
L’exploitation des ressources animales pendant le 9e millénaire en Egée et le statut ambigu des...
26
MANEN C., PERRIN T., GUILAINE J. (eds) 2014 – La transition néolithique en Méditerranée. The Neolithic transition in the Mediterranean. Errance – AEP, 464 p. La Méditerranée constitue un espace privilégié pour étudier la mutation qui fit basculer les anciennes sociétés de chasseurscollecteurs dans la sphère des producteurs de nourriture, agriculteurs et éleveurs néolithiques, car fut à la fois un foyer de transition entre ces deux états et, parallèlement, un espace de diffusion de l’économie nouvelle : deux mécanismes clés pour comprendre le processus d’émergence du monde paysan. C’est d’abord sur les terres de la Méditerranée orientale et sur ses prolongements, depuis le Levant Sud jusqu’à à la Haute Mésopotamie et au Zagros, que des communautés ont progressivement modifié leur organisation sociale, leur cadre symbolique, leur mode de vie pour devenir des sociétés sédentaires et productrices, inaugurant ainsi une ère nouvelle, annonciatrice des temps historiques. Puis ce nouveau système s’est propagé en Méditerranée, favorisant ainsi la conversion à l’économie agricole et pastorale de l’Europe et d’une partie de l’Asie et de l’Afrique. Les mécanismes de cette diffusion furent complexes, entrainant de fréquentes recompositions culturelles et donnant lieu à processus adaptatifs commandés par le double jeu des contraintes environnementales et de la créativité humaine. Cet ouvrage constitue la publication d’un colloque international organisé en avril 2011 au Muséum de Toulouse. Il réunit vingtsix contributions qui dressent le panorama de la recherche actuelle en trois principaux domaines géographiques : le ProcheOrient, les îles de la Méditerranée orientale et la Méditerranée occidentale. The Mediterranean represents an ideal space for studying the transition from the last huntersgatherers to the first farmers. It was both a primal place of transition between these two steps and a space of diffusion of the new economy, two key mechanisms for understanding the process of the emergence of farming. It was first in the eastern Mediterranean, from the southern Levant to upper Mesopotamia and the Zagros, that some communities progressively modified their social organization, their symbolic framework, their way of life to become sedentary and foodproducing societies, thus opening a new era that set the pattern for historical times. Latter on, this new economic system progressively expanded, promoting the farming economy in Europe and in some part of Asia and Africa. The mechanisms of this diffusion were complex and they produced frequent cultural transformations and adaptive processes, both determined by environmental constraints and by human creativity. This book constitutes the proceedings of an international conference held in April 2011 in the Museum of Toulouse. It gathers twentysix papers offering an overview of the current research in three main geographical areas: the NearEast, the eastern Mediterranean islands and the western Mediterranean. http://www.librairieepona.fr/latransitionneolithiqueenmediterranee.html
Transcript of L’exploitation des ressources animales pendant le 9e millénaire en Egée et le statut ambigu des...

MANEN C., PERRIN T., GUILAINE J. (eds) 2014 – La transition néolithique en Méditerranée. The Neolithic transition in the Mediterranean. Errance – AEP, 464 p.
La Méditerranée constitue un espace privilégié pour étudier la mutation qui fit basculer les anciennes sociétés de chasseurs-‐collecteurs dans la sphère des producteurs de nourriture, agriculteurs et éleveurs néolithiques, car fut à la fois un foyer de transition entre ces deux états et, parallèlement, un espace de diffusion de l’économie nouvelle : deux mécanismes clés pour comprendre le processus d’émergence du monde paysan. C’est d’abord sur les terres de la Méditerranée orientale et sur ses prolongements, depuis le Levant Sud jusqu’à à la Haute Mésopotamie et au Zagros, que des communautés ont progressivement modifié leur organisation sociale, leur cadre symbolique, leur mode de vie pour devenir des sociétés sédentaires et productrices, inaugurant ainsi une ère nouvelle, annonciatrice des temps historiques. Puis ce nouveau système s’est propagé en Méditerranée, favorisant ainsi la conversion à l’économie agricole et pastorale de l’Europe et d’une partie de l’Asie et de l’Afrique. Les mécanismes de cette diffusion furent complexes, entrainant de
fréquentes recompositions culturelles et donnant lieu à processus adaptatifs commandés par le double jeu des contraintes environnementales et de la créativité humaine. Cet ouvrage constitue la publication d’un colloque international organisé en avril 2011 au Muséum de Toulouse. Il réunit vingt-‐six contributions qui dressent le panorama de la recherche actuelle en trois principaux domaines géographiques : le Proche-‐Orient, les îles de la Méditerranée orientale et la Méditerranée occidentale. The Mediterranean represents an ideal space for studying the transition from the last hunters-‐gatherers to the first farmers. It was both a primal place of transition between these two steps and a space of diffusion of the new economy, two key mechanisms for understanding the process of the emergence of farming. It was first in the eastern Mediterranean, from the southern Levant to upper Mesopotamia and the Zagros, that some communities progressively modified their social organization, their symbolic framework, their way of life to become sedentary and food-‐producing societies, thus opening a new era that set the pattern for historical times. Latter on, this new economic system progressively expanded, promoting the farming economy in Europe and in some part of Asia and Africa. The mechanisms of this diffusion were complex and they produced frequent cultural transformations and adaptive processes, both determined by environmental constraints and by human creativity. This book constitutes the proceedings of an international conference held in April 2011 in the Museum of Toulouse. It gathers twenty-‐six papers offering an overview of the current research in three main geographical areas: the Near-‐East, the eastern Mediterranean islands and the western Mediterranean.
http://www.librairie-‐epona.fr/la-‐transition-‐neolithique-‐en-‐mediterranee.html

SOMMAIRE
TransiTions en MédiTerranée - ou CoMMenT des Chasseurs devinrenT aGriCulTeurs
Jean Guilaine ....................................................................................................................................................... 9 Un colloque pour en débattre
la MuTaTion ProChe - OrienTale - The near - easT ChanGe
Frédéric abbès .................................................................................................................................................................... 13 Bal’as : un autre scénario de la néolithisation du Proche-Orient
The Bal’as Mountains: a different scenario of the Near Eastern neolithization
danielle stordeur ............................................................................................................................................................. 27 Jerf el Ahmar entre 9500 et 8700 cal. BC. Un village des débuts de l’agriculture. Une société complexe
Jerf el Ahmar between 9500 and 8700 cal. BC. A village at the outset of farming. A complex society
GeorGe Willcox ................................................................................................................................................................. 47 Les premiers indices de la culture des céréales au Proche-Orient
The beginnings of cereal cultivation in the Near East
adrian niGel GorinG-Morris, anna belFer-cohen .......................................................................................................... 59 The Neolithic in the southern Levant: yet another ‘unique’ phenomenon…
Le Néolithique dans le sud du Levant : un autre phénomène « singulier »…
MehMet Özdoğan ............................................................................................................................................................... 74 The Quest for New Criteria in Defining the Emergence and the Dispersal of Neolithic Way of Life
À la recherche de nouveaux critères pour définir l’émergence et la diffusion du mode de vie néolithique
éric coqueuGniot ............................................................................................................................................................... 91 Dja‘de (Syrie) et les représentations symboliques au IXe millénaire cal. BC
Dja’de (Syria) and the symbolic representations during the 9th millennium cal. BC
Miquel Molist ..................................................................................................................................................109 Le processus de consolidation de la néolithisation au Proche-Orient : Apports de l’étude du site de Tell Halula
(vallée de l’Euphrate, Syrie) The process of consolidation of the neolithization in the Near East: evidence from the site of Tell Halula (Euphrates valley, Syria)
la diffusion Par ChyPre, l’ÉGée eT l’adriaTique - The diffusion by CyPrus, eaGean and adriaTiC
Jean-denis ViGne ...............................................................................................................................................125 Nouveaux éclairages chypriotes sur les débuts de la domestication des animaux et sur la néolithisation au Proche-Orient
New insights from Cyprus on the beginning of animal domestication and on the neolithisation in the Near East
Katerina trantalidou .......................................................................................................................................141 L’exploitation des ressources animales pendant le 9e millénaire en Égée et le statut ambigu des suidés
Trends in faunal taxonomic representation during the 9th millennium in the Aegean and the ambiguous status of suids
aMelie scheu, ruth bollonGino, Jean-denis ViGne, anne tresset, norbert benecKe, JoachiM burGer ................165 The spread of domesticated cattle in the Neolithic transition
La diffusion du bœuf domestique durant la transition néolithique
niKos eFstratiou ...............................................................................................................................................173 “MICROHISTORIES” of transition in the Aegean islands. The cases of Cyprus and Crete
« MICRO-HISTOIRES » de la transition dans les îles Égéennes. Les cas de Chypre et de la Crète
adaMantios saMpson .........................................................................................................................................193 The Mesolithic of the Aegean basin
Le Mésolithique du bassin Égéén

Jean-François berGer, GaryFalia Metallinou, Jean Guilaine ........................................................................................ 213 Vers une révision de la transition méso-néolithique sur le site de Sidari (Corfou, Grèce).
Nouvelles données géoarchéologiques et radiocarbone, évaluation des processus post-dépositionnels Reconsidering the mesolithic-neolithic transition at the site of Sidari (Corfu, Greece). New geoarchaeological and radiocarbon data, evaluation of the post-depositional processes
stašo Forenbaher, preston Miracle .............................................................................................................................. 233 Transition to Farming in the Adriatic: a View from the Eastern Shore
La transition vers l’agriculture et l’élevage en Adriatique: une vue des côtes orientales
renata GriFoni creMonesi, GioVanna radi ..................................................................................................................... 243 Du Mésolithique au Néolithique ancien en Italie centrale et méridionale
From the Mesolithic to the Early Neolithic in central and southern Italy
la néoliThisaTion de l’ouesT MédiTerranéen - The WesTern MediTerranean neoliThizaTion
thoMas perrin, didier binder ......................................................................................................................................... 271 Le Mésolithique à trapèzes et la néolithisation de l’Europe sud-occidentale Late Mesolithic trapeze assemblages and the Neolithization of the South-Western Europe
Joseph cesari, patrice courtaud, FrancK leandri, thoMas perrin, claire Manen ................................................... 283 Le site de Campu Stefanu (Sollacaro, Corse-du-Sud) : une occupation du Mésolithique et du Néolithique ancien
dans le contexte corso-sarde Campu Stefanu (Sollacaro, Sourthern Corsica) : a Mesolithic and Early Neolithic settlement in the Corso-sardinian context
carlo luGliè ....................................................................................................................................................307 The Su Carroppu rockshelter within the process of neolithization of Sardinia L’abri sous roche de Su Carroppu et le processus de néolithisation de la Sardaigne
pilar utrilla, raFael doMinGo ..........................................................................................................................327 La transition Mésolithique-Néolithique dans la vallée de l’Èbre The Mesolithic-Neolithic transition in the Ebro valley
carMen olaria..................................................................................................................................................359 The Mesolithic collective burial of Cingle del Mas Nou (Ares del Maestre, Castellón, Spain) L’inhumation collective mésolithique du Cingle del Mas Nou (Ares del Maestre, Castellón, Espagne)
JaVier Fernández-lópez de pablo .......................................................................................................................371 Art traditions, cultural interactions and symbolic contexts during the Neolithic transition in the Eastern Iberian Peninsula Traditions artistiques, interactions culturelles et contextes symboliques de la transition néolithique dans la région
méditerranéenne espagnole
claire Manen ...................................................................................................................................................405 Dynamiques spatio-temporelles et culturelles de la néolithisation ouest-méditerranéenne Spatial, chronological and cultural dynamics of the neolithization in the western Mediterranean
Joan bernabeu aubán, bernat Martí oliVer .....................................................................................................419 The First Agricultural Groups in the Iberian Peninsula Les premiers groupes agropastoraux de la péninsule Ibérique
Marie lacan, François-xaVier ricaut, bertrand ludes, éric crubézy, Jean Guilaine ..........................................439 La néolithisation de l’Europe : apports de l’ADN ancien The neolithization of Europe: evidence from ancient DNA
siMone Mulazzani .............................................................................................................................................453 Le site de SHM-1 (Tunisie) entre le VIIe et le VIe mill. cal. BC. Prémices d’une transition du Capsien supérieur vers le Néolithique The site of SHM-1 (Tunisia) between the 7th and the 6th mill. cal. BC. Evidence of Upper Capsian – Neolithic transition
liste des auteurs ..............................................................................................................................................463

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
141141
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
Les données obtenues à partir de l’analyse des ossements et des mollusques des sites de Maroulas sur l’île de Kythnos dans les Cyclades (8930-8410 cal. BC), de la grotte du Cyclope sur l’île de Youra dans les Sporades du nord (8626-8290 cal. BC) et de la grotte Franchthi dans l’est du Péloponnèse (8400-7650 cal. BC) révèlent que de petites communautés pré-agricoles du IXe millénaire avaient fondé leur subsistance sur les coquil-lages marins, les poissons, les oiseaux et les ongulés. Sur les îles, les suidés introduits étaient sous le contrôle de l’homme. De plus, l’isolement insulaire, sorte d’enclos naturel, a joué sur la taille des animaux.
Dans le présent article, les traits principaux des restes fauniques sont examinés. Les recherches génétiques sur les changements controversés de la morphologie des suidés
L’EXPLOITATION DES RESSOURCES ANIMALES PENDANT LE 9e MILLÉNAIRE EN ÉGÉE ET LE STATUT AMBIGU DES SUIDÉS
TRENDS IN FAUNAL TAXONOMIC REPRESENTATION DURING THE 9th MILLENNIUM IN THE AEGEAN AND THE AMBIGUOUS STATUS OF SUIDS
KaTerina TranTalidou
RÉSUMÉ
progressent. Le carbone stable et les isotopes de l›azote ont été mesurés sur les os de la grotte du Cyclope et les analyses ont prouvé que les suidés avaient plutôt un régime herbivore sans consommation de ressources marines. Les analyses du strontium ont également été appliquées à l’assemblage de Kythnos et ont permis de détecter des déplacements d’hu-mains et d’animaux, mais ces résultats restent à clarifier. Petit à petit, les suidés sont remplacés par les caprinés. Dans la grotte du Cyclope, les chèvres apparaissent très tôt dans la séquence stratigraphique. Leur apparition précoce peut être interprétée, soit en termes d’intrusion depuis une couche sus-jacente, soit comme le résultat de premiers échanges entre des communau-tés élevant des animaux et celles conservant une technologie mésolithique.
Bone and mollusc records stemming from the settlements of Maroulas on Kythnos in the Cyclades (8930-8410 cal. BC), the Cyclops Cave on Youra, in the Northern Sporades (8626-8290 cal. BC) and the Franchthi cave in the Eastern Peloponnese (8400-7650 cal. BC) reveal that the small pre-agricultural communities of the 9th millennium based their meat consumption upon fish-shellfish, birds and ungulates. On islands, suids seemingly have been kept under a certain control, a kind of human domination over them, coupled with the fact that islands are natural enclosures, influencing on the size of the animals.
In this contribution, the main features of the faunal data are examined. Genetic research is in progress in order to investigate
ABSTRACT
the controversial subject of the morphological changes observed for the suids. Stable carbon and nitrogen isotope analyses have been also used on bones from the Cyclops cave and they proved that the boar diet was an herbivore diet without consummation of marine resources. Strontium analysis has also been applied to the assemblage from Kythnos to detect movements of people and animals but the results remain unclear. Progressively, the suids have been replaced by caprins. At Cyclops Cave goats appear quite early within the stratigraphic sequence. They are either intrusive due to post-depositional phenomena or stem from exchange between groups keeping animals and groups with Mesolithic technology.

142142
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
Fig.1 : Carte des sites de la transition Pléistocène-Holocène mentionnés dans le texte (fond de carte M. P. Deilaki - Ephorate for Palaeoanthropology-Speleology).
Map of Late Pleistocene and Early Holocene sites discussed in the text. By M. P. Deilaki (Ephorate for Palaeoanthropology-Speleology).

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
143143
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
Le cadre culturel des groupes du IXe millénaire est défini à partir des caractéristiques technologiques de l’outillage lithique (Runnels et Van Andel, 2003) et repose traditionnellement sur la chronologie relative établie à partir de stratigraphies. La grotte de Zaimis en Attique, actuellement détruite, et celle de Franchthi en Argolide, toutes deux situées dans des péninsules surplombant la mer Égée, figurent parmi les premiers sites explorés. Elles ont livré des couches qui contenaient des industries du Mésolithique, caractérisées par la production de lamelles (Galanidou et Perlès, 2003). La séquence évolutive du Mésolithique comprend le Mésolithique ancien, le Mésolithique récent et le Mésolithique final (fig. 1). Ces données archéologiques confrontées aux mesures radiocarbone 1 datent la séquence mésolithique entre la deuxième moitié du IXe millénaire et la deuxième moitié du VIIe millénaire avant notre ère (Sampson, 2006).
Les analyses polliniques ont permis de mettre en évidence une diversification de la végétation en fonction de paramètres tant géographiques que chronologiques (Bottema, 2003, incluant la littérature plus ancienne). Pour citer un exemple : durant la dernière moitié du IXe et le premier quart du VIIIe millénaire, des plantes herbacées appartenant aux familles des Poaceae, Ranunculaceae et Rosaceae, occupaient l’île de Youra, située dans le nord de l’archipel des Sporades, cette flore indiquant un climat froid et sec (Ioakeim, 2011). Les charbons de bois retrou-vés dans les niveaux 23-24 de la grotte du Cyclope révèlent la présence du genévrier, qui tolère les sols pauvres, calcaires et qui colonisait probablement les falaises de l’île. Espèce survivante
1 Les résultats sont indiqués en BP, parfois en tenant compte des facteurs de correction de l’effet « réservoir marin », et en dates calibrées (Facorellis, 2003 et 2011 ; Facorellis et al., 2010).
LES CAMPEMENTS DU IXe MILLÉNAIRE EN GRÈCE ET DANS LES ÎLES ÉGÉENNES
du Pléistocène, le genévrier cohabitait probablement avec des arbustes du genre Phillyrea-Rhamnus (Ntinou, 2011). Par la suite, la couverture forestière composée de chênes à feuillage persistant et de pins était accompagnée de plantes des genres Phillyrea-Rhamnus, Arbutus, de légumineuses et du gainier (Cercis siliquastrum). Ces espèces étaient présentes dans la partie centre-ouest de l’île, à falaises abruptes (Ioakeim, 2011 ; Ntinou, 2011).
Tous les sites attestent d’un mode de vie de type chas-seur-cueilleur. En effet, les macrorestes identifiés ne révèlent pas la présence de plantes domestiquées (Franchthi : Hansen, 1991 et 1992 ; grotte du Cyclope : Sarpaki, 2011). La question de l’exploitation des plantes par les occupants de la grotte Theopetra est sujette à des discussions, car sa stratigraphie est considérée comme perturbée (Mangafa, 2000 ; Sarpaki, 2011 ; Valamoti et Kotsakis, 2007).
Les amas coquilliers de mollusques terrestres, notamment à Helix figulina, découverts au sein des stratigraphies (Lubell, 2004), montrent qu’il s’agit de restes de consommation. C’est le cas dans les trois couches superposées de coquilles d’escar-gots mises en évidences dans les huttes circulaires de Maroulas sur l’île de Kythnos (Alexandrowicz, 2010) ou encore dans les dépôts de la grotte du Cyclope sur l’île de Youra (Karali, 2011). Mais il est aussi possible que leur exploitation ait varié au cours du temps. Dans les niveaux du Paléolithique final de Franchthi, par exemple, les coquilles d’escargots sont intactes tandis que dans les niveaux mésolithiques elles sont fragmen-tées (Farrand, 2000 et 2003). Enfin, de grandes quantités de restes des coquillages marins sont ubiquistes, surtout dans les sites côtiers.
Les publications concernant les niveaux acéramiques de la Grèce continentale (Boessneck, 1962) et de la Crète (Jarman et Jarman, 1969 ; Isaakidou, 2004) montrent la présence bien éta-blie d’animaux domestiques dans la région égéenne au cours du VIIe millénaire cal. BC. Cependant, l’histoire de la domestication des animaux en Méditerranée orientale est encore mal connue. Actuellement, seul l’environnement de quelques sites prénéo-lithiques a été étudié. Les traits généraux sont les suivants :
- La faune présente pendant le changement climatique de la transition Pléistocène – Holocène était composée d’une grande variété d’espèces. L’Épire est la région qui abrite les vestiges les mieux conservés de la transition Paléolithique–Mésolithique, grâce à des conditions géomorphologiques particulières. La chaîne du Pinde constitue une barrière qui isole le territoire épi-rote du reste de la péninsule. De ce fait, les communications et les affinités culturelles se sont davantage établies avec l’Adriatique et la partie nord-ouest des Balkans qu’avec les zones côtières
de la mer Égée. Dans les vallons de l’arrière-pays, bouquetins et chamois constituaient le gibier par excellence à la fin de l’époque glaciaire (fig. 2). À Klithi dans le bassin du Voidomatis, ils réunissent à eux seuls 95% de la totalité des restes osseux. Les animaux adultes constituent la principale cible des chasseurs, même si 30% des animaux tués sont de jeunes animaux (Gamble, 1999). Les animaux adultes ont également été sélectionnés par les chasseurs qui fréquentaient l’abri de Boila, localisé dans le même défilé (Kotjabopoulou, 2001). Dans le bassin d’Ioannina, à Kastritsa, le cerf prédomine sur les bouquetins et les chamois et représente 65% des fragments osseux récoltés. Ce sont majo-ritairement les jeunes cervidés adultes qui ont été chassés. En revanche, les restes de l’hydrontin (Equus hydruntinus), un animal dont la chasse demeure secondaire, montrent que des jeunes animaux étaient également abattus (Kotjabopoulou, 2001). L’inventaire des espèces chassées à l’abri Grava Gardikiou (altitude 60 m) dans le sud de l’île de Corfou inclut l’aurochs, les
LA FAUNE DU SUD DE LA PÉNINSULE BALKANIQUE DURANT LA TRANSITION PLÉISTOCÈNE-HOLOCÈNE

144144
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
28
84
747
14 14
64 51
6
133
14
5679
11 14
20
202
10
42
1
725
225
6
21
36
64
40
24
1
10
100
1000
10000
Bovida
e
Caprin
ae
Cervus
elap
hus
Capreo
lus ca
preolu
s
Sus sc
rofa
Equus
hydru
ntinu
s
Carnivo
ra
Lago
morpha
Ave
s
Pisces
Other
Kastritsa, stratum 1 : 13400 + 210 BP Klithi : 16000 - 13000 BPBoïla IIIb : 14310 ±100 BP
Fig. 2 : Graphique illustrant la forte fragmentation de la faune du Paléolithique supérieur provenant de trois abris sous roche de l’arrière-pays de l’Épire
(données modifiées d’après Gamble, 1999 ; Kotjabopoulou, 2001).
Illustration of the highly fragmented Upper Palaeolithic bone assemblages stemming from three rock shelter sites in the hinterland of Epirus (modified after Gamble, 1999; Kotzabopoulou, 2001).
Fig. 3 : Matériel osseux (cervidés, équidés) extrêmement fragmenté, provenant d’une grotte anonyme à Schistos, Keratsini, sur le mont Aegaleon, Attique. Dans ce site, on observe une chasse ciblée sur le
petit gibier qui semble constituer la tendance à cette période. Photographie : K.Trantalidou.
Abundant faunal material (Cervids, Equids) extremely fragmented coming from a anonymous cave at Schistos, Keratsini, on mount Aegaleon, Attica. In this site, one can observe the hunting of small game is
the main trend at that period. Photograph: K.Trantalidou.
La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
145145
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
trois espèces de cervidés (cerf, chevreuil, daim), les suidés, les équidés et des espèces d’oiseaux (Sordinas, 1969). Tous ces sites ont été abandonnés vers le début de l’Holocène (Runnels et Van Andel, 2003).
Pendant le Mésolithique, on observe une migration impor-tante des groupes humains vers les sites côtiers. Sidari, site de plein air dans le nord de l’île de Corfou, présente un niveau du Néolithique ancien séparé d’un niveau du Mésolithique (niveau D d’A. Sordinas daté de 7770 ± 340 BP (7515 – 6031 cal. BC) qui se caractérise par l’exploitation des ressources marines et par une industrie microlithique (fouilles de Sordinas : Runnels et Murrey, 2001). Les dernières fouilles entreprises (Berger, Metallinou et Guilaine, ce volume) ont mis en évidence une stratigraphie plus complexe et il est possible que l’horizon mésolithique ne contienne que des restes d’animaux sauvages (faune peu abondante : E. Psathi, communication personnelle). La faune mésolithique de Konispol, une grotte située sur le continent dans le massif de Saraqint, à une altitude de 400 m au bord de la rivière Pavel, en face du détroit de Corfou (Petruso et al., 1996), contenait des restes de bouquetin, de chamois, de cerf élaphe, de sanglier ainsi que de la microfaune. Les restes indiquent une occupation au cours du printemps et de l’été (Leblanc, 1996). Une remarque supplémentaire sur l’économie s’impose : les outils en silex de tous les sites mésolithiques du littoral de la Mer ionienne sont en relation avec des activités de chasse (Runnels et Van Andel, 2003).
Malgré la diversité des espèces, le bouquetin, le cha-mois, l’aurochs, les cervidés et l’âne sauvage constituent les ressources de base durant le Paléolithique supérieur - final. La chasse, le ramassage des escargots, la cueillette, la pêche et la sédentarisation croissante seraient liés à une amélioration du cli-mat. À titre d’exemple, la composition de la faune et le mode de
subsistance des quatre gisements répertoriés (grotte de Schistos, Theopetra, Franchthi, Klissoura I) seront plus particulièrement développés.
LA GROTTE DE SCHISTOS
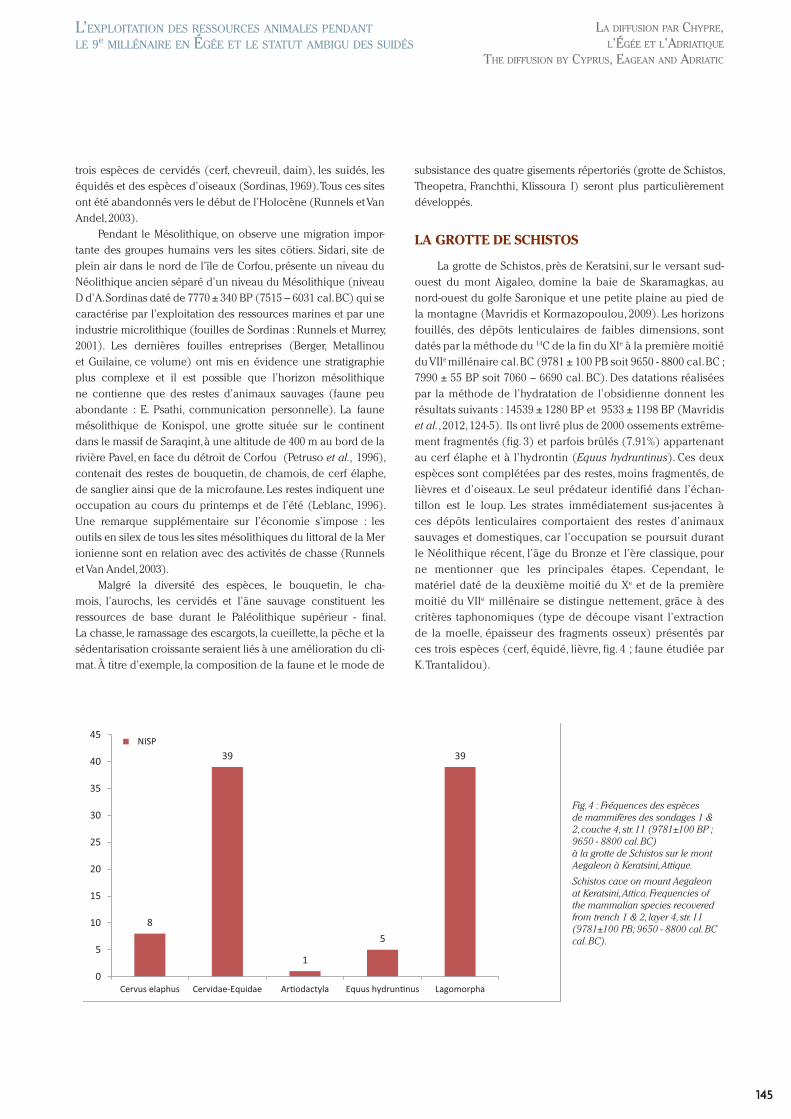
La grotte de Schistos, près de Keratsini, sur le versant sud-ouest du mont Aigaleo, domine la baie de Skaramagkas, au nord-ouest du golfe Saronique et une petite plaine au pied de la montagne (Mavridis et Kormazopoulou, 2009). Les horizons fouillés, des dépôts lenticulaires de faibles dimensions, sont datés par la méthode du 14C de la fin du XIe à la première moitié du VIIe millénaire cal. BC (9781 ± 100 PB soit 9650 - 8800 cal. BC ; 7990 ± 55 BP soit 7060 – 6690 cal. BC). Des datations réalisées par la méthode de l’hydratation de l’obsidienne donnent les résultats suivants : 14539 ± 1280 BP et 9533 ± 1198 BP (Mavridis et al., 2012, 124-5). Ils ont livré plus de 2000 ossements extrême-ment fragmentés (fig. 3) et parfois brûlés (7.91%) appartenant au cerf élaphe et à l’hydrontin (Equus hydruntinus). Ces deux espèces sont complétées par des restes, moins fragmentés, de lièvres et d’oiseaux. Le seul prédateur identifié dans l’échan-tillon est le loup. Les strates immédiatement sus-jacentes à ces dépôts lenticulaires comportaient des restes d’animaux sauvages et domestiques, car l’occupation se poursuit durant le Néolithique récent, l’âge du Bronze et l’ère classique, pour ne mentionner que les principales étapes. Cependant, le matériel daté de la deuxième moitié du Xe et de la première moitié du VIIe millénaire se distingue nettement, grâce à des critères taphonomiques (type de découpe visant l’extraction de la moelle, épaisseur des fragments osseux) présentés par ces trois espèces (cerf, équidé, lièvre, fig. 4 ; faune étudiée par K. Trantalidou).
Fig. 4 : Fréquences des espèces de mammifères des sondages 1 & 2, couche 4, str. 11 (9781±100 BP ; 9650 - 8800 cal. BC) à la grotte de Schistos sur le mont Aegaleon à Keratsini, Attique.
Schistos cave on mount Aegaleon at Keratsini, Attica. Frequencies of the mammalian species recovered from trench 1 & 2, layer 4, str. 11 (9781±100 PB; 9650 - 8800 cal. BC cal. BC).

146146
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
LA GROTTE DE THEOPETRA
En Grèce continentale, trois grottes ont livré une stratigra-phie conséquente, bien que comportant des hiatus importants entre le Paléolithique moyen, la transition Pléistocène-Holocène et le Néolithique. Il s’agit de la grotte de Theopetra en Thessalie, de la grotte de Franchthi et de l’abri Kleissoura 1 en Argolide (Péloponnèse). Dans la grotte Mouta à Theopetra, près des Météores, la séquence sédimentaire observée atteint une épais-seur de 4 m à 6 m dans sa partie centrale. La couche mésolithique possédait une épaisseur de 0,20 m et, d’après les 17 mesures radiométriques (situées entre 9940 - 8550 et 7060 - 6780 cal. BC), elle représentait 1700 à 2000 ans d’occupations discontinues (Kyparisi-Apostolika, 2000 et 2003 ; Adam, 2000 ; Karkanas et Weiner, 2000 ; Facorellis, 2003 ; Facorellis et al., 2001). Tout au long du Paléolithique supérieur et du Tardiglaciaire (14895 ± 181 BP soit 16600 – 15950 cal. BC ; 10972 ± 87 BP soit 11030 – 10870 cal. BC), la faune (299 restes osseux) se composait d’une multitude d’espèces : bovidés (aurochs et bouquetin), cervidés (cerf, che-vreuil et daim dans les niveaux les plus bas), équidés, suidés, carnivores (ours, hyène, chat sauvage, loup, renard, blaireau, cas-tor), léporidés, oiseaux aquatiques ainsi que rapaces et reptiles (Newton, 2000 et 2003). Une soixantaine d’ossements provient des couches mésolithiques (cerf, sanglier, chat sauvage, renard, loup, lièvre, oiseaux, reptiles). Cet assemblage est sujet à discus-sion dans la mesure où 27 des 60 ossements appartiennent à de jeunes ovicaprinés. Il pourrait probablement s’agir d’intrusions à partir des couches supérieures (Newton, ibidem).
Un examen rapide du matériel récolté lors des fouilles de 2002 à 2005 a permis les observations suivantes :
- Les ossements mésolithiques, peu abondants par rapport au nombre d’espèces, et l’horizon archéologique, peu développé, suggèrent la fréquentation du site par un petit groupe d’individus au début de l’Holocène.
- Le matériel est caractérisé par une importante fragmenta-tion en spirale des diaphyses de mammifères de taille moyenne à grande. Ces esquilles en spirale témoignent de l’extraction de la moelle. Les os des lièvres et des oiseaux sont nettement moins fragmentés. Des carapaces de tortues et des os longs de grenouilles sont également présents.
- Les os des grands mammifères sont légèrement concré-tionnés (les sédiments ont probablement ont subi l’action de l’eau ; Karkanas et al., 1999) et/ou brûlés.
- De toute évidence, les vestiges mésolithiques sont inclus dans des dépôts lenticulaires de faible dimension.
- D’un point de vue taphonomique, ces espèces se dis-tinguent par rapport aux ossements des chèvres et des moutons, parfois prélevés dans le même sac plastique lors de la fouille. Tous ont vraisemblablement été retrouvés dans le même mètre carré de la fouille, mais appartiennent à deux périodes diffé-rentes (observations de K. Trantalidou).
LA GROTTE DE FRANCHTHI
La longue séquence stratigraphique des couches du Paléolithique supérieur, du Mésolithique et du Néolithique mise en évidence dans la grotte de Franchthi a permis de distinguer trois niveaux mésolithiques distincts datés entre 9340 ± 160 BP (soit 9150 – 8250 cal. BC) et 9150 ± 100 BP (soit 8700 – 8200 cal. BC) ; 9480 ± 130 BP (soit 9250 – 8450 cal. BC) et 8530 ± 90 BP (soit 7760 – 7350 cal. BC) ; 8190 ± 80 BP (soit 7460 – 7040 cal. BC) et 8020 ± 80 BP (soit 7180 et 6680 cal. BC) ; 15 échantillons ont été prélevés dans une coupe d’une hauteur de 3 m : Perlès, 2001; Facorellis, 2003 ; Stroulia, 2010). Ces niveaux s’individualisent par un changement aussi bien dans l’outillage lithique (phases VII, VIII, IX : Perlès, 1987 et 1990) que dans la faune (Payne, 1975
7
187
17
630
6
797
4 24
639
35
197
606
104
255
2 1
67 65 67
303
103
16
639
18 11
892
0
100
200
300
400
500
600
700
800
900
1000
Capra
sp.
Cervus
elaph
us
Felis s
ilvest
ris
Musteli
dae
Erinace
us eu
ropaeu
s
Alector
is sp.
Sparid
ae
Conge
r con
ger
Pisces
Emys
Testu
do
Microfau
na
Sus scr
ofa
Bos pri
migeniu
s
Equus
hydru
ntinu
s
Ungula
te ind
.
Felis l
ynx
Vulpes
vulpe
s
Carnivo
ra
Lepus
europ
aeus
Small M
ammali
a
Otis ta
rda
Falcon
iform
es
Aves in
d.
Thunn
us thy
nnys
Sphyra
ena s
phyra
ena
Fig. 5 : Spectre faunique des couches du Méso-lithique ancien et récent (c. 8350-7650 cal. BC) de la grotte de Franchthi – modifié d’après Stiner et Munro, 2011.
Faunal spectra from the Lower and Upper Mesolithic layers of Franchthi Cave (c. 8350-7650 cal. BC) - modified after Stiner and Munro, 2011.

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
147147
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
et 1982). Vers la fin du Paléolithique, le régime carné unique-ment axé sur les mammifères terrestres se diversifie en exploi-tant l’environnement marin (fig. 5). Le cerf élaphe, gibier par excellence, et le sanglier sont les ongulés les plus chassés. Le lièvre constitue un complément important dans le régime alimentaire. La pêche s’intensifie vers la fin du Mésolithique et le thon domine les assemblages de restes de poissons (Rose, 1995). L’ensemble indique que la pêche se pratiquait dans des eaux profondes, loin du rivage. Durant le Mésolithique récent, la grotte semble avoir été fréquentée tout au long de l’année (données obtenues à partir des mollusques marins : Shackleton, 1988). Le pourcentage global d’os calcinés contenus dans les couches mésolithiques est de 24%, bien supérieur à celui des dépôts paléolithiques (Stiner et Munro, 2011).
L’ABRI DE KLEISSOURA 1
L’abri de Kleissoura 1 se situe à l’entrée de la gorge du même nom, sur la rive droite de l’actuel ruisseau Berbadiotis. Le site domine la plaine et la baie de l’Argolide (Koumouzelis et al., 2001). Plusieurs complexes sédimentaires en constituent
le remplissage, formé dès le Paléolithique moyen. Une seule date a été obtenue à la base de la séquence mésolithique : 9150 ± 220 BP soit 9200 – 7700 cal. BC (Koumouzelis et al., 2004).
À partir de l’Aurignacien, pendant l’Épigravettien et le Mésolithique, la présence d’une grande variété d’ongulés (fig. 6) reflète la composition du milieu. Dans la mesure où le daim représente la moitié des herbivores de l’assemblage faunique, l’environnement devait correspondre à un milieu ouvert et sec. Puis la chasse s’oriente progressivement vers le petit gibier. L’absence du squelette axial du daim atteste que le découpage avait lieu à un endroit qui n’a pas été fouillé à l’intérieur de la grotte ou en dehors de celle-ci. D’après les âges des cervidés abattus, le site était fréquenté entre la fin de l’été et le début du printemps (Starkovich et Stiner, 2010). Les ossements sont concrétionnés et portent des fractures trans-versales et en spirale. Les cavités médullaires ont été ouvertes pour en extraire la moelle (Starkovich et Stiner, 2010 ; obs. personnelles). Le mode de découpe et de fracturation des os (fig. 7) rappelle celui pratiqué à Theopetra durant la même période. Les deux gisements en Argolide témoignent ainsi de multiples activités sur le plan économique.
Fig. 6 : Péninsule de l’Argolide. Tendances dans l’économie de chasse durant le Mésolithique. Fréquence relative des ossements (données modifiées d’après Starkovich et Stiner, 2010 ; Stiner et Munro, 2011).
Argolid Peninsula. Trends in hunting economy throughout the whole Mesolithic sequence. Relative abundance of the bones (data modified after Starkovich and Stiner, 2010; Stiner and Munro, 2011).

148148
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
Les fouilles effectuées sur les îles de la partie orientale de la mer Égée, par exemple à Keramé sur l’île d’Icarie (Kaczanowska et Kozłowski, 2006 ; Sampson et al., 2009 et à Gavdos en Crète (Kopaka et Matzanas, 2009 ; Plakias : Strasser et al., 2010), n’ayant, pour le moment, livré aucun fragment de faune associé aux pièces lithiques, on décrira plutôt les dépôts datés du Mésolithique et ceux en relation avec les processus de néolithisation dans les îles à l’ouest de la mer Égée (Kythnos et Youra).
LE SITE DE MAROULAS
Les principales caractéristiques de Maroulas, un site de plein air, considérablement érodé, localisé sur le promontoire du même nom, dans le nord-est de l’île de Kythnos, sont les suivantes:
- Les restes de huttes de plan circulaire ou elliptique avec un pavage en dalles de schiste ou en galets ont été révélés (Sampson, ce volume). Leur diamètre, délimité parfois par des dalles verticales, varie entre 3 et 4 m. Une construction semi-enterrée, creusée dans le rocher, comprenant trois sols d’occupation superposés, a également été identifiée (Sampson, 2006). De même, des concentrations de pierres ont été conservées.
Sur une surface de 2000 m², les contours de 31 huttes ont été dégagés. Des constructions similaires ont été mises au jour dans d’autres gisements en Méditerranée orientale (Sampson et al., 2010).
- Vingt-cinq sépultures, couvertes d’ocre rouge, aménagées dans des cuvettes naturelles ou artificielles, creusées dans le substrat géologique en schiste, sous le pavage des cabanes, ont été dégagées. La découverte de plusieurs squelettes acéphales et celle de crânes isolés démontrent la présence d’inhumations secondaires.
- La technologie lithique semble dériver de l’Épigravettien de la Grèce continentale, un trait également constaté pour le matériel de la grotte du Cyclope sur l’îlot de Youra. L’île de Kythnos est relativement proche de l’Attique et des côtes orien-tales du Péloponnèse. Les microlithes y étaient peu abondants, les occupants se consacrant plutôt à la cueillette qu’à la chasse (Kaczanowska et Kozłowski, 2006).
- Le macro-outillage (broyeurs, pierres à aiguiser, meules) ne trouve pas de parallèle dans les sites de la péninsule Balkanique. En revanche, des pièces similaires se retrouvent dans le Natoufien, le Zarzien et le Néolithique précéramique de la vallée du Jourdain ou du littoral syro-palestinien (Sampson et al., 2010).
Fig. 7 : Abri de Kleissoura 1, Argolide. La faune est extrêmement fragmentée. Photographie : K. Trantalidou.
Rock shelter of Kleissoura 1, Argolis. The fauna is highly fragmented. Photograph: K. Trantalidou.
LA FAUNE DES STATIONS MÉSOLITHIQUES DES ÎLES DE L’OUEST DE LA MER ÉGÉE

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
149149
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
- Les 24 datations radiocarbone situent l’occupation du site entre 8930 et 8410 cal. BC (Facorellis et al., 2010).
- Sur les 1193 restes osseux, 655 appartiennent à des pois-sons migrateurs (thon, bonites, maquereaux) et côtiers (fig. 8). Les poissons ont vraisemblablement été conditionnés, fumés ou salés, une activité également attestée dans la grotte du Cyclope sur l’île de Youra, qui a livré une ichtyofaune comparable (Mylona, 2010 et 2011).
- Tous les ossements, humains et fauniques, étaient couverts d’une croûte de couleur grise. Parmi eux, 7.78% des restes de poissons et 21.07% des restes de mammifères et de l’avifaune portaient des traces de brûlures ou étaient complètement cal-cinés. La coloration gris foncé pourrait être due à la présence de sédiments cendreux (Karkanas, communication personnelle).
Après une fréquentation plus ou moins régulière durant environ 500 ans, le site a été abandonné. Il a été à nouveau occupé durant le Bronze ancien, apparemment pour une période courte, si l’on prend en compte l’épaisseur des couches. Le matériel osseux des dépôts postérieurs se distingue facile-ment. Il ne possède pas la même morphologie de fractures et les résidus grisâtres sur les ossements sont absents. De plus, les espèces représentées se limitent aux chèvres et aux moutons (Trantalidou, 2010).
LA GROTTE DU CYCLOPE
La grotte du Cyclope (d’une surface de 50 m x 40 m) est située dans la partie sud-ouest de l’îlot de Youra (au
nord-ouest de la mer Égée). Par endroits, le remplissage atteint une épaisseur comprise entre 3 m et 4,40 m. Ces sédiments ont été accumulés tout au long de l’Holocène (Sampson, 2008 et ce volume). Les observations se référant à cet ensemble mésolithique, appuyées par une importante série de datations absolues, sont les suivantes :
- Les niveaux mésolithiques, qui constituaient l’horizon de base, ont été identifiés sur une hauteur de 1,35 m à 2,85 m (tranchée C, partie ouest et partie est). Sept niveaux d’occupation peuvent être attribués à cet ensemble tandis que cinq niveaux ont été identifiés pour l’ensemble néolithique qui lui succède. Les vestiges archéologiques datés du Néolithique (ancien, moyen et récent, précisément datés) étaient beaucoup plus abondants. Il est fort probable que des niveaux néolithiques aient été détruits par des occupations postérieures d’une plus grande ampleur (piétinement, remaniement des sédiments pour niveler des hori-zons ou pour d’autres constructions, etc.).
D’après les observations faites à la fouille, la nappe argileuse qui forme le sol 2, partout où elle est attestée, semble sceller un tapis épais de coquillages et de restes de poissons. Il s’agit très probablement d’un aménagement qui a perduré pendant plusieurs siècles. Des exemples similaires ont été repérés dans plusieurs grottes (Trantalidou et al., 2011).
2 Elle a été localisée dans les mètres carrés (tranchée, couche, carré, numéro de couche) suivants : Mésolithique ancien : C/E, 21, 1-2 (12)/ C/E, 21, 5-6 (12a) ; C/E, 19, 1 (11) ; C/E, 18, 1-2 (10); C/E, 18, 10 (10b)/ C/W, 12, 4 (7a)/ C/W, 12, 2 (7) - C/W, 11, 4 (7) ; Mésolithique récent : C/W, 10, 1-5 ; C/W, 9, 4 (6)/ C/W, 9, 1&3 (5) ; Mésolithique final : C/E, 18,7a (10a).
37
5
12
1
6
18
5
1 1
67
8
3 1 1
5
35
31
7 10
12
3
25
1 0
10
20
30
40
50
60
70
80
Muraen
idae
Conge
r con
ger
Muraen
idae/C
ongri
dae
Zeus f
aber
Serran
idae
Epinep
helus
sp.
Serran
us cab
rila/sc
riba
Decentr
archu
s labra
x
Umbrina
cirro
sa
Sparid
ae ind
et. sp
.
Dentex
dente
x
Diplod
us sar
gus
Pagrus
pagru
s
Labrid
ae
Trachin
idae
Scombri
dae i
ndet.
sp.
Euthyn
nus a
llette
ratus
Sarda s
arda
Scombe
r japo
nicus/
scom
brus
Thunn
us sp.
Mugilli
dae
Scorpa
enida
e
Pleuron
ectifo
rmes
Fig. 8 : Les différentes espèces de poissons déterminées (données d’après Mylona, 2010) sur le site de Maroulas à Kythnos
Maroulas on Kythnos. Identified fish species (data after Mylona, 2010).

150150
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
Les couches présentaient une forte inclinaison vers le sud et vers l’ouest due à l’affaissement des sédiments. C’est la raison pour laquelle, seuls les mètres carrés au centre ont été sélec-tionnés pour l’ensemble des échantillons prélevés (Sampson, 2008). Toutefois, l’accumulation des sédiments en aval, l’affais-sement des couches, le transport des sédiments par le vent, l’érosion du sol ou le ruissellement sont les conséquences de processus post-sédimentaires parfois rencontrés dans les grottes (Trantalidou, 2011). Dans la grotte de Schistos, un phénomène similaire se manifeste.
- Les 20 datations radiocarbone réalisées sur des charbons de bois et des mollusques ont livré les âges absolus suivants (BP non calibré): 9258 ± 50 BP (C/W, L12, sq.1-2), 9252 ± 31 BP (C/E, L23, sq.7) et 9250 ± 60 BP (C/W, stalagmite, sq.3) ; 9056 ± 28 BP et 8791 ± 23 BP (C/E, L20, sq.5), 8864 ± 37 BP et 8834 ± 20 BP (C/E, L14, sq.1), 8855 ± 28 BP et 8776 ± 19 BP (C/W, L11, sq.4), 9042 ± 24 BP, 9011 ± 22 BP et 8487 ± 22 BP (C/E, L19, sq.5), 8209 ± 47 BP (C/W, L10, sq.1-2), 8754 ± 20 BP et 8624 ± 20 BP (C/W, L9, sq.5), 8773 ± 24 BP et 8283 ± 27 BP (C/E, L17, sq.5), 8761 ± 29 BP (C/E. L15, sq.5), 8218 ± 43 BP (C/W, L8, sq.3-4), 7779 ± 32 BP (C/E, L16, sq.6). Les dates 7803 ± 41 BP et 7971 ± 41 BP se situeraient vers la fin du Mésolithique et marqueraient la transition vers le début du Néolithique (Sampson, 2006 ; Facorellis, 2011).
Des datations effectuées sur des dents des suidés, issues des couches du Mésolithique récent et du Néolithique, ont confirmé les données précédentes réalisées au laboratoire Demokritos, à Athènes. Dans la tranchée C (partie est, couche 19, carré 5) il a été possible d’obtenir trois paires de dates mesurées sur des échantillons de mollusques terrestres, de coquillages marins et de charbons de bois. Les Patella ulyssiponensis Gm. ont livré les dates suivantes, exprimées en cal. BC corrigées de l’effet « réser-voir marin » : 7675 - 7462 (95.4%) / 7590 - 7507 (68.3%) cal. BC (Facorellis, 2011). Une dent de suidé soumise au laboratoire de Durham, issue de la même couche 19 dans le carré 6, a donné le résultat suivant: 7691 ± 98 cal. BC (68.2%). Les fragments de charbons de bois prélevés dans la tranchée B, 4, 4 et une dent ont fourni les dates 5801 - 5641 (95.4%) / 5742 - 5670 (68.3%) et 6151 ± 77 cal. BC (68.2%) (les dates sur les dents nous ont été com-muniquées par L.G. Flink, doctorant à l’université de Durham). Une autre dent confiée au laboratoire de Beta Analytic à Miami (sample IAUF004, analyse : AMS-Standard delivery, traitement à l’extraction du collagène avec alkali) a livré la mesure de 8250 ± 70 BP, Ratio 13C/12C : -21.7o/oo, soit un âge conventionnel de 8300±70 BP soit 7530-7100 cal. BC (communication de D. Hood, R. Hatfield, Chr. Patrick).
Enfin, des dents de chèvres provenant des couches supé-rieures à cet ensemble stratigraphique, attribuées au Mésolithique final, ont été adressées au laboratoire de Beta Analytic à Miami. Ces échantillons ont donné les datations suivantes (Trantalidou, 2003 et inédit) : 7060 ± 80 BP (soit 6065 – 5755 cal. BC), 7120 ± 80 BP (soit 6211- 5838 cal. BC), 7360 ± 50BP (soit 6368-6088 cal. BC, Ratio 13C/12C : -20.3o/oo) et 7440 ± 50 BP (soit 6410-6220 cal. BC).
- Les couches mésolithiques ont avant tout été identifiées par d’importantes concentrations de mollusques, de vertèbres de
poissons et notamment par l’absence de céramique. Les Helix aspersa, gastéropode terrestre le plus abondant dont 17201 coquilles ont été récoltées, totalisent 3325, 2772, 3497 et 726 fragments pour le Mésolithique ancien, le Mésolithique récent, le Néolithique ancien/moyen et le Néolithique récent, pour ne citer que les étapes majeures et les couches non remaniées (Karali, 2011).
- L’industrie lithique se limite à 48 outils en silex pour les niveaux inférieurs (8600 - 7800 cal. BC) ainsi qu’à 116 objets en silex et 12 en obsidienne pour les niveaux supérieurs (7500 - 7000 cal. BC) Selon les chercheurs, les microlithes en obsidienne semblent être façonnés avec des techniques diffé-rentes de celles employées sur le continent grec au cours du Mésolithique. Ces microlithes s’apparenteraient à ceux décou-verts dans les couches néolithiques sus-jacentes (Kaczanowska et Kozłowski, 2006 et 2008).
Une interrogation demeure concernant un éventuel tasse-ment d’une partie des sédiments néolithiques vers les couches inférieures. Des datations par l’hydratation de l’obsidienne ont été effectuées sur trois pièces provenant de la grotte du Cyclope, sur trois pièces provenant de la grotte de Sarakenos en Béotie, sur deux pièces de Keramé, sur une pièce de la station mésolithique de l’île d’Icarie, à l’est de la mer Égée, et sur deux pièces de la grotte de Schistos. D’après les datations obtenues, on peut avancer l’hypothèse de l’exploitation des sources d’obsidienne de Mélos, dans les Cyclades, dès le XIIIe millé-naire BP (Laskaris et al., 2011). Cependant, certaines datations obtenues pour la grotte du Cyclope (10968 ± 640 BP ; 11236 ± 776 BP ; 12017 ± 1875 BP) semblent être en contradiction avec la séquence stratigraphique proposée (Sampson, 2008), car ces pièces proviennent des strates du Néolithique récent. Selon notre avis, de nombreux paramètres peuvent être à l’origine de ces résultats, rendant incertaines ou inutilisables certaines séries d’échantillons.
- Les objets en matières dures animales, pour la plupart des armes de chasse et de pêche, sont au nombre de 79 dont 25 sont datés du Mésolithique ancien. Les pêcheurs et les chas-seurs qui occupaient de manière non régulière la grotte durant la seconde moitié du IXe millénaire façonnaient des outils en os avec une technique beaucoup plus performante que celle utilisée par les groupes contemporains qui fréquentaient la grotte de Franchthi. Le fait que le Néolithique initial ne soit pas présent dans la grotte rend difficile l’approche des aspects concernant l’évolution technologique. Comparée à l’évolution observée sur le continent pour les périodes suivantes, celle de la grotte du Cyclope ne semble pas très marquée (Moundrea-Agrafioti, 2011).
- L’horizon daté entre 8610 et 8300 cal. BC est intéressant d’un point de vue stratigraphique. Il a livré deux outils en silex, un outil en os, au moins 368 ossements de poissons déterminés ainsi que 1454 fragments de mollusques récoltés. Parmi ces derniers, le petit-gris (Helix aspersa) et la patelle (Patella asper-sa) sont les espèces les plus fréquentes, avec respectivement 67.81% et 27.51% pour l’ensemble de cet horizon (Karali, 2011).

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
151151
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
Les seuls ossements de mammifères recueillis appartiennent aux suidés. La rareté des vestiges osseux pourrait refléter les modes de subsistance de l’époque. Toutefois, il est possible que cette vision soit tronquée, car la sédimentation de ce secteur est moins importante que celle du reste des secteurs fouillés.
- Le seul assemblage conséquent, composé de suidés, est celui du Mésolithique ancien, comportant au moins 15 à 17 individus en traduisant les fréquences de restes en nombre minimal d’individus (fig. 9). Ces ossements, plus particulière-ment ceux de la phase la plus ancienne, sont plus dégradés que le reste du matériel osseux du site et montrent des taches de concrétions brunâtres. En revanche, ils sont moins fragmentés que ceux de tous les autres assemblages fauniques décrits et ils correspondent à quatre individus seulement. Par la suite, le
nombre de restes des caprinés est fortement majoritaire. Le pro-fil taphonomique (type de cassures, absence des concrétions) des ossements des chèvres et de moutons provenant de la partie centrale et sommitale de la séquence stratigraphique rappelle celui des sites du Néolithique (avec ou sans céramique).
- Les espèces d’oiseaux (fig. 10) et les mammifères chassés apparaissent en faible nombre. L’homme fut le seul prédateur sur la petite île. Dans la partie inférieure de la séquence, un faible nombre de restes appartenant au cerf, au lièvre et aux mammifères marins a été répertorié (Trantalidou, 2003 et 2011). L’abondance des ossements de poissons et des coquillages marins pourrait ainsi suggérer une exploitation intense des ressources marines durant le Mésolithique récent-final et le Néolithique ancien (Powell, 2011 ; Mylona, 2011).
Fig. 9 : Grotte du Cyclope à Youra. Détermination du sexe fondée sur les différences morphologiques de la forme des canines des suidés (d’après Trantalidou, 2011).
Cyclops Cave on Youra. Determination of the sex based on the morphological differences of the form of the canine teeth of Suids (after Trantalidou, 2011).
25
11
3
1 0 0
1 0
7 7
14
8
0 0
2
0 1
0 1 1
0
5
10
15
20
25
30
F M F M F M F M F M
L .MES UP. MES UP. MES / NEO
LN NEO
Maxilla
Mandible
152152
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
Dans le site de Maroulas sur l’île de Kythnos, l’épaisseur des strates ne dépasse 0,50 m et les ossements sont peu abondants. La faune mammalienne se compose de suidés, de lièvres et de carnivores (fig. 11).
LA FRÉQUENCE DES PARTIES DU SQUELETTE
Les différents segments du squelette des suidés sont plus ou moins présents, à l’exception des os du pied et surtout des phalanges (fig. 12). Dans la grotte du Cyclope, sur l’île de Youra, dans la mesure où les variations ne dépendent pas des modalités du remplissage, du secteur de fouille (20% de la surface totale de la grotte ont été dégagés) ou de la dégradation progressive des os, les effectifs de chaque catégorie anatomique montrent que la tête, le squelette axial et les extrémités constituent la majeure partie de l’assemblage. Il est possible que les parties anatomiques correspondant à l’épaule, au jambon et au jarret aient été dépo-sées ailleurs.
L’ÂGE D’ABATTAGE
En général, les animaux ont été abattus jeunes. On observe que, notamment à Maroulas mais aussi à Youra, 70% des osse-ments proviennent d’individus qui n’avaient pas encore atteint l’âge mature. Toutefois, à Maroulas, certains suidés atteignent
l’âge de 4 ans voire plus. Les classes d’âge représentées dans la figure 13 sont assez proches des structures démographiques connues chez les individus sauvages bien que ces derniers puissent survivre jusqu’à l’âge de 15 ans (Hone et Pedersen, 1980 ; ici : fig.13). Dans le gisement de la grotte du Cyclope, le phéno-mène est encore plus accentué, les âges des animaux chassés ne dépassant pas 2,5 ans. Dans certains sites, les chasseurs–cueil-leurs épipaléolithiques (par exemple dans les montagnes du Taurus occidental) qui se déplaçaient d’une station à l’autre au fil des saisons, pratiquaient une chasse ciblant majoritairement les jeunes bouquetins et chamois (Atici, 2009).
LA MORPHOMÉTRIE
Dans les deux sites, les restes des suidés, indépendamment du sexe, témoignent d’une diminution de la taille par rapport aux sangliers des sites du sud de la péninsule Balkanique, notamment en Thessalie (Pefkakia, Platia Magoula Zarkou) ou en Béotie (grotte de Sarakenos) entre le VIIe et le IIe millénaire. L’épaisseur des os et le cortex sont plus fins que ceux des sangliers adultes du continent (observations macroscopiques). La comparaison avec des individus provenant de sites modernes ou préhistoriques dans la péninsule Balkanique et en Asie Mineure (d’après les données morphométriques de Boessneck, 1962, p. 95-96 ; Jordan, 1975, tabl. 28, 34 ; Driesch et Enderle, 1976, tabl. 13 ; Hinz, 1979, tabl. 8, 12 ; Bökönyi, 1989, tabl. E7 ; Becker, 1991a, tabl. VII ; Payne et Bull 1988,
Fig. 10 : Distribution des trois principales espèces d’oiseaux chassées tout au long de la séquence mésolithique et néolithique de la grotte du Cyclope (en excluant les couches mélangées). Données modifiées d’après Trantalidou, 2011.
Distribution of the three main avian species hunted throughout the Mesolithic and Neolithic sequence of the Cyclops cave (mixed strata are excluded). Data after Trantalidou 2011, modified.
LE STATUT AMBIGU DES SUIDÉS

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
153153
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
493
6 8 7
24
5
2
1 1
7
1
10
100
1000
Sus scr
ofa
Lepus
europ
aeus
Vulpes
vulpe
s
Martes
foina
Aves
NISP MNI
176
68 62 69
6
1313
79
59
16
185
4
1
7 8 10
1
10
100
1000
10000
Skull
Axial sk
eleton
Fore lim
b
Hind Lim
b
Extrem
ities
MAROULAS, LM
YOURA, LM
YOURA, UM
Fig. 11 : Maroulas à Kythnos. Répartition de la faune mammalienne et aviaire (d’après Trantalidou, 2008 et 2010).
Maroulas on Kythnos. Distribution of the mammalian and avian fauna (after Trantalidou, 2008 and 2010).
Fig. 12 : Squelette appendiculaire et axial des suidés. Fréquence des spécimens trouvés à Maroulas et dans la grotte du Cyclope (d’après Trantalidou, 2010 et 2011).
Appendicular and axial skeleton of suids. Frequency of specimens found at Maroulas and Cave of Cyclops (after Trantalidou, 2010 and 2011).
154154
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
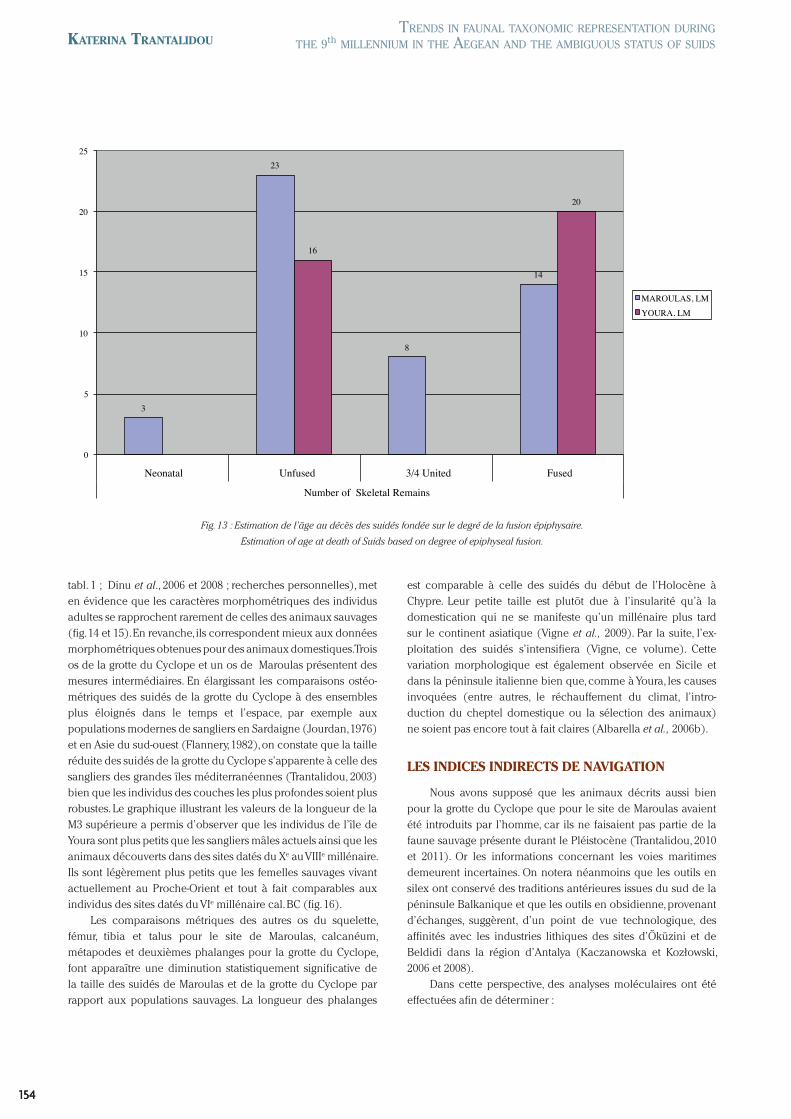
tabl. 1 ; Dinu et al., 2006 et 2008 ; recherches personnelles), met en évidence que les caractères morphométriques des individus adultes se rapprochent rarement de celles des animaux sauvages (fig. 14 et 15). En revanche, ils correspondent mieux aux données morphométriques obtenues pour des animaux domestiques. Trois os de la grotte du Cyclope et un os de Maroulas présentent des mesures intermédiaires. En élargissant les comparaisons ostéo-métriques des suidés de la grotte du Cyclope à des ensembles plus éloignés dans le temps et l’espace, par exemple aux populations modernes de sangliers en Sardaigne (Jourdan, 1976) et en Asie du sud-ouest (Flannery, 1982), on constate que la taille réduite des suidés de la grotte du Cyclope s’apparente à celle des sangliers des grandes îles méditerranéennes (Trantalidou, 2003) bien que les individus des couches les plus profondes soient plus robustes. Le graphique illustrant les valeurs de la longueur de la M3 supérieure a permis d’observer que les individus de l’île de Youra sont plus petits que les sangliers mâles actuels ainsi que les animaux découverts dans des sites datés du Xe au VIIIe millénaire. Ils sont légèrement plus petits que les femelles sauvages vivant actuellement au Proche-Orient et tout à fait comparables aux individus des sites datés du VIe millénaire cal. BC (fig. 16).
Les comparaisons métriques des autres os du squelette, fémur, tibia et talus pour le site de Maroulas, calcanéum, métapodes et deuxièmes phalanges pour la grotte du Cyclope, font apparaître une diminution statistiquement significative de la taille des suidés de Maroulas et de la grotte du Cyclope par rapport aux populations sauvages. La longueur des phalanges
est comparable à celle des suidés du début de l’Holocène à Chypre. Leur petite taille est plutôt due à l’insularité qu’à la domestication qui ne se manifeste qu’un millénaire plus tard sur le continent asiatique (Vigne et al., 2009). Par la suite, l’ex-ploitation des suidés s’intensifiera (Vigne, ce volume). Cette variation morphologique est également observée en Sicile et dans la péninsule italienne bien que, comme à Youra, les causes invoquées (entre autres, le réchauffement du climat, l’intro-duction du cheptel domestique ou la sélection des animaux) ne soient pas encore tout à fait claires (Albarella et al., 2006b).
LES INDICES INDIRECTS DE NAVIGATION
Nous avons supposé que les animaux décrits aussi bien pour la grotte du Cyclope que pour le site de Maroulas avaient été introduits par l’homme, car ils ne faisaient pas partie de la faune sauvage présente durant le Pléistocène (Trantalidou, 2010 et 2011). Or les informations concernant les voies maritimes demeurent incertaines. On notera néanmoins que les outils en silex ont conservé des traditions antérieures issues du sud de la péninsule Balkanique et que les outils en obsidienne, provenant d’échanges, suggèrent, d’un point de vue technologique, des affinités avec les industries lithiques des sites d’Öküzini et de Beldidi dans la région d’Antalya (Kaczanowska et Kozłowski, 2006 et 2008).
Dans cette perspective, des analyses moléculaires ont été effectuées afin de déterminer :
3
23
8
14
16
20
0
5
10
15
20
25
Neonatal Unfused
3/4 United Fused
Number of Skeletal Remains
MAROULAS, LM YOURA, LM
Fig. 13 : Estimation de l’âge au décès des suidés fondée sur le degré de la fusion épiphysaire.
Estimation of age at death of Suids based on degree of epiphyseal fusion.

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
155155
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
Fig. 14 : Longueur maximale de la M3 inférieure d’individus de Youra (8500-7000) - en jaune - comparée à celle de spécimens sauvages modernes d’Anatolie (Kizilcahamam, Anatolie centrale, individu femelle - en violet foncé)
et de la région danubienne (Dubova, individu male - en rose ), du site Mésolithique récent (7200-6300) Icoana, Portes de Fer ainsi que de porcs du Néolithique récent (5e millénaire) et de l’âge du Bronze moyen/récent
(ca 2600-2000/ 2000-1550) de Thessalie - en bleu.
Maximum length of the lower M3 from Youra (8500-7000) – in yellow - compared with those of modern wild specimen from Anatolia (Kizilcahamam, Central Anatolia, female- in dark purple) and the Danubian area
(Dubova, individu male – in pink ), from the Late Mesolithic site of Icoana as well as of Late Neolithic and Bronze Age pigs of Thessaly – in blue.
Fig. 15 : Comparaison de la largeur (Bd) des extrémités distales d’humérus de suidés de l’île de Youra (8500-8300) – en jaune – et de sites du Néolithique et de l’âge du Bronze en Thessalie, Grèce continentale (env. 6800-1550 cal. BC ; 31 valeurs).
En violet, des individus sauvages des sites de Pefkakia et Platia Magoula Zarkou (Néolithique récent, âge du Bronze) et en bleu des individus domestiques d’Achilleion, Argissa, Ayia Sofia et Pefkakia.
Width comparison (Bd) between distal humeri of suids from Youra island (8500-8300) – in yellow - and from Neolithic and Bronze Age sites in Thessaly, continental Greece (approximately 6800-1550 cal. BC; 31 values).In purple, wild specimen from Pefkakia and Platia Magoula Zarkou (Late Neolithic, Bronze age) and in blue, domestic specimen from Achilleion, Argissa,
Ayia Sofia and Pefkakia.
156156
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
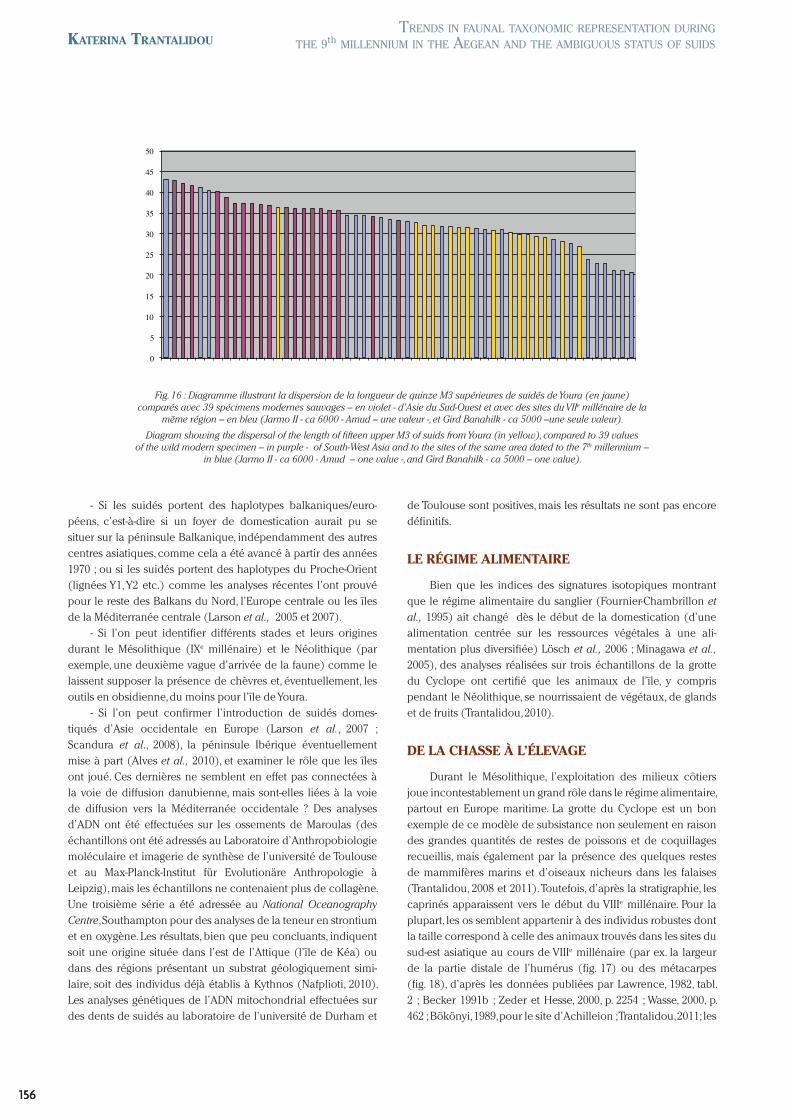
- Si les suidés portent des haplotypes balkaniques/euro-péens, c’est-à-dire si un foyer de domestication aurait pu se situer sur la péninsule Balkanique, indépendamment des autres centres asiatiques, comme cela a été avancé à partir des années 1970 ; ou si les suidés portent des haplotypes du Proche-Orient (lignées Y1, Y2 etc.) comme les analyses récentes l’ont prouvé pour le reste des Balkans du Nord, l’Europe centrale ou les îles de la Méditerranée centrale (Larson et al., 2005 et 2007).
- Si l’on peut identifier différents stades et leurs origines durant le Mésolithique (IXe millénaire) et le Néolithique (par exemple, une deuxième vague d’arrivée de la faune) comme le laissent supposer la présence de chèvres et, éventuellement, les outils en obsidienne, du moins pour l’île de Youra.
- Si l’on peut confirmer l’introduction de suidés domes-tiqués d’Asie occidentale en Europe (Larson et al., 2007 ; Scandura et al., 2008), la péninsule Ibérique éventuellement mise à part (Alves et al., 2010), et examiner le rôle que les îles ont joué. Ces dernières ne semblent en effet pas connectées à la voie de diffusion danubienne, mais sont-elles liées à la voie de diffusion vers la Méditerranée occidentale ? Des analyses d’ADN ont été effectuées sur les ossements de Maroulas (des échantillons ont été adressés au Laboratoire d’Anthropobiologie moléculaire et imagerie de synthèse de l’université de Toulouse et au Max-Planck-Institut für Evolutionäre Anthropologie à Leipzig), mais les échantillons ne contenaient plus de collagène. Une troisième série a été adressée au National Oceanography Centre, Southampton pour des analyses de la teneur en strontium et en oxygène. Les résultats, bien que peu concluants, indiquent soit une origine située dans l’est de l’Attique (l’île de Kéa) ou dans des régions présentant un substrat géologiquement simi-laire, soit des individus déjà établis à Kythnos (Nafplioti, 2010). Les analyses génétiques de l’ADN mitochondrial effectuées sur des dents de suidés au laboratoire de l’université de Durham et
Fig. 16 : Diagramme illustrant la dispersion de la longueur de quinze M3 supérieures de suidés de Youra (en jaune) comparés avec 39 spécimens modernes sauvages – en violet - d’Asie du Sud-Ouest et avec des sites du VIIe millénaire de la
même région – en bleu (Jarmo II - ca 6000 - Amud – une valeur -, et Gird Banahilk - ca 5000 –une seule valeur).
Diagram showing the dispersal of the length of fifteen upper M3 of suids from Youra (in yellow), compared to 39 values of the wild modern specimen – in purple - of South-West Asia and to the sites of the same area dated to the 7th millennium –
in blue (Jarmo II - ca 6000 - Amud – one value -, and Gird Banahilk - ca 5000 – one value).
de Toulouse sont positives, mais les résultats ne sont pas encore définitifs.
LE RÉGIME ALIMENTAIRE
Bien que les indices des signatures isotopiques montrant que le régime alimentaire du sanglier (Fournier-Chambrillon et al., 1995) ait changé dès le début de la domestication (d’une alimentation centrée sur les ressources végétales à une ali-mentation plus diversifiée) Lösch et al., 2006 ; Minagawa et al., 2005), des analyses réalisées sur trois échantillons de la grotte du Cyclope ont certifié que les animaux de l’île, y compris pendant le Néolithique, se nourrissaient de végétaux, de glands et de fruits (Trantalidou, 2010).
DE LA CHASSE À L’ÉLEVAGE
Durant le Mésolithique, l’exploitation des milieux côtiers joue incontestablement un grand rôle dans le régime alimentaire, partout en Europe maritime. La grotte du Cyclope est un bon exemple de ce modèle de subsistance non seulement en raison des grandes quantités de restes de poissons et de coquillages recueillis, mais également par la présence des quelques restes de mammifères marins et d’oiseaux nicheurs dans les falaises (Trantalidou, 2008 et 2011). Toutefois, d’après la stratigraphie, les caprinés apparaissent vers le début du VIIIe millénaire. Pour la plupart, les os semblent appartenir à des individus robustes dont la taille correspond à celle des animaux trouvés dans les sites du sud-est asiatique au cours de VIIIe millénaire (par ex. la largeur de la partie distale de l’humérus (fig. 17) ou des métacarpes (fig. 18), d’après les données publiées par Lawrence, 1982, tabl. 2 ; Becker 1991b ; Zeder et Hesse, 2000, p. 2254 ; Wasse, 2000, p. 462 ; Bökönyi, 1989, pour le site d’Achilleion ; Trantalidou, 2011; les

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
157157
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
mêmes résultats ont été obtenus en utilisant deux variables). La présence des quelques ovicaprinés de plus grande taille dans les couches du Mésolithique final tout comme celle de suidés (fig. 19, fig. 20) mérite d’être remarquées. En effet, les données métriques des ossements de chèvres de Youra se rapprochant de celles des chèvres actuelles sur les plateaux de la chaîne du Zagros
(Lawrence, 1982 ; Zeder, 2001) méritent d’être signalées, car la taille est aussi un indicateur de l’espace que les animaux ont à leur disposition. Une analyse moléculaire effectuée dans le laboratoire de l’université de Mayence confirme cette origine des chèvres et permettra éventuellement de préciser les routes mari-times, les échanges ou d’éventuels déplacements de personnes.
0
5
10
15
20
25
30
35
40
45
50
Iran & Iraq, W, mod.
Iran & Iraq, W, mod.
Ganj Dareh, 8200-7500
BC
Çayönü, Ain Ghazal, Jordan valley,
Ain Ghazal
Ain Ghazal
Youra, Ceast, 17,3
Achilleion, 6000BC
Iran & Iraq, W, mod.
Southern Zagros, W, M, mod.
Southern Zagros, W, M, mod.
Southern Zagros, W, F, mod
Iran & Iraq, W, mod.
Southern Zagros, W, F, mod.
(Körtik Tepe, Tigris river, 10000-9500 BC
Ganj Dareh, Zagros region
Ganj Dareh, 8200-7500 BC
Çayönü, Taurus mountains
Çayönü, 8700-8200 BC
Çayönü, 8200-7500 BC
Çayönü,
Ain Ghazal , 7500-7000BC Fig. 17 : Comparaison de mesures des extrémités distales d’humérus de Capra sp. En violet, chèvres sauvages
modernes (Iran, Iraq, Zagros sud) ; en bleu, Capra aegagrus de sites PPN des montagnes Zagros-Taurus (Körtik Tepe, Tigris river, 10000-9500 / Ganj Dareh, Zagros region, 8200-7500 / Çayönü, Taurus mountains,
PPNA- EPPNB, 8700-8200), de la vallée du Jourdain (Ain Ghazal, PPNB, 7500-7000) et du site néolithique ancien d’Achilleion dans la vallée du Pénée (6300-5600) ; en jaune, ossements de Youra.
Comparison of measurements of distal humerus of Capra sp. In purple, modern wild goats (Iran, Iraq, South Zagros); in blue, Capra aegagrus bones from PPN sites in the Zagros-Taurus mountain area Taurus
(Körtik Tepe, Tigris river, 10000-9500 / Ganj Dareh, Zagros region, 8200-7500 / Çayönü, Taurus mountains, PPNA- EPPNB, 8700-8200), the Jordan valley (Ain Ghazal, PPNB, 7500-7000) and from Early Neolithic Achilleion
in the Peneos valley; in yellow, specimen from Youra.
Fig.18 : Largeurs distales des métacarpes de Capra sp comparées avec celles des métacarpes de Capra aegagrus provenant de sites PPN d’Asie du Sud-Ouest ainsi que de chèvres sauvages modernes.
Distal width of Capra sp metacarpal bones compared to PPN sites from SW Asia and modern Capra aegagrus metacarpal bones.
158158
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
Fig. 19 : Grotte du Cyclope, Youra, Sporades du Nord. État de conservation des ossements de Capra sp. Les humérus de caprins ont été trouvés dans les couches archéologiques suivantes : tranchée C ouest, couche 7 et tranchée C est, couche 16, m2 3.
Photographie : A. Iliakopoulos.
Cave of Cyclops, Youra, Northern Sporades. State of preservation of the Capra component. Caprine humeri were found in the following cultural deposits: Trench C west, layer 7 and Trench C east, Layer 16, sq. 3 (the third specimen).
Photograph: A. Iliakopoulos.
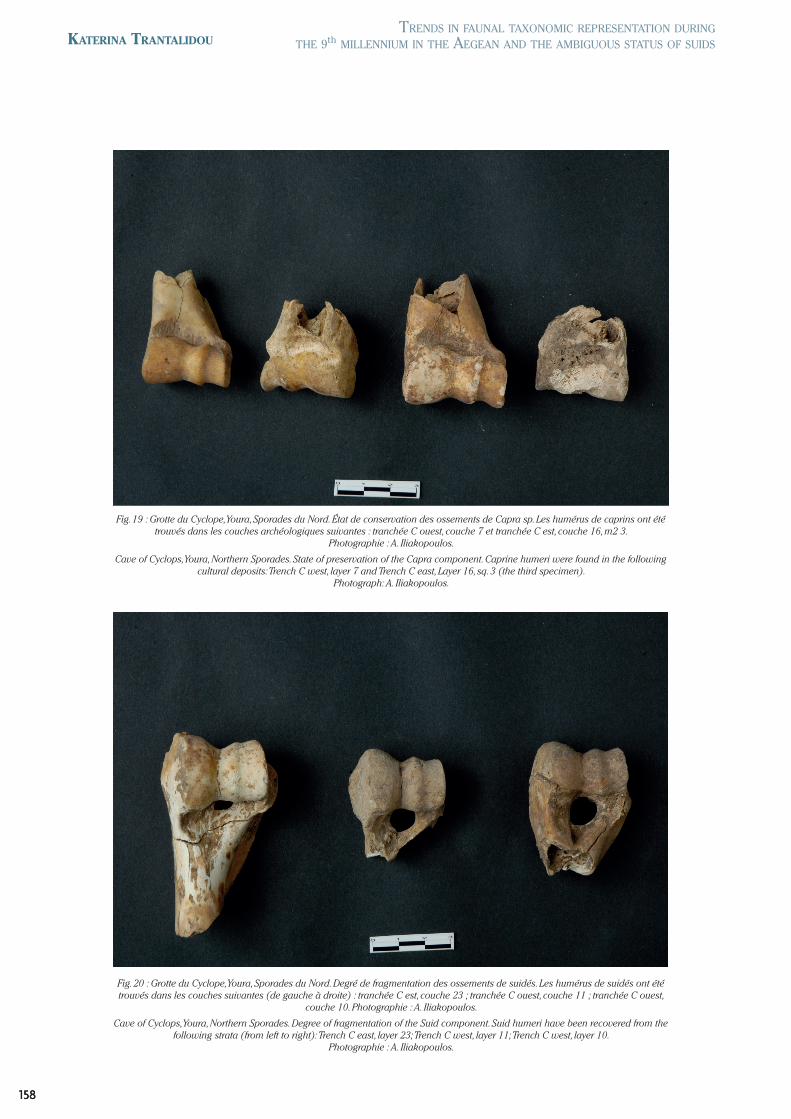
Fig. 20 : Grotte du Cyclope, Youra, Sporades du Nord. Degré de fragmentation des ossements de suidés. Les humérus de suidés ont été trouvés dans les couches suivantes (de gauche à droite) : tranchée C est, couche 23 ; tranchée C ouest, couche 11 ; tranchée C ouest,
couche 10. Photographie : A. Iliakopoulos.
Cave of Cyclops, Youra, Northern Sporades. Degree of fragmentation of the Suid component. Suid humeri have been recovered from the following strata (from left to right): Trench C east, layer 23; Trench C west, layer 11; Trench C west, layer 10.
Photographie : A. Iliakopoulos.

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
159159
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
Dans l’extrémité sud de la péninsule Balkanique, les inves-tigations entreprises dans les sites qui abritaient des groupes contemporains des phases de l’Alleröd et du Préboréal ont per-mis de mettre en évidence des répercussions considérables sur la faune et, par conséquent, sur l’économie et le comportement humain.
Certaines espèces (par exemple l’hydrontin (Equus hydrun-tinus) disparaissent, d’autres se maintiennent sur le continent, mais se font rarissimes. D’autres encore diminuent en taille, phénomène attesté dans les îles, notamment à Chypre dès le Xe millénaire (Vigne et al., 2009) et, dès le IXe millénaire cal. BC sur les îles de Kythnos et de Youra. La présence de populations de suidés de petite taille, isolées sur les îles, est souvent remarquée dans la littérature (Albarella et al., 2006a, b et 2009). À ce pro-pos, le matériel osseux de Maroulas et de la grotte du Cyclope contient des informations précieuses. L’espace limité par les montagnes et les côtes permettait ainsi un certain contrôle sur les animaux exercé par les groupes humains fréquentant les deux sites. Cependant, le matériel de Maroulas est trop fragmenté pour observer un processus de changement morphologique. En ce qui concerne le matériel osseux de la grotte du Cyclope, un certain rétrécissement de la dentition, souvent lié aux chan-gements morphologiques du crâne, n’est visible que dans les couches du Néolithique ancien. Les recherches effectuées dans le site de Çayönü ont révélé un processus de domestication lent, s’étalant sur deux millénaires (Ervynck et al., 2001).
Le ramassage, la cueillette, la chasse et la pêche rythmaient la vie des groupes mésolithiques. Dans certains sites, des osse-ments d’ovicaprinés ont été identifiés dans les dépôts méso-lithiques. L’observation stratigraphique montre que ces éléments fauniques ne sont pas contemporains des dépôts lenticulaires
qui contenaient les objets mésolithiques. Il est fort probable que la formation de ces couches résulte de processus post-dé-positionnels, fréquents dans les grottes. Au lieu de considérer la stratigraphie du site comme intacte, il serait préférable de s’inté-resser aux épisodes successifs et d’évaluer si ces remaniements sont dus à des phénomènes naturels ou à des actions humaines (Mlekuž et al., 2008).
À ce jour, aucune espèce domestique n’est attestée avec certitude dans les sites mésolithiques du sud de la péninsule Balkanique. Toutefois, pour résoudre ce problème, il sera indispensable de disposer d’échantillons fiables, comprenant des analyses de l’ADN ancien et de datations absolues effectuées à partir des ossements pour les associer aux analyses morphométriques et au profil démographique (répartition par classes d’âge, établies à partir de l’usure dentaire et de l’observation des épiphyses) des populations d’animaux (Dobney et Larson, 2006).
Les différences concernant les modes économiques et les stratégies de subsistance observées entre les îles et le continent s’expliquent tout d’abord par leur situation géographique et leur environnement. Elles sont aussi dues à la mobilité des groupes humains qui ont colonisé ces îles en introduisant des animaux et en participant à des réseaux d’échange de produits et d’idées. Si en l’état actuel des recherches, la maîtrise de la navigation au tout début de l’Holocène ne fait aucun doute, l’histoire de la domes-tication des animaux peut contribuer à retracer le parcours des groupes humains dans différentes directions : du sud de la péninsule Balkanique vers Kythnos et Youra, du continent asia-tique vers Youra et ensuite vers d’autres îles méditerranéennes, en direction de l’ouest. Cette hypothèse reste à confirmer ou à infirmer.
CONCLUSION
RemerciementsJ’exprime toute ma reconnaissance aux directeurs des fouilles suivantes : Maroulas et grotte du Cyclope ; grotte de Schistos
(section préhistorique) ; grotte de Theopetra et grotte de Kleissoura : Pr. Ad. Sampson, Dr. F. Mavridis et L. Kormazopoulou, Dr. N. Kyparissi-Apostolika, directeur honoraire des Antiquités, et Dr. M. Koumouzelis pour m’avoir confié leur matériel. Je remercie vivement les organisateurs du congrès pour leur accueil chaleureux et pour avoir bien voulu publier le présent texte.

160160
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
ADAM E. (2000) - Οι ανώτερες παλαιολιθικές λιθοτεχνίες του σπηλαίου Θεόπετρας και η συμβολή τους στην εκτίμηση της χρήσης του σπηλαίου κατά το τελικό Πλειστόκαινο και πρώιμο Ολόκαινο, in N. Kyparissi–Apostolika (dir.), Theopetra Cave. Twelve years of excavation and research 1987 – 1998, Proceedings of the International Conference, Trikala, 1998, Athens, Ministry of Culture, p. 163-171.
ALBARELLA U., MANCONI F., ROWLEY-CONWY P., VIGNE J.-D. (2006a) - Pigs of Corsica and Sardinia: a biometrical re-evaluation of their status and history, Studi di archeozoologia in onore di Alfredo Riedel, Bolzano, Dipdruck, p. 285-302.
ALBARELLA U., TAGLIACOZZO A., DOBNEY K., ROWLEY-CONWY P. (2006b) - Pig hunting and husbandry in prehistoric Italy: a contribution to the domestication debate, Proceedings of the Prehistoric Society, 72 p. 193-227.
ALBARELLA U., DOBNEY K., ROWLEY-CONWY P. (2009) - Size and shape of the Eurasian wild boar (Sus scrofa), with a view to the reconstruction of its Holocene history, Environmental Archaeology, 14 (2), 103-136.
ALEXANDROWICZ S. W. (2010). Malacofauna of the Mesolithic site in the Kythnos island, in A. Sampson, M. Kaczanowska, J. Kozłowski (dir.), Τhe prehistory of the island of Kythnos and the Mesolithic settlement at Maroulas, Krakow, Polish Academy of Sciences and Arts and University of the Aegean, p. 143-145.
ALVES P.C., PINHEIRO IR., GODINHO R., VICENTE J., GORTÁZAR CHR., SCANDURA M. (2010) - Genetic diversity of wild boar populations and domestic pig breeds (Sus scrofa) in South-western Europe, Journal of the Linnean Society, 101, p. 797–822.
ATICI L. (2009) - Implications of Age Structures for Epipaleolithic Hunting Strategies in the Western Taurus Mountains, Southwest Turkey, Anthropozoologica, 44(1), p. 13-39.
BECKER C. (1991a) - Die Tierknochenfunde von der Platia Magoula Zarkou : neue Untersuchungen zu Haustierhaltung, Jagd und Rohstoffverwendung im neolithisch-bronzezeitlichen Thessalien, PZ 66, p. 14-78.
BECKER C. (1991b) - The Analysis of mammalian bones from Basta, a pre-pottery Neolithic site in Jordan, Paléorient 17 (1), p. 59-75.
BOESSNECK J.A. (1962) - Die tierreste aus der Argissa-Magula vom präkeramischen Neolithicum bis zur mittleren Bronzezeit, in V. Milojčič, J. Boessneck, M. Hopf (dir.), Die Deutschen Ausgrabungen auf der Argissa-Magula in Thessalien I, Bonn, Habelt, p. 27-99.
BÖKÖNYI S. (1989) – Fauna, in M. Gimbutas, Sh. Winn, D Shimabuku (dir.), Achilleion. A Neolithic Settlement in Thessaly, Greece, 6400-5600 BC, Los Angeles, Monumenta Archaeologica, p. 315-339.
BOTTEMA S. (2003) - The Vegetation History of the Greek Mesolithic, in N. Galanidou, C. Perlès (dir.), The Greek Mesolithic. Problems and perspectives, London, British School of Athens, p. 32-49.
DINU A., MEIGGS D., BĂLĂSESCU A., BORONEANŢ A., SOFICARU A., MIRIŢOIU N. (2006) - On Men and Pigs: were Pigs Domesticated at Mesolithic Iron Gates of the Danube? Part One: Teeth Metrics, Studii de Preistorie, 3, p. 77-98.
DINU A., BORONEANŢ A., BĂLĂSESCU A., SOFICARU A., MIRIŢOIU N. (2008) - Mesolithic and Neolithic Pigs of the Northern Balkans: Astragali vs. Teeth as Markers of Domestication, Mesolithic Miscellany, 19 (1), p. 7-12.
DOBNEY K., LARSON G. (2006) - Genetics and animal domestication: new windows on an elusive process, J. Zool., 269, p. 261-271.
DRIESCH VON DEN A., ENDERLE K. (1976) - Die Tierreste aus der Agia Sofia Magula in Thessalien, in Vl. Milojçiç Vl., A. von den Driesch, K. Enderle, V. Milojçiç, J. Zumbusch Kl. (dir.), Magulen um Larisa in Thessalien 1966, Bonn, Rudolf Habelt Verlag, p. 15-54.
ERVYNCK A., DOBNEY K., HONGO H., MEADOW R. (2001) - Born free? New evidence for the status of Sus scrofa at Neolithic Çayönü Tepesi (Southeastern Anatolia, Turkey), Paléorient, 27, 2, p. 47–73.
FACORELLIS Y. (2003) - Radiocarbon dating the Greek Mesolithic, in N. Galanidou, C. Perlès (dir.), The Greek Mesolithic. Problems and perspectives, London, British School of Athens, p. 51-67.
FACORELLIS Y. (2011) - Sequential Radiocarbon Dating and Calculation of the Marine Reservoir Effect, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, Bone Tool Industries, Dietary Resources and the Paleoenvironment, and Archeometrical Studies, vol II, Philadelphia, INSTAP, Monograph Series, p. 361-372.
FACORELLIS Y., KYPARISSI-APOSTOLIKA N., MANIATIS Y. (2001) - The cave of Theopetra, Kalambaka: radiocarbon evidence for 50000 years of human presence, in I. Carmi, E. Boaretto (dir.), Proceedings of the 17th International 14C Conference, Radiocarbon 43(2B), p 1029-1048.
FACORELLIS Y., DAMIATA B.N., VARDALA-THEODOROU E., NTINOU M., SOUTHON J. (2010) - AMS Radiocarbon Dating of the Mesolithic Site Maroulas on Kythnos and calculation of the Regional Marine Reservoir, in A. Sampson, M. Kaczanowska, J. Kozłowski (dir.), Τhe prehistory of the island of Kythnos and the Mesolithic settlement at Maroulas, Krakow, Polish Academy of Sciences and Arts and University of the Aegean, p. 127-135.
FARRAND W.R. (2000) - Depositional History of Franchthi Cave: Sediments, stratigraphy and chronology. Fasc. 12, Bloomington, Indiana, Indiana University Press, 152 p.
FARRAND W.R. (2003) - Depositional environments and site formation during the Mesolithic occupations of Franchthi Cave, Peloponnesos, Greece, in N. Galanidou, C. Perlès (dir.), The Greek Mesolithic. Problems and perspectives, London, British School of Athens, p. 69-78.
FLANNERY K.V. (1982) - Early pig domestication in the fertile crescent a retrospective look, in P.E. L. Smith, Braidwood R.J., P. Mortensen (dir.), The Hilly Flanks and beyond: essays on the prehistory of southwestern Asia, Chicago, The Oriental Institute of the University of Chicago, 36, p. 163-187.
FOURNIER-CHAMBRILLON C., MAILLARD D., FOURNIER P. (1995) - Diet of the Wild boar (Sus scrofa L.) inhabiting the Montpellier Garrigue, Ibex (Journal of Mountain Ecology), 3, p. 174-179.
HANSEN J. (1991) - Excavations at Franchthi Cave, Greece. Fasc. 7. The palaeoethnobotany of Franchthi cave, Bloomington & Indianapolis, Indiana University Press, 280 p.
HANSEN J. (1992) - Franchthi cave and the beginnings of agriculture in Greece and the Aegean, in P. Anderson-Gerfaud (dir.), Préhistoire de l’agriculture : nouvelles approches expérimentales et ethnographiques, Paris, CNRS, p. 231-247.
HINZ G. (1979) - Neue Tierknochenfunde aus der Magula Pevkakia in Thessalien. I. Die Nichtwiederkäuer, Munich, Ph. D. thesis.
BIBLIOGRAPHIE

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
161161
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
HONE J., PEDERSEN H. (1980) - Changes in a Feral Pig Population after poisoning, in J.P. Clark (dir.), Proceedings of the 9th Vertebrate Pest Conference, University of Nebraska - Lincoln, p. 176-182.
GALANIDOU N., PERLÈS C. (2003) - An introduction to the Greek Mesolithic, in N. Galanidou, C. Perlès (dir.), The Greek Mesolithic. Problems and perspectives, London, British School of Athens, p. 27-32.
GAMBLE C. (1999) - Faunal exploitation at Klithi: a Late Glacial rockshelter in Epirus, Northwestern Greece, in G.N. Bailey, E. Adam, E. Panagopoulou, C. Perlès, K. Zachos (dir.), The Palaeolithic Archaeology of Greece and Adjacent Areas, Proceedings of the ICOPAG Conference, Ioannina, September 1994, British School at Athens, London (Studies 3), p. 179-187.
IOAKEIM Chr. (2011) - Palynological Evidence, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, Bone Tool Industries, Dietary Resources and the Paleoenvironment, and Archeometrical Studies, vol II, Philadelphia, INSTAP, Monograph Series, p. 289-295.
ISAAKIDOU V. (2004) - Bones from the labyrinth. Faunal Evidence for the Management and the Consumption of Animals at Neolithic and Bronze Age Knossos, Crete. Phd dissertation. UCL. Unpublished Thesis.
JARMAN M.R., JARMAN H.N. (1969) - The fauna and economy of early Neolithic Knossos, in P. Warren, M.R. Jarman, H.N. Jarman, N.J. Shackleton, J.D. Evans (dir.), Knossos Neolithic. Part II. Britisch School of Athens, 63, p. 241-264.
JORDAN B. (1975) - Tierknochenfunde aus der Magula Pevkakia in Thessalien, München, University of Munich, PhD.
JOURDAN L. (1976) - La faune du site gallo-romain et paléochrétien de la Bourse (Marseille), Paris, CNRS, 338 p.
KACZANOWSKA M., KOZŁOWSKI J.K. (2006) - Παλαιολιθικές παραδόσεις, μεσολιθικές προσαρμογές και νεολιθικές καινοτομίες μέσα από το πρίσμα της λιθοτεχνίας, in A. Sampson (dir.), Η προϊστορία του Αιγαίου: Παλαιολιθική, Μεσολιθική, Νεολιθική, Athens, Atrapos, p. 67-87.
KACZANOWSKA M., KOZŁOWSKI J.K. (2008) - Chipped stone artifacts, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, vol 1, Philadelphia, INSTAP, Monograph Series, p. 169-179.
KARALI L. (2011) - Malacological Material, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, vol 1, Philadelphia, INSTAP, Monograph Series, p. 267-287.
KARKANAS P., KYPARISSI-APOSTOLIKA N., BAR-YOSEF O., WEINER ST. (1999) - Mineral assemblages in Theopetra, Greece: a framework for understanding diagenesis in a prehistoric cave, Journal of Archaeological Science, 26 (9), p. 1171-1180.
KARKANAS P., WEINER ST. (2000) - Λιθοστρωματογραφία και διαγένεση των αποθέσεων του σπηλαίου Θεόπετρας Καλαμπάκας, in N. Kyparissi–Apostolika (dir.), Theopetra Cave. Twelve years of excavation and research 1987 – 1998, Proceedings of the International Conference, Trikala, 1998, Athens, Ministry of Culture, p. 37-51.
KOPAKA K., MATZANAS CHR. (2009) - Palaeolithic industries from the island of Gavdos, near neighbour to Crete in Greece, Antiquity 83 (321), http://antiquity.ac.uk/projgall/kopaka321/.
KOTJABOPOULOU E. (2001) - Patterned fragments and fragments of patterns: Upper Palaeolithic rockshelter faunas from Epirus northwestern Greece, Cambridge, University of Cambridge, Unpublished PhD Thesis, 304 p.
KOUMOUZELIS M., GINTER B., KOZŁOWSKI J.K., PAWLIKOWSKI M., BAR-YOSEF O., ALBERT R.M., LITYNSKA-ZAJAC M., STWORZEWICZ E., WOJTAL P., LIPECKI G., TOMEK T., BOCHENSKI Z.M., PAZDUR A. (2001) - The Early Upper Palaeolithic in Greece: The Excavations in Klisoura Cave, Journal of Archaeological Science, 28 (5) p. 515-539.
KOUMOUZELIS M., GINTER B., KOZLOWSKI J.K., KAZIOR B., SOBCZYK K. (2004) - Cave 1 in Klisoura Gorge (Greece), Recherches archéologiques de 1993-1998, Krakow, p. 206-220.
KYPARISSI-APOSTOLIKA N. (2000) - Η ανασκαφή του σπηλαίου Θεόπετρας 1987– 1998, in N. Kyparissi–Apostolika (dir.), Theopetra Cave. Twelve years of excavation and research 1987 – 1998, Proceedings of the International Conference, Trikala, 1998, Athens, Ministry of Culture, p. 17–36.
KYPARISSI-APOSTOLIKA N. (2003) - The Mesolithic in Theopetra Cave: new data on a debated period of Greek prehistory, in N. Galanidou, C. Perlès (dir.), The Greek Mesolithic. Problems and perspectives, London, British School of Athens, p. 189-198.
LARSON G., K. DOBNEY, U. ALBARELLA, M. FANG, E. MATISOO-SMITH, J. ROBINS, S. LOWDEN, H. FINLAYSON, T. BRAND, E. WILLERSLEV, P. ROWLEY-CONWY, L. ANDERSSON, A. COOPER (2005) - Worldwide phylogeography of wild boar reveals multiple centers of pig domestication, Science, 307, 1618-1621.
LARSON G., ALBARELLA U., DOBNEY K., ROWLEY-CONWY P., SCHIBLER J., TRESSET A., VIGNE J.-D., EDWARDS C.J., SCHLUMBAUM A., DINU A., BǍLǍÇSESCU A., DOLMAN G., TAGLIACOZZO A., MANASERYAN N., MIRACLE P., VAN WIJNGAARDEN-BAKKER L., MASSETI M., BRADLEY D.G., COOPER A. (2007) - Ancient DNA, pig domestication, and the spread of the Neolithic into Europe, Proceedings of the National Academy of Sciences of the United States of America, 104 (39), p. 15276-15281.
LASKARIS N., SAMPSON A., MAVRIDIS F., LIRITZIS I., (2011) Late Pleistocene/Early Holocene seafaring in the Aegean: new obsidian hydration dates with the SIMS-SS method, Journal of Archaeological Science, 38, p. 2475-2479.
LAWRENCE B. (1982) - Principal Food Animals at Çayönü, in L.S. Braidwood, R.J. Braidwood (dir.), Prehistoric Village Archaeology in South-Eastern Turkey, Oxford, Archaeopress (BAR International Series 138), p.175-199.
LEBLANC L. (1996) - Zooarchaeological report from Konispol Cave, Albania, Canadian Zooarchaeology, 10, p. 11-15.
LÖSCH S., GRUPE G., PETERS J. (2006) - Stable isotopes and dietary adaptations in humans and animals at pre-pottery Neolithic Nevalli Çori, southeast Anatolia, American Journal of Physical Anthropology, 131, p. 181-193.
LUBELL D. (2004) - Prehistoric edible land snails in the circum-Mediterranean: the archaeological evidence, in J.-P. Brugal, J. Desse (dir.), Petits animaux et sociétés humaines. Du complémentarisme alimentaire aux ressources utilitaires, XXIVe rencontres internationales d’archéologie et d’histoire d’Antibes, Antibes, Éditions APDCA, p. 77-98.
MANGAFA M. (2000) - Η εκμετάλλευση των φυτών από τη Μέση Παλαιολιθική έως τη Νεολιθική Περίοδο. Από την καρποσυλλογή στην καλλιέργεια, in N. Kyparissi–Apostolika (dir.), Theopetra Cave. Twelve years of excavation and research 1987 – 1998, Proceedings of the International Conference, Trikala, 1998, Athens, Ministry of Culture, p. 163-171.

162162
Trends in faunal Taxonomic represenTaTion during The 9th millennium in The aegean and The ambiguous sTaTus of suidsKaterina trantalidou
ΜAVRIDIS F., ΚORMAZOPOULOU L. (2009) - Aνώνυμο σπήλαιο Σχιστού Κερατσινίου. Η ανασκαφή των ετών 2006-2007. Τα πρώτα στοιχεία της έρευνας, Αρχαιολογικά Ανάλεκτα εξ Αθηνών, 40-41, p. 13-23.
ΜAVRIDIS F., LASKARIS Ν., PAPADEA Α., ΑPOSTOLIKAS Ο., ΤRANTALIDOU Κ., ΚOTZAMANI G., KARKANAS P., MANIATIS Υ., LIRITZIS Ι., ΚORMAZOPOULOU L. (2012) - Aνώνυμο σπήλαιο Σχιστού Κερατσινίου: Χρονολόγηση δειγμάτων Οψιανού από στρώματα του ανώτερου Πλειστοκαίνου και των αρχών του Ολόκαινου με τη μέθοδο SIMS-SS, in N. Zacharias (dir.), Archaeological Research and New Technologies, Proceedings of the 2nd Symposium, Kalamata 2010, University of Peloponnesus, p. 121-131.
MINAGAWA M., MATSUI A., ISHIGURO N. (2005) - Patterns of prehistoric boar Sus scrofa domestication, and inter-islands pig trading across the East China Sea, as determined by carbon and nitrogen isotope analysis, Chemical Geology, 218, p. 91-102.
MLEKUŽ D., BUDJA M., PAYTON R., BONSALL CL. (2008) - “Mind the Gap”. Caves, Radiocarbon Sequences, and the Mesolithic–Neolithic Transition in Europe. Lessons from the Mala Triglavca Rockshelter Site, Geoarchaeology, 23, 3, p. 398-416.
MOUNDREA-AGRAFIOTI A. (2011) - The Mesolithic and Neolithic Bone Implements, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, Bone Tool Industries, Dietary Resources and the Paleoenvironment, and Archeometrical Studies, vol II, Philadelphia, INSTAP, Monograph Series, p. 3-49.
MYLONA D. (2010) - Mesolithic Fishers at Maroulas, Kythnos: the Fish Bones, in A. Sampson, M. Kaczanowska, J. Kozłowski (dir.), Τhe prehistory of the island of Kythnos and the Mesolithic settlement at Maroulas, Krakow, Polish Academy of Sciences and Arts and University of the Aegean, p. 151-161.
MYLONA D. (2011) - Fish Vertebrae, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, Bone Tool Industries, Dietary Resources and the Paleoenvironment, and Archeometrical Studies, vol II, Philadelphia, INSTAP, Monograph Series, p. 237-266.
NAFPLIOTI A. (2010) - The Mesolithic Occupants of Maroulas on Kythnos: Skeletal Isotope Ratio Signatures of their Geographic, in A. Sampson, M. Kaczanowska, J. Kozłowski (dir.), Τhe prehistory of the island of Kythnos and the Mesolithic settlement at Maroulas, Krakow, Polish Academy of Sciences and Arts and University of the Aegean, p. 207-215.
NEWTON S. (2000) - Theopetra cave and the Palaeolithic – Mesolithic transition in southern Europe, in E. Kotjabopoulou, Y. Hamilakis, P. Halstead, C. Gamble, P. Elefanti (dir.), Zooarchaeology in Greece. Recent advances, Athens, British School at Athens Studies 9, p. 115-122.
NEWTON S. (2003) - The Mesolithic fauna from Theopetra Cave, in N. Galanidou, C. Perlès (dir.), The Greek Mesolithic. Problems and perspectives, London, British School of Athens, p. 199-205.
NTINOU M. (2011) - Charcoal Analysis, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, Bone Tool Industries, Dietary Resources and the Paleoenvironment, and Archeometrical Studies, vol II, Philadelphia, INSTAP, Monograph Series, p. 297-314.
PAYNE S. (1975) - Faunal change at Franchthi cave from 20,000 BC-3,000 BC, in A.T. Clason (dir.), Archaeozoological Studies, Amsterdam, North Holland Publishing Co., p. 120-131.
PAYNE S. (1982) - Faunal evidence for environmental/climatic change at Franchthi cave (Southern Argolid, Greece), 25,000 BP-5,000 BP. Preliminary results, in J.L. Bintliff, W. van Zeist (dir.), Palaeoclimates, Palaeoenvironments and Human Communities in the Eastern Mediterranean Region in Later Prehistory, Oxford, Archaeopress (BAR International Series, 133), p. 133-137.
PAYNE S., BULL G. (1988) - Components of Variation in Measurements of Pig Bones and Teeth, and the Use of Measurements to Distinguish Wild from Domestic Pig Remains, ArchaeoZoologia, 2, p. 27-65.
PERLÈS C. (1987) - Excavations at Franchthi Cave, Greece. Fasc. 3. Les industries lithiques taillées de Franchthi (Argolide, Grèce). Tome I. Présentation générale et industries paléolithiques, Indiana University Press, 355 p.
PERLÈS C. (1990) - Excavations at Franchthi Cave, Greece. Fasc. 5. Les industries lithiques taillées de Franchthi (Argolide, Grèce). Tome. II. Les industries du Mésolithique et du Néolithique initial, Indiana University Press, 288 p.
PERLÈS C. (2001) - The early Neolithic in Greece: the first farming communities in Europe, Cambridge, Cambridge University Press, 356 p.
PETRUSO K.M., KORKUTI M., BEJKO L., BOTTEMA S., ELLWOOD B. B, HANSEN J. M., HARROLD F.B., RUSSELL N. (1996) - Konispol Cave, Albania: A Preliminary Report on Excavations and Related Studies, 1992-1994, Iliria, 26, 1-2, p. 183-224.
POWELL J. (2011) - Non-Vertebral Fish Bones, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, Bone Tool Industries, Dietary Resources and the Paleoenvironment, and Archeometrical Studies, vol II, Philadelphia, INSTAP, Monograph Series, p. 151-235.
ROSE M. (1995) - Fishing at Franchthi Cave, Greece: changing environments and patterns of exploitation, Old World Archaeology Newsletter, 18, p. 21-26.
RUNNELS C.N., MURRAY P. (2001) - Greece Before History: an Archaeological Companion and Guide, Stanford, Stanford University Press, 224 p.
RUNNELS C. N., VAN ANDEL T. H. (2003) - The early Stone Age of the Nomos of Preveza: Landscape and Settlement Landscape archaeology in southern Epirus, Greece, in J. Wiseman, K.L. Zachos (dir.), Landscape archaeology in southern Epirus, Greece I, Hesperia suppl., 32, p. 47-134.
SAMPSON A. (2006) - H Mεσολιθική περίοδος στο αιγαίο, in Sampson (dir.), Η προϊστορία του Αιγαίου: Παλαιολιθική, Μεσολιθική, Νεολιθική, Athens, Atrapos, p. 25-66.
SAMPSON A. (2008) - Cave Setting and Stratigraphy, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, vol 1, Philadelphia, INSTAP, Monograph Series, p. 1-16.
SAMPSON A., KACZANOWSKA M., KOZŁOWSKI J. (2009) - The first Mesolithic site in the eastern part of the Aegean Basin: excavations into the site Kerame I on the island of Ikaria in 2008, Rocznik Polskiej Akademii Umiejtnosci, 1958, p. 321-329.
SAMPSON A., KACZANOWSKA M., KOZŁOWSKI J. (2010) - Τhe prehistory of the island of Kythnos and the Mesolithic settlement at Maroulas, Krakow, Polish Academy of Sciences and Arts and University of the Aegean, 215 p.

La diffusion par Chypre, L’ÉgÉe et L’adriatique
the diffusion by Cyprus, eagean and adriatiC
163163
L’expLoitation des ressources animaLes pendant Le 9e miLLénaire en égée et Le statut ambigu des suidés
SARPAKI A. (2011) - Archaeobotanical Seed Remains, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, Bone Tool Industries, Dietary Resources and the Paleoenvironment, and Archeometrical Studies, vol II, Philadelphia, INSTAP, Monograph Series, p. 315-324.
SCANDURA M., IACOLINA L., CRESTANELLO B., PECCHIOLI E., DI BENEDETTO M.F., RUSSO V., DAVOLI R., APOLLONIO M., BERTORELLE G. (2008) - Ancient vs. recent processes as factors shaping the genetic variation of the European wild boar: are the effects of the last glaciation still detectable? Molecular Ecology, 17 (7), p. 1745-1762.
SHACKLETON J.C. (1988) - Excavations at Franchthi Cave, Greece, fasc. 4. Marine Molluscan Remains from Franchthi Cave, Bloomington, Indiana, Indiana University Press, 208 p.
SORDINAS A. (1969) - Investigations in the prehistory of Corfu during 1964-66, Balkan Studies, 10/2, p. 393-424.
STARKOVICH B.M., STINER M.C. (2010) - Upper Palaeolithic Animal Exploitation at Klissoura cave 1 in Southern Greece: Dietary trends and Mammal Taphonomy, Eurasian Prehistory, 7 (2), p. 107–132.
STINER M.C., MUNRO N.D. (2011) - On the evolution of diet and landscape during the Upper Paleolithic through Mesolithic at Franchthi Cave (Peloponnese, Greece), Journal of Human Evolution, 60, p. 618-636.
STRASSER T. F., PANAGOPOULOU E., RUNNELS C.N., MURRAY P.M., THOMPSON N., KARKANAS P., MCCOY F.W., WEGMANN K.W (2010) - Stone Age Seafaring in the Mediterranean: Evidence from the Plakias Region for Lower Palaeolithic and Mesolithic Habitation of Crete, Hesperia, 79, p. 145-190.
STROULIA A. (2010) - Excavations at Franchthi Cave, Greece, fasc. 14. Flexible stones: ground stone tools from Franchthi Cave, Bloomington, Indiana, Indiana University Press, 242 p.
TRANTALIDOU K. (2003) - Faunal remains from the earliest strata of the Cave of Cyclope, Youra, in N. Galanidou, C. Perlès (dir.), The Greek Mesolithic. Problems and perspectives, London, British School of Athens, p. 143-172.
TRANTALIDOU K. (2008) - Glimpses of Aegean Island Communities during the Mesolithic and Neolithic periods: The zooarchaeological point of view, in N. Brodie, J. Doole, G. Gavalas, C. Renfrew (dir.), Ορίζων. A Colloquium on the Prehistory of the Cyclades, Mc Donald Institute for Archaeological Research, University of Cambridge, p. 19-27.
TRANTALIDOU K. (2010) - Dietary Adaptations of Coastal People in the Aegean Archipelago during the Mesolithic Period: The Macrofauna Assemblages of Maroulas on Kythnos, in A. Sampson, M. Kaczanowska, J. Kozłowski (dir.), Τhe prehistory of the island of Kythnos and the Mesolithic settlement at Maroulas, Krakow, Polish Academy of Sciences and Arts and University of the Aegean, p. 163-177.
TRANTALIDOU K. (2011) - From Mesolithic Fisherman and Fish Hunters to Neolithic Goat herders: The transformation of an island economy in the Aegean, in A. Sampson (dir.), Τhe Cyclops Cave on the island of Youra, Greece. Mesolithic and Neolithic networks in the Northern Aegean Basin, Bone Tool Industries, Dietary Resources and the Paleoenvironment, and Archeometrical Studies, vol II, Philadelphia, INSTAP, Monograph Series, p. 53-150.
TRANTALIDOU K., BELEGRINOU EL., ANDREASEN N. (2011) - Pastoral societies in the southern Balkan Peninsula: The evidence from caves occupied during the Neolithic and the Chalcolithic era, The Phenomena of Cultural Borders and Border Cultures across the Passage of the Time, Trnava University, Anodos 10 (2010), p. 295-320.
VALAMOTI S.-M., KOTSAKIS K. (2007) - Transitions to agriculture in the Aegean: the archaeobotanical evidence, S. Colledge, J. Connoly (dir.), The origins and spread of domestic plants in southwest Asia and Europe, London, University College, Institute of Archaeology, p. 75-90.
VIGNE J.-D. (2011) - Les suinés (Sus scrofa), in J. Guilaine, F. Briois, J.-D. Vigne (dir.), Shillourokambos. Un établissement néolithique pré-céramique à Chypre. Les fouilles du Secteur 1, Paris, Errance & École française d’Athènes, p. 919-969.
VIGNE J.-D., ZAZZO A., SALIÈGE J.-F., POPLIN F., GUILAINE J., SIMMONS A. (2009) - Pre-Neolithic wild boar management and introduction to Cyprus more than 11,400 years ago, Proceedings of the National Academy of Sciences of the United States of America, 106, 38, p. 16131-16138.
WASSE A. (2000) - The development of goat and sheep herding during the Levantine Neolithic. Unpublished PhD thesis, Univeristy of London.
ZEDER M. A. (2001) - A Metrical Analysis of a Collection of Modern Goats (Capra hircus aegargus and Capra hircus hircus) from Iran and Iraq: Implications for the Study of Caprine Domestication, Journal of Archaeological Science, 28, p. 61-79.
ZEDER M. A., HESSE B. (2000) - The Initial Domestication of Goats (Capra hircus) in the Zagros Mountains 10,000 Years Ago, Science, 287, p. 2254-2257.




















