projeto Dinâmica Biológica de Fragmentos Florestais (PDBFF), situada a 80 km da
cidade de Manaus, Brasil. A área é caracterizada por fragmentos florestais de terra firme
de tamanhos diferentes, circundados por uma área de floresta secundária, com
aproximadamente 30 anos de sucessão. As áreas de florestas secundárias são originadas
a partir das áreas desmatadas para o estabelecimento de pastagens, que após alguns anos
de uso foram abandonadas.
Busca por abrigos, riqueza e abundância de sementes e plântulas
Nós percorremos uma trilha de 2,6 km fazendo uma varredura de três metros para cada
lado a partir do ponto central, cobrindo uma área de 1,56 ha. Durante o nosso
deslocamento fizemos uma busca ativa pelos abrigos de morcegos. Esses abrigos são
conspícuos no ambiente devido a sua aparência peculiar, um aglomerado de folhas
sobrepostas, com forma cônica. Em geral eles são formados por folhas simples e
grandes em árvores com alturas de 0,4 a 4 m. Classificamos as plantas usadas como
abrigo em morfotipos. Abaixo de cada abrigo colocamos uma parcela de 1 m², tomando
como ponto central o caule da planta com o abrigo. A dois metros desta primeira parcela
foi estabelecida uma parcela controle, sempre em direção oposta à inclinação do terreno,
para evitar amostrar sementes que pudessem ter rolado da área abaixo do abrigo.
Dentro de cada parcela coletamos todas as plântulas com altura máxima de 50
cm e todas as sementes encontradas. Em seguida, levamos essas amostras ao laboratório
e classificamos em morfotipos, baseados nas características morfológicas.

16
Análises estatísticas
Para testarmos se a dispersão feita por morcegos sob os abrigos aumentou a riqueza e a
abundância de plântulas e sementes, realizamos testes-t pareados considerando o
número de indivíduos e de morfoespécies das plântulas e sementes como variáveis
dependentes, e os locais, abrigo e controle, como as variáveis explicativas.
Resultados
Nós encontramos 10 plantas usadas como abrigo por morcegos que pertenciam à pelo
menos quatro morfoespécies. Os abrigos estavam em alturas que variaram entre 1,5 e
3,8 m e se encontravam em três hábitats distintos: dois indivíduos na capoeira, um
indivíduo na borda entre o fragmento florestal e a matriz de capoeira e sete indivíduos
no interior do fragmento florestal. Encontramos nove morfoespécies de sementes e 50
de plântulas. O número total de morfotipos encontrados para sementes e plântulas foi de
14 e 131, respectivamente (Tabela 1). Para plântulas, 47% (n=50) das espécies foram
exclusivas dos locais com abrigos. De modo geral, encontramos um pequeno número de
sementes em ambos os ambientes, sendo que somente 35% (n=7) das parcelas tiveram
pelo menos uma semente.

17
Tabela 1: Número de morfoespécies (média ± desvio padrão) e de indivíduos (média ±
desvio padrão) para sementes e plântulas localizadas abaixo e fora dos abrigos usados
por morcegos em uma área da Amazônia Central.
Nº de espécies Nº de indivíduos
Sementes
Abrigo 4 (0,7 ± 1,23) 6 (0,55 ± 1,5)
Controle 6 (0,42 ±1,72) 8 (0,73 ± 1,1)
Total 9 (0,81 ± 1,94) 14 (1,2 ± 1,95)
Plântulas
Abrigo 35 (4,2 ± 2,28) 61 (7 ± 6,54)
Controle 29 (5,9 ± 2,43) 70 (4,8 ± 1,67)
Total 50 (7.81 ± 3,55) 131 (11,9 ± 8,35)
O número de indivíduos de plântulas não diferiu entre os locais abaixo e fora do
abrigo (t=0,83; gl=9; p=0,42), o mesmo foi observado para o número de morfoespécies
(t=0,62; gl=9; p=0,54).
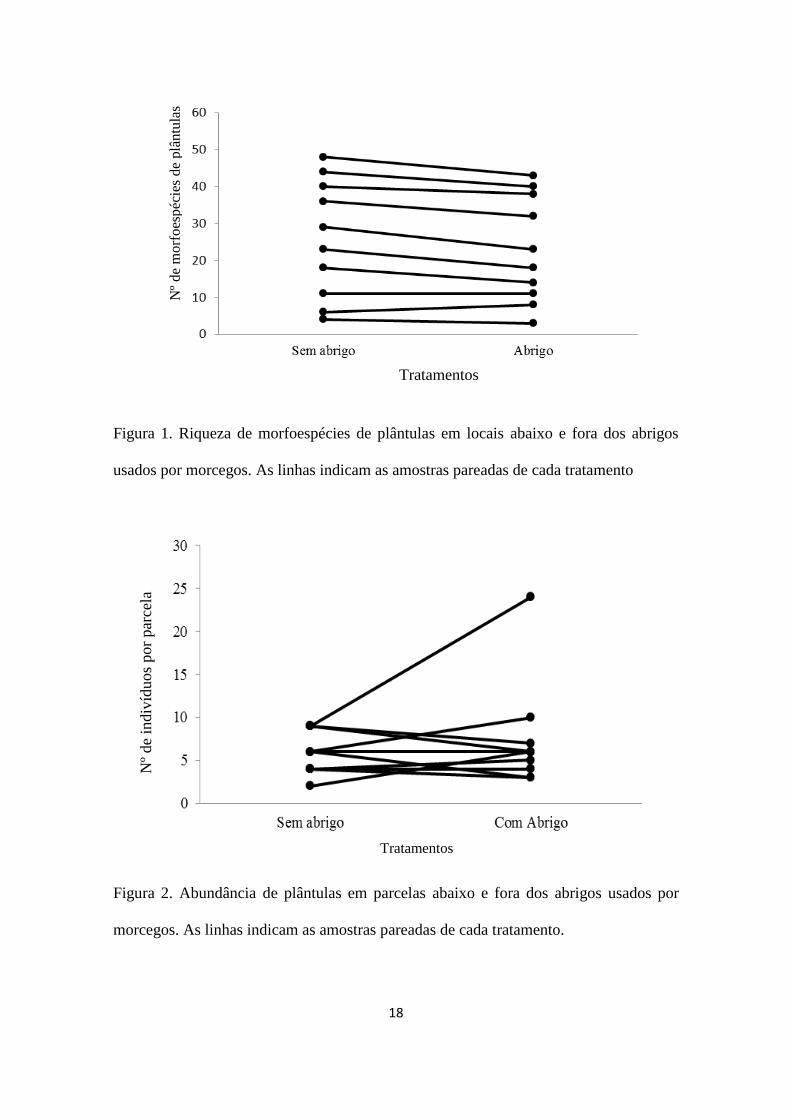
18
Figura 1. Riqueza de morfoespécies de plântulas em locais abaixo e fora dos abrigos
usados por morcegos. As linhas indicam as amostras pareadas de cada tratamento
Figura 2. Abundância de plântulas em parcelas abaixo e fora dos abrigos usados por
morcegos. As linhas indicam as amostras pareadas de cada tratamento.
Tratamentos
Tratamentos
Nº
de
mo
rfo
esp
écie
s d
e plâ
ntu
las
Nº
de
indiv
íduos
por
par
cela

19
Discussão
Apesar de morcegos agirem como dispersores de sementes (Mendellín & Gaona 1999),
nós não encontramos evidências de que os abrigos podem aumentar a riqueza e
abundância de plântulas. Portanto, morcegos não atuam como uma espécie capaz de
aumentar a diversidade de plantas abaixo dos locais em que eles se abrigam. Como
morcegos carregam alimento para o abrigo e defecam nesses locais (Galindo-Gonzalez
1998), eventualmente as sementes liberadas pelos morcegos são predadas ou sofrem
dispersão secundária. Por outro lado, o fato de 47% das plântulas terem ocorrido
exclusivamente abaixo dos abrigos, indica que apesar dos morcegos não aumentarem a
diversidade local, eles trazem espécies oriundas de outras áreas.
A dispersão secundária das sementes poderia ser efetuada por espécies de
besouros. Muitas espécies de coleópteros da família Scarabaeidae são conhecidas por
utilizar fezes de mamíferos como sítios de oviposição (Andersen 2001, Andersen 2002).
Um comportamento comum desses besouros é deslocar porções de fezes para longe dos
locais originais de deposição. Nesse sentido, é possível que existam besouros associados
as fezes abaixo do abrigo que sejam responsáveis pela remoção das sementes. Além dos
besouros, formigas também podem estar associadas à dispersão secundária. As formigas
podem levar as sementes para longe ou, ainda, podem predar sementes provenientes da
dispersão realizada por vertebrados (Passos & Oliveira, 2002). Em ambos os casos, não
haveria uma acumulação de sementes e plântulas sob locais usados como abrigos por
morcegos.
Embora a maioria dos estudos tem dado destaque à dispersão realizada por
morcegos, aparentemente essa importância está associada à regeneração de florestas em
sucessão (Melo 2009). Como nosso estudo foi realizado predominantemente em uma
floresta madura, pode ser que o efeito da dispersão secundária seja maior que a

20
dispersão realizada por morcegos. Assim, o aumento na diversidade de plantas
decorrente do uso de abrigos por morcegos pode ser dependente da idade da floresta.
Agradecimentos
Agradecemos aos professores orientadores, Leão do Norte (Felipe Melo) e Paulo Mau
Estefano, ao Thiago Xexéu Kloss pelas revisões e ao professor Paulo Rainbow Master
Enrique pelo auxílio na morfotipagem das plântulas. Aos professores, colegas e
monitores pelas observações relevantes levantadas nas discussões. Aos colaboradores
do curso EFA, D. Eduarda, Sr. Jorge e Sr. Léo.
Referências
Andersen, E. 2001. Effects of dung presence, dung amount and secondary dispersal by
dung beetles on the fate of Micropholis guyanensis (Sapotaceae) seeds in Central
Amazonia. Journal of Tropical Ecology, 17:61-78.
Andersen, E. 2002. Dung beetles in a Central Amazonian rainforest and their ecological
role as secondary seed dispersers. Ecological Entomology, 27:257-270.
Galindo-Gonzalez, J. 1998. Dispersion de semillas por murciélagos: su importancia en
la conservacion y regeneracion del bosque tropical. Acta Zoologica Mexicana,
73:57-74.
Galindo-Gonzalez, J., S. Guevara & V. J. Sosa, V. J. 2000. Bat and bird- generated seed
rains at isolated trees in pastures in a tropical rainforest. Conservation Biology,
14:1693-1703.
Hampe, A., J.L. García-Castaño, E.W. Schupp & P. Jordano. 2008. Spatio-temporal
dynamics and local hotspots of initial recruitment in vertebrate-dispersed trees.
Journal of Ecology, 96:668-678.

21
Jordano, P. 2000. Fruits and frugivory. pp. 125-165. In: Seeds: The Ecology of
Regeneration in Plants Community.
Medellin, R. & O. Gaona. 1999. Seed dispersal by bats and birds in forest and disturbed
habitats of Chiapas, México. Biotropica, 31:478-485.
Melo, F.P.L., B. Rodriguez-Herrera, R. Chazdon, R. Medellin & G. Ceballos. 2009.
Small tent-roosting bats promote dispersal of large-seeded plants in a
Neotropical forest. Biotropica, 41:737-743.
Passos, L. & P.S. Oliveira. 2002. Ants affect the distribution and performance of
seedlings of Clusia criuva, a primarily bird-dispersed rain forest tree. Journal of
Ecology, 90:517-528.
Stoner, K. 2000. Leaf selection by the tent-making bat Artibeus watsoni in Asterogyne
martiana palms in southwestern Costa Rica. Journal of Tropical Ecology,
16:151-157.

22
A cobertura foliar por liquens e briófitas produz custos para Duguetia cf.
flagellaris (Annonaceae)?
Daniel Passos, Bianca Silva, Cibele Paiva & Vidal Carrascosa
Introdução
Nos sub-bosques florestais, a disponibilidade de luz representa um recurso limitante
para o crescimento e desenvolvimento das plantas (Schulze et al. 2002). Por isso,
mesmo pequenas reduções na incidência luminosa podem diminuir a taxa fotossintética
e, consequentemente prejudicar o desenvolvimento das plantas.
Em florestas tropicais úmidas ocorre o fenômeno da epifilia, um tipo de
interação na qual briófitas, algas, fungos e liquens crescem sobre a superfície das folhas
de angiospermas (também chamadas de forófitos), podendo cobri-las total ou
parcialmente (Richards 1996; Gentil & Menezes 2011). Essa cobertura pode reduzir a
área de captação de luz, e consequentemente, a taxa fotossintética, gerando custos para a
planta.
Sob uma condição estressante gerada por uma baixa taxa fotossintética, a planta
pode diminuir a quantidade de recursos investidos em defesa, tornando-se mais
suscetível a herbivoria (Schulze et al. 2002). Além disso, se a produção de carbono é
menor, a massa foliar específica da planta também será reduzida (Larcher 1986). Então,
é possível que a planta fragilizada, devido à menor taxa fotossintética, possua maiores
chances de ser consumida por herbívoros.
Uma vez que as epifilas são comuns em sub-bosques de florestas tropicais
pluviais (Zartman & Ilkou-Borges 2007), o entedimento dos seus efeitos sobre as
plantas onde elas se desenvolvem pode revelar aspectos importantes acerca da dinâmica
das plantas de sub-bosque (Jordan et al. 1980). Sendo assim, a pergunta que direcionou

23
esta pesquisa foi: como a epifilia se relaciona com alocação de carbono e consumo
foliar no forófito? Nossa hipótese foi que um incremento da epifilia diminui a taxa
fotossintética e a defesa da planta contra herbivoria. Nossa expectativa é que quanto
maior a proporção da área de cobertura por epífilas na folha, menor será a massa foliar
específica e maior será a proporção de área foliar consumida.
Métodos
Conduzimos o estudo em um fragmento de floresta de terra firme de 100 ha, na Fazenda
Dimona, Manaus, Brasil. Usamos como modelo de estudo 10 indivíduos de Duguetia
cf. flagellaris (Anonnaceae), uma espécie típica de sub-bosque, abundante no interior do
fragmento (Ribeiro et al. 1999). Para cada individuo, coletamos aleatoriamente cinco
folhas, nas quais medimos as seguintes variáveis: massa foliar específica, proporção da
cobertura foliar por epífilas na face adaxial e a proporção da área foliar consumida.
Medimos a proporção da cobertura por epífilas nas folhas e a proporção da área
de consumo foliar utilizando uma malha de pontos distantes 5 mm entre si.
Confeccionamos a malha em uma folha plástica transparente e a colocamos sobre a
superfície de cada folha. Em seguida, contamos os pontos que sobrepunham a área
foliar consumida e a área ocupada por epífilas. Dividimos estas estimativas pelo numero
total de pontos sobrepostos na superfície total da folha (incluindo as áreas perdidas por
consumo) para obter as proporções relativas da área foliar consumida e da área de
cobertura por epífilas. Para medir a massa específica foliar, removemos todas as epífilas
presentes na folha. Em seguida, desidratamos um fragmento foliar (2 x 2 cm) em forno
médio, por 1 h e pesamos o material desidratado em uma balança de 1 mg de precisão.
Calculamos a massa foliar específica ao dividir o peso seco pela área foliar desidratada.
Posteriormente, calculamos a média de cada atributo das cinco folhas, para obter os

24
valores médios de proporção de área de cobertura por epifilas, proporção área de
consumo foliar e massa foliar específica para cada indivíduo.
Para testar a previsão de que a proporção de área de cobertura por epífilas se
relaciona negativamente com a massa foliar específica em cada indivíduo, fizemos uma
regressão linear simples. Consideramos a massa foliar específica média como variável
resposta e a média da proporção da área de cobertura por epífilas como variável
preditora. Para testar a previsão de que plantas com maior proporção da área de
cobertura foliar por epífilas apresentam maior proporção de área com consumo foliar,
fizemos uma correlação de Spearman.
Resultados
Não houve relação entre a média da proporção da área de cobertura por epífilas e a
media da massa foliar específica (R2
= 0,31; F(1,9) = 3,99; p = 0,08; Figura 1). Também
não houve associação entre a média da proporção de consumo foliar e a média da
proporção da área de cobertura por epifilas em cada indivíduo (N=10, Rs = 0,55; p =
0,10; Figura 2).

25
Figura 1. Relação entre a massa foliar específica média (mg/cm2) e a proporção média
da área de cobertura por epífilas em indivíduos da planta Duguetia cf. flagellaris
localizados num fragmento florestal na Fazenda Dimona, Manaus, Brasil.
Figura 2. Relação entre a proporção média de área com consumo foliar e proporção
média da área de cobertura por epífilas em indivíduos da planta Duguetia cf. flagellaris
localizados num fragmento florestal na Fazenda Dimona, Manaus, Brasil.
0,12 0,14 0,16 0,18 0,20 0,22 0,24 0,26 0,28 0,30 0,32 0,34 0,36 0,38
Proporção Média de Área Coberta por Epífilas
5
6
7
8
9
10
Mass
a F
oliar
Esp
ecíf
ica M
édia
(mg/c
m2)
y = 6,4879 + 5,094*x
0,12 0,14 0,16 0,18 0,20 0,22 0,24 0,26 0,28 0,30 0,32 0,34 0,36 0,38
Proporção Média de Área Coberta por Epífilas
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
Pro
po
rção
Méd
ia d
e Á
rea d
e C
on
sum
o F
oli
ar

26
Discussão
O aumento da cobertura por epífilas não apresentou relação com a massa foliar
específica, evidenciando que sombreamento foliar causado pelas epífilas não afeta a
eficiência fotossintética das folhas do forófito. Além disso, maiores proporções de
cobertura por epífilas não afetaram o consumo foliar, demonstrando que o
sombreamento não torna as folhas mais suscetíveis a herbivoria.
Em florestas, a pouca quantidade de luz que atravessa o dossel constitui um fator
limitante para o desenvolvimento de muitas plantas (Raven 1992). Assim, é possível
que D. cf. flagellaris, por ser típica de sub-bosque, apresente adaptações a ambientes
com essas restrições, como folhas grandes, taxa reduzida de troca foliar e ponto de
compensação luminosa baixo (Schulze et al. 2002). Nessa situação, o sombreamento
foliar causado pelas epífilas pode não ser suficiente para prejudicar as plantas.
Muito do que se conhece sobre a redução de luz no sub-bosque de floretas
úmidas está associado à captação de luz pelas plantas no dossel (Schulze et al. 2002).
No entanto, como a epifilia é comum no sub-bosque de florestas pluviais tropicais
(Zartman & Ilkou-Borges 2007), é possível que a ocorrência destes organismos sobre as
folhas de plantas do sub-bosque constitua um componente historicamente relacionado à
adaptação dessas plantas a baixa disponibilidade de luz. Consequentemente, a presença
desses organismos sobre as folhas não afetaria negativamente seu metabolismo.
Agradecimentos
Agradecemos ao professor Márcio Zikán pela orientação ao longo de todo o projeto, ao
Sr. Leonardo por nos guiar em campo e encontrar os indivíduos de Duguetia cf.
flagellaris, ao prof. Michael Hopkins por confirmar a identificação do objeto de estudo
e, ao prof. Paulo Cardoso pelas valiosas contribuições na versão final deste manuscrito.

27
Referências
Berrie, G.K. & J.M.O. Eze. 1975. The relationship between an epiphyllous liverwort
and host leaves. Annals of Botany, 39:955-963.
Gentil, K.C.S. & C.R. Menezes. 2011. Levantamento de briófitas bioindicadoras de
perturbação ambiental do campus Marco Zero do Equador da UNIFAP. Biota
Amazônia, 1:63-73.
Jordan, C., F. Golley, & J. Hall. 1980. Nutrient scavenging of rainfall by the canopy of
the Amazonian rain forest. Biotropica, 12:61-66.
Larcher, W. 1986. Ecofisiologia vegetal. São Paulo: Editora Pedagógica Universitária.
Raven, P.H., R.F. Evert & S.E. Eichhorn. 1992. Biologia vegetal. Rio de Janeiro:
Guanabara Koogan.
Ribeiro, J.E.L.S, M.J.G. Hopkins, A. Vicentini, C.A. Sothers, M.A.S. Costa, J.M. Brito,
M.A.D. Souza, L.H.P. Martins, L.G. Lohmam, P.A.C.L. Assunção, E.C. Pereira,
C.F. Silva, M.R. Mesquita & L.C. Procópio. 1999. Flora da reserva Ducke: Guia
de identificação das plantas vasculares de uma floresta de terra firme na
Amazônia Central. Manaus: INPA.
Richards, P.W. 1996. The tropical rain forests. Cambridge: Cambridge University Press.
Schulze, E., E. Beck, K. Müller-Hohenstein. 2002. Plant ecology. Bayreuth: Springer.
Zartman, C.E. & A.L. Ilkou-Borges. 2007. Guia para as briófitas epífilas da Amazônia
Central. Manaus: INPA.

28
A riqueza de plantas aumenta a riqueza de herbívoros, mas não dos predadores
Renata Pimentel Rocha, Caroline Marques Dracxler, Leonardo Oliveira, Bruno
Travassos de Britto
Introdução
A riqueza de plantas é apontada como um importante determinante da diversidade
animal (Langellotto & Denno 2004). O aumento no número de espécies de plantas pode
contribuir para uma paisagem mais diversificada, com estruturas morfológicas variáveis
para abrigar artrópodes e mais recursos alimentares para artrópodes herbívoros (Tews et
al. 2004).
Para evitar o ataque por artrópodes herbívoros, as plantas utilizam mecanismos
de defesa física ou química (Godfray et al. 1999, Novotny & Basset 2005). Tais
mecanismos representam uma barreira contra herbivoria que irá selecionar somente os
herbívoros capazes de se alimentar dessas plantas. Essa relação gera maior
especialização tanto por parte das plantas quanto dos herbívoros (Jaenike 1990), o que
leva a uma relação entre riqueza de plantas e dos herbívoros (Dinnage et al. 2006).
Artrópodes predadores, por sua vez, não se alimentam de plantas e por isso não
são diretamente afetados pelo tipo de defesa das plantas. Esses artrópodes usam as
plantas como abrigo, local para forrageio, oviposição e proteção. Esses tipos de uso
estão relacionados com aspectos da arquitetura da planta, como quantidade e formato
dos ramos e das folhas. Dessa forma, o aumento da riqueza de plantas pode ter efeito
menos acentuado sobre os predadores, pois sua relação com as plantas não está ligada
ao número de espécies e sim com a arquitetura da planta (Robinson, 1981).
Como riqueza de espécies de plantas pode estar associada de forma diferente
com artrópodes herbívoros e predadores, nós investigamos se o aumento na riqueza de

29
plantas afeta de maneira distinta a riqueza de herbívoros e predadores. Esperamos que o
aumento no número de plantas resulte em aumento na riqueza de artrópodes herbívoros
com mais intensidade do que na riqueza de artrópodes predadores.
Métodos
Área de Estudo
Nós realizamos o estudo na fazenda Dimona, uma ARIE gerenciada pelo Projeto de
Dinâmica Biológica de Fragmentos Florestais (PDBFF), em conjunto com o ICMBio. A
área é localizada a 80 km a norte de Manaus, na Amazônia Central, Brasil (02°20’ N,
60°05’ O). A paisagem é composta por uma matriz de vegetação secundária contendo
fragmentos de floresta do tipo tropical pluvial. A pluviosidade anual varia de 1900 a
2300 mm, com temperatura média de 26°C (Marques & Filho 1981).
Coleta de plantas e artrópodes
Coletamos os dados em dois ambientes distintos (interior de mata e capoeira) para
aumentar variação na diversidade de plantas. Em cada ambiente, amostramos sete
pontos, distantes 15 m entre si, ao longo de um transecto. Em cada ponto, amostramos
sete arbustos com altura entre um e dois metros. Para selecionar os arbustos, sorteamos
números de um a oito em um dado, correspondentes a oito direções distribuídas
igualmente em 360°. O número sorteado corresponde à direção para escolha da primeira
planta a ser amostrada. Os arbustos seguintes foram escolhidos por meio de novos
sorteios da direção. Ensacamos e coletamos dois galhos de cada planta selecionada, nos
quais realizamos 10 batimentos para coletar os artrópodes. Após a coleta dos insetos, as
plantas foram identificadas ao nível de família por especialistas e em seguidas foram

30
morfotipadas. Em laboratório separamos artrópodes herbívoros e predadores através de
características estruturais do aparato bucal e distinguimos os morfotipos.
Análise de Dados
Para testar o efeito da riqueza de plantas sobre a riqueza de herbívoros e predadores,
usamos uma Análise de Variância (ANOVA). A variável dependente riqueza de
predadores e herbívoros foi transformada para escala logarítmica para se ajustar à
distribuição normal.
Resultados
Nós amostramos um total de 69 plantas distribuídas em 24 famílias, das quais
Violaceae, Annonaceae e Melastomataceae foram as mais representativas. A quantidade
de morfoespécies de plantas variou entre 5 e 7 (Média = 6 ± 0,89) em cada ponto de
amostragem. A riqueza de plantas aumentou com a riqueza de herbívoros (F(2, 7)= 11,22;
p= 0,01; Figura 1), mas não com a riqueza dos predadores (F(2, 7)= 1,38; p= 0,31; Figura
2).
A intensidade da relação entre plantas e herbívoros foi diferente do que o
observado entre plantas e predadores. O aumento no número de herbívoros em função
da riqueza de plantas foi em média 0,45. Os predadores, por sua vez, tiveram aumento
médio de 0,15 espécies à medida que a riqueza de plantas aumentou.

31
Figura 1. Relação entre a riqueza (morfotipos) de plantas e a riqueza de morfotipos de
herbívoros em uma área na Amazônia Central. Quadrados representam médias e barras
o desvio padrão.
Figura 2. Relação entre a riqueza (morfotipos) de plantas e a riqueza de morfotipos de
predadores em uma área na Fazenda Dimona (80 km a norte de Manaus) - Amazônia
Central. Quadrados representam médias e barras o desvio padrão.

32
Discussão
Os resultados mostraram que o aumento da riqueza dos herbívoros foi relacionado com
a riqueza das plantas. Isso indica uma especialização alimentar nos herbívoros, o que
resulta em vínculo direto entre riqueza de plantas e herbívoros. A diversidade de
predadores, por outro lado, não teve relação com a diversidade de plantas, o que indica
que variações estruturais nas plantas e até mesmo a riqueza de herbívoros não afetaram
a presença dos predadores nesses locais (Greenquist & Rovner 1976).
O aumento da riqueza na comunidade de plantas está relacionado com aumento
da heterogeneidade de recursos alimentares, defesas químicas e físicas (Dinnage et al.
2012). Essa heterogeneidade leva ao aumento da riqueza de herbívoros, pois esses
artrópodes tendem a se especializar no consumo de determinadas plantas (Jaenike
1990). A especialização dos herbívoros ocorre principalmente em resposta a defesas
químicas da planta, uma vez que contornar essa proteção demanda grande
especialização fisiológica para lidar com compostos químicos específicos (Mello &
Silva-Filho 2002). As estratégias contra defesas físicas, por outro lado, podem ser
menos específicas, pois uma característica do herbívoro, como a presença de mandíbula
forte para consumir uma folha, pode ser usada em diferentes espécies de plantas
(Crawley 1997).
A relação entre plantas e predadores se baseia, sobretudo, em características
estruturais da planta. Diferentes espécies de plantas podem apresentar arquitetura
semelhante e essa complexidade estrutural pode variar pouco entre espécies,
principalmente entre plantas do mesmo gênero. Por isso as adaptações morfológicas dos
artrópodes predadores podem ser menos especializadas e a capacidade de uso de recurso
é independente da riqueza de plantas.

33
Além disso, a diversidade de predadores pode ser mais relacionada com a
riqueza dos herbívoros do que diretamente com a riqueza de plantas. Como predadores
são generalistas, a quantidade de alimento disponível deve ser mais relevante do que a
diversificação desse alimento. Dessa forma, é possível que o aumento na riqueza de
herbívoros também aumente sua abundância, o que poderia levar a um aumento na
abundância de predadores. Essa relação da quantidade de herbívoros com a dos
predadores pode ser mais comum em ambientes com alta diversidade de plantas, onde a
riqueza de plantas e, consequentemente, dos herbívoros é alta.
Referências
Balvanera P. & E. Aguirre. 2006. Tree diversity, environmental heterogeneity, and
productivity in a Mexican tropical dry forest. Biotropica, 38:479-491
Crawley, M.J. 1997. Plant ecology. Oxford: Blackwell publishing
Dinnage, R., M.W. Cadotte, N.M. Haddad, G.M. Crutsinger & D. Tilman. 2012.
Diversity of plant evolutionary lineages promotes arthropod diversity. Ecology
Letters, 15:1308-1317.
Downes, B. J., P. S. Lake, E. S. G. Schreiber & A. Glaister. 1998. Habitat structure and
regulation of local species diversity in a stony, upland stream. Ecological
Monographs, 68:237-257.
Godfray, H.C.J., O. T. Lewis & J. Memmott. 2005. Studying insect diversity in the
tropics. Philosophical Transaction of Royal Society of London B, 354:1811-1824.
Halaj, J., D. W. Ross & A. R. Moldenke. 2000. Importance of habitat structure to the
arthropod food-web in Douglas-Fir Canopies. Oikos, 90:139-152.
Jaenike, J. 1990. Host specialization in phytophagous insects. Annual Review of
Ecology, Evolution and Systematics, 21:243-273

34
Langellotto, G.A. & R.F. Denno. 2004. Responses of invertebrate natural enemies to
complex-structured habitats: a meta-analytical synthesis. Oecologia, 139:1-10.
Marques-Filho, A. O., Ribeiro, M. N. G., Santos, H. M., Santos J. M. (1981) Estudos
climatológicos da Reserva Florestal Ducke – Manaus-AM. Acta
Amazonica,11:759-768.
Mello, M.O. & M.C. Silva-Filho. 2002. Plant-insect interactions: an evolutionary arms
race between two distinct defense mechanisms. Brazilian Journal of Plant
Physiology, 14:71-81.
Novotny V. & Y. Basset. 2005. Host specificity of insect herbivores in tropical forests.
Proceedings of The Royal Society B, 272:1083-1090
Ødegaard, F., O.H. Diserud & K. Østbye. 2005. The importance of plant relatedness for
host utilization among phytophagous insects. Ecology Letters, 8:612-617.
Robinson, J. V. 1981. The effect of architectural variation in habitat on a spider
community: an experimental field study. Ecology, 63: 73-80
Romero, G.Q. & J. Vasconcellos-Neto. 2005. The effects of plant structure on the
spatial and microspatial distribution of a bromeliad-living jumping spider
(Salticidae). Journal of Animal Ecology, 74:12-21.
Scherber, C., N. Eisenhauer, W.W. Weisser, B. Schmid, W. Voigt & M. Fischer. 2010.
Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity
experiment. Nature, 468:553-556.
Tews, J., U. Brose, V. Grimm, K. Tielbörger, M.C. Wichmann, M. Schwager & F.
Jeltsch. 2004. Animal species diversity driven by habitat heterogeneity/diversity: the
importance of keystone structures. Journal of Biogeography, 31:79-92.
Weiblen, G.D., C.O. Webb, V. Novotny, Y. Basset, & S.E. Miller. 2006. Phylogenetic
dispersion of host use in a tropical insect herbivore community. Ecology, 87:62-75.

35
Espécies de animais aquáticos e semiaquáticos se distribuem entre poças
temporárias independentemente da qualidade do habitat
Mariana Velasque, Ana Claudia Rorato, Ludmila Rattis, Randolpho Gonçalves Dias
Terceiro
Introdução
A qualidade do habitat pode ser entendida como a soma das condições ambientais
(bióticas e abióticas) e dos recursos que estão disponíveis para os organismos que vivem
em um determinado local (Begon et al. 2006). Perturbações naturais ou de origem
antrópica podem promover a deterioração dessas condições e recursos, tornando o
habitat menos favorável para algumas espécies. Como resultado, espécies menos
tolerantes poderão ser as primeiras a ser extintas localmente (Atmar & Patterson 1993;
Duffy 2003). Assim, habitats de menor qualidade devem comportar número menor de
espécies, que correspondem a um subconjunto previsível de espécies altamente
tolerantes que também são encontradas em habitats de maior qualidade (Patterson &
Atmar 1986; Ulrich & Gotelli 2007; Jacquemyn et al. 2007). Essa perda ordenada de
espécies em comunidades, associada à deterioração da qualidade do habitat, produz um
padrão conhecido como aninhamento (Atmar & Patterson 1993; Lomolino 1996).
Poças temporárias formadas pelo acúmulo de água no interior de florestas
tropicais podem ser exemplos de habitats que sofrem deterioração gradual de qualidade
(Espírito-Santo et al. 2009). A formação dessas poças ocorre devido ao alagamento de
áreas adjacentes ao leito de riachos em decorrência da alta pluviosidade. Ao cessar o
alagamento, formam-se poças temporárias em pontos do solo com concavidade
acentuada, que são ocupadas por uma série de organismos aquáticos e semi-aquáticos
(Rodríguez & Lewis 1997). O volume de água nas poças pode ser interpretado como um

36
indicador da qualidade do habitat, pois determina as condições e recursos disponíveis
para os organismos que as habitam (Ceotto et al. 2001). Poças com volume menor
provavelmente possuem condições abióticas mais instáveis, com propriedades físicas e
químicas alterando-se mais rapidamente que poças com volume maior. Adicionalmente,
poças menores podem apresentar menor quantidade de recursos como a própria água,
matéria orgânica particulada e alterações nas condições como oxigênio dissolvido,
aumento temperatura e redução do pH.
Na floresta amazônica é comum a formação de poças temporárias próximas a
riachos de primeira ordem conhecidos localmente como igarapés. Essas poças são
ocupadas por uma série de espécies com diferentes graus de dependência da água
(Espírito-Santo et al. 2009). Portanto, elas constituem um sistema ideal para investigar o
padrão de composição de espécies em diferentes condições de qualidade de habitat.
Nossa hipótese é que as espécies de animais encontradas nas poças de menor qualidade
constituem subconjuntos das espécies de animais presentes em poças de maior
qualidade.
Métodos
Área de estudo
Realizamos o estudo ao longo de um riacho de cerca de 3 m de largura que cruza uma
área de capoeira presente na Fazenda Dimona (ARIE Projeto Dinâmica Biológica de
Fragmentos Florestais), localizada a cerca de 80 km ao norte de Manaus, Amazonas,
Brasil (2°25’ S, 59°50’ O). Durante chuvas intensas, o nível de água do riacho sobe e as
pequenas depressões do entorno são alagadas. Logo após a chuva, o nível do riacho
abaixa e a água retida nessas depressões dá origem a pequenas poças temporárias que

37
contém organismos aquáticos e semiaquáticos, tais como pequenos peixes, anuros e
pequenos artrópodes.
Estimativa da qualidade do habitat
Usamos o volume de água presente nas poças como um indicador de qualidade de
habitat. Porém, escolhemos poças estruturalmente semelhantes em área para
controlarmos o efeito do tamanho sobre a riqueza de espécies. Em cada poça, medimos
o comprimento e a largura como os pontos mais distantes nas bordas ao longo dos eixos
longitudinal e transversal. Calculamos a profundidade média a partir de cinco medidas
tomadas em pontos arbitrários ao longo da poça. Multiplicamos as medidas das três
dimensões para estimar o volume (Figura 1).
Figura 1. Método de mensuração das dimensões das poças temporárias ao redor de um
riacho na floresta amazônica. As setas contínuas apontam os pontos mais distantes ao
longo dos eixos transversal e longitudinal. As setas pontilhadas representam locais
hipotéticos de medida de profundidade da poça com os quais se obteve a profundidade
média.

38
Riqueza e composição das comunidades animais
Coletamos indivíduos macroscópicos de espécies aquáticas e semiaquáticas por toda a
coluna d’água até a camada mais superficial do sedimento com o uso de uma peneira de
malha de 3 mm2. Triamos folhas, galhos, sedimentos e organismos em laboratório e, em
seguida, separamos os indivíduos encontrados em morfoespécies. Utilizamos o número
de morfoespécies presente nas poças temporárias como medida de riqueza.
Análise dos dados
Para testar se as espécies nas poças com menor volume de água compunham
subconjuntos das espécies em poças com maior volume, construímos uma matriz de
presença/ausência de espécies (linhas) por poças (colunas) arranjadas em ordem
decrescente de volume. Calculamos o grau de aninhamento observado a partir da matriz
de presença/ausência de morfoespécies por poça usando a métrica NODF (um acrônimo
para Nestedness metric based on Overlap and Decreasing Fill; Almeida-Neto et al
2008). Fizemos as análises no programa Aninhado versão 3.0 Bangu (Guimarães &
Guimarães 2006).
Resultados
Identificamos 18 morfoespécies distribuídas em 13 ordens, sendo Coleoptera a ordem
mais abundante (Figura 2). O volume das poças variou de 0,084 m3
a 0,549 m3, com
média ± DP de 0,306 ± 0,169 m3. A composição de morfoespécies não foi influenciada
pelo volume das poças, pois poças de menor volume de água não representaram um
subconjunto das espécies das poças de maior volume (NODF = 13,14; p = 0,117).

39
Espécies
Po
ças
C
ole
op
tera
1
Co
leo
pte
ra 2
Lar
va
de
inse
to 1
Ch
iro
no
mid
ae
Pei
xe
1
Co
leo
pte
ra 3
Pei
xe
2
Lar
va
de
inse
to 2
Lar
va
de
inse
to 3
Cru
sta
ceo
1
Cru
sta
ceo
2
Cru
sta
ceo
3
Bla
tod
ea
Lar
va
de
inse
to 4
Co
leo
pte
ra 4
Co
leo
pte
ra 5
Co
leo
pte
ra 6
Pip
a c
arv
alh
oi
1 (0,549 m³)
2 (0,446 m³)
3 (0,430 m³)
4 (0,363 m³)
5 (0,271 m³)
6 (0,205 m³)
7 (0,100 m³)
8 (0,084 m³)
Figura 2. Matriz de presença-ausência de animais macroscópicos em poças temporárias
ao longo de um igarapé na Amazônia Central. As linhas representam as poças, dispostas
em ordem decrescente de volume e as colunas representam as espécies. Células
marcadas de preto representam presença das espécies nas poças, células em branco a
ausência. Marcações em negrito indicam as morfoespécies estritamente aquáticas.
Discussão
O padrão de composição das espécies encontrado não indica uma perda ordenada de
espécies entre as poças em função do volume de água. Consequentemente, é possível
que as mudanças nas características físico-químicas associadas à degradação das poças
não afetem grupos específicos de espécies que abandonariam gradualmente os locais
com condições mais adversas.
Dois terços das morfoespécies encontradas possui capacidade de deslocamento
entre poças. Consequentemente, é possível que eles transitem recorrentemente entre as
poças, evitando que um padrão aninhado seja formado. Vieira (2010) demonstrou que

40
uma espécie de peixe que ocorre nesse sistema abandona as poças em função da
presença de predadores. Nesse sentido, é possível que os organismos transitem entre as
poças em função de interações bióticas como predação ou competição.
Em um estudo conduzido no mesmo local, Souza et al. (2005) encontraram um
padrão aninhado entre as comunidades das poças localizadas a diferentes distancias do
riacho. Uma vez que a distancia pode determinar o acesso dos organismos a área fonte
de espécies, talvez a comunidade responda mais a esse fator do que a variações de
qualidade. De fato, em outro trabalho envolvendo o efeito de degradação de poças em
sistemas inundáveis, nenhum padrão de aninhamento foi encontrado (Zatz et al. 2008).
Por fim, ainda é possível que o efeito da degradação varie de acordo com o grupo de
organismos. Souza et al. (2005) encontraram principalmente espécies de vertebrados,
enquanto nós encontramos predominância de insetos. Eventualmente, por apresentarem
um ciclo de vida mais curto, os insetos podem ser menos dependentes da degradação
das poças quando comparados aos vertebrados que podem depender da qualidade local
durante toda a vida.
Agradecimentos
Gostaríamos de agradecer à Esther Sebastián e ao monitor barbudo Marcos Costa Vieira
pela orientação e ajuda em campo. Aos coordenadores Paulo Estefano e Paulo Enrique
pela ajuda na triagem e orientação, ao Leo Marajó pela ajuda em campo, ao Glauco por
revisar esse trabalho 23 vezes e ao Paulo Enrique, por tirar leite de pedra.

41
Referências
Almeida M.N., P. Guimarães, P.R. Guimarães, R.D. Loyola & W. Ulrich. 2008. A
consistent metric for nestedness analysis in ecological systems: reconciling
concept and measurement. Oikos, 117:1227-1239.
Atmar, W. & B.D. Patterson. 1993. The measure of order and disorder in the
distribution of species in fragmented habitat. Oecologia, 96:373-382.
Begon, M.; C.R. Townsend & J.L. Harper. 2006. Ecology: From Individuals to
ecosystems. Blackwell Publishing: Oxford.
Ceotto, P.C.; A.M.F. Pacheco; L.M. Scoss; M. Camargo & P.S. Pinheiro. 2001. Efeito
do isolamento em poças sobre a estrutura da comunidade de macroinvertebrados
bentônicos no arquipélago das Anavilhanas, AM. Em: Livro do curso de campo
“Ecologia da Floresta Amazônica” (E. Venticinque & J. Zuanon, eds.).
PDBFF/INPA, Manaus.
Duffy, J.E. 2003. Biodiversity loss, trophic skew and ecosystem functioning. Ecology
Letters, 6:680–687.
Guimarães, P.R. & P. Guimarães. 2006. Improving the analyses of nestedness for large
sets of matrices. Environmental Modelling and Software, 21:1512-1513.
Espírito-Santo, H. M. V.; Magnusson, W. E.; Zuanon, J.; F. P. Mendonça & V. Lemes
2009. Seasonal variation in the composition of fish assemblages in small
Amazonian forest streams: evidence for predictable changes. Freshwater
Biology, 54:536–548.
Jacquemyn, H., O. Honnay & T. Pailler. 2007. Range size variation, nestedness and
species turnover of orchid species along an altitudinal gradient on Reunion
Island: implications for conservation. Biological Conservation, 136:388-397.

42
Lomolino, M.V. 1996. Investigating causality of nested-ness of insular communities:
selective immigrations or extinctions? Journal of Biogeography, 23:699–703.
Patterson, B.D. & W. Atmar 1986. Nested subsets and the structure of insular
mammalian faunas and archipelagos. Biological Journal of the Linnean Society,
28:65-82.
Rodríguez, M.A. & W.M. Jr Lewis. 1997. Structure of fish assemblages along
environmental gradients in floodplain lakes of the Orinoco River. Ecological
Monographs, 67:109–128.
Souza, A.C., D.P. Lima-Junior, G. Schussler, V.T.Cardoso, W.R. Almeida. 2005. Existe
uma sequência na perda de espécies em poças temporárias? In: Livro do curso de
campo “Ecologia da Floresta Amazônica” (Machado G. & H.Nascimento EDS)
PDBFF/INP:Manaus
Ulrich, W. & N.J. Gotelli. 2007a. Null model analysis of species nestedness patterns.
Ecology, 88:1824-1831.
Vieira, C. Sinais químicos de predadoresinterferem na permanência de R. compressus
(Cyprindontiformes: Rivulidae) em poças. In: Livro do curso de campo
“Ecologia da Floresta Amazônica” (Peixoto, P.E.C. & P.E.D. Bobrowiec EDS)
PDBFF/INP:Manaus
Zatz, C., A. Miranda, B. Azambuja, C. Cordeiro & G. Sinimbu. 2008. A estrutura física
de poças marginais influencia a estrutura da comunidade de macroinvertebrados
aquáticos? In: Livro do curso de campo “Ecologia da Floresta Amazônica”
(Machado, G. & J.L.C. Camargo EDS.) PDBFF/INP:Manaus

43
O tamanho da domácia foliar determina sua ocupação por ácaros?
Tatiane Gomes Calaça Menezes, Daniela Pinto Coelho, Lorena Costa Pinto e Artur
Alcantara Madeira
Introdução
Seleção de habitat é uma escolha feita pelos organismos por características específicas
do ambiente (Morris 1992). Tem como objetivo minimizar efeitos prejudiciais e/ou
maximizar ganhos para os indivíduos (Morin 2011). Em geral, os ganhos são
determinados pela disponibilidade de alimento e de parceiros reprodutivos, enquanto os
efeitos negativos estão relacionados ao risco de predação enquanto forrageiam (Sih
1980, Morin 2011).
Os organismos podem usar diferentes tipos de informações para selecionar um
habitat. Por exemplo, a rã Hyla chrysoscelis deposita diferentes quantidades de ovos no
substrato, de acordo a presença de predadores e competidores que habitam o local
(Resetarits & Wilbur, 1989). Já os crustáceos anfípodes selecionam as macroalgas
marinhas que habitam através de características como, por exemplo, a coloração da alga
que lhe permita ser críptico, diminuindo o risco de predação (Hacker & Steneck 1990).
Um problema inerente a estudos de seleção de habitat em sistemas naturais é o
excesso de correlações entre os fatores que podem determinar a escolha de habitats. Tais
correlações dificultam a identificação da característica que de fato é selecionada pelo
organismo (e.g. Resetarits & Wilbur, 1989). Por exemplo, no estudo com H.
chrysoscelis várias espécies de predadores e competidores podem estar ocorrendo no
mesmo ambiente, tornando difícil separar o efeito de cada espécie sobre a escolha do
local de oviposição pela rã. Nesse sentido, a busca por abordagens in situ que sofram
pouco efeito de confundimento entre variáveis podem ser esclarecedoras.

44
Plantas com domácias foliares habitadas por ácaros representam um sistema
natural particularmente interessante para remover efeitos de correlações espúrias entre
características do ambiente e a seleção de habitat por animais. Domácias são estruturas
localizadas nas axilas das nervuras das folhas formadas por uma cavidade ou tufo de
pelos que servem como local de refúgio e reprodução para ácaros (Norton et al. 2000).
Nesse contexto, em uma mesma folha o tamanho da domácia é um dos poucos atributos
que varia consideravelmente ao longo do substrato relativamente uniforme da folha.
Portanto, folhas podem representar um sistema que permite avaliar com mais clareza
como uma característica específica pode determinar a seleção de habitat em um sistema
natural.
O tamanho da domácia pode determinar sua qualidade para os ácaros. Domácias
maiores apresentam maior número e adensamento de pelos, o que pode aumentar a
segurança do refúgio. Nesse trabalho objetivamos responder qual o efeito do tamanho
da domácia na ocupação por ácaros. Nossa hipótese é que o tamanho da domácia é
positivamente relacionado à sua probabilidade de ocupação por ácaros. Desse modo,
esperamos que domácias de maior área sejam mais ocupadas por ácaros em relação às
domácias de menor área.
Métodos
Área de estudo
Realizamos o estudo em uma floresta secundária localizada na fazenda Dimona (02° 20’
S – 60º 06’ O), a aproximadamente 90 Km ao norte da cidade de Manaus, Brasil
(Bierregaard & Gascon 2001). Usamos como modelo de estudo a planta Buchenavia
tomentosa (Combretaceae), que apresenta domácias formadas por tufos de tricomas,

45
localizados na interseção da venação principal com as veias secundárias da face abaxial
das folhas (Ribeiro et al. 1999).
Coleta e análise dos dados
Coletamos 10 folhas de diferentes ramos de dois indivíduos de B. tomentosa. No
laboratório, fotografamos as domácias situadas no lado esquerdo da venação principal
da folha, utilizando uma régua como escala métrica. A partir dessas fotografias,
calculamos a área de cada domácia utilizando o programa Image Tool.
Após fotografar as folhas, usamos uma lupa para vistoriar todas as domácias a
procura de ácaros ou indícios de sua ocupação (ovos ou detritos). Classificamos os
ácaros encontrados em morfo-espécies de acordo com o formato e coloração de seus
corpos. Para testar a relação entre a área da domácia e a presença de ácaros, utilizamos
uma regressão logística. Consideramos a área da domácia como variável preditora e a
presença de ácaros como variável resposta.
Resultados
Encontramos 83 domácias, com média de 8,3 ± 0,48 (média ± desvio padrão) domácias
por folha. Registramos ácaros ou indícios de sua ocupação em 49 domácias (59%).
Identificamos quatro morfo-espécies de ácaros, sendo que todas tinham tamanhos
visualmente similares entre si. Duas ou três morfo-espécies co-ocorreram em 25% das
folhas. No entanto, a ocupação por ácaros não esteve associada à área foliar (χ2
= 0,725;
gl = 1; p = 0,394, Figura 1).
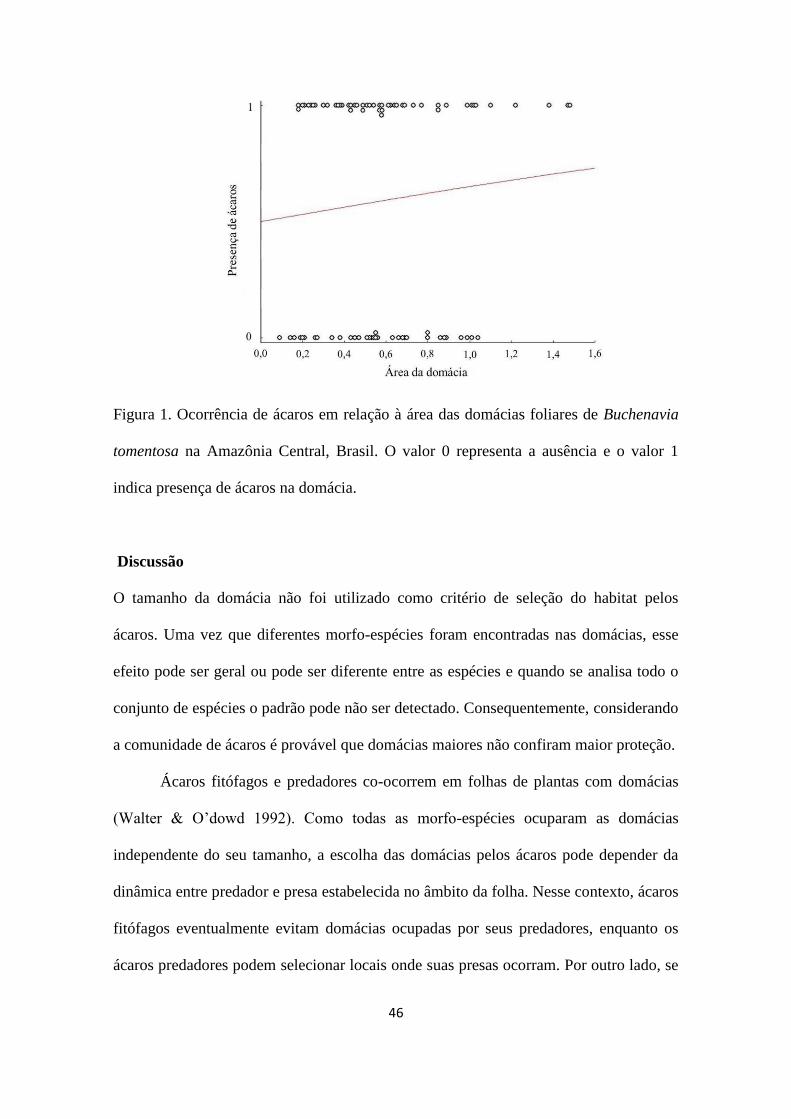
46
Figura 1. Ocorrência de ácaros em relação à área das domácias foliares de Buchenavia
tomentosa na Amazônia Central, Brasil. O valor 0 representa a ausência e o valor 1
indica presença de ácaros na domácia.
Discussão
O tamanho da domácia não foi utilizado como critério de seleção do habitat pelos
ácaros. Uma vez que diferentes morfo-espécies foram encontradas nas domácias, esse
efeito pode ser geral ou pode ser diferente entre as espécies e quando se analisa todo o
conjunto de espécies o padrão pode não ser detectado. Consequentemente, considerando
a comunidade de ácaros é provável que domácias maiores não confiram maior proteção.
Ácaros fitófagos e predadores co-ocorrem em folhas de plantas com domácias
(Walter & O’dowd 1992). Como todas as morfo-espécies ocuparam as domácias
independente do seu tamanho, a escolha das domácias pelos ácaros pode depender da
dinâmica entre predador e presa estabelecida no âmbito da folha. Nesse contexto, ácaros
fitófagos eventualmente evitam domácias ocupadas por seus predadores, enquanto os
ácaros predadores podem selecionar locais onde suas presas ocorram. Por outro lado, se

47
não existe uma relação com os ácaros predadores, o mesmo não pode ser dito para seus
ovos. Sabe-se que os ovos dos ácaros predadores são mais sensíveis à dessecação que
ácaros fitófagos (Agrawal 1997). Esses ovos podem ser em parte beneficiados pelo
microclima no interior da domácia. Logo uma associação entre deposição dos ovos e
tamanho da domácia poderia ser encontrada. O adulto por sua vez, não ocorreria
preferencialmente associado a domácias grandes por não sofrer esse efeito da
dessecação como ocorre nos ovos.
Se o tamanho da domácia não está associado a nenhum benefício específico para
os ácaros e as demais características da folha são relativamente homogêneas, os ácaros
podem se distribuir homogeneamente pela folha, proporcionando o mesmo nível de
defesa ao longo de todo o limbo foliar. Logo, a variação do tamanho da domácia não
deve ser resultado de benefícios associados à atração dos ácaros para regiões específicas
da folha, mas deve ser um padrão resultante do acaso.
Agradecimentos
Ao Michael Hopkins pela orientação e vivência do tema e ao Paulo Enrique pela valiosa
contribuição no desenho experimental, coleta e análise dos dados.
Referências
Agrawal, A.A. 1997. Do leaf domatia mediate a plant-mite mutualism? An experimental
test of the efects on predators and herbivores. Ecological entomology, 22:371-376.
Bierregaard, R.O. & C. Gascon. 2001. The Biological Dynamics of Forest Fragments
Project: overview and history of a long-term conservation project, pp. 5–12. In:
Lessons from Amazonia: the ecology and conservation of a fragmented forest

48
(Bierregaard, R.O., T. Gascon, T.E. Lovejoy & R. Mesquita, eds). New Haven: Yale
University Press.
Hacker, S.D. & R.S. Steneck. 1990. Habitat architecture and the abundance and body-
size-dependent habitat selection of a phytal amphipod. Ecology, 71:2269-2285.
Morin, P. 2011. Community ecology. 2ª ed. West Sussex: Wiley-Blackwell.
Morris, D.W. 1992. The role of habitat selection in landscape ecology. Evolutionary
Ecology, 6:357-359.
Norton A.P., G. English-Loeb, D. Gadoury & R.C. Seem. 2000. Mycophagous mites
and foliar pathogens: leaf domatia mediate tritrophic interactions in grapes. Ecology,
81:490–499.
Resetarits, W.J. & H.M, Wilbur. 1989. Choice of oviposition site by Hyla chrysoscelis:
role of predators and competitors. Ecology, 70:220-228.
Ribeiro, J.E.L.S. & N. Marquete. 1999. Combretaceae, pp.459-461. In: Flora da
Reserva Ducke: guia de identificação das plantas vasculares de uma floresta de terra
firme na Amazônia Central (Ribeiro, J.E.L., M.J.G. Hopkins, A.Vincentini,
C.A.Sothers, M.A. da S. Costa, J.M. de Brito, M.A.D. de Souza, L.H.P. Martins,
L.G. Lohmann, P.A.C.L. Assunção, E.C. Pereira, C.F. da Silva, M.R. Mesquita &
L.C. Procópio, eds.). Manaus: INPA.
Rosenzweig, M.L. 1981. A theory of habitat selection. Ecology, 62:327-335.
Sih, A. 1980. Optimal behavior: can foragers balance two confliting demands? Science,
210:1041-1043.
Walter, D.E. & D.J. O’Dowd. 1992. Leaf morphology and predators: effect of leaf
domatia on the abundance of predatory mites (Acari: Phytoseiidae). Environmental
Entomology, 21:478-484.

49
Dimona – projetos livres
Alocação diferencial de recurso em Vismia guianensis (Clusiaceae) associada a
condições de luminosidade
Randolpho Gonçalves Dias Terceiro, José Hidasi Neto, Sarah Freitas, Lorena Costa
Pinto
A teoria da proporção de recursos prediz que diferentes condições de luminosidade
determinam uma alocação diferencial de recursos em espécies vegetais. Em florestas
tropicais, as bordas florestais têm incidência luminosa maior que no interior do sub-
bosque. Assim, investigamos a existência de alocação diferencial de recursos nesses
dois ambientes por uma planta pioneira. Nossa hipótese é que, em ambientes
sombreados, uma espécie pioneira investe em folhas de menor qualidade, porém mais
eficientes na captação de luz, enquanto, em ambientes abertos, o investimento é em
folhas de maior qualidade. Para isso, comparamos a massa foliar específica (MFE) de
20 indivíduos de Vismia guianensis, sendo 10 indivíduos em uma borda de fragmento
florestal e 10 no sub-bosque de uma floresta secundária. A MFE foi 63% maior em
indivíduos em áreas abertas (média=0,019±0,004 g/cm2) que em indivíduos em áreas
fechadas (média=0,011±0,003 g/cm2 ) (F=1,586; p<0,001; gl=18). Nossos resultados
corroboram a hipótese que a planta é capaz de investir recursos de forma diferenciada
dependendo das condições de luminosidade. Em ambientes sombreados, o recurso é
limitado e a planta parece investir preferencialmente em folhas mais eficientes para a
captação de luz, o que poderia resultar em um aumento no crescimento vertical. Em
contrapartida, o alto investimento em folhas de maior qualidade em plantas da área de

50
borda, possivelmente, está relacionado a estratégias contra a perda de água, uma vez
que, na borda, a água é um recurso mais limitante que luz.

51
Distribuição espacial em Selaginella amazonica (Selaginellaceae) em função da
umidade
Leonardo Queiroz de Oliveira, Bianca Ferreira da Silva, Mariana Velasque &
Joselândio Corrêa Santos
Indivíduos que ocupam locais com alta disponibilidade de recursos podem investir
menos em dispersão a longas distâncias, pois já ocupam locais com boas condições de
desenvolvimento. Em plantas com capacidade de reprodução clonal, como pteridófitas,
a redução do investimento em dispersão por propágulos pode possibilitar o aumento na
reprodução vegetativa, gerando uma agregação de plantas ao redor da planta mãe. Na
Amazônia Central, pteridófitas ocorrem em áreas de baixio e platô. Essas áreas
apresentam diferenças de umidade, com áreas de baixio sendo mais úmida que áreas de
platô. Dessa forma, nossa hipótese e que o padrão de distribuição espacial da pteridófita
Selaginella amazonica será mais agregado em áreas mais úmidas. Em cada área
construímos cinco parcelas de 10 m entre si. Em cada parcela, medimos a distância de
cada planta ao seu vizinho mais próximo de S. amazonica. Se a nossa hipótese for
verdadeira, esperamos que a distância média entre os indivíduos de S. amazonica seja
menor nos baixios e maior em áreas de platô. O padrão de distribuição de S. amazonica
foi similar entre as áreas (t = 1,61; gl = 8; p = 0,14). Isso indica que a planta investe nas
duas formas de reprodução em qualquer local. Talvez isso funcione como uma
estratégia de espalhamento de risco que assegure prole com alta chance de
sobrevivência próxima da planta mãe, mas que garanta descendentes em áreas distantes
que não sofreriam efeitos prejudiciais imprevisíveis próximos das agregações de onde
eles partiram.

52
O investimento em reprodução não diminui o investimento em defesa em Miconia
dispar (Melastomataceae)
Daniela Coelho, Ludmila Rattis, Tatiane Menezes & Cibele Paiva
Os organismos dispõem de quantidade limitada de energia de modo que, aumentar o
investimento em uma atividade requer a diminuição do investimento em outra. O
deslocamento de recursos para uma atividade onerosa como a reprodução poderia levar
a uma demanda conflitante entre reprodução e defesa em plantas, o que implicaria em
maior susceptibilidade aos herbívoros. Observamos indivíduos da planta Miconia dispar
com ramos reprodutivos e vegetativos que exibiam sinais de herbivoria e nos
perguntamos se haveria uma demanda conflitante entre defesa e reprodução. Nossa
hipótese é de que ramos reprodutivos seriam mais suscetíveis à herbivoria que ramos
vegetativos. Para avaliá-la coletamos dois ramos adjacentes com e sem infrutescência de
17 indivíduos. Previmos que a intensidade de herbivoria seria maior nos ramos
reprodutivos. Em cada ramo, estimamos a intensidade de herbivoria das folhas usando
um índice de seis categorias: 0%; 1 – 6%; 6 -12%; 12 – 25%; 25 – 50% e 50 – 100% de
dano foliar. A intensidade de herbivoria entre ramos reprodutivos (média ± dp:
1,9±0,70) e vegetativos (1,6±0,65) foi semelhante (t=1,50; gl=16 p=0,15). Isso indica
que o investimento em reprodução não leva a uma diminuição do investimento em
defesa. A estratégia de vida de uma espécie pioneira como Miconia dispar implica em
investimento preferencial em reprodução e crescimento, dado que seu ciclo de vida é
curto. Consequentemente, investir em defesa pode não ser muito importante. Nesse
caso, a demanda conflitante não deveria ocorrer.

53
Por que o hemíptero Jalisus ossae (Reduviidae) não desencadeia resposta defensiva
da formiga Pheidole minutula em Maieta poeppigii (Melastomataceae)?
Ana Carolina Vieira Pires, Ana Claudia Rorato Vitor, Caroline Marques Dracxler e
Daniel Passos
A planta mirmecófita Maieta poeppigii apresenta mutualismo com formigas Pheidole
minutula. As formigas nidificam nas domáceas foliares e atacam diversos organismos
em resposta ao estímulo mecânico provocado pelo pouso deles sobre a planta. Contudo,
indivíduos do hemíptero Jalisus ossae que habitam essas plantas não são atacados pelas
formigas. Para entender esse fato, propomos a hipótese de que o hemíptero não provoca
estímulo mecânico suficiente para desencadear resposta defensiva. Simulamos pousos
de organismos de diferentes tamanhos sobre a folha de 16 indivíduos de M. poeppigii,
utilizando objetos de mesma densidade com 3, 5, 10 e 15 mm de comprimento, além
dos próprios hemípteros. Contamos as formigas sobre a folha e trinta segundos após a
soltura dos objetos, contabilizamos o recrutamento adicional, esperando que houvesse
aumento na mobilização das formigas com o aumento do tamanho do objeto.
Analisamos as diferenças no recrutamento entre os tratamentos usando GLM com
distribuição de erros Poisson e análise de contraste, considerando cada planta como
bloco. A quantidade de formigas mobilizadas foi afetada pelo comprimento do objeto
(χ2
(4,60)=97,31; p=0,013), indicando maior recrutamento de formigas em estímulos
mecânicos maiores. Os tratamentos com hemípteros e objetos de 3 e 5 mm
desencadearam respostas defensivas menores que 10 e 15 mm (χ2
(1,78)=103,42;
p=0,011). Isso indica que existe um limiar entre 6 e 9 mm que desperta maior
recrutamento de formigas. Por desencadear resposta similar aos objetos de 3 e 5 mm,

54
sugerimos que o impacto do pouso do hemíptero não é suficiente para ser detectado
pelas formigas em M. poeppigii.

55
Recrutamento de formigas na mirmecófita Hirtella sp.: estudo em plantas com
diferentes níveis de estresse
Bruno Travassos, Artur Madeira, Vidal Carrascosa, Renata Rocha
Interações de formigas com plantas mirmecófitas é um mutualismo em que os insetos
beneficiam-se do abrigo e área de forrageio, enquanto a planta é protegida contra
herbívoros. Nesse sistema, plantas debilitadas atraem menos herbívoros, que representa
menos alimento para a colônia de formigas. Dessa forma, pode haver recrutamento mais
intenso de formigas diante da presença de um herbívoro, fazendo com que plantas sob
maior estresse sejam, indiretamente, mais protegidas pelas formigas. Nosso objetivo foi
avaliar se a resposta anti-herbivoria das formigas em Hirtella sp. (Chrysobalanaceae)
aumenta em relação ao nível de estresse da planta. Acreditamos que mais formigas
serão recrutadas em plantas mais debilitadas (menor Massa Foliar Específica-MFE).
Selecionamos 17 plantas na fazenda Dimona, ARIE PDBFF, Amazônia Central. Em
cada planta escolhemos uma folha com pelo menos uma formiga para que ela recrutasse
mais formigas após dano foliar. Para medir intensidade de reação das formigas,
contamos o número de formigas recrutadas quatro minutos após cortarmos um pedaço
da folha. Medimos MFE para indicar estresse das plantas. A quantidade de formigas
recrutadas após o dano variou entre 0 e 25 (Média ±DP=9,8±7) e não foi associada à
MFE (F(1,15)=3,05, p=0,1), que variou entre 3,4 e 9 mg/cm2
(Média±DP=5,6±1,4).
Estudos indicam que herbivoria mais intensa causa maior recrutamento de formigas
para defender a planta. Como nossa simulação de herbivoria teve intensidade igual, a
resposta das formigas não diferiu. O recrutamento de formigas provavelmente evoluiu
em resposta à intensidade do dano causado por herbívoros e não pela condição ou
qualidade da planta hospedeira.

56
Várzea – projetos orientados
A seleção de habitats por cupins em áreas alagadas não visa redução dos custos de
deslocamento até o solo
Sarah Freitas Magalhães, Caroline Marques Dracxler, Renata Pimentel Rocha e
Randolpho Dias Terceiro
Introdução
As estratégias para obtenção de alimento envolvem custos para os animais em termos de
tempo e energia. Segundo a teoria de forrageamento ótimo, como diferentes espécies
têm habilidades distintas na captação de recursos, a seleção natural deveria favorecer
estratégias que maximizem o ganho energético e reduzam os custos associados à sua
obtenção (MacArthur & Pianka 1966). Assim, reduzir o tempo de busca pelo alimento e
concentrar esforços em recursos de maior qualidade eleva o lucro energético
(MacArthur & Pianka 1966).
Os fatores que determinam os lucros e os custos para o organismo, em geral
dependem da sua estratégia de forrageio. Forrageadores de área central, por exemplo,
possuem um local fixo como refúgio, de onde saem para buscar alimento e para onde
retornam depois do forrageio (Chase 1998). Sendo assim, quanto maior a distância do
local de forrageio até o refúgio central, maior deve ser a exposição a predadores e o
gasto energético associado ao deslocamento (Chase 1998).
Cupins são um exemplo de organismos que forrageiam a partir de uma área
central, pois constroem seus ninhos como refúgio e usam túneis para acessar o recurso
nas proximidades (Araújo et al. 2011). Nas áreas alagáveis da Floresta Amazônica

57
existem diversos cupins arborícolas que constroem seus ninhos em troncos próximos ao
dossel e descem para forragear na serapilheira. Como essas áreas são periodicamente
inundáveis, os ninhos de cupins ficam sujeitos ao risco de submersão. Assim, selecionar
áreas de construção do ninho acima do nível atingido pela inundação pode ser vital para
a sobrevivência da colônia. Porém, se os ninhos forem construídos muito acima do nível
máximo de inundação, o gasto energético para forragear no solo após a vazante também
deverá ser grande.
Dentro das regiões alagadas é comum que existam áreas com diferentes
elevações em relação ao nível da água. Como as áreas topograficamente mais altas
possuem menor altura do nível da água em relação ao solo, os cupins que estabelecerem
suas colônias nesses locais, poderiam minimizar a distância do ninho até a serapilheira.
Portanto, nossa hipótese é de que os cupins constroem seus ninhos em árvores de forma
a reduzir o risco de inundação e, ao mesmo tempo, minimizar o gasto energético para
obtenção de alimento. Se a nossa hipótese for verdadeira, esperamos que o número de
ninhos seja maior em locais mais elevados em relação à altura máxima de inundação.
Métodos
Área de estudo
Conduzimos o estudo em uma área de floresta periodicamente inundável, sob influência
dos rios Negro e Solimões, localizada próxima à cidade de Manaus, Brasil. Nessa área
há um ciclo anual de secas e cheias. Durante o período de cheia, que corresponde a
aproximadamente 160 dias, a cota máxima de inundação pode atingir 28 m, enquanto a
cota mínima durante a seca varia de 17 a 19 m (Bittencourt & Amadio 2007).

58
Coleta dos dados
Estabelecemos quatro blocos contendo três parcelas de 20 x 10 m em cada um,
totalizando 12 parcelas. Distribuimos os blocos em diferentes níveis topográficos,
paralelamente a um lago. A distância entre cada bloco e cada parcela foi de 20 e 10 m,
respectivamente. Em cada parcela, medimos a altura máxima alcançada pela inundação
observando marcas presentes nas árvores. Adicionalmente, contamos o número de
ninhos de cupim e registramos a distância de cada cupinzeiro em relação ao nível da
inundação. A fim de controlar uma possível influência da quantidade de troncos sobre o
número de ninhos, contamos o número de troncos que estavam acima do último nível de
inundação.
Para avaliar se o número de ninhos de cupins está associado à elevação
topográfica, realizamos uma análise de covariância (ANCOVA). Consideramos o
número de ninhos como variável resposta, a altura da parcela em relação ao nivel
maximo da água como variável preditora, o número de troncos e os blocos como co-
variáveis.
Resultados
Encontramos 27 ninhos de cupins. A altura máxima da inundação variou entre 4,3 e 6,4
m e o número de árvores entre 7 e 32. O número de ninhos de cupim não esteve
associado à altura do nível de inundação (Tabela 1; Figura 1).

59
Tabela 1. Análise de covariância do número de ninhos de cupins em relação à altura da
parcela removendo os efeitos dos blocos e do número de troncos em uma área alagável
próxima a Manaus, Brasil.
Efeito gl F p
Bloco 3 0,14 0,93
Tronco 1 0,16 0,70
Altura 1 0,05 0,84
Resíduo 6
Figura 1. Relação entre o número de ninhos de cupins localizados em uma floresta
alagável próxima à cidade de Manaus, Brasil (representado pelos resíduos de um
modelo linear entre o número de ninhos de cupins em relação aos blocos e ao número de
troncos) e a altura da inundação (m).
Discussão
A ausência de relação entre o número de ninhos e a altura da inundação indica que os
cupins não ajustam a construção dos ninhos de forma a minimizar o gasto energético
para obtenção de alimento. Em um ambiente periodicamente inundável, estocar
alimento pode ser mais importante para os cupins do que a proximidade com o recurso.

60
Nesse sentido, as reservas provenientes do estoque podem garantir que os cupins
resistam a um longo tempo de cheia, mesmo quando os ninhos se encontram a grandes
distâncias do solo. Visto que a altura máxima da inundação varia anualmente
(Bittencourt & Amadio 2007) e que cupins são forrageadores de área central, perder ou
realocar a área central pode ser mais custoso que construir túneis mais extensos para
chegar até a serapilheira. Assim, a construção dos ninhos acima da altura máxima pode
assegurar a persistência das colônias, evitando a sua submersão, ainda que isso gere um
custo maior para obtenção de alimento.
Interações entre cupins ou com seus predadores parecem afetar fortemente as
características das colônias (Araújo et al. 2007). Portanto, é possível que o
estabelecimento das colônias seja mais dependente dessas interações. De fato, em outras
espécies de animais, a pressão de predação impede que estratégias de forrageio que
maximizem o ganho de energia sejam adotadas (Verdolin 2006).
Agradecimentos
Agradecemos ao Cassiano Rosa pela orientação e aos professores Jack Sparrow
(Fabrício), Paulinho Bom e Monitor Barbudo (Marcos) pela revisão e comentários.
Referências
Araújo, A.P.A., C. Galbiati & O. De Souza. 2007. Neotropical termite species (Isoptera)
richness declining as resource amount rises: food or enemy-free space
constraints? Sociobiology, 49:1-14.
Araújo, A.P.A., F.S. Araújo & O. De Souza. 2011. Resource suitability affecting
foraging area extension in termites (Insecta, Isoptera). Sociobiology, 57:1-14.

61
Bittencourt, M.M & S.A. Amadio. 2007. Proposta para identificação rápida dos
períodos hidrológicos em áreas de várzea do rio Solimões-Amazonas nas
proximidades de Manaus. Acta Amazonica, 37:303-308.
Chase, J.M. 1998. Central-place forager effects on food web dynamics and spatial
pattern in northern California meadows. Ecology, 79:1236-1245.
MacArthur, R.H. & E.R. Pianka. 1966. On optimal use of a patchy environment.
American Naturalist, 100:603-609.
Verdolin, J.L. 2006. Meta-analysis of foraging and predation risk trade-offs in terrestrial
systems. Behavioural Ecology and Sociobiology, 60:457-464.

62
O crescimento vertical da macrófita Eichhornia crassipes (Pontederiaceae) é maior
em ambientes com menor disponibilidade de água
José Hidasi, Artur Madeira, Cibele Paiva e Daniel Passos
Introdução
Plasticidade fenotípica é a variação que existe em determinadas características
morfológicas, anatômicas ou fisiológicas em resposta a variações nas condições
ambientais (Justo et al. 2005). Ela possibilita que organismos se estabeleçam em
ambientes com condições ambientais variáveis (Kemp & Jones 2001). Nesses casos, é
de se esperar que a mudança nos atributos confira maior chance de sobrevivência
quando ocorrem modificações ambientais.
Na região amazônica ocorrem pulsos de inundação que consistem em variações
sazonais no nível da água (Junk 1997). Durante os períodos em que esse nível está mais
baixo ocorre a formação de lagos, onde macrófitas aquáticas ficam isoladas. Nesses
sistemas, os lagos perdem água gradativamente durante toda a fase de vazante. Portanto,
macrófitas que apresentam estratégias para contornar as condições hídricas adversas
podem ser favorecidas, resistindo até o próximo período de inundação.
Uma macrófita comum na Amazônia é a Eichhornia crassipes, que possui
tecidos especializados em seus pecíolos, responsáveis por sua flutuabilidade (Lopes et
al. 2009). As folhas dessa planta são arredondadas na parte basal que fica em contato
direto com a água e afilam gradativamente ao longo do pecíolo no sentido vertical.
Porém, existe uma variação entre indivíduos dessa espécie quanto a essa característica
morfológica nos pecíolos de suas folhas. Alguns indivíduos apresentam pecíolos
estreitos e longos enquanto outros são largos e curtos. Pecíolos largos e curtos podem
estar relacionados com a maior flutuabilidade por aumentarem a área de contato da

63
folha com a água. Por outro lado, pecíolos estreitos e longos podem apresentar maior
área fotossinteticamente ativa devido à maior superfície em contato com raios
luminosos.
Como as condições hídricas de um lago podem variar ao longo do tempo, é
possível que as variações morfológicas nas folhas de E. crassipes representem o ajuste
da planta em resposta à disponibilidade de água. Portanto, nossa hipótese é que E.
crassipes apresenta maior crescimento vertical em regiões menos alagadas. Esperamos
que os indivíduos tenham pecíolos mais estreitos e longos quando encostados à margem
do lago (que representa um ambiente com menor disponibilidade de água), e mais largos
e curtos no centro do lago (onde a disponibilidade de água não é restritiva).
Métodos
Realizamos o estudo em um lago, localizado em uma área de várzea, próxima ao
encontro dos rios Negro e Solimões (3° 09’ S-59° 54’ O), Manaus, AM - Brasil.
Utilizamos como modelo de estudo a macrófita Eichhornia crassipes (Pontederiaceae).
Os indivíduos analisados se encontravam em dois locais distintos: na superfície da água
próximos ao centro do lago ou apoiados no solo da margem.
Coletamos 19 indivíduos de E. crassipes, 10 no centro e nove na margem do
lago. Em cada indivíduo, medimos o comprimento e o diâmetro de três pecíolos por
indivíduo. Posteriormente, calculamos um índice peciolar ao dividir o comprimento
médio pelo diâmetro médio dos três pecíolos coletados. Sendo assim, um maior índice
peciolar representa uma planta com pecíolos estreitos e compridos, e um menor índice
representa uma planta com pecíolos largos e curtos. Para testar se o índice peciolar está
relacionado com diferentes condições de alagamento utilizamos um teste de Mann-
Whitney.

64
Resultados
O índice peciolar das plantas no centro do lago foi de 3,99 ± 1,05 (média ± desvio
padrão), enquanto o das plantas na margem foi de 11,66 ± 8,86 (U = 13,5; p < 0,01;
Figura 1). A variabilidade do índice peciolar para plantas localizadas às margens do
lago foi cerca de oito vezes maior que a encontrada para as plantas que estavam
flutuando na região central do lago.
Figura 1. Índice peciolar de Eichhornia crassipes no centro e na margem de um lago na
várzea do Rio Negro, Manaus, Brasil. Pontos representam as medianas e as barras
representam os quartis de 25% e 75%.
Discussão
A conformação estrutural dos pecíolos de E. crassipes foi distinta nas diferentes
condições de alagamento. O fato de indivíduos de E. crassipes encontrados na margem
terem apresentado pecíolos mais estreitos e longos em relação a indivíduos no centro do
lago, indica um maior investimento dos indivíduos na margem em crescimento vertical.
Centro Margem
Localização no lago
2
4
6
8
10
12
14
16
Índic
e pec
iola
r

65
Sendo assim, a variação morfológica do pecíolo pode constituir uma resposta à
ambientes com condições hídricas distintas.
A plasticidade fenotípica na forma do pecíolo pode refletir um investimento
diferencial em capacidade de flutuação ou área fotossinteticamente ativa. Uma vez que
investir na flutuabilidade não traz vantagens a indivíduos localizados nas margens dos
lagos, a estratégia neste caso pode ser mais direcionada para o crescimento dos pecíolos
e aumento da fotossíntese. Durante os períodos de seca, o aumento da área
fotossinteticamente ativa da planta possibilitaria um maior acúmulo de reservas que
podem permitir a sobrevivência da planta até que o nível de água suba novamente.
Eichhornia crassipes apresenta ampla ocorrência nas regiões tropicais alagáveis
(Sculthorpe 1985). Isso pode ser um reflexo de seu sucesso em se estabelecer em locais
com condições ambientais variáveis (Junk & Piedade 1997; Sculthorpe 1985). Sendo
assim, caso a plasticidade na morfologia da planta seja uma adaptação a ambientes que
secam periodicamente, esperamos que não ocorra essa variação morfológica em regiões
com corpos d’água perenes.
Agradecimentos
Agradecemos aos professores Aline e Paulo Enrique pelas discussões e ideias. Além
disso, agradecemos ao monitor Thiago pela orientação e ajuda no campo.
Referências
Hu C., L. Zhang, D. Hamilton, W. Zhou, T. Yang & D. Zhu. 2007. Physiological
responses induced by copper bioaccumulation in Eichhornia crassipes (Mart.).
Hydrobiologia, 579:211-218.

66
Junk, W.J. 1997. General aspects of floodplain ecology with special reference to
Amazonian floodplain: Ecology of a pulsing system, pp. 3-20. In: The Central
Amazon Floodplain (W.J. Junk). Berlin: Springer.
Junk, W.J. & M.T. Piedade. 1997. Plant life in the floodplain with special reference to
herbaceous plants, pp. 147-181. In: The Central Amazon Floodplain (W.J. Junk).
New York: Springer-Verlag.
Justo, C.F., A.M. Soares, M.L. Gavilanes & E.M. Castro. 2005. Leaf anatomical
plasticity of Xylopia brasiliensis Sprengel (Annonaceae). Acta Botanica
Brasilica, 19:112-123.
Kemp, D. & R.E. Jones. 2001. Phenotypic plasticity in field populations of the tropical
butterfly Hypolimnas bolina (L.). Biological Journal of the Linnean Society,
72:33-45.
Lopes, A., S.M. Rosa-Osman & M.T.F. Piedade. 2009. Effects of crude oil on survival,
morphology, and anatomy of two aquatic macrophytes from the Amazon
floodplains. Hydrobiologia, 636:295-305.
Sculthorpe, C.D. 1985. The biology of aquatic vascular plants. Konigstein: Koeltz
Scientific Books.

67
Ocorrência de grupos funcionais de peixes em dois rios Amazônicos com diferentes
transparências de água
Lorena Pinto, Ludmila Rattis, Tatiane Calaça & Bruno Travassos
Introdução
As espécies podem ser agrupadas conforme características funcionais compatíveis com
condições ambientais específicas. Essas condições, ou filtros ambientais, selecionam
espécies que ocorrerão no ambiente local, a partir de um conjunto regional de espécies
(Wiescher et al. 2012). Com isso, os filtros operam selecionando as espécies com
características adequadas ao ambiente, de modo que o conjunto de espécies em uma
localidade é o resultado dessa seleção (Keddy 1992).
Por causa das restrições impostas pelos filtros ambientais, algumas
características tornam-se mais comuns nas espécies que compõem a comunidade. Essas
características ou atributos funcionais podem ser morfológicos, fisiológicos e
comportamentais e atuam no aumento do desempenho do indivíduo, com efeitos sobre o
crescimento, reprodução e sobrevivência (Violle et al. 2007). Quando diferentes
espécies compartilham mais de um desses atributos formam-se grupos funcionais
(Keddy 1992).
Em ambientes aquáticos, características físicas e químicas da água, como
temperatura, condutividade, acidez, nutrientes dissolvidos e transparência, podem agir
como filtros ambientais (Bronmark & Hansson 2005, Rodriguez & Leweis1997).
Dentre essas características, a diferença de transparência da água pode selecionar
organismos com características sensoriais que favoreçam sua orientação nesse ambiente.
Nos peixes, por exemplo, os olhos são responsáveis pela capacidade de orientação
visual em águas transparentes e órgãos receptores como o aparelho labirinto, barbilhões

68
e a linha lateral são os principais órgãos quimio e tatorreceptores, que auxiliam na
orientação não visual em águas de baixa transparência (Pough et al. 2008). Esses tipos
de orientações sensoriais podem influenciar nas relações interespecíficas, como
predação e competição, afetando assim, a sobrevivência e o sucesso reprodutivo das
populações nos ambientes com transparências distintas.
Na Amazônia Central, os rios são agrupados com base nas características físico-
químicas que determinam a transparência da água. Os rios de águas pretas, como o rio
Negro, apresentam águas mais transparentes devido à baixa quantidade de material em
suspensão, enquanto que rios de água branca, como o rio Solimões, são mais turvos
devido ao excesso de sedimentos (Junk & Furch 1985).
A variação na transparência dos rios pode representar um importante filtro
selecionador de espécies, contribuindo para a estruturação da comunidade de peixes.
Neste estudo, nós analisamos a influência da transparência da água dos rios Negro e
Solimões sobre a ocorrência de grupos funcionais de peixes, de acordo com o tipo de
orientação no ambiente. Nossa hipótese é que espécies de peixes com orientação visual
serão favorecidas no rio de maior transparência, enquanto as espécies do grupo
funcional sem orientação visual serão favorecidas no rio com a água mais turva.
Esperamos encontrar maior riqueza de espécies visuais no rio Negro (rio com maior
transparência) e maior riqueza de espécies não visuais no rio Solimões (rio com menor
transparência).
Métodos
Nós realizamos esse estudo durante o período de seca, na região da confluência entre os
rios Negro e Solimões, próximo a Manaus, Brasil. Escolhemos oito pontos de coleta nas
margens dos rios, sendo quatro no Negro e quatro no Solimões. Em um mesmo rio, os

69
pontos estavam 500 m distantes entre si, com exceção de um dos pontos no rio Negro,
que estava a 1.388 m do ponto mais próximo. Entre os rios a menor distância foi de
1.000 metros.
Em cada ponto, coletamos os peixes com uma passada de rede de arrasto de 10
m de comprimento e malha de 5 mm. Contabilizamos todos os indivíduos coletados e os
identificamos no nível de espécie. Com o auxílio de um especialista, classificamos em
visualmente orientados (ou visuais) os peixes com olhos bem desenvolvidos, e como
não orientados pela visão (não visuais), os peixes que apresentavam barbilhão, olhos
pequenos e sistema de linha lateral desenvolvido.
Para testar o efeito da transparência da água sobre a riqueza de peixes dos dois
grupos funcionais, nós realizamos uma Análise de Variância (ANOVA) hierárquica.
Consideramos os grupos funcionais (visual e não visual) e os rios (Negro e Solimões)
como variáveis preditoras e o número de espécies de peixes como resposta. Os oito
pontos de coleta foram aninhados de acordo com o rio, sendo quatro no Rio Negro (alta
transparência) e quatro no Rio Solimões (baixa transparência).
Resultados
Nós capturamos 293 indivíduos, sendo 254 no rio Solimões e 39 no rio Negro. Os
indivíduos pertenciam a 32 espécies, sendo que destas 23 foram encontradas no rio
Solimões (15 exclusivas), 16 no rio Negro (sete exclusivas). Do total das espécies, nove
(28,1%) foram compartilhadas entre os dois rios. A maioria dos indivíduos coletados
(64,5%, n= 189) pertencia a apenas três espécies que ocorreram em ambos os rios:
Pimelodus blochii (Pimelodidae), Hemidoras stenopeltis e Plagioscion squamosissimus.
Outras 10 espécie somaram 29,4% (n=86) dos indivíduos amostrados (Figura 1).

70
Figura 1. Abundância relativa das 13 espécies de peixes mais abundantes, separadas
pelo tipo de orientação (visual e não visual), coletadas em oito pontos nos rios Negro e
Solimões, Amazonas, Brasil.
Classificamos 21 espécies como visuais e 12 como não visuais. No rio Solimões,
encontramos 14 espécies visuais (41,7% dos indivíduos coletados) e nove não visuais e
no rio Negro, 11 espécies foram visuais (56,4% dos indivíduos) e cinco não visuais. O
número de espécies orientadas e não orientadas pela visão não foi diferente entre os dois
rios (F1, 7 = 0,52; p = 0,49; Figura 2).

71
Figura 2. Riqueza de espécies de peixes com orientação visual e não visual nos Rio
Negro e Solimões, próximo a Manaus, Brasil. Os círculos representam as médias de
riqueza dos peixes orientados visualmente e os quadrados representam as médias dos
peixes não orientados visualmente para os dois rios. As barras verticais mostram o
intervalo de confiança de 95%. A linha tracejada indica a tendência de mudança na
riqueza para os peixes visualmente orientados e a linha contínua a tendência para os
peixes orientados não visualmente.
Discussão
A transparência da água não foi um filtro ambiental para os grupos funcionais de peixes
visuais e não visuais, uma vez que as espécies desses dois grupos ocorreram em ambos
os rios. É possível, que outros aspectos morfológicos, como o sistema de linha lateral ou
fisiológicos, como tolerância a diferentes valores de pH, temperatura e condutividade
(McFall-Ngai 1990), possibilitem o fluxo dos organismos dos dois grupos funcionais
entre os rios.

72
Independente das diferenças nas características da água, estratégias morfo-
fisiológicas podem ser favorecidas para o uso de ambos os tipos de rios. Por exemplo,
espécies com orientação não visual capturadas nos dois rios, como Pimelodus blochii
(Família Pimelodidae) e Hemidoras stenopeltis, apresentavam, além dos barbilhões,
olhos bem desenvolvidos indicando que essas espécies podem usar o tipo de orientação
mais adequada para o nível de transparência do rio. Assim, uma espécie de peixe
poderia usar os barbilhões para a percepção química e tátil em rios com pouca
transparência e a percepção visual em ambientes com maior entrada de luz.
A priori, seria razoável esperar que peixes visualmente orientados fossem
favorecidos em águas transparentes, característica considerada vantajosa tanto para fugir
de predadores como para aproximação às presas (McFall-Ngai 1990; Zuanon et al.
2006). Já os peixes com orientação química e tátil não seriam favorecidos nas águas
transparentes devido à competição com predadores adaptados à transparência, que
seriam potenciais competidores dominantes (Zuanon et al. 2006). No entanto,
independente da orientação sensorial, a riqueza de espécies aumentou no rio Solimões,
ambiente de maior produtividade por causa da alta quantidade de sedimentos em
suspensão (Junk & Furch 1985). Durante o período de seca, peixes de rios de água preta
perdem gordura corporal devido à escassez de alimento (Junk 1985). Dessa forma, a
escolha de um ambiente mais produtivo nessa época pode ser crucial para a
sobrevivência e o sucesso reprodutivo na próxima estação. Peixes que possuem maior
tolerância às diferentes características dos rios, devido a atributos morfo-fisiológicos,
podem migrar para ambientes mais favoráveis, seja para sua proteção contra predadores
ou para obter mais alimento.

73
Agradecimentos
Agradecemos a todos os professores e colegas que colaboraram de alguma forma para o
desenvolvimento desse trabalho. No entanto, nosso muito obrigado mais que especial é
dedicado ao Dr. Jansen Zuanon, que com todo carinho e dedicação, nos levou para o
mundo mágico dos peixes e fez nosso dia ser extremamente divertido!
Referências
Brönmark, C. & L.A. Hansson. 2005. The biology of lakes and pounds. New
York: Oxford University Press.
Junk, W.J. & K. Furch. 1985. The physical and chemical properties of Amazonian
waters and their relationship with the biota, pp.3-17. In: Amazonia: key
environments (G.T. Prance & T. Lovejoy, eds). Oxford: Pergamon Press.
Junk, W.J. 1985. Temporary fat storage, an adaptation of some fish species to the water
level fluctuations and related environmental changes of the Amazon system.
Amazoniana, 9: 315-351.
Keddy, P.A. 1992. Assembly and response rules: two goals for predictive community
ecology. Journal of Vegetation Science, 3: 157-164.
McFall-Ngai, M.J. 1990. Crypsis in the pelagic environment. American Zoology,
30: 175-188.
Pough, F.H., C.M. Janis & J.B. Heiser. 2008. A vida dos vertebrados. 4a ed. São
Paulo: Atheneu Editora.
Rodríguez, M.A. & W.M. Lewis. 1997. Structure of fish assemblages along
environmental gradients in floodplain lakes of the Orinoco River. Ecological
Monographs, 67:109-128.

74
Violle, C., M.L. Navas., D.Vile., E. Kazakou., C. Fortunel., I. Hummel & E. Garnier.
2007. Let the concept of trait be functional! Oikos, 116: 882-892.
Wiescher, P.T., J.M.C. Pearce-Duvet & D.H. Feener. 2012. Assembling an ant
community: species functional traits reflect environmental filtering. Oecologia,
169: 1063-1074.
Zuanon, J., F.A., Bockmann & I. Sazima. 2006. A remarkable sand-dwelling fish
assemblage from central Amazonia, with comments on the evolution of
psammophily in South American freshwater fishes. Neotropical Ichthyology, 4:107-
118.

75
Plantas não sincronizam a produção de frutos de acordo com a vazante em uma
comunidade de várzea no Rio Solimões
Ana Claudia Rorato Vitor, Leonardo Oliveira, Vidal Carrascosa, Mariana Velasque
Introdução
A dispersão é o movimento pelo qual os organismos se distribuem no ambiente. Em
plantas, a dispersão possibilita um distanciamento entre os indivíduos podendo
minimizar a competição intra-específica (Odum & Barret 2007), principalmente entre a
prole e o indivíduo parental (Connell 1971; Janzen 1970). Dependendo do vetor
responsável pela dispersão das sementes, ela pode ser categorizada como zoocórica
(feita por animais), anemocórica (feita pelo vento) ou hidrocórica (quando a semente é
transportada pela água).
Em áreas sujeitas a regimes de inundações periódicas é comum encontrar
estratégias de dispersão de sementes associadas aos pulsos de inundação. Esses pulsos
são resultantes do aumento do nível de grandes rios que alagam a floresta adjacente
(Ferreira 1991). Em função disso, é comum a ocorrência de estratégias de resistência ao
período de cheia na flora local (Junk 1989). Em particular, a fenologia das plantas
normalmente é sincronizada com os eventos de inundação a fim de otimizar a dispersão
das sementes pela água (Adalardo de Oliveira & Daly 2001).
Apesar da sincronização da frutificação com o pico da cheia favorecer a chance
de um propágulo atingir as cotas mais elevadas, isso não seria necessariamente benéfico
para todas as espécies de plantas. Durante a vazante, as cotas mais altas são expostas
primeiro, além de sofrer menor deposição de sedimentos que as cotas mais baixas. Isso
resulta em um gradiente de variação direcional nas condições e recursos das cotas mais
altas para as mais baixas. Como diferentes espécies de plantas possuem necessidades

76
distintas para se desenvolverem adequadamente (Ferreira 1997), para uma planta que se
desenvolve melhor em cotas mais baixas, a liberação de suas sementes no pico da cheia
aumentaria a chance da deposição ocorrer numa cota de inundação mais elevada. Por
outro lado, se a planta ajusta a sua frutificação de acordo com o nível do rio durante a
vazante, a chance de sua semente atingir uma área mais baixa e mais adequada para o
seu estabelecimento seria maior.
As áreas de várzea na bacia Amazônica são um exemplo de um sistema que
sofre a influência do regime sazonal de inundação. Os pulsos de inundação são
determinantes na composição das espécies de plantas, gerando uma sincronização na
produção dos frutos (Kubitzki & Ziburski 1994). Nesse sentido, nos propomos a avaliar
a hipótese de que existe uma sincronia da frutificação das plantas com a redução gradual
do nível das águas. O que resultaria em uma distribuição espacial estratificada das
sementes em diferentes níveis topográficos, pois, a liberação sequencial dos frutos
levaria a uma troca de espécies ao longo dos diferentes níveis topográficos de acordo
com a cota de inundação, mas não levaria a uma troca de espécies dentro de um mesmo
nível. Sendo assim, esperamos encontrar maior similaridade de morfoespécies de
sementes dentro do mesmo nível topográfico, porém em níveis topográficos diferentes
esperamos encontrar baixa similaridade das morfoespécies de sementes.
Métodos
Coleta de dados
Desenvolvemos o estudo em uma praia arenosa na várzea do Rio Solimões, Manaus,
Brasil. Estabelecemos cinco transectos de 200 m paralelos à margem do rio,
representando cinco níveis topográficos distintos. O primeiro transecto estava adjacente
ao nível da água, e o último distante a aproximadamente 50 m acima. Este espaçamento

77
garantia um aumento gradual da altitude dos transectos em relação ao nível da água. Em
cada transecto instalamos 10 parcelas de 10x2 m, com uma distância de 10 m entre si
(Figura 1). Em cada parcela realizamos uma busca ativa de sementes na superfície do
solo. Coletamos as sementes encontradas, e posteriormente as classificamos em
morfoespécies usando características morfológicas externas, como forma, rugosidade,
coloração e presença e disposição de nervuras.
Análises
Para avaliar a similaridade das morfoespécies de sementes entre parcelas e entre níveis
topográficos utilizamos o índice de similaridade de Jaccard, que calcula a porcentagem
de espécies compartilhadas em diferentes locais. Calculamos dois índices de
similaridade: um (βp) representa a média dos índices entre pares de parcelas vizinhas
dentro do mesmo nível topográfico, enquanto o outro (βn) representa a média dos índices
entre pares de parcelas vizinhas entre níveis topográficos diferentes. Este cálculo gerou
quatro valores médios de βn e nove valores médios de βp. Posteriormente, calculamos a
média dos quatro valores de βn (βnmédio) e a média dos nove valores de βp (βpmédio) para
então calcular a razão de similaridade βpmédio / βnmédio. Essa razão representa quantas
vezes, em média, a similaridade de espécies dentro de uma cota é maior que a
similaridade média de espécies entre as cotas distintas. Para testar se a razão de
similaridade observada é diferente de uma razão esperada ao acaso, geramos uma
distribuição nula da razão de similaridade usando de 10000 aleatorizações das matrizes
de presença e ausência das espécies nas parcelas.

78
Figura 1. Esquema da distribuição das parcelas em relação ao rio Solimões, Manaus,
Brasil, (a linha 1 se refere às parcelas mais próximas ao rio). βp representa a
similaridade de espécies entre pares de parcelas imediatamente vizinhas dentro do
mesmo nível topográfico e βn representa a similaridade de espécies entre pares de
parcelas imediatamente vizinhas entre os diferentes níveis.
Resultados
Identificamos 83 morfoespécies distribuídas ao longo das cinco níveis topográficos de
inundação. O índice de similaridade de Jaccard indicou um compartilhamento de 13%
das espécies entre as parcelas de um mesmo nível topográfico (dp = 15%), e um
compartilhamento de 8% das espécies entre níveis topográficos diferentes (dp = 10%).
O valor da razão entre os índices médios de similaridade (βpmédio/ βnmédio) foi de 1,614.
De acordo com o modelo nulo, não encontramos evidências de que a similaridade de
morfoespécies de sementes entre os diferentes níveis topográficos seja menor do que a
similaridade entre as amostras do mesmo nível topográfico (p=0,29).
βp
βn

79
Discussão
A composição das espécies de sementes depositadas ao longo do gradiente de inundação
não apresentou um padrão regular de troca de morfoespécies entre as diferentes cotas de
inundação. Isso indica que as plantas não apresentaram sincronia de frutificação com a
redução gradual do nível da água. Provavelmente todas as plantas frutificam no pico da
cheia, o que produziria o mesmo padrão de troca de sementes ao longo do gradiente
altitudinal (Kubitzki & Ziburski 1994).
Os rios amazônicos possuem uma vazante acelerada, podendo reduzir
rapidamente o nível de água em curtos intervalos de tempo (Junk 1997). Isso implica
em uma diferença curta de tempo em que as plantas ficam submersas nas diferentes
cotas topográficas (Adalardo de Oliveira & Daly 2001). Como o intervalo de descida da
água é curto, eventualmente mesmo pequenos desvios no tempo de frutificação das
plantas que exibissem sincronização poderiam ser suficientes para a maioria das
sementes ser depositada em um local inadequado. Nesse contexto, a liberação das
sementes desde o pico da cheia poderia assegurar que parte delas fosse depositada nas
cotas adequadas independentemente de variações imprevisíveis na taxa de redução do
nível da água.
Na várzea, a composição de plantas adultas varia ao longo das cotas topográficas
de inundação (Junk 1989). Uma vez que os nossos resultados indicam que as sementes
se distribuem ao acaso ao longo do gradiente topográfico, é provável então que o padrão
de troca de espécies existentes para a comunidade de plantas adultas seja gerado por um
processo que atua após a chegada das sementes. Eventualmente, as sementes
depositadas em cotas inadequadas não se desenvolvem ou as plântulas não persistem.

80
Agradecimentos
Gostaríamos de agradecer ao professor Felipe pela orientação, ajuda em campo e na
triagem das morfoespécies. Ao professor Paulo Enrique pela ajuda nas análises. Ao
monitor Marcos pela ajuda e discussão. E ao motorista do barco que nos levou à campo.
Referências
Adalardo de Oliveira, A. & D.C. Daly. 2001. Florestas do Rio Negro. São Paulo:
Schwarcz.
Connell, J.H. 1971. The role of natural enemies in prevent competitive exclusion in
some marine animals and rainforest trees, pp. 298-312. In: Dynamics of
populations (P.J. den Boer & G.R. Gradwell, eds.) Wageningen: Pudoc.
Ferreira, L.V. 1991. O efeito do período de inundação na zonação de comunidades,
fenologia e regeneração em uma floresta de igapó na Amazônia Central.
Dissertação de mestrado, Instituto Nacional de Pesquisa da Amazônia, Manaus.
Ferreira, L.V. 1997. Effects of the duration of flooding on species richness and floristic
composition in three hectares in the Jaú National Park in floodplain forests in
central Amazonia. Biodiversity Conservation, 6:1353–1363.
Junk, W.J. 1989. Flood tolerance and tree distribution in central Amazonian floodplains,
pp. 47–64. In: Tropical forests: botanical dynamics, speciation and diversity
(Holm- Nielsen LB, Nielsen IC & Balslev H, eds). New York: Academic Press.
Junk, W.J. 1997. General aspects of floodplain ecology with special reference to
Amazonian floodplains, pp. 3-17. In: The central Amazon floodplain ecology of
pulsing system (W.J. Junk, ed.). Berlin: Springer.
Janzen, D.H. 1970. Herbivores and the number of tree species in tropical forest. The
American Naturalist, 104:501-528.

81
Kubitzki, K. & A. Ziburski. 1994. Seed dispersal in flood plain forests of Amazonia.
Biotropica, 2:30-43.
Odum, E.P. & G.W. Barrett. 2007. Fundamentos da ecologia. São Paulo: Cengage
Learning.

82
Respostas comportamentais em presas de ninfas de Odonata (Subordem:
Anisoptera)
Bianca Silva, Joselândio Santos, Ana Carolina Vieira & Daniela Coelho
Introdução
A predação é uma força seletiva que pode moldar a morfologia, o comportamento e a
distribuição das presas no ambiente (Verdolin 2006). A simples presença do predador
pode afetar as atividades da presa causando modificações comportamentais, tais como o
incremento de vigilância e/ou redução de atividade de forrageio da presa (Rypstra et al.
2007; Thaler et al. 2012). Assim, a percepção do risco de predação pela presa pode
induzir respostas antipredatórias que diminuem a probabilidade de ataque do predador
(Bednekoff 2007; Tollrian & Harvell 1999). Estudos com aranhas e visitantes florais,
por exemplo, mostram que as espécies de visitantes reconhecem e evitam flores com
presença das aranhas (Golçalves-Souza et al. 2008).
A resposta comportamental da presa depende da orientação sensorial do
predador (Schmidt & Amézquita 2001). Para predadores orientados visualmente, a
movimentação da presa é importante para o sucesso de captura. Assim, na presença de
um predador orientado visualmente, as presas deveriam reduzir sua movimentação de
modo a reduzir o risco de serem percebidas pelo predador.
Em lagos, ninfas de Odonata são predadores generalistas abundantes e
orientados visualmente, que promovem o aumento do risco de predação nesses sistemas
(Bednekoff 2007). Diante disso, o objetivo do nosso trabalho foi avaliar a influência do
risco de predação sobre o comportamento de presas de ninfas de Odonata em uma
comunidade aquática. Esperamos que em locais onde os predadores estejam presentes as
presas reduzam a movimentação.

83
Métodos
Para avaliar o efeito do predador sobre o comportamento das presas, montamos um
experimento com grupos experimentais, nos quais variamos a presença do predador da
seguinte forma: (1) controle, sem o predador: destinado a avaliar como as presas se
comportam naturalmente na ausência do predador; (2) tratamento, com predador:
destinado a avaliar como a presença do predador influencia o comportamento das
presas.
Como sistema de estudo, utilizamos ninfas de Odonata (Anisoptera) (n=20
indivíduos) como predadores. Como presas, utilizamos larvas de Ephemeroptera (n=10
indivíduos) e peixes, de aproximadamente 10 mm, do gênero Triportheus (n=10
indivíduos). Essas espécies eram abundantes e com menor porte que as ninfas de
Odonata. Sendo assim, essas espécies possuem alta probabilidade de servirem como
recurso para as ninfas. Coletamos os organismos utilizados neste estudo com puçás e
peneiras no lago da Comunidade Catalão, cidade de Careiro, Amazonas, Brasil.
Para cada tipo de presa, montamos um experimento independente com cinco
réplicas em cada grupo experimental. Utilizamos bandejas com dimensões de 20 x 30 x
10 cm e nelas despejamos 900 ml de água proveniente do lago onde os indivíduos foram
coletados. Liberamos a ninfa de Odonata e esperamos 2 min até sua aclimatação. Após
este período, liberamos a presa na mesma bandeja e aguardamos 1 min para a sua
aclimatação. Em seguida, iniciamos as observações da movimentação das presas por um
período de 2 min. Consideramos cada movimento realizado pela presa intercalado por
uma pequena pausa como um deslocamento. Quantificamos a movimentação das presas
usando o número de deslocamentos. Para cada espécie de presa, utilizamos um teste t-
Student para avaliar possíveis diferenças entre as médias do número deslocamentos nos
dois grupos experimentais.

84
Resultados
Não houve diferença entre a média do número de deslocamentos por indivíduos de
Ephemeroptera na presença e na ausência do predador (t = -0,88; gl = 8; p = 0,40;
Figura 1).
Figura 1: Número de deslocamentos das ninfas de Ephemeroptera em grupos
experimentais com e sem ninfas de Odonata. Os pontos representam o número médio de
deslocamentos, e as barras representam o desvio padrão.
Para indivíduos de Triportheus sp., a média do número de deslocamentos por
indivíduos na presença e na ausência do predador foi semelhante. (t = -0,21; gl = 8; p =
0,84; Figura 2).

85
Figura 2: Número de deslocamento dos indivíduos de Triportheus sp. em grupos
experimentais com e sem ninfas de Odonata. Os pontos representam o número médio de
deslocamentos, e as barras representam o desvio padrão.
Discussão
Encontramos que as presas não manifestaram diferenças comportamentais diante da
presença das ninfas de Odonata. No meio aquático, presas podem ser guiadas por
estímulos químicos emitidos pelos predadores (Schmidt & Amézquita 2001). Em
situações em que tais sinais encontram-se muito diluídos no meio, a presa pode não
perceber a presença do predador, não alterando, portanto, seu comportamento. Dessa
forma, é possível que as larvas de Ephemeroptera, orientadas quimicamente, tenham
sido incapazes de perceber a presença das ninfas de Odonata. Em presas visualmente
orientadas, o reconhecimento do predador pode ocorrer com base em características
conspícuas do predador (Gonçalves-Souza et al. 2008). A ausência de tais
características pode dificultar o reconhecimento do predador pela presa. Assim, é

86
possível que os indivíduos de Triportheus sp. tenham sido incapazes de perceber as
ninfas de Odonata, já que elas apresentam padrão de coloração críptico (Oliveira 2007).
Uma vez que a percepção de risco pela presa é crucial para a manifestação da
sua resposta antipredatória, sugerimos estudos com tratamentos que variem a
abundância de predadores no meio e considerem também sinais químicos de
coespecíficos mortos. Esses fatores podem influenciar a percepção do risco pelas presas
e afetar sua resposta comportamental (Schmidt & Amézquita 2001). Dessa forma,
estudos que considerem esses fatores poderão ajudar na compreensão da intensidade
mínima do sinal de predação necessária para que as presas apresentem uma resposta
comportamental à presença de predadores.
Agradecimentos
Agradecemos aos professores Tiago Souza e Paulo Estefano que nos orientaram na
escolha e desenvolvimento do projeto e as revisões do professor Glauco, sabendo que
sem a discussão que proporcionaram não teria sido possível a construção do trabalho.
Referências
Bednekoff , P.A. 2007. Foraging in the face of danger, pp 305-329. In: Foraging:
behavior and ecology (Stephens D.W., J.S. Brown, R.C. Ydenberg, eds.).
Chicago: The University of Chicago Press.
Gonçalves-Souza, T., P.M. Omena, J.C. Souza & G.Q. Romero. 2009. Trait-mediated
effects on flowers: artificial spiders deceiver pollinators and decrease plant
fitness. Ecology, 89:240-243.
Oliveira, D.E. 2007. Existe uma demanda conflitante entre a melanina alocada para o
sistema imune e para a camuflagem em larvas da libélula Micrathyria catenata

87
(Odonata: Libellulidae)? In: Livro da Ecologia da Floresta Amazônica (G.
Machado & J.L.C. Camargo, eds). Manaus: PDBFF / INPA.
Rypstra, A.L., J.M. Schmidt, B.D. Reif, J. DeVito & M.H. Persons. 2007. Tradeoffs
involved in site selection and foraging in a wolf spider: effects of substrate
structure and predation risk. Oikos, 116:853-863.
Schmidt, B.R. & A. Amézquita. 2001. Predator-induced behavioural responses: tadpoles
of the neotropical frog Phyllomedusa tarsius do not respond to all predators.
Herpetological Journal, 11:9-15.
Thaler, J.S., S.H. McArt & I. Kaplan. 2012. Compensatory mechanisms for
ameliorating the fundamental trade-off between predator avoidance and
foraging. PNAS, 109:12075-12080.
Tollrian, R. & C.D. Harvell (eds). 1999. The ecology and evolution of inducible
defenses. Princeton: Princeton University Press.
Verdolin, J.L. 2006. Meta-analysis of foraging and predation risk trade-offs in terrestrial
systems. Behavioral Ecology Sociobiology, 60:457-464.

88
Várzea – projetos livres
A herbivoria em Eichhornia crassipes (Pontederiaceae) não depende do estresse
hídrico
Cibele Paiva, Lorena Pinto, Mariana Velasque, Ana Carolina Vieira Pires, Leonardo
Oliveira
Em condições favoráveis plantas podem investir em produção de compostos de defesa,
mas em condições adversas, a produção desses compostos é limitada. Avaliamos se
macrófitas em condições de estresse hídrico são mais atacadas por herbívoros.
Esperamos que a quantidade de água nos tecidos da folha seja inversamente relacionada
com a área foliar perdida por herbivoria. Realizamos esse estudo em um lago do Rio
Solimões, próximo a Manaus. Amostramos três folhas de 16 indivíduos localizados na
borda e no interior do lago. Calculamos a área foliar perdida pela proporção entre área
total da folha e área removida. A área foliar perdida em plantas da borda do lago foi
maior (27,6 ± 21,3 cm) comparada às plantas do interior (9,5 ± 6,44) (t = 2,6; gl = 14;
p= 0,02). A localização das macrófitas foi considerada como indicativo de estresse
hídrico, pois indivíduos da borda do lago tiveram 30,45% menos água no tecido foliar
comparado as do interior. Amostramos uma área de 2,4 cm2
das folhas e quantificamos
a água presente nos tecidos através da diferença do peso úmido e seco da amostra. A
quantidade de água nas folhas não teve relação com a área foliar perdida por herbivoria
(F = 0,41; gl = 14; p = 0,21; r2
= 0,06). Isso indica que os herbívoros não selecionaram
os indivíduos de macrófita pelo nível de estresse hídrico da folha. Possivelmente a
localização da macrófita próximo a borda do lago facilitou o acesso dos herbívoros
aumentando a quantidade de folha predada.

89
A transparência da água como filtro ambiental para a pigmentação de espécies de
peixes em rios amazônicos
Ludmila Rattis, Daniel Passos, Daniela Coelho, Joselândio Santos e Renata Rocha
Características ambientais selecionam a ocorrência de espécies de acordo com o grau de
tolerância dos indivíduos gerando um efeito de filtragem ambiental. Em rios de águas
transparentes, peixes de tegumento opaco podem ser desfavorecidos por serem mais
facilmente detectados e consumidos por predadores visualmente orientados. Portanto,
esperamos que a riqueza de espécies de peixes opacos seja menor em rios com águas
transparentes quando comparada à de rios de águas turvas. Amostramos a ictiofauna
próximo à confluência dos rios Negro (mais transparente) e Solimões (turvas) em
Manaus. Selecionamos sete pontos de coleta nas margens de cada rio, distanciados 500
m entre si, onde coletamos peixes com rede de arrasto de 10 m de comprimento e malha
de 5 mm. Colocamos os peixes sobre uma folha branca com pontos pretos e
consideramos translúcidos aqueles que permitiam a visualização dos pontos através de
seu tegumento. Comparamos a riqueza de peixes translúcidos e opacos entre os dois rios
com uma ANOVA em blocos, considerando as amostras de cada rio como um bloco.
Como esperado, a riqueza de espécies de peixes opacos foi menor no rio Negro (18
espécies) do que no rio Solimões (25 espécies) (F(1,12)=14,7; p=0,002). Portanto, o
padrão encontrado condiz com o processo de filtragem por predação mais intensa sobre
peixes opacos em águas translúcidas. Para testar o processo em si, sugerimos a
realização de experimentos comparando a eficiência de captura de presas opacas e
translúcidas por predadores visualmente orientados ora em aquários contendo água do
rio Negro ora do rio Solimões.

90
Complexidade estrutural do habitat influenciando o padrão de frequência relativa
de espécies em comunidades de formigas
Vidal Carrascosa, Randolpho Dias Terceiro, Bruno Travassos, José Hidasi & Bianca
Ferreira
Habitats estruturalmente mais complexos proporcionam maior variedade de nichos,
permitindo a coexistência de um número maior de espécies com abundâncias mais
similares. Em ambientes alagáveis da Amazônia, a inundação promove uma diferença
de estrutura da vegetação entre áreas que permanecem alagadas por diferentes períodos,
o que pode influenciar as comunidades de formigas. Nossa hipótese é que existirá
menor equidade na abundância relativa de espécies de formigas em áreas com menor
complexidade estrutural. Conduzimos o estudo em duas áreas adjacentes que
permanecem alagadas por diferentes períodos: uma por mais tempo, que possui
vegetação herbácea, e outra por menos tempo, com estrutura arbórea. Em cada área,
instalamos um transecto de 100 m e a cada 10 m depositamos dois tipos de iscas
(sardinha e goiabada) para atrair formigas. Coletamos e morfotipamos as formigas e
calculamos a frequência de registro de cada morfoespécie nas armadilhas. Encontramos
oito morfoespécies, duas no ambiente herbáceo e seis no arbóreo. Nenhuma das
espécies ocorreu nas duas áreas. A frequência de registro diferiu entre as morfoespécies
no ambiente herbáceo, havendo uma morfoespécie sete vezes mais frequente que a
outra. No ambiente arbóreo as frequências relativas de ocorrência foram mais similares.
A presença de poucas espécies encontradas exclusivamente no ambiente herbáceo
indica que esse local pode ser mais restritivo e que eventualmente a espécie mais
abundante è especializada a esse tipo de ambiente. Sendo assim, as poucas espécies de

91
formigas capazes de colonizar esse local podem apresentar alta abundância devido à
baixa competição com outras espécies nesse local.

92
Plantas sujeitas ao alagamento sazonal possuem menor variação no tamanho das
folhas
Caroline Marques Dracxler, Artur Alcântara Madeira, Tatiane Gomes Calaça Menezes,
Sarah Freitas Magalhães e Ana Claúdia Rorato
Estratégias como investimento em área foliar para otimizar a captação de energia podem
auxiliar as plantas a ocupar e persistir em ambientes sazonais. Em florestas tropicais,
plantas podem apresentar plasticidade fenotípica na morfologia foliar em resposta a
diferentes características ambientais. Em plantas sujeitas ao alagamento sazonal, folhas
no topo da planta são as últimas a submergir e as primeiras a emergir durante o ciclo
hidrológico. Assim, seria esperado que a área foliar dos ramos mais altos de uma planta
de área alagável seja maior que a dos ramos mais baixos, de modo a maximizar a
fotossíntese durante a vazante. Nós usamos uma espécie de Myrtaceae como modelo de
estudo em uma área de várzea próxima à Manaus, Amazonas, para avaliar se existe
variação no tamanho da folha em diferentes estratos da planta. Coletamos cinco ramos
de 19 indivíduos adultos em duas alturas da planta (1,5-2,0 m e 3,5-4,0 m) e
mensuramos a área foliar média a partir de 10 folhas de cada estrato. A média da área
foliar no estrato inferior da planta foi de 3,73±0,82 cm² (Média±DP) e de 3,72±1,01cm²
no estrato superior. Não houve relação entre a área foliar e sua posição na planta (t=-
0,03; gl=18; p=0,97). Os resultados indicam que a morfologia foliar não representa uma
vantagem para resistir à inundação sazonal. Independente da forma e do tamanho da
folha, a atividade metabólica das plantas é reduzida devido à hipóxia ou anóxia no
entorno das raízes durante o período da cheia, o que impossibilita a fotossíntese.

93
Anavilhanas – projetos orientados
A coloração alar é um indicativo da qualidade dos machos em Diastatops sp.
(Odonata: Libelullidae)
Daniela P. Coelho, Leonardo Oliveira, José Hidasi e Joselândio C. Santos
Introdução
Muitas espécies de animais possuem machos com características exageradas que, em
princípio, conferem uma menor chance de sobrevivência, pois os expõe a maiores riscos
de predação. Supostamente tais características foram selecionadas por aumentarem a
chance do macho ser escolhido pelas fêmeas. No entanto, isso gera uma questão sobre
por que fêmeas escolheriam machos com características que confeririam menor
sobrevivência. Nesse sentido, foi proposta a hipótese de indicação de qualidade que
postula que para as fêmeas, características chamativas expressas nos machos seriam um
indicativo de maior habilidade de sobrevivência e escape a predação e, portanto, um
indicativo de parceiros de maior qualidade. Desde que a qualidade seja geneticamente
determinada, os benefícios dessa escolha serão herdados pela prole (Contreras-Garduño
et al. 2006; Contreras-Garduño et al. 2008; Schultz & Fincke 2009).
Os atributos indicativos de qualidade do indivíduo podem variar entre as
espécies, tais como chifres de queratina em escarabeídeos, chifres córneos em alguns
mamíferos, patas posteriores superdesenvolvidas em alguns crustáceos e, ainda, a
pigmentação expressa nas asas em alguns grupos de artrópodes (Emlen 2008). Em
particular, o padrão conspícuo de cores nas asas pode indicar melhor saúde e estado
nutricional do macho (e.g. Contreras-Garduño et al. 2006; Contreras-Garduño et al.
2008). Dessa forma, machos mais conspícuos exibem características que indicam

94
indiretamente maiores reservas energéticas, melhor resposta imunológica ou maior
massa muscular (Andersson 2006; Schultz & Fincke 2009).
Libélulas da espécie Diastatops sp. (Anisoptera: Libelullidae) presentes no
arquipélago de Anavilhanas possuem dimorfismo sexual com relação à coloração das
asas. Machos exibem coloração alar avermelhada, enquanto às fêmeas apresentam uma
coloração predominantemente marrom. Esse dimorfismo sexual indica que existe algum
tipo de pressão de seleção sexual nos machos. Uma possibilidade para isso é que as
fêmeas avaliem os parceiros sexuais com base na coloração alar. Nesse sentido, nossa
hipótese é de que machos mais conspícuos possuem melhor qualidade.
Métodos
Coleta de dados
Realizamos o estudo no arquipélago de Anavilhanas, próximo à base do lago do Prato,
Amazonas, Brasil. Coletamos 14 machos da libélula Diastatops sp. localizados em
manchas de sol no sub-bosque de uma área de mata entre as 8:30 h e 10:45 h.
Dissecamos cada macho para a retirada de suas asas posteriores, tórax e abdômen.
Fizemos fotografias das asas dos indivíduos amostrados e usamos o programa Adobe
Photoshop para calcular a proporção de cor vermelha na asa como a razão entre o
número de pixels vermelhos e o número total de pixels na asa.
Avaliamos a qualidade dos machos em relação a três variáveis: (1) atividade do
sistema imunológico, (2) quantidade relativa de gordura e músculo no tórax e (3)
quantidade relativa de gordura e músculo no abdômen. Imediatamente após a captura de
cada macho, inserimos um implante feito com um fio de náilon (1 cm de comprimento)
na região abdominal para estimar a habilidade imunológica. Este implante permaneceu
durante um período de 2,5 h. Após a remoção do implante, colocamos o fio de náilon

95
sobre um papel milimetrado e utilizamos os 5 mm finais para a quantificação da
proporção de área do fio coberta por melanina. Para esse calculo, consideramos que
cada milímetro do implante correspondia a uma unidade. Para cada unidade, atribuímos
um valor que poderia variar de zero a dois: zero correspondia à ausência de depósitos de
melanina, um correspondia à presença de depósito de melanina em apenas um dos lados
do fio de náilon e dois correspondia a depósitos de melanina em ambos os lados (Figura
1). Caso houvesse deposição de melanina na ponta do implante, adicionávamos um ao
somatório. A medida de intensidade de deposição de melanina era o somatório do
número de unidades dividido por 11, que era o número máximo de unidades
potencialmente cobertas por melanina. Como a resposta imunológica em libélulas é
manifestada pela deposição de melanina na superfície do implante (Contreras-Garduño
et al. 2006), esperávamos que machos com maior proporção de cor vermelha na asa
apresentassem maior porcentagem de área coberta por melanina no implante.
Figura 1. Desenho esquemático da medida de deposição de melanina em um implante,
que representa a resposta imunológica de libélulas da espécie Diastatops sp. Das 11
quadrículas totais, seis apresentam pontos melanizados, o que corresponde a 54,5% de
melanização no implante.
Para estimar a quantidade de gordura relativa acumulada no tórax, calculamos a
sua densidade. Para esse cálculo, medimos a massa torácica em uma balança semi-

96
analítica (precisão de 1 mg) e dividimos o valor obtido pelo seu volume. Para o cálculo
do volume, consideramos que o tórax possui a forma de um paralelepípedo. Assim,
estimamos o seu volume pela fórmula: volume do tórax = altura x largura x
comprimento do tórax. Já para abdômen, consideramos que ele possui forma cilíndrica e
estimamos o volume pela fórmula: volume do abdômen = comprimento x raio x valor
de . Como uma maior massa de lipídeos diminui a densidade, indivíduos com alta
quantidade de gordura terão valores mais baixos de densidade.
Análise dos dados
Nossa variável preditora foi proporção de cor vermelha na asa e nossas variáveis
respostas foram densidade do tórax, densidade do abdômen e proporção de área
melanizada do implante feito no abdômen das libélulas. Com isso, fizemos três
regressões simples, relacionando a proporção de cor vermelha com cada variável
resposta.
Resultados
A porcentagem média (± desvio padrão) de cor vermelha nas asas dos machos de
Diastatops sp. foi de 31,7 ± 8,8 %. A proporção de área melanizada do implante foi de
0,41 ± 0,25 %. A densidade média do tórax foi de 0,0006 ± 0,0008 g/mm³ e do
abdômen foi de 0,0002 ± 0,00004 g/mm³.
A proporção de vermelho na asa não esteve associada à proporção de área
coberta por melanina do implante (F = 0,048; P < 0,829; R² = 0,004) ou à densidade do
tórax (F = 3,637; P < 0,081; R² = 0,232). Por outro lado, a proporção de vermelho na
asa esteve positivamente relacionada com a densidade do abdômen (F = 27,523; p <
0,001; R² = 0,697; Figura 2).

97
(A)
(B)
(C)

98
Figura 2. Relação entre a proporção de vermelho nas asas de machos da libélula
Diastatops sp. e a porcentagem de área melanizada (A), densidade do tórax (B) e
densidade do abdômen (C).
Discussão
A ausência de relação entre proporção de área vermelha nas asas e a proporção de
cobertura por melanina indica que a conspicuidade alar de machos de Diastatops sp. não
é uma pista da intensidade da resposta imune. Em relação à densidade do tórax e a
proporção de vermelho nas asas, o direcionamento dessa relação se deu de acordo com a
tendência esperada pelas nossas previsões. No entanto, uma vez que a relação não
aparentou ser muito acentuada, é possível que o investimento em gordura dos machos
também não seja sinalizado pela conspicuidade das asas. Por outro lado, a relação
positiva entre densidade do abdômen e a proporção de vermelho nas asas indica que a
conspicuidade é um indicativo de um maior investimento em musculatura abdominal.
A resposta imunológica em libélulas é mediada por melanina, que é produzida a
partir dos estoques de gordura do macho (Siva-Jothy 2000). Porém, é possível que esses
atributos variem de acordo com a idade ou com o estado nutricional do adulto (e.g.
Peixoto & Benson 2008). Uma vez que a asa possui um padrão de coloração fixo, se
essas características de qualidade variarem nos adultos, a asa não seria um indicativo
confiável desses atributos. Consequentemente, a escolha dos machos com asas
conspícuas pelas fêmeas de forma a selecionar indivíduos com maior resistência imune
ou investimento em gordura no tórax não deveria ser favorecida.
A musculatura das libélulas, por sua vez, é primariamente determinada pelas
condições de desenvolvimento da larva e, portanto, não deve variar ao longo da vida de
um macho adulto (Córdoba-Aguilar 2008). Consequentemente, como o abdômen não

99
tem a função de acúmulo de gordura, a sua densidade deve variar pouco com a
senescência. Dessa forma, a informação sobre o investimento muscular no abdómen
fornecida pela conspicuidade das asas seria um indicativo confiável da sua qualidade.
Machos com maior musculatura abdominal eventualmente podem se aquecer mais
rapidamente durante o voo. Isso pode permitir maior sucesso durante confrontos
territoriais e consequentemente, possibilitar o aumento do sucesso reprodutivo (e.g.
Schultz & Fincke 2009).
Em geral, as características sexuais secundárias dos machos de libélulas indicam
qualidade associada a reservas energéticas e resposta imune (Contreras-Garduño et al.
2006). Nossos resultados mostraram que outros traços de qualidade podem ser
sinalizados. Talvez a expressão dos atributos de qualidade sinalizados pelas
características sexuais secundárias ocorra principalmente em características
determinadas pelas larvas e que se mantenham constantes ao longo da vida do adulto,
desde que elas confiram maior sucesso reprodutivo ao indivíduo.
Agradecimentos
Agradecemos à orientação do Glauco Machado, por todo seu conhecimento e paciência
em nos ensinar a contar esta história. Agradecemos também ao Paulinho Bom pelo
auxílio em campo e revisão do manuscrito
Referências
Andersson, M. 2006. Condition-dependent indicators in sexual selection: development
of theory and tests, pp. 255-270. In: Essays in Animal behaviour (J.R. Lucas &
L.W. Simmons, eds.). California: Elsevier Academic Press.

100
Bateman, A.J. 1948. Intra-sexual selection in Drosophylla. Heredity, 2:349-368.
Contreras-Garduño, J., J. Canales-Lazcano & A. Cordoba-Aguilar. 2006. Wing
pigmentation, immune ability, fat reserves and territorial status in males of the
rubyspot damselfly, Hetaerina americana. Journal of Ethololy, 24:165–173.
Contreras-Gardunño, J., B.A. Buzatto, M.A. Serrano-Meneses, K. Nájera-Cordero & A.
Córdoba-Aguilar. 2008. The size of the red wing spot of the American rubyspot
as a heightened condition-dependent ornament. Behavioral Ecology, 26:724-
732.
Córdoda-Aguilar. A.2008. Dragonflies & Damselflies. Oxford: Oxford Biology.
Emlen, D. J. 2008. The Evolution of Animal Weapons. Annual Review of Ecology,
Evolution and Systematics, 39:387–413.
Peixoto, P.E.C. & W.W. Benson. 2008. Body mass and not wing length predicts
territorial success in a tropical satyrine butterfly. Ethology, 114:1069–1077.
Schultz, T.D. & O.M. Fincke. 2009. Structural colours create a flashing cue for sexual
recognition and male quality in a Neotropical giant damselfly. Functional
Ecology, 23:724–732.

101
A escolha do local de oviposição de um lepidóptero é influenciada pela
disponibilidade de recurso
Vidal Carrascosa, Sarah Freitas Magalhães, Renata Pimentel Rocha e Tatiane Gomes
Calaça Menezes
Introdução
Habitats distintos podem oferecer diferentes quantidades de alimento e apresentar
diferenças quanto ao risco de predação. Assim, o crescimento, a reprodução e a
sobrevivência dos indivíduos dependem do habitat em que eles vivem. Em muitos
animais, a seleção natural favoreceu a capacidade de avaliar diferentes habitats e
selecionar aqueles mais vantajosos. Dessa forma, a seleção de habitat por um individuo
é um importante fator que determina o seu sucesso reprodutivo (Sih 1980, Doak 2006).
Um componente importante da seleção de habitat é determinar o local adequado
para o desenvolvimento da progênie. A prole se beneficia da escolha de habitats com
alta qualidade, tais como aqueles com grande quantidade de abrigos contra a predação
ou grande quantidade de alimento disponível.
Em alguns grupos de animais, a seleção de habitat deve ser mais criteriosa,
como aqueles cuja progênie apresenta baixa capacidade de locomoção. Um exemplo de
animais com mobilidade reduzida são os lepidópteros. Durante a fase imatura, as
lagartas tendem a permanecer próximas ao local de nascimento. Assim, as plantas onde
os parentais ovipositam representam o primeiro recurso para a prole (Heisswolf et al.
2005) . Desse modo, para garantir o desenvolvimento da progênie, podemos inferir que
o sítio de oviposição deve apresentar alimento abundante para permitir o
desenvolvimento das lagartas até a fase de pupa (Kemp 1998).

102
Nas áreas sob a influência do rio Negro, existem diversas praias com gramíneas
que são utilizadas como alimento por lepidópteros. Essas plantas ocorrem em diferentes
padrões de distribuição, com densidades distintas. Nós questionamos se a
disponibilidade de recurso é um fator de seleção do local de oviposição em uma espécie
de lepidóptero. Nossa hipótese é que o lepidóptero deve ovipositar preferencialmente
em locais onde há uma alta disponibilidade de alimento para sua prole. Assim,
prevemos encontrar lagartas preferencialmente em locais onde as plantas são maiores e
mais adensadas.
Métodos
Conduzimos o estudo no Parque Nacional de Anavilhanas, Amazonas, Brasil. Em uma
praia onde observamos lagartas consumindo gramíneas, estabelecemos uma parcela de
50 x 8 m. A cada 5 m estabelecemos duas subparcelas de 30 x 30 cm, distantes 7 m
entre si, totalizando 20 parcelas. Em cada subparcela, registramos a presença ou
ausência de lagartas e medimos a densidade (nº de plantas por cm2) e a altura das
plantas. Consideramos a presença de lagartas como um indicativo do local de
oviposição, pois uma vez que a capacidade de locomoção das lagartas é limitada, o local
onde eclodem não deveria diferir muito de onde foram encontradas.
Para testar se a presença de lagartas estava associada com a densidade e a altura
das plantas, fizemos uma regressão logística para cada variável, utilizando a densidade e
altura média das plantas como variáveis preditoras e a presença/ausência de lagartas
como variável resposta.

103
Resultados
Registramos lagartas em 10 parcelas, sendo que o número máximo de lagartas numa
mesma parcela foi cinco. A quantidade média (± DP) de plantas nas subparcelas com
lagartas foi de 53,9 ± 26,9 e a altura média de 22,1 ± 2,6 cm. Nas parcelas sem lagartas,
a quantidade média de plantas foi 20,1 ± 21,7 e altura média foi 18,9 ± 3,2 cm. A
presença de lagartas foi mais frequente em locais com maior densidade de plantas (χ² =
7,12; gl = 1; p < 0,01; Figura 1) e com plantas mais altas (χ² = 5,30; gl = 1; p = 0,02;
Figura 2).
Figura 1. Relação entre a presença/ausência de lagartas e a densidade de gramíneas em
uma praia do arquipélago de Anavilhanas, Brasil. Os números 0 e 1 indicam ausência e
presença de lagartas, respectivamente.
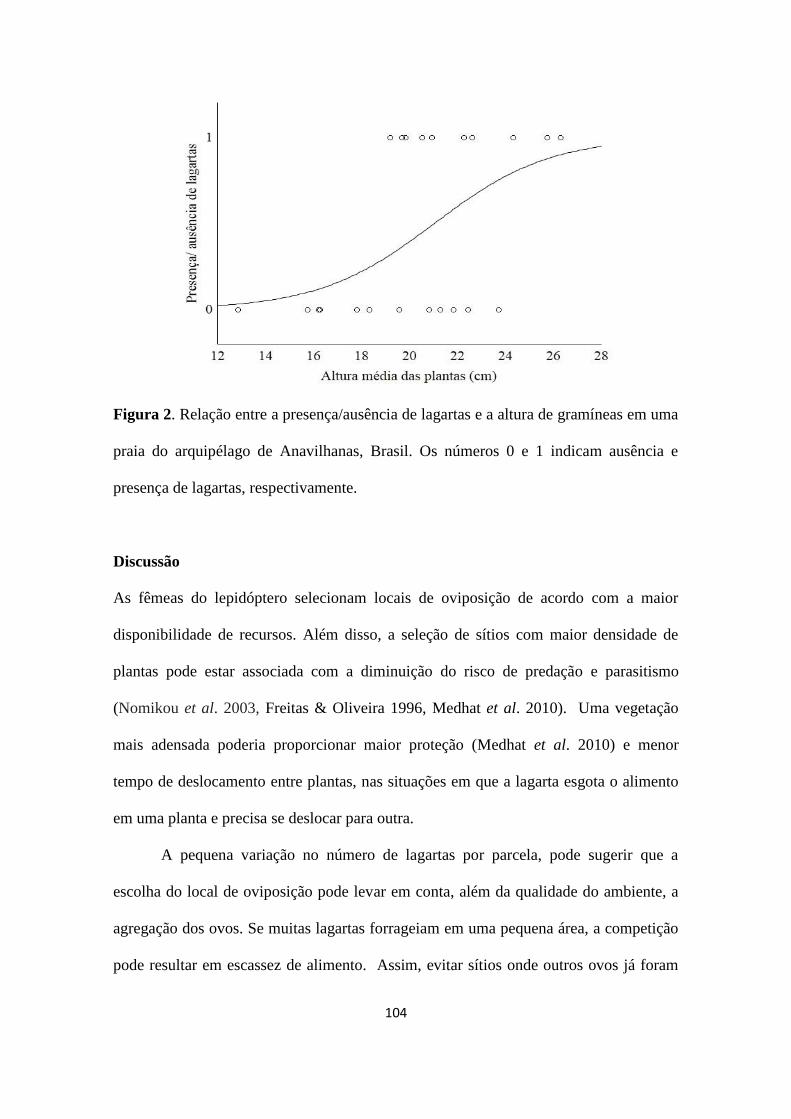
104
Figura 2. Relação entre a presença/ausência de lagartas e a altura de gramíneas em uma
praia do arquipélago de Anavilhanas, Brasil. Os números 0 e 1 indicam ausência e
presença de lagartas, respectivamente.
Discussão
As fêmeas do lepidóptero selecionam locais de oviposição de acordo com a maior
disponibilidade de recursos. Além disso, a seleção de sítios com maior densidade de
plantas pode estar associada com a diminuição do risco de predação e parasitismo
(Nomikou et al. 2003, Freitas & Oliveira 1996, Medhat et al. 2010). Uma vegetação
mais adensada poderia proporcionar maior proteção (Medhat et al. 2010) e menor
tempo de deslocamento entre plantas, nas situações em que a lagarta esgota o alimento
em uma planta e precisa se deslocar para outra.
A pequena variação no número de lagartas por parcela, pode sugerir que a
escolha do local de oviposição pode levar em conta, além da qualidade do ambiente, a
agregação dos ovos. Se muitas lagartas forrageiam em uma pequena área, a competição
pode resultar em escassez de alimento. Assim, evitar sítios onde outros ovos já foram

105
depositados pode conferir vantagens ao minimizar a competição por recursos entre as
lagartas (Janz 2002).
Nossa abordagem de seleção de habitat em um ambiente dominado por uma
única espécie difere da maioria dos trabalhos sobre o tema, que são executados em
locais com maior heterogeneidade ambiental. Constatamos que lepidópteros são capazes
de selecionar o ambiente favorável para o desenvolvimento da prole mesmo um
ambiente com pouca variação estrutural da vegetação. Isso evidencia a refinada
capacidade de seleção do local de oviposição que esses organismos possuem.
Agradecimentos
Agradecemos a Marcos Vieira, Thiago Kloss e Paulo Estefano pela orientação, ajuda e
correções durante o desenvolvimento deste projeto.
Referências
Doak, P., P. Kareiva & J. Kingsolver. 2006. Fitness consequences of choosy oviposition
for a time-limited butterfly. Ecology, 87:395-408.
Freitas, A.V.L. & P.S. Oliveira. 1996. Ants as Selective Agents on Herbivore Biology:
Effects on the Behaviour of a Non-Myrmecophilous Butterfly. Journal of Animal
Ecology, 65:205-228.
Heisswolf, A., E. Obermaier & H.J. Poethke. 2005. Selection of large host plants for
oviposition by a monophagous leaf beetle: nutritional quality or enemy-free space?
Ecological Entomology, 30:299-306.
Janz, N. 2002. Evolutionary ecology of oviposition strategies, pp. 349-376. In:
Chemoecology of insect eggs and egg deposition (M. Hilker & T. Meiners, eds.).
Berlin:Blackwell.

106
Kemp, D.J. 1998. Oviposition behaviour of post-diapause Hypolimnas bolina (L.)
(Lepidoptera : Nymphalidae) in tropical Australia. Australian Journal of Zoology,
46:451-459.
Medhat, M.S., B.S. Hanssond & P. Anderson. 2010. Does risk of egg parasitism affect
choice of oviposition sites by a moth? A field and laboratory study. Basic and
Applied Ecology, 11:135–143.
Nomikou, M., A. Janssen, M.W. Sabelis. 2003. Herbivore host plant selection: whitefly
learns to avoid host plants that harbour predators of her offspring. Behavioural
Ecology,136:484-488.
Sih, Andrew. 1980. Optimal behavior: Can foragers balance two conflicting demands?
Science, 20:1041-1043.

107
Diferentes períodos de inundação influenciam resistência de folhas em
plantas no igapó
Bruno Travassos, Ana Claudia Rorato, Mariana Velasque e Ana Carolina Vieira Pires
Introdução
Os padrões locais de riqueza e abundância dos organismos são determinados por fatores
bióticos e abióticos, que favorecem a ocorrência das espécies mais tolerantes às
condições locais, e excluindo as espécies não tolerantes (Townsend et al. 2006). Filtro
ambiental é o processo através do qual condições bióticas e/ou abióticas locais,
restringem a sobrevivência de determinado grupo de organismos, fazendo com que o
número de espécies de um local seja menor do que o número de espécies presente na
região. De maneira geral, um filtro seleciona apenas organismos que possuam atributos
que lhes permitem sobreviver sob certas condições restritivas de um local.
Os organismos selecionados em um determinado local terão atributos morfo-
fisiológicos comuns que favorecem a sobrevivência desses organismos (Lebrija-Trejos
et al. 2010). Esses atributos são chamados de atributos funcionais (Cornelissen et al.
2003, Cianciaruso et al. 2009, Violle et al. 2007). A limitação da variação do estado dos
atributos funcionais pode ser ocasionada por distúrbios que modificam condições
ambientais periodicamente funcionando como filtros ambientais.
As florestas de igapó no baixo Rio Negro estão sujeitas a pulsos de inundação
sazonais. Esses pulsos têm amplitude média de 10 m e podem durar até sete meses
(Goulding et al. 1988). Os longos períodos de inundação podem restringir a ocorrência
de certas espécies, selecionando aquelas que possuem atributos funcionais que conferem
tolerância à inundação (Shongart et al. 2002). Às margens do rio, a comunidade de
plantas presentes em áreas de menor altitude estão sujeitas a um maior período de

108
inundação. Já nas áreas de maior altitude, o período de inundação é mais curto e as
árvores mais altas podem não ser totalmente recobertas pela água. Como as plantas
submersas ficam mais expostas à abrasão e ao apodrecimento dos seus tecidos (Parolin
2009), o ambiente que passa mais tempo submerso é mais restritivo que o ambiente que
passa menos tempo submerso.
Resistência mecânica das folhas pode ser considerada um atributo funcional
vantajoso para plantas que passam longo período de submersão (Junk et al. 1989, Junk
1997). Portanto, plantas com folhas mais resistentes seriam selecionadas positivamente
nas áreas mais baixas, que ficam mais tempo inundadas. Assim, a variação total dos
atributos funcionais encontrados na comunidade regional deve estar mais bem
representada nas áreas mais altas do que nas áreas mais baixas. O objetivo do nosso
estudo foi investigar se em áreas inundadas há seleção de plantas com folhas mais
resistentes. Nossas previsões são: (i) a força necessária para a ruptura das folhas será
maior em áreas sujeitas a maior tempo de inundação; (ii) a variação da força de ruptura
das folhas será maior em áreas com menor tempo de inundação.
Métodos
Área de estudo
Conduzimos nosso estudo no Parque Nacional do Arquipélago de Anavilhanas,
localizado no baixo Rio Negro, na Amazônia brasileira (02º47’ S - 60º48’ O). Essa
bacia está sujeita a inundações semestrais que podem durar até 270 dias (Junk 2000).
Devido à elevada amplitude de variação no nível da água, as árvores localizadas
próximo à margem do rio ficam total ou parcialmente submersas durante o período da
cheia.

109
Coleta de dados
Selecionamos duas áreas de floresta, uma em cada margem do rio. As duas áreas tem
regimes de inundação distintos: uma das áreas é mais elevada e, portanto, fica menos
tempo inundada que a outra área. Em cada área demarcamos uma parcela de 200 m2,
dentro da qual coletamos um ramo de todos os indivíduos arbóreos com altura mínima
de 1 m. Posteriormente, morfotipamos as plantas através de atributos morfológicos das
folhas e selecionamos uma folha madura de cada morfotipo para medir a resistência das
folhas. Para isso confeccionamos um instrumento para medir a força necessária para
rasgar a folha. Esse aparelho consistiu em um dinamômetro com um Erlenmeyer
acoplado na extremidade inferior. Recortamos uma tira de tamanho padronizado (5 cm
x 1 cm) do limbo de cada folha e fixamos uma extremidade superior da tira a um ponto
fixo. Penduramos o dinamômetro com Erlenmeyer na parte inferior da folha. Em
seguida despejamos a água no recipiente, com velocidade constante até que a folha se
rompesse e registramos o volume de água necessário para essa ruptura. O volume de
água foi convertido em força, sendo que um litro de água equivale a 10000 N.
Análise dos dados
Utilizamos o teste-t para analisar a diferença entre a média da força de fratura das folhas
provindas das áreas com diferentes cotas de inundações. Para testar a diferença das
variâncias da força de fratura nas duas áreas usamos o teste de Levene.
Resultados
A força de fratura das folhas foi maior para as folhas provindas da área mais baixa do
que as folhas da área mais alta e com menor tempo de inundação (t=1,62; gl=58;
p=0,03). A força média necessária para a ruptura das folhas na área de maior altitude foi

110
4143,3 ± 1222,3 N e na área mais alta foi 3396,7 ± 1443,2 N. A variação da força de
ruptura das folhas não foi diferente entre as duas áreas (F(1,58)=2,59; p=0,11; Figura 1).
Figura 1. Força necessária para fraturar folhas de plantas provindas de terrenos de
diferentes elevações no Arquipélago de Anavilhanas. Cada ponto representa uma folha
de cada morfotipo. As médias da força necessária para fratura de folha em cada área
estão representadas pelas barras horizontais.
Discussão
Plantas que passam mais tempo submersas apresentaram folhas mais resistentes do que
aquelas que ficam submersas por períodos mais curtos. Isso pode indicar uma resposta a
diferentes pressões entre os dois ambientes. Assim, plantas que investem muito em
resistência foliar, de maneira geral têm um crescimento mais lento (Coley et al. 1986).
Portanto, sacrificar a taxa de crescimento em uma área onde a resistência foliar não é
vital pode ser uma desvantagem. Em plantas que se encontram submersas por menos
tempo, o investimento em folhas mais resistentes pode reduzir o investimento em outras
Bai
xaAlta
0
2000
4000
6000
8000
Elevação do terreno
Fo
rça d
e f
ratu
ra (
N)

111
características, como crescimento rápido. Para plantas, o crescimento rápido implica em
um aumento na capacidade de competição por luz (Hubbel et al. 1999) e no caso das
plantas em áreas alagadas, a altura da planta pode significar escapar da submersão.
Desta forma, a produção de folhas menos resistentes em plantas que ficam menos tempo
submersas seria favorecida, permitindo alocação de recursos para outras funções.
Enquanto nas áreas baixas, o regime de inundação favorece plantas que investem mais
em folhas resistentes, pois durante os eventos de cheia, as folhas poderiam ser
danificadas ou até mesmo perdidas, comprometendo a aquisição de energia na época em
que ficam emersas.
Comunidades vegetais que estão presentes em locais que sofrem diferentes
intensidades de inundação podem estar submetidas a restrições semelhantes, mas que
selecionam atributos em sentidos opostos. Em áreas que ficam muito tempo inundadas,
o pulso de inundação funciona como filtro selecionando plantas com folhas resistentes.
Já em áreas que ficam pouco tempo inundadas, é provável que a competição por luz
esteja funcionando como filtro, restringindo a variação na resistência das folhas em
outro sentido. Nesse caso o filtro estaria atuando na seleção de plantas com crescimento
rápido, que são melhores competidoras por luz, mas consequentemente têm folhas
menos resistentes.
Nossos resultados levantam a questão de como se distribuem as espécies no
Igapó. As duas cotas de inundação podem conter conjuntos de espécies distintos cujas
espécies possuem folhas com diferentes níveis de resistência. Assim, em áreas com
estresse hídrico maior, espécies com folhas mais resistentes estariam sendo
selecionadas. A segunda hipótese é que as duas cotas de inundação possuem o mesmo
conjunto de espécies, porém os indivíduos dessas espécies são plásticos o suficiente

112
para expressarem diferentes resistências de folhas conforme a exposição ao estresse
hídrico diferentes (Metamoros & Moreno 2001).
Agradecimentos
Gostaríamos de agradecer a Catá pela orientação, ajuda em campo e na triagem das
morfoespécies. À pesola quebrada, à Dona Eduarda por ceder nosso “Erlenmeyer” e ao
Baby pelas 43 correções impressas do nosso trabalho.
Referências
Belyea, L.R. & J. Lancaster. 1999. Assembly rules within a contingent ecology. Oikos,
86:402-416.
Cianciaruso, M.V., I.A. Silva & M.A. Batalha. 2009. Diversidades filogenética e
funcional: novas abordagens para a ecologia de comunidades. Biota Neotropica,
9:93-103.
Coley, P.D., J.P. Bryant & F. S. Chapin III. 1985. Resource availability and plant
antiherbivory defense. Science, 230:895-899
Cornelissen, J.H., C. Lavorel, E. Garnier, S. Díaz, N. Buchmann, D.E. Gurvich, P.B.
Reich, H. ter Steege, H.D. Morgan, M.G.A. van der Heijden, J.G. Pausas, & H.
Poorter. 2003. Handbook of protocols for standardised and easy measurement of
plant functional traits worldwide. Australian Journal of Botany, 51:335-380.
Goulding, M., M.L. Carvalho & E.G. Ferreira. 1988. Rio Negro, Rich Life in Poor
Water. Netherlands: SPB Academic Publishing.
Hubbell, S. P., R.B. Foster, S.T.O’Brien, K.E. Harms, R. Condit, B. Wechsler, S.J.
Wright & S. Loo de Lao. 1999. Light-gap disturbances, recruitment limitation,
and tree diversity in a neotropical forest. Science, 286:554.

113
Junk, W.J., P.B. Bayley & R.E. Sparks. 1989. The flood pulse concept in river-
floodplain systems. Special Publications of the Canadian Journal of Fisheries
and Aquatic Sciences, 106:110-127.
Lavorel, S., S. McIntyre, J. Landsberg & D. Forbes. 1997. Plant functional
classifications: from general groups to specific groups based on response to
disturbance. Trends in Ecology and Evolution, 12:474-478.
Lavorel, S. & E. Garnier. 2002. Predicting changes in community composition and
ecosystem functioning from plant traits: revisiting the Holy Grail. Functional
Ecology, 16:545-556.
Lawton, J.H. 1983. Plant architecture and the diversity of phytophagous insects. Annual
Review of Entomology, 28:23-39.
Lebrija-Trejos, E., E.A. Pérez-García, J.A. Meave, F. Bongers & L. Poorter. 2010.
Functional traits and environmental filtering drive community assembly in a
species-rich tropical system. Ecology, 91:386-398.
Metamoros, R. & M. Moreno. 2001. La plasticidad de las plantas. Elementos, 41:39-43.
Parolin, P. 2009. Submerged in darkness: adaptations to prolonged submergence by
woody species of the Amazonian floodplains. Annals of Botany, 103:359-376.
Schongart, J., M.T. Piedade, S. Ludwighausen, V. Honans & M. Worbes. 2002.
Phenology and stem growth periodicity of tree species in Amazonian floodplain
forests. Journal of Tropical Ecology, 18:581–597.
Townsend, C.R., M. Begon & J.L. Harper. 2006. Fundamentos em ecologia. Porto
Alegre: Artmed.
Violle, C., M.L. Navas, D. Vile, E. Kazakou, C. Fortunel, I. Hummel & E. Garnier.
2007. Let the concept of trait be functional! Oikos, 116:882-892.

114
Formigas que planam, nadam? Relação entre queda direcional e capacidade de
natação em três espécies de formigas de uma área alagável na Amazônia Central
Lorena Pinto, Caroline Marques Dracxler, Randolpho Dias Terceiro & Cibele Paiva
Introdução
O pulso de inundação favorece a ocorrência e a distribuição de plantas e animais
adaptados a ambientes sazonalmente alagados (Junk et al.1989). Em animais, algumas
estratégias morfológicas (desenvolvimento de órgãos respiratórios), fisiológicas (maior
habilidade de flutuação) e comportamentais (queda direcional) permitem o sucesso de
alguns grupos nesses ambientes (Adis & Junk 2002, Yanoviak et al. 2005 ). Entre os
invertebrados de solo, as estratégias comportamentais podem envolver uma migração
vertical, para substratos mais elevados, ou migração horizontal, que é o deslocamento
para porções de terra firme não inundáveis. Tais movimentos sazonais favorecem o
restabelecimento das populações após o período de inundação (Adis & Junk 2002).
Espécies arborícolas apresentam estratégias que impedem ou diminuem o
contato com as áreas alagadas. Algumas formigas que nidificam e forrageiam em
árvores conseguem realizar queda direcional. Esse comportamento consiste em um
redirecionamento corporal para a árvore durante uma queda, possibilitando o retorno ao
ninho e reduzindo a chance de predação na superfície da água (Yanoviak et al. 2011).
Essa estratégia aparenta ter diferentes origens evolutivas, mas fortes evidências sugerem
que a origem da queda direcional está principalmente relacionada com florestas
alagadas, como uma estratégia para diminuir a pressão de predação nesse ambiente e
sobreviver à inundação (Yanoviak et al. 2005).
Por outro lado, as espécies associadas ao solo, à serapilheira e a moitas de
vegetação estão entre as espécies mais afetadas pela inundação (Philpott et al. 2010).

115
Assim, formigas de solo que ocorrem em áreas inundadas desenvolveram formas de
lidar com as alterações sazonais desse ambiente. Alguns mecanismos de sobrevivência
incluem evacuar os ninhos para áreas mais altas ou para as árvores (Adis 2001,
Ballinger et al. 2007, Lude et al. 1999), ou formar aglomerados compostos pela rainha,
soldados e prole para serem carregados pela corrente d’água até uma área seca. Ainda
que a capacidade de nado não seja um comportamento comum em formigas, algumas
espécies desenvolveram formas de se deslocar sobre a água no período de inundação.
Isso favorece a continuação do forrageamento mesmo quando seu habitat é inundado
(Adis 1982).
Como formigas arborícolas têm menor contato com a água no período de
inundação quando comparado as formigas de solo, nós avaliamos a capacidade de nado
em formigas arborícolas que direcionam a queda. Nós esperamos que espécies
arborícolas com queda direcional comprovada, tenham uma capacidade de natação
menos desenvolvida. Dessa forma, essas espécies demorariam mais tempo para sair da
água quando comparadas as formigas de solo.
Métodos
Área de estudo
Realizamos esse estudo no Parque Nacional de Anavilhanas (PNA), no município de
Novo Airão, Amazônia Central. A área, banhada pelo Rio Negro, é caracterizada por
diversas ilhas de florestas de igapó periodicamente inundadas.
Espécies estudadas
Nós selecionamos três espécies de formiga como modelo de estudo, de acordo com a
disponibilidade e com o hábito de nidificação. Cephalotes atratus (Myrmicinae) é uma

116
formiga de dossel, que constrói ninhos no topo das árvores e forrageia frequentemente
nos galhos em busca de pólen, néctar e exsudato de homópteros (Yanoviak et al. 2005).
Camponotus sp. (Formicinae) é uma espécie que forrageia na vegetação, com ninhos
polidômicos (ninhos fragmentados em diversos domos), próximos ao nível do solo. Já
foi descrito que a espécie C. atratus e que algumas espécies do gênero Camponotus
apresentam capacidade de direcionar a queda e retornar ao tronco após cair de um ramo
no dossel (Yanoviak et al. 2005). Formigas do gênero Atta (Myrmicinae) constroem
grandes ninhos no solo e forrageiam nas árvores em busca de folhas. Essas formigas
dificilmente mudam seus ninhos de lugar, mantendo-se fixas por grande parte do tempo.
Além disso, forrageiam na vegetação e não apresentam capacidade de queda direcional.
Coleta das formigas e realização do experimento
Realizamos busca ativa por formigas em uma ilha do PNA. Como os ninhos de C.
atratus são construídos no dossel, coletamos indivíduos que estavam forrageando na
vegetação. Indivíduos de Atta sp. e Camponotus sp. foram coletados diretamente dos
ninhos, encontrados em troncos caídos ou no solo.
Após a coleta das formigas em campo, conduzimos um experimento em
laboratório para testar a capacidade de nado das diferentes espécies. Para isso,
estabelecemos um círculo de 10,6 cm de diâmetro no centro de uma bandeja de 15 x 10
cm² com água. O experimento consistiu em quantificar o tempo que cada formiga levou
para se deslocar para fora do círculo. Realizamos o experimento com 20 formigas de
cada espécie.

117
Análises estatísticas
Nós comparamos o tempo de deslocamento para saída do circulo entre as três espécies
usando uma Análise de Variância (ANOVA). Em seguida, fizemos comparações
planejadas para buscar diferenças no tempo de deslocamento para fora do círculo entre
os pares de espécies.
Resultados
O tempo médio que as formigas levaram para sair do círculo de 10,6 cm de diâmetro foi
de 44,1 ± 25,87 s em C. atratus, 31,2 ± 28,9 s em Atta sp. e 3,6 ± 7,6 s em Camponotus
sp. O tempo de deslocamento para fora do círculo do experimento diferiu entre as três
espécies (F(2,57) =16,58; p<0,01; Figura 1). O tempo foi similar entre C. atratus e Atta
sp. (t=1,8; gl=1; p=0,08) e foi cerca de oito vezes maior em Atta sp. quando comparado
com Camponotus sp. (t=3,84; gl=1; p<0,01).
Figura 1. Tempo de deslocamento (em segundos) das formigas Cephalotes atratus, Atta
sp. e Camponotus sp. para fora do círculo de 10,6 cm de diâmetro utilizado no
experimento que avaliou a capacidade de natação das formigas. As barras representam
médias e as linhas verticais o desvio padrão.

118
Discussão
As espécies que apresentam queda direcional, C. atratus e Camponotus sp., tiveram
respostas opostas quanto a capacidade de natação, quando comparadas com a espécie
Atta sp. Dessa forma, a queda direcional não determinou a capacidade de nado dessas
duas espécies. Outros fatores como hábito de nidificação, características de forrageio e
estratégias de sobrevivência à inundação sejam importantes na diferenciação da
capacidade de nado entre formigas.
As espécie C. atratus e Atta sp. não apresentaram capacidade de nado, ou foi
pouco desenvolvida. O comportamento de queda direcional e a construção de ninhos e
forrageio no dossel diminuem as chances de contato com o solo e a água durante a
cheia. Isso pode justificar a baixa capacidade de nado dessa espécie. Já as formigas Atta
sp. constroem ninhos únicos no solo, estando sujeitas a inundação. Nesse período, elas
ficam em contato direto com a água e a única maneira da colônia sobreviver é
abandonar o ninho e colonizar novas áreas secas (Lude et al. 1999). Dentre as
estratégias para escapar do alagamento pode ocorrer a formação de uma aglomerado de
indivíduos, com a rainha e a prole no centro, que flutuam até uma área longe da
inundação, ou um possível investimento em indivíduos alados para iniciar uma nova
colônia em outro local. Essas estratégias não incluem o nado, de modo que os
indivíduos que não migram, podem morrer afogados ou predados por peixes na lâmina
d’água.
Embora Camponotus sp. realize queda direcional sua capacidade de nado foi
superior a Atta sp., contrariando nossa hipótese. Os indivíduos de Camponotus sp.
acessam o dossel para forragear e seus ninhos são polidômicos, com domos de uma
mesma colônia distribuído em diferentes regiões de uma mesma árvore ou distribuído
em várias árvores próximas ao solo. A princípio, a construção desse tipo de ninho é uma

119
estratégia de forrageamento, pois aumenta a área ocupada pela colônia e o território para
exploração dos recursos (Peeters & Molet 2010). É possível, que esse tipo de estratégia
tenha favorecido a capacidade de nado nessa espécie, pois possibilita o acesso dos
indivíduos aos domos da colônia que estão acima do nível da água em uma mesma
árvore ou nas árvores próximas.
Assim, a diferença na resposta de natação entre as três espécies pode ter ocorrido
devido a um conjunto de fatores e não apenas pela capacidade de queda direcional das
duas espécies arborícolas. As diferentes estratégias de sobrevivência, como
deslocamento na água para acessar o ninho ou deslocar indivíduos para fora do
alagamento, além de hábitos distintos de forrageamento e nidificação, podem ser
determinantes no comportamento de nado de cada espécie.
Agradecimentos
Agradecemos ao Jack Sparrow (Fabrício Baccaro) por todo o aprendizado e pela viagem
ao mundo das formigas e ao nosso barqueiro Carlos pela ajuda nas coletas.
Referências
Adis J. 2001. Amazonian arthropods (terrestrial). Pages:.249–260. In: Encyclopedia of
Biodiversity (S.A. Levin, ed), Vol. 1, San Diego: Academic Press.
Adis, J. & W.J. Junk. 2002.Terrestrial invertebrates inhabiting lowland river floodplains
of Central Amazonia and Central Europe: a review. Freshwater Biology, 47:711–
731.
Ballinger, A., P.S. Lake & R. Mac Nally. 2007. Do terrestrial invertebrates experience
floodplains as landscape mosaics? Immediate and longer-term effects of flooding
on ant assemblages in a floodplain forest. Oecologia, 152:227–38.

120
Bush, G.L. 1994. Sympatric speciation in animals: new wine in old bottles. Trends in
Ecology & Evolution, 9:285–288.
Holt, R.D., R. Gomulkiewicz & M. Barfield. 2003. The phenomenology of niche
evolution via quantitative traits in a ‘black-hole’ sink. Proceedings of the Royal
Society London B, 270:215–224.
Junk, W., P.B. Bayley & R.E. Sparks. 1989. The flood pulse concept in river-
floodplain-systems. Canadian Special Publications for Fisheries and Aquatic
Sciences, 106:110–127.
Junk, W.J. 1997. General aspects of floodplain ecology with special reference to
Amazonian floodplains. pages 3-20. In: The central Amazon floodplain – ecology
of a pulsing systems (W.J. Junk, ed). Springer-Verlag.
Krivan, V. 2003. Ideal free distributions when resources undergo population dynamics.
Theoretical Population Biology, 64:25–38.
Lude, A., M. Reich, & H. Plachter. 1999. Life strategies of ants in unpredictable
floodplain habitats of Alpine rivers (Hymenoptera: Formicidae). Entomologia
Generalis, 24:75–91.
Peeters, C. & M. Molet. 2010. Colonial reproduction and life histories, pp.159-162. In:
Ant ecology (L. Lori, C.L. Parr & K.L. Abbott, eds). Oxford: Oxford University
Press.
Philpott, S.M., I.Perfecto, I. Armbrecht & C.L. Parr. 2010. Ant Diversity and Function
in Disturbed and Changing Habitats, pp.137-142. In: Ant ecology (L. Lori, C.L.
Parr & K.L. Abbott, eds). Oxford: Oxford University Press.
Yanoviak S.P., R. Dudley & M. Kaspari. 2005. Directed aerial descent in canopy ants.
Nature, 433:624–626.

121
Yanoviak, S.P, Y. Munk & R. Dudley. 2011. Evolution and ecology of directed aerial
descent in arboreal ants. Integrative and Comparative Biology, 51:944–956.

122
O tempo de submersão de plântulas de Dalbergia sp. gera um investimento
diferencial entre os sistemas aéreo e radicular?
Artur Madeira, Bianca Silva, Daniel Passos, Ludmila Rattis
Introdução
O pulso de inundação é uma elevação periódica no nível da água em regiões alagáveis
que separa o ambiente em dois períodos bastante distintos: o período da cheia e o
período da seca (Junk et al. 2010). Essa divisão resulta em mudanças que geram uma
pressão seletiva sobre os organismos daquele ambiente, por exemplo, por alterar a
disponibilidade de oxigênio no solo durante o período de cheia. Solos inundados se
tornam anóxicos ou hipóxicos dentro de horas, como resultado do consumo do oxigênio
por raízes e microorganismos, e pela baixa difusão de oxigênio na água (Parolin et al.
2004). Essas condições contrastantes favoreceram a evolução de adaptações e
estratégias que permitiram o estabelecimento de espécies nesses ambientes (Junk et al.
2010).
Uma adaptação comum de espécies de plantas em ambientes alagáveis é a
sincronia da produção e liberação de frutos na época em que o período de cheia está no
auge, favorecendo estratégias de dispersão como a hidrocoria (Wittmann et al. 2007).
As sementes são então liberadas ao mesmo tempo na água, sendo depositadas no solo à
medida que o nível da água começa a baixar. Assim, uma semente depositada em um
local mais baixo permanece mais tempo na água e isto pode afetar o investimento
diferencial em partes da planta.
Ao germinar, o embrião começa a se desenvolver utilizando reservas de energia
e nutrientes armazenados no cotilédone até que ele produza folhas e comece a fazer
fotossíntese (Malavasi & Malavasi 2001, Parolin 2002). Em resposta à pressão causada

123
por uma possível submersão no próximo período de cheia, as plântulas desenvolveram
duas estratégias principais que não são mutuamente exclusivas, evitar ou tolerar a
submersão total (Parolin 2002).
Durante a cheia, locais mais baixos serão os primeiros a serem cobertos e os
últimos a serem descobertos pela água, estando sujeitos a um maior tempo de
submersão, enquanto locais mais altos ficarão submersos por menos tempo. Assim, o
maior investimento no sistema aéreo permitirá que a plântula fique o mais alta possível
antes que o próximo período de cheia comece, mantendo suas folhas acima do nível da
água por mais tempo (Parolin 2002). Por outro lado, o maior investimento no sistema
radicular permitirá que a plântula se fixe mais firmemente, evitando sua remoção
durante o período de submersão, e que ela absorva água e nutrientes do solo com maior
eficiência.
Nas margens do Rio Negro sujeitas à inundaçào (igapó), plântulas de Dalbergia
sp. podem ser encontradas tanto em locais mais altos como em locais mais baixos. As
plântulas das regiões mais baixas ficarão mais tempo submersas e por isso seria
vantajoso para elas investir mais no sistema aéreo. Nestas regiões baixas, plântulas mais
altas demorariam mais para serem totalmente submersas e, quando o nível da água
começasse a baixar novamente, elas seriam descobertas pela água mais cedo do que
plântulas mais baixas. Por outro lado, plântulas de regiões mais altas ficariam
submersas por menos tempo pois já se encontram em um local menos sujeito à
inundação e, por isso, deveriam investir mais no sistema radicular, melhorando sua
fixação e absorção de água e nutrientes do solo.
Nosso objetivo foi avaliar se existe investimento diferencial em crescimento
aéreo e radicular dependente do tempo de inundação ao qual a plântula será submetida.

124
Esperamos que plântulas localizadas em locais mais altos investirão mais no sistema
radicular e plântulas localizadas em locais mais baixos investirão mais no sistema aéreo.
Métodos
Realizamos o trabalho no arquipélago de Anavilhanas, localizado no Rio Negro,
Amazonas, Brasil. Selecionamos dois locais, um mais baixo e sujeito a um maior tempo
de inundação (cota baixa), e outro mais alto, que passa menos tempo sujeito à inundação
(cota alta). A diferença de altitude entre as duas cotas foi aproximadamente 2 m.
Em cada cota coletamos 33 plântulas de Dalbergia sp. (Leguminosae). Medimos
o comprimento do epicótilo e o comprimento da radícula de cada indivíduo. Para medir
o comprimento do epicótilo, consideramos a distância da parte aérea desde o ponto de
saída do cotilédone até a primeira folha formada. Consideramos o tamanho da radícula
como o comprimento da raiz principal. Então, calculamos a razão entre o comprimento
do epicótilo e o comprimento da radícula (razão epicótilo-radícula). Realizamos um
teste t unicaudal para avaliar se a razão epicótilo-radícula (variável resposta) varia entre
as cotas de inundação alta e baixa (variável preditora). Especificamente, testamos a
previsão de que a razão epicótilo-radícula é maior na cota baixa.
Resultados
Nossa análise demonstrou que a razão epicótilo-radícula das cota de inundação mais alta
não foi menor do que a razão da cota baixa (t = 1,30; gl = 64; p = 0,10; Figura 1).

125
Figura 1. Razão epicótilo-radícula entre cotas de inundação (baixa e alta) em plântulas
de Dalbergia sp. de uma floresta de igapó no arquipélogo de Anavilhanas, Manaus,
Brasil. Pontos e barras representam média ± desvio padrão.
Discussão
Não houve efeito do tempo de submersão das plântulas sobre o investimento nos
sistemas aéreo e radicular em Dalbegia sp.. Isso indica que o investimento das reservas
do cotilédone em fixação e crescimento se dá de forma semelhante, independente do
local de estabelecimento.
Plântulas estão sujeitas à submersão mesmo nos locais mais altos. Assim, é
provável que as plântulas de Dalbergia sp., independente da cota, invistam mais em
estratégias de tolerância à submersão. Por outro lado, as plântulas também podem
investir em crescimento rápido, independente do local em que se encontram, de forma a
reduzir ao máximo o tempo de submersão de suas folhas no próximo período de
inundação (Parolin 2002). Portanto, mesmo que as plântulas invistam mais em
estratégias de crescimento, elas o fazem de maneira similar em ambas as cotas.
Baixa Alta
Cota de Inundação
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
Raz
ão E
pic
óti
lo/R
adíc
ula

126
Se as plântulas não respondem ao tempo de inundação de formas diferentes em
cotas altas e baixas, os indivíduos de cotas baixas serão afetados pela inundação antes e
permanecerão assim por mais tempo do que indivíduos de cotas altas. Isso acarretará em
um menor tempo para crescimento nas cotas baixas. Assim, a estratégia de tolerância
deve ser mais importante para as plântulas, enquanto que a estratégia de escape do
efeito prolongado de inundação pode ser mais importante no estágio adulto, quando
ocorre a produção de sementes. Considerando que a cota em que a semente será
depositada depende do nível da água, é mais vantajoso liberar os frutos no auge do
período de cheia aumentando as chances da semente ser depositada em uma cota mais
alta.
Concluímos que não houve investimento diferencial nos sistemas aéreo e
radicular em Dalbergia sp. entre cotas com uma diferença de até 2 m de altura. Porém
esse resultado não invalida a possibilidade de que o efeito exista entre cotas com
maiores diferenças de altura.
Referências
Junk, W.J., M.T.F. Piedade, F. Wittmann, J. Schongart & P. Parolin (eds). 2010.
Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable
Management. New York: Springer.
Malavasi, U.C. & M.M. Malavasi. 2001. Influência de tamanho e do peso da semente na
germinação e no estabelecimento de espécies de diferentes estágios da sucessão
vegetal. Floresta e Ambiente, 8:211-215.
Parolin, P. 2002. Submergence tolerance vs. escape from submergence: two strategies
of seedling establishment in Amazonian flood plains. Environmental and
Experimental Botany, 48:177-186.

127
Parolin, P., O. De Simone, K. Haase, D. Waldhoff, S. Rottenberger, U. Kuhn, J.
Kesselmeier, B. Kleiss, W. Schmidt, M.T.F. Piedade & W.J. Junk. 2004. Central
Amazonian floodplain forests: tree adaptations in a pulsing system. The Botanical
Review, 70:357-380.
Wittmann, A.O., M.T.F. Piedade, P. Parolin & F. Wittmann. 2007. Germination in four
low-várzea tree species of Central Amazonia. Aquatic Botany, 86:197-203.

128
Anavilhanas – projetos livres
A borboleta Glutophrissa drusilla (Pieridae) prefere cruzar rios mais largos entre
as manchas florestais
Mariana Velasque, Ana Carolina Vieira Pires, Tatiane G. C. Menezes, Renata Pimentel
Rocha e Ludmila Rattis
A migração é um padrão de movimentação direcional de grande número de indivíduos
de uma mesma espécie. Durante esse trajeto, o deslocamento por áreas com poucos
refúgios pode ser prejudicial para os organismos, devido ao aumento na exposição a
predadores. Assim, organismos que têm o comportamento de selecionar rotas de
migração que minimizem o risco de predação deveriam ser favorecidos. No
Arquipélago de Anavilhanas, observamos indivíduos da borboleta Glutophrissa drusilla
atravessando braços do rio Negro de diferentes larguras entre fragmentos de mata, em
voo direcionado ao sul, o que indica um comportamento migratório. Uma vez que
braços mais largos do rio aumentam a exposição da borboleta a possíveis predadores,
propomos a hipótese de que elas preferem cruzar braços mais estreitos. Para isso,
realizamos observações a cada 30 min , entre 9 h e 12 h, em um braço mais largo do rio
(533 m) e outro mais estreito (80 m). Registramos em média 1,5 vezes mais borboletas
cruzando o braço largo do rio (F(1,5)=35,7; p=0,002). Observamos quatro ataques de
aves apenas no braço mais longo, todos sem captura. A maior frequência de borboletas
sobrevoando o leito largo indica que há um direcionamento da rota de forma a
atravessar áreas abertas mais extensas entre fragmentos de mata. Dessa forma,
sugerimos que essas áreas abertas poderiam proporcionar maior visibilidade,
possibilitando o escape de ataques de predadores. Porém ,ainda que rios largos

129
permitam maior visibilidade, a utilização dos fragmentos de floresta entre os braços de
rio poderia servir como pontos de descanso.

130
O tamanho das pernas de formigas em áreas alagáveis influencia sua capacidade
de deslocamento na água?
Caroline Marques Dracxler, Lorena Costa Pinto, Vidal Carrascosa, Daniela Pinto
Coelho e Sarah Freitas Magalhães
Em ambientes alagáveis, formigas de solo entram periodicamente em contato com a
água e precisam se deslocar para reestabelecer seus ninhos. No entanto, a capacidade de
deslocamento na água varia entre diferentes espécies de formigas. De forma geral,
quanto maior o tamanho do tórax das formigas, proporcionalmente maior é o tamanho
de suas pernas, o que pode facilitar o deslocamento na água durante o período de
inundação. Assim, esperamos que formigas com menor relação entre perna e tórax
apresentarão menor velocidade de deslocamento sobre a lâmina d’água. Testamos a
capacidade de deslocamento de oito espécies (seis gêneros) de formigas de solo
coletadas em uma área de igapó no Parque Nacional de Anavilhanas.
Experimentalmente, estabelecemos um círculo de 22,5 cm de diâmetro em uma bacia
com água e registramos a velocidade de deslocamento para fora do círculo de três
indivíduos de cada espécie. Calculamos a proporção perna/tórax de cada indivíduo e
obtivemos as médias para cada espécie. A velocidade mínima e máxima registrada foi
0,22 cm/s e 8,25 cm/s, respectivamente, e a proporção perna/tórax variou de 1,73 a 3,24.
Não houve relação entre a proporção perna/tórax das formigas e a velocidade de
deslocamento (F(1,6)=0,26; p=0,63). Apesar dessas espécies viverem em áreas
inundáveis, o tamanho das pernas parece não fornece vantagem no deslocamento sobre
a superfície da água. No entanto, é possível que certas características morfológicas,
como cerdas maiores ou presença de cera na superfície do tegumento favoreçam maior
flutuação e maior eficiência de deslocamento na água.

131
Pernas para que te quero? Função das pernas na natação da formiga Camponotus
sp. (Formicidae)
Randolpho Dias-Terceiro, Artur Madeira, Daniel Passos, Cibele Paiva & Leonardo
Oliveira
Formigas de ambientes periodicamente alagáveis ocasionalmente podem entrar em
contato com a água e, portanto, indivíduos capazes de nadar podem aumentar sua
sobrevivência em relação àqueles destituídos da capacidade natatória. Operárias de
formigas do gênero Camponotus são capazes de nadar movimentando as pernas
anteriores e mantendo as posteriores estáticas e paralelas à superfície d’água. Nossa
hipótese é de que as pernas anteriores são responsáveis pela propulsão enquanto as
posteriores exercem função de direcionamento do nado. Portanto, com a remoção das
pernas anteriores o nado será mais lento e linear, enquanto que com a remoção das
pernas posteriores o nado será rápido, porém sem direção. Coletamos indivíduos de
Camponotus sp. e aplicamos três tratamentos: remoção das antenas (controle), das
pernas posteriores e das anteriores. Medimos a velocidade (cm/s) e a linearidade do
nado de 15 formigas por tratamento. Indivíduos do grupo controle apresentaram maior
velocidade (F(2,42) = 21,15; p < 0,01) e linearidade (F(2,42) = 3,51; p = 0,04) no nado
quando comparado aos indivíduos dos demais tratamentos. Entretanto, os tratamentos
de remoção das pernas anteriores e posteriores não diferiram entre si quanto à
velocidade ou linearidade do nado. Tanto a remoção das pernas anteriores quanto
posteriores implicou em menor eficiência natatória, sugerindo que as funções de
propulsão e direcionamento do nado não são realizadas independentemente por um par
de pernas em particular.

132
Variação da capacidade de armazenamento de água na planta Psychotria sp.
mediada por filtro ambiental
Ana Claudia Rorato, Joselândio Santos, Bianca Ferreira, José Hidasi Neto & Bruno
Travassos
Filtros ambientais podem gerar mortalidade diferencial em indivíduos com
características de tolerância distintas. Em um determinado conjunto de condições,
sobrevivem indivíduos que são funcionalmente semelhantes (convergência). Bordas
florestais apresentam maior incidência luminosa, favorecendo maior dessecação, em
comparação ao interior florestal. Investigamos o efeito da borda no armazenamento
hídrico em Psychotria sp. (Rubiaceae). Nossa hipótese é que a borda leva a uma
convergência na capacidade de armazenamento hídrico. Coletamos 40 indivíduos, 20 na
borda e 20 no interior florestal. Para cada indivíduo, calculamos a diferença entre a
massa foliar túrgida e a massa seca (DMF) para quatro folhas. Previmos que, na borda,
Psychotria sp. apresentaria maior média, porém menor variabilidade em DMF.
Comparamos as médias de DMF entre as áreas e os desvios padrões observados de
DMF com o desvio esperado em um modelo nulo. As médias de DMF foram
semelhantes (F=38,31; p=0,48; gl=38). Entretanto, o desvio de DMF foi menor que o
esperado na borda (p < 0,01), mas não no interior (p = 0,89). Assim, as condições na
borda atuam como filtros na convergência de armazenamento hídrico em Psychotria sp.,
porém esse armazenamento não foi maior nesse ambiente. Isso indica que indivíduos
com potenciais de armazenamento muito baixos ou muito elevados são eliminados na
borda. Indivíduos com baixo armazenamento hídrico seriam desfavorecidos devido à
alta taxa de dessecação. Por outro lado, potenciais de armazenamento muito elevados

133
podem estar associados a folhas pouco esclerificadas e mais susceptíveis à herbivoria,
que seriam desfavorecidas na borda.

134
Km 41 – projetos orientados
A aranha Faiditus subflavus (Theridiidae) selecionam indivíduos de Maieta
guianensis (Melastomataceae) com menor dano foliar?
Leonardo Oliveira, Renata Rocha, José Hidasi e Ana Carolina Vieira Pires
Introdução
A seleção de habitat pode ser definida como a escolha que os indivíduos fazem por
locais que favorecem sua sobrevivência e reprodução (Orians 1991). Essa relação pode
ocorrer de acordo com diversos fatores, como menor risco de predação e parasitismo.
Em locais com elevada pressão de predação, os indivíduos podem selecionar habitats
específicos (Ricklefs & Schluter 1993) que apresentam componentes capazes de reduzir
o risco de predação para o adulto e/ou a prole. Assim, o processo de escolha do habitat é
um fator que pode determinar o sucesso reprodutivo do indivíduo (Sih 1980).
Diversas aranhas maximizam o sucesso reprodutivo por meio da escolha de
habitats de melhor qualidade, que podem ser compreendidos como sítios mais seguros,
com maior disponibilidade de recursos ou que apresentam estruturas para a fixação da
teia (Allan 1995). O processo de escolha do habitat pode envolver sentidos visuais,
táteis ou olfativos. Por exemplo, a aranha Misumena vatia (Thomisidae) utiliza pistas
visuais e táteis para selecionar melhores sítios de forrageio, o que aumenta o sucesso
reprodutivo dos indivíduos dessa espécie (Fritz & Morse 1985, Greco & Kevan 1994).
Além disso, alguns grupos de aranhas como as que vivem em plantas, podem usar
indicativos indiretos de qualidade do habitat, como o dano físico provocado por
herbívoros. Isso pode indicar a existência de intensa atividade de insetos na planta, que

135
podem servir tanto para alimentação como para a proteção das aranhas (Romero &
Vasconselos-Neto 2007).
Essas pistas indiretas podem ser particularmente importantes para aranhas que
habitam plantas mirmecófitas. Nesse sistema, as plantas possuem estruturas que
abrigam formigas, as quais patrulham a planta em busca de herbívoros (Vasconcelos
1991). Dessa forma, danos físicos provocados por herbivoria podem indicar baixa
capacidade de proteção da planta pelas formigas.
No ambiente amazônico, a aranha Faiditus subflavus (Theridiidae) apresenta
uma relação íntima com a mirmecófita Maieta guianenses (Melastomataceae). As
formigas da espécie Pheidole minutula (Myrmicinae), que residem nessa planta
oferecem proteção para a aranha adulta e para a prole por meio do patrulhamento da
superfície, no qual ela afasta possíveis predadores (Santos 2007) e parasitas das aranhas.
Indivíduos de M. guianensis são encontrados exibindo diferentes níveis de dano foliar
por herbivoria. Assumindo que folhas com menos herbivoria podem indicar para as
aranhas que as formigas oferecem boa defesa, avaliamos se as aranhas se estabelecem
preferencialmente em plantas mais protegidas por formigas e consequentemente com
menos herbivoria. Esperamos que a presença de aranhas seja maior nas plantas com
menor número de folhas consumidas.
Métodos
Área de estudo
Realizamos a coleta de dados em uma área de floresta contínua na Reserva do Km 41
próxima ao município Rio Preto da Eva, na Amazônia Central, Brasil. O local é
classificado como uma ARIE (Área de Relevante Interesse Ecológico), gerido pelo
ICMbio e pelo Projeto Dinâmica Biológica de Fragmentos Florestais (PDBFF).

136
Coleta de dados
Selecionamos 20 indivíduos de M. guianensis próximo a um igarapé, sendo 10
indivíduos com aranhas e 10 indivíduos sem aranhas. Restringimos nossa coleta para
amostrar apenas plantas que continham entre 10 e 20 domáceas, pois abaixo de 10
domáceas as plantas são muito pequenas e acima de 20 existe alta taxa de ocorrência de
F. subflavus (Santos 2007). Para obter o dano foliar, contamos em cada planta o número
total de folhas e a quantidade de folhas com dano.
Análise dos dados
Testamos a previsão de que a presença de aranhas é maior nas plantas com menor
número de folhas consumidas. Para isso, comparamos a diferença de ajuste entre um
modelo saturado e reduzido. O modelo saturado conteve a ocorrência de aranhas como
variável dependente e as variáveis número total de folhas e número de folhas
consumidas como variáveis independentes. O modelo reduzido foi obtido por meio da
remoção da variável número de folhas consumidas do modelo saturado.
Resultados
Encontramos uma grande variação no número de folhas danificadas. As plantas que
continham aranhas apresentavam entre 5 e 19 folhas consumidas, com o número total de
folhas variando de 23 a 49. As plantas sem aranhas tiveram de 3 a 17 folhas
consumidas, com o número total de folhas variando entre 16 e 33. O número de folhas
consumidas não explicou a presença e ausência de aranhas na planta (χ²= 0,57; gl= 1; p=
0,45; Figura 1).

137
Figura 1. Presença e ausência de aranhas F. subflavus em função da quantidade de
folhas consumidas da planta M. guianensis em uma floresta contínua, Amazônia central,
Manaus, Brasil. O “0” no eixo vertical indica ausência e “1” indica presença.
Discussão
Ao contrário do esperado, as fêmeas de F. subflavus não selecionaram plantas com
menor herbivoria para construir sua teia, indicando que a herbivoria não é um indicativo
de segurança. A herbivoria modifica a arquitetura da planta no momento em que o
herbívoro remove a biomassa vegetal das folhas. Assim é possível inferir que a
herbivoria reduza o substrato utilizado pela aranha para ancoragem da teia, o que
implica em ausência da aranha na planta. Entretanto, a aranha F. subflavus utiliza a base
das folhas para a construção da teia (Kasper 2006). No nosso estudo, a base das folhas
permaneceu intacta, pois os danos foliares se concentraram na parte anterior da folha,
sugerindo que, nesse caso, a herbivoria não reduz o substrato necessário para a aranha
se estabelecer. Assim, sugerimos que o padrão de herbivoria observado não é suficiente
para impedir o estabelecimento das teias de F. subflavus.

138
O tamanho da planta por outro lado é apontado como uma das principais
características que influenciam positivamente as comunidades de artrópodes (Kuppers
1989). No caso específico do estudo, sabemos que a altura da planta está relacionada
com a quantidade de formigas (Santos 2007). Como as formigas protegem a aranha,
podemos sugerir que o tamanho da planta pode indicar para a aranha a quantidade de
formigas nas plantas, o que indica maior proteção. Além disso, podemos esperar que
quanto maior a planta maior a possibilidade de encontrar sítios para ancoragem da teia.
De forma geral, podemos concluir que a herbivoria não é indicativo de qualidade de
habitat para F. subflavus e que outros fatores, como o tamanho da planta, pode
influenciar na seleção do habitat.
Agradecimentos
Agradecemos a Thiago Kloss e Paulo Bobrowiek pelo auxílio em campo e orientações
na redação do manuscrito.
Referências
Allan, J.D. 1995. Stream ecology. Structure and function of running waters. London:
Chapman & Hall.
Fritz, R.S. & D.H. Morse. 1985. Reproductive success and foraging of the crab spider
Misumena vatia. Oecologia, 65:194-200.
Greco, C.F. & P.G. Kevan. 1994. Contrasting patch choosing by anthophilous ambush
predators: vegetation and floral cues for decision by a crab spider (Misumena
vatia) and males and females of an ambush bug (Phymata americana). Canadian
Journal of Zoology, 72:1583-1588.

139
Kasper, D. 2006. Fatores ambientais que influenciam a ocorrência e abundância de
aranhas Faiditus subflavus (Theridiidae) em plantas do gênero Maieta
(Melastomataceae). In: Curso de campo ecologia da floresta amazônica (G.
Machado & J.L. Camargo, eds.). INPA/PDBFF, Manaus.
Kuppers, M. 1989. Ecological significance of above-ground architectural patterns in
woodplants: a question of cost-benefit relationship. Trend in ecology & Evolution,
4: 375-379.
Orians, G.H. 1991. Preface. The American Naturalist, 137:S1-S4.
Ricklefs, R.E. & D. Schluter. 1993. Species diversity in ecological communities.
Chicago: University of Chicago.
Romero, G.Q. & J. Vasconselos-Neto. 2007. Aranhas sobre plantas: dos
comportamentos de forrageamento às associações específicas, pp.67-88. In:
Ecologia e comportamento de aranhas (M.O. Gonzaga, A.J. santos & H.F.
Japyassú, eds.). Rio de janeiro: Interciência.
Sih, A. 1980. Optimal behavior: can foragers balance two conflicting demands?
Science, 210:1041-1043.
Santos, G.R. 2007. Babás ou guarda-costas: a formiga Pheidole minutula (Myrmicinae)
protege os filhotes ou as fêmeas da aranha Faiditus subflavus (Theridiidae)? In:
Curso de campo ecologia da floresta amazônica (G. Machado & J.L. Camargo,
eds.). INPA/PDBFF, Manaus.
Vasconcelos, H.L. 1991. Mutualism between Maieta guianensis Abul. A
myrmecophytic melastome, and one of its ant inhabitants: and protection against
insect herbivores. Oecologia, 87:295-298.

140
Importância relativa do espaço e do ambiente na explicação da diversidade beta:
um teste da Teoria do Nicho x Teoria Neutra em pequena escala
Daniela Coelho, Bruno Travassos, Randolpho Dias-Terceiro e Cibele Paiva
Introdução
A distribuição das espécies no espaço vem sendo discutida desde o século XIX, com os
trabalhos de Humboldt e outros geógrafos. Eles estabeleceram os fundamentos da
biogeografia moderna e basearam seus trabalhos nos diferentes padrões de distribuição
das espécies e comunidades (McIntosh 1991) que hoje é estudado pela ecologia de
comunidades (Gravel et al. 2006). Existem duas correntes teóricas que explicam a
distribuição das espécies de forma distintas, a teoria de nicho e a teoria neutra.
A teoria de nicho assume que os ambientes são heterogêneos, e possuem
recursos e condições distribuídos de maneira diferenciada no espaço. Ainda, esta teoria
assume que as espécies possuem diferentes necessidades e habilidades para a obtenção
desses recursos. Portanto, espécies somente devem ocorrer em locais onde existam
condições adequadas e recursos suficientes para sua sobrevivência. De acordo com essa
teoria, quanto mais diferentes forem dois hábitats em condições ambientais, mais
dissimilares suas comunidades devem ser, ou seja, maior a diversidade-β entre eles
(MacArthur & MacArthur 1961).
A teoria neutra assume que as espécies são ecologicamente equivalentes e
possuem igual probabilidade de ocupar um local no espaço (Hubbell 2001). Nessa
situação, o que explicaria a ocorrência das espécies seriam processos de contingência
histórica, extinções estocásticas e habilidade de dispersão dos organismos (Whitfield
2002). Uma vez que a habilidade de dispersão é limitada, a probabilidade de uma
espécie se dispersar para outro lugar reduz quanto mais distante for esse lugar

141
(MacArthur & Wilson 1967). Dessa forma, uma decorrência lógica dessa teoria é que
quanto mais distantes as comunidades estão no espaço, mais dissimilares elas devem
ser, ou seja, apresentarão maior diversidade-β.
Alguns autores argumentam que essas duas teorias não são precisamente
antagônicas, mas sim complementares (Gravel et al. 2006). É provável que a escala
espacial tenha um papel importante no poder de previsão dessas duas teorias (Levin
1992). Os modelos derivados da teoria neutra parecem ter melhor aceitação em escalas
regionais (e. g. MacArthur & Wilson 1967; Kadmon & Allouche 2007). Entretanto, em
escalas mais restritas, o modelo neutro pode não ser muito eficaz para explicar
mudanças na composição de espécies. Por outro lado, as previsões da teoria de nicho
parecem robustas mesmo em escalas finas.
Avaliamos os efeitos do espaço e do ambiente sobre a diversidade-β.
Especificamente, hipotetizamos que a diversidade-β varia com a diferença entre
ambientes. Prevemos que, mesmo em pequena escala, ambientes distintos possuam
maior dissimilaridade do que ambientes semelhantes. Por outro lado, a diversidade beta
não deve variar em pequenas distâncias, pois a dispersão seria suficiente para
homogeneizar a composição de espécies dentro de escalas pequenas.
Métodos
Realizamos o estudo na reserva florestal do Km 41, na área do Projeto de Dinâmica
Biológica de Fragmentos Florestais, a cerca de 80 km ao norte de Manaus. Escolhemos
uma área de mata fechada cortada por uma estrada, que possui dois ambientes com
características distintas, borda e mata fechada. Estabelecemos quatro blocos onde os
dois ambientes estavam contidos. Estes blocos eram distantes 80 m.

142
Em cada bloco fizemos cinco pontos de amostragem ativa de plantas
pertencentes a três famílias (Arecaceae, Melastomataceae e Polypodiaceae) durante 1,5
min. Para cada bloco consideramos o primeiro ponto amostral realizado na borda como
ponto focal. A partir desse ponto, definimos quatro pontos em distâncias diferentes na
borda e no interior da mata, dois deles a 20 m (pontos 1 e 3) e os outros dois a 60 m
(pontos 2 e 4) do ponto focal (Figura 1). As plantas coletadas em cada ponto foram
levadas ao laboratório e morfotipadas.
Análise dos dados
Para cada bloco calculamos a diversidade-β do ponto focal em relação aos outros quatro
pontos amostrais, obtendo quatro valores de diversidade-β. Fizemos a comparação entre
os pontos através do índice de dissimilaridade de Jaccard. Realizamos uma Análise de
Variância (ANOVA) de dois fatores em blocos para testar o efeito dos tipos de ambiente
Figura 2. Delineamento amostral dentro de cada bloco. O ponto focal está indicado pela
letra ‘F’. Os dois pontos de distância diferentes na borda estão indicados pelos números
1 e 2. Os pontos amostrais na mata estão indicados pelos números 3 e 4.

143
(borda, mata) e da distância (perto [20 m], longe [60 m]) sobre a diversidade-β (variável
resposta).
Resultados
No levantamento foram registradas 29 morfoespécies de plantas, sendo 17
morfoespécies de Melastomataceae, seis de Arecaceae e seis de Polypodiaceae. As
espécies de Melastomataceae tiveram maior ocorrência nos pontos estabelecidos nas
bordas. Por outro lado, as espécies de Arecaceae e Polypodiaceae foram mais
abundantes no interior da mata.
A ANOVA indicou que não houve efeito da interação entre ambiente e distância
sobre a diversidade-β (r²=0,02; F(1,11) = 0,97; p = 0.34; Figura 2). A diversidade-β foi
maior entre locais de ambientes distintos do que entre locais de mesmo ambiente (r2 =
0,68; F(1,11) = 27,63; p < 0,001). No entanto, a dissimilaridade entre locais do mesmo
ambiente em função da distância não diferiu (r2 = 0,02; F(1,11) = 1,14; p = 0,31) (Figura
2).

144
Figura 2. Efeitos do ambiente e espaço sobre a dissimilaridade de espécies de plantas
(famílias Melastomataceae, Arecaceae e Polypodiaceae) entre sítios na borda e dentro
da mata. Os círculos preenchidos indicam a média da dissimilaridade entre os pontos
focais (borda) e os pontos na mata. Os círculos vazios indicam a média da
dissimilaridade entre os pontos focais (borda) e os pontos na borda. As barras verticais
indicam o erro padrão.
Discussão
A variação do tipo de ambiente explicou a variação na diversidade β em pequena escala.
Em contrapartida, a diversidade-β não apresentou variação em relação ao mesmo
ambiente em distâncias distintas. Este resultado indica que variações ambientais foram
mais importantes que a distância espacial na determinação da ocorrência de espécies,
como previsto pela teoria do nicho.
Como ambientes de borda diferem quanto ao microclima em relação ao interior
de mata, variações na disponibilidade de recursos, como luz e nutrientes, e condições,
como umidade e modificação do regime dos ventos, podem afetar o estabelecimento das

145
espécies de plantas (Murcia 1995; Primack & Rodrigues 2001). Estudos prévios
indicam que grupos diferentes de espécies vegetais são adaptados a viverem em
condições distintas, como aquelas de borda e interior (Murcia 1995).
Apesar da variação na diversidade-β não ter sido explicada pela distância,
conforme previsto pela teoria neutra, é possível que ela seja capaz de explicar padrões
de distribuição de espécies em escalas maiores. Em escalas menores, como a utilizada
neste estudo, a capacidade de dispersão das espécies não representa um fator limitante
para a colonização. Ou seja, as espécies são capazes de dispersar seus diásporos por
toda a área de estudo, fazendo com que a composição de espécies seja homogênea.
A intensidade relativa de variação ambiental e espacial deve ser determinante
sobre qual processo (filtro ambiental, dispersão) é mais importante na determinação do
padrão de diversidade beta. Sendo assim, se o gradiente de variação ambiental for maior
que o gradiente espacial, como o observado no presente estudo, processos associados ao
nicho devem prevalecer. Por outro lado, se a variação espacial for maior que a variação
ambiental, os padrões de dispersão podem se tornar mais importantes para determinar a
troca de espécies.
Referências
Gravel, D., C.D. Canham, M. Beaudet & C. Messier. 2006. Reconciling niche and
neutrality: the continuum hypothesis. Ecology Letters, 9:399-409.
Hubbell, S.P. 2001.The Unified Neutral Theory of Biodiversity and Biogeography.
Niche Theory. The American Naturalist, 170:443-454Levin, S.A. 1992. The problem of
pattern and scale in ecology. Ecology, 73:1943-1967.
MacArthur, R. H. & J. W. MacArthur. 1961. On bird species diversity. Ecology,
42:594-598.

146
MacArthur, R.H. & E.O. Wilson. 1967. The Theory of Island Biogeography. Princeton
University Press, Princeton.
McIntosh, R.P. 1991. Concept and terminology if homogeneity and heterogeneity in
ecology. Ecological Studies, 86:24-46.
Murcia, C. 1995. Edge effects in fragmented forest: implications for conservation.
Trends in Ecology and Evolution, 10:58-62.
Primack, R.B. & E. Rodrigues. 2001. Biologia da Conservação. Londrina: E. Rodrigues.
Princeton University Press, Princeton, NJ.
Whitfield, J. 2002. Neutrality versus the niche. Nature, 417:480-481.

147
Interação formiga-planta: a abundância da planta não afeta o número de formigas
mutualistas com que ela interage
Sarah Freitas Magalhães Silva, Ana Claudia Rorato, Artur Alcântara Madeira e Mariana
Velasque
Introdução
As interações interespecíficas são uma das principais forças estruturadoras das
comunidades, influenciando a distribuição e abundância das espécies (Jones et al.
1994). Essas interações podem ocorrer de forma específica ou difusa. Interações
específicas ocorrem quando pares de espécies interagem exclusivamente entre si. Já as
interações difusas ocorrem quando uma espécie pode interagir com várias outras (Morin
2011).
Em sistemas de interações difusas, diferentes espécies têm diferentes números de
interações. Dois mecanismos foram propostos para explicar o número de interações
realizadas por uma espécie. É possível que espécies com determinados atributos sejam
capazes de atrair um grande número de espécies que podem interagir com elas. Por
outro lado, se a preferência por determinados atributos não existir, as espécies mais
abundantes seriam aquelas que apresentam um maior número de associações com outras
espécies (Vázquez & Aizen 2003).
Insetos e plantas interagem de formas espécífica e difusa. Plantas que
apresentam nectários extraflorais (NEF) oferecem substâncias nutritivas às formigas,
que forrageiam sobre suas folhas e em troca atuam na defesa contra herbívoros (Becerra
&Venable 1989). Em alguns casos, a interação formiga-planta é mediada por afídeos
que sugam a seiva da planta, secretam uma solução açucarada que atrai formigas para as
plantas (Engel et al. 2001). Em regiões tropicais, essas interações entre formigas e

148
plantas raramente são específicas, ocorrendo de forma facultativa e/ou oportunista, e
portanto difusa (Rico-Gray & Oliveira 2007).
Observando formigas associadas a NEFs ou afídeos, nos questionamos sobre o
que determina a quantidade de formigas com as quais uma planta dotada de NEFs ou
afídeos pode interagir. Nossa hipótese é de que, quanto mais abundante for uma espécie
de planta, maior é o número de espécies de formigas com as quais ela interage. Assim,
esperamos que quanto maior o número de indivíduos de uma determinada espécie de
planta, maior o número de espécies de formigas observadas em associação com ela.
Métodos
Área de estudo
Conduzimos o estudo na área de borda de uma floresta contínua de terra firme
localizada na Reserva do Km 41 (02° 24’S e 59° 44’O) administrada pelo Projeto
Dinâmica Biológica de Fragmentos Florestais (PDBFF), localizada a cerca de 80 km ao
norte de Manaus, Brasil.
Coleta dos dados
Para descrever as interações entre formigas e plantas, coletamos ao longo de um
transecto de cerca de 100 m, 20 plantas que apresentavam interação com formigas, via
nectários extraflorais ou afídeos. Consideramos que uma formiga estava interagindo
com a planta quando ela estava forrageando próxima ao nectário ou ao afídeo. Por
apresentarem uma interação com alta especialização, plantas mimercófitas e suas
formigas associadas foram desconsideradas. Para reduzir a probabilidade de amostrar
plantas visitadas pela mesma colônia de formigas e inflar os resultados das associações,
coletamos indivíduos a cada 5 m no transecto. Uma vez que ao buscar quantificar as

149
associações, coletar em plantas próximas poderia alterar o resultado das interações, pois
as relações poderiam ser em função da colônia e não pela abundância da espécie da
formiga.
Para quantificar a abundância de plantas, delimitamos 20 parcelas de 25m², que
distavam 5 m entre si. Em cada parcela coletamos todas as plantas que apresentavam
nectários extraflorais ou afídeos. Posteriormente, classificamos as plantas e as formigas
em morfoespécies, identificamos o gênero de algumas formigas quando possível. Se
uma planta ocorreu nas amostragens de interação formiga-planta, mas não estava
presente na amostragem de abundância, consideramos sua abundância como zero e
incluímos esse valor na análise. Para avaliar se a abundância de uma espécie de planta
determina o número das associações com as formigas, realizamos uma regressão linear
entre a abundância de cada espécie de planta e o número de formigas observadas em
associação com cada planta.
Resultados
Amostramos 20 plantas, sendo 13 de espécies diferentes. Dentre as formigas,
encontramos 13 espécies diferentes, sendo que as mais abundantes foram
Crematogaster sp (quatro ocorrências), Azteca sp (três ocorrências) e uma formiga não
identificada (três ocorrências). Não houve relação entre a abundância das plantas e o
número de espécies de formigas que interagiram com elas (F(1,11)=2,9, p=0,12, R2=0,21).
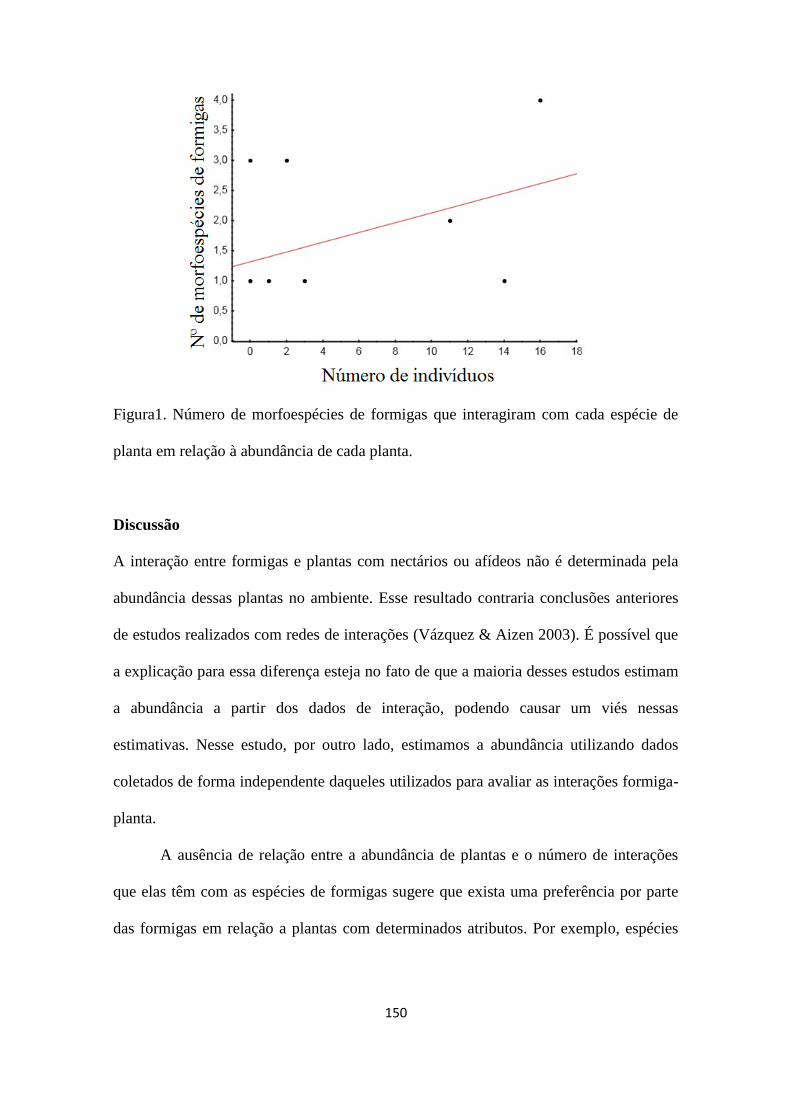
150
Figura1. Número de morfoespécies de formigas que interagiram com cada espécie de
planta em relação à abundância de cada planta.
Discussão
A interação entre formigas e plantas com nectários ou afídeos não é determinada pela
abundância dessas plantas no ambiente. Esse resultado contraria conclusões anteriores
de estudos realizados com redes de interações (Vázquez & Aizen 2003). É possível que
a explicação para essa diferença esteja no fato de que a maioria desses estudos estimam
a abundância a partir dos dados de interação, podendo causar um viés nessas
estimativas. Nesse estudo, por outro lado, estimamos a abundância utilizando dados
coletados de forma independente daqueles utilizados para avaliar as interações formiga-
planta.
A ausência de relação entre a abundância de plantas e o número de interações
que elas têm com as espécies de formigas sugere que exista uma preferência por parte
das formigas em relação a plantas com determinados atributos. Por exemplo, espécies

151
de plantas com maior volume e qualidade do nectar podem atrair maior número de
espécies de formigas (Elias 1983).
Uma explicação alternativa para a ausência de relação entre a abundância e o
número de interações está relacionada com a proteção contra a herbivoria que as
formigas conferem às plantas. Assim, se as plantas dependem dessa defesa (Guimarães
Jr. et al. 2006), é possível que aquelas menos abundantes invistam em mecanismos que
as tornem mais atrativas para as formigas, compensando assim, o efeito da abundância.
Agradecimentos
Agradecemos ao Marcos Vieira e ao Paulo Enrique pela orientação do projeto e ajuda
para fazer o manuscrito.
Referências
Becerra, J.X.I. & D.L. Venable. 1989. Extrafloral nectaries - a defense against ant-
homoptera mutualisms. Oikos, 55:276-280.
Elias, T.S. 1983. Extrafloral nectarines: their structure and distribution. Pp. 174-203. In:
Bentley, B. & Elias, T. (Ed.). The Biology of Nectaries. Columbia University
Press, New York.
Engel, V., M.K. Fischer, F.L. Wackers & W. Volkl. 2001. Interactions between
extrafloral nectaries, aphids and ants: are there competition effects between plant
and homopteran sugar sources? Oecologia, 129:577-584.
Guimarães Jr. P.R., V, Rico-Gray, S.F. Reis & J.N Thompson. 2006. Asymmetries in
specialization in ant–plant mutualistic networks. Proceedings of the Royal
Society B, 273:2041-2047.

152
Jones, C.G.; J.H. Lawton, & M. Shackak. 1994. Organisms as ecosystem engineers.
Oikos, 69: 373-386.
Morin, P. J. 2011. Community Ecology. New Jersey: Blackwell Publishing.
Rico-Gray, V & P.S. Oliveira. 2007. The ecology and evolution of ant-plant
interactions. Chicago: The University of Chicago Press.
Vázquez, D.P. & M.A. Aizen. 2003. Null model analyses of specialization in plant
pollinator interactions. Ecology, 84:2493–-2501.

153
Onde os similares também têm vez: árvores com morfologia foliar semelhante
coexistem em uma floresta secundária
Caroline Marques Dracxler, Joselândio Corrêa Santos, Ludmila Rattis e Bianca Ferreira
da Silva
Introdução
A exclusão competitiva é um processo pelo qual espécies mais competitivas excluem
outras espécies que adquirem de forma menos eficiente o mesmo recurso (Gause 1934).
Processos de segregação de nicho fazem com que algumas espécies coexistam por meio
de mecanismos que reduzem ao máximo a sobreposição do uso do recurso (Abrams
1975, MacArthur & Levins 1967). Nesse sentido, se duas espécies muito similares
ocupassem o mesmo local, a espécie menos competitiva seria excluída pela espécie
mais competitiva. Mas se a espécie menos competitiva fosse suficientemente diferente
em relação à espécie mais competitiva, ambas seriam mantidas no sistema. Assim,
mecanismos de seleção podem limitar a similaridade entre os indivíduos que ocorrem
em uma comunidade, favorecendo a coexistência de espécies ecologicamente distintas.
A segregação de nicho entre espécies pode ser ainda mais limitada em plantas,
pois elas utilizam os mesmos recursos básicos, como dióxido de carbono, água, luz e
nutrientes, gerando sobreposição no uso de recursos. Esses recursos podem ocorrer de
forma especialmente limitada em florestas tropicais que ocorrem em solos pobres em
nutrientes, como as florestas de terra firme na Amazônia Central (Chauvel 1983). Em
florestas secundárias na Amazônia, a disponibilidade de recursos como água e
nutrientes pode ser ainda mais restrita (Mesquita 2000), intensificando as relações
competitivas entre as plantas. Dessa forma, modificações nas estratégias de obtenção e
captação dos recursos permitiriam que um maior número de espécies coexistisse.

154
Algumas características das plantas, como a morfologia foliar, refletem as
estratégias de captação e uso de recursos. A arquitetura das folhas e o seu
posicionamento nas plantas, por exemplo, indicam como a planta capta luz e retém
calor. Folhas grandes são capazes de reter mais calor e, consequentemente, transpiram
mais do que folhas menores. Isso leva à maior perda de água pela planta e,
consequentemente, à maior absorção de água e nutrientes no solo.
Como a limitação por similaridade permitiria que plantas que utilizam os
mesmos recursos básicos coexistissem, avaliamos se plantas em uma mesma
comunidade utilizam os recursos de forma diferencial. Assim, esperamos que duas
plantas vizinhas apresentem morfologias foliares distintas.
Métodos
Realizamos o estudo em uma floresta secundária de terra firme próxima à Reserva do
Km 41, na Amazônia Central, situada a 80 km ao norte de Manaus. Selecionamos uma
área de capoeira em uma fazenda abandonada há cerca de 30 anos, localizada às
margens da estrada ZF3.
Amostramos 16 pares de árvores ao longo de três transectos de 50 m paralelos à
estrada. A cada 10 m ao longo dos transectos, selecionamos um indivíduo focal e seu
vizinho mais próximo, ambos com circunferência à altura do peito (CAP) maior que 10
cm. Buscamos pares de árvores de modo a maximizar a variação da distância entre
vizinhos e então medimos a distância entre as árvores de cada par. Além disso, somente
selecionamos pares de indivíduos que possuíam ramos à altura máxima aproximada de
5 m. Em seguida, amostramos três ramos de cada indivíduo com o auxílio de um podão,
e selecionamos duas folhas no meio de cada ramo, totalizando seis folhas por indivíduo.
Descartamos as folhas com marcas de herbivoria nas margens das folhas. Medimos o

155
comprimento e a largura de cada folha, a fim de obter a média do índice de forma
(índice de forma = comprimento / largura) e a área da folha (área da folha = π *
comprimento * largura) por indivíduo. Em seguida, os dois índices foram padronizados,
gerando valores entre zero e um, representando os menores e maiores valores,
respectivamente.
Através dos valores padronizados de atributo, calculamos a distância euclidiana
entre os vizinhos gerando assim um índice de dissimilaridade para cada par. Os valores
do índice de dissimilaridade e a distância entre os vizinhos também foram padronizados
entre 0 e 1, a partir dos quais realizamos um teste de canto vazio. Esse teste demarca um
limite mínimo esperado para a observação dos pontos na análise, delimitando uma área
vazia no canto inferior esquerdo do gráfico. Assim, esperamos que o valor do índice de
dissimilaridade mínima diminua com o aumento da distância entre os vizinhos.
Resultados
A distância média entre os vizinhos foi de 1,44 m (DP=±0,89). Ao contrário do
esperado, plantas mais próximas umas das outras não apresentaram maior índice de
dissimilaridade (p=0,29; Figura 1).

156
Figura 1. Relação entre o índice de dissimilaridade e a distância do vizinho mais
próximo. De acordo com a hipótese de limitação por similaridade, os pontos deveriam
estar acima da linha pontilhada.
Discussão
Os resultados sugerem que indivíduos localizados próximos uns aos outros podem ter
modos similares de obtenção de recurso, logo, não houve limitação por similaridade.
Considerando que há um tipo ótimo de folha que maximiza a competitividade das
plantas em um dado ambiente, plantas com folhas parecidas ao tipo ótimo seriam fortes
competidoras, ao passo que plantas com folhas diferentes do tipo ótimo seriam menos
competitivas. Diferentes cenários de capacidade competitiva das espécies podem
explicar como plantas pouco dissimilares coexistiriam.
Em um primeiro cenário, quando dois indivíduos com o tipo de folha ótimo
ocorrem próximos, um não consegue excluir o outro por serem competidores
equivalentes. Em um segundo cenário, indivíduos com folhas diferentes do tipo ótimo
Distância do vizinho mais próximo
Dis
sim
ila
rid
ad
e
0 0,2 0,4 0,6 0,8 1,0
00
,20
,40
,60
,81
,0

157
ocorrendo próximos também não conseguiriam se excluir, por estabelecerem uma
relação competitiva equivalente entre si.
As relações competitivas entre indivíduos também são dependentes das
condições ambientais e da disponibilidade de recursos. Em ambientes dinâmicos, como
florestas em regeneração (Mesquita 2000), essas relações de competição poderiam ser
alteradas. Desse modo, a estocasticidade ambiental poderia levar a uma alternância
entre a hierarquia competitiva das espécies, ora favorecendo as espécies competidoras
mais fortes, ora as mais fracas. Assim, a longo prazo, as capacidades competitivas das
plantas equivaler-se-iam, permitindo sua coexistência (Wright 2002).
Nesse sentido, quando duas plantas com capacidades competitivas diferentes se
estabelecem próximas, seria esperado que a melhor competidora excluiria a
competidora mais fraca, por não serem competitivamente equivalentes. Assim, é
possível que exista um limite de dissimilaridade ou um intervalo de similaridade entre
organismos que coexistem.
Agradecimentos
Agradecemos ao Marcel Baby pela orientação e ajuda na construção das ideias e ao
professor Adriano Melo pela ajuda com as análises estatísticas.
Referências
Abrams, P. 1975. Limiting similarity and the form of the competition coefficient.
Theoretical Population Biology, 8:356-375.
Chauvel, A. 1983. Os latossolos amarelos, álicos e argilosos dentro dos ecossistemas
das bacias experimentais do INPA e da região vizinha. Acta Amazonica, 12:47-60.

158
Gause, G.F. 1934. The struggle for existence. Williams & Wilkins, Baltimore. New
York: Halfner.
MacArthur, R. H. & R. Levins. 1967. The limiting similarity, convergence, and
divergence of coexisting species. The American Naturalist, 101:377-385.
Mesquita, R.C.G. 2000. Management of advanced regeneration in secondary forests of
the Brazilian Amazon. Forest Ecology and Management, 130:131-140.
Wright, S.J. 2002. Plant diversity in tropical forests: a review of mechanisms of species
coexistence. Oecologia, 130:1-14.

159
Plantas com folhas menores impedem o estabelecimento de minas e galhas
Daniel Passos, Tatiane Calaça, Lorena Pinto e Vidal Carrascosa
Introdução
O consumo de plantas por outros organismos é uma das relações bióticas mais
freqüentes em ambientes terrestres (Schoonhoven et al. 2005). Estas interações
ecológicas estão associadas a processos co-evolutivos, nos quais os herbívoros
desenvolvem estratégias de ataque, enquanto as plantas desenvolvem defesas para
combatê-los (Carmona et al. 2011). A relação dos herbívoros com as plantas pode
ocorrer por duas vias principais: predação, que acarreta na morte de um indivíduo
vegetal, ou parasitismo, quando apenas parte do organismo vegetal é consumido
(Millard & Way 2011).
Entre os organismos que se alimentam de plantas, existem parasitas que se
estabelecem no interior dos tecidos vegetais, denominados endofíticos (Ribeiro et al.
1999). Estes parasitas são classificados em dois grupos: os minadores e os galhadores.
Minadores são insetos que se estabelecem entre as epidermes das folhas, consumindo o
mesófilo foliar. Já os galhadores, que podem ser representados por bactérias, fungos e
insetos, causam alterações anatômicas na folha, induzindo tumores e drenando recursos
floemáticos da planta (Schoonhoven et al. 2005).
As plantas apresentam diversas estratégias para combater a herbivoria (Carmona
et al. 2011). Estas estratégias podem atuar isoladamente ou em conjunto para impedir a
infecção de parasitas ou reduzir os danos causados por eles aos tecidos vegetais (Millard
& Way 2011). Entre estas formas de defesa, existem mecanismos físicos, como o
espessamento da cutícula, químicos, como a produção de metabólitos secundários, e
fisiológicos como a abscisão da folha infectada por parasitas (Silva & Batalha 2011).

160
A abscisão foliar constitui uma estratégia fisiológica para a eliminação de folhas
danificadas (Raven et al. 1992). Na perspectiva da planta, os custos para eliminação de
uma folha podem variar de acordo com a quantidade de recursos investidos na mesma.
A perda de folhas pequenas, produzidas com menor investimento de recursos, seria
menos custosa do que a perda de folhas grandes. Já na perspectiva do parasita, a
infecção de uma folha pequena pode ser menos favorável, pois a quantidade de tecidos
consumidos pelos minadores e de nutrientes drenados pelos galhadores também seria
menor, prejudicando seu desenvolvimento (Schoonhoven et al. 2005). O tempo de
suscetibilidade à infecção também pode afetar a ocorrência de parasitas. Este período
coincide com a duração do crescimento da folha (Aide 1992). Folhas maiores demoram
mais para se desenvolver e podem ser mais vulneráveis ao parasitismo.
O tamanho foliar acarreta custos distintos para a planta e para o parasita. Nesse
estudo, analisamos se o tamanho da folha influencia a ocorrência de parasitas
endofíticos. Propomos a hipótese de que espécies com folhas maiores têm maior
probabilidade de serem parasitadas. Dessa forma, esperamos maior ocorrência de
parasitas endofíticos com o aumento do tamanho da área foliar.
Métodos
Nós desenvolvemos este estudo em uma floresta tropical de terra firme, inserida na Área
de Relevante Interesse Ecológico do PDBFF reserva km 41, localizada a 80 km ao norte
de Manaus - AM, Brasil. Coletamos folhas de todos os indivíduos de espécies lenhosas
ao longo de um transecto de 400 m na borda de uma estrada. Posteriormente,
morfotipamos as plantas e inspecionamos as folhas à procura de galhas e minas.
Consideramos a ausência de parasitas em um morfotipo quando pelo menos cinco
indivíduos não apresentaram evidências de parasitismo. Os morfotipos com pelo menos

161
um indivíduo com galhas ou minas foram consideradas parasitadas. Calculamos a área
foliar média para cada espécie e para este cálculo, usando a menor unidade destacável
da planta (folha, folíolo ou foliólulo). Para testar se o tamanho da área foliar influencia a
ocorrência de galhas ou minadores na folha, fizemos regressões logísticas utilizando
como variável preditora o logaritmo neperiano da área foliar e como variável resposta a
presença ou ausência de galhas e de minas.
Resultados
Nós coletamos ramos de 261 indivíduos classificados em 45 morfo-espécies de plantas.
Dessas, 27 apresentaram galhas, 14 apresentaram minas e 11 morfo-espécies
apresentaram ambos os tipos de parasitas endofitícos. A amplitude de área foliar variou
de 0,01 a 718,87 cm2.
A presença dos galhadores teve uma relação positiva com a área foliar (χ2 =
14,35, g.l. = 1, p < 0,01, Figura 1) e o tamanho foliar mínimo para a ocorrência desses
parasitas foi de 2,14 cm2. Um padrão similar foi encontrado para parasitas minadores (χ
2
= 10,68, g.l. = 1, p < 0,01, Figura 2), entretanto, o tamanho foliar mínimo para o seu
estabelecimento foi de 24 cm2.
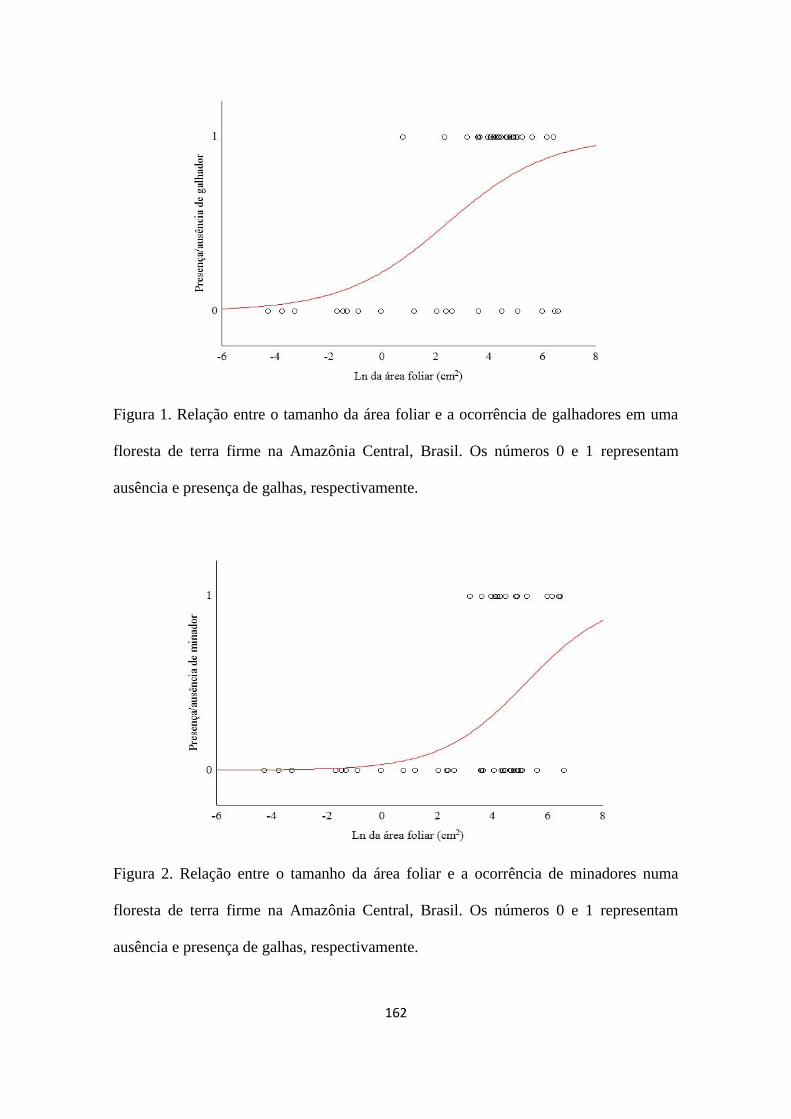
162
Figura 1. Relação entre o tamanho da área foliar e a ocorrência de galhadores em uma
floresta de terra firme na Amazônia Central, Brasil. Os números 0 e 1 representam
ausência e presença de galhas, respectivamente.
Figura 2. Relação entre o tamanho da área foliar e a ocorrência de minadores numa
floresta de terra firme na Amazônia Central, Brasil. Os números 0 e 1 representam
ausência e presença de galhas, respectivamente.

163
Discussão
A ocorrência de parasitismo por minadores e galhadores foi positivamente associada
com o tamanho da folha. Folhas são mais vulneráveis ao ataque de parasitas durante o
período de crescimento (Aide 1992). Por isso, folhas maiores seriam mais suscetíveis à
infecção, por terem um tempo de desenvolvimento maior.
O tamanho foliar mínimo pode ter sido selecionado para ocorrência de parasitas
endofíticos. Os galhadores se estabeleceram em folhas a partir de 2,14 cm² de área
foliar, enquanto os minadores ocorreram em folhas 11 vezes maior. O tipo de recurso
explorado difere entre os parasitas, enquanto os galhadores drenam os nutrientes dos
vasos floemáticos, os minadores consomem os tecidos estruturais da folha, dependendo
assim de uma área foliar maior.
A ausência de galhadores e minadores em folhas pequenas poderia ser resultante
da rápida abscisão das folhas pequenas logo após a infecção por parasitas, devido ao
baixo custo de sua perda. Neste caso, o parasitismo ocorre, mas o parasita não se
desenvolve até a fase adulta por causa da abscisão foliar. Uma forma de demonstrar isso
seria monitorar o desenvolvimento de folhas pequenas para evidenciar se a abscisão
foliar ocorre como resposta a infecção por parasitas. Por outro lado, a exclusão de
parasitas de folhas menores pode ter ocorrido ao longo do tempo evolutivo. Nas
linhagens de plantas parasitadas que tiveram diminuição do tamanho da folha, seus
parasitas podem não ter tolerado esta redução do tamanho da folha e se extinguiram.
O estabelecimento em folhas menores parece não ser vantajoso para parasitas
endofíticos. Entretanto, o processo que gerou esta estratégia anti-predatória nas plantas
ainda não foi explicado. A redução da área foliar pode ter se desenvolvido como uma
adaptação contra o parasitismo foliar, entretanto, pode também ser devido a um

164
processo de exaptação, em que essa redução foliar se deu em resposta a outras pressões
seletivas, mas que confere vantagem em relação ao parasitismo.
Referências
Aide, M.T. 1992. Dry season leaf production: an escape from herbivory. Biotropica
24:532-537.
Carmona, D., Lajeunesse, M.J. & Johnson, T.J. 2011. Plant traits that predict resistence
to herbivores. Functional Ecology, 25:358-367.
Millard, P. & Way, D.A. 2011. Tree competition and defense against herbivores:
currency matters when counting the cost. Tree Physiology, 31:579-581.
Raven, P.H., R.F. Evert & S.E. Eichhorn. 1992. Biologia vegetal. Rio de Janeiro:
Guanabara Koogan.
Ribeiro, J.E.L.S, M.J.G. Hopkins, A. Vicentini, C.A. Sothers, M.A.S. Costa, J.M. Brito,
M.A.D. Souza, L.H.P. Martins, L.G. Lohmam, P.A.C.L. Assunção, E.C. Pereira,
C.F. Silva, M.R. Mesquita & L.C. Procópio. 1999. Flora da reserva Ducke: Guia
de identificação das plantas vasculares de uma floresta de terra firme na
Amazônia Central. Manaus: INPA.
Schoonhoven, L.M., Van Loon, J.J.A., Dicke, M. 2005. Insect-plant biology.New York:
Oxford University Press.
Silva, D.M. & Batalha, M.A. 2010. Defense syndromes against herbivory in a cerrado
plant community. Plant Ecology, 212:181-193.

165
Km 41 – projetos individuais
Diferença no comportamento de forrageio de formigas associadas a nectários
extraflorais e exsudato de afídeos
Ana Carolina Vieira Pires
Introdução
O mutualismo é uma interação reciprocamente benéfica entre duas espécies, na qual os
ganhos para cada parceiro são quantificados em termos de sobrevivência e sucesso
reprodutivo (Bronstein 2009). Essa interação pode ser obrigatória, na qual as espécies
estão vitalmente associadas, ou facultativa, em que as espécies sobrevivem mesmo sem
estarem associadas. No mutualismo facultativo a interação pode acontecer sempre entre
as mesmas espécies ou pode haver variação nas espécies associadas (mutualismo
difuso).
A associação entre plantas mirmecófilas e formigas representa um exemplo de
mutualismo difuso. Nesse mutualismo as plantas oferecem uma recompensa não
especializada para atrair formigas, através de nectários extraflorais (NEFs) (Del-Claro
2004, Rosumek et al. 2009). Essas estruturas são glândulas secretoras de néctar
localizadas em órgãos que não estão envolvidos diretamente na polinização (Bronstein
et al. 2006). As formigas associadas a plantas com NEFs exibem um comportamento
agressivo em relação aos herbívoros, afetando positivamente a aptidão da planta, ao
diminuir os danos causados por fitófagos em órgãos vegetativos e reprodutivos (Del-
Claro et al. 1996).

166
Uma associação semelhante à de plantas com NEFs e formigas ocorre quando
algumas plantas estão associadas a hemípteros que se alimentam da seiva do floema e
liberam um exsudato composto por uma mistura de açúcar, aminoácidos, amidos e
proteínas (Auclair 1963 apud Del-Claro & Oliveira 1999). Algumas formigas se
alimentam desse exsudato liberado pelos hemípteros e os defendem contra predadores e
parasitoides e, portanto, acabam atuando indiretamente na vigilância das plantas
(Moreira & Del-Claro 2005). O patrulhamento de formigas tanto em plantas com NEFs
quanto em plantas com hemípteros diminui a herbivoria, aumenta a aptidão da planta e
aumenta a sobrevivência de hemípteros (Moreira & Del-Claro 2005, Styrsky & Eubanks
2010).
Ainda que as plantas sejam um recurso abundante e atrativo, a quantidade de
nitrogênio nos tecidos vegetais é baixa e a obtenção desse nutriente parece ser um
problema para os insetos herbívoros (Schowalter 2006, Speight et al. 2008). Nesse
sentido, é possível que o comportamento de formigas que defendem as plantas com
NEFs ou com hemípteros secretores de exsudato contra o ataque de outros insetos,
dependa da qualidade do recurso defendido e da demanda nutricional das formigas.
Como esses recursos podem ter concentrações distintas de nitrogênio, o ataque de
formigas a esses insetos pode representar um ganho extra do nutriente limitante.
Portanto, formigas submetidas a uma maior limitação de nitrogênio devem investir mais
na predação sobre herbívoros, devido ao seu maior teor de nitrogênio.
Embora as formigas possam mudar sua dieta em resposta a déficits nutricionais
(Wilder & Eubanks 2010), esse comportamento nunca foi investigado em sistemas
naturais. Uma vez que o exsudato de hemípteros contém mais nitrogênio que o néctar de
NEFs (P. E. Peixoto, com. pess.), avalio a hipótese de que formigas que se alimentam
do recurso vegetal de menor conteúdo nitrogenado predem mais prontamente outros

167
insetos presentes na planta. Assim, espero que as formigas associadas a NEFs demorem
menos tempo para remover os herbívoros do que aquelas que se alimentam de exsudato,
bem como recrutarão maior quantidade de indivíduos para removê-lo.
Métodos
Área de estudo
Realizei esse estudo na borda de uma floresta criada por uma estrada ao longo da
reserva do Km 41, localizada a 80 km ao norte de Manaus, na Amazônia Central, Brasil.
O local está inserido em uma ARIE (Área de Relevante Interesse Ecológico), gerida
pelo ICMBio e pelo Projeto de Dinâmica Biológica de Fragmentos Florestais (PDBFF).
Coleta dos dados
Selecionei 20 indivíduos de diferentes espécies de plantas que possuíam nectários
extraflorais e não possuíam hemípteros e 20 indivíduos com hemípteros secretando
exsudato açucarado e sem NEFs. Em metade dos indivíduos amostrados com NEFs e
metade dos com exsudato havia presença de formigas do gênero Crematogaster,
enquanto na outra metade das plantas de cada grupo havia formigas do gênero Azteca.
Em nenhuma das plantas amostradas havia a presença simultânea dos dois gêneros de
formigas. As formigas de ambos os gêneros são dominantes, agressivas e recrutam
operárias para defender o recurso vegetal do qual se alimentam (Holldobler & Wilson
1990).
Para simular a presença de um herbívoro, coloquei um cupim na folha mais
próxima ao recurso (NEF ou exsudato) onde não havia presença de formiga. Essa folha
distava aproximadamente 5 cm do recurso. Após depositar a isca, cronometrei o tempo
da chegada da formiga até o cupim, o tempo que a formiga levou para remover o cupim

168
após encontrá-lo e o número de formigas que removeram o cupim. O tempo máximo
estipulado para a observação foi de 5 min. Para as formigas que não removeram o
cupim, mas permaneceram sobre ele em processo de remoção durante o tempo de
observação, considerei como tempo máximo de 300 s. Excluí das análises, as amostras
nas quais as formigas não responderam à presença do cupim.
Análises dos dados
Para avaliar a diferença na resposta de formigas quanto ao recurso patrulhado (nectários
extraflorais ou hemípteros) utilizei análise de variância (ANOVA) de dois fatores.
Minhas variáveis respostas foram o tempo de chegada das formigas ao cupim, o tempo
de remoção do cupim da folha, o tempo total entre a deposição do cupim até a sua
remoção e a quantidade de formigas que atuaram na remoção. Considerei as seguintes
variáveis como preditoras: tipo de recurso oferecido (nectários extraflorais e exsudado)
e o gênero da formiga (Crematogaster e Azteca). Transformei os dados de tempo de
chegada e de quantidade de formigas que removeram o cupim com logaritmo para
homogeneizar as variâncias.
Resultados
Das 20 plantas com NEFs, em três não houve resposta de Crematogaster sp. e em uma
não houve resposta de Azteca sp., apesar de ambas permanecerem patrulhando os ramos
durante as observações. Em relação às plantas que possuíam hemípteros e eram
patrulhadas por Crematogaster sp., apenas em duas as formigas não chegaram até o
cupim. Todas as demais formigas responderam à presença do cupim durante as
observações.

169
De forma geral, as formigas do gênero Crematogaster demoravam mais de 5
min para a remoção completa do cupim da folha. Isso ocorreu em cinco plantas com
NEFs e nas 10 plantas que tinham hemípteros. Por outro lado, apenas em uma planta
com NEFs e em uma planta com hemípteros que era patrulhada por Azteca sp. a
resposta foi superior a 5 min para a remoção completa do cupim.
Formigas despenderam tempos similares para chegar até o cupim em plantas
com NEFs ou com hemípteros (F(1,34)=2,94; p=0,23; Figura 1). Porém, o tempo de
chegada foi diferente entre os gêneros de formigas (F(1,34)=4,47; p=0,04). A resposta de
formigas do gênero Azteca sp. foi 3,5 vezes mais rápida que as de Crematogaster sp. em
plantas com NEFs e 1,8 vezes mais rápida em plantas com hemípteros. Não houve
interação entre os dois fatores (gênero da formiga e tipo de recurso defendido) em
relação ao tempo de chegada (F(1,34)=3,31; p=0,08).
Figura 1. Tempo de chegada de formigas dos gêneros Crematogaster (●) e Azteca (■)
até o cupim em plantas com os recursos: NEF = nectários extraflorais; HEM=hemíptero
na reserva do km 41, Manaus, Brasil. Os símbolos representam as médias do tempo de
resposta e as barras verticais representam o intervalo de 95% de confiança.

170
O tempo que a formiga despendeu até a remoção completa do cupim da folha foi
diferente dependendo da combinação entre os recursos e entre os gêneros das formigas
(F(1,34)=6,65; p=0,01; Figura 2). Formigas do gênero Crematogaster demoram mais
tempo para remover o cupim quando estavam em plantas com hemípteros do que
quando estavam em plantas com NEFs. Já as formigas do gênero Azteca apresentaram o
mesmo tempo de remoção para ambos os tipos de recurso e esse tempo foi menor do
que o despendido por Crematogaster sp. apenas em plantas que possuíam hemípteros.
Figura 2. Tempo despendido para remoção do cupim por formigas dos gêneros
Crematogaster (●) e Azteca (■) em ambos os recursos: NEF = nectários extraflorais;
HEM=hemíptero na reserva do km 41, Manaus, Brasil. Os símbolos representam as
médias do tempo de remoção e as barras verticais representam o intervalo de 95% de
confiança.
Considerando o tempo que durou toda a ação, desde que coloquei o cupim sobre
a folha até sua remoção completa pelas formigas, não houve diferença no tempo de

171
resposta das formigas entre os recursos defendidos (F(1,34)=0,01; p=0,94). Porém,
formigas do gênero Crematogaster eram mais lentas para executar toda a ação quando
comparadas às do gênero Azteca (F(1,34)=40,11; p<0,001; Figura 3). Não houve interação
entre o gênero de formigas e o tipo de recurso com relação ao tempo total de ação
(F(1,34)=2,40; p=0,13).
Figura 3. Tempo total da ação de formigas dos gêneros Crematogaster (●) e Azteca (■)
em ambos os recursos: NEF = nectários extraflorais; HEM=hemíptero na reserva do km
41, Manaus, Brasil. Os símbolos representam as médias do tempo total e as barras
verticais representam o intervalo de 95% de confiança.
A quantidade de indivíduos de formigas dos dois gêneros mobilizados para a
remoção do cupim foi semelhante (F(1,34)=1,90; p=0,18; Figura 4). A mobilização de
formigas entre os dois tipos de recursos defendidos também foi similar (F(1,34)=3,43;
p=0,07). Não houve interação entre esses dois fatores (F(1,34)=1,15; p=0,29).

172
Figura 4. Número de formigas dos gêneros Crematogaster (●) e Azteca (■) mobilizadas
na remoção do cupim em ambos os recursos: NEF = nectários extraflorais;
HEM=hemíptero na reserva do km 41, Manaus, Brasil. Os símbolos representam as
médias do número de formigas e as barras verticais representam o intervalo de 95% de
confiança.
Discussão
As formigas que patrulhavam nectários extraflorais e exsudato de hemípteros
apresentaram respostas similares quanto ao tempo de chegada até o cupim, tempo total
da ação e número de formigas mobilizadas. Dessa forma, diferentes tipos de recursos
não determinam a intensidade da resposta das formigas. A única exceção ocorreu em
relação ao tempo despendido para a remoção completa do cupim, na qual
Crematogaster sp. foi mais lenta quando estava patrulhando hemípteros. Eventualmente
essas formigas que se alimentam de exsudato podem ter sua preferência por herbívoros
reduzida. Assim, as formigas podem realizar uma avaliação da qualidade nutricional da
presa depois de encontrá-la e demorar mais para removê-la.

173
O folhiço de florestas tropicais é úmido e rico em nitrogênio que encontra-se
homogeneamente distribuído (Yanoviak & Kaspari 2000). Como espécies de
Crematogaster e Azteca forrageiam tanto na planta quanto no solo (Holldobler &
Wilson 1990), elas podem procurar por alimentos que complementem sua dieta. Assim,
independente da qualidade do recurso explorado nas plantas (NEFs ou exsudato), as
formigas podem apresentar condições nutricionais semelhantes, gerando respostas
similares com relação à presença de herbívoros.
Wilder & Eubanks (2010), ao desenvolverem um experimento em laboratório,
mostraram que a diferença na preferência da presa por formigas é mediada pela sua
demanda nutricional. Com base nisso, esses autores sugeriram que as plantas poderiam
manipular as formigas quanto ao herbívoro que as ataca. Os resultados encontrados
nesse estudo não corroboraram essa hipótese, indicando que em ambientes naturais,
recursos de qualidade nutricional distintos, como NEFs e exsudato de hemípteros, não
afetam o comportamento de forrageio das formigas. Como nesses sistemas naturais,
formigas dispõem de uma variedade de fontes alternativas de alimento, é possível que
elas adaptem suas estratégias de forrageio para compensar eventuais defasagens
nutricionais oriundas do forrageio nos NEFs ou no exsudato de hemípteros.
Agradecimentos
Após trinta anos de convivência, isolados na Amazônia, chega a ser difícil saber como
agradecer a cada um que fez parte da turma do EFA 2012. Começando por Paulinho
Bom (Rainbow Master) e Paulinho Mau (Batman) pela incrível oportunidade de
participar do curso e por todo apoio durante esses dias. Encarar trinta dias com essa
galera e acordar de bom humor não é para qualquer um. Ao Seu Jorge e Dona Eduarda

174
pela paciência e pela preocupação em fazer a soja nossa de cada dia. O curso não seria o
mesmo sem o vasto cardápio de todo dia (dá-lhe suco de caju!).
A todos os professores que fizeram a diferença nesses dias: Felipe, Thiago
Toyoyo, Richard, Mike, Esther, Cassiano, Glauco, Catá, Dé, Jack Sparrow, Marcel
Baby, Jansen e Aline. O conhecimento que vocês me passaram será para toda a vida! E
aquele abraço especial aos monitores Thiago Gloss Xexéu e Marcos Barbudinho. Valeu
pela ajuda em todos os momentos de necessidades. Mais do que monitores, vocês são
parte da turma.
E o que falar das pessoas dessa turma? Depois de trinta anos de convivência é
difícil pensar em passar um dia sem escutar as fofocas no café da manhã, saber quem
dormiu torto na rede e quem está devendo relatório. Cada um de vocês acrescentou um
pouquinho em mim. Primeiro agradeço ao Daniel Lagartão pelo carinho, atenção e
ajuda em todos os momentos. Obrigada pelos momentos felizes que compartilhamos.
Agradeço à Alanis (isso, aham) e Carol da Silva pela parceria, amizade e conversas de
confessionário. Aos mineiros Renatinha Pocket (e os segredos divididos), Sarah
Baranga, Joseph, Léo Mr. Bigodinho, Mari Exú (e seu jeitinho de mãe professora) e
Ludmilla Miss-Kessy (e os comentários sem noção de cada dia). Valeu por fazerem me
sentir mais perto de casa. À Dani Atoladinha, que com toda sua maturidade e
criatividade cativou a todos. Ao Vidal e Tatu, um estrangeiro e um tradutor (para hablar
español basta colocar la língua na ponta de los dientes e hablar con fuerza, certo?). Aos
nobres Conde Harry Potter e Condessa de Hogwartts (também conhecida por Lorrhayne
Xaiene Vanessão). Visitarei vocês na Hungria. À Lore Pole Dance e sua risada
contagiante. Você tem futuro, gata! À Tati (e suas árvres) e seu jeito carinhoso de
chamar todas as meninas de vadia. Ao Bruno Baba Pau que curtia minhas músicas mais
do que eu mesma. À Bia Bis-kate e Randolpho Hobbit pelos bons momentos.

175
A vocês, todo o amor que houver nessa vida!
Referências
Bronstein. J.L., R. Alarcón & M. Geber. 2006. The evolution of plant–insect
mutualisms. New Phytologist, 172:412-428.
Bronstein, J.L. 2009. The evolution of facilitation and mutualism. Journal of Ecology,
97:1160-1170.
Del-Claro, K., V. Berto & W. Réu. 1996. Herbivore deterrence by visiting ants
increases fruit-set in an extrafloral nectary plant Qualea multiflora
(Vochysiaceae) in cerrado vegetation. Journal of Tropical Ecology, 12:887-
892.
Del-Claro, K. & P.S. Oliveira. 1999. Ant-homoptera interactions in a neotropical
savanna: the honeydew-producing treehopper, Guayaquila xiphias
(Membracidae), and its associated ant fauna on Didymopanax vinosum
(Araliaceae). Biotropica, 31:135-144.
Del-Claro, K. 2004. Multitrophic relationships, conditional mutualisms, and the study of
interaction biodiversity in tropical savannas. Neotropical Entomology, 33:665-
672.
Holldobler, B. & E.O. Wilson. 1990. The ants. Harvard University Express: Harvard.
Moreira, V.S.S. & K. Del-Claro. 2005. The outcomes of an ant-treehopper association
on Solanum lycocarpum St. Hill: increased membracid fecundity and reduced
damage by chewing herbivores. Neotropical Entomology, 34:881-887.
Rosumek, F.B., F.A.O Silveira, F.S. Neves, N.P.U. Barbosa, L. Diniz, Y. Oki, F.
Pezzini, G.W. Fernandes & T. Cornelissen. 2009. Ants on plants: a meta-
analysis of the role of ants as plant biotic defenses. Oecologia, 160:537-549.

176
Schowalter, T.D. 2006. Insect ecology, an ecosystem approach. Elsevier: London.
Speight, M.R., M.D. Hunter & A.D. Watt. 2008. Ecology of insects, concepts and
applications. Blackwell Publishing: Oxford.
Styrsky, J.D. & M.D. Eubanks. 2010. A facultative mutualism between aphids and an
invasive ant increases plant reproduction. Ecological Entomology, 35:190-199.
Yanoviak, S.P. & M. Kaspari. 2000. Community structure and the habitat templet: ants
in the tropical forest canopy and litter. Oikos, 89:259-266.
Wilder, S.M. & M.D. Eubanks. 2010. Extrafloral nectar content alters foraging
preferences of a predatory ant. Biology Letters, 6:177-179.

177
Convergência de características funcionais foliares entre uma floresta em
regeneração e uma floresta primária na Amazônia Central
Ana Claudia Rorato
Introdução
A floresta Amazônica tem sido alvo de elevadas taxas de desmatamento, principalmente
para a formação de áreas de campos de cultivo ou de pastagens. No entanto, essas áreas
são geralmente abandonadas após um período de cinco a dez anos de uso devido ao
declínio na produtividade (Mesquita 2001). Isso resulta em um mosaico de áreas
florestais em processo de regeneração (Quesada et al. 2009). As florestas em
regeneração diferem dos remanescentes de floresta primária em relação à estrutura
vertical, às condições ambientais e à composição de espécies (Laurence 1998, Mesquita
2001). A floresta em regeneração apresenta estrutura vertical composta por um dossel
mais baixo e menos denso e um sub-bosque mais aberto, quando comparada à floresta
primária. Essa diferença permite maior penetração de luz, o que acarreta na elevação da
temperatura do ar e do solo (Benitez-Malvido 1998, Casenave et al. 1995, Kapos et.al
1997).
O aumento da temperatura resulta na diminuição da umidade relativa do ar e do
solo, acompanhada de uma diminuição da disponibilidade de água. Embora nas florestas
em regeneração a maior entrada de luz favoreça a fotossíntese, o ambiente relativamente
mais seco é mais propício à perda de água (Casenave et al.1995). Em contrapartida, nas
florestas primárias as plantas estão sujeitas a uma menor incidência de luz direta no solo
e por isso não sofrem tanto com a diminuição da umidade relativa. (Casenave et
al.1995, Kapos et al. 1997). A estruturação das comunidades vegetais é influenciada
pelo conjunto de condições abióticas (temperatura, umidade e disponibilidade de

178
recursos) e bióticas (competição, predação, mutualismo) que selecionam quais grupos
de espécies são capazes de crescer e se reproduzir num local (Diamond 1975, Lavorel
& Garnier 2002).
As diferenças abióticas existentes entre as florestas em regeneração e as florestas
primárias favorecem grupos de plantas com estratégias ecológicas distintas. Áreas
desmatadas ou em estádios iniciais de sucessão são colonizadas por espécies que são
favorecidas pela elevada incidência luminosa. Com o avanço da sucessão nas florestas
em regeneração, espécies tardias podem se estabelecer, resultando em uma sobreposição
de espécies de estádios sucessionais iniciais e tardios (Mesquita 2001).
A regulação entre a taxa de assimilação de CO2 e a perda de água nas plantas
pode ser feita através de mecanismos morfológicos e/ou fisiológicos ligados às
estratégias de captação de luz e armazenamento de água das plantas. Dessa forma,
características foliares das plantas variam em resposta a diferentes condições do
ambiente e disponibilidade de recursos (Tyree 2007). Dentre as características
relacionadas às funções foliares destacam-se a área foliar, a forma foliar, o número de
folhas por ramo, a massa foliar e a capacidade de acúmulo de água (Cianciaruso et al.
2009, Cornelissen et al. 2003).
Plantas com área foliar maior possuem maior taxa de transpiração, e
consequentemente maior taxa fotossintética e perda de água. O mesmo ocorre para a
forma das folhas. Folhas mais arredondadas aumentam a taxa de perda de água quando
comparadas a folhas mais alongadas. Portanto, a redução da área foliar e a produção de
folhas mais alongadas pode ser considerada como uma estratégia contra a perda de
água. Por outro lado, folhas mais espessas podem conter o mesmo número de células e
de cloroplastos e, assim, manter a taxa fotossintética elevada, com uma menor
superfície de transpiração. A capacidade de armazenamento de água nas folhas permite

179
que plantas resistam à baixa disponibilidade de água (Pimentel 1998, Taiz & Zeiger
2002). Desse modo, a diferença na disponibilidade de água e quantidade de luz que
chega no sub-bosque devem levar a combinações particulares de características
funcionais foliares relacionadas à captação de luz e armazenamento de água.
O objetivo desse estudo foi entender de que maneira a diversidade funcional de
características foliares relacionadas ao armazenamento de água e a captação de luz em
espécies arbóreas varia entre uma floresta em regeneração e uma floresta primária.
Assim, proponho duas hipóteses: (1) na floresta em regeneração os indivíduos arbóreos
juvenis do sub-bosque possuem maior capacidade de armazenamento de água nas
folhas; (2) na floresta primária os indivíduos juvenis otimizam a captação de luz através
da forma e tamanho das folhas. As minhas previsões foram: (i) os indivíduos juvenis na
floresta em regeneração podem armazenar mais água nas folhas do que os indivíduos da
floresta primária; (ii) na floresta em regeneração as folhas são menores e mais
alongadas, enquanto na floresta primária as folhas são maiores e mais arredondadas;
(iii) a variação das características funcionais foliares (área foliar, forma foliar e
quantidade de água armazenada) entre os indivíduos é maior na floresta em
regeneração do que na floresta primária.
Métodos
Área de estudo
Esse estudo foi desenvolvido na AIRE PDBFF sítio Km 41 (2°24’S, 59°43’O), co-
gerida pelo Projeto Dinâmica Biológica de Fragmentos Florestais (PDBFF) e ICMBIO.
A reserva está situada a 80 km ao norte de Manaus e abrange uma área de
aproximadamente 10.000 ha de floresta contínua de terra firme.

180
Os dados para esse estudo foram coletados em dois tipos de floresta. A primeira
foi uma floresta primária contínua, enquanto a segunda foi uma floresta em regeneração
que possui por volta de 30 anos. Essa floresta em regeneração é resultado da conversão
de uma área de floresta primária em pastagem que após alguns anos de uso foi
abandonada.
Características funcionais das folhas
Para comparar as características funcionais foliares (área foliar, forma foliar e DMF
(diferença entre massa túrgida e massa seca) nas duas florestas, coletei folhas de
indivíduos arbóreos juvenis. Em cada tipo de floresta amostrei seis pontos distantes 20
m entre si ao longo de um transecto. Para cada ponto amostrei 15 indivíduos de espécies
diferentes, e coletei três folhas para a mensuração das características funcionais
foliares. As folhas foram coletadas nos dois ramos mais altos para diminuir variações
devido à idade da folha. A altura dos indivíduos também foi padronizada entre 0,5 m e
1,5 m para diminuir variações em relação a idade dos indivíduos. Cada indivíduo foi
considerado uma amostra e cada ponto uma réplica. Para cada indivíduo os valores das
características das folhas foram somados e a média entre eles foi assumida para o
indivíduo.
A partir das folhas coletadas medi as três características foliares: DMF
(diferença entre a massa túrgida foliar e a massa seca), área foliar e forma foliar. O
DMF foi usado como medida da capacidade de acúmulo de água nas folhas e foi obtido
através da diferença entre a massa túrgida foliar e a massa seca. A massa túrgida foliar
média de cada indivíduo foi inferida pesando um pedaço de 4 cm2
de cada folha que
ficou imerso em água por volta de duas horas. Após imersão em água, esses pedaços de
folhas foram secos em um forno e pesados novamente para obtenção da massa seca

181
média de cada indivíduo. A área da folha foi calculada através da fórmula da elipse (π x
raio maior x raio menor / 4). A forma foliar foi obtida através da razão comprimentomédio
/ larguramédia das três folhas coletadas para cada planta. Se o valor dessa razão é próximo
de 1 a forma foliar é arredondada, o que aumenta a captação de luz e consequentemente
a perda de água. Por outro lado, quanto maior o valor dessa razão mais alongada é a
folha, fazendo com que a transpiração e a perda de água sejam menores.
Análises
Para avaliar a diferença nas características funcionais foliares entre a floresta em
regeneração e a floresta primária realizei um teste t para cada variável medida. Para
testar se a variação das características funcionais foliares entre os indivíduos da mesma
floresta foi maior na floresta em regeneração realizei o teste de homogeneidade de
variâncias de Levene.
Resultados
Os resultados indicam que não existe diferença entre a área foliar dos indivíduos da
floresta em regeneração e da floresta primária (t = 0,84; gl = 180; p = 0,40 Figura 1). A
média da área foliar foi de 87,52 ± 81,84 cm2 (média ± dp) para a floresta em
regeneração e 78,30 ± 64,25 cm2 para a floresta primária. Do mesmo modo, o DMF não
diferiu entre a floresta em regeneração e a floresta primária (t = 0,080; gl = 174; p =
0,93 Figura 2). A média do DMF para a floresta em regeneração e para a floresta
primária foi de 0,121 g e 0,122 g, respectivamente. A forma foliar também não
apresentou diferença entre as duas florestas (t = 0,29; gl = 183; p = 0,77 Figura 3), com
média de 2,92 para as folhas da floresta em regeneração e 2,89 para folhas da floresta
primária. Isso indica que as folhas são em geral mais alongadas, pois o comprimento foi

182
em média três vezes maior do que a largura. Para o teste de homogeneidade das
variâncias também não houve diferenças para as características área foliar (F = 0,01; p =
0,92), DMFmédio (F = 0,80; p = 0,37) e forma foliar (F = 0,90; p = 0,34) entre a floresta
em regeneração e a floresta primária.
Figura 1. Área foliar média de indivíduos arbóreos juvenis de uma floresta em
regeneração e uma floresta primária localizadas na Amazônia Central, Brasil. As barras
verticais representam o desvio padrão da área foliar.
Floresta primária Floresta em regeneração
Tipo de floresta
-20
0
20
40
60
80
100
120
140
160
180
Áre
a fo
liar
(cm
2)
183
Figura 2. DMF médio (diferença entre a massa foliar túrgida e a massa seca) dos
indivíduos arbóreos juvenis de uma floresta em regeneração e uma floresta primária
localizadas na Amazônia Central, Brasil. As barras verticais representam o desvio
padrão do DMF.
Figura 3. Forma foliar média calculada através da razão comprimento médio / largura média
para os indivíduos arbóreos juvenis em uma floresta em regeneração e uma floresta
primária na Amazônia Central, Brasil. As barras verticais representam o desvio padrão.
Floresta primária Floresta em regeneração
Tipo de floresta
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
DM
F (
g)
Floresta em regeneração Floresta primária
Tipo de floresta
1,8
2,0
2,2
2,4
2,6
2,8
3,0
3,2
3,4
3,6
3,8
Form
a f
oli
ar
(cm
)

184
Discussão
Os resultados indicam que não existem diferenças relacionadas às características
funcionais área foliar, DMF e forma foliar entre a floresta em regeneração e a floresta
primária. Desse modo, é possível inferir que os indivíduos recrutados nas duas florestas
apresentam estratégias funcionais semelhantes relacionadas à captação de luz e
armazenamento de água nas folhas. Mesmo em diferentes condições de disponibilidade
de água e incidência de luz, que são recursos essenciais às plantas, não houve
divergência nessas características funcionais foliares entre as duas florestas.
O crescimento e a reprodução das plantas dependem essencialmente da
disponibilidade de água e de luz. No entanto, a transpiração é um processo inevitável,
pois a absorção do CO2 pelos estômatos para a fotossíntese permite também a perda de
água (Pimentel 2004, Tyree 2007). Para regular o balanço entre taxa fotossintética e
perda de água, mecanismos fisiológicos que atuam nos estômatos garantem a eficiência
no uso da água. Mecanismos como a abertura e fechamento dos estômatos podem atuar
de maneira diferencial nas plantas dependendo das condições ambientais. Esse
mecanismo funciona como um ajuste para diminuir a perda de água das folhas, mesmo
com um aumento na área foliar (Tyree 2007). O controle da abertura estomática garante
que a folha alcance uma taxa fotossintética suficiente para sua demanda, evitando a
perda excessiva de água (Taiz & Zeiger 2002, Tyree 2007). Essa regulação pode manter
os estômatos abertos durante um período mais curto do dia em ambientes com
temperaturas elevadas, como nas florestas em regeneração (Taiz & Zeiger 2002). Essa
capacidade de ajuste explica o fato de que as plantas da floresta em regeneração são
capazes de manter folhas com área, forma e capacidade de armazenamento de água
semelhante às folhas encontradas na floresta primária.

185
Características morfológicas foliares que permitem atingir altas taxas
fotossintéticas aparentemente tem maior valor para a sobrevivência das plantas, quando
comparadas as características morfológicas relacionadas à conservação de água
(Pimentel 2004). Desse modo, a manutenção das características foliares que aumentam
a capacidade de captação de luz, como o aumento da área foliar, é favorecida mesmo
levando a uma maior perda de água. Assim, compensações fisiológicas podem levar a
uma convergência nas características funcionais morfológicas foliares entre florestas
com condições ambientais distintas.
Agradecimentos
Primeiramente gostaria de agradecer aos coordenadores Paulo Estefano D. Bobrowiec e
Paulo Enrique Cardoso Peixoto que são as pessoas responsáveis pela realização desse
curso. Incrível a dedicação e paciência de vocês com essa cambada de gonorantes.
Quando eu disse na carta de intenção que ficaria muito feliz em participar desse curso,
não imaginava que seria tanto! Sou grata pela forma com que vocês mudaram e
enriqueceram a minha percepção da ciência durante esses 30 dias. Em segundo lugar
gostaria de agradecer aos monitores Tiago Gloss e Marcos Barbudo que foram
extremamente dedicados e atenciosos em ajudar, e nunca se negaram a discutir qualquer
teoria, por mais mirabolante que ela fosse. Vocês foram fundamentais para o curso!
Agradeço também a todos os professores que enriqueceram o curso durante
todas as etapas e foram fundamentais para a realização de todos os projetos.
Agradecimentos vão também para todas as pessoas que me ajudaram na
realização do meu projeto individual. Entre elas: Paulinho Mau, que me acompanhou no
campo durante minha coleta de dados e foi o revisor do relatório, você foi fundamental
para levá-lo para o céu. A professora Ana que com toda paciência revisou meu relatório.

186
Ao Paulinho Bom que me ajudou e esclareceu minhas ideias várias vezes, e que mediu
782854649 folhas para mim, mesmo eu sendo uma racha. Ao Marcel que foi
fundamental para que eu conseguisse realizar esse projeto, me ajudando em campo, na
medição das folhas e na pesagem de 8 mil pedaços de folhas. Além de discutir comigo
todas as ideias do projeto. A minhas amigas Tati, da Silva, Carolzinha pé-de-ouro,
Bianca, e ao monitor Marcos que me ajudaram na triagem das 79821961893 folhas.
Especialmente a Tati que foi responsável por arrebanhar a maioria dessas pessoas. Aos
monitores Tiago e Marcos que me ajudaram a elaborar essa ideia. E ao Joseph que me
acompanhou e me ajudou no campo na primeira coleta!
Um agradecimento em especial vai para a Dona Eduarda e para o Seu Jorge, que
carinhosamente preparam nossas refeições!
E por fim, gostaria de agradecer imensamente a todos os amigos de cada canto
do Brasil que tive a enorme honra de conhecer. Cada um de vocês, com suas
particularidades, personalidades e sotaques me ensinaram “que o ser humano é ridículo”
(Joseph 2012). Queridos, já sinto saudades mesmo ainda estando com vocês! Não
citarei nomes em especial, porque acabaria escrevendo o de todos, sem exceção!
Obrigada por compartilharem comigo os momentos felizes, os momentos de agonia
durante os PO’s e PL’s, os momentos de alegria durante os PO’s e PL’s, os momentos
de conversas construtivas, bem como os de conversas inúteis. O suco de caju e a
goiabada, e as festas insanas! Ohhh igarafest!
Referências
Benitez-Malvido, J.1998. Impact of forest fragmentation on seedling abundance in a
tropical rainforest. Conservation Biology, 12: 380-389.

187
Casenave, J.L., J.P. Pelotto & J. Protomastro. 1995. Edge-interior differences in
vegetation structure and composition in a Chaco semi-arid forest, Argentina.
Forest Ecology and Management, 72: 61-69.
Cianciaruso, M.V., I.A. Silva & M.A. Batalha. 2009. Diversidades filogenética e
funcional: novas abordagens para a ecologia de comunidades. Biota Neotropica,
9: 93-103.
Cornelissen, J.H.C., S. Lavorel, E. Garnier, S.Díaz, N.Buchmann, D.E.Gurvich, P.B.
Reich, H. ter Steege, H.D. Morgan, M.G.A.van der Heijden, J.G. Pausas, & H.
Poorter. 2003. Handbook of protocols for standardized and easy measurement of
plant functional traits worldwide. Australian Journal of Botany, 51: 335-380.
Diamond, J.M. 1975. Assembly of species communities, pp. 332-359. In: Ecology and
evolution of communities (M.L. Cody & J.M. Diamond, eds.). Cambridge:
Harvard University Press.
Kapos V., E. Wandelli, J.L. Camargo & G. Ganade. 1997. Edge-related changes in
environment and plant responses due to forest fragmentation in central Amazonia,
pp.33-44. In: Tropical forest remnants: ecology, management, and conservation of
fragmented communities (W.F. Laurance & R.O. Bierregaard, eds.). Chicago: The
University of Chicago Press.
Laurance, W., L.V. Ferreira, J.M. Rankin-de-Merona, S.G.W. Laurance, W.R.
Hutchings, & T.E. Lovejoy. 1998. Effects of forest fragmentation on recruitment
patterns in Amazonian tree communities. Conservation Biology, 12: 460-464.
Lavorel, S., & E. Garnier. 2002. Predicting changes in community composition and
ecosystem functioning from plant traits: revisiting the Holy Grail. Functional
Ecology, 16: 545–556.

188
Mesquita, R.C.M., K. Ickes, G. Ganade, G.B. Williamson. 2001. Alternative sucessional
pathways following deforestation in the Amazon Basin. Journal of Ecology, 89:
528-537.
Pimentel, C. 2004. A relação da água com a planta. Seropédica, RJ: Edur, 191p.
Quesada, M., G.A. Sanchez-Azofeifa, M. Alvarez-Anorve, K.E. Stoner, L. Avila-
Cabadilla, J. Calvo-Alvarado, A. Castillo, M.M. Espirito-Santo, M. Fagundes,
G.W. Fernandes, J. Gamon, M. Lopezaraiza-Mikel, D. Lawrence, L.P.C.
Morellato, J.S. Powers, F.D. Neves, V. Rosas- Guerrero & R. Sayago, G.
Sanchez-Montoya. 2009. Succession and management of tropical dry forests in
the Americas: review and new perspectives. Forest Ecology and Management,
258: 1014–1024.
Tyree, M.T. 2007. Water relation and hydraulic architecture, pp.175-212. In: Functional
plant ecology (F.I. Pugnaire & F. Valladares, eds). Boca Raton: CRC Press.
Taiz, L. & E. Zeiger. 2002. Fisiologia vegetal. Porto Alegre: Artmed.l
Carlos Pimentel
UFRURALRJJA Relação da
Planta

189
Distribuição de galhas em folhas de Vismia japurensis (Clusiaceae): existe uma
seleção do local de oviposição?
Artur Alcantara Madeira
Introdução
A seleção de habitat consiste na escolha ativa dos organismos por características
específicas do ambiente (Morris 1992). Essa seleção geralmente tem como objetivo
minimizar efeitos negativos ou maximizar os efeitos positivos para os indivíduos
(Morin 2011). Os efeitos negativos podem ser, por exemplo, referentes ao risco de
predação e à competição, e os efeitos positivos, podem ser relacionados à alimentação e
reprodução (Sih 1980, Morin 2011). Dessa forma, a escolha do habitat tem papel
importante para moldar a comunidade, atuando como um dos principais mecanismos
que permitem às espécies coexistirem (Rosenzweig 1981). Mesmo assim, para qualquer
habitat selecionado, o organismo necessariamente irá interagir com outros organismos
daquele local, seja com interações positivas, negativas ou neutras.
Estudos sobre interações inseto-planta consideram que a riqueza de espécies e a
abundância de indivíduos em comunidades de insetos herbívoros é influenciada pelas
características de suas plantas hospedeiras, como área de distribuição e complexidade
estrutural. A qualidade, a quantidade e a disponibilidade de recursos oferecidos pela
planta hospedeira também exercem papel fundamental sobre a diversidade de insetos
herbívoros (Strong et al. 1984). Por exemplo, os vasos condutores de uma planta
transportam, além de água e outras substâncias, a seiva. Então organismos que
necessitam dela como alimento podem selecionar locais da planta onde a exploração
desses recursos seja mais fácil. Tecidos de maior valor nutricional aumentam o
desempenho da prole desses insetos, pois permitem que as larvas se desenvolvam com

190
mais eficiência e rapidez. Dessa forma, a seleção natural deve agir sobre o processo de
oviposição de insetos herbívoros, como os insetos galhadores, para otimizar a
performance da prole (Fritz et al. 2003).
Quando os insetos galhadores ovipõem nas plantas, eles induzem a formação de
galhas. As galhas são estruturas compostas de tecidos vegetais dentro dos quais a larva
se alimenta. Galhas se distinguem de outros abrigos criados por insetos, como folhas
enroladas, por causarem uma diferenciação e crescimento de tecidos da própria planta.
Além disso, provocam um direcionamento de nutrientes da planta para si, e por isso
contêm tecidos que são mais nutritivos do que tecidos sem galhas (Stone & Schonrogge
2003, Motta et al. 2005).
Em Vismia japurensis mais de uma galha pode ocorrer em uma mesma folha, a
partir desse fato, verifiquei se: (i) existe uma preferência de oviposição nas regiões de
nervuras das folhas ao invés das regiões entre nervuras, (ii) se as galhas foliares que se
encontram nas regiões de nervuras são maiores do que galhas encontradas nas regiões
entre nervuras, e (iii) se as galhas que se encontram nas partes mais distais das nervuras
secundárias, ocorrem apenas se houver uma outra galha na região mais basal da mesma
nervura. Minhas hipóteses foram que: (i) os indivíduos parentais devem selecionar
locais melhores para ovipor, de forma que existirá um maior número de galhas nas
regiões de nervuras do que nas regiões entre nervuras, (ii) as galhas das regiões de
nervuras serão maiores por estarem mais próximas da fonte de recurso, e (iii) se houver
galhas nas regiões mais distais de uma nervura, esta estará mais distante da nervura
central, e portanto mais distante dos recursos, por haver uma ou mais galhas nas regiões
mais basais da mesma nervura.

191
Métodos
Realizei o estudo na Reserva do Km 41 (02° 24’S e 59° 44’O), 80 km ao norte de
Manaus, Amazonas, Brasil. A reserva é formada por floresta de terra firme e faz parte
da Área de Relevante Interesse Ecológico (ARIE), administrada pelo Projeto Dinâmica
Biológica de Fragmentos Florestais (PDBFF) e pelo Instituto Chico Mendes de
Conservação da Biodiversidade (ICMBio). Na borda da floresta coletei, por busca ativa,
todas as folhas de indivíduos de Vismia japurensis (Clusiaceae) que encontrei com a
presença de galhas da mesma espécie, obtendo no total 32 folhas.
Após a coleta, levei as folhas para o laboratório, onde medi o comprimento e a
largura de todas as galhas cujas larvas já haviam eclodido. Essas medidas foram
utilizadas para calcular a área de cada galha com a fórmula de área de uma elipse
(Comprimento x Largura x π). Em seguida, classifiquei as galhas de acordo com sua
posição na folha: região de intersecção de nervuras, região de nervura e região entre
nervuras.
Para avaliar se existe uma preferência pelas regiões de nervura, comparei a
frequência de galhas encontradas nessas regiões com a frequência esperada se essa
distribuição fosse aleatória. Para saber a frequência esperada, determinei a porcentagem
da área foliar ocupada por nervuras utilizando o programa ImageTools (versão 3.0,
UTHSCSA, 2002). Obtive a porcentagem média da área foliar ocupada por nervuras, a
partir de cinco folhas escolhidas aleatoriamente da amostra.
Utilizei um teste qui-quadrado para avaliar se existe uma seleção pelas regiões
de nervura ao invés das regiões entre nervuras. Para relacionar a área da galha com sua
posição na folha utilizei uma ANOVA, seguida de um teste – t (posteriori) para
entender como as categorias avaliadas se relacionavam. E para analisar se as galhas de

192
regiões mais distais das nervuras secundárias ocorriam sozinhas ou associadas a outra
galha na mesma nervura, utilizei novamente o teste qui-quadrado.
Resultados
Encontrei 260 galhas nas folhas de V. japurensis, sendo que destas, 20 estavam
localizadas nas regiões de intersecção de nervuras, 131 estavam localizadas nas regiões
das nervuras e 109 estavam localizadas nas regiões entre nervuras. As nervuras
corresponderam a 9% da área foliar total e foram ocupadas por 151 das 260 galhas
(58%). De acordo com os dados, haviam mais galhas presentes nas regiões de nervura
do que o esperado se a distribuição delas na folha fosse ao acaso (χ2 = 781,48; gl = 1; p
< 0,01).
As galhas das três regiões analisadas apresentaram diferenças em relação à sua
área (F(2) = 10,53; p < 0,01) (Figura 1). As galhas das regiões de intersecção de nervuras
e das regiões de nervura não diferiram em área (t = -0,50; gl = 1; p = 0,62), porém as
galhas das regiões de intersecção de nervuras e das regiões de nervura foram maiores do
que as das regiões entre nervuras em relação à área (t = 4,55; gl = 1; p <0,01).

193
Figura 1. Tamanho médio das galhas em relação à região na folha. Pontos representam a
média e as barras representam o intervalo de confiança.
Vinte e uma galhas estavam presentes na porção da nervura secundária mais
próxima à borda da folha (8,1%). Destas, 14 ocorreram sozinhas e sete estavam
acompanhadas por uma outra galha na porção mais próxima à nervura central. As galhas
das nervuras secundárias que estavam mais distantes da nervura central não estavam
associadas à presença de outras galhas na mesma nervura (χ2 = 2,33; gl = 1; p = 0,13).
Discussão
A distribuição das galhas nas folhas de V. japurensis não foi aleatória. Os indivíduos
parentais selecionaram regiões de nervura para oviposição, e as galhas dessas regiões
foram maiores do que as galhas localizadas nas regiões entre nervuras. Larvas dentro de
galhas podem se alimentar de substâncias foto-assimiladas e nutrientes. Essas
substâncias são transportadas dos sítios de produção ou armazenamento (fontes), para
os sítios de uso ativo ou crescimento da planta (dreno). Ao se acoplarem no sistema

194
vascular das plantas hospedeiras, as galhas induzem um direcionamento desses recursos
para si, criando um tipo de dreno artificial (Larson & Whitham 1997). Outras galhas se
alimentam de carboidratos e lipídios das plantas, além de drenar compostos secundários,
utilizados nas camadas externas das galhas. Estas camadas conferem uma proteção extra
contra parasitas e predadores (Motta et al. 2005). Assim, as regiões de nervura das
folhas devem possibilitar um melhor acesso às substâncias utilizadas pelas galhas,
permitindo um melhor desenvolvimento larval. Por isso, essas regiões são um local
melhor para oviposição e, portanto, são escolhidas com maior frequência (Sih 1980).
As galhas das porções distais das nervuras secundárias não estavam mais
distantes da nervura central em decorrência da presença de outras galhas na mesma
nervura. A frequência de ocorrência de galhas nas porções distais das nervuras
secundárias foi extremamente baixa. É provável que essa região esteja muito distante da
nervura central e, assim, longe dos recursos alimentares. Então, os indivíduos parentais
escolhem ovipor em lugares melhores (Morin 2011), independente da presença ou
ausência de outras galhas na mesma nervura.
Considerando os resultados desse estudo, é possível concluir que existe uma
seleção de habitat para o sistema estudado. Indivíduos parentais de insetos galhadores
apresentam uma preferência em ovipor nas regiões de nervura das folhas de Vismia
japurensis, já que estas são mais próximas aos vasos que transportam a seiva da planta.
Assim, as larvas dessas regiões se alimentam e se desenvolvem melhor, eclodindo com
um tamanho maior, o que provavelmente confere a estes indivíduos melhores chances
de sobrevivência e maior sucesso reprodutivo.

195
Agradecimentos
Agradeço aos professores Paulo Estefano Dineli Bobrowiec e Paulo Enrique Cardoso
Peixoto, aos monitores Thiago Gechel Kloss e Marcos Costa Vieira e ao colega
Randolfo Gonçalves Dias Terceiro pela ajuda e pelas sugestões no desenvolvimento
desse trabalho, bem como durante a triagem das amostras coletadas. Agradeço também
a todos os professores e colegas do curso de campo Ecologia da Floresta Amazônica de
2012 e ao “Seu Jorge” e “Dona Eduarda” pelos momentos de descontração, sem os
quais tudo teria sido muito mais difícil.
Referências
Fritz, R.S., B.A. Crabb & C.G. Hochwender. 2003. Preference and performance of a
gall-inducing sawfly: plant vigor, sex, gall traits and phenology. Oikos,
102:601-613.
Larson, K.C. & T.G. Whitham. 1997. Competition between gall aphids and natural plant
sinks: plant architecture affects resistance to galling. Oecologia, 109:575-582.
Morin, P. 2011. Community Ecology. West Sussex: Wiley-Blackwell.
Morris, D.W. 1992. The role of habitat selection in landscape ecology. Evolutionary
Ecology, 6:357-359.
Motta, L.B., J.E. Kraus, A. Salatino & M.L.F. Salatino. 2005. Distribution of
metabolites in galled and non-galled foliar tissues of Tibouchina pulchra.
Biochemical Systematics and Ecology, 33:971-981.
Rosenzweig, M.L. 1981. A Theory of Habitat Selection. Ecology, 62:327-335.
Sih, A. 1980. Optimal Behavior: Can foragers balance two confliting demands? Science,
210:1041-1043.

196
Stone, G.N & K. Schönrogge. 2003. The adaptive significance of insect gall
morphology. Trends in Ecology and Evolution, 18:512-522.
Strong, D.R., J.H. Lawton & T.R.E. Southwood. 1984. Insects on plants: community
patterns and mechanisms. Oxford: Blackwell Scientific Publications.
Wilcox, D., B. Dove, D. McDavid & D. Greer. 2002. ImageTools. UTHSCSA. Texas,
EUA.

197
Quando uma ajuda é bem-vinda: briófitas facilitam o estabelecimento de plantas
sobre troncos caídos
Bianca Ferreira da Silva
Introdução
Interações positivas entre plantas encontram-se difundidas em diversas comunidades e
interferem fortemente na dinâmica dessas comunidades (Bruno et al. 2003). A
facilitação é uma interação positiva em que a presença de determinadas plantas cria
condições ambientais que permitem que outros indivíduos se estabeleçam no local
(Callaway 1995). Esse processo ocorre quando condições ambientais impõem fortes
restrições para o estabelecimento de plantas (Maestre & Cortina 2004). Na sucessão
ecológica, a facilitação ocorre quando as plantas de um estágio sucessional precedente
geram condições para o estabelecimento das plantas do próximo estágio (Brooker 2008;
Clements 1916).
Os mecanismos através dos quais uma planta facilita o estabelecimento de outro
indivíduo podem ser indiretos ou diretos (Baumeister & Callaway 2006). Os
mecanismos indiretos incluem proteção contra herbívoros e o aumento de visitas de
polinizadores e dispersores (Acuña-Rodríguez et al. 2006; Callaway 1995). Já os
mecanismos diretos são a atenuação das condições abióticas, para espécies menos
resistentes. Essa atenuação pode estar associada, por exemplo, à diminuição da
exposição à radiação e às altas temperaturas (Holzapfel et al. 2006; Castro et al. 2004),
ao aumento da oxigenação do solo (Callaway & King 1996) e ao aumento da
quantidade de nutrientes (Gómez-Aparicio et al. 2005).
O processo de facilitação é bastante estudado em ambientes áridos e semi-áridos
(Holzapfel et al. 2006; Flores & Jurado 2003; Castro et al. 2002), onde as condições

198
ambientais são bastante restritivas. Entretanto, florestas tropicais também apresentam
limitações à germinação e ao estabelecimento de plantas, como presença de patógenos e
baixa fertilidade dos solos (Richards 1996). Nesses ambientes, briófitas geram manchas
de ocorrência ao se propagarem assexuadamente (Vanderpoorten & Goffinet 2009).
Essas manchas constituem um substrato que pode reter água e nutrientes (Bates 2009),
enquanto outros locais da floresta podem ter esses nutrientes carreados pela chuva.
Assim, essas manchas de briófitas podem ser sítios propícios para o desenvolvimento de
outras plantas. Manchas de briófitas são particularmente comuns sobre a superfície de
troncos caídos no interior da mata. Sobre um mesmo tronco ocorrem áreas com
manchas de briófitas e áreas sem a presença delas. Nesse contexto, investiguei o papel
das briófitas no estabelecimento de indivíduos de outras plantas sobre troncos caídos.
Minha hipótese é que briófitas atuam como facilitadoras para o estabelecimento desses
indivíduos. Assim, espero que outras plantas além das briófitas ocorram mais
frequentemente em áreas do tronco cobertas por briófitas do que nas áreas sem briófitas.
Métodos
Realizei o estudo em um trecho de floresta contínua na Amazônia Central, localizado na
Área de Relevante Interesse Ecológico do Km 41, pertencente ao Projeto Dinâmica
Biológica de Fragmentos Florestais (PDBFF) do Instituto Nacional de Pesquisas na
Amazônia (INPA). Ao longo de um trecho de 1400 m, procurei por troncos caídos e
parcialmente cobertos por briófitas. Em cada tronco avaliado delimitei duas parcelas
paralelas de 15 cm², sendo uma no centro de uma mancha de briófitas e outra, na área
sem briófitas mais próxima ao lado da mancha. Para cada área delimitada verifiquei a
presença de outras plantas. Considerei todos os indivíduos de plantas presentes na
parcela cujo ponto de enraizamento se encontrasse dentro de um mesmo tratamento –

199
com briófitas ou sem briófitas – na superfície do tronco. A fim de analisar o efeito da
presença de briófitas sobre a ocorrência de outras plantas, realizei um teste de qui-
quadrado de McNemar.
Resultados
Amostrei um total de 80 troncos. As plantas encontradas eram plântulas, juvenis e
alguns adultos de lenhosas, trepadeiras e epífitas. A média do número de espécies nas
parcelas com briófitas foi 0,99 ± 0,86, podendo ocorrer até três espécies diferentes. A
média do número de indivíduos em parcelas com briófitas foi 1,19 ± 1,17, variando de
zero a cinco indivíduos de plantas em cada parcela. Em parcelas sem briófitas, a média
do número de espécies de plantas foi 0,33 ± 0,61, variando de zero a duas espécies
distintas, e a média do número de indivíduos por parcelas foi 0,36 ± 0,72, com, no
máximo, três indivíduos.
Dos 80 pares de parcelas, 55 tinham a presença de outras plantas na parcela com
briófitas. Desses 55 pares, apenas 11 tinham plantas na parcela sem briófitas (Figura 1).
Dos 25 pares em que não ocorreram outras plantas na parcela com briófitas, 16
possuíam plantas na parcela sem briófitas (Figura 1). A presença das briófitas afetou
positivamente a presença de outras plantas (χ2=21,81; gl=1; p<0,01).

200
Figura 3. Proporção de troncos encontrados para cada uma das combinações possíveis
de resultados. Cada tronco contém uma parcela com briófitas e uma parcela sem
briófitas. Cada parcela pode ou não conter outras espécies de plantas. “1” indica a
presença e “0” indica a ausência de outras espécies de plantas em cada parcela. O
asterisco indica a combinação que seria mais frequente caso as briófitas facilitassem o
estabelecimento de outras plantas.
Discussão
A presença de briófitas favorece a ocorrência de plantas de outras espécies sobre a
superfície de troncos. Essa facilitação pode ocorrer devido à criação de condições
ambientais favoráveis para o estabelecimento. Assim, o desenvolvimento desses
organismos parece depender das alterações ambientais provocadas pela presença das
briófitas, como microclima mais úmido, substrato para fixação das plântulas e retenção
e acúmulo de nutrientes. A atenuação das condições ambientais por um grupo de
espécies parece ser mesmo um fator determinante para que indivíduos de outras
espécies se desenvolvam (Baumeister & Callaway 2006).

201
A facilitação gerada por briófitas em superfícies de troncos pode estar
relacionada a mecanismos indiretos, como o estabelecimento em sítios onde a
competição é menor. No solo, a elevada abundância de plântulas e a proximidade a
adultos aumentam os níveis de competição, dado que os envolvidos retiram de uma
mesma área de solo os nutrientes para o próprio desenvolvimento e manutenção (Chen
et al. 2010; Connel 1971; Janzen 1970). A competição com indivíduos adultos, que são
competitivamente mais fortes, pode resultar em um aumento da taxa de mortalidade das
plântulas (Chen et al. 2010). Assim, a presença das briófitas sobre a superfície do tronco
pode fornecer às plântulas um sítio mais favorável ao estabelecimento, por apresentar-se
distante do solo, livrando-as da intensa competição por recursos ali existente. Após a
decomposição do tronco, esses indivíduos podem alcançar o solo com um tamanho em
que o risco de mortalidade já seja bastante reduzido.
Os troncos caídos de árvores nos quais as briófitas ocorriam possuíam
superfícies com rugosidade diferente. Reentrâncias nos troncos podem facilitar a
fixação e o desenvolvimento das plântulas, já que as reentrâncias são locais de acúmulo
de solo e serapilheira. Portanto, estudos que considerem o possível efeito sinérgico de
fatores como presença das briófitas e a rugosidade dos troncos poderiam esclarecer a
importância relativa desses fatores na facilitação da ocorrência de plantas em florestas
tropicais.
Agradecimentos
Agradeço aos organizadores do EFA pela oportunidade de participar do curso e aos
coordenadores, Paulinhos -Rainbow Master e Mauravilhoso - por todos os puxões de
orelha bem dados ao longo desse mês. Aos monitores, Barbudo e Gloss, pelas conversas
e orientações. Ao Paulinho (Rainbow Master) por todos os abraços (argh!) nos

202
momentos de desespero. Agradeço a Samara (Tati) pela companhia no campo, pelas
dicas para o trabalho e pelos abraços às “ávores”. Agradeço também ao Marcel por
escutar minhas dúvidas e discutir o trabalho comigo e pelo enorme carinho e paciência
na correção desse trabalho. Ao Marcos (monitor Barbudo chato e [não] favorito) por ser
sempre crítico, me fazendo prestar atenção nos detalhes do desenvolvimento do
trabalho. Agradeço a todos os amigos do EFA, que durante um mês aguentaram toda a
minha variação de humor e ainda me proporcionaram muitos momentos de risos e
descontração, dos quais lembrarei sempre. Obrigada à Amazônia, por me mostrar um
mundo a ser explorado e discutido em seus processos e por ter aqui descoberto essas
pessoas maravilhosas, com as quais percebo que nem tudo é trabalho e que o estresse
deve ser deixado de lado para que a vida possa ser realmente vivida.
Referências
Acuña-Rodríguez, I.S., L.A. Cavieres & E. Gianoli. 2006. Nurse effect in seedling
establishment: facilitation and tolerance to damage in the Andes of central Chile.
Revista Chilena de Historia Natural, 79:329-336.
Bates, J.W. 2009. Mineral nutrition and substratum ecology, pp. 299-356. In: Bryophyte
biology (B. Goffinet & A.J. Shaw, eds.). Cambridge: Cambridge University
Press.
Baumeister, D. & R.M. Callaway. 2006. Facilitation by Pinus flexilis during succession:
a hierarchy of mechanisms benefits other plant species. Ecology, 87:1816-1830.
Brooker, R.B., F.T. Maestre, R.M. Callaway, C.L. Lortie, L.A. Cavieres, G. Kunstler, P.
Liancourt, K. Tielbörger, J.M.J. Travis, F. Anthelme, C. Armas, L. Coll, E.
Corcket, S. Delzon, E. Forey, Z. Kikvidze, J. Olofsson, F. Pugnaire, C.L.
Quiroz, P. Saccone, K. Schiffers, M. Seifan, B. Touzard & R. Michalet. 2008.

203
Facilitation in plant communities: the past, the present, and the future. Journal of
Ecology, 96:18-34.
Bruno, J.F., Stachowicz, J. J. & Bertness, M. D. 2003. Inclusion of facilitation into
ecological theory. Trends in Ecology & Evolution, 18:119-125.
Callaway, R.M. 1995. Positive interactions among plants. Botanical Review, 61:306-
349.
Callaway, R.M. & L. King. 1996. Oxygenation of the soil rhizosphere by Typha
latifolia and its facilitative effects on other species. Ecology, 77:1189-1195.
Castro, J., R. Zamora, J.A. Hódar & J.M. Gómez. 2002. Use of Shrubs as Nurse Plants:
A New Technique for Reforestation in Mediterranean Mountains. Restoration
Ecology, 10:297-305.
Castro, J., R. Zamora, J.A. Hódar, J.M. Gómez & L. Gómez-Aparicio. 2004. Benefits of
Using Shrubs as Nurse Plants for Reforestation in Mediterranean Mountains: A
4-Year Study. Restoration Ecology, 12:352-358.
Chen, L., X. Mi, L.S. Comita, L. Zhang, H. Ren & K. Ma. 2010. Community-level
consequences of density dependence and habitat association in a subtropical
broad-leaved forest. Ecology Letters, 13:695-704.
Clements, F.E. 1916. Plant succession: an analysis of the development of vegetation.
Washington: Carnegie Institution of Washington.
Connell, J.H. 1971. On the role of natural enemies in preventing competitive exclusion
in some marine animals and in rain forest trees, pp. 298-312. In: Dynamics of
Populations (P.J. Den Boer & G. Gradwell, eds). Wageningen: Centre for
Agricultural Publishing and Documentation.
Flores, J. & E. Jurado. 2003. Are nurse-protégé interactions more common among
plants from arid environments? Journal of Vegetation Science, 14:911-916.

204
Gómez-Aparicio, L., F. Valladares, R. Zamora & J.L. Quero. 2005. Response of tree
seedlings to the abiotic heterogeneity generated by nurse shrubs: an
experimental approach at different scales. Ecography, 28:757-768.
Holzapfel, C., K. Tielbörger, H.A. Parag, J. Kigel & M. Sternberg. 2006. Annual plant–
shrub interactions along an aridity gradient. Basic and Applied Ecology, 7:268-
279.
Janzen, D.H. 1970. Herbivores and number of tree species in tropical forests. American
Naturalist, 104:501-528.
Maestre, F.T. & J. Cortina. 2004. Do positive interactions increase with abiotic stress?
A test from a semi-arid steppe. Proceedings of the Royal Society London B
(Suppl.) 271:S331–S333.
Richards, P.W. 1996. The tropical rain forest: an ecology study. Cambridge: Cambridge
University Press.
Vanderpoorten, A. & B. Goffinet. 2009. Introduction to Bryophytes. Cambridge:
Cambridge University Press.

205
As formigas não utilizam sinal químico de formigas de outros territórios para
melhorar sua eficiência de forrageio
Bruno Travassos
Introdução
Formigas são organismos que se orientam basicamente por sinais químicos (Wilson
1958). As formigas não usam somente os sinais químicos do ambiente para se orientar,
como também produzem seus próprios sinalizadores que as ajudam a coordenar funções
no formigueiro (Wilson 1962). Os sinalizadores químicos das formigas estão envolvidos
em quase todas as funções vitais do formigueiro. Por exemplo, no forrageio por patrulha
e recrutamento das formigas de ninho fixo, algumas formigas têm a função de patrulhar
determinada área em busca de alimento. Quando essas formigas encontram algum tipo
de alimento, elas marcam o recurso com um sinalizador químico e voltam para o
formigueiro. Essa trilha química é utilizada pelo restante das formigas para encontrar
esse alimento mais rapidamente (Burkhardt 1983).
Algumas formigas são capazes de reconhecer o sinal químico de formigas de
outro formigueiro e até de outra espécie (Mintzer 1980). Isso já foi demonstrado tanto
para formigas parasitas sociais que seguem a trilha de suas hospedeiras, quanto em
formigas escravas que seguem a trilha das formigas que as dominam (Wilson 1955). Em
outros casos, formigas não parasitas já foram reportadas utilizando trilhas de outras
espécies (Hölldobler e Wilson 1990). Hölldobler e Wilson (1990) argumentam que
quando não existe parasitismo, o comportamento de seguir a trilha de outras formigas
talvez não tenha significado adaptativo. Entretanto, para formigas que usam recursos
semelhantes, utilizar um sinal químico de outras formigas para acessar esses recursos
pode ser muito vantajoso, já que isso pouparia tempo de patrulha.

206
No forrageio por patrulha e recrutamento, o maior investimento de energia é na
patrulha (Stephen & Krebs 1986). Patrulhar implica em gasto de energia para o
deslocamento além da exposição à predação e a intempéries ambientais. Portanto,
qualquer mecanismo que aumente a eficiência de patrulha representa uma economia
significativa de energia e, portanto, fornece uma vantagem grande para o formigueiro
(Lighton et al. 1987).
O objetivo deste estudo foi avaliar se formigas utilizam o sinal químico de
outras formigas para melhorar a eficiência do forrageio. Minha expectativa é que as
formigas de um território encontrem mais rapidamente alimentos que já tenham sido
marcados quimicamente por formigas de outros territórios.
Métodos
Área de estudo
Eu realizei o estudo em uma floresta contínua de terra firme localizada na reserva
florestal do km 41 (02° 24’S – 59° 44’O), cogerida pelo Projeto Dinâmica Biológica de
Fragmentos Florestais (PDBFF) e pelo Instituto Chico Mendes de Conservação da
Biodiversidade (ICMBio). Essa área fica localizada a cerca de 80 km a norte de
Manaus, Amazonas, Brasil.
Preparação das iscas
Eu utilizei iscas ricas em proteína e gordura que consistiam de pequenos pedaços de
papel (2 x 2 cm) embebidos em óleo de sardinha, sobre os quais depositei pequenos
pedaços de sardinha. As iscas foram divididas em dois tipos: iscas sem sinal químico de
formigas e iscas com sinal químico de formigas. Para obter as iscas com sinal químico
de formigas eu preparei iscas conforme descrito acima e deixei essas iscas sobre o

207
folhiço. As iscas foram visitadas por formigas por pelo menos cinco minutos para
garantir que houvesse depósito de sinalizadores químicos (Agosti & Agosti 2002).
Depois desse tempo eu removi todas as formigas que estavam na isca e imediatamente
levei a isca para outro território.
Delineamento amostral
Eu coloquei 36 iscas (18 com sinal químico de formigas, 18 sem sinal) intercaladas por
tipo e espaçadas pelo menos 10 m ao longo de uma trilha na floresta. O espaçamento de
10 m de distância entre uma isca e outra garantiu que as formigas atraídas por essas
iscas fossem de formigueiros diferentes (Alonso & Agosti 2002) e portanto pudessem
ser consideradas como unidades amostrais independente.
Para avaliar o tempo que as formigas levaram para acessar a isca, eu registrei o
tempo entre a colocação da isca no folhiço até o momento em que cinco formigas
dominantes da mesma espécie haviam sido recrutadas. Considerei recrutamento quando
uma formiga de determinada espécie encontrava a isca, saía do local e algum tempo
depois mais formigas da mesma espécie chegavam à isca. Eu comparei o tempo que as
formigas levaram para encontrar essas iscas previamente marcadas com sinal químico
de outra formiga com iscas controle sem sinal químico.
Análise dos dados
Para testar a previsão de que iscas previamente marcadas com sinal químico de
formigas são detectadas mais rapidamente por formigas de outros territórios eu utilizei
uma análise de variância (ANOVA). O tempo necessário para recrutamento foi a
variável resposta e o tipo de isca (com sinal químico ou sem sinal químico) foi a

208
variável preditora. Eu desconsiderei unidades amostrais nas quais não houve
recrutamento por nenhuma espécie em até 20 minutos.
Resultados
Dentre as 36 iscas colocadas, não houve recrutamento em sete iscas (19%) (cinco em
iscas sem sinal químico das formigas, duas em iscas com sinal químico). A média
(±desvio padrão) do tempo de recrutamento de formigas para as iscas sem sinal químico
foi de 8,27±3,05 min e a média do tempo de recrutamento para as iscas com sinal
químico foi de 6,07±3,2 min. Não houve diferença entre os dois tipos de isca quanto ao
tempo de recrutamento de formigas (F1,28 = 3,51; p = 0,07; Figura 1).
Figura 1. Tempo de recrutamento de formigas em iscas sem sinal químico e com sinal
químico de outras formigas amostradas na reserva florestal do km 41, Manaus –
Amazonas, Brasil.
Isca sem sinal químico
de outras formigas
Isca com sinal químico de
outras formigas

209
Discussão
A presença do sinal químico de outras formigas no alimento não aumenta a eficiência
do forrageamento de formigas. Isso indica que as formigas não utilizam a pista química
de outras formigas para encontrar alimento dentro do seu território. É possível que para
as formigas encontrem um determinado alimento mais rapidamente, elas dependam não
só da sinalização química do próprio alimento, mas também da existência de uma trilha
química que as leve até ele. Sem uma trilha química até o alimento, a operária depende
da dispersão de voláteis químicos no ar para encontrar o alimento (Stephen & Krebs,
1986). Nessa situação, se o alimento também dispersa voláteis químicos no ar, a
operária poderia encontrá-lo com a mesma rapidez independente de ele estar marcado
com sinal químico de outra formiga.
Outra possível explicação para o padrão encontrado pode estar associada à
especialização no comportamento das castas das formigas. Em muitas espécies de
formiga que usam a estratégia de patrulha e recrutamento para forragear, as operárias
forrageiras são altamente especializadas em buscar alimento. Para alguns grupos existe
um nível de especialização até para tipos diferentes de alimento, por exemplo, algumas
operárias somente conseguem detectar alimentos ricos em proteína e outras somente
alimentos ricos em açúcar (Oster & Wilson 1978). Essa especialização é um mecanismo
que torna a busca por alimento muito mais eficiente (Seeley 1985). É possível que o
reconhecimento do sinal químico de outras espécies não represente uma maior
eficiência no forrageio e, portanto, esse comportamento não teria sido selecionado. Essa
explicação é condizente com o argumento de Hölldobler e Wilson (1990) no qual eles
afirmam que o reconhecimento do sinal químico de formigas de outra espécie pode não
ter significado adaptativo.

210
Agradecimentos
Agradeço aos brows Tiko Jordão e André Mendonça por terem me incentivado
(praticamente obrigado) a me inscrever no EFA. A minha namorada Camila
Hohlenwerger, por ter me ajudado com toda logística e organização da viagem, por ter
apoiado minha vinda incondicionalmente independentemente dos boatos a respeito do
EFA e por ter cedido um lugar quentinho para minha mente se reconfortar nos parcos
momentos de descanso. A todos os amigos que fiz durante o curso, em especial (em
ordem de aparecimento) a Leonardo por ter me aconselhado com palavras sábias e
convincentes sobre qual deveria ser meu real aprendizado no curso; a Randolpho que a
despeito das piadas de 8ª série sempre esteve lá pra dizer o quanto ele me acha legal; a
Lorena por ter tomado conta de todos nós e cujo carinho pôde ser percebido até por um
cara chato como eu; ao Conde José Hidasi por ter rido de todas as piadas de nerds. A
todos os professores que souberam lidar com o gênio desse “aluno feral”, ensinando-lhe
a aceitar a possibilidade de erro e, portanto, aprender muito mais.
Referências
Agosti, D., J.D. Majer, L.E. Tennant de Alonso & E. Schultz. 2002. Measuring and
monitoring biological diversity: standard methods for ants. Washington D.C.:
Smithsonian Institution Press.
Burkhardt, J.F. 1983. Foraging strategies in ants: the influence of colony size on the
organization of an ant colony. Tese de mestrado, Duke University.
Davies, N.B. & A.I. Houston. 1984. Territory economics, pp. 148-169. In: Behavioural
ecology: an evolutionary approach, 2nd
ed (J.R. Krebs & N.B. Davies, eds.).
Sunderland: Sinauer associates.

211
Hölldobler, B. & E. O. Wilson. 1990. The ants. Berlin: Springer-Verlag Berlin
Heidelberg.
Lighton, J.R.B., G.A. Bartholomew & D.H. Feener. 1987. Energetics of locomotion and
load carriage and a model of the energy cost foraging in the leaf-cutting ant Atta
colombica Guér. Physiological Zoölogy, 60:524-537.
Mintzer, A. 1980. Simultaneous use of foraging trail by two leafcutter ant species in
Sonoran desert. Journal of the New York Entomological Society, 88:102-105.
Oster, G.F. & E.O. Wilson. 1978. Caste and ecology of the social insects. New Jersey:
Princeton University Press.
Seeley, T.D. 1985. Honeybee ecology: a study of adaptation in social life. New Jersey:
Princeton University Press.
Stephens, D.W. & J.R. Krebs. 1986. Foraging theory. New Jersey: Princeton University
Press.
Wehner, R. 1987. Spatial organization of foraging behaviour in individually searching
desert ants, Cataglyphis (Sahara Desert) and Ocymyrmex (Namib Desert), pp. 15-
42. In: From individual to collective behaviour in social insects (Experimentia
Suplement, vol. 54) (Pasteels, J.M. & J.L. Deneubourgs, eds.). Bassel: Birkhäuser
Verlag.
Wilson, E.O. 1955. Division of labor in a nest of slave-making ant Fomica wheeleri
Creighton. Psyche, 60:130-133.
Wilson, E.O. 1958. A chemical releaser of alarm and dig behaviour in the ant
Pogonomyrmex badius (Latreille). Psyche, 65:41-51.
Wilson, E.O. 1962. Chemical communication among workers of the fire ant Solenopsis
saevissima (Fr. Smith), 1: The organization of mass-foraging; 2: An information

212
analysis of the odour trail; 3: The experimental induction of social responses.
Animal Behaviour, 10:134-164.

213
Agregação de palmeiras não aumenta o consumo por herbívoros
em plântulas de Attalea sp.
Caroline Marques Dracxler
Introdução
A herbivoria exerce um papel potencialmente regulador das comunidades vegetais
(MacDougall 2008). A magnitude dos efeitos dos herbívoros sobre as plantas pode
variar de acordo com as partes consumidas, com a fase de desenvolvimento da planta e
com as espécies de herbívoros envolvidos (Harper 1977). A herbivoria em folhas, por
exemplo, pode reduzir a capacidade fotossintética da planta, levando a alterações na
aptidão e a possíveis consequências demográficas para as plantas (García & Ehrlén
2009). Além disso, o porte dos herbívoros pode determinar o impacto sobre a planta,
que pode variar desde pequenos danos às folhas até o consumo do indivíduo inteiro.
As plantas representam um recurso altamente diverso e variável no espaço para
os herbívoros (Marques et al. 2000, Pimentel 1961). Os padrões de abundância e
distribuição espacial das plantas podem influenciar a herbivoria, e seus efeitos devem
variar de acordo com o grau de especialização dos herbívoros. Os efeitos negativos da
herbivoria sobre as plantas podem ser maiores em estádios ontogenéticos mais jovens, o
que pode reduzir o estabelecimento de novos indivíduos e limitar o crescimento
populacional (Harper 1977, Medina 2007). A hipótese de concentração de recursos
postula que herbívoros espécie-específicos são mais propensos a encontrar, permanecer
e consumir a planta hospedeira localizada em manchas de indivíduos coespecíficos do
que em indivíduos isolados (Root 1973). De fato, diversos estudos demonstram que
plantas hospedeiras altamente adensadas sofrem maiores níveis de herbivoria por

214
animais especialistas, tornando-se mais suscetíveis (Frederickson & Gordon 2007,
Pimentel 1961, Tahvanainen & Root 1972).
Embora a especialização de herbívoros seja um tema central em interações
herbívoros-plantas, estudos recentes mostram que herbívoros são raramente monófagos
e apresentam dietas que incluem plantas filogeneticamente próximas, as quais dividem
características nutricionais e pistas de alimentação similares (Novotny et al. 2002,
Novotny & Basset 2005, Ødegaard et al. 2005, Weiblen et al. 2006). Nesse sentido, a
chance de um mesmo herbívoro consumir duas espécies diferentes de plantas aumenta
com a proximidade filogenética entre elas (Novotny & Basset 2005). Portanto, assim
como previsto pela hipótese de agregação de coespecíficos, a agregação de plantas do
mesmo gênero ou da mesma família pode aumentar a chance de uma planta ser
consumida.
Palmeiras (Arecaceae) são um grupo monofilético abundante que representam
um recurso importante para herbívoros. Em florestas tropicais, as palmeiras estão entre
as plantas dominantes e são um importante componente estrutural das florestas (Scariot
1999). Como diversas espécies de palmeiras co-ocorrem, essas plantas podem ser
particularmente suscetíveis ao efeito da agregação de indivíduos aparentados sobre a
intensidade de herbivoria. Minha hipótese é que a intensidade de herbivoria não é
influenciada exclusivamente pela densidade de indivíduos coespecíficos, e deve
aumentar também com a concentração de palmeiras de outras espécies. Assim, ao
considerar plântulas de Attalea sp., espero que quanto maior o número de vizinhos da
mesma família, tanto coespecíficos quanto interespecíficos, maior será a área total
consumida por herbívoros.

215
Métodos
Realizei o estudo em uma floresta de terra firme na reserva do Km 41 (2°24’ S–59°43’
O), localizada a 80 km ao norte de Manaus, Brasil. A reserva faz parte da Área de
Relevante Interesse Ecológico, cogerida pelo Projeto Dinâmica Biológica de
Fragmentos Florestais (PDBFF) e abrange cerca de 10.000 ha de mata contínua.
Para avaliar a influência da agregação de indivíduos da mesma espécie e de
indivíduos filogeneticamente próximos sobre a intensidade de herbivoria, selecionei 25
plântulas focais de Attalea sp. com até 1 m de altura e com folíolos ainda unidos. A fim
de contabilizar as plântulas de Attalea sp. e de outras espécies de palmeiras localizadas
no entorno dos indivíduos selecionados, estabeleci um raio de 3 m a partir de cada
indivíduo focal, totalizando uma área de 28,26 m². Dentro da circunferência, busquei
por plântulas e juvenis da família Arecaceae e registrei a quantidade de indivíduos de
Attalea sp. (espécie focal) e das outras espécies de palmeiras.
Calculei a intensidade de herbivoria nas plântulas focais de Attalea sp.
registrando a área consumida em cada folha da plântula com o auxílio de um plástico
milimetrado. Somei a área consumida de todas as folhas para obter a área total
consumida por plântula. Identifiquei a herbivoria a partir de marcas características
deixadas pelos herbívoros, desde pequenos furos até a remoção parcial das folhas. Para
controlar os efeitos do tamanho das folhas sobre a área foliar consumida, medi o
comprimento e largura de todas as folhas de cada plântula. A partir dessas medidas,
calculei a área (área da folha=π*comprimento*largura/4) de cada folha, obtendo assim a
área foliar total de cada plântula. Para avaliar a influência do número de indivíduos de
Attalea sp., de outras espécies de palmeiras e da área foliar total das plântulas sobre a
intensidade de herbivoria na plântula focal de Attalea sp., realizei uma regressão
múltipla.

216
Resultados
A área foliar média consumida das plântulas focais foi 99,97 ± 104,75 cm² (média ±
desvio padrão), o que correspondeu a 7% da área foliar média das plântulas, a qual foi
de 1504,52 ± 1552,27 cm². O número médio de plântulas e juvenis coespecíficos
vizinhos ao indivíduo focal de Attalea sp. foi de 2,44 ± 2,08 e o de outras espécies da
família Arecaceae foi de 19,04 ± 4,92. A área foliar consumida por plântula foi
positivamente relacionada apenas com a área foliar total das plântulas de Attalea sp.
(F(1,22)=83,66; p<0,001; R²=0,58), mas não foi afetada pelo número de vizinhos
coespecíficos (F(1,22) =1,20; p = 0,28; R²=0,04; Figura 1) nem pelo número de vizinhos
de outras espécies de palmeiras (F(1,22) = 0,11; p = 0,74; R²=0,005; Figura 2).
Figura 1. Relação entre a área total consumida por herbívoros de Attalea sp. e o número
de vizinhos coespecíficos em uma floresta de terra firme na Amazônia Central, Brasil.
A área total consumida por herbívoros é representada pelos resíduos da regressão
múltipla entre a área total consumida por herbívoros, o número de vizinhos
interespecíficos e a área foliar total de plântulas de Attalea sp..
-2 0 2 4 6 8 10
Número de vizinhos coespecíficos
-100
-80
-60
-40
-20
0
20
40
60
80
100
Res
ídu
os
de
um
a re
gre
ssão
mú
ltip
la e
ntr
e a
área
to
tal
con
sum
ida
po
r h
erb
ívo
ros,
o n
úm
ero
de
viz
inh
os
inte
resp
ecíf
ico
s e
a ár
ea f
oli
ar t
ota
l d
e p
lân
tula
s d
e
Att
ale
a s
p.

217
Figura 2. Relação entre a área total consumida por herbívoros de Attalea sp. e o número
de vizinhos de outras espécies de palmeiras (Arecaceae) em uma floresta de terra firme
na Amazônia Central, Brasil. A área total consumida por herbívoros é representada
pelos resíduos da regressão entre a área consumida por herbívoros, o número de
vizinhos coespecíficos e a área foliar total de plântulas de Attalea sp..
Discussão
A intensidade de herbivoria em plântulas de Attalea sp. não foi dependente da densidade
de indivíduos de palmeiras. Por outro lado, a área foliar representa um recurso
disponível para os herbívoros e quanto maior a folha, maior foi a intensidade de
herbivoria. Embora todas as plântulas focais apresentassem sinais de herbivoria, a área
consumida por herbívoros foi relativamente baixa, mantendo-se em torno de 7% da área
foliar.
A abundância dos indivíduos de Attalea sp. não afetou a herbivoria sobre as
plântulas focais. Como a densidade de Attalea sp. foi baixa (0,08 palmeiras/m²) em
relação à densidade total de palmeiras no sub-bosque (0,8 palmeiras/m²), é possível que
-2 0 2 4 6 8 10
Número de vizinhos coespecíficos
-100
-80
-60
-40
-20
0
20
40
60
80
100
Res
ídu
os
de
um
a re
gre
ssão
mú
ltip
la e
ntr
e a
área
to
tal
con
sum
ida
po
r h
erb
ívo
ros,
o n
úm
ero
de
viz
inh
os
inte
resp
ecíf
ico
s e
a ár
ea f
oli
ar t
ota
l d
e p
lân
tula
s d
e
Att
ale
a s
p.

218
indivíduos de Attalea sp. não formem manchas de coespecíficos suficientemente
grandes para atrair mais herbívoros. Tendo em vista que a maioria dos herbívoros é
considerada generalista (Novotny et al. 2002, Novotny & Basset 2005, Ødegaard et al.
2005, Weiblen et al. 2006), eles não devem responder à densidade de uma única
espécie, o que ajuda a explicar a ausência de resposta ao adensamento da Attalea sp..
A presença de indivíduos de outras espécies de palmeiras também não
influenciou a intensidade de herbivoria em plântulas de Attalea sp. A aglomeração do
recurso está ligada à capacidade dos herbívoros de se deslocar e de encontrar o recurso
(Baraza et al. 2007). Alguns estudos demonstram que insetos herbívoros, como
borboletas, têm elevada capacidade de voo e podem alcançar até 200 m de distância
(Hanski et al. 2006, Merckx & Dyck 2007). Assim, é possível que a elevada densidade
de palmeiras no sub-bosque (0,7 palmeiras/m²) não represente agrupamentos isolados,
especialmente para herbívoros com boa capacidade de deslocamento. Como plântulas e
juvenis de outras espécies de palmeiras foram muito frequentes, os herbívoros têm igual
acesso a diferentes espécies de palmeiras hospedeiras por todo o sub-bosque. Isso dilui
possíveis efeitos da aglomeração de indivíduos aparentados sobre a herbivoria em
Attalea sp. De fato, a baixa herbivoria indicada pela proporção de área foliar consumida
das plântulas, fortalece a ideia que a herbivoria é diluída entre os diferentes indivíduos
de palmeiras. Assim, é possível que padrões diferentes de herbivoria sejam observados
somente para plantas raras e associadas a herbívoros especialistas, os quais devem
responder fortemente à aglomeração de recurso.
Agradecimentos
“O leito dos rios fartou-se e inundou de água doce a amargura do mar. Numa enchente
amazônica, numa explosão atlântica!” (MC Chicão). Quero agradecer imensamente,

219
com todo o coração, a oportunidade de participar do EFA 2012! A cada uma das
pessoinhas que eu conheci, agradeço muito! Ao Felipe, moranguinho do nordeste, e ao
Toyoyo pelos bilhetinhos das primeiras semanas e pela companhia no contrabando de
bebida nas madrugadas (Ops, falei ou pensei?). À Esther pela companhia, foi ótimo te
ver de novo! Ao Richard Gere e ao Mike por seus sonhos. Às lembranças que a festinha
sensacional da Dimonia vai deixar (beber água é a solução!). À Atoladinha (Dani) pelo
dia seguinte, que recolheu minhas coisas! Ao Jansen, à Aline e ao Cassiano por encarar
o calor com a gente na Várzea. Ao Jack Sparrow (Fabrício), companheiro de balde, ao
Dé e à Catá pela excelente companhia na fase mais bonita do curso! Sentirei falta de
vocês três! Aos promoters da festa do barco em pleno Rio Negro, nunca vou esquecer
essa festa! Ao Baby (Marcel), pela companhia, ajuda e por fazer toda e qualquer frase
ter duplo sentido! Ao Adriano, Glauco, à Ana e ao Jorge pelos aprendizados. À Manô
pela chegada no finzinho do curso e pela simpatia! Às meninas, minhas amigas tão
queridas, que me fizeram companhia. À Lore Pole Dance, pelas cervejas escondidas,
pelos banhos de balde, por tudo! Você é um doce de menina e vai fazer uma falta que dá
dó! À Tati das Áveres (somo nozes) pelas loucuras e por me ensinar que xingamentos
são amor! Ai, que saudade que eu já tô! À Aninha Alanis Maria Bethânia pela
companhia e fofocas desde o início, minha vizinha oficial de rede! Você é um amor,
sentirei muitas saudades! (sss)Sarah Baranga, você é uma calhooorda! Que será de mim
sem você, sem as risadas e os quase-choros?! Daaaani, pra bom entendedor uma letra
basta! Obrigada pela companhia, loucura, e seus quase-infartos nos Igarafests!! À
Pezinho de Ouro (Carol Brabuleta) pelas conversas e companhia! Já te conhecia e não
sabia! À Shay (Condessa de Hogwarts) por ser única! Menina, sorte do Painha ter você
por perto! Vidal querido, meu espanhol favorito, obrigada pela companhia em todas as
festinhas piratas e também nas oficiais! Ao Tatu (Vidal Pocket) por ser sempre tão feliz

220
e “cotête”, por nunca negar uma cerveja e por me ajudar no campo! Ao JosEFA pelas
caretas e por ser tão engraçado, inclusive quando fica de mau humor! A todos as outras
pessoas por tudo, de verdade, vocês são sensacionais! Aos monitores tão queridos,
Beluguinha e Marcos, por nos aturar! Marcos, por estar sempre presente nas festinhas
piratas! Beluguinha Pomerano (Thiago Xexéu), além de monitor, você foi uma fonte de
diversão esse mês! Que bom que veio e ainda trouxe a Sarinha Baranga! E como não
poderia deixar de ser, um obrigada especial aos Paulinhos, que seguraram a barra o mês
todo, sempre de bom humor e preparados para ouvir nossas besteiras, os relatórios
dignos de asco e os fiascos das apresentações. Mas vocês nunca desistiram, e sempre
muito pacientes, elogiavam todo e qualquer avanço, o que nesse mundo paralelo é um
estímulo imeeeenso! E como se não bastasse serem exemplos e pessoas absolutamente
sensacionais, vocês ainda fizeram de TUDO pra gente ter a melhor experiência das
nossas vidas, com as festas mais incríveis de todas, vídeos hilários que acabavam com a
tensão, piadas e recadinhos, banhos de sol e a cerveja nossa de todo dia! Paulinho
Morcegão “Mauravilhoso”, muito obrigada por tudo, pela companhia nas festas e pelo
estímulo sempre! Rainbow Master (Paulinho Bom), não sei nem o que dizer (será se?)!
Obrigada pela companhia, conversas, revelações, fofocas, por conseguir ficar em
silêncio (ou não) alguns segundos enquanto eu tentava medir a herbivoria nas
palmeirinhas, e por ser tão Paulinho Rainbow Master! Adooooro! Obrigada por tudo,
sempre!!!!!!!!!
Referências
Baraza, E., R. Zamora, J.A. Hódar & J.M. Gómez. 2007. Plant–herbivore interaction:
beyond a binary vision, pp. 481–514. In: Functional plant ecology (F.I. Pugnaire
& F. Valladares, eds.). Florida: CRC Press.

221
Coley, P.D. & J.A. Barone. 1996. Herbivory and plant defenses in tropical forests.
Annual Review of Ecology and Systematics, 27:305–335.
Dinnage, R., M.W. Cadotte, N.M. Haddad, G.M. Crutsinger & D. Tilman. 2012.
Diversity of plant evolutionary lineages promotes arthropod diversity. Ecology
Letters, 15:1308–1317.
Frederickson, M.E. & D.M. Gordon. 2007. The devil to pay: a cost of mutualism with
Myrmelachista schumanni ants in ‘devil’s gardens’ is increased herbivory on
Duroia hirsuta trees. Proceedings of the Royal Society of London B, 274:1117–
1123.
García, M.B. & J. Ehrlén. 2009. Evaluación de los efectos demográficos y evolutivos de
las interacciones planta-animal mediante modelos matriciales, pp. 95–111. In:
Ecología y evolución de interacciones planta-animal: conceptos y aplicaciones
(R. Medel, M.A. Aizen & R. Zamora, eds.). Santiago de Chile: Editorial
Universitaria.
Harper, J.L. 1977. Population biology of plants. London: Academic Press.
MacDougall, A.S. 2008. Herbivory, hunting, and long-term vegetation change in
degraded savanna. Biological conservation, 141:2174–2183.
Marques, E.S.A., P.W. Price, & N.S. Cobb. 2000. Resource abundance and insect
herbivore diversity on woody fabaceous desert plants. Environment Entomology,
29:696–703.
Medina, E. 2007. Tropical forests: diversity and function of dominant life-forms,
pp.313–350. In: Functional plant ecology (F.I. Pugnaire & F. Valladares, eds.).
Florida: CRC Press.

222
Novotny, V., Y. Basset, S.E. Miller, G.D. Weiblen, B. Bremer, L. Cizek, & P. Drozd.
2002. Low host specificity of herbivorous insects in a tropical forest. Nature,
416:841–844.
Novotny, V. & Y. Basset. 2005. Host specificity of insect herbivores in tropical forests.
Proceedings of the Royal Society of London B, 272:1083–1090.
Ødegaard, F., O.H. Diserud & K. Østbye. 2005. The importance of plant relatedness for
host utilization among phytophagous insects. Ecology Letters, 8:612–617.
Pimentel, D. 1961. The influence of plant spatial patterns on insect population. Annals
of the Entomological Society of America, 54:61–69.
Root, R.B. 1973. Organization of a plant-arthropod association in simple and diverse
habitats: the fauna of collards (Brassica oleracea). Ecological Monographs,
43:95–124.
Scariot, A. 1999. Forest fragmentation effects on palm diversity in central Amazonia.
Journal of Ecology, 87:66–76.
Tahvanainen, J.O. & R.B. Root. 1972. The influence of vegetational diversity on the
population ecology of a specialized herbivore, Phyllotreta cruciferae
(Coleoptera: Chrysomelidae). Oecologia, 10:321–342.
Weiblen, G.D., C.O. Webb, V. Novotny, Y. Basset, & S.E. Miller. 2006. Phylogenetic
dispersion of host use in a tropical insect herbivore community. Ecology, 87:62–75.

223
Interação mutualista entre afídeos e formigas não altera a eficácia de defesa da
mirmecófita Tococa sp. por formigas
Cibele Paiva
Introdução
O mutualismo é uma associação em que espécies diferentes convivem para obter
benefícios mútuos (Vasconcelos 1991). Este tipo de interação é muito comum entre
insetos e plantas, em especial nas plantas mirmecófitas, que são plantas que interagem
com formigas (Benson 1985). As mirmecófitas possuem estruturas denominadas
domáceas que são usadas como abrigo por colônias de formigas e outros insetos. Já as
formigas conferem proteção às plantas consumindo fungos patógenos e herbívoros
(Fonseca 1994).
O forrageio das formigas em mirmecófitas reduz a taxa de herbivoria
(Vasconcelos 1991), de modo que, sem essa interação, as mirmecófitas não chegariam à
maturidade reprodutiva (Fonseca & Ganade 1996). Essas formigas exibem um
comportamento de recrutamento em massa para atacar os herbívoros que consomem a
planta (Vasconcelos 2001).
As formigas que ocorrem em mirmecófitas também podem exibir interações
mutualistas com outros insetos. Isso ocorre, por exemplo, com afídeos, que ao consumir
a seiva da planta secretam uma substância açucarada rica em carboidratos (Schurch
2000). As formigas consomem esse composto, podendo inclusive, se alimentar do
próprio afídeo (Flatt & Weisser 2000). Em troca as formigas oferecem proteção aos
afídeos contra inimigos naturais (Vasconcelos 1991). Essa relação entre afídeo e
formiga é obrigatória e o afídeo é levado à domácea pelas formigas (Wimp & Whitham
2001).

224
As formigas que interagem com afídeos adquirem uma alta quantidade de
exudato. Isso pode diminuir a procura por alimento, acarretando em menor forrageio das
formigas nas plantas e menor defesa contra herbívoros. Sendo assim, avaliei se a
quantidade de afídeos na domácea influencia a eficácia de defesa das plantas pelas
formigas. Espero que o aumento no número de afídeos nas domáceas diminua a
quantidade de formigas que patrulham a folha e consequentemente aumente a área das
folhas consumida por herbívoros.
Métodos
Realizei o presente estudo em uma área de floresta de terra firme na reserva florestal do
km 41, localizada 80 km ao norte de Manaus, Brasil. Esta reserva é uma Área de
Relevante Interesse Ecológico (ARIE) co-gerida pelo Projeto Dinâmica Biológica de
Fragmentos Florestais (PDBFF) e pelo Instituto Chico Mendes de Conservação da
Biodiversidade (ICMBio).
Em 18 plantas de Tococa sp. selecionei quatro folhas que apresentavam
diferentes graus de herbivoria. Em cada folha, quantifiquei durante 30 segundos o
número de indivíduos da formiga Azteca sp. que patrulhavam a folha. Após a contagem
das formigas, coletei as folhas e abri as domáceas para quantificar o número de
formigas e afídeos presentes em seu interior. Para calcular a proporção da folha
consumida por herbívoros, coloquei a folha sobre um papel milimetrado e quantifiquei a
área do limbo que havia sido removida. Posteriormente, medi o comprimento e a largura
da folha e calculei a área da folha utilizando a fórmula de área da elipse. A proporção de
área consumida foi a razão entre a área do limbo removida e a área da total da folha.
Para testar a previsão de que o aumento no número de pulgões nas domáceas diminui o
forrageio pelas formigas e aumenta a herbivoria, fiz duas análises de covariância

225
utilizando a planta, a quantidade de afídeos e a quantidade de formigas na domácea
como variáveis preditoras e a porcentagem da folha consumida por herbívoros e o
número de formigas patrulhando as folhas como variáveis resposta.
Resultados
Em cada domácea o número médio (± desvio padrão) de formigas foi de 48,6 ± 40,3
com variação de uma a 233 formigas por domácea. Em cada folha observei em média
6,6 ± 6,4 formigas patrulhando, variando de uma a 29 formigas em cada folha. Em cada
domácea encontrei em média 5,7 ± 11,2 afídeos com variação de um a 76 afídeos por
domácea. A porcentagem média da área da folha consumida por herbívoros foi de 6,4%
± 7,5, variando entre um a 31,2%. Não houve relação entre a quantidade de afídeos
presentes nas domáceas e o número de formigas patrulhando as folhas (F(1,64)= 0,95; p =
0,33; Figura 1), e nem entre a quantidade de afídeos nas domáceas e a porcentagem das
folhas consumidas por herbívoros (F(1,64)= 0,95; p = 0,33; Figura 2).
226
Figura 1. Relação entre os resíduos da análise de covariância entre o número de
formigas patrulhando a folha, o número de formigas na domácea e a planta e o número
de afídeos por domácea.
Figura 2. Relação entre os resíduos de uma análise de covariância entre a porcentagem
da folha consumida por herbívoros, o número de operárias na domácea e a planta e
número de afídeos por domáceas.

227
Discussão
O número de afídeos presentes nas domáceas não influenciou o patrulhamento da folha
pelas formigas nem a área das folhas consumidas por herbívoros. Isso indica que a
interação mutualista entre formigas e afídeos não interfere na eficácia de defesa da
mirmecófita pelas formigas associadas a ela. É possível que a presença de afídeos nas
domáceas não represente uma quantidade significativa de recurso alimentar para que as
formigas diminuam a capacidade de forrageio. O tipo de recurso oferecido pelos afídeos
é diferente do tipo do recurso obtido pelo forrageio em busca de herbívoros. Os afídeos
fornecem carboidrato para as formigas enquanto os herbívoros constituem proteína para
a colônia (Morim 2011). Assim, mesmo que haja mais afídeos, as formigas continuam
dependendo do patrulhamento para captura de herbívoros a fim de garantir alimento
suficiente para a colônia.
A associação mutualista entre formigas e mirmecófitas é bastante estreita, o que
é evidenciado pela presença de estruturas vegetais especificas para essa interação
(domáceas), por comportamentos específicos das formigas (e.g patrulhamento e defesa
contra herbívoros) e pelo fato das formigas dependerem dessa interação para atingir a
maturidade reprodutiva (Fonseca & Ganade 1996). É possível, portanto, que o
comportamento de patrulha e defesa desempenhado pelas formigas seja fixado
evolutivamente e ocorra independentemente da necessidade de obtenção de outros
recursos pelas formigas.
Agradecimentos
Agradeço imensamente a Deus por permitir viver essa experiência única que levarei por
toda a minha vida!!! A minha família pelo apoio e por acreditarem no meu sonho,

228
minhas irmãs Cintia Paiva e Simara Fernanda, minha mãe e minha sobrinha Sophia amo
muito vocês e obrigada por entenderem minha ausência.
A Universidade Estadual de Feira de Santana pelo apoio e, ao PDBFF/INPA por
oferecer um curso tão maravilhoso e enriquecedor;
A todos que fizeram parte do Curso de Ecologia da Floresta Amazônica em 2012, mas
de uma forma muito especial aos Paulinhos “Painha” (pessoa mais fantástica não
conheço) e ao “Malravilhoso” por todas as sugestões e ajuda durante o curso;
Ao André Junqueira que além de excelente revisor ainda é um artista nato,
obrigadãooo!!
A Mariana Velasque, amiga e parceira de todas as horas, nossa amizade se estende as
“arvres” da floresta amazônica (risos) obrigada por tudo!!
A Sarinha por todos os momentos compartilhados (isso incluiu Xexeu) risos;
A rainha dos“bilhetim” Dany Zoo, pessoa especial e talentosa
A José Hidasi , por tanto carinho e timidez adorável;
A todas as meninas da coletividade feminina (Taty pernambucana arretada e prestativa,
Lory a manauara mais doce que conheci, a Carol Draxler carioca sangue bom,
Carolzinha pé de ouro, Bianca, Renatinha, Claudinha).
Aos colegas Daniel, Bruno, Leo, Tatu, Randolfo, Vidal, Joselandio por tanta animação!
Aos monitores Marcus e Thiago Kloss por sempre estarem prontos para ajudar!!!!
Referências
Benson, W.W. 1985. Amazon antplants, pp. 239-266. In: Amazonia (G.T. Prance &
T.E.C.P. Lovejoy, eds). New York: Pergamon Press.
Flatt, T. & W. Weisser. 2000. The effects of mutualistic ants on aphid life history traits.
Ecology, 81:3522-3529.

229
Fonseca, C.R. 1994. Herbivory and the long‐lived leaves of an Amazonian ant‐tree.
Journal of Ecology, 82: 833‐ 842.
Izzo, T.J. 2005. Recolonização de capoeiras com diferentes histórias de uso por plantas
mirmecófitas e suas formigas associadas. Tese de doutorado. Instituto Nacional de
Pesquisas da Amazônia, Manaus.
Morim, P.J. 2011. Community Ecology. Blackwell Science. New Jersey.
Schurch, S., M. Pfunder & B. A. Roy 2000. Effects of ants on the reproductive success
of Euphorbia cyparissias and associated pathogenic rust fungi. Oikos 88: 6-12.
Vasconcelos, H.L. 1991. Mutualism between Maieta guianensis Aubl., a
myrmecophytic melastome, and one of its ant inhabitants: ant protection against
insect herbivores. Oecologia, 87: 295‐298.
Vasconcelos, H.L. & D.W. Davidson, 2000. Relationship between plant size and ant
associates in two amazonian ant-plants. Biotropica, 32: 100-111.
Wimp. G. & T.Whitham. 2001. Biodiversity consequences of predation and host plant
hybridization on an aphid–ant mutualism. Ecological Society of America, 440-
452.

230
Pequenos camuflados e grandes esquivos? O tamanho corpóreo não atua na
seleção de habitat em peixes de riachos amazônicos
Daniel Passos
Introdução
A seleção de habitat é a escolha de um ambiente adequado à sobrevivência e reprodução
por um organismo (Morris 2003). Para que esse processo ocorra, é necessário que
indivíduos sejam capazes de perceber a heterogeneidade do ambiente em que vivem e
selecionar áreas de melhor qualidade (MacArthur & Pianka 1966). A escolha destes
locais específicos pode ser influenciada por diversos fatores, como as condições
abióticas do ambiente, a disponibilidade de recursos e as relações interespecíficas, como
a competição e a predação (Allan 1995).
A predação, em particular, pode atuar como pressão seletiva sobre o
comportamento das presas, induzindo mudanças na escolha do habitat por elas (Endler
1991, Lima 1998). Nestes casos, as presas podem reduzir o risco de serem predadas ao
escolher habitats nos quais o acesso de predadores é dificultado (Wooster & Sih 1995)
ou se tornando crípticas em resposta ao principal sentido sensorial do predador
(Edmunds 1974). Especificamente para presas suscetíveis a predadores visualmente
orientados, a seleção de substratos com coloração similar a sua pode aumentar sua
camuflagem e reduzir a chance de serem detectadas.
Além da camuflagem visual, o tamanho do corpo da presa também pode afetar a
sua chance de detecção pelo predador (e.g. Ware 1973). Maiores dimensões corpóreas
podem aumentar o contraste visual entre a presa e o substrato e, consequentemente,
influenciar sua detecção por predadores visualmente orientados (Edmunds 1974). Neste
sentido, indivíduos maiores podem ser mais facilmente percebidos que os menores e,

231
portanto, devem selecionar ambientes com coloração mais similar à de seu tegumento
para reduzir a chance de serem predados.
A ictiofauna de ambientes aquáticos de pequeno porte, como os riachos
amazônicos, sofre forte influência da pressão predatória (Lowe-McConnell 1975,
Mendonça et al. 2005). Nestes sistemas, os peixes usam uma grande variedade de
substratos (Mortatti 2004) e isto pode estar relacionado à busca por ambientes nos quais
as presas se tornem menos visíveis para os predadores. Nesse sentido, visto que peixes
maiores podem ser mais facilmente detectados por predadores externos ao riacho,
proponho a hipótese de que peixes maiores são mais seletivos quanto à escolha do
habitat do que peixes menores. Se essa hipótese for verdadeira, espero que o tamanho da
área dorsal do peixe esteja positivamente relacionado à similaridade de coloração entre
o seu tegumento e o substrato em que ele se encontra. Além disso, espero que, em um
dado morfotipo, o tamanho da área dorsal do indivíduo esteja positivamente relacionado
à similaridade de coloração entre o seu tegumento e o substrato em que ele se encontra.
Métodos
Área de estudo
Realizei este estudo em um riacho de segunda ordem, localmente chamado de igarapé,
com águas claras e rasas, em uma floresta de terra firme na Amazônia brasileira. A área
específica de estudo (2° 25´ S, 59° 45´ O) está inserida na reserva do Km 41, localizada
a 80 km ao norte de Manaus - AM, Brasil, pertencente a uma Área de Relevante
Interesse Ecológico (ARIE) co-gerida pelo Projeto Dinâmica Biologica de Fragmentos
Florestais (PDBFF) e pelo Instituto Chico Mendes de Conservação da Biodiversidade
(ICMBio). . O clima da região é tropical úmido, com temperatura média anual de 26,7
ºC e pluviosidade superior a 2000 mm por ano (Lovejoy & Bierregaard 1990).

232
Coleta de dados
Realizei buscas ativas noturnas à procura de peixes ao longo de um trecho de 50 m no
igarapé, vasculhando os bancos de areia branca, os aglomerados de folhiço do leito, bem
como suas poças adjacentes. Os peixes do igarapé, em sua maioria diurnos, não alteram
o uso do habitat entre dia e noite (observação pessoal). Portanto, realizei coletas à noite
pela maior facilidade em capturar indivíduos inativos. Registrei fotograficamente, em
vista dorsal e a 25 cm da superfície da água, cada peixe encontrado e seu substrato
adjacente. Em seguida, coletei cada indivíduo com uso de um rapiché de 50 cm² e malha
de 1 mm². Posteriormente, classifiquei os peixes em morfotipos e medi o comprimento
(mm) e a largura (mm) de cada indivíduo capturado.
Para estimar a área dorsal dos peixes, utilizei a fórmula da área da elipse (Ap=
π*C/2*L/2), onde “Ap” é a área dorsal do peixe (mm²), “C” é o comprimento (mm) e
“L” é a largura (mm). Para estimar a similaridade de coloração entre os peixes e os
substratos adjacentes, calculei a quantidade média de pixels vermelhos, verdes e azuis
(RGB) na superfície dorsal dos peixes e em seis amostras de substrato (1 cm² cada).
Distribuí sistematicamente as amostras de substrato ao redor do peixe, a uma distância
fixa equivalente a largura do indivíduo (Figura 1). Posteriormente, calculei o módulo da
diferença de RGB entre a média das seis amostras de substrato e a média da superfície
dorsal do peixe. Desta forma, quanto menor a diferença em RGB, maior a similaridade
de coloração entre o peixe e o substrato.

233
Figura 1. Esquema do método de cálculo da similaridade de coloração entre os peixes e
os substratos adjacentes. Círculos cinzas representam amostras de substrato de 1 cm².
Barras pretas representam a largura do peixe, bem como a distância aproximada entre as
amostras de substrato e o peixe.
Análises estatísticas
Para avaliar a similaridade de coloração entre os peixes e o substrato adjacente, fiz uma
correlação de Pearson. Para testar a relação entre área dorsal e a similaridade de
coloração entre peixe e substrato, fiz duas regressões lineares simples. Na primeira,
utilizei a área dorsal média de cada morfotipo como variável preditora e a diferença
média de RGB entre peixe e substrato como variável resposta. Na segunda, utilizei a
área dorsal dos indivíduos do morfotipo mais abundante como variável preditora e a
diferença média de RGB entre cada indivíduo e seu respectivo substrato como variável
resposta.
1 cm²

234
Resultados
Coletei 41 indivíduos classificados em nove morfotipos. A coloração dos morfotipos foi
correlacionada com a coloração do ambiente (r = 0,84; p < 0.01). Não houve relação
entre a área dorsal média dos morfotipos e as diferenças médias de RGB entre os peixes
e seus respectivos substratos (R² = 0,02; F(1,7) = 0,17; p = 0,69; Figura 2). Também não
houve relação entre a área dorsal média e as diferenças médias de RGB entre os peixes e
seus respectivos substratos, quando removi os morfotipos representados por apenas um
indivíduo (R² = 0,01; F(1,4) = 0,04; p = 0,86).
Figura 2. Relação entre área dorsal média e a diferença média de RGB entre os
morfotipos de peixes e seus substratos adjacentes, em um igarapé na reserva do Km 41,
Amazonas, Brasil. Os números sobre os pontos representam o número de indivíduos de
cada morfotipo.
O morfotipo mais abundante (N = 10) apresentou área dorsal média de 275,14 ±
117,94 mm² (média ± desvio padrão). Não houve relação entre a área dorsal dos
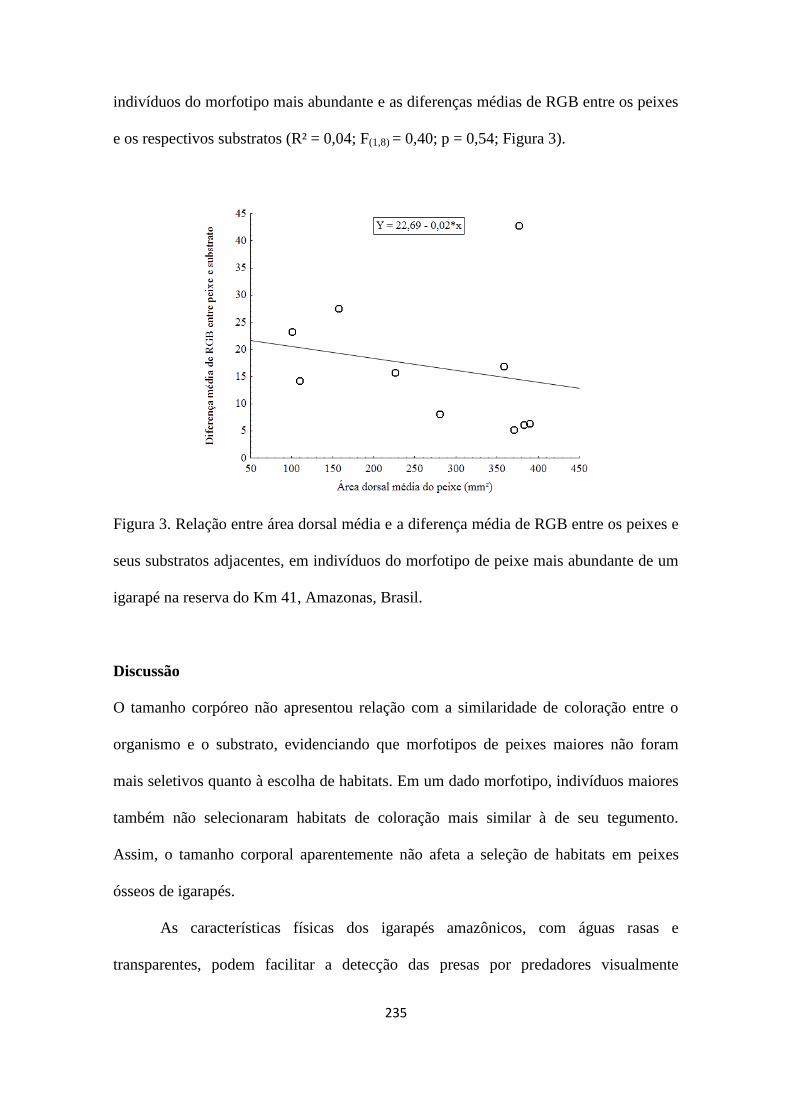
235
indivíduos do morfotipo mais abundante e as diferenças médias de RGB entre os peixes
e os respectivos substratos (R² = 0,04; F(1,8) = 0,40; p = 0,54; Figura 3).
Figura 3. Relação entre área dorsal média e a diferença média de RGB entre os peixes e
seus substratos adjacentes, em indivíduos do morfotipo de peixe mais abundante de um
igarapé na reserva do Km 41, Amazonas, Brasil.
Discussão
O tamanho corpóreo não apresentou relação com a similaridade de coloração entre o
organismo e o substrato, evidenciando que morfotipos de peixes maiores não foram
mais seletivos quanto à escolha de habitats. Em um dado morfotipo, indivíduos maiores
também não selecionaram habitats de coloração mais similar à de seu tegumento.
Assim, o tamanho corporal aparentemente não afeta a seleção de habitats em peixes
ósseos de igarapés.
As características físicas dos igarapés amazônicos, com águas rasas e
transparentes, podem facilitar a detecção das presas por predadores visualmente

236
orientados, como aves piscívoras da família Alcedinidae (Sick 1997). Se a chance de
detecção for constantemente alta nesses ambientes, os peixes podem apresentar intensa
seletividade de habitat, independente do seu tamanho corpóreo. Por outro lado, os
peixes maiores podem apresentar estratégias alternativas para reduzir os riscos da
predação. Neste sentido, uma vez que a movimentação da presa pode contribuir para o
escape de predadores (Ware 1973), é possível que peixes maiores apresentem maior
capacidade de deslocamento que peixes menores e, portanto, sejam aptos a fugir mais
eficientemente do ataque de predadores.
A intensa seletividade de habitat pelas presas e o uso de estratégias defensivas
alternativas por presas maiores podem ter implicações sobre a dinâmica dos peixes em
níveis de organizações maiores. Se peixes de maior tamanho de fato usam o
deslocamento como principal estratégia de escape da predação, então isso deve refletir
em mudanças ontogenéticas no padrão de uso do espaço em uma dada espécie, como
também em uma maior substituição de espécies de grande porte entre habitats com
diferentes tipos de substratos.
Agradecimentos
À coordenação do “Projeto Dinâmica Biológica de Fragmentos Florestais” (PDBFF) e
do curso “Ecologia da Floresta Amazônica” (EFA), pela oportunidade de participar
desta experiência inesquecível e pela enorme contribuição à minha formação
profissional e cultural. Aos coordenadores, professores, monitores, funcionários e
colegas do EFA 2012 pelo surpreendentemente agradável convívio durante a realidade
paralela que é esta imersão social no coração da Amazônia. Em particular, aos
professores Mike Hopkins, Jansen Zuanon e Glauco Machado pelo enorme exemplo de
excelentes profissionais e pelas imensuráveis contribuições à minha formação

237
intelectual. Principalmente, ao estimado prof. Paulo Enrique Peixoto por toda a
competência, dedicação e por todos os ensinamentos transmitidos ao longo do curso,
que tornaram esta experiência de vida indubitavelmente mais engrandecedora (VÔCE se
garante “Libélula”). Aos colegas de curso, pelo respeito, atenção e companheirismo
durante todas as fases do EFA 2012, tornando a vivência cotidiana cordialmente
tolerável nestes intensos 30 dias. Em especial aos mais próximos, Ana Carolina Vieira,
Daniela Coelho, Ana Cláudia Rorato, Vidal Carrascosa, José Hidasi e Arthur Madeira,
pela competência, organização e responsabilidade, que tornaram nossa convivência fácil
e afetuosa durante a execução dos projetos.
Especificamente quanto a este projeto, agradeço ao prof. Jansen Zuanon, pela
orientação sobre a fascinante ictiofauna amazônica e por me instigar a trabalhar com os
peixes de igarapés no projeto individual. Aos profs. Paulo Enrique Peixoto, André
Junqueira, Paulo Bobrowiec e Marcel Vaz e aos monitores Marcos Vieira e Thiago
Kloss pelas sugestões e críticas à atual versão deste trabalho. Em especial, à Carol Pires,
pela prazerosa companhia durante as pescarias, pela dedicação na realização das coletas
e principalmente por toda a atenção nos momentos adversos.
Referências
Allan, J.D. 1995. Stream ecology: Structure and function of running waters.
London:Chapman & Hall.
Edmunds, M. 1974. Why are there good and poor mimics? Biological Journal of the
Linnean Society, 70:459-466.
Endler, J.A. 1991. Variation in the appearance of guppy color patterns to guppies and
their predators under different visual conditions. Vision Research, 31:587-608.

238
Lima, S.L. 1998. Nonlethal effects in the ecology of predator-prey interactions.
Bioscience, 48:25-34.
Lovejoy, T.E. & R.O. Bierregaard. 1990. Central amazonian forests and the minimal
critical size of ecosystems project, pp. 60‐71. In: Four neotropical rainforests
(A.H. Gentry, ed.). London: Yale University Press.
Lowe-McConnell, R.H. 1975. Fish communities in tropical freshwaters: their
distribution, ecology and evolution. London: Longman.
MacArthur, R.H. & E.R. Pianka. 1966. On optimal use of a patchy environment.
American Naturalist, 100:603-609.
Mendonça, F.P., W.E. Magnusson & J. Zuanon. 2005. Relationships between habitat
characteristics and fish assemblages in small streams of central Amazonia.
Copeia, 2005:751-764.
Morris, D.W. 2003. Toward an ecological synthesis: a case for habitat selection.
Oecologia, 136:1-13.
Mortatti, A.F. 2004. Colonização por peixes no folhiço submerso: implicações das
mudanças na cobertura florestal sobre a dinâmica da ictiofauna de igarapé de
terra firme, na Amazônia Central. Dissertação de Mestrado, INPA/UA, Manaus.
Reis, R.E., S.O. Kullander, J. Ferraris & J. Carl. 2003. Check list of the freshwater
fishes of south and central America. Porto Alegre: EDIPUCRS.
Sick, H. 1997. Ornitologia brasileira. Rio de Janeiro: Nova Fronteira.
Ware, D.M. 1973. Risk of epibenthic prey to predation by rainbow trout (Salmo
gairdneri). Journal of Fish Research Board, 30:787-797.
Wooster, D. & A. Sih. 1995. A review of the drift and activity responses of stream prey
to predator presence. Oikos, 73:3-8.

239
A herbivoria em mirmecófitas afetando a resposta da colônia de formigas
Daniela Pinto Coelho
Introdução
O comportamento exibido por animais diante do risco de serem predados é uma
resposta condicionada a pressão exercida pelo predador (Kemp & Jones 2001). Diante
de uma ameaça, os organismos tendem a avaliar os custos do investimento energético
em fugir e abandonar o recurso ou em ficar e defendê-lo (Lima 1998, Sansom et al.
2009). Reações de formigas que têm suas colônias ameaçadas, por exemplo, irão
depender de alguns fatores. Dentre eles, a frequência e a intensidade do distúrbio, além
da capacidade de investimentos de recursos da colônia na geração de uma nova colônia
ou na produção de novos indivíduos para incrementar sua defesa (Lima 1998, Philpott
et al. 2010).
Colônias de formigas podem interagir de forma mutualística com determinadas
espécies de plantas, conhecidas como mirmecófitas. Nesta relação, as formigas
conferem proteção para planta contra ação de insetos fitófagos e, em troca, a planta
oferece abrigo e locais de oviposição para as formigas (Janzen 1966, Christianini &
Machado 2004). Estes abrigos, chamados de domáceas, são estruturas infladas e com
orifícios que podem se situar em diferentes partes da planta (Frederickson 2005).
A ocupação da planta mirmecófita pelas formigas ocorre em um indivíduo ainda
jovem. As formigas aladas ocupam uma das domáceas e ovipositam, dando início à
colônia, com posterior ocupação das demais domáceas existentes (Janzen 1966). A
colônia consiste basicamente de três castas: rainhas, operárias e soldados. As rainhas
são as maiores formigas da colônia, sendo aladas e responsáveis pela geração de novos
indivíduos. As operárias são formigas menores, mais abundantes e responsáveis pelo

240
patrulhamento da planta e pela liberação de feromônio de alarme contra herbívoros. Os
soldados são indivíduos maiores que as operárias, com grandes mandíbulas e que
respondem à liberação do feromônio das operárias sendo responsáveis pelo ataque aos
invasores. A relação entre soldados e operárias é bastante estreita, já que é o
patrulhamento eficiente das operárias implicará no sucesso de resposta dos soldados à
herbivoria. Vale ressaltar, que o investimento na produção de soldados pode ser bastante
custoso para colônia, já que eles são recrutados apenas quando há a ação de herbívoros
(Hölldobler & Wilson 1990).
Folhas mais jovens (que neste trabalho serão chamadas de apicais, por situarem-
se na região mais apical do ramo), oferecem maior contribuição para a produtividade da
planta por meio de seus tecidos fotossintéticos mais eficientes. Porém, estas folhas
tendem a ser mais atrativas para herbívoros o que faz com que a ocupação das domáceas
e as ações de defesa da colônia de formigas sejam mais intensas em folhas apicais do
que em folhas mais antigas (denominadas neste trabalho de folhas basais, por situarem-
se em posições mais basais no ramo). Logo, se o sucesso da colônia está diretamente
relacionado com a qualidade da planta, os esforços em melhorar o patrulhamento e as
ações de defesa podem ocorrer de forma mais eficiente em folhas apicais. Dessa forma,
folhas basais ficam mais suscetíveis à predação do que folhas apicais (Christianini &
Machado 2004).
Sendo a herbivoria uma força que afeta a integridade da colônia de formigas em
mirmecófitas e levando-se em consideração o sistema de proteção por formigas descrito
anteriormente, este trabalho pretende estudar se a proporção de formigas soldados e
operárias de Pheidole minutula em folhas apicais de Maieta guianensis está relacionado
à herbivoria em folhas basais. Acredita-se que o investimento na geração de soldados
pela colônia em folhas apicais é positivamente relacionado à herbivoria em folhas

241
basais. Sendo assim, quanto maior a área de herbivoria em folhas basais, maior será a
quantidade de soldados presentes nas domáceas de folhas apicais.
Métodos
Área de estudo
O estudo foi realizado na Fazenda Esteio (Km 41), localizada a cerca de 80 km ao norte
de Manaus, Amazonas, Amazônia Central, Brasil. A reserva esta inserida na Área de
Relevante Interesse Ecológico (ARIE) sendo co-gerenciada pelo Projeto Dinâmica
Biológica de Fragmentos Florestais (PDBFF) e pelo Instituto Chico Mendes (ICMBio).
A vegetação é formada por floresta ombrófila densa. O clima é do tipo tropical úmido
de acordo com a classificação de Köppen (Peel et al. 2007) com temperatura média
anual de 26,7˚C e pluviosidade anual variando entre 1900 e 2500 mm (Lovejoy &
Bierregaard 1990).
Coleta de dados
Foram amostrados 37 indivíduos de Maieta guianensis ao longo das margens de
um Igarapé. De cada indivíduo foi retirado um ramo inteiro da porção superior da
planta. Em cada ramo, foram retiradas as duas folhas apicais e as folhas basais. As
amostras apicais e basais de cada indivíduo foram acondicionadas separadamente em
sacos de papel para que não houvesse a migração de formigas de uma amostra para
outra. Posteriormente, o material foi processado em laboratório.
Para mensuração dos indivíduos de Pheydole minutula em suas respectivas
castas, as domáceas das folhas apicais foram abertas e os indivíduos quantificados. A
mensuração da área foliar de folhas basais de M. guianensis foi realizada através do
cálculo da área média foliar por meio da fórmula da elipse [(R x r x π) / 4], onde R é o
raio maior e r o raio menor. Para o cálculo da área média de herbivoria em folhas basais,

242
foi utilizada uma malha transparente preenchida com quadrículas de 0,25 cm2. A malha
era colocada sobre a folha e a estimativa da quantidade de dano foliar era contabilizada
somando-se da área das quadrículas correspondentes.
Análises estatísticas
Para avaliar se a herbivoria nas folhas basais era positivamente relacionada à quantidade
de formigas soldados nas folhas apicais foi feita uma regressão linear múltipla
utilizando a quantidade de formigas operárias como covariável. Essa medida foi tomada
com o objetivo de controlar o efeito da quantidade de formigas operárias na quantidade
de formigas soldados.
Resultados
Dos 37 ramos coletados de Maieta guianensis, um não apresentou ocupação por
formigas, sendo, portanto, excluído das análises. Foi analisado um total de 72 domáceas
em folhas apicais e 123 folhas basais. A porcentagem média de herbivoria nos ramos
com folhas basais foi de 15,52% (mínimo = 1,41%; máximo = 52,85%). A quantidade
total de formigas operárias encontradas em todas as domáceas de folhas apicais
analisadas foi de 1.135 (mínimo = 10; máximo = 86) e a de formigas soldados foi de
241 (mínimo = 1; máximo = 23).
Em todas as domáceas analisadas, com exceção de dois ramos (que serão
denominados de discrepantes), havia a presença de formigas operárias, soldados e ovos.
Estes ramos discrepantes apresentaram apenas formigas operárias. Vale ressaltar, que as
folhas basais destes ramos discrepantes apresentaram elevado tamanho de área foliar
(22,13 cm2 e 25,08 cm
2, respectivamente; média do total de indivíduos = 22,14 cm
2).
Um deles também apresentou a segunda maior taxa de herbivoria encontrada (52,8%).

243
Por haver esta discrepância de ocupação em relação ao padrão observado, foram feitas
duas análises. Uma contendo a presença destes dois ramos discrepantes e outra os
excluindo. Na análise com a presença dos ramos discrepantes, a quantidade de formigas
soldados presentes nas domáceas de ramos apicais não foi explicada pela porcentagem
de herbivoria nos ramos basais (F(1,33) = 2,809; p = 0,103). Porém, com a retirada destes
da análise, foi encontrada uma relação positiva entre a porcentagem de herbivoria nos
ramos basais e a quantidade de formigas soldados nas domáceas de ramos apicais (F(1,31)
= 10,843; p = 0,002), como previsto no estudo (Figura 1).
Figura 1: Relação positiva entre a área de herbivoria (cm2) em folhas basais e o resíduo
de uma regressão linear entre a quantidade de operárias pela quantidade de soldados da
formiga Pheidole minutula presentes em domáceas de folhas apicais da planta
mirmercófita Maieta guianensis.
Discussão
Na presença dos ramos discrepantes, a resposta da colônia diante da herbivoria em
folhas basais não se manifesta através do investimento na produção de soldados. Como

244
a resposta da colônia é condicionada à pressão de herbivoria, esta ausência de relação
mostra que a herbivoria não seria uma pressão suficientemente forte para que a colônia
desloque recursos no investimento em soldados (Hölldobler & Wilson 1990). Porém,
estes dois ramos discrepantes apresentaram um padrão diferenciado dos demais ramos
coletados. Ambos tinham elevado tamanho de área foliar, o que pode estar associado à
idade mais avançada destas plantas. Além disso, um destes ramos também apresentou a
segunda maior taxa de herbivoria, mostrando que a colônia de formigas poderia não
mais estar defendendo a planta de forma eficiente (Izzo 2005). Sendo assim, acredita-se
que estas colônias poderiam estar em processo de evasão das duas plantas coletadas.
Por outro lado, sem considerar os ramos discrepantes, a relação positiva entre a
herbivoria em folhas basais e a proporção de soldados e operárias em folhas apicais
mostra que a ação anterior da herbivoria pode ser o fator preponderante para o
investimento posterior da colônia na geração de soldados. Dessa forma, a evasão da(s)
rainha(s) para fundar uma nova colônia, torna-se uma estratégia mais custosa do que
investir na produção de soldados. Esta questão pode ser corroborada pelo fato de que,
das 37 plantas examinadas neste trabalho apenas uma não apresentou ocupação por P.
minutula. Isso pode indicar que existe uma pequena disponibilidade de sítios vazios
para fundação de uma nova colônia, o que aumentaria a vantagem na produção de
soldados.
Como evidências prévias indicam uma forte co-dependência do sistema
Pheydole – Maieta, o impacto negativo do abandono da planta pela colônia pode refletir
em uma redução do sucesso reprodutivo da planta. Isso mostra que o resultado
encontrado pela análise sem os ramos discrepantes faz mais sentido, pois estes dois
ramos discrepantes destoaram do padrão encontrado e já descrito pela literatura
(Vasconcelos 1999). Para evitar o abandono pela colônia de formigas, a planta pode

245
investir na produção de folhas com domáceas novas para que ocorra um aumento do
tamanho da colônia que ocupa aquele indivíduo e, por conseguinte, aumento da
proteção da planta contra herbivoria (Janzen 1966, Christianini & Machado 2004).
Dessa forma, sendo a herbivoria um fator que afeta a integridade da colônia de
formigas, o investimento na produção de soldados para minimizar a ação de herbívoros
nas novas folhas produzidas é a estratégia de investimento mais vantajosa da colônia de
P. minutula.
Agradecimentos
Agradecimentos eternos ao INPA e ao PDBFF pela chance única e inesquecível de
participar deste curso sensacional!!! A intensidade de tudo que vivi aqui não será
refletida apenas em minha vida profissional, isso é fato!!! Hipotetizo (falo do jeito que
eu quiser, o relatório é meu!) que Rainbow master jamais se esquecerá de mim! E agora,
quem irá me arrasar 25 horas por dia? Paulinho, não te largarei jamais, vou ser que nem
mucuim “ni” você! Feira de Santana que me aguarde!!! Só um detalhe, “sou brasileira e
não desisto nunca”! Venha cá, o que seriam dos meus dias sem as mega risadas
provocadas pelas frases delicadas (só que ao contrário) e hiper criativas do Paulinho
“Mau”ravilhoso? Paulinhos, vocês preencheram o meu “eu” de alegria! Vocês são o “+”
de minha vidinha “+ ou -”! A todos os meus queridos novos amigos, que há um mês eu
nem fazia ideia que existiam, não consigo imaginar minha vida sem vocês! Comunidade
feminina, nossa força não pode morrer, nossa pegada agora vai ser bilhetinho virtual!!!
Em especial, à condessa Lohaynne Chayenne Vanessão (a musa inspiradora dos meus
cordéis), Morceguete da Silva, vulgo “Miss Balde” (minha cuidadeira pós festas,
companheira de rede, escrevedora de bilhetinhos e parceira da linguagem das letras),
Lori “Polidance” (irmã de alma), Tati “arvéres” (não resisti”), Sarah Baranga

246
(mooooorro de amor) e Mari Exú, já tô morrendo de saudade meninas!!! Comunidade
cueca, vocês são as fofuras de minha vida! Em especial, meu querido e agora pra
sempre novo amigo Daan, Joseph, meu ursinho pimpão (só vintchi), Vidal (meu
estrangeiro preferido) e Harry (não aguentava mais reclamar de suas roupas mal lavadas
e mal estendidas), por vocês, foi identificação à primeira vista! A todos os professores,
em especial Felipe e Toyoyo! Aos monitores Barbudinho (Nhéééun) e Beluga (meu
potinho de ninho solei, Ass: Cachorrona), vocês são meu máximo denominador
comum!!! Como tenho juízo, Manô foférrima, mal te conheci mas a baianidade tá no
sangue!!! Hahahaha... sou foda e você também! O EFA também não seria o mesmo sem
as comidas “deliça” da Dona Eduarda e, óóóóbeveo, sem Seu Jorge, afinal, quem me
ajudaria a acordar Morceguete da Silva na pegada da escrotidão, além de gelar nossa
cerveja pro Igarafest, fazer suco de caju todo dia e cuidar da gente com tanta fofura e
delicadeza (comigo ele foi fofo, se não foi com você, deita na BR)!!! Por fim, jamaaaais
esqueceria da pessoa que se tornou a mais importante na minha vida acadêmica neste
último ano. Você fez uma diferença absurda! Tô “Glauco-dependente”! Por fim, quero
ir pro CÉU e dar “mortal de cuestas” caraaaai!!!
Referências
Christianini, A.V. & G. Machado. 2004. Induced biotic responses to herbivory and
associated cues in the Amazonian ant-plant Maieta poeppigii. Entomologia
Experimentalis et Applicata, 112:81– 88.
Frederickson, M.E. 2005. Ant species confer different partner benefits on two
neotropical myrmecophytes. Oecologia, 143:387–395.
Hölldobler, B. & E.O. Wilson. 1990. The ants. London: Harward University Press.

247
Izzo, T.J. 2005. Recolonização de capoeiras com diferentes histórias de uso por plantas
mirmecófitas e suas formigas associadas. Tese de doutorado, Instituto Nacional
de Pesquisas da Amazônia, Manaus.
Janzen, D.H. 1966. Coevolution of mutualism between ants and acacias in Central
America. Internation Journal of Organic Evolution, 20:249-275.
Kemp, D.J. & R.E. Jones. 2001. Phenotypic plasticity in field populations of the tropical
butterfly Hypolimnas bolina (L.) (Nymphalidae). Biological Journal of the
Linnean Society, 72:33–45.
Lima, S.L. 1998. Nonlethal Effects in Ecology of Predator-Prey Interaction. Bioscience,
48:25-34.
Lovejoy T.E. & R.O. Bierregaard. 1990. Central Amazonian forests and the minimum
critical size of ecosystem project, pp. 60-71. In: Four neotropical rainforest
(Gentry, A.H., ed.). New Haven: Yale University Press.
Peel, M.C., B.L. Finlayson & T.A. McMahon. 2007. Updated world map of the
Köppen-Geiger climate classification. Hydrology and Earth System Sciences,
11:1633–1642.
Philpott, S.M., I. Perfecto, I. Armbrecht & C.L. Parr. 2010. Ant diversity and function
in disturbed and changing habitats, pp. 137-156. In: Ant Ecology (L. Lach, C.L.
Parr & K.L. Abbott eds.). Oxford University Press.
Sansom, A., J. Lind & W. Cresswell. 2009. Individual behavior and survival: the roles
of predator avoidance, foraging success, and vigilance. Behavioral Ecology,
20:1168–1174.
Vasconcelos, H.L. 1999. Effects of forest disturbance on the structure of ground-
foraging ant communities in central Amazonia. Biodiversity and Conservation,
8:409-420.

248
Habitats com maior luminosidade não representam melhores territórios de
acasalamento para machos da libélula Chalcopteryx scintilans (Odonata:
Polythoridae)
José Hidasi Neto
Introdução
Uma parte representativa dos animais possui o comportamento de brigar por territórios
que apresentam condições e recursos que possam aumentar sua sobrevivência (Adams
2001). Algumas espécies também utilizam territórios como locais de exibição e
acasalamento, atraindo indivíduos do sexo oposto para que possam copular (Contreras-
Garduño et al. 2006). Ainda, é sabido que os locais defendidos apresentam variações
em sua qualidade (Arnott & Elwood 2008). Sendo assim, como os territórios de maior
qualidade são mais disputados, é de se esperar que eles sejam ocupados por indivíduos
mais fortes.
Diversas características de um habitat podem determinar sua qualidade como um
território. Um fator que normalmente determina a escolha de um sítio de defesa são as
condições abióticas do local (Arnott & Elwood 2008). Em ambientes florestais, por
exemplo, habitats com maior incidência luminosa, como as clareiras, podem ser bons
territórios para o estabelecimento de espécies ectotérmicas (Velde et al. 2011). Além
disso, no caso de animais visualmente orientados, a alta luminosidade nas clareiras
poderia facilitar a detecção de parceiros de alta qualidade (e.g. Bergman & Wiklund
2009).
As libélulas (Odonata) constituem um grupo em que diversas espécies
apresentam comportamento territorial. Esses insetos são comumente encontrados em
locais ensolarados, pois dependem dos raios solares para termoregulação. Além disso,

249
as libélulas são organismos orientados primariamente pela visão (Córdoda-Aguilar
2008). Portanto, é possível que ambientes com maior incidência luminosa representem
territórios de maior qualidade para as libélulas, especialmente em ecossistemas com o
predomínio de áreas sombreadas, por facilitar a termorregulação e aumentar a chance de
detecção de parceiros sexuais.
Uma espécie de libélula comum no sub-bosque de florestas na Amazônia é
Chalcopteryx scintilans (Odonata: Polythoridae). Os machos dessa espécie possuem
uma coloração avermelhada conspícua em suas asas anteriores que provavelmente está
associada a um indicativo de qualidade do macho (Fitztephens & Getty 2000). Esse
indicativo pode servir como um aviso contra possíveis competidores e como uma pista
de qualidade para fêmeas, por estar relacionado a traços que determinam a saúde dos
machos (e.g. Contreras-Garduño et al. 2006). Outro indicativo de qualidade dos machos
é a sua densidade corporal, uma vez que esta é inversamente proporcional à quantidade
de reserva energética e, portanto, à capacidade de briga e de defesa do território
(Marden & Waage 1990; Contreras-Garduño et al. 2006). Portanto, machos de maior
qualidade em C. scintilans, devem ser aqueles que apresentam coloração mais
conspícua nas asas e que possuem menor densidade corporal.
Machos de C. scintilans são comumente encontrados próximos a manchas de sol
criadas por pequenas aberturas no dossel ao longo de riachos. É possível que machos
com melhor capacidade de briga se estabeleçam e protejam territórios com maior
incidência luminosa, pois estes permitiriam maior estabilidade em sua termorregulação
e maior facilidade de detecção de fêmeas, o que poderia aumentar o seu sucesso
reprodutivo (Bergman & Wiklund 2009). Portanto, minha hipótese é que áreas com
maior disponibilidade de luz representam territórios de melhor qualidade para machos
de libélula da espécie C. scintilans. Se essa hipótese for verdadeira, espero que machos

250
de C. scintilans encontrados em áreas com maior abertura do dossel apresentem asas
com maior intensidade de vermelho e menor densidade corporal.
Métodos
Realizei o estudo em um riacho localizado em uma área de floresta de terra firme
próximo ao acampamento do km 41 (02º 24’ S - 59º 44’ O), localizado em uma Área de
Relevante Interesse Ecológico (ARIE) cogerida pelo Projeto Dinâmica Biológica de
Fragmentos Florestais (PDBFF) e pelo Instituto Chico Mendes de Conservação da
Biodiversidade (ICMBio), a cerca de 80 km ao norte de Manaus, Brasil. Busquei
ativamente por machos de C. scintilans em machas de sol ao longo do riacho durante
dois dias entre as 10 h e as 14 h, o que aparentemente é o horário de maior atividade da
espécie. Coletei 14 indivíduos e fotografei o dossel do local onde cada indivíduo foi
encontrado. Duas clareiras apresentavam mais de um indivíduo. Nesses locais, coletei
uma única libélula e a considerei como uma amostra.
Posteriormente, utilizei as fotos do dossel para estimar a incidência luminosa dos
locais onde coletei as libélulas. Para isso, transformei as colorações das fotos em preto e
branco e calculei a abertura do dossel como a razão entre a quantidade de pixels brancos
e a quantidade total de pixels na imagem. Sendo assim, valores mais altos da proporção
de pixels brancos representam áreas com maior incidência luminosa.
Após a coleta, dissequei cada indivíduo para a retirada de suas asas, tórax e
abdômen. Em seguida, fotografei a asa anterior esquerda dos indivíduos e as usei para
calcular a média de vermelho na asa. Para isso, calculei a média de pixels vermelhos ao
longo das asas, visto que ela representa uma medida indireta da qualidade do macho
(quanto mais vermelha a asa, maior a sua qualidade). Realizei os cálculos da abertura do
dossel e da média de vermelho nas asas usando o programa ‘Image J’.

251
Para estimar a quantidade relativa de gordura acumulada nos indivíduos, calculei
a densidade corporal dos mesmos. Para esse cálculo, medi a massa torácica e abdominal
em uma balança semi-analítica (precisão de 1 mg) e dividi a soma dos valores obtidos
pela soma dos volumes do tórax e do abdômen. Para o cálculo dos volumes, considerei
que o tórax possuía a forma de um paralelepípedo e que o abdômen possuía a forma de
um cilindro. Dessa forma, estimei o volume do tórax pela fórmula “volume = altura x
largura x comprimento do tórax”, e o volume do abdômen pela fórmula “volume =
comprimento x raio x ”. Para testar se manchas de sol maiores têm maior qualidade
para machos de C. scintilans, realizei duas regressões lineares utilizando a abertura do
dossel como variável independente, e a média de vermelho nas asas e a densidade
corporal como variáveis dependentes.
Resultados
A proporção média de abertura do dossel foi de 16,03 ± 5,93% (média ± desvio padrão),
enquanto a média de vermelho nas asas de C. scintilans foi de 141,24 ± 27,04 pixels e
de densidade corporal média foi de 0,0004 ± 0,0001g/mm³. Não houve relação entre a
abertura do dossel e a média de vermelho na asa (F(1,12) = 0,128; p = 0,726; R² = 0,011;
Figura 1), nem entre a abertura do dossel e a densidade corporal (F(1,12) = 0,08; p = 0,78;
R² = 0,007; Figura 2).

252
Figura 1. Relação entre a abertura do dossel e a média de intensidade de vermelho na
asa anterior esquerda de machos da libélula C. scintilans em um riacho na reserva
florestal do km 41, Manaus, Brasil.
Figura 2. Relação entre a abertura do dossel e a densidade corporal de machos da
libélula C. scintilans em um riacho na reserva florestal do km 41, Manaus, Brasil.
6 8 10 12 14 16 18 20 22 24 26 28
Abertura do dossel (%)
90
100
110
120
130
140
150
160
170
180
190
Méd
ia d
e v
erm
elh
o (
pix
els
)
y = 133,718+0,469*x
6 8 10 12 14 16 18 20 22 24 26 28
Abertura do dossel (%)
90
100
110
120
130
140
150
160
170
180
190
Méd
ia d
e v
erm
elh
o (
pix
els
)
y = 133,718+0,469*x

253
Discussão
A ausência de relação entre a abertura do dossel e a média de vermelho nas asas e a
densidade do tórax indica que o tamanho da clareira não é um indicativo de um bom
território para C. scintilans. Sendo assim, machos de maior qualidade para brigas e
acasalamento, que possuem maior reserva energética (Contreras-Garduño et al. 2006) e
coloração mais conspícua nas asas (Schultz & Fincke 2009), não preferem clareiras
maiores.
O fato do tamanho da mancha de sol não estar associado à qualidade do macho
que a ocupa indica que territórios com maior intensidade luminosa não são melhores
para o encontro com fêmeas. Já foi observado que a presença de troncos caídos pode
explicar a distribuição dos machos na floresta, uma vez que esses troncos são sítios de
oviposição para as fêmeas e, provavelmente, são territórios defendidos pelos machos
(Resende 2002). Talvez, os machos de C. scintilans sejam capazes de entrar em
atividade sob menores temperaturas corporais e sejam capazes de manter a temperatura
corpórea ideal ao usar as pequenas faixas de sol que penetram nos territórios. O
investimento preferencial pode ser voltado para a proteção de territórios que
representem bons sítios de oviposição para fêmeas.
Vários animais determinam como melhores territórios aqueles que apresentam
uma maior quantidade de recursos disponíveis (Adams 2001). Nesse contexto, como
encontrei que a luminosidade não é um bom preditor de qualidade de território para
machos de C. scintilans, é possível que outros recursos, como sítios de oviposição,
sejam mais importantes para explicar a distribuição de machos na floresta (Arnott &
Elwood 2008). Dessa forma, é possível que a qualidade de territórios dessa espécie seja
determinada principalmente pela presença de sítios que apresentem condições

254
favoráveis para o desenvolvimento de suas larvas, mesmo que elas não sejam ideais
para os adultos.
Agradecimentos
De uma forma geral, agradeço ao José Luís, aos Paulinhos Mãe e Pai, aos monitores
Gloss e Barbudo, aos professores convidados e aos amigos do EFA 2012 por uma das
melhores experiências da minha vida, que não se limitou ao aperfeiçoamento do
raciocínio e escrita científica, mas também desencadeou uma mudança em minha
postura crítica no meio científico e social. Além disso, poucos tiveram ou terão a chance
de conhecer, estudar e pesquisar uma região com tamanha biodiversidade e beleza como
é a floresta amazônica.
Vou agradecer a algumas pessoas específicas (não que sejam mais especiais que
as outras). Dani, muito obrigado pela ajuda com quase tudo nesse curso. Isso vale para a
escrita de relatório, as minhas roupas que você lavou ou me ajudou por pena de mim e
às críticas negativas sobre as minhas habilidades para tarefas caseiras. ‘Anjo’ (quem
quer que você seja), muito obrigado pela bermuda, foi uma ótima ajuda. Mari, Renata e
Vidal, não me esquecerei da visita ao teatro e do açaí horrível que a gente tomou.
Paulinho e Cheyene, vocês se merecem. Seu Jorge e Dona Eduarda, valeu por tudo. Vou
sentir falta do café, almoço e jantar, e do suco de caju e goiabada. Agradeço também ao
professor convidado André por motivos burocráticos e ao meu orientador Marcus
Cianciaruso por ter me incentivado a me inscrever no EFA 2012.
Gostaria de terminar os agradecimentos com algumas besteiras que nós falamos
durante o curso: “Mhiau!” (to rindo agora dessa coisa ridícula), “Áveres”, “Vocês
querem Partoba?!”, “Isso aí ta ó... uma B%$!a!”, “Tinha dois pinguins” e “Pô, Idásio,
pega a sua cueca!”. Tenho certeza que nos encontraremos nos congressos da vida e

255
usaremos essas expressões novamente em conjunto. Um grande abraço para todos
vocês.
Referências
Adams, E.S. 2001. Approaches to the study of territory size and shape. Annual Review
of Ecology and Systematics, 32:277-303.
Arnott, G. & R.W. Elwood. 2008. Information gathering and decision making about
resource value in animal contests. Animal Behaviour, 76:529-542.
Bergman, M. & C. Wiklund. 2009. Visual mate detection and mate flight pursuit in
relation to sunspot size in a woodland territorial butterfly. Animal Behaviour,
78:17-23.
Contreras-Garduño, J., J. Canales-Lazcano & A. Córdoba-Aguilar. 2006. Wing
pigmentation, immune ability, fat reserves and territorial status in males of the
rubyspot damselfly, Hetaerina americana. Journal of Ethology, 24:165–173.
Córdoba-Aguilar, A. 2008. Dragonflies & damselflies: model organisms for ecological
and evolutionary research. Oxford: Oxford Biology.
Fitztephens, D.M. & T. Getty. 2000. Colour, fat and social status in male damselfly
Calopteryx maculate. Animal Behaviour, 60:451-455.
Marden, J.H. & J.K. Waage. 1990. Escalated damselfly territorial contests are energetic
wars of attrition. Animal Behaviour, 39:954-959.
Resende, D.C. 2002. Comportamento territorial de Chalcopteryx scintilans
(Odonata:Polythoridae). In: Curso de campo ecologia da floresta amazônica
(Zuanon, J. & E. Venticinque, eds.). Manaus: INPA/PDBFF.

256
Schultz, T.D. & O.M. Fincke. 2009. Structural colours create a flashing cue for sexual
recognition and male quality in a Neotropical giant damselfly. Functional
Ecology, 23:724-732.
Velde, L.V., C. Turlure & H.V. Dyck. 2011. Body temperature and territory selection
by males of the speckled wood butterfly (Pararge aegeria): what makes a forest
sunlit patch a rendezvous site? Ecological Entomology, 36:161-169.

257
Mutualismo de defesa entre plantas mirmecófitas e formigas reduz o investimento
em defesas estruturais em espécies de Melastomataceae
Joselândio Corrêa Santos
Introdução
A principal causa de perda de área foliar em plantas de ambientes tropicais é a
herbivoria por insetos (Coley & Barone 1996). Estima-se que esses artrópodes são
responsáveis por cerca de 70% da área foliar consumida anualmente, ocasionando um
forte impacto no sucesso reprodutivo e influenciando o padrão de alocação de recursos
em plantas (Herms & Mattson 1992). Desse modo, há uma constante pressão seletiva
sobre as plantas em favor de mecanismos que minimizem a herbivoria nas folhas.
Basicamente, as plantas utilizam três tipos de defesas contra herbívoros, as
químicas, as estruturais e as biológicas, que podem ocorrer simultaneamente em um
mesmo indivíduo (Coley & Barone 1996). As plantas defendidas quimicamente utilizam
subprodutos do seu metabolismo como substâncias deterrentes, inibidoras ou tóxicas
aos herbívoros, tornando o tecido foliar impalatável ou de baixa qualidade para
consumo. Já as defesas estruturais são aquelas em que ocorre a produção ou
enrijecimento de estruturas já existentes nas folhas, por meio do espessamento das
paredes celulares, do aumento da quantidade de fibras ou do acúmulo de cristais
minerais (e.g. oxalatos de cálcio) em tecido das folhas, aumentando assim a dureza
foliar (Johnson 2011). Os mecanismos de defesa biológica envolvem associações da
planta com outros organismos, tais como formigas, que ocasionam um impacto negativo
sobre as populações de herbívoros, e assim reduzem o dano causado por eles.
Dentro das defesas biológicas, a mais bem estudada é a interação tri-trófica entre
planta, herbívoro e formigas (Cornelissen & Fernandes 2001). Em muitas espécies de

258
plantas, existem estruturas que são utilizadas por formigas como sítios de forrageio e
abrigo (Fáveri et al. 2008). Plantas que estabelecem este tipo de associação são
chamadas de mirmecófitas (Ribeiro et al. 1999). A presença constante de formigas
sobre a planta diminui o consumo de suas estruturas pelos herbívoros, uma vez que eles
são utilizados como alimento pelas formigas (Fáveri et al. 2008).
A produção de defesas pelas plantas demanda recursos que poderiam ser
utilizados para outras funções, tais como a reprodução e crescimento (Herms & Mattson
1992). Entretanto, o risco de consumo constante pelos herbívoros a que as plantas estão
sujeitas torna necessário o investimento em defesas mesmo sem a presença do
herbívoro. Em situações que as plantas são protegidas por outros organismos de maneira
efetiva, como nas interações entre os predadores de herbívoros e as plantas, estas
deveriam diminuir o investimento em defesas químicas e estruturais e direcionar esses
recursos para outras funções (Cornelissen & Fernandes 2001).
Os gêneros Maieta e Tococa, ambos pertencentes à família Melastomataceae,
são exemplos de mirmecófitas que ocorrem na Amazônia Central (Ribeiro et al. 1999).
Além desses dois gêneros, a família Melastomataceae está representada na Amazônia
Central por outros dez gêneros, dos quais somente mais um (Cliedemia) possui
domácias, colonizadas por formigas, na base de suas folhas (Ribeiro et al. 1999).
Associações entre plantas e formigas apresentam diferenças no grau de proteção
conferida pelas formigas. Espécies de plantas que abrigam formigas do gênero Pheidole
(e.g. espécies do gênero Maieta) são mais bem protegidas que plantas defendidas por
outros gêneros de formigas. Isso ocorre porque o tempo de resposta de Pheidole à
presença do herbívoro é inferior ao tempo de resposta de espécies de outros gêneros.
Dessa forma, o objetivo desse estudo foi determinar se plantas com diferentes níveis de
proteção por formigas investem diferencialmente em outras defesas contra herbívoros.

259
Minha hipótese é que o investimento em defesas físicas deveria diminuir com o
aumento da eficiência da defesa biológica.
Métodos
Área de estudo
Realizei o estudo na Área de Relevante Interesse Ecológico Projeto Dinâmica Biológica
de Fragmentos Florestais (ARIE PDBFF), no sítio Km-41 que está localizado a 80 km
de Manaus – AM. A vegetação é caracterizada como floresta tropical úmida de terra-
firme e a região possui precipitação média anual de 2200 mm e temperatura média anual
de 26,7ºC, com duas estações bem definidas: de dezembro a junho (estação chuvosa) e
de julho a novembro (estação seca) (Radam-Brasil 1978).
Espécies focais
Para o estudo, utilizei quatro espécies pertencentes à família Melastomataceae: Maieta
sp. (n=10), Tococa sp. (n=10), Miconia sp. (n=10) e Henriettella sp. (n=10). Maieta sp.
possui domácias que são geralmente ocupadas por formigas do gênero Pheidole. Em
Tococa sp., a colonização das domácias é feita por formigas dos gêneros Azteca spp. e
Crematogaster spp. As plantas das espécies Miconia sp. e Henriettella sp. não possuem
domácias em suas folhas e também não há nenhuma evidência de que elas possuam
associação com formigas.
As espécies foram ordenadas quanto à presença ou ausência de defesa biológica
e quanto à eficiência dessa defesa. A eficiência da defesa biológica foi determinada por
meio de ensaios experimentais realizados em campo. Neste ensaio, utilizei cupins sobre
as folhas das plantas como forma de simular a presença de um herbívoro. Determinei se
o tempo necessário para acumular cinco formigas junto ao cupim diferia entre as

260
mirmecófitas. Repeti o ensaio nos 20 os indivíduos de mirmecófitas por três vezes,
sempre em ramos diferentes. Em Maieta sp. o tempo médio de resposta foi 67±16 s
(média ± desvio padrão), já em Tococa sp. o menor tempo foi 107 s. Somente em dois
indivíduos de Tococa sp. presenciei o ataque ao cupim durante os 3 min. de observação.
Massa específica e dureza foliar
Em cada planta, coletei três folhas, preferencialmente de ramos diferentes e com idades
semelhantes. Padronizei a idade das folhas amostrando sempre as que estavam no
terceiro nó mais recente do ramo. Todas as plantas se encontravam no sub-bosque de
uma floresta madura.
A constituição das folhas pode variar quanto à quantidade de cada substância
utilizada para a sua constituição (lignina, celulose, hemicelulose e fibra). A maneira
como estes compostos estão distribuídos nos tecidos foliares (parênquimas, nervuras,
epiderme) também pode variar e isso pode refletir no grau de dureza foliar. Dessa
forma, medidas da quantidade desses compostos alocados na folha fornecem estimativas
do investimento em defesas estruturais. Segundo Cornelissen et al. (2003), a massa
foliar específica (MFE) e a resistência à perfuração (FPF) estão associadas ao
investimento em defesa estrutural nas plantas.
Estimei a força necessária para perfurar o limbo foliar utilizando um
Perfurômetro. Esse aparelho consiste em uma haste de madeira que, em uma de suas
extremidades, possui um arame com 1,5 mm de diâmetro e na outra, um funil de papel,
sendo ambos fixados por fita adesiva. Para estimar a força necessária para romper a
folha, posicionei a extremidade com o arame do Perfurômetro sobre a folha mantendo o
ângulo reto entre eles. Depois, adicionei areia ao funil até o momento em que a
extremidade que estava sobre a folha atravessou o limbo foliar. Em seguida, pesei a

261
areia acumulada no funil em uma balança semianalítica e, ao valor da massa da areia no
funil, adicionei o valor da massa do Perfurômetro. Multipliquei o resultado dessa soma
pelo valor da aceleração da gravidade no local (aproximadamente 10m/s2). Desse modo,
obtive uma estimativa da força (em N) necessária para perfurar cada folha. Fiz três
repetições por folha e obtive uma média de FPF por planta.
Para o cálculo de Massa Foliar Específica (MFE), cortei um fragmento foliar
com 4 cm2 de área de cada folha e em seguida os desidratei até a estabilizar a massa.
Com uma balança semianalítica, obtive a massa dos fragmentos foliares. Calculei a
MFE dividindo a massa do fragmento foliar por sua área, obtendo assim o valor de
massa acumulada por unidade de área.
Análise estatística
Para avaliar se o os atributos foliares, MFE e FPF, diminuem com o aumento da
eficiência da defesa biológica, utilizei análise de variância de um fator (ANOVA-One
Way). Previ que as plantas sem domácias, Miconia sp. e Henriettella sp., teriam uma
maior MFE e FPF que Tococa sp., que por sua vez deveria ter maior MFE e FPF que
Maieta sp.
Resultados
A força necessária para perfurar a lâmina foliar foi maior nas espécies sem associação
com formigas do que nas plantas mirmecófitas (F=33,16; gl=3; p<0,001; Figura 1).
Entretanto, não houve diferença entre as espécies mirmecófitas.

262
Figura 4. Valores médios da força necessária para perfurar as folhas de espécies com
diferentes níveis de proteção por formigas. As espécies Miconia sp. e Henrietella sp.
não apresentam associação com formigas e Tococa sp. e Maieta sp. são mirmecófitas.
As barras representam o desvio padrão. F=33,16; gl=3; p<0,001; n=10.
A massa foliar específica diferiu entre as mirmecófitas e as espécies sem
associação com formigas (F=33,16; gl=3; p<0,001; Figura 2). As mirmecófitas tiveram
menor massa por unidade de área. Porém, assim como ocorreu com a FPF, a MFE não
diferiu entre as espécies desse grupo.

263
Figura 5: Valores médios de massa foliar específica de espécies com diferentes níveis
de proteção por formigas. As espécies Miconia sp. e Henrietella sp. não apresentam
associação com formigas e Tococa sp. e Maieta sp. são mirmecófitas. As barras
representam o desvio padrão. F=58,25; gl=3; p<0,001; n=10.
Discussão
A associação com formigas por parte das mirmecófitas resultou em um menor
investimento em defesas estruturais. Essa redução de investimento em defesas
estruturais pode permitir que as plantas invistam mais recursos em crescimento e
reprodução (Herms & Mattson 1992).
Ao contrário do esperado, mirmecófitas com níveis diferentes de proteção por
formigas não diferiram quanto ao investimento em defesas estruturais. Em algumas
espécies de plantas, existem diferenças de coloração entre folhas jovens, que são
avermelhadas, e as maduras, que são verdes. Dominy et al. (2002) sugere que a
coloração avermelhada em folhas jovens seja devida ao acúmulo de substâncias tóxicas
aos animais. As folhas da espécie de Tococa analisada apresentam na superfície abaxial

264
uma coloração arroxeada, o que pode sinalizar a presença desses compostos. Investir em
defesas químicas em vez de defesas físicas pode ser vantajoso, uma vez que os
compostos químicos podem ser recuperados das folhas durante o processo de abscisão
foliar (Cornelissen & Fernandes 2001). Compostos associados a defesa estrutural, por
outro lado, uma vez depositados não podem ser realocados para outro fim. Assim, é
possível que, em vez de investir em defesas estruturais, as plantas de Tococa sp.
invistam em defesas químicas contra herbívoros.
As plantas defendidas por formigas investem menos em defesas estruturais
contra os herbívoros. Em situações que a colonização das domácias por formigas não
ocorre, estas plantas estariam mais susceptíveis a um aumento no consumo pelos
herbívoros, o que poderia afetar diretamente a sua sobrevivência, crescimento e
reprodução. Logo, quanto maior eficiência em defesa biológica maior grau dependência
em relação proteção à das formigas.
Agradecimentos
Agradeço a equipe de organização do Curso de Ecologia da Floresta Amazônica – 2012
(EFA-2012), Paulo Mau, Paulo Rainbow, Thiago Beluga, Marcos Barbixa e ao Zé Luís,
pela grande oportunidade proporcionada por meio deste curso. Aos colegas, pelos bons
momentos e discussões que compartilhamos e por mostrar, que apesar das diferenças, é
possível ser uma turma e não somente uma “cambada”. Agradeço ainda a todos aqueles
que contribuíram e se empenharam de alguma forma em prol do nosso aprendizado, aos
professores temporários, D. Eduarda e o Sr. Jorge, motoristas e barqueiros. Ainda sou
muito grato ao PPG-ECO INPA e aos meus orientadores que me incentivaram a
participar do EFA-2012, Beto Vicentini, Zé Luís e Robyn Burnham. Agradeço aos

265
revisores anônimos (Marcos Barbixa e Ana Andrade) pelas sugestões, que com certeza
ajudaram-me a escrever com clareza e qualidade.
Referências
Coley, P.D., & J.A. Barone. 1996. Herbivory and plant defenses in tropical forests.
Annual Review of Ecology, Evolution and Systematics, 27:305–335.
Cornelissen, T.G., & G.W. Fernandes. 2001. Induced defences in the neotropical tree
Bauhinia brevipes (Vog.) to herbivory: effects of damage-induced changes on leaf
quality and insect attack. Trees, 15:236–241.
Cornelissen, J.H.C., S. Lavorel, E. Garnier, S. Díaz, N. Buchmann, D.E. Gurvich, P.B.
Reich, et al. 2003. A handbook of protocols for standardised and easy
measurement of plant functional traits worldwide. Australian Journal of Botany,
51:335-380.
Cornelissen, T.G., & Fernandes, G.W. 2001. Defence, growth and nutrient allocation in
the tropical shrub Bauhinia brevipes (Leguminosae). Austral Ecology, 26:246–253.
Dominy, N.J., P.W. Lucas, L.W. Ramsden, P. Riba-Hernandez, K.E. Stoner & I.M.
Turner. 2002. Why are young leaves red? Oikos, 98:63–176.
Fáveri, S.B., H.L. Vasconcelos & R. Dirzo. 2008. Effects of Amazonian forest
fragmentation on the interaction between plants, insect herbivores, and their natural
enemies. Journal of Tropical Ecology, 24:57–64.
Herms, D.A., & W.J. Mattson. 1992. The dilemma of the plants: to grow or defend. The
quarterly review of biology, 67:283–335.
Johnson, M.T.J. 2011. Evolutionary ecology of plant defences against herbivores.
Functional Ecology, 35:305–311.

266
Radam-Brasil.1978. Programa Nacional: Levantamento de Recursos Naturais (18th
ed.). Rio de Janeiro, Brasil: Ministério de Minas e Energia, Departamento Nacional
de Produção Mineral.
Ribeiro, J. E. L., M.J.G. Hopkins, A. Vicentini, C.A. Sothers, M.A.S. Costa, J.M. Brito,
M.A.D Souza. 1999. Flora da Reserva Ducke - guia de identificação das plantas
vasculares de uma floresta de terra-firme na Amazônia Central (1 ed). Manaus-
AM: Editora INPA. p. 816.

267
A condição alimentar das larvas de formigas-leão influencia no tamanho e no
tempo de construção do funil de areia?
Leonardo Queiroz de Oliveira
Introdução
Os organismos utilizam os recursos alimentares para realizarem funções vitais e
comportamentais, como a manutenção do metabolismo, crescimento, forrageio,
construção de abrigos e armadilhas (Reynolds 1990). Cada uma dessas funções
possuem diferentes níveis de investimento energético, que estão relacionados com a
quantidade de recursos alimentares que o indivíduo consome (Reynolds et al. 1985).
Por exemplo, a quantidade de energia utilizada para a manutenção do metabolismo em
atividade é pequena quando comparado com a quantidade de energia utilizada para o
crescimento do indivíduo e da prole.
Nas populações é comum que existam indivíduos com diferentes condições
nutricionais, onde os mais alimentados serão provavelmente maiores e irão reproduzir
com maior frequência (Gribbin & Thompson 1990). Esses indivíduos provavelmente
terão mais energia para se deslocar no ambiente e procurar sítios com maior
disponibilidade de recursos.
Predadores que utilizam armadilhas para captura de presas normalmente
investem muita energia na construção e manutenção das estruturas da armadilha (Lucas
1985). Assim, é provável que indivíduos em melhores condições nutricionais invistam
mais energia na construção de armadilhas maiores, podendo aumentar a eficiência na
captura de presas.
As larvas das formigas-leão Myrmeleon brasiliensis (Neuroptera:
Myrmeleontidae) são predadoras que permanecem enterradas no fundo de funis cônicos

268
de areia a espera de pequenos insetos de solo que eventualmente caem na armadilha
(Penny & Arias 1982). Considerando que as larvas de formigas-leão investem energia
na construção dos funis, o objetivo desse trabalho foi avaliar se indivíduos de M.
brasiliensis sobre diferentes condições alimentares constroem funis de tamanhos
distintos e em velocidades diferentes. Espero que larvas com melhores condições
alimentares façam funis de maior diâmetro e demorem mais tempo para iniciar a
construção do funil, dado que possuem energia extra para buscar sítios mais adequados
para o estabelecimento dos funis e para construir armadilhas maiores.
Métodos
Realizei o estudo na Área de Relevante Interesse Ecológico do Projeto Dinâmica
Biológica de Fragmentos Florestais (ARIE PDBFF) no sítio km 41, localizado a 80 km
ao norte de Manaus, AM - Brasil. No acampamento da reserva selecionei 40 indivíduos
de formigas-leão onde se encontra uma alta concentração de larvas, principalmente nos
terrenos secos e arenosos. Ainda no solo, dividi as larvas selecionadas em dois
tratamentos, cada tratamento contendo 20 indivíduos. No primeiro tratamento alimentei
as larvas com dois cupins. A primeira alimentação aconteceu na noite anterior ao
experimento e a segunda alimentação aconteceu uma hora antes do início do
experimento. No segundo tratamento não alimentei as larvas selecionadas por um
período de 6 horas antes do início do tratamento.
No dia seguinte coletei do solo a porção de areia de cada funil selecionado
juntamente com sua respectiva larva. A porção de areia foi peneirada e a larva de
formiga leão coletada e separada de acordo com o tratamento que cada larva recebeu.
Preparei quatro bandejas com areia seca e peneirada, sendo duas bandejas para o
tratamento alimentado e as outras duas bandejas para o tratamento sem alimentação.

269
Assim, para cada tratamento havia duas bandejas com 10 indivíduos em cada.
Fotografei as bandejas de areia em intervalos de 30 min para verificar o início de
formação dos funis das formigas-leão. Numerei os funis em ordem crescente à medida
que se formavam para determinar em qual tempo aquele indivíduo iniciou a construção
do funil de areia. O experimento se estendeu por 07:00 h até que não houvesse mais
mudanças de posição ou expansão da área do funil o que indicava que a construção dos
funis estava concluída.
Para quantificar os parâmetros de interesse (tamanho e tempo de construção do
funil), medi o diâmetro dos funis com um paquímetro e removi as larvas do
experimento. Para realizar as medidas de tamanho das larvas, fotografei cada indivíduo
e medi a distância entre os olhos com o auxílio do software Image J. Utilizei a distância
entre os olhos como medida de tamanho porque o comprimento do abdômen dos
animais alimentados estava maior devido à alimentação extra que receberam. Para
determinar se existiu diferença entre os tratamentos (alimentados e não alimentados)
para cada variável dependente (tamanho do funil e tempo de formação do funil) utilizei
uma ANCOVA, considerando a distância entre os olhos como covariável.
Resultados
O diâmetro dos funis variou de 1,0 a 5,7 cm, enquanto o tempo de formação dos funis
variou de 90 a 300 min. As formigas-leão com alimentação extra e sem alimentação não
construíram funis de tamanhos diferentes (F(1, 34) = 0,24; p = 0,62; Figura 1). Da mesma
forma, o tempo de formação dos funis não foi diferente entre os tratamentos testados
(F(1, 34) = 2,34; p = 0,13; Figura 2).

Figura 1. Tamanho médio do funil feito por formigas-leão nos tratamentos alimentado e
não alimentado. As barras representam os intervalos de confiança de 0,95.
Figura 2. Tempo médio de formação do funil feito por formigas-leão nos tratamentos
alimentado e não alimentado. As barras representam os intervalos de confiança de 0,95.
Discussão
As larvas de formigas-leão com alimentação extra não construíram funis de areia
maiores e não demoraram mais tempo na construção da armadilha. Dessa forma, os

271
resultados sugerem que a quantidade extra de alimento fornecida para as larvas de
formigas-leão não foi convertida em energia extra para a construção de funis maiores ou
para o deslocamento a procura de locais mais favoráveis, indicando que essa energia
provavelmente foi acumulada.
Evolutivamente, a divisão do ciclo de vida em diferentes estágios de
desenvolvimento, passando por ovo, larva e adulto está relacionado com o maior
acúmulo de reservas energéticas nos estágios larvais, que irão refletir em adultos mais
vigorosos (Werner 1986). A construção do funil de fato requer um alto investimento
energético das formigas-leão (Lucas 1985). Portanto, é provável que as larvas não
gastem mais energia aumentando o tamanho da armadilha, uma vez que o objetivo é
acumular reservas energéticas para o estágio adulto (Werner 1986). Além disso, a
construção de funis de areia muito grandes podem resultar na captura de animais
maiores, como formigas solitárias, que ao invés de servirem de alimento para as larvas,
podem atacá-las (observação pessoal). Da mesma forma, animais grandes podem
danificar a estrutura do funil o que resulta em mais gasto energético para a reconstrução.
É provável que exista um tamanho ótimo para o funil de areia, proporcional ao
tamanho da larva, onde os custos energéticos relacionados com a construção não
superem os ganhos relacionados com a alimentação (Kramer 2001). Assim, indivíduos
em melhores condições alimentares provavelmente manterão os funis no tamanho
ótimo, mas gastarão proporcionalmente menos energia na construção do funil,
resultando em maior acúmulo de reservas. Por outro lado, os indivíduos que estão com
déficit alimentar provavelmente vão alocar proporcionalmente mais energia para a
manutenção do tamanho ótimo do funil. Entretanto, a energia resultante para o acúmulo
de reservas será menor do que naquelas larvas em melhores condições alimentares.

272
O fato das larvas de formigas-leão que receberam alimentação extra não
investirem mais tempo a procura de locais para construção de suas armadilhas, pode
indicar que não é vantajoso aumentar o tempo de deslocamento. Considerando que a
única maneira das larvas acessarem os recursos alimentares é por meio da construção
dos funis de areia (Crowley & Linton 1999) o aumento no tempo para iniciar a
construção do funil pode implicar em redução da captura de alimento. Além disso, o
aumento do tempo para iniciar a construção, está relacionado com maior tempo de
deslocamento. Esse fato pode aumentar o risco do indivíduo ser canibalizado (Lucas
1989; Matsura & Takano 1989), ou de ser detectado por outros predadores (Best &
Gennaro 1985). Dessa forma, é provável que as larvas não invistam muita energia para
o deslocamento, pois o risco de predação e a redução no acúmulo de reservas podem
comprometer o desenvolvimento e sobrevivência dos indivíduos.
Agradecimentos
Agradeço ao professor Paulo Bobrowiek e Paulo Enrique Cardoso Peixoto pelo auxílio
durante todas as fases do projeto. Agradeço ao Raimundo de Souza (Seu Jorge), a
Eduarda Costa pelo auxílio durante a preparação do experimento e aos demais colegas
pelas sugestões construtivas durante o decorrer do trabalho.
Referências
Best, T.L & A.L. Gennaro. 1985. Food habits of the western whiptail lizard
(Cnemidophorus tigris) in south-eastern New Mexico. Great Basin Naturalist,
45:527-534.
Craig, C.L. 1987. The ecological and evolutionary interdependence between web
architecture and web silk spun by orb web weaving spider. Biological Journal

273
of the Linnean Society, 30:135-162.
Crowley, P.H. & M.C. Linton. 1999. Antlion foraging: tracking prey across space and
time. Ecology, 80:2271-2282
Gribbin, S. D. & Thompson, D. J. 1990. Asymmetric intraspecific competition among
larvae of the damselfly Ischnura elegans (Zygoptera: Coenagrionidae).
Ecological Entomology, 15:37-42.
Kramer, D. L. 2001. Foragin behavior, pp. 232-246. In: Evolutionary Ecology Concepts
and Case Studies (C. W. Fox, D. A. Roff, D. J. Fairbairn, eds.). Oxford: Oxford
University Press.
Lucas, J. R. 1985. Metabolic rates and pit-construction costs of two antlion species.
Journal of Animal Ecology, 54:295-309.
Lucas, J. R. 1989. Differences in habitat use between two pit building antlion species:
causes and consequences. American Midland Naturalist, 121:84-98.
Matsura, T & H. Takano. 1989. Pitt-relocation of antlion larvae in relation to their
density. Researches on population Ecology, 31:225-234.
Reynolds, S.E., S.F. Nottingham & A.E. Stephens. 1985. Food and water economy and
Its relation to growth in fifth-instar larvae of the tobacco hornworm, Manduca
sexta. Journal of Insect Physiology, 31:119-127.
Reynolds, S.E. 1990. Feeding in caterpillars: maximizing or optimizing food
acquisition?, pp106-118. In: Animal Nutrition and Transport Processes (J.
Mellinger, eds.). Basel: Karger.
Penny, N. D. & J. Arias. 1982. Insects of an Amazon forest. Columbia University Press,
New York.
Werner, E. E & J. F. Gilliam. 1984. The ontogenetic niche and species interactions in
size-structured populations. Annual Review of Ecology and Systematics, 15:393-

274
425.
Werner, E. E. 1986. Amphibian metamorphosis, growth rate, predation risk and the
optimal size at transformation. American Naturalist, 128:319-341.

275
Relação entre tamanho corporal, respostas de defesa e posição na teia de
indivíduos de uma espécie de aranha (Pholcidae) que vive em agregações na
Amazônia Central
Lorena Costa Pinto
Introdução
Em várias populações naturais os indivíduos interagem recorrentemente com seus co-
específicos, especialmente indivíduos vizinhos ou parentais, no tempo e no espaço,
formando grupos sociais (Mazer & Damuth 2001). O comportamento social dentro
desses grupos pode variar desde cooperação e altruísmo até competição e disputa por
recursos entre os organismos (Stevenson 2006). Da mesma forma, existem diferentes
níveis de organização, como os sistemas eussociais, que consistem na associação de
organismos aparentados que convivem em cooperação e possuem divisões de tarefas
(Wilson & Holldobler 2005) ou sistemas de agregados de organismos não aparentados,
que geralmente não apresentam divisões de tarefas e nos quais os indivíduos podem
competir intensamente por espaço e recursos.
Uma das principais explicações para a ocorrência de agregações em organismos
não aparentados é a busca por proteção contra inimigos naturais. A teoria do rebanho
egoísta, proposta por Hamilton (1971), postula que os animais em grupos sociais podem
diminuir o risco individual de predação ao ocorrerem em agregados. No entanto, dentro
dos grupos os indivíduos que se estabelecerem no centro do grupo, deslocando seus co-
específicos para periferia, podem reduzir ainda mais o risco de predação. Dessa forma,
os indivíduos centrais, em geral os mais fortes, são mais beneficiados pela proteção
conferida pelo grupo quando comparado aos indivíduos da periferia (Gross &
MacMillan 1981).

276
Em aranhas construtoras de teias, o comportamento de agregação entre co-
específicos não aparentados pode ser uma estratégia selecionada para minimizar os
riscos individuais de predação ou parasitismo (Gonzaga 2007). No entanto, mesmo
agregadas, essas aranhas podem adotar estratégias adicionais de defesa. Dentre elas,
existe o abandono da teia, que consiste em atirar-se no solo ou na vegetação em
situações de risco (Blackledge & Pickett 2000) e o comportamento de vibrar o corpo
por alguns minutos ou até por alguns dias (restrito às famílias Pholcidae e Araneidae), o
que dificulta a identificação e captura da aranha na teia pelo predador (Jackson et al.
1993, Heuts et al. 2001). O comportamento de vibrar o corpo, em particular, pode ser
utilizado como uma resposta rápida de defesa, mas como essa estratégia não envolve
saída da teia, os riscos de predação e parasitismo do indivíduo podem ser altos. Já a
fuga da teia pode gerar custos distintos para os indivíduos, dependendo de sua posição
no grupo. Nas aranhas que ocupam o centro da teia, o abandono poderia acarretar na
perda do local seguro contra a predação, que poderia ser ocupado por outro indivíduo.
Já para as aranhas da periferia, a saída da teia seria a resposta mais esperada, pois esses
indivíduos não possuem posições fixas no grupo e podem retornar facilmente para a teia
após a ameaça.
Em uma espécie de aranha pertencente à família Pholcidae, que ocorre na
Amazônia central, os indivíduos podem ocorrer em agregações com co-específicos em
uma mesma teia ou em teias conectadas. As teias dessa espécie têm formato
tridimensional, com superfície arredondada, convexa e com o centro da teia localizado
entre os fios de sustentação. Devido a essas características, o centro da teia pode ser o
local mais seguro contra predadores e parasitas, de modo que os indivíduos maiores
podem se comportar de acordo com o postulado pela teoria do rebanho egoísta
(Hamilton 1971), dominando essa região e deslocando os menores para a periferia da

277
teia. Uma vez que os indivíduos da periferia estariam mais suscetíveis aos riscos de
predação e parasitismo, é possível que as estratégias de defesa adotadas por esses
indivíduos incluam respostas rápidas e diferentes das apresentadas pelos indivíduos do
centro da teia. Assim, meu objetivo foi avaliar se o tamanho dos indivíduos determina
sua posição espacial na teia e se elas respondem de forma diferenciada a um estímulo de
predação/parasitismo de acordo com sua posição. Minhas hipóteses são que (a) os
indivíduos localizados no centro da teia serão maiores, (b) os indivíduos localizados na
periferia da teia vão responder mais rapidamente ao estímulo de predação quando
comparado ao indivíduo central e (c) aranhas localizadas na periferia da teia terão maior
propensão a abandoná-la quando comparadas às aranhas do centro. Para essas hipóteses
minhas previsões são que (a) aranhas do centro da teia apresentarão maior largura do
cefalotórax que seus pares na periferia, (b) aranhas centrais precisarão de uma maior
quantidade de estímulos que as periféricas para apresentar alguma resposta de defesa e
(c) aranhas localizadas na periferia da teia apresentarão maior frequência de abandono,
enquanto aranhas do centro apresentarão maior freqüência do comportamento de vibrar
o corpo sem abandonar a teia.
Métodos
Área e objeto de estudo
Realizei o estudo ao longo de três riachos (localmente conhecidos como igarapés) na
reserva do km 41, localizada a 80 km ao norte de Manaus, Brasil (02°25′ S - 59°43′ O).
Essa reserva é composta por uma floresta tropical de terra firme e faz parte de uma Área
de Relevante Interesse Ecológico – ARIE, sob co-gestão do Projeto Dinâmica Biológica
de Fragmentos Florestais – PDBFF (Gascon & Bierregaard 2001).

278
Nos igarapés procurei por 30 teias com agregações de uma espécie de aranha
(Pholcidae), com cefalotórax e abdômen vermelhos e pernas pretas com marcas
vermelhas. Os indivíduos dessa espécie podem ocorrer solitários ou em agregados que
podem variar de dois a nove indivíduos (obs. pess.).
Simulação de predação/parasitismo e medidas de tamanho corporal
No intuito de obter as respostas de defesa apresentada pelos indivíduos em diferentes
posições na teia, realizei simulações de estímulos de predação ou parasitismo em duas
aranhas de cada teia. Para isso, identifiquei o indivíduo mais próximo e mais distante do
centro da teia e simulei a presença de um predador/parasita tocando cada indivíduo na
superfície dorsal, como um espeto de 24 cm de comprimento e 3 mm de largura por
baixo da teia. Para minimizar o efeito da ordem de manipulação da teia nas respostas
dos indivíduos, eu alternei as simulações entre os indivíduos do centro e da periferia
entre as teias. Em seguida, contabilizei o número de toques necessários para cada aranha
apresentar uma resposta de defesa. Considerei como respostas o abandono da teia ou a
vibração corporal.
Para verificar se havia diferença de tamanho entre os indivíduos do centro e da
periferia da teia, coletei os dois indivíduos testados após a simulação, para a medição do
cefalotórax em laboratório. Para isso, fotografei cada indivíduo e calculei a largura do
cefalotórax com o programa Image Tool (Versão 3.0, UTHSCSA).
Análises
Utilizei um teste t pareado para analisar se existia diferença na largura do cefalotórax
entre aranhas do centro e da periferia da teia. Para testar se a quantidade de estímulos
necessários para uma resposta de defesa era diferente entre aranhas do centro e da

279
periferia da teia, usei um modelo linear generalizado com dispersão de erros de Poisson.
Para isso, usei a posição na teia (centro ou periferia) como variável preditora, número de
estímulos (toques) como variável resposta e cada teia como bloco. Como o tamanho
pode influenciar nas taxas de predação e parasitismo (Rayor & Uetz 1993) e,
consequentemente, nas respostas apresentadas pelas aranhas, utilizei a largura do
cefalotórax como co-variável para controlar esse efeito. Para testar se aranhas da
periferia e do centro da teia apresentavam respostas de defesa diferentes (vibração do
corpo ou fuga da teia) utilizei um teste qui-quadrado de McNemar, que considera o
pareamento entre as aranhas da periferia e do centro de cada teia.
Resultados
As aranhas localizadas no centro das teias eram maiores que os indivíduos da periferia,
com cefalotórax 20% mais largo (t = 2,66; gl = 29; p < 0,01; Figura1). A largura média
do cefalotórax dos indivíduos do centro da teia foi de 2,1 mm (DP = 0,40 mm),
enquanto a largura média do cefalotórax dos indivíduos da periferia foi de 1,84 mm
(DP=0,39 mm).

280
Figura 1. Relação entre a largura do cefalotórax e a posição dos indivíduos na teia de
uma espécie de aranha (Pholcidae), na reserva do Km 41-PDBFF, Amazonas, Brasil.
Cada linha conecta o valor de largura do cefalotórax do indivíduo do centro e seu par da
periferia de uma mesma teia.
As aranhas do centro e da periferia da teia apresentaram diferenças quanto ao
número de toques necessários para responder a simulação do predador/parasita
(χ2=14,4; gl=1; p<0,01; Figura 2). A diferença entre as médias de estímulos entre as
aranhas do centro e da periferia foi de 1,83 toques. As aranhas do centro responderam
em média com 4,5 toques (DP=5,3) e as aranhas periféricas com 2,6 toques (DP=2,1).

281
Figura 2. Relação entre o número de estímulos necessários para exibição de resposta de
defesa (fuga da teia ou vibração corporal) e a posição dos indivíduos na teia de uma
espécie de aranha (Pholcidae), na reserva do Km 41-PDBFF, Amazonas, Brasil. Cada
linha conecta o valor do número de estímulos do indivíduo do centro e da periferia de
uma mesma teia.
As aranhas do centro e da periferia da teia apresentaram frequências similares no
tipo de resposta de defesa frente ao estímulo de predação/parasitismo (χ2=1,23; gl=1;
p=0,27). O comportamento de vibrar o corpo foi mais comum tanto em aranhas do
centro da teia (76,7% das simulações), quanto nas aranhas da periferia (60% das
simulações).
Discussão
Conforme o esperado, houve diferença entre os tamanhos das aranhas centrais e
periféricas, indicando que quando agregadas, as aranhas apresentam segregação
espacial, com os indivíduos maiores localizados no centro da teia. Além disso, a
quantidade de estímulos de predação necessários para incitar uma resposta também

282
diferiu de acordo com a posição na teia. O menor número de estímulos necessários para
desencadear uma resposta de defesa nas aranhas da periferia pode ter ocorrido devido à
maior vulnerabilidade desses indivíduos à predação e ao parasitismo. No entanto, o tipo
de resposta de defesa dos indivíduos foi semelhante. A preferência pelo comportamento
de vibrar o corpo pode indicar que o abandono da teia é arriscado para todas as aranhas,
independente de sua posição na agregação.
A diferença no tamanho das aranhas de acordo com a posição na teia indica uma
relação de dominância entre esses indivíduos (e.g. Rayor & Uetz 1990). Permanecer na
parte central do grupo pode ser uma vantagem devido ao menor risco de predação (e.g.
Alves-Costa & Gonzaga 2001). Eventualmente, o indivíduo dominante pode defender o
centro da teia contra indivíduos vizinhos e deslocar os menores e mais fracos para a
periferia. Alternativamente, é possível que indivíduos da periferia sejam aranhas que se
estabeleceram depois que a primeira fundou sua teia. No entanto, uma vez que mesmo
aranhas grandes podem ser predadas na periferia (Rayor & Uetz 1993), é provável que
as grandes optem por se localizar nas áreas centrais.
A posição da aranha na teia também teve influência na quantidade de estímulos
necessários para as aranhas apresentarem uma resposta de defesa. A maior propensão
dos indivíduos da periferia a responder aos estímulos pode ocorrer pelo fato dessa área
ser mais suscetível aos ataques de predadores e parasitas. Se esses indivíduos estão
constantemente em contato com diferentes situações de risco, uma resposta rápida para
minimizar os danos causados pelos inimigos naturais seria favorecida (Rayor & Uetz
1993). Em contrapartida, as aranhas centrais por serem menos suscetíveis a predação ou
parasitismo podem desencadear respostas apenas sob estímulos mais intensos.
Ao contrário do esperado, tanto as aranhas centrais quanto as aranhas periféricas
apresentaram preferencialmente o comportamento de vibrar o corpo como primeira

283
resposta de defesa. Por mais que essa estratégia demande um alto custo energético, pois
só é eficiente se o indivíduo conservar a vibração corporal até eliminar o estímulo
responsável pela resposta (Heuts 2001), o abandono da teia pode ocasionar maiores
riscos de predação (Gonzaga 2007). Assim, é possível que o comportamento de
abandono da teia seja utilizado apenas como uma última resposta, quando as outras
estratégias, como vibração corporal, não forem suficientes para afastar os predadores e
parasitas.
A diferença de tamanho, associada com a posição da aranha na teia, sugere que a
teoria do rebanho egoísta (Hamilton 1971) se aplica a essa espécie. O beneficio obtido
pela posição central na teia deve ocasionar disputas entre os indivíduos do grupo, de
modo que o indivíduo central é mais favorecido. No entanto, a ocorrência de um grande
número de agregados indica que os indivíduos da periferia também são beneficiados de
alguma forma. Talvez mesmo na periferia o risco de predação seria ainda menor quando
comparado com indivíduos solitários. Por fim, se o risco de predação/parasitismo de
fato afeta a propensão de uma aranha emitir uma resposta de defesa, é possível que os
indivíduos dessa espécie, quando estão solitários na teia, respondam a predação de
maneira semelhante aos indivíduos da periferia do agregado, pois estão frequentemente
mais sujeitos a situações de risco.
Agradecimentos
Agradeço aos predadores, Tatu e Hobbit, pela ajuda imprescindível nas coletas! Sem
vocês eu ficaria mais uns três dias andando sozinha por aí!! E ao Harry Potter, que com
sua mágica resolveu todos os problemas relativos às análises deste trabalho! Agradeço
também aos revisores Marcel (Baby) e Paulinho Rainbow por toda a paciência e pelo
direcionamento neste trabalho! Com relação ao EFA ... nossa é difícil escrever em

284
poucas palavras como essa experiência foi importante na minha vida profissional e
pessoal. Agradeço de coração a todos os organizadores e professores que passaram por
esse curso, especialmente ao Paulinho Morcego e Paulinho Rainbow pela oportunidade
de vivenciar o mundo paralelo do EFA. (obs: Você é incrível dona bicha! Obrigada por
ser essa luz de alegria nesse curso!). A dona Eduarda e seu Jorge, que com sua comida
com gostinho de casa da mamãe, fizeram nossa vida mais feliz! E a todos os colegas do
curso, especialmente a Carol (Da SILVA), parceira dos tráficos de cerveja e farra de
todos os dias, Dani (ATOLADINHA) que com seus projetos artísticos deixou o clima
do curso sempre descontraído, Tati (Árveres) que com seu jeito arretado de
pernambucana deixava sempre as discussões mais quentes e divertidas e a Cibele
(Lohaynne, Shayenne, Vanessão, Condessa ...) que toda atrapalhada e falante alegrou o
ambiente de uma forma incomparável. Aos que eu não detalhei, fica aqui meu carinho e
saudades por tudo que nós vivenciamos nesse um mês de casamento forçado! Obrigada
por fazerem desse mês o período mais incrível da minha vida! Valeu galera! Amo
vocês! Ôpa ... eu falei ou pensei!
Referências
Alves-Costa, C.P. & M.O. Gonzaga, 2001. Prey capture and spatial distribution of
Philoponella vittata (Araneae, Uloboridae) in host webs. Ethology, Ecology and
Evolution, 13:239-246.
Blackledge, T.A. & K.M. Pickett. 2000. Predatory interactions between mud-dauber
wasps (Hymenoptera, Sphecidae) and Argiope (Aranae, Araneidae) in captivity.
Journal of Arachnology, 28:211-216.
Gascon, C. & R.O. Bierregaard. 2001. The biological dynamics of forest fragments
project, pp. 31–42. In: Lessons from Amazonia, the ecology and conservation of a

285
fragmented forest (R. O. Bierregaard, C. Gascon, T.E. Lovejoy & R.C.G.
Mesquita, eds.). Yale University Press: New Haven.
Gonzaga, O.G. 2007. Inimigos naturais e defesas contra predação e parasitismo em
aranhas, pp. 209-237. In: Ecologia e comportamento de aranhas (O.G. Gonzaga,
A.J. Santos & H.F. Japyassú, Eds.). Rio de Janeiro: Interciência.
Gross, M.R. & A.M. MacMillan. 1981. Predation and the evolution of colonial nesting
in bluegill sunfish (Lepomis macrochirus). Behavioral Ecology and Sociobiology,
8:163-174.
Hamilton, W.D. 1971. Geometry for the selfish herd. Journal of Theory Biology,
31:295-311.
Heuts, B.A., M. Witteveldt, L.M. Dionisio Pires & F. van Wageningen. 2001. Long-
duration whirling of Pholcus phalangioides (Aranae, Pholcidae) is specifically
elicited by salticid spiders. Behavioral Proceedings, 55:27-34.
Jackson, R.R., E.M. Jakob, M.B. Willey, G.E. Campbell. 1993. Anti-predator defence
of a web-building spider Holocnemus pluchei (Aranae, Pholcidae). Journal of
Zoology, 229: 347-352.
Mazer, S.J. & J. Damuth. 2001. Evolutionary significance of variation, pp.16-26. In:
Evolutionary ecology: concepts and case studies (C.W. Fox, D.A. Roff & D.J.
Fairbairn, eds.). Oxford: Oxford University Press.
Rayor, L.S. & G.W. Uetz. 1990. Trade-offs in foraging success and predation risk with
spatial position in colonial spiders. Behavioral Ecology and Sociobiology, 27:77-
85.
Rayor, L.S. & G.W. Uetz. 1993. Ontogenetic shifts within the selfish herd: predation
risk and foraging trade-offs change with age in colonial web-building spiders.
Oecologia, 95:1- 8.

286
Stevenson, R.L. 2006. Doing adaptive things, pp. 75-85. In: Discovering evolutionary
ecology: bringing together ecology and evolution (P.J. Mayhew, ed.). Oxford:
Oxford University Press.
Wilson, E.O. & B.H. Holldobler. 2005. Eusociality: origin and consequences. PNAS,
20:13367–13371.

287
A influência da agregação espacial de Maieta guianensis (Melastomataceae) sobre
a intensidade da herbivoria
Ludmila Rattis
Introdução
Os padrões de distribuição espacial dos indivíduos podem ser decorrentes de diversos
processos que afetam a reprodução, a sobrevivência e a movimentação. Existem três
principais padrões de distribuição: aleatório, quando a probabilidade de um organismo
ocupar qualquer ponto no espaço é igual; uniforme, quando os organismos se distribuem
de modo a estabelecer intervalos regulares entre si, e o padrão agregado, quando os
indivíduos tendem a ocorrer em grupos (Begon et al. 2006). Existe uma variação no
nível de isolamento dos indivíduos, tanto na distribuição aleatória quanto na
distribuição agregada.
O nível de isolamento entre os indivíduos pode trazer consequências positivas e
negativas como resultado de interações intra e interespecíficas. A competição por
recursos pode ser maior quando os indivíduos ocorrem em agrupamentos,
principalmente para organismos sésseis, em que a interferência direta entre indivíduos é
mais provável (Begon et al. 2006). Para algumas espécies, o agrupamento pode resultar
em menor pressão por predação, pela diluição do efeito entre indivíduos (Hamilton
1971), como ocorre em agrupamentos de ungulados ou de algumas aves florestais (Holt
2006). Existem também consequências do nível de isolamento mediadas por interações
mutualísticas. Plantas e seus polinizadores e dispersores podem se beneficiar quando as
plantas se estabelecem agrupadas. Nessa situação, polinizadores e dispersores gastariam
menos energia se deslocando entre as plantas (Schoonhoven et al. 2005). Presumindo
que os polinizadores e dispersores sejam capazes de optar por forrageamento

288
preferencial, plantas ocorrendo em agrupamentos deveriam se beneficiar de serviços de
polinização e dispersão mais eficientes que plantas ocorrendo isoladas. Por outro lado, é
possível que as consequências do nível de isolamento sejam mediadas por outros tipos
de interações mutualísticas.
Mutualismos de defesa contra herbivoria em plantas, geralmente envolvem a
associação entre a planta e o predador capaz de se alimentar dos herbívoros (Schaller
2008). A interação entre a planta Maieta guianensis (Melastomataceae) e a formiga
Pheidole minutula (Hymenoptera: Formicidae) configura um exemplo de mutualismo
de defesa. A formiga se alimenta principalmente de insetos capturados sobre as folhas
de M. guianensis e da seiva do floema liberada por coccídeos mantidos no interior de
estruturas especializadas, denominadas domáceas. As domáceas são estruturas
localizadas nas bases das folhas de M. guianensis e são usadas para o estabelecimento
de colônias das formigas (Vasconcelos 1991; Agrawal & Rutter 1998). A formiga P.
minutula por sua vez, oferece um sistema de defesa para a planta, através de defesa
induzida contra herbívoros. Quando a planta sofre herbivoria, compostos secundários
voláteis são liberados pela planta. Esses compostos são percebidos pelas formigas, que
se deslocam para o local para atacar o possível herbívoro (Capurucho, 2010). Em
situações em que os indivíduos ocorrem em agrupamentos, os compostos liberados pelo
dano causado a uma planta deveriam produzir respostas de defesa por parte das
formigas associadas a indivíduos vizinhos.
Nesse estudo, investiguei a hipótese de que indivíduos de P. minutula
associadas a indivíduos de M. guianensis são capazes de responder ao dano provocado
por herbívoros em indivíduos vizinhos de M. guianensis. Se a hipótese for verdadeira,
espero que 1) aumente o número de formigas ocorrendo em determinada folha, quando
a folha de uma planta vizinha sofrer o dano; 2) que a probabilidade de recrutamento

289
diminua com o aumento da distância entre folhas e 3) que plantas estabelecidas a menos
de 1 m de outra tenham menor porcentagem de área foliar consumida que plantas
estabelecidas a mais de 1 m.
Métodos
Área de estudo
Realizei este estudo na Área de Relevante Interesse Ecológico (ARIE) do Km 41 (2°24’
S-59°44’ O) pertencente ao Projeto Dinâmica Biológica de Fragmentos Florestais
(PDBFF-INPA). Essa área está localizada no município de Rio Preto da Eva, a cerca de
80 km ao norte de Manaus, AM, Brasil (Dobrovolski, 2010).
Sistema de estudo
Na área de estudo, ocorrem indivíduos de M. guianensis próximo ao riacho do interior
da floresta. Considerei agrupados os indivíduos estabelecidos a menos de 1 m de
distância de outro indivíduo de M. guianensis. Descartei observações feitas em folhas
cujas domáceas estavam colonizadas por formigas da espécie Crematogaster sp. que
apresentam comportamento distinto de P. minutula.
Recrutamento na planta vizinha e distância de resposta
Para avaliar se as formigas respondiam ao dano provocado na folha da planta
vizinha, fiz um experimento utilizando pares de indivíduos agrupados de M. guianensis.
Registrei o número de formigas P. minutula sobre uma folha de um dos indivíduos
(folha focal). Após observá-la durante um minuto cortei a porção distal da folha de um
vizinho mais próximo com uma tesoura e registrei o número de indivíduos presentes na
folha durante um minuto. Observei um par por agrupamento em 29 agrupamentos
distintos. Para avaliar se havia diferença no número de formigas sobre a folha focal

290
antes e depois da aplicação do dano na folha vizinha, utilizei um teste t pareado. Para
avaliar se havia relação entre a probabilidade de recrutamento das formigas e a distância
entre as folhas do par, utilizei uma regressão logística entre a ocorrência de
recrutamento (variável resposta) e a distância entre as folhas (variável preditora).
Nível de isolamento e área foliar consumida
Para avaliar se havia diferenças no ataque de herbívoros entre as plantas agrupadas e
não agrupadas contei o número total de folhas de 11 indivíduos agrupados e 11
indivíduos não agrupados. Em cada indivíduo, coletei todas as folhas que apresentavam
dano. Em laboratório, calculei a área foliar baseando-me na área da elipse ((R * r * π) /
4), em que R é o raio maior (metade do comprimento da folha) e r é o raio menor
(metade da largura da folha).
Estimei a proporção de área foliar consumida com o auxílio de papel
milimetrado: com a folha sobre o papel milimetrado, contei quantos quadrados com 25
mm2 de área coincidiam sobre buracos na folha. Calculei a proporção de área foliar
consumida para cada folha dividindo a área foliar consumida pela área total da folha.
Para cada indivíduo, calculei a proporção média de área foliar consumida como a média
das proporções consumidas de cada folha. Utilizei um teste t para avaliar se havia
diferença na proporção média de área foliar consumida entre indivíduos agrupados e
não agrupados.
Resultados
Recrutamento na planta vizinha
O número de formigas presentes sobre a folha focal aumentou após a aplicação do dano
na folha vizinha (N=29; Média ± dp= 0,59±1,27; p=0,019; Figura 1).

291
Figura 1. Número de formigas Pheidole minutula presente sobre folhas de Maieta
guianensis antes e depois da aplicação de dano a uma folha vizinha (N=29).
Distância de resposta
A probabilidade de recrutamento de formigas P. minutula aumentou com a distância
entre as folhas de indivíduos de Maieta guianensis (χ2= 5,36; gl=1; p= 0,020; Figura 2).

292
Figura 2. Relação entre a probabilidade de recrutamento por Pheidole minutula em
folhas não danificadas próximas a folhas danificadas em um experimento envolvendo
indivíduos de Maieta guianensis estabelecidos a diferentes distâncias (N=29).
Nível de isolamento e área foliar consumida
A proporção de área foliar consumida nas plantas não agrupadas (N=11; Média ± dp:
11,4 ± 10,4) foi maior que a proporção de dano nas folhas das plantas agrupadas (N=11;
Média ± dp: 8,1 ± 5,4; F(1,20) = 3,7; p=0,049; Figura 3).

293
Figura 3. Proporção média de área foliar consumida em indivíduos de Maieta
guianensis espacialmente agrupados e não agrupados (N=22).
Discussão
As formigas P. minutula respondem ao dano foliar em plantas vizinhas àquela que
ocupam. Além disso, indivíduos de M. guianensis sofrem menor intensidade de
herbivoria quando ocorrem próximos a outros indivíduos da mesma espécie.
Provavelmente essa redução na intensidade de herbivoria está associada ao
recrutamento que ocorre nas folhas vizinhas àquela que sofreu o dano. A presença de
um herbívoro em uma folha é percebida pelas formigas que habitam as folhas vizinhas
de maneira que estas se mobilizem para a defesa. Essa resposta pode afastar eventuais
herbívoros das plantas vizinhas àquela onde ocorreu o dano. Assim, nas agregações de
M. guianensis, o ataque parece gerar uma reação sinérgica entre as formigas, o que pode
resultar na diminuição do risco de predação entre indivíduos que ocorrem próximos uns
aos outros.
Assim como demonstrei para o sistema M. guianensis/Pheidole minutula,
Agrawal (1998) demonstrou que formigas do gênero Azteca presentes em folhas da

294
planta Cecropia obtusifolia respondem a estímulos causados pela liberação de
compostos voláteis em folhas danificadas de plantas vizinhas. Os dois casos mostram
que a interação entre estratégias de defesa direta (quando a planta produz defesa física
e/ou química) e indireta (quando a planta estabelece interações de defesa com
organismos de outras espécies) (Schaller 2008) pode aumentar a eficiência da defesa em
plantas agrupadas.
Observei, durante o experimento, que o número de formigas recrutado na folha
focal era menor que o número recrutado na folha danificada (obs. pessoal). Isso pode
indicar que a resposta ao estímulo químico começa com poucas formigas percorrendo a
folha em busca da causa da liberação do volátil. Ao percorrer a folha e não encontrar
dano ou herbívoro, não há a mesma mobilização massiva que é vista na folha
danificada. Talvez os compostos liberados estimulem o patrulhamento inicial, mas para
que mais formigas sejam recrutadas é preciso que as primeiras encontrem o substrato
danificado, ou o herbívoro.
Curiosamente, a probabilidade de o dano ocorrido em uma folha vizinha resultar
em recrutamento em uma folha que não foi atacada diminui em função da distância
entre as folhas. Peixoto (2005) também não observou resposta das formigas P. minutula
ao dano em folhas vizinhas em M. guianensis, em um experimento em que considerou
apenas distâncias curtas. Uma explicação para esse fenômeno se baseia no efeito
diferenciado provocado pelos compostos voláteis sobre as colônias de formigas de
plantas vizinhas localizadas a diferentes distâncias. Pressupondo que exista uma
variação individual na composição dos voláteis, as formigas poderiam reconhecer
compostos vindos das plantas mais próximas e não responder a eles. Tal fenômeno se
baseia na familiaridade entre vizinhos (Temeles, 1994), que resulta do contato frequente
das colônias com os voláteis de plantas muito próximas. A partir de certa distância, as

295
formigas não teriam essa familiaridade com os compostos voláteis emitidos e não o
reconheceriam como pertencente à planta que ocupam (Temeles, 1994). Essa percepção
diferencial de voláteis por formigas de plantas vizinhas pode favorecer um nível mínimo
de isolamento, dado que plantas muito próximas não seriam beneficiadas pela defesa
promovida pelas formigas de colônias vizinhas. Tal defesa adicional ocorreria apenas a
partir de certa distância entre as plantas. Apesar da resposta contra o ataque de
herbívoros ser mais eficiente em agrupamentos, um mecanismo favorecendo um nível
mínimo de isolamento garantiria menor pressão competitiva intraespecífica em
agrupamentos de plantas (Coley & Barone, 1996). É possível testar se, de fato, há
mecanismos de reconhecimento entre vizinhos muito próximos. Para testar qual a
distância mínima e máxima de percepção de compostos voláteis por P. minutula, pode-
se alocar extrato foliar de M. guianensis embebido em algodão a diferentes distâncias
das folhas e observar a quais distâncias há recrutamento. Se o nível de isolamento
implica em competição intraespecífica acentuada (Flügge et al. 2012), mas em aumento
da defesa contra predadores, o nível de isolamento na escala da população deveria
depender da importância relativa desses dois fatores.
Agradecimentos
Agradeço aos colegas que ajudaram na coleta de dados: Marcos, o monitor predileto,
Dani Coelho, Joseph-EFA, Tatu, Tati, Lorena e Paulinho Bom. Agradeço ao Paulinho
Bom e ao Paulinho Mau-ravilhoso por todas as vezes que olharam para trás quando eu
os gritava com o computador nas mãos: “você pode tirar uma dúvida? É pequenininha!”
Obrigada por não me afogar no Igarapé.
Agradeço também à Catá e ao Marcos por revisar meu relatório 32 vezes. Só
consegui conceber, executar e escrever meu projeto individual graças ao apoio da plebe

296
efana, coordenadores, professores e monitores. Nem de longe eu poderia imaginar o
quanto um curso de campo poderia mudar minha vida profissional e pessoal. Caramba!
Não acredito até agora!
Agradeço especialmente aos pacientes professores que corrigiram meus projetos
e responderam minhas perguntas esquisitas.
Algumas pessoas foram especialmente importantes, sem as quais eu teria ficado
doida com tantas mudanças: Marcos, Renatinha, Sr. Jorge, Paulinho’s, Glauco, Daniel,
Xexéu, Felipe, André, Catá, Marcel, meu Anjo, Leo Marajó e o bem-humorado
tripulante do barco Regional, Sr. Antônio. Valeu, valeu muito e ainda vai valer por
muito tempo. Fingers crossed!
Referências
Agrawal, A.A. 1998. Leaf damage and associated cues induce aggressive ant
recruitment in a neotropical ant-plant. Ecology 79:2100-2112.
Agrawal, A.A. & M.T. Rutter. 1998. Dynamic anti-herbivore defense in ant-plants: the
role of induced reponses. Oikos 83:227-236
Begon, M., C.R. Townsend & J.L. Harper. 2006. Ecology: from individuals to
ecosystems. Oxford: Blackwell Publishing.
Capurucho, J.M.G. 2010. Pheidole minutula (Hymenoptera: Formicinae) captura presas
usando compostos voláteis liberados por Maieta guianensis (Melastomataceae)
como pista da presença de herbívoros. Em: Livro do curso de campo “Ecologia
da Floresta Amazônica” (Peixoto, P.E.C. & Bobrowiec, P.E.D. eds.).
PDBFF/INPA, Manaus.
Coley, P.D. & J.A. Barone. 1996. Herbivory and plant defenses in tropical forests.
Annu. Rev. Ecol. Syst 27:305–35

297
Dobrovolski, R. 2010. Ausência de efeito de borda sobre a demografia e a diversidade
funcional do banco de plântulas emuma área contínua de floresta amazônica.
Em: Livro do curso de campo “Ecologia da Floresta Amazônica” (Peixoto,
P.E.C. & Bobrowiec, P.E.D. eds.). PDBFF/INPA, Manaus.
Flügge, A.J., S.C. Olhede & D.J. Murrel. 2012. The memory of spatial patterns: changes
in local abundance and aggregation in a tropical forest. Ecology 93(7):1540–
1549
Hamilton, W.D. 1971. Geometry for the selfish herd. Journal of Theoretical Biology
31:295–311
Holt, R. D. 2009. Predation and Community Organization. In: Levin, S.A. The
Princeton guide to Ecology. Princeton University Press, New Jersey.
Peixoto, P.E.C. 2005. Respostas comportamentais da formiga Pheidole minutula
(Hymenoptera: Formicidae) a estímulos químicos e biológicos: compostos
voláteis são identificados pelas formigas? Em: Livro do curso de campo
“Ecologia da Floresta Amazônica” (Machado, G. & Nascimento, H. eds.).
PDBFF/INPA, Manaus.
Schaller, A. 2008. Induced plant resitence to herbivory. Berlim: Springer.
Schoonhoven, L.M., L.L.A. van Loon & M. Dicke. 2005. Insects and flowers:
mutualism per excellence. In: Insect plant biology. 2nd
. Oxford: New York.
Vasconcelos, H.L. 1991. Mutualism between Maieta guianensis Aubl., a
myrmecophytic melastome, and one of its ant inhabitants: ant protection against
insect herbivores. Oecologia 87:295-298

298
Tricomas foliares auxiliam formigas na detecção de herbívoros em plantas
mirmecófita?
Mariana Velasque
Introdução
Formigas e plantas geralmente interagem, mas raramente formam relações mutualísticas
(Schurch et al. 2000). Entretanto, algumas plantas desenvolveram estruturas atrativas
para estes insetos, otimizando assim o sistema de defesa anti-herbivoria (Bronstein
1994). Neste sistema, as formigas patrulham a planta em busca de recompensas como
alimentos ou abrigos, removendo os herbívoros e potenciais competidores de recursos.
Essa relação é chamada de mirmecofilia e é um exemplo extremo de interação
mutualística, uma vez que os organismos envolvidos desenvolveram relações
obrigatórias e simbióticas (Beattie 1985, Benson 1985, Davidson & McKey 1993).
Nestas plantas, ocorreu o desenvolvimento de estruturas complexas e especializadas,
conhecidas como domáceas, onde as formigas estabelecem e abrigam suas colônias
(Beattie 1985, Benson 1985, Vasconcelos 1991). Em contrapartida, as formigas
removem os herbívoros da planta hospedeira durante a patrulha, conferindo proteção à
planta (Vasconcelos 1991).
O custo energético para manter a colônia patrulhando pode ser alto. Por isso a
patrulha é exercida por poucos indivíduos. Esta limitação no número de patrulheiras
pode reduzir a eficiência para encontrar os herbívoros, já que estes ocorrem em baixa
densidade e de forma imprevisível (Brouat et al., 2000). Isso pode resultar na seleção de
sistemas mais rápidos e eficientes de sinalização da presença de herbívoros para as
formigas. Isso auxilia a resposta defensiva e leva as formigas ao local de atividade do
herbívoro (Brouat et al., 2000). Como formigas apresentam um elaborado sistema de

299
recepção química (Hölldobler & Wilson 1990), é possível que compostos químicos
voláteis emitidos pelas partes danificadas nas folhas seja uma importante forma de
recrutar mais formigas (Agrawal 1998, Brouat et al., 2000). De forma semelhante,
distúrbios mecânicos causados por herbívoros também podem intensificar a atividade
anti-herbivoria da colônia (Vasconcelos & Casimiro 1997).
Distúrbios mecânicos podem ser amplificados quando a planta possui estruturas
como tricomas. Na Amazônia Central é comum encontrar plantas mirmecófitas que
possuem tricomas nas folhas (Hölldobler & Wilson 1990). Estas estruturas estão
associadas à resistência da planta contra a herbivoria e a redução da perda de água pela
transpiração (Ehrlinger 1984, Choinski & Wise 1999, Benz & Martin 2006). Como
várias espécies de plantas com tricomas ocorrerem no interior das florestas, a
disponibilidade de água parece não ser um recurso limitante o que justifique a utilidade
dos tricomas para evitar a perda de água. Da mesma forma, esta estrutura parece não
estar ligada à defesa física da planta à herbivoria, visto que estas plantas são
intensamente patrulhadas por formigas (Vasconcelos 1991). Estes fatos sugerem que a
presença de tricomas em plantas mirmecófitas pode não estar associada às funções
usuais do tricoma, mas relacionados à outra função como aumento da sensibilidade da
formiga ao herbívoro.
Neste sentido, o presente estudo teve como objetivo avaliar se tricomas foliares
presentes em Tococa sp. amplificam os estímulos mecânicos gerados pelos herbívoros,
otimizando a detecção destes pelas formigas. Realizei um experimento de retirada dos
tricomas para avaliar se na ausência desta estrutura, o tempo de encontro da formiga
com o objeto que causa o estímulo mecânico aumenta.

300
Métodos
Eu realizei o estudo em uma floresta tropical úmida de terra firme, localizada na
Reserva do Km 41(2°26´S, 59°45´O), situada 80 km ao norte de Manaus, AM, Brasil. O
local é considerado uma Área de Relevante Interesse Ecológico (ARIE), co-gerida pelo
Projeto Dinâmica Biológica de Fragmentos Florestais (PDBFF). Segundo a
classificação de Köppen (Peel et al. 2007), a região apresenta o clima tropical úmido,
com temperatura média anual de 26,7 °C e com precipitação anual variando entre 1900
e 2300 mm (Bierregaard et al. 2001).
Mirmecófitas do gênero Tococa (Melastomataceae) apresentam domáceas
localizadas entre a folha e o pecíolo que são frequentemente ocupadas por formigas,
além de tricomas em toda face adaxial da folha. Marquei 12 indivíduos de Tococa sp.
presentes no interior e na borda da reserva. Em cada planta, selecionei uma folha com a
menor área predada por herbívoros. Em um dos lados da folha (separados a partir da
nervura central), removi todos os tricomas, raspando cuidadosamente com uma lâmina
de barbear. Após a raspagem, mantive a planta sem estímulos induzidos por 24h, a fim
de reduzir o estresse imposto às colônias de formigas e diminuir a liberação de
compostos voláteis pela planta. Ao término deste período estimulei a saída de formigas
das domáceas para patrulhar sobre as folhas manipuladas, dando leves batidas sobre as
folhas. Quando o número de formigas sobre a folha era maior do que dois, passei uma
pinça com ritmo constante sobre um dos lados das folhas, de forma a simular um
pequeno herbívoro caminhando sobre a planta. O movimento foi mantido até que as
formigas encontrassem a pinça. O tempo de encontro da formiga com a pinça foi
cronometrado. Este procedimento foi realizado em ambos os lados da superfície adaxial
da mesma folha. Para avaliar a diferença no tempo de resposta das formigas em relação
ao estímulo simulado nos lados com e sem tricoma da folha, utilizei um teste t pareado.
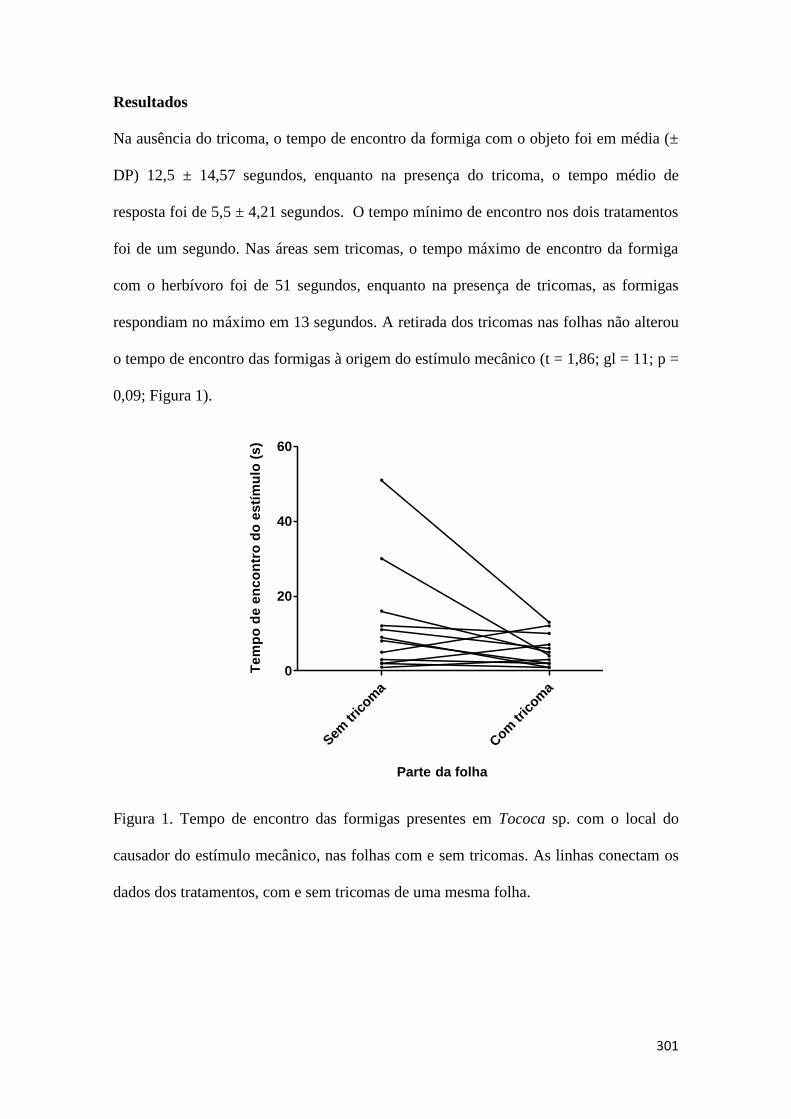
301
Resultados
Na ausência do tricoma, o tempo de encontro da formiga com o objeto foi em média (±
DP) 12,5 ± 14,57 segundos, enquanto na presença do tricoma, o tempo médio de
resposta foi de 5,5 ± 4,21 segundos. O tempo mínimo de encontro nos dois tratamentos
foi de um segundo. Nas áreas sem tricomas, o tempo máximo de encontro da formiga
com o herbívoro foi de 51 segundos, enquanto na presença de tricomas, as formigas
respondiam no máximo em 13 segundos. A retirada dos tricomas nas folhas não alterou
o tempo de encontro das formigas à origem do estímulo mecânico (t = 1,86; gl = 11; p =
0,09; Figura 1).
Figura 1. Tempo de encontro das formigas presentes em Tococa sp. com o local do
causador do estímulo mecânico, nas folhas com e sem tricomas. As linhas conectam os
dados dos tratamentos, com e sem tricomas de uma mesma folha.
Sem
tric
oma
Com
tric
oma
0
20
40
60
Parte da folha
Te
mp
o d
e e
nc
on
tro
do
es
tím
ulo
(s
)

302
Discussão
Diferente do esperado, a presença do tricoma não auxiliou a detecção da formiga,
podendo não estar associado à amplificação dos estímulos mecânicos causados por
herbívoros. Nas florestas, estímulos mecânicos como vento e galhos que caem de
estratos superiores são frequentes. Desta forma, as formigas seriam estimuladas
constantemente e teriam um grande gasto energético, inviabilizando a detecção de
herbívoros por estímulos mecânicos. Assim, as formigas possivelmente possuem
mecanismos que identificam estímulos mecânico específico do herbívoro para incitar a
ação de defesa.
Insetos, em especial formigas, apresentam um reconhecimento químico muito
desenvolvido (Holldobler & Wilson 1990). Estímulos mecânicos nem sempre são
facilmente identificáveis quanto à origem do dano, quando comparados aos químicos.
Neste caso, a liberação de um composto volátil pela planta após sofrer danos, pode ser
um estímulo mais intenso para as formigas do que estímulos mecânicos. A combinação
destes dois estímulos (mecânico e químico) é mais eficiente no mecanismo de defesa
anti-herbivoria.
Em interações mirmecófita, podem existir formigas parasitas no sistema, que
levam a uma variação na qualidade da defesa anti-herbivoria em função da espécie da
formiga hospedeira (Suarez et al. 1998, Gaume & McKey 1999, Bruna et al. 2004).
Desta forma, é possível que nestas plantas, existam tipos de defesas anti-herbivoria
complementares às formigas mirmecófilas. Nesta situação os tricomas podem atuar
juntamente com dureza e arquitetura foliar na defesa estrutural da planta, atuando como
uma barreira física contra a herbivoria. Isso reduz o consumo das folhas pelos
herbívoros (Coley & Barone 1996).

303
Agradecimentos
Gostaria de agradecer ao INPA e ao PDBFF pela oportunidade de realizar este curso.
Ao Programa de Pós-graduação em Ecologia e Conservação de Recursos Naturais da
Universidade Federal de Uberlândia por custear minhas despesas com passagens aéreas
e alimentação. Ao Paulinho Mauravilhoso pelas horas de conversas e puxões de orelha.
Ao Paulinho rainbow máster por carinhosamente me lembrar que tenho relatórios para
entregar (seus capetas, demônios!!!) e nas correções o quanto eu sou gonorrrrrante!!! A
minha mais nova grande amiga Lorrhaynie Shayennie Vanessão por ser minha vizinha
de rede, confidente e por tocar horas de terror no campo comigo.
Aos monitores Xexeu e Barbudinho por criarem novas categorias de vômitos nos
relatórios e mostrarem que existe vida após os geradores. Ao Seu Jorge (porra!!!) e a
Dona Eduarda pela disposição e pelos bolos. As pessoas (não lembro quem) que me
cederam o barbeador para depilar a Amazônia. A minha vizinha de rede Sarahhh que
dava colo toda vez que eu precisava. A Tati-quebra-Ávres por me animar com um
carinhoso abraço.
Ao Vidal, Vidalzinho (Tatu), Bruno, Carol, Aninha, Ludimila, Bianca, Conde,
Joseph, Lore, Renatinha, Leo e Randolfo, por me aturarem durante esse mês. Jamais
imagina que naquele dia no micro-ônibus, nos tornaríamos tão próximos. Obrigada por
compartilharem comigo essa loucura que foi o EFA.
Referências
Agrawal A.A. 1998. Leaf damage and associated cues induce aggressive ant recruitment
in a neotropical ant-plant. Ecology 79:2100–12
Beattie, A.J. 1985. The evolutionary ecology of ant-plant mutualisms. Cambridge:
Cambridge University Press.

304
Benson, W.W. 1985. Amazon ant-plants. In: Amazonia (T.E. Lovejoy ed.). Oxford:
Pergamon Press.
Benz, B.W., C.E. Martin. 2006. Foliar trichomes, boundary layers, and gas exchange in
the species of epiphytic Tillandsia (Bromeliaceae). J Plant Physiol, 163:648–656.
Bierregaard, R.O., C. Gascon, T.E. Lovejoy & R.C.G. Mesquita. 2001. Lessons from
Amazonia: the ecology and conservation of a fragmented forest. London: Yale
University press.
Bronstein, J.L. 1994. Our current understanding of mutualism. The Quarterly Review of
Biology, 69:31-51.
Brouat C., D. McKey, J.M. Bessiere, L. Pascal & M. Hossaert-McKey. 2000. Leaf
volatile compounds and the distribution of ant patrolling in an ant-plant protection
mutualism: preliminary results on Leonardoxa (Fabaceae: Caesalpinioideae) and
Petalomyrmex (Formicidae: Formicinae). Acta Oecol, 21:349–357.
Bruna, E.M., D.M. Lapola & H.L. Vasconcelos. 2004. Interspecific variation in the
defensive responses of obligate plant-ants: experimental tests and consequences
for herbivory. Oecologia, 138:558–565.
Choinski, J.S. & R.R. Wise. 1999. Leaf growth and development in relation to gas
exchange in Quercus marilandica Muenchh. J Plant Physiol,154:302–309.
Coley, P.D.& J.A. Barone. 1996. Herbivory and plant defenses in tropical forests.
Annual Review of Ecology in Systematics 27:305-335
Davidson D.W. & D. McKey. 1993. The evolutionary ecology of antplant relationships.
J Hymenopt Res, 2:13–83
Ehrlinger, J. 1984. Ecology and physiology of leaf pubescence in North American
desert plants. In: Biology and chemistry of plant trichomes (Rodriguez, E., P.L.
Healey & I. Mehta eds). New York: Plenum Press.

305
Gaume, L. & D. McKey. 1999. An ant-plant mutualism and its hostspecific parasite:
activity rhythms, young leaf patrolling, and effects on herbivores of two specialist
plant-ants inhabiting the same myrmecophyte. Oikos, 83:130–140
Hölldobler, B. & E.O. Wilson. 1990. The Ants. Massachusetts: Belknap Press.
Peel, M.C., B.L. Finlayson & T.A. Mcmahon. 2007. Updated world map of the Koppen-
Geiger climate classification. Hydrology and Earth System Sciences, 11:1633–
1644.
Schurch, S., M. Pfunder & B. A. Roy. 2000. Effects of ants on the reproductive success
of Euphorbia cyparissias and associated pathogenic rust fungi. Oikos, 88:6-12
Suarez, A.V., C. de Moraes & A. Ippolito. 1998. Defense of Acacia collinsii by an
obligate and nonobligate ant species: the significance of encroaching vegetation.
Biotropica, 30:480–482
Vasconcelos, H.L. 1991. Mutualism between Maieta guianensis Aubl. a myrmecophytic
meslatome, and one of its ant inhabitants: ant protection against insect herbivores.
Oecologia, 87:295–298
Vasconcelos, H.L. & A.B.Casimiro. 1997. Influence of Azteca alfari Ants on the
Exploitation of Cecropia Trees by a Leaf-Cutting Ant. Biotropica, 29:84-92.

306
Folha consumida é menos querida? Seleção de habitat de Aranhas Faiditus
subflavus (Theridiidae) por folhas de Maieta guianensis (Melastomataceae)
Randolpho Gonçalves Dias Terceiro
Introdução
A teoria de seleção de habitat assume que a seleção por tipos de habitat ocorre quando
há uma maximização da aptidão individual (Revilla et al. 2000). Sendo assim,
indivíduos que selecionem melhor o habitat aumentarão sua chance de sobrevivência e
consequentemente sua reprodução (Orians 1991).
A decisão de indivíduos quanto à seleção de habitats disponíveis, pode ser
influenciada por interações e pela disponibilidade de recursos (Revilla et al. 2000),
como alimento e abrigo. Ambientes com maior quantidade de recursos, como maior
diversidade de alimento e mais sítios para construção de abrigo devem ser favorecidos
na escolha de habitat, pois ele aumenta o sucesso reprodutivo dos indivíduos.
Em aranhas, a disponibilidade de presas e sítios favoráveis para sua instalação,
são fatores determinantes para a seleção do habitat (Souza 2007). Dentre os sítios
favoráveis utilizados por estas espécies, destacam-se a serrapilheira e as plantas, que são
usadas para instalação de suas teias. Nas plantas, a arquitetura pode proporcionar locais
que são usados como abrigo para evitar predadores, locais de forrageamento, encontro
de parceiros sexuais, acasalamento e oviposição, além de oferecer proteção contra
dessecação e condições extremas de temperatura (Kuppers 1989). As aranhas
selecionam as plantas através de informações visuais, táteis e olfativas (Romero &
Vasconselos-Neto 2007). Sendo assim, danos físicos nas folhas (visualmente
perceptíveis) provocados por herbívoros podem ser indicativos indiretos de qualidade
do habitat, pois podem sugerir a existência de intensa atividade de herbívoros na planta.

307
Na Amazônia Central, a aranha Faiditus subflavus é associada com plantas
mirmecófitas do gênero Maieta guianensis (Kasper 2006). Estas plantas possuem
estruturas denominadas domáceas que podem servir de abrigo e sítios para o
estabelecimento de formigas que, em troca, defendem a planta contra ataque de
herbívoros (Romero & Izzo 2004). Duas espécies de formiga são encontradas
associadas a plantas do gênero Maieta, a Pheidole minutula e a Crematogaster laevis
(Vasconcelos 1993). As aranhas Faiditus subflavus costumam habitar a face abaxial das
folhas de Maieta guianensis, próximo às entradas das domáceas (Kasper 2006), sendo
também protegidas pelas formigas contra possíveis predadores e parasitas (Santos
2007). Sendo assim, a seleção de folhas para o estabelecimento de Faiditus subflavus,
deve levar em consideração os danos físicos provocados por herbivoria, pois eles podem
indicar uma baixa eficiência de proteção da planta pelas formigas, o que resulta em
prejuízos para a aranha.
Baseado no fato de que aranhas Faiditus subflavus se estabelecem em Maieta
guianensis, e que as formigas presentes nestas plantas protegem as ootecas e os
indivíduos de F. subflavus, proponho a hipótese de que Faiditus subflavus irão
selecionar folhas sem a presença de herbivoria. Assim, espero que a frequência de
permanência de indivíduos de Faiditus subflavus será maior em folhas intactas do que
em folhas danificadas.
Métodos
Área de estudo
Conduzi o estudo em uma área de floresta contínua na Reserva do Km 41, localizada 80
km ao Norte de Manaus, na Amazônia Central, Brasil. O local é classificado como uma
Área de Relevante Interesse Ecológico (ARIE). A ARIE é administrada pelo Instituto

308
Chico Mendes de Conservação da Biodiversidade (ICMBio) e pelo Projeto de Dinâmica
Biológica de Fragmentos Florestais (PDBFF). A vegetação local é predominantemente
de florestas de terra firme.
Delineamento amostral
Coletei 38 indivíduos adultos de Faiditus subflavus em indivíduos de Maieta
guianensis. Cada indivíduo de F. subflavus foi inserido em diferentes indivíduos de M.
guianensis distribuídos aleatoriamente no ambiente. Antes de instalar o experimento
removi os outros indivíduos de F. subflavus que estavam naturalmente presentes na
Maieta guianensis. Do total de F. subflavus, inseri 19 indivíduos em folhas com
evidência de herbivoria e 19 em folhas intactas. Para o experimento foram utilizadas
folhas apicais dos indivíduos de M. guianensis. Marquei as folhas em que coloquei os
indivíduos de F. subflavus com esmalte amarelo, e etiquetei os indivíduos de M.
guianensis para posterior identificação. Após 24 h verifiquei o estabelecimento dos
indivíduos de F. subflavus nas folhas em que foram inseridos. Contabilizei o número de
indivíduos que permaneceram nas folhas intactas e danificadas.
Análise dos dados
Comparei a seleção de F. subflavus por folhas intactas e danificadas com um teste de
Qui-quadrado. Para tanto, utilizei a frequência de permanência de F. subflavus (variável
resposta) nas folhas consumidas e não consumidas (variável preditora).
Resultados
Do total de 19 indivíduos inseridos em folhas intactas, nove permaneceram na folha
(47,37%) e 10 não permaneceram (52,63%). Nas folhas consumidas, 15 indivíduos

309
(78,95%) não permaneceram, e quatro indivíduos (21,05%) permaneceram nas folhas. A
diferença entre as frequências de seleção de F. subflavus para permanência em folhas
intactas e consumidas não foi significativa (χ2 = 2,92; gl = 1; p = 0,08; Figura 1).
Discussão
Os resultados indicam que F. subflavus não seleciona as folhas em relação ao dano
foliar. Como a formiga oferece proteção para as ootecas das aranhas contra predadores
(Santos 2007), a simples ocorrência das formigas pode sinalizar a qualidade do habitat
para as aranhas. Sendo assim, a presença de formigas parece ser mais importante que o
dano foliar, no processo de seleção de habitat por F. subflavus. Ainda, o padrão de
distribuição de F. subflavus ocorre preferencialmente em plantas com uma alta
quantidade de domáceas (Kasper, 2006). Esse fato pode estar associado à quantidade de
formigas estabelecidas na planta hospedeira (Santos 2007).
Figura 1. Frequência de permanência da aranha Faiditus subflavus em folhas
consumidas e intactas de Maieta guianensis.

310
Além disso, a aranha F. subflavus utiliza a base das folhas para a construção da
teia (Kasper 2006). Sendo assim, provavelmente o dano foliar observado na região
anterior da folha, não modifica o local para fixação das teias de F. subflavus (Oliveira et
al. 2012). Esse fato reforça que a seleção de habitat não possui relação com o dano
foliar, pois independente da área foliar consumida a aranha possui espaço para fixar a
teia. Por fim, é possível que a seleção de habitat para o estabelecimento de Faiditus
subflavus em Maieta guianensis se deva a presença de domáceas com formigas, sendo o
aspecto de herbivoria na folha um fator irrelevante.
Agradecimentos
Gostaria de agradecer a todos os colegas que participaram do curso do EFA 2012! Em
especial às pessoas que me ajudaram neste trabalho: Artur (Tatu ou Vidalzinho) pela
ajuda em campo, escrita do texto e titulo do trabalho (VLW BROTHER); Leonardo
(Bigode de novela das seis) por me ajudar a coletar as aranhas; Lorena (Lore) por me
ajudar a encontrar as Maietas e pela companhia na trilha “M”, a trilha mais punk do
KM-41; e aos monitores Manoela (Manu) e Tiago (Xexeu) pela paciência em corrigir
este manuscrito e ideias!! Obrigado galera!
Referências
Kasper, D. 2006. Fatores ambientais que influenciam a ocorrência e abundância de
aranhas Faiditus subflavus (Theridiidae) em plantas do gênero Maieta
(Melastomataceae). In: Curso de campo ecologia da floresta amazônica (G.
Machado & J.L. Camargo, eds.). INPA/PDBFF, Manaus.

311
Kuppers, M. 1989. Ecological significance of above-ground architectural patterns in
Woody plants: a question of cost-benefit relationships. Trends in Ecology &
Evolution, 4:375:379.
Oliveira, L., R. Rocha, J. Hidasi & A. C. V. Pires. 2012. A aranha Faiditus subflavus
(Theridiidae) selecionam indivíduos de Maieta guianensis (Melastomataceae)
com menor dano foliar? In: Curso de campo ecologia da floresta amazônica (G.
Machado & J.L. Camargo, eds.). INPA/PDBFF, Manaus.
Orians, G.H. 1991. Preface. The American Naturalist, 137:1-4.
Revilla, E., F. Palomares & M. Delibes. 2000. Defining key habitats for low density
populations of Eurasian badgers in Mediterranean environments. Biological
Conservation, 95:269-277
Romero, G.Q. & T. J. Izzo. 2004. Leaf damage induces ant recruitment in the
Amazonian ant-plant Hirtella myrmecophila. Journal of Tropical Ecology,
20:675–682.
Romero, G.Q. & J. Vasconselos-Neto. 2007. Aranhas sobre plantas: dos
comportamentos de forrageamento às associações específicas, pp.67-88. In:
Ecologia e comportamento de aranhas (M.O. Gonzaga, A.J. santos & H.F.
Japyassú, eds.). Rio de janeiro: Interciência.
Santos, G.R. 2007. Babás ou guarda-costas: a formiga Pheidole minutula (Myrmicinae)
protege os filhotes ou as fêmeas da aranha Faiditus subflavus (Theridiidae)? In:
Curso de campo ecologia da floresta amazônica (G. Machado & J.L. Camargo,
eds.). INPA/PDBFF, Manaus.
Souza, A.L.T. 2007. Influência de estrutura do habitat na abundância e diversidade de
aranhas. pp.29-43. In: Ecologia e comportamento de aranhas (M.O. Gonzaga, A.J.
Santos & H.F. Japyassú, eds.). Rio de janeiro: Interciência.

312
Vasconcelos, H.L. 1993. Ant colonization of Maieta guianensis seedlings, an Amazon
ant plant. Oecologia, 95:439-443.

313
Quanto menor o guerreiro, maior a armadura: defesa estrutural em plantas com
diferentes tamanhos foliares
Renata Pimentel Rocha
Introdução
Plantas e insetos herbívoros representam grande parte da diversidade terrestre conhecida
(Farrel et al. 1992). A interação entre esses grupos de organismos é marcada por
processos co-evolutivos, nos quais os herbívoros desenvolvem estratégias de ataque,
enquanto as plantas desenvolvem defesas para combatê-los (Carmona et al. 2011).
As estratégias de defesa das plantas se baseiam em mecanismos que afetam
negativamente a performance ou preferência do herbívoro por determinada planta
(Schaller 2008). Essas estratégias compreendem características morfológicas para
defesa física da planta, como espinhos, tricomas, ceras, rigidez da epiderme, além de
compostos de defesa química, como metabólitos secundários e proteínas que reduzem a
digestibilidade (Schaller 2008). Esses mecanismos de defesa podem ser constitutivos ou
induzidos mediante a ação do herbívoro (Schaller 2008).
Diante da diversidade de estratégias de proteção das plantas, herbívoros
especialistas são mais eficientes em contornar as defesas das plantas e obter alimento, o
que os torna capazes de crescer e se reproduzir mais rapidamente (Coley & Barone
1996). Um grupo de herbívoros altamente especializado é o dos parasitas endofíticos
(Schoonhoven et al. 2005). Esses insetos ovipõem nas folhas e suas larvas se
estabelecem e desenvolvem no interior do tecido foliar. Os insetos minadores se
alimentam consumindo o mesófilo foliar. Os galhadores induzem a formação de
tumores que fornecem nutrientes, alimento e abrigo para os insetos (Schoonhoven et al.
2005).

314
O tamanho da folha é uma das características da planta que podem afetar o ataque
de parasitas. Folhas maiores são mais frequentemente atacadas por parasitas endofíticos
do que folhas menores (Passos et al. 2012). Folhas menores oferecem menos alimento
para o inseto, o que pode limitar seu crescimento, desenvolvimento e reprodução, uma
vez que esses insetos não podem migrar para outras folhas. Dessa forma, é possível que
insetos que colonizassem folhas muito pequenas tenham sido selecionados
negativamente e que, por isso, haja um tamanho foliar mínimo a partir do qual os
insetos conseguem se estabelecer (Passos et al. 2012).
Plantas com folhas pequenas, por terem menor tendência a serem parasitadas,
deveriam investir menos em mecanismos de defesa, de modo a alocar maior volume de
recursos em crescimento e reprodução. Já plantas com folhas maiores, que sofreriam
maior pressão de parasitismo, deveriam investir mais em mecanismos de defesa. Tendo
em vista que as defesas estruturais da planta conferem à folha maior massa por unidade
de área (Cornelissen 2003), espero que a massa foliar por área seja menor em plantas
com folhas pequenas do que em plantas com folhas grandes.
Métodos
Desenvolvi este estudo em uma floresta tropical de terra firme, na borda da estrada ZF-
3, próxima à reserva do Km 41(02°40' S - 59°44' O), a 80 km ao norte de Manaus,
Brasil. Essa reserva está inserida na Área de Relevante Interesse Ecológico (ARIE) do
Projeto Dinâmica Biológica de Fragmentos florestais (PDBFF).
Coletei ramos de todos os arbustos e árvores com até 2 m de altura, ao longo de
um transecto de 500 m de comprimento, na borda da estrada ZF3. Classifiquei os ramos
em morfotipos e calculei a área foliar da menor unidade destacável de cada planta
(folha, folíolo ou foliólulo). Na unidade foliar selecionada, medi o comprimento (c) e a

315
maior largura (l) com uma régua e com esses valores calculei a área (A) usando a
fórmula da área de uma elipse (A = π * c/2 * l/2). Em seguida, selecionei uma folha ou
conjunto de folíolos ou foliólulos de cada morfotipo, tendo como critério aquele que
apresentasse o mínimo de área foliar danificada ou predada. Extraí três amostras de 4
cm2 de cada folha selecionada, evitando a nervura central. Em folhas pequenas, nas
quais o procedimento não foi possível, utilizei a folha inteira. Sequei as amostras em um
forno convencional, por 15 minutos e, em seguida, pesei com o auxílio de uma balança
de alta precisão. Para obter a massa foliar por área (MFA) dividi o valor da massa seca
obtida pela área da amostra pesada. Classifiquei as plantas amostradas em duas
categorias, plantas com folhas pequenas (abaixo de 24 cm2) e plantas com folhas
grandes (acima de 24 cm2). Para testar se a MFA de folhas pequenas é menor que a de
folhas grandes, usei o teste de Mann-Whitney.
Resultados
Coletei 50 morfotipos de plantas, dos quais 21 pertenciam à categoria de folhas
pequenas e 30 à categoria de folhas grandes. A área foliar média (±DP) foi de 6,92 ±
5,56 cm2 para as plantas com folhas pequenas e 201,89 ± 221,43 para as plantas com
folhas grandes. A MFA média (±DP) para as plantas com folhas pequenas foi 0,0176 ±
0,0183 g/cm2
e 0,0059 ± 0,0015 g/cm2
para as plantas com folhas grandes. Os valores de
MFA diferem entre plantas com folhas pequenas e plantas com folhas grandes (U=197;
Z=-2,26; p=0,02). Ao contrário do esperado, folhas pequenas apresentaram MFA maior
que folhas grandes (Figura 1).
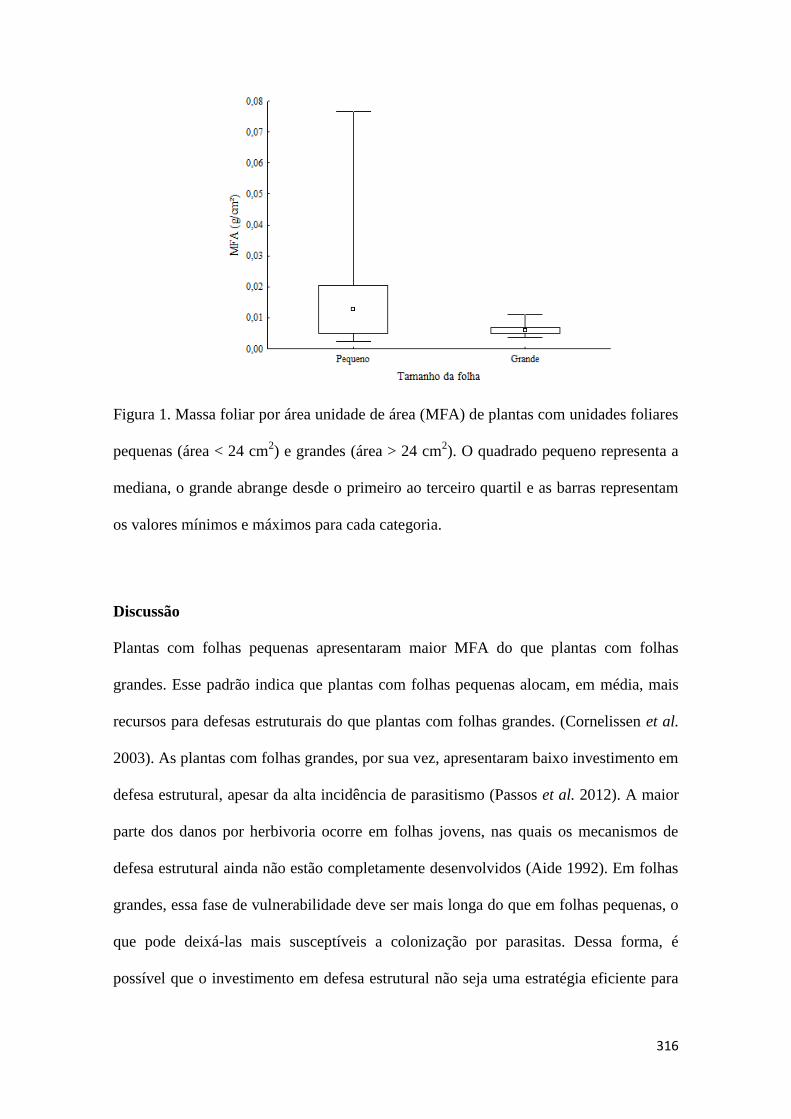
316
Figura 1. Massa foliar por área unidade de área (MFA) de plantas com unidades foliares
pequenas (área < 24 cm2) e grandes (área > 24 cm
2). O quadrado pequeno representa a
mediana, o grande abrange desde o primeiro ao terceiro quartil e as barras representam
os valores mínimos e máximos para cada categoria.
Discussão
Plantas com folhas pequenas apresentaram maior MFA do que plantas com folhas
grandes. Esse padrão indica que plantas com folhas pequenas alocam, em média, mais
recursos para defesas estruturais do que plantas com folhas grandes. (Cornelissen et al.
2003). As plantas com folhas grandes, por sua vez, apresentaram baixo investimento em
defesa estrutural, apesar da alta incidência de parasitismo (Passos et al. 2012). A maior
parte dos danos por herbivoria ocorre em folhas jovens, nas quais os mecanismos de
defesa estrutural ainda não estão completamente desenvolvidos (Aide 1992). Em folhas
grandes, essa fase de vulnerabilidade deve ser mais longa do que em folhas pequenas, o
que pode deixá-las mais susceptíveis a colonização por parasitas. Dessa forma, é
possível que o investimento em defesa estrutural não seja uma estratégia eficiente para

317
plantas com folhas grandes. Além disso, quanto mais reforçada for a estrutura da folha,
maior é seu peso e, consequentemente, maior o custo para sua sustentação. Nesse caso,
é provável que plantas com folhas grandes invistam preferencialmente em mecanismos
de defesa induzida diante de ataques por parasitas.
A tendência de maior MFA nas plantas com folhas pequenas indica que o menor
parasitismo nelas (Passos et al. 2012) deve ocorrer devido a seu investimento em defesa
estrutural. Por outro lado, a variação nos valores de MFA para esse grupo pode indicar o
emprego de diferentes estratégias diante do ataque de parasitas. As plantas com maior
MFA são aquelas que investem muito defesas estruturais, enquanto as que apresentam
menor MFA são aquelas em que esse investimento é pequeno. Folhas pouco protegidas
em geral têm vida útil curta e uma capacidade fotossintética alta (Wright et al. 2002).
Sendo assim, não é vantajoso para a planta investir em defesa estrutural em folhas com
vida útil curta, mas sim em obtenção de energia e crescimento.
Agradecimentos
Ao Paulinho Peixoto (Rainbow master) pela idéia do projeto e orientação no
delineamento amostral. Ao Paulinho Bobrowiec (Batman) pelas explicações e
discussões ao longo do desenvolvimento do projeto. Aos dois Paulinhos pela
oportunidade de participar do curso, pela atenção e apoio constantes, orientação, e
ensinamentos. Ao Marcell Caritavas (Baby) pela ajuda constante ao longo do projeto,
desde a identificação dos morfotipos, discussão das idéias e resultados, correções
afiadas, além de todas as piadinhas e implicâncias. Ao Vidal Carrascosa pela eficiente
ajuda durante a coleta, apoio constante nos momentos complicados e alto astral
garantido em todos os momentos. Ao Daniel Passos (Lagartão) pelas excelentes
contribuições para o trabalho com discussões e conselhos, além da animação

318
contagiante. A Tatiane Menezes (Samara), Joselândio Santos (Joseph), José Hidasi
(Harry Potter) e Artur Madeira (Tatu) pela ajuda ao longo do projeto. Aos monitores,
Thiago Kloss (Beluga Xexéu Gloss) e Marcos Vieira (Barbudo) pela presença
constante, apoio, ensinamentos, críticas incisivas, correções maldosas, além dos ótimos
momentos de convivência. A todos colegas alunos, monitores e professores do EFA
2012 pelos momentos únicos e inesquecíveis.
Referências
Aide, M.T. 1992. Dry season leaf production: an escape from herbivory. Biotropica,
24:532-537.
Carmona, D., M.J. Lajeunesse & M.T.J. Johnson. 2011. Plant traits that predict
resistance to herbivores. Functional Ecology, 25:358–367
Coley, P.D. & J. A. Barone. 1996. Herbivory And Plant Defenses In Tropical Forests.
Annual Review of Ecology Evolution and Systematics, 27:305-35.
Cornelissen, J.H.C., S. Lavorel, E. Garnier, S. Díaz, N. Buchmann, D.E. Gurvich, P.B.
Reich, H. ter Steege, H.D. Morgan, M.G.A. van de Heijden, J.G. Pausas & H.
Poorter. 2003. A handbook of protocols for standardised and easy measurement of
plant functional traits worldwide. Australian Journal of Botany, 51:335-380.
Farrell, B.D., C. Mitter & D.J. Futuyma. 1992. Diversification at the insect-plant
interface. BioScience, 42:34-42.
Passos, D.. T. Calaça, L. Pinto & V. Carrascosa. 2012. Plantas com folhas menores
impedem o estabelecimento de minas e galhas In: Curso de campo ecologia da
floresta amazônica (Peixoto, P.E.C & B.E.D Bobrowiec, eds.). INPA/PDBFF,
Manaus.
Schaller, A. 2008. Induced plant resitence to herbivory. Berlim: Springer.

319
Schoonhoven, L.M., Van Loon, J.J.A., Dicke, M. 2005. Insect-plant biology.New York:
Oxford University Press.
Wright, I.J., M. Westoby & P.B. Reich. 2002. Convergence towards higher leaf mass
per area in dry and nutrient-poor habitats has different consequences for leaf life
span. Journal of Ecology, 90: 534–543.

320
A resposta de captura em uma aranha orbitela aumenta com a redução do valor
da presa
Sarah Freitas Magalhães Silva
Introdução
A obtenção de alimento envolve gastos de tempo e energia para os animais. A teoria do
forrageamento ótimo (MacArthur & Pianka 1966) afirma que estratégias que otimizem
o balanço entre os custos e benefícios do forrageio devem ser favorecidas pela seleção
natural. A minimização de custos pode estar associada a uma redução no tempo de
busca e manipulação da presa, enquanto a maximização dos benefícios pode estar
associada a um aumento na qualidade nutricional da presa (MacArthur & Pianka 1966).
Uma das estratégias possíveis de forrageio que minimiza o tempo de busca por
presas é a do tipo senta-e-espera, caracterizada por um baixo gasto energético (Huey &
Pianka 1981). Além de minimizar o gasto energético do forrageio, animais que adotam
essa estratégia têm uma redução no risco de predação, já que ficam menos expostos aos
predadores. Por outro lado, a obtenção diária de energia é baixa, quando comparada à de
forrageadores que buscam ativamente por suas presas (Pough et al. 1999).
Em aranhas, o forrageio senta-e-espera está associado à produção de teias, que
permitem que elas detectem e subjuguem suas presas sem a necessidade de se
deslocarem em busca delas (Vieira et al. 2007). Aranhas têm a capacidade de conhecer
características da presa, como a massa ou o grau de atividade (Suter 1978), com base na
vibração provocada em suas teias (Vieira et al. 2007). Por serem capazes de distinguir
suas presas, as aranhas podem ajustar as formas de captura de acordo com o tipo de
presa (Vieira et al. 2007).

321
O uso de teias para obter alimento é bastante disseminado entre vários grupos de
aranhas, e a estrutura das teias apresenta uma ampla variação (Vieira et al. 2007).
Algumas espécies de aranha da família Araneidae constróem teias orbiculares dotadas
de abrigos que as protegem de possíveis predadores (Gonzaga 2007). Observei uma
espécie de aranha da família Araneidae que constrói um abrigo na parte superior da teia,
feito por fio de seda e detritos de matéria orgânica. Assumindo que essas aranhas têm a
capacidade de discriminar suas presas, avaliei se o comportamento de forrageio de uma
espécie de aranha dessa família pode ser mediado pelo valor da presa. O valor de uma
presa é dado pela razão entre o tamanho da aranha e o tamanho da presa. Assim, quanto
maior o valor da presa, maior deveria ser a probabilidade de as aranhas deixarem o
abrigo para capturar a presa. Desse modo, testei a previsão de que diante de uma presa
de tamanho fixo no centro da teia, aranhas menores saem do abrigo para obter o recurso
com mais frequência que aranhas maiores.
Métodos
Conduzi o estudo em uma floresta contínua de terra firme localizada na Reserva do Km
41 (02°24’ S – 59°44’ O) administrada pelo Projeto Dinâmica Biológica de Fragmentos
Florestais (PDBFF), localizada a cerca de 80 km ao norte de Manaus, Amazônia
Central, Brasil. A vegetação é do tipo floresta ombrófila úmida. A temperatura média
anual é de 26,7 ºC, com precipitação média anual de 2.186 mm, sendo julho a setembro
o período mais seco, com precipitação de aproximadamente 100 mm/mês (Lovejoy &
Bierregaard 1990).
Na área de estudo, busquei ativamente por teias de uma espécie de aranha da
família Araneidae. As teias são caracterizadas por um abrigo construído na parte
superior da teia. Para testar a minha previsão, forneci cupins, de um mesmo ninho e de

322
comprimentos semelhantes, como presas para as aranhas. Escolhi cupins como presas
porque tinham tamanho semelhante ao das aranhas menores, de modo que mesmo as
aranhas menores deveriam ser capazes de manipular a presa. Em 23 teias, coloquei um
cupim no ponto mais central de cada teia e observei, por 10 minutos, a resposta das
aranhas na presença da presa. Esse experimento foi realizado no período noturno. Após
o experimento em campo, coletei todos os indivíduos de aranha para medir o
comprimento total (da cabeça até o final do abdómen) com o auxílio de um paquímetro,
usei essa medida, devivo ao diminuto tamanho dos indivíduos. Para avaliar se a
probabilidade de deixar o abrigo diminui conforme aumenta o tamanho da aranha,
realizei uma regressão logística, sendo o comprimento das aranhas a variável preditora e
a saída ou não do abrigo em direção à presa, a variável resposta.
Resultados
O comprimento das aranhas variou de 1,16 a 6,61 mm. A partir da distribuição dos
dados, considerei duas categorias de tamanho: menores, com comprimento variando
entre 1,16 a 2,84 mm e maiores, com comprimento de 5,1 a 6,61 mm. A probabilidade
de a aranha sair do abrigo aumenta à medida que aumenta o comprimento do
cafalotórax da aranha (χ²=15,909; gl=1; p<0,01; Figura 1). Embora não tenham deixado
o abrigo para capturar a presa, as aranhas menores tentaram pescar as presas de dentro
do abrigo, através dos fios de seda, sem obter sucesso.

323
Figura 1. Regressão logística da resposta das aranhas à presença de uma presa (cupim)
na teia em função do comprimento das aranhas. 1 corresponde à saída do abrigo e 0 à
permanência no abrigo.
Discussão
A probabilidade de a aranha sair do abrigo aumentou conforme diminuiu o valor da
presa. De acordo com MacArthur & Pianka (1966), o ganho líquido de um item
alimentar se dá pela razão entre o conteúdo energético e o tempo de manipulação.
Assim, embora as presas utilizadas no experimento representem um conteúdo altamente
energético para aranhas pequenas, elas podem representar um custo de manipulação
maior para aranhas menores do que para aranhas maiores.
Uma explicação alternativa está relacionada ao fato de que estratégias de
forrageio podem sofrer mudanças decorrentes da pressão de predação (Krebs & Davies
1993). Gonzaga & Sobczak (2007) demonstraram que aranhas maiores podem ser mais
efetivas em se defender de predadores. Uma vez que deixar o abrigo significa se expor a
potenciais predadores, isso pode explicar o fato de as aranhas maiores terem deixado o
abrigo em busca da presa com mais frequência do que as aranhas menores.

324
Meus resultados mostraram que aranhas maiores, provavelmente mais velhas,
capturaram presas com mais frequência que aranhas menores, provavelmente mais
jovens. Assim, é possível que aranhas mais jovens tenham outras estratégias de
forrageio, como se alimentarem de presas que caiam mais próximas ao abrigo ou que
sejam menores e menos custosas para manipular.
Agradecimentos
Agradeço ao PDBFF/INPA por oferecer um curso de alta qualidade e extremamente
importante para a formação de um pesquisador. Aos Paulinhos, que parecem enxergar
nesse curso uma missão e por isso compartilham tanto conhecimento conosco! Ao
Rainbow Master- mamis de coração- obrigada por ensinar com tanta diversão, por puxar
a orelha com tanto carinho, tornando o processo um pouco mais fácil. Por sempre nos
lembrar de que tudo que foi vivido aqui não irá se repetir, nos fazendo tentar aproveitar
cada momento, como treinar a sensualidade da reconquista, dançar funk na mesa e
deixar a dignidade na rede! Ao Paulinho Batman- papis de coração- obrigada por
conseguir disfarçar todo o stress e ainda conseguir conversar coisas da vida e falar
besteira também! Ao Thiago namorado, que me incentivou a me inscrever para o curso
e não deixou que eu desistisse! Ao Thiago amigo, que suportou todas as minhas crises
de enxaqueca e existenciais! Ao Thiago monitor, que me acompanhou pacientemente no
campo durante esse PI, que foi executado durante a noite! Aos amigos Joseph e
Leozinho, que facilitaram muito a minha vida durante esse tempo por aqui... Juntos
conseguimos mudar o critério de seleção do EFA: tem que ser de Montes Claros!
À dupla dinâmica Shayenne Lohrraynny e Mari Exu, vizinhas de rede e
companheiras de uma relação poligâmica... Muito obrigada por cada risada que dei com
vocês! À Tati (Ávres), pelas brincadeiras e conversas sobre saudade. À Carol (Da Silva)

325
pelas conversas de besteira e de coisa séria também! À Dani Atoladinha por cada
bilhetinho do amor e cada verso de cordel, que nos desestressaram tanto nos momentos
de desespero! À Renatinha Pocket, por sempre me deixar com a auto-estima elevada,
dizendo que sempre estou cheirosa, até mesmo no 40 e úmido! À Loris, organizadora
das festas, por ser um doce de pessoa e pela risadinha que faz a gente dar risada! À
todos os colegas de turma do EFA 2012 que foram minha família durante esses 30 anos,
obrigada por tornarem esse tempo tão feliz e inesquecível! Ao monitor barbudo e
bipolar que, apesar de não ser meu monitor favorito, teve muita paciência comigo (ou
pelo menos fingiu ter) e me ensinou muito! Ao Marcel (Baby) pelas correções no
manuscrito! Ao Moranguinho do Nordeste, que nos ensinou muito com muita diversão!
Aos professores orientadores de POs: Felipe Melo, Cassiano Rosa, Thiago Kloss e
Marcos Vieira. Aos demais professores que acrescentaram novos conhecimentos das
mais diversas áreas: Toyoyo, Cassiano, Dé, Catá, Jack Sparrow, Marcel, Glauco,
Jansen, Manu, Ana, Aline, Ester, Mike, Márcio e Adriano. À Dona Eduarda pelo olhar
carinhoso de vó e por garantir a nossa energia a cada dia! Ao Seu Jorge pela
“delicadeza” e pelo suco de caju gelado de todo santo dia! Por último, mas jamais
menos importante, agradeço aos amigos Alline Mendes, Maria Luiza e Thiago Silva,
que estão cuidando da minha vida lá fora enquanto estou aqui vivendo nesse universo
paralelo!
Referências
Gonzaga, M.O. 2007. Inimigos naturais e defesas contra predação e parasitismo em
aranhas, pp. 209-237. In: Ecologia e comportamento de aranhas (M.O. Gonzaga,
A.J. Santos & H.F. Japyassú, eds.). Rio de Janeiro: Editora Interciência.

326
Gonzaga, M.O. & J.F. Sobczak. 2007. Parasitoid-induced mortality of Araneus
omnicolor (Araneae, Araneidae) by Hymenoepimecis sp. (Hymenoptera,
Ichneumonidae) in southeastern Brazil. Naturwissenschaften, 94:223-227.
Huey, R.B & E.R. Pianka. 1981. Ecological consequences of foraging mode. Ecological
Society of America, 62:991-999.
Krebs, J.R. & N.B. Davies. 1993. An introduction to behavioural ecology. Oxford:
Blackwell Publishing.
Lovejoy, T.E. & R.O. Bierregaard. 1990. Central Amazonian forests and the minimal
critical size of ecosystems project, pp. 60‐71. In: Four Neotropical Rainforests
(A.H. Gentry, ed.). London: Yale University Press.
MacArthur, R.H. & E.R. Pianka. 1966. On optimal use of a patchy environment.
American Naturalist, 100:603-609.
Pough, F.H, C.M. Janis & J.B. Heiser. 1999. A vida dos vertebrados (2ª edição).
Atheneu: São Paulo.
Suter, R.B. 1978. Cyclosa turbinate (Araneae: Araneidae): prey discrimination via web-
borne vibration. Behavioral Ecology and Sociobiology, 3:283-296.
Vieira, C., H.F. Japyassú, A.J. Santos & M.O. Gonzaga. 2007. Teias e forrageamento,
pp. 45-65. In: Ecologia e comportamento de aranhas (M.O. Gonzaga, A.J.
Santos & H.F. Japyassú, eds.). Rio de Janeiro: Editora Interciência.

327
Árvores grandes limitam o tamanho de suas vizinhas?
Tatiane Gomes Calaça Menezes
Introdução
Interações entre plantas tem um papel importante no estabelecimento, crescimento e
desenvolvimento das espécies vegetais. Essas interações podem ser positivas ou
negativas e atuam em diferentes fases do ciclo de vida (Ewel & Hiremath 2005).
Interações positivas são aquelas em que pelo menos uma das espécies que interage é
beneficiada, sem prejuízos à outra. Exemplo disso é a facilitação, que ocorre quando a
presença de uma espécie altera as condições físicas do ambiente, como o microclima,
proporcionando um sítio favorável ao estabelecimento de outra espécie (Ricklefs 2003).
Já as interações negativas são aquelas em que uma das espécies envolvida na interação é
prejudicada. Por exemplo, a competição por recursos comuns, onde uma espécie ou
indivíduo limita ou impede o crescimento de uma planta vizinha já estabelecida (Ewel
& Hiremath 2005).
O crescimento e desenvolvimento das plantas são influenciados por fatores
ambientais como disponibilidade de luz, água e nutrientes (Poorter 2005). Esses fatores
agem em conjunto, e a intensidade da influência de cada um deles sobre as espécies
varia entre os diferentes tipos de ambiente. Em ambientes onde os recursos são
sazonais, como as florestas secas, a disponibilidade de água pode representar o maior
fator limitante ao crescimento e desenvolvimento das plantas quando comparado a
outros recursos. Porém, em ambientes sem mudanças sazonais marcantes, como nas
florestas tropicais úmidas, recursos como água e luz deixam de ser limitantes ao
estabelecimento das espécies e as interações entre espécies passam a ter maior

328
importância na determinação de quais espécies conseguem se desenvolver (Lieberman
& Lieberman 2007).
Em florestas tropicais, muitas interações planta-planta resultam em impactos
negativos para pelo menos uma das espécies, pois elas competem por recursos comuns
(Ewel & Hiremath 2005). Nestas florestas altamente diversas, interações entre plantas
são bastante frequentes e sua coexistência depende basicamente da disponibilidade de
espaço físico (Lieberman & Lieberman 2007). Nestes ambientes, a influência de uma
planta estabelecida sobre suas vizinhas pode ser alta. Entre árvores, a competição por
recursos, como a radiação solar, pode resultar em um padrão de variação de tamanho
entre espécies próximas espacialmente. Isso se deve a penetração diferencial de luz no
interior da floresta que varia em função da abertura do dossel (Brown 1993). A intensa
competição por luz gera uma corrida ascendente em direção ao dossel, onde este recurso
é abundante, maximizando o potencial de crescimento das plantas (Westoby 1998).
Dessa forma, árvores maiores diminuem a quantidade de luz que as árvores, logo abaixo
da influência de sua copa, recebem, e podem limitar o crescimento destes indivíduos.
Desse modo, é razoável esperar que a influência de uma árvore sobre o
crescimento de árvores vizinhas localizadas abaixo da sua copa seja maior com o
aumento do tamanho desta árvore. Assim, meu objetivo foi avaliar se a presença de uma
árvore grande tem influência sobre o tamanho das árvores vizinhas, numa floresta
tropical onde a radiação solar no sub-bosque é um recurso limitado por espécies
maiores. Se a relação de interferência planta – planta é verdadeira, espero que (i) com o
aumento da circunferência à altura do peito (CAP) das árvores focais, a variação do
CAP das árvores vizinhas seja menor e que (ii) com o aumento da distância da árvore
focal, o CAP das árvores vizinhas seja maior.

329
Métodos
Área de estudo
Realizei o estudo na Área de Relevante Interesse Ecológico do Projeto Dinâmica
Biológica de Fragmentos Florestais (PDBFF), no sítio km 41. A área está localizada na
Amazônia Central à aproximadamente 80 km ao norte da cidade de Manaus, Brasil. O
clima da região é tropical úmido de acordo com a classificação de Köppen (Peel et al.
2007), a precipitação média anual é de 2.186 mm e a temperatura média anual é de 26,7
ºC (Lovejoy & Bierregaard 1990). A vegetação da área é caracterizada como floresta
tropical úmida de terra firme e pelo relevo podemos identificar três habitats: platô,
vertente e baixio. Os platôs ocupam os locais mais altos e planos, com solos argilosos e
bem drenados. Os baixios ocupam as áreas de relevo mais baixo, sujeitas a pulsos de
inundação. E as vertentes são faixas de transição entre essas áreas de maior e menor
altitude (Ribeiro et al. 1999). Para fins de controle da variação de características
ambientais, utilizei apenas áreas de platô neste estudo.
Coleta dos dados
Esta reserva possui trilhas a cada 100 m nas direções norte-sul e leste-oeste. Utilizei 3
trilhas como transecto e em cada trilha percorri cerca de 300 metros. A cada 10 m me
distanciei pelo menos 7 m da trilha e selecionei o indivíduo de maior CAP como árvore
focal. Para cada planta focal, marquei nas direções norte, sul, leste e oeste, a árvore
vizinha mais próxima. No total, utilizei 80 conjuntos de árvores, cada um composto por
uma árvore focal e quatro árvores vizinhas. Mensurei a circunferência à altura do peito
(CAP) de cada árvore amostrada e medi a distância da árvore focal para cada planta
vizinha selecionada. Utilizei como critério de seleção da árvore focal o CAP mínimo de
15 cm e para as vizinhas, a altura mínima de 1,5 e/ou CAP de 3,5 m, com o objetivo de

330
evitar plantas jovens que ainda não se estabeleceram completamente. Utilizei o CAP da
árvore focal e das árvores vizinhas como indicativo do tamanho das plantas, uma vez
que esta variável está intimamente relacionada com a altura das árvores.
Análise dos dados
Para testar se o tamanho de uma árvore vizinha era explicado pela distância ou pelo
CAP da árvore focal fiz uma regressão múltipla. Para isso, sorteei apenas uma árvore
vizinha por conjunto. Para testar se a árvore focal influencia o tamanho das árvores
vizinhas, fiz uma ANCOVA utilizando o CAP da árvore focal como variável preditora e
o desvio padrão do CAP de cada conjunto de árvores vizinhas como variável resposta.
Esta variável resposta expressa uma medida da variação de tamanho da amostra de
árvores vizinhas. Para essa análise, usei a distância da árvore focal como co-variável.
Resultados
O CAP das árvores focais variou entre 15 e 290 cm, com média e desvio padrão de
78,83 ± 55,74 cm. As árvores vizinhas apresentaram CAP entre 3,5 e 130 cm com
média e desvio padrão de 19,44 ± 19,42 cm. A distância das árvores vizinhas à árvore
focal variou entre 0,01 e 6 m. O CAP das árvores focais não influenciou o tamanho de
suas vizinhas (F(1,77) = 2,36; p = 0,128), porém o CAP das árvores vizinhas foi maior
com o aumento da distância da árvore focal (F(1,77) = 8,78; p = 0,004; Figura 1).

331
Figura 1. Relação entre a distância da árvore focal com os resíduos da regressão entre o
CAP da árvore focal e CAP de sua vizinha direta, em uma floresta de terra firme na
Amazônia Central, Manaus, Brasil.
O desvio padrão do CAP das árvores vizinhas foi maior em árvores focais de
maior CAP (F(1,77) = 7,56; p = 0,007; Figura 2).
Figura 2. Relação entre a circunferência à altura do peito (CAP) das árvores focais com
o desvio padrão do CAP das árvores vizinhas diretas na Amazônia Central, Manaus,

332
Brasil. Cada ponto refere-se a um bloco formado por uma focal e quatro árvores
vizinhas.
Discussão
Árvores maiores não representam um impedimento ao desenvolvimento das plantas
vizinhas. Isso foi evidenciado pelo aumento da variação do CAP das árvores vizinhas
com o aumento do CAP da árvore focal, contrariando minha previsão. Entretanto,
árvores maiores limitam o crescimento das árvores vizinhas, já que existe uma relação
entre o tamanho do CAP das árvores vizinhas e o aumento da distância da árvore focal.
Uma possível explicação para esse padrão é que uma árvore grande pode consumir boa
parte da luz e nutrientes disponíveis no ambiente próximo, diminuindo a disponibilidade
desses recursos para plantas que estão sob a influência de sua copa. Desse modo,
árvores de diferentes tamanhos e variadas estratégias de crescimento e captação de
recursos podem se estabelecer tentando se desenvolver sob a influência espacial dessa
árvore de grande porte.
Diferente do esperado, com o aumento do tamanho da árvore focal, a variação
do tamanho das árvores vizinhas também aumentou. À medida que a copa de uma
árvore se eleva, a influência de seu sombreamento parece diminuir. Árvores maiores
permitem lacunas por onde a radiação solar pode penetrar. Isso pode explicar a
formação de um gradiente vertical de luminosidade (Poorter 2005). Assim, árvores
maiores podem abrigar uma maior variação de tamanho entre suas vizinhas próximas,
devido a um gradiente de luminosidade decorrente da penetração diferencial de luz na
floresta através da sua copa. Esse fenômeno pode levar a um gradiente de tamanho de
árvores.

333
Diante desse gradiente de árvores de diferentes tamanhos pode se estabelecer
uma sucessão em pequena escala. Quando a árvore maior perece por queda ou morte
provocada por qualquer perturbação, tem início uma corrida pela luz (Lieberman &
Lieberman 2007). A árvore de maior tamanho entre os vizinhos deve ocupar a lacuna
aberta, deixando seu nicho, que passa a ser ocupado pela árvore de tamanho inferior a
ela. O que se vê então, é uma sucessão oportunista, onde lacunas vão sendo abertas e
preenchidas de acordo com o gradiente de tamanho das árvores. Portanto, embora uma
árvore grande não represente um obstáculo ao desenvolvimento de suas vizinhas
menores, ela controla o processo de sucessão em pequena escala das árvores sob sua
influência direta.
Agradecimentos
Agradeço aos coordenadores do EFA 2012 (Paulo Enrique, Paulo Estefano e Zé) pela
oportunidade de poder desfrutar dos encantos da Amazônia de uma forma tão intensa e
marcante. Com certeza, esta foi uma experiência marcante, construtiva e encantadora
jamais vivida antes. Aos monitores Thiago e Marcos, pelas conversas e prontidão a tirar
dúvidas sempre. A todos os professores que nos transmitiram o melhor de si nestes 30
dias: Ester, Felipe (Moranguinho do Nordeste), Márcio (Richard Gere), Mike, Tiago
(Toyoyo), Jansen, Aline, Catá, André, Fabrício (Jack Sparrow), Adriano, Marcel e
Glauco. À Dona Eduarda e seu Jorge, pelas comidinhas gostosas e carinho para comigo
em todo este período. Em especial, ao Paulinho Rainbow, por todo exemplo de
dedicação e amor pelo que faz que pude testemunhar nestes 30 dias. Teu exemplo me
inspira profundamente, obrigada por todos os ensinamentos. Ao Paulinho Mau, por toda
a paciência e críticas que só contribuíram para minha formação.

334
Aos amigos que fiz. Obrigada por tornarem estes 30 dias tão maravilhosos e
incríveis. Vocês tem lugar em meu coração para sempre: À Lorena (Lori), Ana Rorato
(Maria Betânia), Mari (Exú), Sarah, Carol (Missbalde na cerveja quente), Cibele
(Condessa Lorraynie), Ana Carol (Pezinho de ouro), Renatinha (Pockett), ao Joselândio
(Joseph). Obrigada pelas conversas, pelas brincadeiras, resenhas, risadas, pelo carinho e
atenção, amo todos vocês. A todos os colegas do EFA 2012 com quem convivi este
mês: Daniel (Lagartão), Artur (Tatu), Dani (Atoladinha), Bianca, Ludmilla, Bruno,
Vidal, Hidasi (Harry Potter), Léo (Mr. Bigodinho) e Randolpho (Hobbit). Ao meu
anjo(a), pelos presentinhos e bilhetinhos que me alegraram no decorrer destes dias.
Agradeço a todos que me ajudaram na elaboração e execução deste projeto,
assim como nas análises e discussões. Aos professores, Adriano Melo, Marcel Bebê,
Paulinho Rainbow e Paulinho Mau e aos monitores Marcos (monitor Barbudo) e Thiago
(Xexéu) pela ajuda na concepção da ideia. À Bianca, pela ajuda no trabalho de campo, à
Missbalde no Igarapé e Rainbow Master, por ajudar a planilhar os dados. Ao Paulinho
Rainbow, pela análise dos dados. Ao Marcel, Carol (Missbalde nos mícuins) e Paulinho
Mau pelas discussões. Aos corretores Ana (alto clero) e Xexéu, pelas preciosas
intervenções.
Referências
Brown, N. 1993. The implications of climate and gap microclimate for seedling growth
conditions in a Bornean lowland rain forest. Journal of Tropical Ecology,
9:153–168.
Ewel, J.J. & A.J. Hiremath. 2005. Plant-plant interactions in tropical forests. In: Biotic
interactions in the tropics: Their role in the maintenance of species diversity

335
(Burslem, D., M. Pinard & S. Hartley, eds.), Ecological Reviews. New York:
Cambrindge University Press.
Lieberman, M. & D. Lieberman. 2007. Nearest-neighbor tree species combinations in
tropical forest: the role of chance, and some consequences of high diversity.
Oikos, 116:377-386.
Lovejoy, T.E. & R.O. Bierregaard. 1990. Central Amazonian forests and the minimal
critical size of ecosystems project, pp. 60‐71. In: Four Neotropical Rainforests
(A.H. Gentry, ed.). London: Yale University Press.
Peel, M.C., B.L. Finlayson & T.A. Mcmahon. 2007. Updated work map of the Köppen
–Geiger climate classification. Hydrology and Earth System Sciences, 11:1633-
1644.
Poorter, L. 2005. Resource capture and use by tropical forest tree seedlings and their
consequences for competition. In: Biotic Interations in the tropics: Their role in
the maintenance of species diversity (Burslem, D., M. Pinard & S. Hartley,
eds.), Ecological Reviews. New York: Cambrindge University Press.
Ribeiro, J.E.L.S., M.J.G. Hopkins, A. Vicentini,C.A. Sothers, M.A. Costa, J.M. Brito,
M.A.D. Souza, L.H.P. Martins, L.G. Lohmann, P.A.C.L. Assunção, E.C.
Pereira, C.F.D. Silva, M.R. Mesquita & L.C. Procópio. 1999. Flora da Reserva
Ducke: guia de identificação das plantas vasculares de uma floresta de terra-
firma na Amazônia Central. Manaus: INPA.
Ricklefs, R.E. 2003. A economia da natureza. Rio de Janeiro: Guanabara Koogan.
Westoby, M. 1998. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and
Soil, 199:213-227.

336
Qual é a influência da qualidade do macho da espécie Chalcopteryx scintilans
(Odonata: Polythoridae) no tempo de luta por um território?
Vidal C. Carrascosa
“Não é a espécie mais forte que sobrevive, nem a mais inteligente, mas sim a que
melhor responde às mudanças.” (Charles Darwin)
Introdução
Machos de muitas espécies animais frequentemente brigam pela posse de territórios de
acasalamento (Arnott & Elwood 2008). Nessas brigas, os machos vencedores
normalmente são aqueles com melhor capacidade de luta (Mesterton-Gibbons et al.
1996). Portanto, possuir um território é um indicativo de maior qualidade do macho, já
que ele foi capaz de vencer o dono anterior do território (Mardem & Cobb 2004,
Plaistow & Siva-Jothy 1996).
A capacidade de luta do macho pode ser determinada por vários fatores como
tamanho corporal (Mesterton-Gibbons et al. 1996), quantidade de gordura estocada
(Mesterton-Gibbons et al. 1996, Plaistow & Siva-Jothy 1996), qualidade do sistema
imunológico (Mardem & Cobb 2004) ou quantidade de confrontos vencidos (Keil &
Watson 2010). Porém, a característica determinante da capacidade de luta pode
depender dos comportamentos empregados durante os confrontos. Em combates com
contato físico, por exemplo, traços determinantes de força devem ter um grande peso na
definição do vencedor. Por exemplo, aranhas que possuem mais de 15% de massa em
relação aos seus oponentes normalmente são as vencedoras dos confrontos com contato
físico (Keil & Watson 2010). Por outro lado, em brigas sem contato físico, a
persistência na luta pode se tornar mais importante que a força para determinar a vitória.

337
Assim traços como a quantidade de gordura estocada podem determinar o vencedor
(e.g. Plaistow & Siva-Jothy 1996).
Em algumas espécies, existem características que, apesar de não determinarem a
capacidade de luta, podem estar correlacionadas com os traços que determinam a
chance de vitória do macho, podendo atuar como pistas indiretas da qualidade do
oponente. A pigmentação, em particular, é uma característica muito conspícua em
machos de diversas espécies (Contreras-Garduño et al. 2006, Schultz & Fincke 2009).
Nas libélulas, por exemplo, a quantidade de pigmento das asas está correlacionada com
a qualidade dos machos bem como sua capacidade de ganhar brigas (Contreras-Garduño
et al. 2006). Assim, em confrontos sem contato físico, a pigmentação de um macho
pode ser uma característica que indique a sua quantidade de reserva energética, caso as
brigas sejam decididas com base na capacidade de persistência.
Machos de libélula geralmente apresentam colorações conspícuas e usam a visão
como principal sentido sensorial (Córdoba-Aguilar 2008). Em muitas espécies os
machos defendem um território de acasalamento, sendo que os indivíduos com sistema
imunológico mais eficiente (Contreras-Garduño et al. 2006) e com maior quantidade de
gordura estocada (Plaistow & Siva-Jothy 1996) tendem a vencer os confrontos. Um
sistema imunológico mais eficiente indica uma maior resistência ao ataque por
patógenos e assegura maior eficiência no embate (Mardem & Cobb 2004). O estoque de
gordura, por sua vez, proporciona ao indivíduo uma maior quantidade de energia, que
garante uma maior persistência na luta (Fitzstephens & Getty 2000). Uma característica
perceptível pelo coespecífico, correlacionada com a gordura e imunologia, é a cor do
indivíduo (Fitzstephens & Getty 2000). Produzir pigmentos acarreta um custo
energético alto. Consequentemente, a quantidade de pigmentos produzidos por um

338
indivíduo tende a ser um estimador confiável da sua capacidade de luta (Fitzstephens &
Getty 2000).
A espécie Chalcopteryx scintilans é comum em riachos da floresta amazônica.
Os machos apresentam cor acobreada nas asas, que reflete vermelho quando a luz incide
nela, enquanto fêmeas apresentam cor preta nas asas. Machos desta espécie
frequentemente brigam pela posse de territórios de acasalamento ao longo dos riachos.
Porém, não se sabe qual característica determina a vitória. Nesse sentido, minha
hipótese é que a quantidade de gordura estocada nos machos determina a capacidade de
luta. Dessa forma, dado que a quantidade de pigmento na asa indica qualidade, espero
que machos de C. scintilans com maior intensidade de vermelho nas asas invistam mais
tempo na luta pelo território.
Métodos
Área de estudo
Realizei o estudo em um riacho de pequena ordem (localmente conhecido como
igarapé) na reserva do Km 41, inserida na área de relevante interesse ecológico (ARIE)
cogerida pelo Projeto Dinâmica Biológica de Fragmentos Florestais (PDBFF) e
localizada a aproximadamente 80 km ao norte de Manaus, Brasil. A vegetação é
formada por floresta ombrófila densa de terra firme (Ribeiro et al. 1999).
Organismo de estudo
Indivíduos da espécie C. scintilans são frequentemente encontrados em igarapés de
floresta contínua, entre 10:00 h e 14:00 h. Os territórios defendidos pelos machos
geralmente estão localizados em uma pequena mancha de sol associada a uma árvore
caída sobre o curso d´água. O comportamento de briga desta espécie consistente em

339
uma série de manobras aéreas (Resende 2002). A briga começa quando um macho se
posiciona na frente do outro e flexiona as asas posteriores para baixo, exibindo a
coloração interna fortemente acobreada. Um macho permanece tentando deslocar o
outro para trás gerando um movimento de vai-e-vem e mudando o ritmo do bater das
asas, embora os embates tenham pouco o nenhum contato físico (Resende 2002).
Avaliação do investimento nas brigas
Realizei o experimento entre os dias 1 e 2 de novembro de 2012. Para testar o
comportamento do macho ante a invasão do território por outro macho, fixei um macho
recentemente morto na ponta de um galho com cola branca, cuidando para que suas asas
ficassem estendidas e visíveis. Após um período de uso, o indivíduo modelo começava
a se degradar (ex. a asa quebrava). Por isso, substituí o indivíduo modelo três vezes para
manter a aparência mais similar à encontrada na natureza.
Após preparar a haste com o modelo morto, busquei ativamente por machos de
C. scintilans ao longo do igarapé. Ao encontrar um indivíduo, eu aproximava o modelo
morto ao macho de libélula. Com a aproximação, o macho territorial normalmente
alçava voo e se aproximava do modelo, batendo as asas de forma mais rápida e às vezes
investindo fisicamente contra o modelo. Considerei essa mudança comportamental
como início da briga e medi o tempo que cada indivíduo permaneceu efetuando esse
padrão de voo. Como o modelo não era removido do território, o fim da disputa ocorria
quando o macho vivo atingia seu limite de investimento. Como controle, aproximei um
galho sem nenhum individuo na frente de três machos, não obtendo resposta em
nenhum dos casos.
Após medir o tempo, capturei o indivíduo com um puçá entomológico e
transportei-o até o laboratório para medir a intensidade de vermelho nas asas anteriores.

340
No laboratório dissequei os indivíduos e fotografei as asas anteriores de cada um nas
mesmas condições de luminosidade. Calculei a quantidade de vermelho da asa anterior
esquerda de cada macho usando o programa Corel PHOTO-PAINT X5 2010, que
fornecia a quantidade média de vermelho. Posteriormente, usei um modelo linear geral
para testar se o tempo investido na luta pelos machos (variável resposta) estava
associado à quantidade de vermelho nas asas (variável preditora). Usei cada modelo de
macho como covariável categórica para controlar os possíveis efeitos do modelo sobre o
comportamento de briga dos machos.
Resultados
Estimulei o combate de 18 machos localizados em um total de 15 territórios. O tempo
médio (±DP) de briga foi de 10,37±9,32 s e a quantidade média de vermelho (±DP) das
asas foi de 135,3±24,6 pixels. Não encontrei relação entre o tempo de briga e a
quantidade de vermelho das asas (F(1,14)=0,66; p=0,43; Figura 1). Os modelos também
não afetaram diferencialmente as respostas dos machos (F(2,14)=1,98; p=0,17).

341
Figura 1. Relação entre o resíduo de um modelo linear geral entre tempo de luta e
modelos de machos usados no experimento e quantidade de vermelho das asas em
machos da espécie de libélula C. scintilans em um igarapé da na ARIE PDBFF,
Amazônia Central, Brasil.
Discussão
Indivíduos da espécie C. scintilans de maior qualidade (maior pigmentação nas asas)
não investiram mais tempo na luta pelo território. Considerando que os modelos
utilizados representam rivais que nunca desistem da luta, é possível que a quantidade de
energia não determine a chance de vitória, e consequentemente não seja representada
pela coloração da asa. Se a quantidade de reservas energéticas fosse um importante
determinante da qualidade dos machos, indivíduos com maior pigmentação deveriam ter
brigado por mais tempo (e.g. Contreras-Garduño et al 2006, Fitzstephens & Getty
2000).
Se a gordura não determina a persistência na luta, outras características como
tamanho (Mesterton-Gibbons et al. 1996), qualidade do sistema imune (Mardem &
Cobb 2004) ou quantidade de músculo (Keil & Watson 2010), podem determinar a
90 100 110 120 130 140 150 160 170 180 190
Quantidade de vermelho (pixels)
-15
-10
-5
0
5
10
15
20
Res
idu
os
do
mo
del
o l
inea
r g
eral
en
tre
tem
po
da
dis
pu
ta
em r
ela
ção
ao
s m
od
elo
s d
e m
ach
os
usa
do
s n
o e
xp
erim
ento

342
chance de vitória. Estas características não estão relacionadas com uma maior duração
da disputa já que não representam estoques de energia utilizados durante o confronto.
Além disso, o fato de existir contato físico com o modelo, mesmo que esporádico, pode
indicar que o tamanho do indivíduo eventualmente determina a chance de vitória. Um
indivíduo de maior tamanho poderia causar injurias maiores no oponente ou resistir
mais ao recebimento de golpes.
Alguns modelos que tratam das regras de resolução de brigas sugerem que o
confronto se divide em várias fases ao longo da disputa pelo território, o que implica na
mudança de comportamento dos machos durante o confronto (Arnott & Elwood 2008).
Possivelmente a coloração da asa é um fator importante no início da briga, quando os
machos podem se avaliar. Logo, se a gordura fosse uma característica importante
deveria haver, mesmo que nessa fase inicial, uma relação entre coloração das asas e
duração da disputa. Porém, como o comportamento do modelo não mudou,
eventualmente, o macho rival não deu seguimento às fases seguintes da luta.
Eventualmente, mudanças comportamentais em confrontos escalonados possam indicar
quais características, que não a gordura, seriam mais prováveis para determinar a chance
de vitória.
Agradecimentos
Bom, gostaria começar agradecer as pessoas que permitiram a realização deste trabalho,
a principal Paulinho Rainbow pela sugestão de trabalho, apoio e correção, além de ser
fonte de admiração e inspiração de como ser legal e eficiente numa mesma pessoa, em
grande parte desencadeador da minha transformação como pesquisador no decorrer
deste mês. A Catá, por alem de revisar o trabalho me proporcionar um exemplo de
profissionalidade difícil de esquecer. A Hidasi por a companhia no campo, a ajuda nas

343
análises e o compartilhamento de experiências que tivemos ao longo do curso. Aos
meus revisores extraoficiais Sarah, Draxler e Léo, que se ofereceram para ajudar ao
gringo escrever um manuscrito com menos vomito, a Lorena e Renatinha por ajudar na
localização dos bichos alem de amenizar minha passagem por aqui. A o alto clero,
Paulinho Vampiro, pela troca de ensinamentos e a paciência tida. Aos monitores
Marcos e Thiago, por se dispor a ajudar com um sorriso sempre que precisei. Ao resto
do alto clero que me motivou e inspirou durante este curso, dentre eles Glauco, Marcel,
André, Fabrício, Felipe, Marcio, Jansen, Ester, Adriano... por demonstrar alegria e
disposição ao vir para o curso, transmitindo além de conhecimentos experiências e
inspirações tanto laborais como pessoais na troca de aparentemente nada. A aquela
pessoa que faz 21 anos teve a ideia, vontade e sacrifico de começar este curso de campo
que tantas vidas mudaram.
Por ultimo, a todos aqueles desconhecidos que ao longo do mês virarem amigos,
família, parceiros e até confidentes, gostei de compartilhar o leite condensado com
vocês. Ao Tatu, Randolpho, Daniel, Cibele, Mari, Tati, Joseph, Ana, Lud, Danizoo,
Bianca, Bruno, Ana Carol (os que já citei não repito!)... Cada uno de vosotros ya ocupa
un lugar en mi vida.
Terminando gostaria citar um refrão da música que acho apropriada para este
momento chamada “A hora do adeus” do Luis Gonzaga, com o que me identifico:
“...eu agradeço,
ao povo brasileiro,
norte, centro e sul inteiro
onde reina o baião...”
Obrigado galera!

344
Referências
Arnott, G. & R.W. Elwood. 2008. Information gathering and decision making about
resource value in animal contests. Animal behavior, 76:529-542.
Briffa, M. & R.W. Elwood. 2001. Motivational change during shell fights in the hermit
crab Pagurus bernhardus. Animal Behavior, 62:505-510.
Contreras-Garduño, J., J. Canales-Lazcano & A. Córdoba-Aguiar. 2006. Wing
pigmentation, immune ability, fat reserves and territorial status in males of the
rubyspot damselfly, Hetaerina americana. Journal of Ethology, 24:165-173.
Córdoba-Aguilar, A. 2008. Dragonflies and Damselflies: model organisms for
ecological and evolutionary research. Oxford:Oxford University Press.
Fitzstephens, D.M. & T. Getty. 2000. Colour, fat and social status in male damselflies,
Calopteryx maculata. Animal Behavior, 60:851-855.
Keil, L.P. & P.J. Watson. 2010. Assessment of self, opponent and resource during male-
emale contests in the sierra dome spider, Neriene litigiosa: Linyphiidae. Animal
Behavior, 80:809-820.
Mardem, J.H. & J.R. Cobb. 2004. Territorial and mating success of dragonflies that
vary in muscle power output and presence of gregarine gut parasites. Animal
behavior, 68:857-865.
Mesterton-Gibbons, M.; J.H. Marden & L.A. Dugatkin. 1996. On wars of attrition
without assessment. Journal of Theoretical Biology, 181:65-83.
Plaistow, S. & M.T. Siva-Jothy. 1996. Energetic constraints and male mate-securing
tactics in the damselfly Calopteryx splendens xanthostoma (Charpentier).
Proceedings of The Royal Society of London B, 263:1233-1239.

345
Resende, C.D. 2002. Comportamento territorial de Chalcopteryx scintilans (Odonata:
Polythoridae). In: Livro do Curso de Campo “Ecologia da Floresta Amazônica”
(Zuanon, J. & E. Venticinque, eds.).Manaus: INPA.
Ribeiro, J.E.L.S., M.J.G. Hopkins, A. Vicentini, C.A. Sothers, M.A. Costa, J.M. Brito,
M.A.D. Souza, L.H.P. Martins, L.G. Lohmann, P.A.C.L. Assuncao, E.C. Pereira,
C.F.D. Silva, M.R. Mesquita & L.C. Procopio. 1999. Flora da Reserva Ducke:
guia de identificação das plantas vasculares de uma floresta de terra-firma na
Amazônia Central. Manaus: INPA.
Schultz, T.D. & O.M. Fincke. 2009. Structural colors create a flashing cue for sexual
recognition and male quality in a Neotropical giant damselfly. Functional
Ecology, 23:724-732.




















