
Leucine transport in membrane vesicles fromChironomus riparius larvae displays a m�lange of...
12
Archives of Insect Biochemistry and Physiology 48:51–62 (2001) © 2001 Wiley-Liss, Inc. Leucine Transport in Membrane Vesicles From Chironomus riparius Larvae Displays a Mélange of Crown-Group Features Paolo Parenti, 1 * Matilde Forcella, 1 Anna Pugliese, 1 Roberto Giacchini, 1 Bruno Rossaro, 2 and Giorgio M. Hanozet 1 1 Department of Environmental Sciences, University of Milan-Bicocca, Milano, Italy 2 Department of Biology, University of Milano, Milano, Italy Leucine uptake into membrane vesicles from larvae of the midge Chironomus riparius was studied. The membrane prepa- ration was highly enriched in typical brush border membrane enzymes and depleted of other membrane contaminants. In the absence of cations, there was a stereospecific uptake of L-leu- cine, which exhibited saturation kinetics. Parameters were de- termined both at neutral (K m 33 ± 5 mM and V max 22.6 ± 6.8 pmol/ 7s/mg protein) and alkaline (K m 46 ± 5 mM and V max 15.5 ± 2.5 pmol/7s/mg protein) pH values. At alkaline pH, external sodium increased the affinity for leucine (K m 17 ± 1 mM) and the maxi- mal uptake rate (V max 74.0 ± 12.5 pmol/7s/mg protein). Stimula- tion of leucine uptake by external alkaline pH agreed with lumen pH measurements in vivo. Competition experiments in- dicated that at alkaline pH, the transport system readily ac- cepts most L-amino acids, including branched, unbranched, and a-methylated amino acids, histidine and lysine, but has a low affinity for phenylalanine, b-amino acids, and N-methylated amino acids. At neutral pH, the transport has a decreased af- finity for lysine, glycine, and a-methylleucine. Taken together, these data are consistent with the presence in midges of two distinct leucine transport systems, which combine characters of the lepidopteran amino acid transport system and of the sodium-dependent system from lower neopterans. Arch. Insect Biochem. Physiol. 48:51–62, 2001. © 2001 Wiley-Liss, Inc. Key words: amino acid transport; membrane vesicles; midge larvae; Chir- onomus riparius Abbreviations used: CAPS = 3-cyclohexylamino-1-propan- sulfonic acid; HEPES = 4-(2-hydroxyethyl)-1-piperazine- ethansulfonic acid; MES = 4-morpholineethansulfonic acid; TEP = transepithelial electrical potential; TMA = tetram- ethylammonium. Contract grant sponsor: Italian MURST. *Correspondence to: Dr. Paolo Parenti, Dipartimento di Scienze dell’Ambiente e del Territorio, P.zza della Scienza 1, 20126 Milano, Italy. E-mail: [email protected] Received 30 October 2000; Accepted 25 March 2001 INTRODUCTION The insect midgut is the site of nutrients up- take and the cell membrane of the enterocytes contains either passive diffusion systems or cat- ion-driven symporter (for a recent review, see Wolfersberger, 2000). Most of what we know of the molecular physiology of nutrient absorption in insect midgut comes from studies on brush bor- der membrane vesicles from lepidopteran larvae,
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Leucine transport in membrane vesicles fromChironomus riparius larvae displays a m�lange of...

Archives of Insect Biochemistry and Physiology 48:51–62 (2001)
© 2001 Wiley-Liss, Inc.
Leucine Transport in Membrane Vesicles FromChironomus riparius Larvae Displays a Mélange
of Crown-Group FeaturesPaolo Parenti,1* Matilde Forcella,1 Anna Pugliese,1 Roberto Giacchini,1 Bruno Rossaro,2
and Giorgio M. Hanozet1
1Department of Environmental Sciences, University of Milan-Bicocca, Milano, Italy2Department of Biology, University of Milano, Milano, Italy
Leucine uptake into membrane vesicles from larvae of themidge Chironomus riparius was studied. The membrane prepa-ration was highly enriched in typical brush border membraneenzymes and depleted of other membrane contaminants. In theabsence of cations, there was a stereospecific uptake of L-leu-cine, which exhibited saturation kinetics. Parameters were de-termined both at neutral (Km 33 ± 5 mM and Vmax 22.6 ± 6.8 pmol/7s/mg protein) and alkaline (Km 46 ± 5 mM and Vmax 15.5 ± 2.5pmol/7s/mg protein) pH values. At alkaline pH, external sodiumincreased the affinity for leucine (Km 17 ± 1 mM) and the maxi-mal uptake rate (Vmax 74.0 ± 12.5 pmol/7s/mg protein). Stimula-tion of leucine uptake by external alkaline pH agreed withlumen pH measurements in vivo. Competition experiments in-dicated that at alkaline pH, the transport system readily ac-cepts most L-amino acids, including branched, unbranched, anda-methylated amino acids, histidine and lysine, but has a lowaffinity for phenylalanine, b-amino acids, and N-methylatedamino acids. At neutral pH, the transport has a decreased af-finity for lysine, glycine, and a-methylleucine. Taken together,these data are consistent with the presence in midges of twodistinct leucine transport systems, which combine charactersof the lepidopteran amino acid transport system and of thesodium-dependent system from lower neopterans. Arch. InsectBiochem. Physiol. 48:51–62, 2001. © 2001 Wiley-Liss, Inc.
Key words: amino acid transport; membrane vesicles; midge larvae; Chir-onomus riparius
Abbreviations used: CAPS = 3-cyclohexylamino-1-propan-sulfonic acid; HEPES = 4-(2-hydroxyethyl)-1-piperazine-ethansulfonic acid; MES = 4-morpholineethansulfonic acid;TEP = transepithelial electrical potential; TMA = tetram-ethylammonium.
Contract grant sponsor: Italian MURST.
*Correspondence to: Dr. Paolo Parenti, Dipartimento diScienze dell’Ambiente e del Territorio, P.zza della Scienza 1,20126 Milano, Italy. E-mail: [email protected]
Received 30 October 2000; Accepted 25 March 2001
INTRODUCTION
The insect midgut is the site of nutrients up-take and the cell membrane of the enterocytescontains either passive diffusion systems or cat-ion-driven symporter (for a recent review, seeWolfersberger, 2000). Most of what we know ofthe molecular physiology of nutrient absorptionin insect midgut comes from studies on brush bor-der membrane vesicles from lepidopteran larvae,

52 Parenti et al.
where several K+-dependent amino acid transportsystems have been identified and characterized(Hanozet et al., 1980; Parenti et al., 1992; Henni-gan et al., 1993; Reuveni and Dunn, 1994; Partha-sarathy et al., 1994; Giordana et al. 1998; Castagnaet al., 1998). Within living (that is crown-group)neopterans, several mechanisms of amino acid up-take have evolved. A K+-driven transport systemis probably present in Hemiptera (Silva and Terra,1994) other than Lepidoptera, whereas cation-in-dependent transport systems have been describedin the coleopteran Leptinotarsa decemlineata(Reuveni et al., 1993). In contrast, Na+ gradient-dependent transport system has been found in thecockroach Blabera gigantea (Parenti et al., 1986)and it is possibly present also within orthopteroids(unpublished observations).
Many dipteran larvae share with lepidopt-eran larvae the trait of a highly alkaline midgutlumen (Dadd, 1975; Dow, 1986). Recent resultsindicate that midgut alkalization in mosquito lar-vae is achieved by molecular mechanisms that aresimilar in lepidopteran and dipteran larvae(Zhuang et al., 1999). Phylogenetic analysis is con-sistent with the hypothesis that this charactermight have arisen in a common ancestor ofDiptera and Lepidoptera (Clark, 1999). Althoughthis closeness suggests similarities in the epithe-lial transport of ions and metabolites, it has beensuggested that the transepithelial electrical po-tential (TEP) is generated in dipterans by funda-mentally different mechanisms from those thatgenerate the potential difference in lepidopteranlarvae (Clark et al., 1999). In the latter, an elec-trogenic active transport mechanism (Harvey andNedergaard, 1964), constituted by an H+-translo-cating V-ATPase and an electrophoretic K+/nH+
antiport, is responsible for the generation of a lu-men-positive TEP (Wieczorek et al., 1989). A V-ATPase has also been localized in the midgut offreshwater mosquitoes (Zhuang et al., 1999), butits contribution to TEP generation has not beenestablished. Two features have been claimed tocontribute to a model for midge larvae: an elec-trogenic sodium pump secreting sodium into theblood and a high degree of independence of anionand cation transport (Wright, 1975). To investi-gate which driving forces are needed for solutetransport in dipteran larvae midgut, we havecharacterized the amino acid uptake into mem-
brane vesicles prepared from the midge Chiron-omus riparius. Our results show the presence inthese dipteran larvae of transport systems thatcombine characters found in several neopteranlineages.
MATERIALS AND METHODSAnimals
Chironomus riparius larvae were collected inthe wild or purchased alive from a wholesale baitand fish stocking farm (Eschematteo, Parma,Italy). Larvae were maintained in large vesselsin a 2-cm water layer at 4°C and reared withTetramin or yeast powder.
Chemicals
L-[4,5-3H]-leucine 152 Ci/mmol was pur-chased from Amersham International plc, UK.Amino acids and derivatives, the protonophorecarbonyl-cyanide p-trifluoromethoxyphenylhydra-zone (FCCP), were from Sigma (St. Louis, MO).
Membrane Preparation
Membrane vesicles were prepared from ei-ther fresh or frozen 4th instar larvae that hadbeen stored in liquid nitrogen by the following mi-nor modification of the method described byWolfersberger et al. (1987). Briefly, larvae (roughly3 g for experiment) were suspended with 9 vol-umes of a hypotonic Tris buffer (Tris-HCl 17 mM,pH 7.4, mannitol 100 mM, EGTA 5 mM) and ho-mogenized at 4°C in a Waring-mixer, set at 11,400rpm, for two 2-min periods, separated by a 2-mincooling interval. The suspension was then filteredthrough two layers of surgical gauze and 1-mlaliquots were withdrawn for enzyme assays. Tothe recovered volume, an equal volume of 24 mMMgCl2 was added. The resulting mixture was thor-oughly stirred for 15 min in an ice bath and thencentrifuged at 2,500g for 15 min at 4°C. The re-sulting pellet was discarded and the supernatantcentrifuged at 48,000g for 20 min at 4°C. The pel-let was resuspended in 0.5 homogenate volumeof ice cold Tris buffer and homogenized in a glassand teflon Potter-Helvehjem, two 6-strokes at2,000 rpm, separated by 1 min in ice. An equalvolume of 24 mM MgCl2 was added, the mixturestirred for 15 min in an ice bath and centrifugedas above at low speed and then at high speed.

Amino Acid Transport in Midge Larvae 53
The final pellet was resuspended in 1 vol homo-genate buffer, homogenized in Potter-Helvehjemas before, and centrifuged again at 48,000g for20 min at 4°C. Unless otherwise stated, the ob-tained pellet was resuspended in HEPES1-Tris10 mM, pH 7.4, mannitol 100 mM, and repre-sented the membrane preparation used for trans-port experiments. The recovery was about 0.8 mgmembrane proteins/g fresh larvae. For transportexperiment, the suspension was made at a pro-tein concentration of 2–3 mg/ml as assayed ac-cording to Bradford (1976) using bovine serumalbumin as standard.
Amino Acid Transport Measurements
Tracer experiments were performed by incu-bating 1 volume of vesicles and 4 volumes of a ra-dioactive cocktail buffered at different final pHvalues, containing 30 µCi L-[3H]leucine and the sub-stances specified in the figure legends. Uptakes werecarried out according to the rapid filtration tech-nique as described elsewhere (Giordana et al., 1982).
Electron Microscopy
The membrane pellet was fixed in 3% glut-araldehyde in 0.1 M sodium cacodylate buffer, pH7.3, for 2 h and 30 min. Afterward, it was washedin cacodylate buffer and post-fixed in 1% OsO4/0.05M HCl/veronal buffer at pH 7.4. The pellet was de-hydrated in ethanol and embedded in Epon 812.Membranes were stained with 4% uranyl acetate.
Enzyme AssaysAssays of the activity of each enzyme were
performed in duplicate on the day of the prepa-ration. Alkaline phosphatase was assayed at pH8.0 using p-nitrophenylphosphate as substrateand leucine aminopeptidase was measured at pH7.4 using L-leucine-p-nitroanilide as substrate. Cy-tochrome c oxidase was measured according toSmith (1955). Na,K-ATPase was assayed accord-ing to Quigley and Gotterer (1969) using theFiske-Subbarow reagent for phosphate determi-nation as modified by Cross et al. (1978), exceptthat, before assay, samples were incubated 30 minat 20°C with 0.06 mg deoxycholic acid/mg pro-teins. Na,K-ATPase activity represents the dif-ference between the activity measured in theabsence and presence of 1 mM ouabain. All as-says were performed at 25°C in a Cary3 Spectro-photometer, recorded on a personal computer, and
analyzed by the Cary WinUV application softwarefor Windows 98.
pH Measurements In Vivo
To estimate the lumen pH, single larvae weredissected alive on a microscopic slide. The intes-tine was isolated and examined under a Leitz ste-reomicroscope in a drop of a saturated solutionof each of the following indicators, Thymol Blue,Phenolphthalein, and Cresol Red. Segments werecompared visually with standards prepared insteps of approximately 0.5 pH units.
Calculations
Experimental data of uptake measurementsare given ± S.E. of triplicate determinations andwere analyzed by computer and plotted using amultiparameter, iterative, nonlinear regressionprogram based on the Marquardt-Levenberg al-gorithm (Sigma Plot 5.0, Jandel, CA). The calcu-lated constants are given ± S.D.
RESULTS
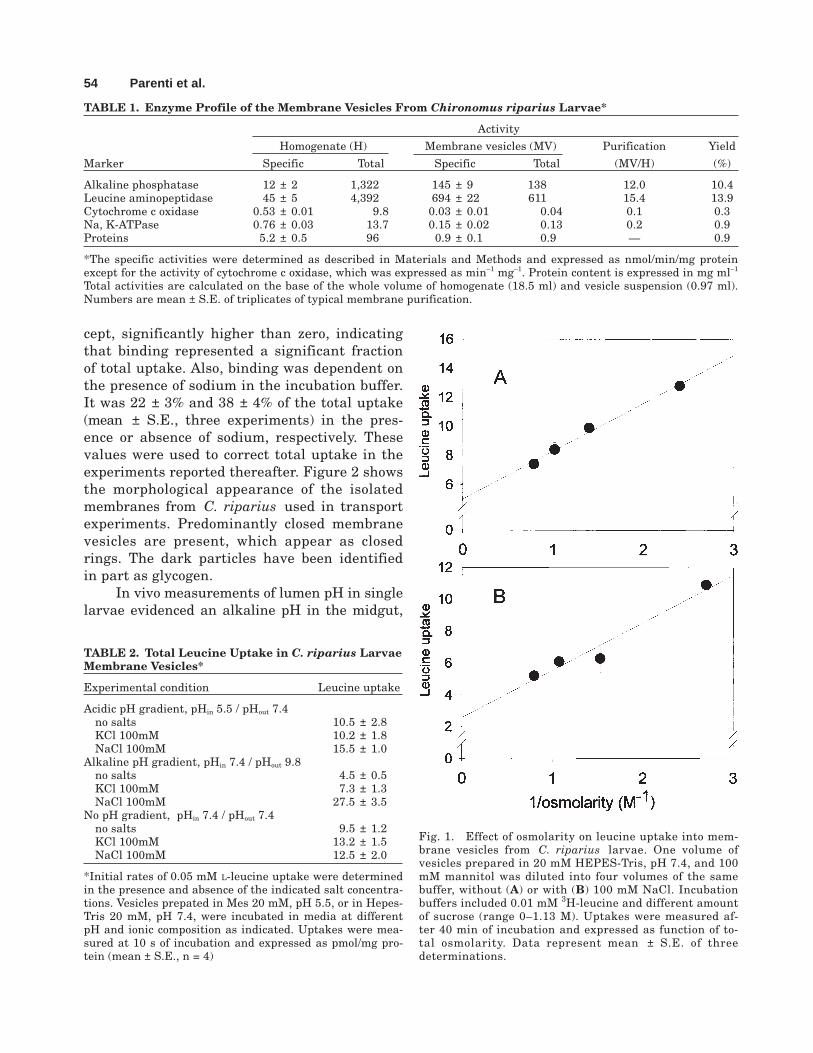
The enzyme profile of the membrane suspen-sion obtained from Chironomus riparius larvaewas consistent with a preparation constitutedmainly of brush border membranes. Table 1shows that typical brush border marker enzymeswere highly enriched in our preparation, whereasmarker enzymes of the other cellular compart-ments had an enrichment factor lower than 1.The high degree of purity of the preparation wasaccompanied by a relatively low yield of totalmembrane proteins, which is probably due to themulti-step purification protocol. The enrichmentof marker enzymes encouraged us to undertakea full characterization of amino acid transportin the membrane preparation. Preliminary tracerexperiments showed a significant leucine uptake,which depended on both medium pH and thepresence of sodium ions (Table 2). To evaluatewhich part of the total uptake actually repre-sents merely binding to the membranes, the ef-fect of medium osmolarity on leucine uptake wasdetermined. As expected for a suspension repre-sented by closed vesicles, leucine uptake linearlydecreased as osmolarity increased (Fig. 1). In ourconditions, extrapolation to infinite osmolarity(i.e., intravesicular volume null) gives an inter-
54 Parenti et al.
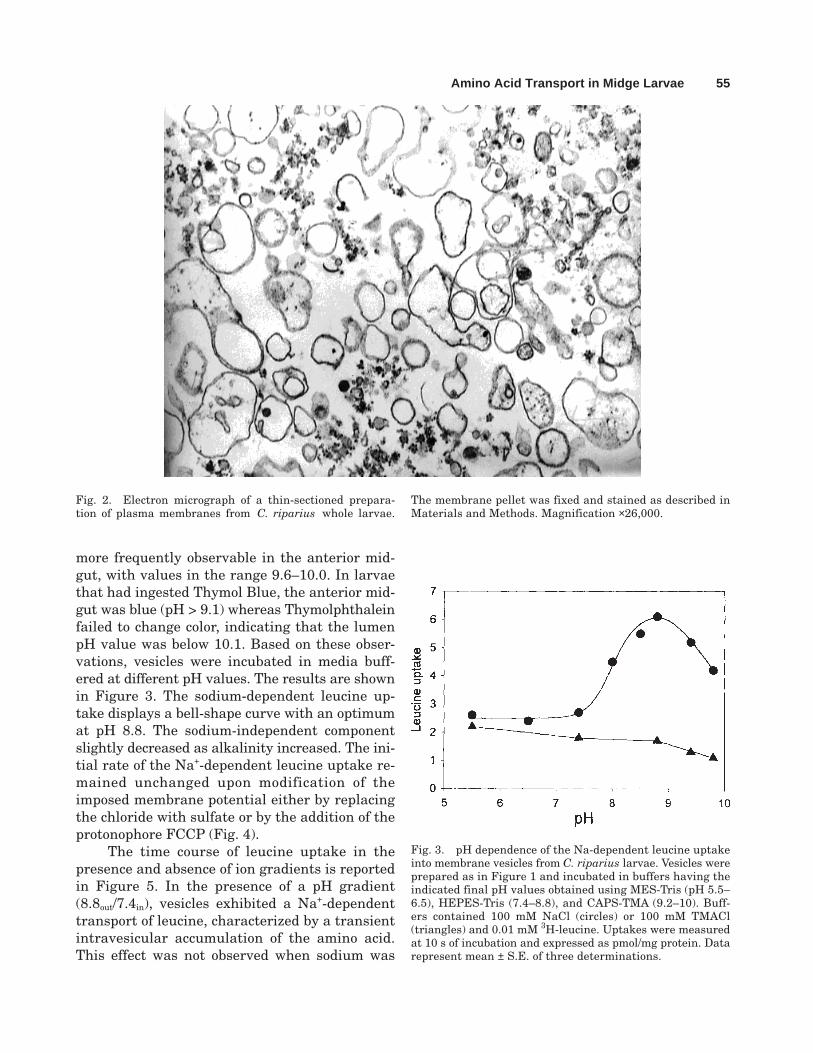
cept, significantly higher than zero, indicatingthat binding represented a significant fractionof total uptake. Also, binding was dependent onthe presence of sodium in the incubation buffer.It was 22 ± 3% and 38 ± 4% of the total uptake(mean ± S.E., three experiments) in the pres-ence or absence of sodium, respectively. Thesevalues were used to correct total uptake in theexperiments reported thereafter. Figure 2 showsthe morphological appearance of the isolatedmembranes from C. riparius used in transportexperiments. Predominantly closed membranevesicles are present, which appear as closedrings. The dark particles have been identifiedin part as glycogen.
In vivo measurements of lumen pH in singlelarvae evidenced an alkaline pH in the midgut,
Fig. 1. Effect of osmolarity on leucine uptake into mem-brane vesicles from C. riparius larvae. One volume ofvesicles prepared in 20 mM HEPES-Tris, pH 7.4, and 100mM mannitol was diluted into four volumes of the samebuffer, without (A) or with (B) 100 mM NaCl. Incubationbuffers included 0.01 mM 3H-leucine and different amountof sucrose (range 0–1.13 M). Uptakes were measured af-ter 40 min of incubation and expressed as function of to-tal osmolarity. Data represent mean ± S.E. of threedeterminations.
TABLE 1. Enzyme Profile of the Membrane Vesicles From Chironomus riparius Larvae*
ActivityHomogenate (H) Membrane vesicles (MV) Purification Yield
Marker Specific Total Specific Total (MV/H) (%)
Alkaline phosphatase 12 ± 2 1,322 145 ± 9 138 12.0 10.4Leucine aminopeptidase 45 ± 5 4,392 694 ± 22 611 15.4 13.9Cytochrome c oxidase 0.53 ± 0.01 9.8 0.03 ± 0.01 0.04 0.1 0.3Na, K-ATPase 0.76 ± 0.03 13.7 0.15 ± 0.02 0.13 0.2 0.9Proteins 5.2 ± 0.5 96 0.9 ± 0.1 0.9 — 0.9
*The specific activities were determined as described in Materials and Methods and expressed as nmol/min/mg proteinexcept for the activity of cytochrome c oxidase, which was expressed as min–1 mg–1. Protein content is expressed in mg ml–1
Total activities are calculated on the base of the whole volume of homogenate (18.5 ml) and vesicle suspension (0.97 ml).Numbers are mean ± S.E. of triplicates of typical membrane purification.
TABLE 2. Total Leucine Uptake in C. riparius LarvaeMembrane Vesicles*
Experimental condition Leucine uptake
Acidic pH gradient, pHin 5.5 / pHout 7.4no salts 10.5 ± 2.8KCl 100mM 10.2 ± 1.8NaCl 100mM 15.5 ± 1.0
Alkaline pH gradient, pHin 7.4 / pHout 9.8no salts 4.5 ± 0.5KCl 100mM 7.3 ± 1.3NaCl 100mM 27.5 ± 3.5
No pH gradient, pHin 7.4 / pHout 7.4no salts 9.5 ± 1.2KCl 100mM 13.2 ± 1.5NaCl 100mM 12.5 ± 2.0
*Initial rates of 0.05 mM L-leucine uptake were determinedin the presence and absence of the indicated salt concentra-tions. Vesicles prepated in Mes 20 mM, pH 5.5, or in Hepes-Tris 20 mM, pH 7.4, were incubated in media at differentpH and ionic composition as indicated. Uptakes were mea-sured at 10 s of incubation and expressed as pmol/mg pro-tein (mean ± S.E., n = 4)
Amino Acid Transport in Midge Larvae 55
more frequently observable in the anterior mid-gut, with values in the range 9.6–10.0. In larvaethat had ingested Thymol Blue, the anterior mid-gut was blue (pH > 9.1) whereas Thymolphthaleinfailed to change color, indicating that the lumenpH value was below 10.1. Based on these obser-vations, vesicles were incubated in media buff-ered at different pH values. The results are shownin Figure 3. The sodium-dependent leucine up-take displays a bell-shape curve with an optimumat pH 8.8. The sodium-independent componentslightly decreased as alkalinity increased. The ini-tial rate of the Na+-dependent leucine uptake re-mained unchanged upon modification of theimposed membrane potential either by replacingthe chloride with sulfate or by the addition of theprotonophore FCCP (Fig. 4).
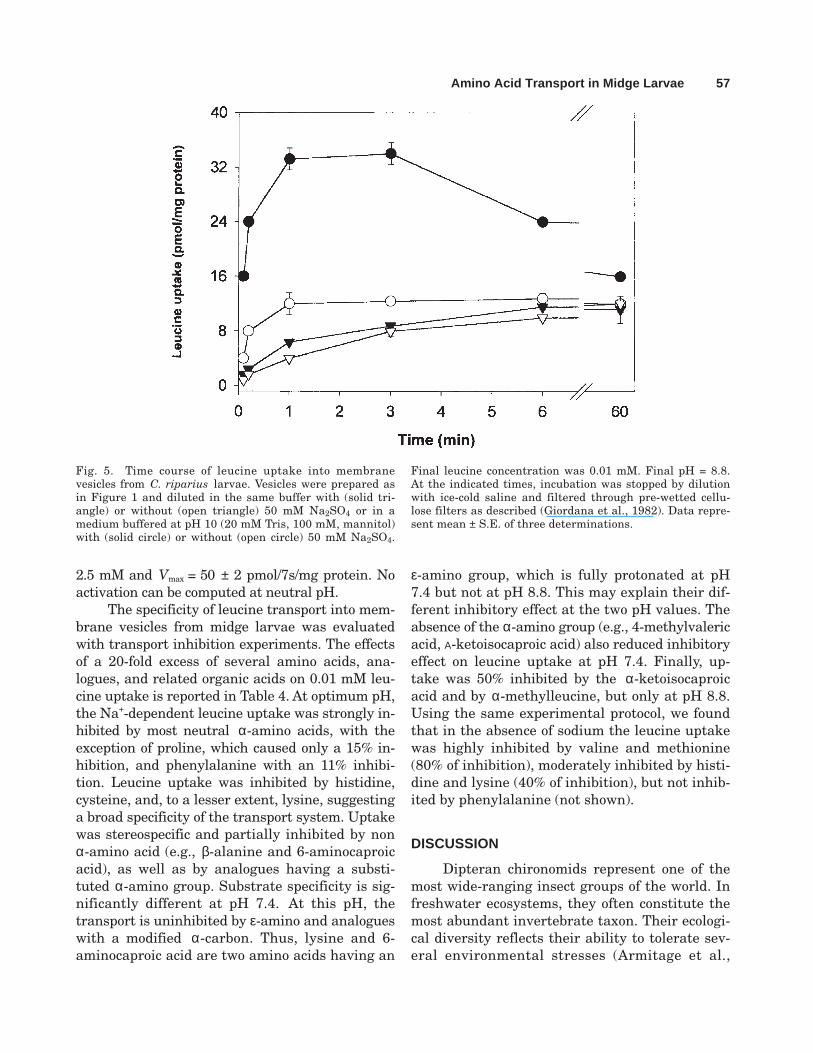
The time course of leucine uptake in thepresence and absence of ion gradients is reportedin Figure 5. In the presence of a pH gradient(8.8out/7.4in), vesicles exhibited a Na+-dependenttransport of leucine, characterized by a transientintravesicular accumulation of the amino acid.This effect was not observed when sodium was
Fig. 2. Electron micrograph of a thin-sectioned prepara-tion of plasma membranes from C. riparius whole larvae.
The membrane pellet was fixed and stained as described inMaterials and Methods. Magnification ×26,000.
Fig. 3. pH dependence of the Na-dependent leucine uptakeinto membrane vesicles from C. riparius larvae. Vesicles wereprepared as in Figure 1 and incubated in buffers having theindicated final pH values obtained using MES-Tris (pH 5.5–6.5), HEPES-Tris (7.4–8.8), and CAPS-TMA (9.2–10). Buff-ers contained 100 mM NaCl (circles) or 100 mM TMACl(triangles) and 0.01 mM 3H-leucine. Uptakes were measuredat 10 s of incubation and expressed as pmol/mg protein. Datarepresent mean ± S.E. of three determinations.

56 Parenti et al.
replaced by potassium or in the absence of addedsalts. At neutral pH, leucine uptake was not en-hanced by the presence of Na+ salts or K+ salts.
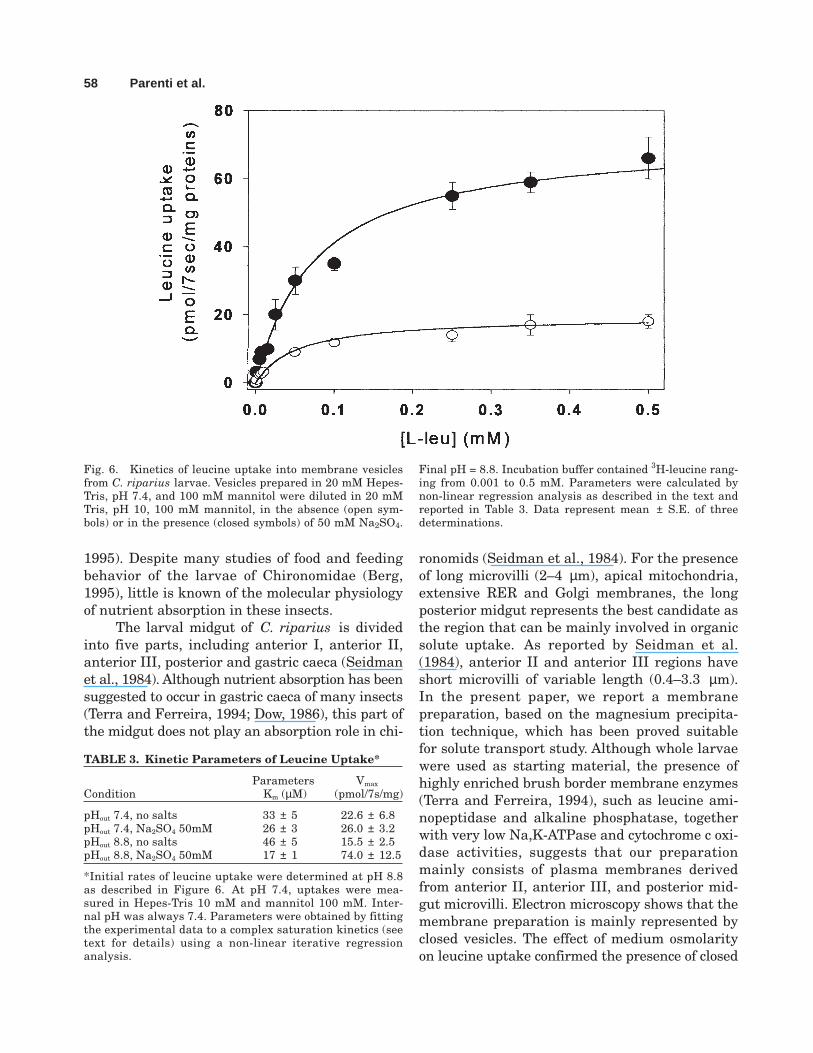
Measurements of leucine uptake as a func-tion of external leucine concentration producedsaturation kinetics both in the presence and ab-sence of sodium (Fig. 6) either at neutral or alka-line external pH (Table 3). Kinetic parameterswere calculated by fitting the experimental datato the following equation:
v0 = Vmax · [Leu]/ (Kleu + [Leu]) + C · [Leu] (1)
where v0 is the initial rate of leucine uptake; Vmax,the maximal initial rate; Kleu is the Michaelis-Menten constant for leucine; and C is the diffu-sional component. From data reported in Table3, it is evident that sodium mainly affects themaximal velocity, i.e., the turnover number, whichrepresents the rate-limiting step for membranetransporters. The Vmax value is 4.8-fold higher in
the presence of Na+ than in its absence. Sodiumalso affects the Kleu value, which is 2.7fold lowerthan that measured in the absence of salts. Pas-sive transport represented by the C value remainsunchanged. Leucine kinetics at pH 7.4 was a satu-rable process with Kleu and Vmax values not af-fected by the presence of sodium.
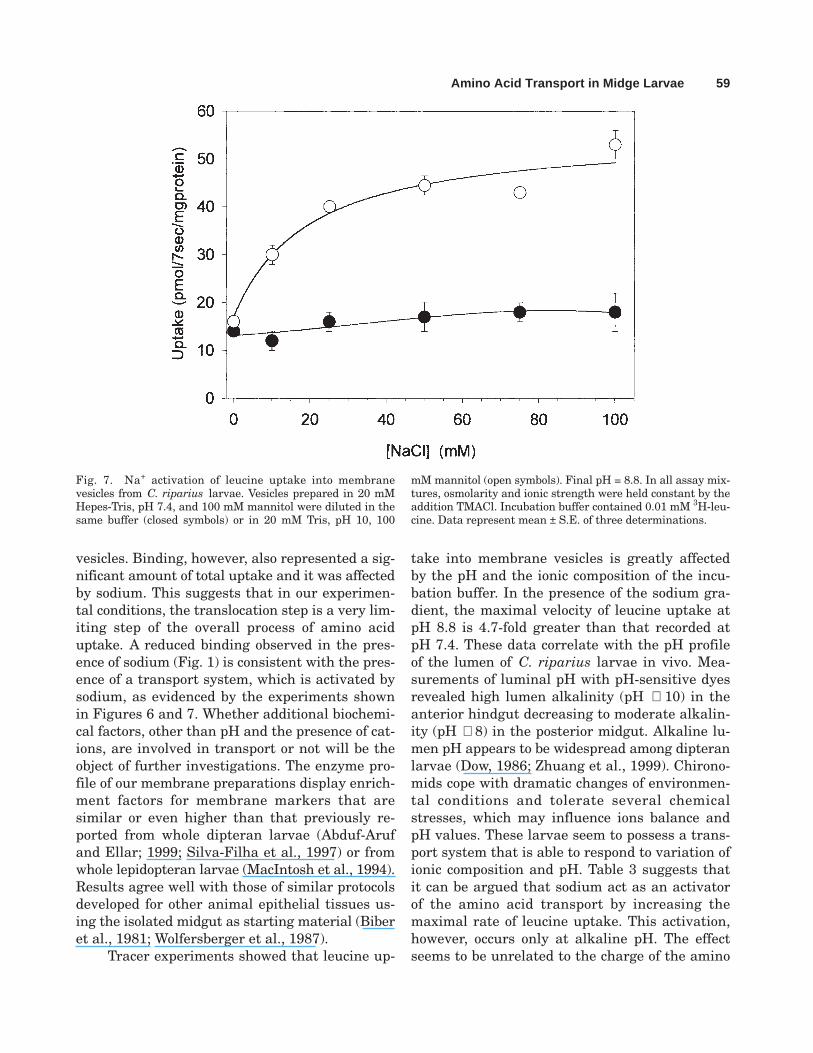
The Na+ dependence of leucine uptake wasexplored further by measuring the Na+ activationcurve at a fixed leucine concentration (0.01 mM).An experiment is illustrated in Figure 7. As thecis Na+ increased from 0 to 100 mM, there is ahyperbolic activation of leucine uptake when thepHout is 8.8 but not when it is 7.4. Data were fit-ted with the following equation:
v0 = Vmax · [Na]/ (KNa + [Na]) + C′ (2)
where v0, Vmax, and KNa are as usually defined,whereas C′ represents the Na+-independent leucineuptake. In the experiment illustrated, KNa = 14.5 ±
Fig. 4. Effect of sodium on leucine uptake into membranevesicles from C. riparius larvae. Vesicles were prepared anddiluted as in Figure 1. Incubation buffers were made with20 mM HEPES-Tris, pH 7.4, 100 mM mannitol or 20 mMCAPS, pH 11, 100 mM mannitol, with (+) 100 mM NaCl or
50 mM Na2SO4 or with (–)100 mM TMACl or 50 mMTMA2SO4. Incubation buffers included 0.01 mM 3H-leucine.Final pH = 9.8. One assay contained 80 µM FCCP. Datarepresent mean ± S.E. of three determinations.
Amino Acid Transport in Midge Larvae 57
2.5 mM and Vmax = 50 ± 2 pmol/7s/mg protein. Noactivation can be computed at neutral pH.
The specificity of leucine transport into mem-brane vesicles from midge larvae was evaluatedwith transport inhibition experiments. The effectsof a 20-fold excess of several amino acids, ana-logues, and related organic acids on 0.01 mM leu-cine uptake is reported in Table 4. At optimum pH,the Na+-dependent leucine uptake was strongly in-hibited by most neutral α-amino acids, with theexception of proline, which caused only a 15% in-hibition, and phenylalanine with an 11% inhibi-tion. Leucine uptake was inhibited by histidine,cysteine, and, to a lesser extent, lysine, suggestinga broad specificity of the transport system. Uptakewas stereospecific and partially inhibited by nonα-amino acid (e.g., β-alanine and 6-aminocaproicacid), as well as by analogues having a substi-tuted α-amino group. Substrate specificity is sig-nificantly different at pH 7.4. At this pH, thetransport is uninhibited by ε-amino and analogueswith a modified α-carbon. Thus, lysine and 6-aminocaproic acid are two amino acids having an
ε-amino group, which is fully protonated at pH7.4 but not at pH 8.8. This may explain their dif-ferent inhibitory effect at the two pH values. Theabsence of the α-amino group (e.g., 4-methylvalericacid, Α-ketoisocaproic acid) also reduced inhibitoryeffect on leucine uptake at pH 7.4. Finally, up-take was 50% inhibited by the α-ketoisocaproicacid and by α-methylleucine, but only at pH 8.8.Using the same experimental protocol, we foundthat in the absence of sodium the leucine uptakewas highly inhibited by valine and methionine(80% of inhibition), moderately inhibited by histi-dine and lysine (40% of inhibition), but not inhib-ited by phenylalanine (not shown).
DISCUSSION
Dipteran chironomids represent one of themost wide-ranging insect groups of the world. Infreshwater ecosystems, they often constitute themost abundant invertebrate taxon. Their ecologi-cal diversity reflects their ability to tolerate sev-eral environmental stresses (Armitage et al.,
Fig. 5. Time course of leucine uptake into membranevesicles from C. riparius larvae. Vesicles were prepared asin Figure 1 and diluted in the same buffer with (solid tri-angle) or without (open triangle) 50 mM Na2SO4 or in amedium buffered at pH 10 (20 mM Tris, 100 mM, mannitol)with (solid circle) or without (open circle) 50 mM Na2SO4.
Final leucine concentration was 0.01 mM. Final pH = 8.8.At the indicated times, incubation was stopped by dilutionwith ice-cold saline and filtered through pre-wetted cellu-lose filters as described (Giordana et al., 1982). Data repre-sent mean ± S.E. of three determinations.
58 Parenti et al.
1995). Despite many studies of food and feedingbehavior of the larvae of Chironomidae (Berg,1995), little is known of the molecular physiologyof nutrient absorption in these insects.
The larval midgut of C. riparius is dividedinto five parts, including anterior I, anterior II,anterior III, posterior and gastric caeca (Seidmanet al., 1984). Although nutrient absorption has beensuggested to occur in gastric caeca of many insects(Terra and Ferreira, 1994; Dow, 1986), this part ofthe midgut does not play an absorption role in chi-
ronomids (Seidman et al., 1984). For the presenceof long microvilli (2–4 µm), apical mitochondria,extensive RER and Golgi membranes, the longposterior midgut represents the best candidate asthe region that can be mainly involved in organicsolute uptake. As reported by Seidman et al.(1984), anterior II and anterior III regions haveshort microvilli of variable length (0.4–3.3 µm).In the present paper, we report a membranepreparation, based on the magnesium precipita-tion technique, which has been proved suitablefor solute transport study. Although whole larvaewere used as starting material, the presence ofhighly enriched brush border membrane enzymes(Terra and Ferreira, 1994), such as leucine ami-nopeptidase and alkaline phosphatase, togetherwith very low Na,K-ATPase and cytochrome c oxi-dase activities, suggests that our preparationmainly consists of plasma membranes derivedfrom anterior II, anterior III, and posterior mid-gut microvilli. Electron microscopy shows that themembrane preparation is mainly represented byclosed vesicles. The effect of medium osmolarityon leucine uptake confirmed the presence of closed
Fig. 6. Kinetics of leucine uptake into membrane vesiclesfrom C. riparius larvae. Vesicles prepared in 20 mM Hepes-Tris, pH 7.4, and 100 mM mannitol were diluted in 20 mMTris, pH 10, 100 mM mannitol, in the absence (open sym-bols) or in the presence (closed symbols) of 50 mM Na2SO4.
Final pH = 8.8. Incubation buffer contained 3H-leucine rang-ing from 0.001 to 0.5 mM. Parameters were calculated bynon-linear regression analysis as described in the text andreported in Table 3. Data represent mean ± S.E. of threedeterminations.
TABLE 3. Kinetic Parameters of Leucine Uptake*
Parameters VmaxCondition Km (µM) (pmol/7s/mg)
pHout 7.4, no salts 33 ± 5 22.6 ± 6.8pHout 7.4, Na2SO4 50mM 26 ± 3 26.0 ± 3.2pHout 8.8, no salts 46 ± 5 15.5 ± 2.5pHout 8.8, Na2SO4 50mM 17 ± 1 74.0 ± 12.5
*Initial rates of leucine uptake were determined at pH 8.8as described in Figure 6. At pH 7.4, uptakes were mea-sured in Hepes-Tris 10 mM and mannitol 100 mM. Inter-nal pH was always 7.4. Parameters were obtained by fittingthe experimental data to a complex saturation kinetics (seetext for details) using a non-linear iterative regressionanalysis.
Amino Acid Transport in Midge Larvae 59
vesicles. Binding, however, also represented a sig-nificant amount of total uptake and it was affectedby sodium. This suggests that in our experimen-tal conditions, the translocation step is a very lim-iting step of the overall process of amino aciduptake. A reduced binding observed in the pres-ence of sodium (Fig. 1) is consistent with the pres-ence of a transport system, which is activated bysodium, as evidenced by the experiments shownin Figures 6 and 7. Whether additional biochemi-cal factors, other than pH and the presence of cat-ions, are involved in transport or not will be theobject of further investigations. The enzyme pro-file of our membrane preparations display enrich-ment factors for membrane markers that aresimilar or even higher than that previously re-ported from whole dipteran larvae (Abduf-Arufand Ellar; 1999; Silva-Filha et al., 1997) or fromwhole lepidopteran larvae (MacIntosh et al., 1994).Results agree well with those of similar protocolsdeveloped for other animal epithelial tissues us-ing the isolated midgut as starting material (Biberet al., 1981; Wolfersberger et al., 1987).
Tracer experiments showed that leucine up-
take into membrane vesicles is greatly affectedby the pH and the ionic composition of the incu-bation buffer. In the presence of the sodium gra-dient, the maximal velocity of leucine uptake atpH 8.8 is 4.7-fold greater than that recorded atpH 7.4. These data correlate with the pH profileof the lumen of C. riparius larvae in vivo. Mea-surements of luminal pH with pH-sensitive dyesrevealed high lumen alkalinity (pH ∼ 10) in theanterior hindgut decreasing to moderate alkalin-ity (pH ∼ 8) in the posterior midgut. Alkaline lu-men pH appears to be widespread among dipteranlarvae (Dow, 1986; Zhuang et al., 1999). Chirono-mids cope with dramatic changes of environmen-tal conditions and tolerate several chemicalstresses, which may influence ions balance andpH values. These larvae seem to possess a trans-port system that is able to respond to variation ofionic composition and pH. Table 3 suggests thatit can be argued that sodium act as an activatorof the amino acid transport by increasing themaximal rate of leucine uptake. This activation,however, occurs only at alkaline pH. The effectseems to be unrelated to the charge of the amino
Fig. 7. Na+ activation of leucine uptake into membranevesicles from C. riparius larvae. Vesicles prepared in 20 mMHepes-Tris, pH 7.4, and 100 mM mannitol were diluted in thesame buffer (closed symbols) or in 20 mM Tris, pH 10, 100
mM mannitol (open symbols). Final pH = 8.8. In all assay mix-tures, osmolarity and ionic strength were held constant by theaddition TMACl. Incubation buffer contained 0.01 mM 3H-leu-cine. Data represent mean ± S.E. of three determinations.

60 Parenti et al.
acid, since optimum pH of transport activity isreached below the pK of the amino group. i.e., whenleucine is still mostly in the zwitterionic form. Toexplain the combined effects of sodium and pH, thepresence of more than one transport system can behypothesized. As shown in Table 4, transport inhi-bition experiments are better interpreted with thepresence of two different transport systems, a so-dium- and pH-dependent one and a sodium- andpH-independent one. As a matter of fact, severalamino acids and analogues, including glycine, lysine,α-methylleucine, leucine methyl ester, α-ketoiso-caproic acid, and 6-aminocaproic acid have a mark-edly different inhibitory effect on leucine transportat the two medium pH values. Based on this con-clusion, we have calculated the Na+-dependent leu-cine transport by subtracting the values in theabsence of Na+ from those in the presence of Na+,as reported in Figure 6. The difference produces anew set of data that fits a hyperbolic saturationcurve having the following kinetic parameters: Vmax
= 55.6 ± 5.6 pmol/7s/mg protein; Km = 25 ± 5 µM.Therefore, at pH 8.8 the Na+-dependent system
doubles its maximal velocity, without changing theaffinity for the substrate. Its catalytic efficiency(Vmax/ Km) is significantly higher than that of theNa+-independent system at the same pH: 2.02 vs.0.35, respectively).
Leucine transport systems in C. riparius lar-vae share the trait of a broad specificity typical ofthe corresponding amino acid transport systemfound in lepidopteran larval midgut (Parenti etal., 2000). This result agrees with a close rela-tionship between the orders of Diptera and Lepi-doptera as proposed by Clark (1999). Transportstudies demonstrate that both midge and cater-pillars have pH-dependent and Na+ gradient-de-pendent amino acid transport systems in theirmidgut epithelia (Sacchi et al., 1994; Henniganand Wolfersberger, 1989). In lepidopteran larvae,however, sodium can not be used as a driver invivo, due to its very low concentration. It was hy-pothesized, however, that this might represent amechanism to ensure sodium intake in an epithe-lium without a sodium pump (Sacchi et al., 1994).Possibly in relation to using leaves as a source offood, caterpillars have evolved the ability of ex-ploiting the K+-electrochemical gradient as a driv-ing force for amino acid accumulation in vivo(Giordana et al., 1982, 1998). Thus, even thougha sound phylogenetic relationship cannot be con-structed on the basis of our limited knowledge oftransport mechanisms in insects, the results pre-sented in this paper may contribute to discrimi-nate between primitive and derived characters.We can speculate that the fully sodium-dependenttransport mechanism represents a plesiomorphiccharacter among neopterans and it is shared bybasal (Parenti et al., 1986) and more evolved(present study) lineages. An apomorphic conditionappears to be the potassium-dependent transportfound in Lepidoptera, whereas the reverse to amore primitive status would be the cation-inde-pendent mechanism found in Coleoptera (Reuveniet al., 1993) and possibly in Hymenoptera (Has-zonits and Crailsheim, 1990). Interestingly, trans-port systems in coleopterans can be stimulatedby monovalent cations, but neither sodium nor po-tassium are able to drive uphill transport of aminoacids (Neal et al., 1996). The variety of foodsources used by Diptera together with their di-versified feeding behavior is a stimulus to increaseour knowledge of nutrient absorption in insects.
TABLE 4. Inhibition of Leucine Uptake by AminoAcids and Analogues*
Inhibitor pH 7.4 pH 8.8
Control 100 100L-leucine 23 20D-leucine 95 97α-methyl-L-leucine 107 45L-leucine methyl ester 96 54Leucinol 98 83α-ketoisocaproic acid 106 554-methylvaleric acid 65 58Glycine 122 40L-alanine 30 37L-valine 28 25L-proline 118 85L-methionine 22 28L-cysteine 37 45L-phenylalanine 71 89L-histidine 65 52L-lysine 93 566-aminocaproic acid 108 55β-alanine 117 96BCH 63 52Acivicin 85 78L-glutamic acid 95 94L-aspartic acid 105 100
*The initial rate of 0.01 mM leucine uptake was measuredat pH 7.4 and at pH 8.8 in the presence of 100 mM NaCland 0.2 mM of the indicated compounds. Numbers repre-sent the residual activity (%) referred to a control in thepresence of 0.2 mM mannitol. BCH=2-amino-2-norbornane-carboxylic acid.

Amino Acid Transport in Midge Larvae 61
ACKNOWLEDGMENTS
We are grateful to Prof. Aurelio Bairati andDr. Mariarosa Gioria for the preparation of theelectron micrograph. This work was supported bygrants from the Italian MURST (ex 60% and, inpart, PRIN, Bioenergetica e trasporto di mem-brana).
LITERATURE CITED
Abduf-Rauf M, Ellar DJ. 1999. Isolation and characteriza-tion of brush border membrane vesicles from wholeAedes aegypti larvae. J Invert Pathol 73:45–51.
Armitage PD, Cranston PS, Pinder LCV, editors. 1995. TheChironomidae. The biology and ecology of non-bitingmidges. London: Chapman & Hall, 572 p.
Berg MB. 1995 Larval food and feeding behaviour. In:Armitage P, Cranston PS, Pinder LCV, editors. TheChironomidae. The biology and ecology of non-bitingmidges. London: Chapman & Hall. p 136–168.
Biber J, Stieger B, Haase W, Murer H. 1981. A high yieldpreparation for rat kidney brush border membranes.Different behaviour of lysosomal markers. BiochemBiophys Acta 647:169–176.
Bradford MM. 1976. A rapid and sensitive method for thequantitation of protein utilizing the principle of dye-protein binding. Anal Biochem 72:248–254.
Castagna M, Shayakul C, Trotti D, Sacchi VF, Harvey WRHediger MA. 1998. Cloning and characterization of apotassium-coupled amino acid transporter. Proc NatlAcad Sci USA 95:5395–5400.
Clark TM. 1999. Evolution and adaptive significance of lar-val midgut alkalinization in the insect superorderMecopterida. Chem Ecol 25:1945–1960.
Clark TM, Koch A, Moffett DF. 1999. The anterior and pos-terior “stomach” regions of larval aedes aegypti mid-gut: regional specialization of ion transport andstimulation by 5.hydroxytryptamine. J Exp Biol202:247–252.
Cross JW, Briggs WR, Dohrman UC, Ray PM. 1978. Auxinreceptors of maize coleoptile membranes do not haveATPase actvity. Plant Physiol 61:581–594.
Dadd RH. 1975. Alkalinity within the midgut of mosquitolarvae with alkaline-active digestive enzymes. J In-sect Physiol 21:1847–1853.
Dow JAT. 1986. Insect midgut function. Adv Insect Physiol19:187–328.
Giordana B, Sacchi VF, Hanozet GM. 1982. Intestinal aminoacid absorption in lepidopteran larvae. BiochemBiophys Acta 692:81–88.
Giordana B, Leonardi MG, Casartelli M, Consonni P, ParentiP. 1998. K+-neutral amino acid symport of Bombyxmori larval midgut: a system operative in extreme con-ditions. Am J Physiol 274:R1361–R1371.
Hanozet GM, Giordana B, Sacchi VF. 1980. K+-dependentphenylalanine uptake in membrane vesicles isolatedfrom the midgut of Philosamia cynthia larvae. BiochimBiophys Acta 596:481–486.
Harvey WR, Nedergaard S. 1964. Sodium-independent activetransport of potassium in the isolated midgut of cecro-pia silkworm. Proc Natl Acad Sci USA 51:481–846.
Haszonits O., Crailsheim K. 1990. Uptale of L-leucine intoisolated enterocytes of the honeybee (Apis melliferaL.) depending on season. J Insect Physiol 36:835–842.
Hennigan BB, Wolfersberger MG. 1989. Intestinal aminoacid absorption in tobacco hornworm larvae is stimu-lated by potassium and sodium but not rubidium orlithium. Arch Insect Biochem Physiol 11:21–28.
Hennigan BB, Wolfersberger MG, Harvey WR. 1993. Neu-tral amino acid symport in larval Manduca sextamidgut brush border membrane vesicles deducedcation-dependent uptake of leucine, alanine and phe-nylalanine. Biochim Biophys Acta 1148:216–222.
MacIntosh SC, Lidster BD, Kirkham CL. 1994. Isolationof brush border membrane vesicles from whole dia-mondback moth (Lepidoptera: Plutellidae) larvae. JInvert Pathol 63:97–98.
Neal, JJ, Wu D, Hong YS, Reuveni M. 1996. High affinitytransport of histidine and methionine across Laptin-otarsa decemlineata midgut brush border membrane.J insect Physiol 42:328–325.
Parenti P, Sacchi FV, Hanozet GM, Giordana B. 1986. Na-dependent uptake of phenylalanine in the midgutof a cockroach Blabera gigantea. J Comp Physiol B156:549–556.
Parenti P Villa M, Hanozet GM. 1992. Kinetics of leucinetransport in brush border membrane vesicles fromLepidopteran Larvae midgut. J Biol Chem 267:15391–15397.
Parenti P, Forcella M, Pugliese A, Casartelli M, GiordanaB, Leonardi MG, Hanozet GM. 2000. Substrate speci-ficity of the brush border K+-leucine symport ofBombyx mori larval midgut. Insect Biochem Mol Biol30:243–252.
Parthasarathy R, Xie T, Wolfersberger MG, Harvey WR.1994.Substrate structure and amino acid/K+ symport inbrush-border membrane vesicles from larval Manducasexta midgut. J Exp Biol 197:237–250.
Quigley JP, Gotterer GS. 1969. Properties of a high specificactivity, (Na+-K+)-stimulated ATPase from rat intesti-nal mucosa. Biochim Biophys Acta 173:456–468.

62 Parenti et al.
Reuveni M, Dunn PE. 1994. Proline transport into brushborder membrane vesicles from the midgut of Manducasexta larvae. Comp Biochem Physiol 107A:685–691.
Reuveni M, Hong YS, Dunn PE, Neal JJ. Leucine transportinto brush border membrane vesicles from guts ofLeptinotarsa decemlineata and Manduca sexta. CompBiochem Physiol 104A:267–272.
Sacchi VF, Parenti P, Perego C, Giordana B. 1994. Interac-tion between Na+ and the K+-dependent amino acidtransport in midgut brush border membrane vesiclesfrom Philosamia cynthia larvae. J Insect Physiol 40:69–74.
Seidman LA, Bergtrom G, Remsen CC. 1984. Structure ofthe larval midgut of the fly Chironomus thummi andits relationship to sites of cadmium sequestration. Tis-sue Cell 18:407–418.
Silva CP, Terra WR. 1994. Digestive and absorptive sitesalong the midgut of the cotton seed sucker bugDysdicercus peruvianus (Hemiptera: Pyrrhocoridae).Insect Biochem Mol Biol 24:493–505.
Silva-Filha, MH, Nielsen-Leroux C, Charles J-F. 1997. Bind-ing kinetics of Bacillus sphaericus binary toxin to mid-gut brush-border membranes of Anopheles and Culexsp. mosquito larvae. Eur J Biochem 247:754–761.
Smith L. 1955. Spectrophotometric assay of cytochrome coxidase. In: Glick T, editor. Methods in biochemicalanalysis. New York: Interscience. 2:427–434.
Terra WR, Ferreira C. 1994. Insect digestive enzymes: prop-
erties, compartmentalization and function. Comp Bio-chem Physiol 109B:1–62.
Terra WR, Ferreira C, DeBianchi AG. 1979. Distributionof digestive enzymes among the endo- and ectoperi-throphic spaces and midgut cells of Rhynchosciaraand its physiological significance. J Insect Physiol25:487–494.
Wieczorek, H. Weerth S, Schindlbeck M, Klein U. 1989. Avacuolar-type proton pump in a vesicles fraction en-riched with potassium transporting plasma mem-branes from tobacco hornworm midgut. J Biol Chem264:11143–11148.
Wolfersberger MG. 2000. Amino acid transport in Insects.Ann Rev Entomol 45:111–120.
Wolfersberger MG, Luethy P, Maurer A, Parenti P, SacchiFV, Giordana B, Hanozet GM. 1987. Preparation andpartial characterization of amino acid transportingbrush border membrane vesicles from the larval mid-gut of the cabbage butterfly Pieris brassicae. CompBiochem Physiol 86A:301– 308.
Wright DA. 1975. The raltionship between transepithelialsodium movement and potential difference in the larvaof Camptochironomus tentans (Fabr.) and some obser-vations on the accumulation of other ions. J Exp Biol62:157–174.
Zhuang ZP, Linser PJ, Harvey WR. 1999. Antibody to H+ V-ATPase subunit E colocalizes with portasomes in al-kaline larval midgut of a freshwater mosquito (Aedesaegypti L.). J Exp Biol 202:2449–2460.




















