
Wing venation and Distal-less expression in Heliconius butterfly wing pattern development

lable at ScienceDirect
Arthropod Structure & Development 44 (2015) 1e9
Contents lists avai
Arthropod Structure & Development
journal homepage: www.elsevier .com/locate/asd
Lehr's fields of campaniform sensilla in beetles (Coleoptera):Functional morphology. II. Wing reduction and the sensory field
Leonid Frantsevich a, *, Stanislav Gorb b, Vladimir Radchenko c, Dmytro Gladun c,Alexey Polilov d, Lyubov Cherney e, Vassily Browdy f, Maxim Kovalev g
a Department of Ethology and Social Biology of Insects, Schmalhausen-Institute of Zoology, B. Chmielnicki Street 15, 01601 Kiev, Ukraineb Department of Functional Morphology and Biomechanics, Christian-Albrechts-University of Kiel, Am Botanischen Garten 1-9, D-24098 Kiel, Germanyc Institute for Evolutionary Ecology, Acad. Lebedev Street 37, 03143 Kiev, Ukrained Department of Entomology, Biological Faculty, Lomonosov Moscow State University, 1-12 Leninskie Gory, 119991 Moscow, Russiae Department of Scientific Fundamental Collection, Schmalhausen-Institute of Zoology, B. Chmielnicki Street 15, 01601 Kiev, Ukrainef Department of Zoology, Dragomanov Pedagogical University, Pirogov Street 9, 01601 Kiev, Ukraineg Department of Zoology, Institute of Biology, National Shevchenko-University, Volodymirska Street, 64, 01601 Kiev, Ukraine
a r t i c l e i n f o
Article history:Received 14 February 2014Received in revised form22 October 2014Accepted 24 October 2014Available online 1 November 2014
Keywords:Insect flightColeopteraCampaniform sensillaWing reductionElytra
* Corresponding author:E-mail address: [email protected] (L. Frantsevic
http://dx.doi.org/10.1016/j.asd.2014.10.0031467-8039/© 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
Loss of the flight ability and wing reduction has been reported for many taxa of Coleoptera. If elytra areclosed, their roots are clenched between the tergum and the pleuron, forces applied to the elytra can notbe transmitted to the field of campaniform sensilla situated on the root. That is why it is plausible toassume that the field becomes redundant in non-flying beetles. We examined the relationships betweenthe hind wing reduction and characters of this mechanosensory field in beetles of six families. Wemeasured the size of the elytron, that of the hind wing and counted the number of sensilla in the sensoryfield. Mesopterous non-flying beetles retain one half to one third of sensilla present in macropterousspecies of the same body size. Further reduction of the sensory field in brachypterous species is obvious,but sensilla are still present in insects with strongly reduced wings, as long as their elytra are separableand mesothoracic axillaries are present. Complete loss of sensilla coincides with the existence of apermanent sutural lock. However, some beetles with permanently locked elytra and absence of axillariesstill retain few campaniform sensilla. A very special case of an extreme wing modification in feather-wingbeetles is considered. No sensilla were revealed either on the root of the elytron or on the basal segmentof such fringed wings in flying ptiliid species.
© 2014 Elsevier Ltd. All rights reserved.
1. Introduction
The present paper is the second of three articles on the structureand size of the field of campaniform sensilla (CFS) situated on theroot of the elytron in the elytron-to-body articulation. The field ofCFS has been initially described by Lehr (1914), this is why wedesignate this sensory field as Lehr's field. Our first paper treatedthe morphology of the root and composition of Lehr's field, as wellas allometric relationships between the size of the elytron and thenumber of sensilla within this field in flying beetles (Frantsevichet al., 2014). The present paper considers the correlation betweenthe grade of hind wing reduction in non-flying beetles and thereduction in number of sensilla within the field.
h).
Loss of flight ability and wing reduction was often reportedamong Coleoptera. Elytra in non-flying beetles are permanentlyclosed. The root is tightly clenched between the tergum and pleu-ron and thus obtains only a negligible portion of forces acting onthe elytra. In such a situation, the non-signaling Lehr's sensory fieldbecomes redundant. Functionality of Lehr's field in non-flyingbeetles may be potentially preserved only in such a rare case,when the elytra still are able to separate, open and close themselvesactively, not for the function of flight but rather for righting. Whathas happened with Lehr's field in the evolution of non-flying bee-tles, which gradually lost their flight ability? One can hypothesizedifferent degrees of the reduction of the field depending on thedegree of wing reduction.
The early stage of loss of the flight ability appears as underde-velopment of direct wing muscles during metamorphosis ordegeneration of those muscles in the second half of the imaginallife. The wings themselves and the external appearance of the

Nomenclature
CFS campaniform sensillumP0 probability of the null hypothesisRWE ratio between the wing and elytral lengthSEM scanning electron microscopeTLM transmitting light microscopy
L. Frantsevich et al. / Arthropod Structure & Development 44 (2015) 1e92
beetle remain usually unaffected at this stage. Further stages of thereduction are (1) degeneration of direct and indirect wing muscles(Tietze, 1963; Davydova, 1966; Hammond, 1985), (2) shortening ofwings, and (3) loss of vein sclerotization up to scale-like residues,which sometimes are difficult to recognize. Reduction of wingstakes place in parallel to the reduction of the metanotum and to thereduction of wing nerves together with muscles (Rüschkampf,1927), as well as to the diminution of metathoracic and the firstabdominal spiracles. Further details on such events may be ob-tained in articles listed below according to the order of beetlefamilies:
Dytiscidae (Leech, 1942; Jackson, 1956);Carabidae (Schulze, 1912; Oertel, 1924; Darlington, 1936; Tietze,1963; Smith, 1964);Ptiliidae (Kolbe, 1920; Dybas, 1978; Taylor, 1981);Silphidae (Parkhomenko, 2002);Staphylinidae (Blum, 1979);Geotrupidae (Geotrupes: Suzuki et al., 2001; Lethrus: Nikolaev,1987);Scarabaeidae (Duncan, 2002);Lampyridae (Davydova, 1966);Coccinellidae (Hammond, 1985);Tenebrionidae (Bouchard and Gorb, 2000; Cherney, 2005);Meloidae (Melo€e proscarabaeus: Linnaeus, 1758, p. 419);Cerambycidae (Plavilschikov, 1936, 1958);Chrysomelidae (Kolbe, 1920; Rüschkampf, 1927; Brovdiy, 1973;Furth, 1980; Samuelson, 1996);Curculionidae (Jackson, 1928; Nazarenko, 1996).
Wing reduction may be a stable character of only females, e.g. infireflies (Lampyridae) (Davydova, 1966) and in the desert long-hornbeetle Prionus komarovi (Cerambycidae) (Plavilschikov, 1936).However, in most cases, the reduction affects both sexes equallyand is polymorphic: macropterous and even well flying specimenscoexist together with brachypterous ones. Sometimes strictdimorphism with only two morphs is caused by experimentallyproven Mendelian mutation (Jackson, 1928; Darlington, 1936;Leech, 1942; Lindroth, 1946 cited by Smith, 1964; Taylor, 1981;Hammond, 1985), but many transitional morphs have beenobserved among more or less brachypterous ones in other species(Rüschkampf, 1927).
The reduction process involves elytra as well. Their protectivefunction is even reinforced by the strengthening of the linkagebetween the elytra and body and between the elytra themselves.Many elytral structures are also affected.
(1) The anterolateral corner of the elytron becomes roundinstead of rectangular, a hillock on this corner, enclosing thewing base, disappears.
(2) The epipleuron becomes broad.(3) Mesothoracic axillary plates become small or entirely
disappear.(4) Frictional sutural lock between the elytra, functioning ac-
cording to the plug-and-socket principle is transformed into
the permanent lock with the snapping plug element. Plugand socket elements in tenebrionids may even be doubled ortripled (Fiori, 1975).
(5) Co-opted fields of microtrichial fasteners on the elytra and onthe body, which passively link closed elytra with the thoraxor the abdomen in flying beetles, remain in severalbrachypterous and apterous leaf beetles (Samuelson, 1996),as well as in the non-flying tenebrionid Tabarus montanus(Bouchard and Gorb, 2000).
(6) Elytra become shorter in the females of P. komarovi and of thefire-fly Luciola mingrelica or disappear completely in thelarviform females of the fire-fly Lampyris noctiluca, whereasin L. orientalis female elytra are still present in the form ofsmall triangular plates.
(7)The size of mesothoracic flight muscles in Ptinidae decreases,but muscles do not disappear completely (Smith, 1964).
Does Lehr's field of CFS follow these morphological changescaused by thewing reduction? Our approach to this problem differsfrom the one used in the previous study on CFS allometry(Frantsevich et al., 2014). In the present paper, we again selected aseries of taxonomically related beetles, but now specimens ofapproximately the same size were chosen, in order to minimize thescaling effect. Partial or complete reduction of Lehr's field wasfound in all flightless beetles studied. However, the analysis of theintraspecific polymorphism of these structures was not the subjectof the present paper.
2. Methods
Morphological methods, microscopy techniques, and quantita-tive analysis were essentially the same as previously used(Frantsevich et al., 2014), with two additions.
Wings were spread in a drop of 96� ethyl alcohol on a glass slidewith the aid of a fine needle. After evaporation of the alcohol, thespread wing kept its shape even in large beetles. Wings werephotographed against a blue background, in order to enhancecontrast of brown sclerotized parts. Their lengths were measuredfrom the images and compared with the length of correspondingelytra. Foldable wings of flying beetles are longer than their elytra:those wings are called macropterous. Strongly reduced short wingrudiments are called brachypterous. Intermediate ones, which arealmost of the same size as the elytra, are called mesopterous.
For inspection of sutural locks, we used the following method:elytra were macerated in 10% KOH, perisutural stripes were excisedin a wax tray in a drop of ethyl alcohol. Stripes were placed intoanother tray with smooth wax; a bit of melted wax was drippedonto the stripe. The stripe was cut with a razor blade into slicesabout 120e150 mm thick. Cross-sections were put into a drop ofglycerol and photographed in transmission light microscope (TLM).
3. Results
57 beetle species of 12 families have been inspected for thisarticle, 34 species were non-flying. Their taxonomic position andnomenclature are listed in Table S1, whereas quantitative dataconcerning processed materials are shown as Table S5, both inSupplementary Materials.
3.1. Wing reduction in diving beetles
Flight in Dytiscidae is used for migration from onewater body toanother one (Bowker, 2002; Miguelez and Valladares, 2008).Dytiscus marginalis was capable of continuous flight for a distanceof up to 25 km in a wind tunnel (Schneider, 1986). However, few

L. Frantsevich et al. / Arthropod Structure & Development 44 (2015) 1e9 3
flightless species with reduced wings do exist among diving bee-tles. Fig. S2 in Supplementary Materials compares three relatedspecies from the subfamily Agabinae (nomenclature by Zaitsev,1953): the flying macropterous Gaurodytes bipustulatus, non-flying macropterous Eriglenus undulatus (Jackson, 1956) and non-flying brachypterous Gaurodytes bifarius (kessleri auct.). It is easyto split elytra manually in all three species.
In Fig. 1, the number of CFS in Lehr's field is plotted versus theratio of the hind wing length to the elytron length. The ratio be-tween the elytral areas in two macropterous species is two-fold,hence, according to allometric relationships between the size ofthe elytron and the number of CFS in flying beetles (Frantsevichet al., 2014), one would expect 40 � √2 ¼ 28 CFS in E. undulatus,but this non-flying species has only 19 CFS. E. undulatus and G.bifarius are of the same body and elytral size, but the brachypterousG. bifarius bears only 11 CFS. Correlation between the relative winglength and the number of CFS is 0.667, non-significant at the singledegree of freedom (Fig. 1 left panel).
3.2. Wing reduction in ground beetles
For comparison, we selected representatives of two closely-related genera, Calosoma and Carabus, which are of similar bodysize and comprise a series of species from having fully operatingwings to those with strong rudimentation.
Calosoma sycophanta is able to fly, but some specimens do notfly (the initial stage of wing reduction) and have reduced apodemesof both basalare and subalare (Tietze, 1963). Cal. inquisitor ismacropterous, but incapable of flight (“fliegt überhaupt nicht mehr”,Oertel, 1924). Our Carabus sample is represented by five non-flyingspecies e from mesopterous to utmost brachypterous ones (Figs. 1right panel and 2). Wings in flying, or at least macropterous Calo-soma, are by half longer than the elytra. The mesopterous wing inCarabus granulatus is as long as the elytron, other species showrudiments from 50 to 5 % of the elytral length.
The number of CFS is rather modest in macropterous groundbeetles of medium size and the elytral area of 75e120 mm2 (28e30CFS in three inspected Calosoma including Cal. chinense), ifcompared with numbers over 50e100 CFS in many, even smallerflying dytiscids, lucanids, melolontines, coccinelids, chrysomelidsetc. (Frantsevich et al., 2014). Correlation between the relative winglength and the number of CFS is 0.974, P0< 0.1%. Wing reduction inCarabus renders only partial loss of sensilla even in species withresidual scales instead of wings Car. granulatus retains axillaryplates, they are lost from the smallest members of the series (Car.
Fig. 1. Decrease of the number of CFS in Lehr's field with the loss of ability to fly and wingundulatus, (c) G. bifarius. Right panel e Carabidae: (a) Calosoma sycophanta, (b) Cal. inquisitocoriaceus. Abbreviations: N e number of CFS, RWE e ratio between the wing length and elytrflying beetles, hollow circles e non-flying beetles.
hungaricus, Car. coriaceus), the elytra of which are fused with therim of the mesotergum.
We inspected several non-flying ground beetles, including twomedium-sized specimens of Pterostichus melanarius and one smallP. strenuus. The latter onewasmacropterous, a rare event noticed byLindroth (1949; cited in Smith, 1964). The relative wing lengthcomprised 30e60% and 130% respectively, and the number of CFSe5e16 and 17, the latter number is rather high, when we take intoaccount that the elytral area in P. strenuuswas 6e8 times smaller, ifcompared to specimens of the former species.
Macropterous non-flying Zabrus tenebrioides of medium bodysize retains two CFS. Both medium-sized Cychrus aeneus and largeAnthia mannerheimi with tiny scales instead of wings exhibit notrace of CFS. A field of rough bristles on the antero-lateral rim of themesonotum, seen in A. mannerheimi, which was previously sug-gested to be a sensor of vertical positioning of the elytron in Calo-soma (Schneider, 1978), was present in the same shape in allinspected species of Carabus.
3.3. Wing modification in featherwing beetles
Ptiliidae belong to most miniature insects (Polilov, 2008; Polilovand Beutel, 2009). In flying species, wing modification ismanifested by the reduction of the membranous part of the wingand by the substitution of the latter with a fan of setae. In a 0.5 mmlong specimen, the elytron was 0.35 mm long (Fig. 3A). The lengthand width of the root in the dorsal aspect comprised 10 by 7 mm(Fig. 3C). Thorough inspection of the root of the elytron in Primor-skiella anodonta at high magnification in all possible aspects(Fig. 3CeF) did not reveal any trace of CFS.
The wing, about 1 mm long, consists of a stem equipped withabout fifty long primary setae which in turn bear small secondarysetulae, analogs of barbs and barbicels in a bird's feather, but verysparse. Setae form a round fan, a sort of sievewhich is empty at 90%of the fan's outline area. The stem is divided into a few serial seg-ments. In the resting state, the wing is folded four times under theelytron (Fig. 3B), all setae are passively bent to their segments. Thebasal segment of the wing also contains no trace of CFS, only fric-tional microstructures.
3.4. Wing reduction in carrion beetles
Predaceous and necrophagous Silphidae, searching fordispersed food sources, are usually able to fly, whereas wingreduction appears in phytophagous and saprophagous beetles
reduction. Left panel e Agabus (Dytiscidae): (a) Gaurodytes bipustulatus, (b) Eriglenusr, (c) Carabus granulatus, (d) Car. arvensis, (e) Car. cancellatus, (f) Car. hungaricus, (g) Car.on length, in %. Data were measured on single representative specimens. Filled circles e

Fig. 2. Elytron, wings or wing rudiments (top panel) and Lehr's fields on the elytra (bottom panel, SEM) in Calosoma and Carabus of comparable body and elytral size: (a) Cal.sycophanta, (c) Car. granulatus, (d) Car. arvensis, (e) Car. cancellatus, (f) Car. hungaricus, (g) Car. coriaceus. Scale bars 10 mm (top panel), 50 mm (bottom panel).
L. Frantsevich et al. / Arthropod Structure & Development 44 (2015) 1e94
relying on less fragmented locations of food sources (Parkhomenko,2002). We selected three medium body size species for each of twotypes of foraging within the subfamily Silphinae (Fig. S3 in Sup-plementary Materials). Flying species have wings substantially
Fig. 3. Fore and hind wings in a miniature beetle Primorskiella anodonta (Ptiliidae). (A) elytro(C), ventral face (D), medial face (E), lateral face (F). No pits or slits were revealed on a sm
longer than their elytra, whereas flightless species exhibited partial(mesopterous) wing reduction: the wing length comprised over50% of the length of the elytron. Despite some variability of thebody size, the number of CFS inmesopterous species was about two
n; (B) trunk with folded pinnate wings; (CeF) elytral root in various aspects: dorsal faceooth surface of the elytral root. Scale bars 100 mm (A, B), 5 mm (CeE), 5 mm (F).
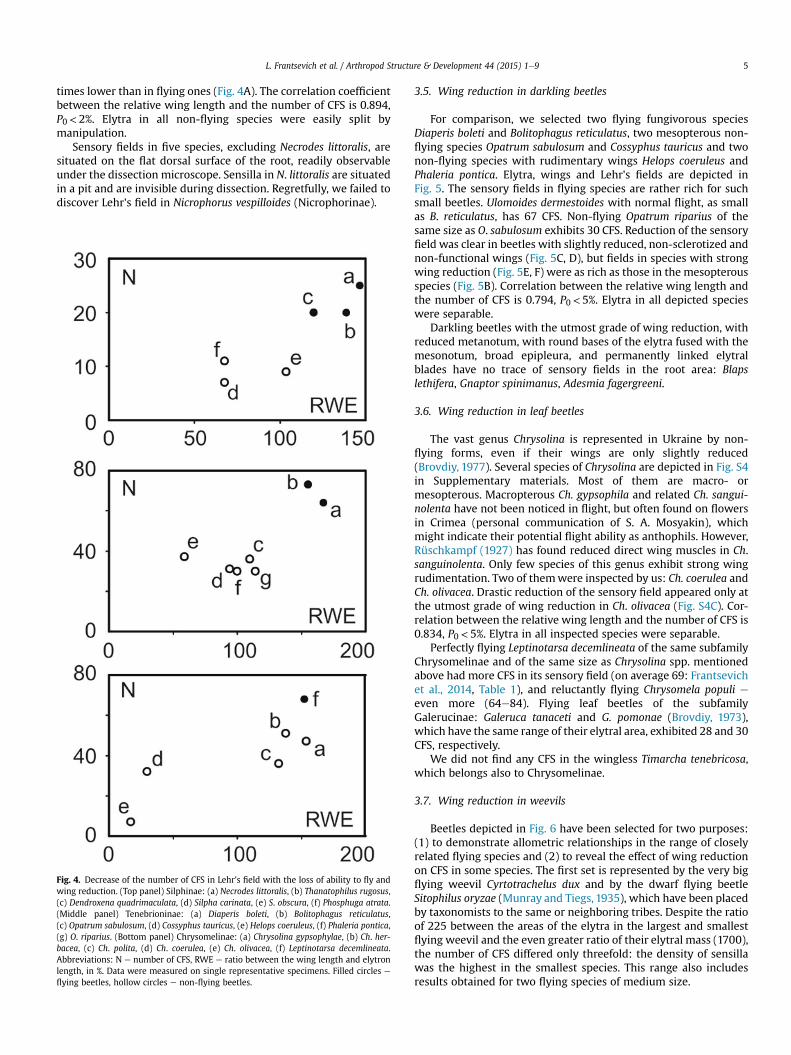
L. Frantsevich et al. / Arthropod Structure & Development 44 (2015) 1e9 5
times lower than in flying ones (Fig. 4A). The correlation coefficientbetween the relative wing length and the number of CFS is 0.894,P0< 2%. Elytra in all non-flying species were easily split bymanipulation.
Sensory fields in five species, excluding Necrodes littoralis, aresituated on the flat dorsal surface of the root, readily observableunder the dissection microscope. Sensilla in N. littoralis are situatedin a pit and are invisible during dissection. Regretfully, we failed todiscover Lehr's field in Nicrophorus vespilloides (Nicrophorinae).
Fig. 4. Decrease of the number of CFS in Lehr's field with the loss of ability to fly andwing reduction. (Top panel) Silphinae: (a) Necrodes littoralis, (b) Thanatophilus rugosus,(c) Dendroxena quadrimaculata, (d) Silpha carinata, (e) S. obscura, (f) Phosphuga atrata.(Middle panel) Tenebrioninae: (a) Diaperis boleti, (b) Bolitophagus reticulatus,(c) Opatrum sabulosum, (d) Cossyphus tauricus, (e) Helops coeruleus, (f) Phaleria pontica,(g) O. riparius. (Bottom panel) Chrysomelinae: (a) Chrysolina gypsophylae, (b) Ch. her-bacea, (c) Ch. polita, (d) Ch. coerulea, (e) Ch. olivacea, (f) Leptinotarsa decemlineata.Abbreviations: N e number of CFS, RWE e ratio between the wing length and elytronlength, in %. Data were measured on single representative specimens. Filled circles e
flying beetles, hollow circles e non-flying beetles.
3.5. Wing reduction in darkling beetles
For comparison, we selected two flying fungivorous speciesDiaperis boleti and Bolitophagus reticulatus, two mesopterous non-flying species Opatrum sabulosum and Cossyphus tauricus and twonon-flying species with rudimentary wings Helops coeruleus andPhaleria pontica. Elytra, wings and Lehr's fields are depicted inFig. 5. The sensory fields in flying species are rather rich for suchsmall beetles. Ulomoides dermestoides with normal flight, as smallas B. reticulatus, has 67 CFS. Non-flying Opatrum riparius of thesame size as O. sabulosum exhibits 30 CFS. Reduction of the sensoryfield was clear in beetles with slightly reduced, non-sclerotized andnon-functional wings (Fig. 5C, D), but fields in species with strongwing reduction (Fig. 5E, F) were as rich as those in the mesopterousspecies (Fig. 5B). Correlation between the relative wing length andthe number of CFS is 0.794, P0< 5%. Elytra in all depicted specieswere separable.
Darkling beetles with the utmost grade of wing reduction, withreduced metanotum, with round bases of the elytra fused with themesonotum, broad epipleura, and permanently linked elytralblades have no trace of sensory fields in the root area: Blapslethifera, Gnaptor spinimanus, Adesmia fagergreeni.
3.6. Wing reduction in leaf beetles
The vast genus Chrysolina is represented in Ukraine by non-flying forms, even if their wings are only slightly reduced(Brovdiy, 1977). Several species of Chrysolina are depicted in Fig. S4in Supplementary materials. Most of them are macro- ormesopterous. Macropterous Ch. gypsophila and related Ch. sangui-nolenta have not been noticed in flight, but often found on flowersin Crimea (personal communication of S. A. Mosyakin), whichmight indicate their potential flight ability as anthophils. However,Rüschkampf (1927) has found reduced direct wing muscles in Ch.sanguinolenta. Only few species of this genus exhibit strong wingrudimentation. Two of themwere inspected by us: Ch. coerulea andCh. olivacea. Drastic reduction of the sensory field appeared only atthe utmost grade of wing reduction in Ch. olivacea (Fig. S4C). Cor-relation between the relative wing length and the number of CFS is0.834, P0< 5%. Elytra in all inspected species were separable.
Perfectly flying Leptinotarsa decemlineata of the same subfamilyChrysomelinae and of the same size as Chrysolina spp. mentionedabove had more CFS in its sensory field (on average 69: Frantsevichet al., 2014, Table 1), and reluctantly flying Chrysomela populi eeven more (64e84). Flying leaf beetles of the subfamilyGalerucinae: Galeruca tanaceti and G. pomonae (Brovdiy, 1973),which have the same range of their elytral area, exhibited 28 and 30CFS, respectively.
We did not find any CFS in the wingless Timarcha tenebricosa,which belongs also to Chrysomelinae.
3.7. Wing reduction in weevils
Beetles depicted in Fig. 6 have been selected for two purposes:(1) to demonstrate allometric relationships in the range of closelyrelated flying species and (2) to reveal the effect of wing reductionon CFS in some species. The first set is represented by the very bigflying weevil Cyrtotrachelus dux and by the dwarf flying beetleSitophilus oryzae (Munray and Tiegs, 1935), which have been placedby taxonomists to the same or neighboring tribes. Despite the ratioof 225 between the areas of the elytra in the largest and smallestflying weevil and the even greater ratio of their elytral mass (1700),the number of CFS differed only threefold: the density of sensillawas the highest in the smallest species. This range also includesresults obtained for two flying species of medium size.

Fig. 5. Elytra, wings and elytral Lehr's fields in darkling beetles of the subfamily Tenebrioninae: flying macropterous Diaperis boleti, 64 CFS (A) and Bolitophagus reticulatus, 73 CFS(B); non-flying mesopterous Opatrum sabulosum, 36 CFS (C) and Cossyphus tauricus, 31 CFS (D); brachypterous Helops coeruleus, 37 CFS (E) and Phaleria pontica, 30 CFS (F). Scale bars5 mm for macrophotographs, 20 mm for TLM microphotographs.
L. Frantsevich et al. / Arthropod Structure & Development 44 (2015) 1e96
The second set is represented by two species with rudimentarywings: S. granarius as small as S. oryzae and the medium sizedLiparus glabrirostris (Nazarenko, 1996). S. granarius exhibits aston-ishingly moderate loss in CFS number, taking into account the
Table 1Types of the sutural lock in non-flying beetles.
Species Type of the wing Type of the lock N CFS
Anthia mannerheimi Brachypterous Permanent 0Calosoma inquisitor Macropterous Split 28Calosoma sycophanta Flying Split 30Carabus arvensis Mesopteous Split 7Carabus cancellatus Brachypterous Split 4Carabus coriaceus Brachypterous Permanent 5Carabus granulatus Macropterous Permanent 16Carabus hungaricus Brachypterous Permanent 3Cychrus aeneus Apterous Permanent 0Zabrus tenebrioides Macropterous Permanent 2Lethrus apterus Apterous Permanent 0Adesmia fagergreeni Apterous Permanent 0Blaps lethifera Apterous Permanent 0Gnaptor spinimanus Apterous Permanent 0Melo€e proscarabaeus Apterous Absent 0Dorcadion fulvum Apterous Split 0Timarcha tenebricosa Apterous Permanent 0Liparus glabrirostris Brachypterous Split 25
bigger size of the elytra in comparison to S. oryzae. Elytra in S.granariuswere easily separable. Despite strong wing rudimentation(Nazarenko, 1996) and accompanying reduction of the metanotum,as well as round shape of the elytra in L. glabrirostris, this weevil hasa considerable number of CFS. It also retained flattened mesotho-racic axillaries.
3.8. Wing reduction in other groups
There are several genera with completely reduced wings amongfamilies with predominantly flying beetles. We inspected Lethrusapterus in Geotrupidae, Dorcadion fulvum in Cerambycidae, Melo€eproscarabaeus in Meloidae. We did not find the CFS in these species.
3.9. Linkage between closed elytra in non-flying beetles
Closed elytra are locked together along the sutura by a frictionalor snap lock built according to the plug-and-socket principle: theventral sutural ridge of one elytron enters into the space betweenthe dorsal and ventral ridges of the opposite elytron. Cross-sectionsof several sutural edges, paired or single, are depicted in Fig. 7. If aplug is narrow enough, the lock is able to split, which is the case inall flying beetles. Such split pair for the macropterous Calosoma

Fig. 6. Campaniform sensilla in Lehr's field in weevils (Dryophthorinae): (A) Cyrtotrachelus dux, >70 CFS, (B) Cleonus achates, 44 CFS, (C) Hylobius abietis, 64 CFS, (D) Sitophilus oryzae,26 CFS, (E) S. granarius, 20 CFS, (F) Liparus glabrirostris, 25 CFS. Outlines of elytra are inserted in SEM photographs in their natural size (AeC, F) or magnified 5:1 (D, E); black bars in(D, E) show the natural length of small elytra. Wings in (E, F) are strongly reduced, though CFS are still present. Horizontal scale bar 10 mm in (C) is common to all SEM photographs,vertical scale bar 10 mm in (C) is common to all outlines of the elytra.
L. Frantsevich et al. / Arthropod Structure & Development 44 (2015) 1e9 7
inquisitor is illustrated in Fig. 7a. On the contrary, if a plug elementis swollen like a fist or bulb, it fits into the round space betweenridges of the opposite elytra and is snapped there forever. Thepermanent lock fixing elytra in thewingless Blaps lethifera is shownin Fig. 7d. This figure illustrates also splitted or snapped profiles inseveral other non-flying beetles.
We inspected the construction of the sutura in 17 species ofnon-flying beetles, plus flying Cal. sycophanta for comparison. Dataon the type of the lock, the state of the hind wing, and the numberof CFS are pooled together in Table 1. Permanent locks are com-bined mostly with brachypterous or apterous wing types, whereassplit locks occur not only in combinations with macropterous (aswell as with flying) wings, but even with strongly reduced wings.Elytra with narrow plug elements in Dorcadion fulvum and Liparusglabrirostris belong morphologically to the split type, but it is ratherdifficult to split them manually during dissection. Elytra in Melo€eproscarabaeus are diverging sidewards and do not form any sutura.
Fig. 7. Temporal and permanent sutural locks in non-flying beetles: cross-sections ofsutural ridges. (a) Plug and socket split pair in macropterous Calosoma inquisitor;(b) split ridges in brachypterous Carabus cancellatus; (c) split ridges in brachypterousLiparus glabrirostris; (d) plug and socket permanent pair in wingless Blaps lethifera;(e) bulbous permanent plug in macropterous Car. granulatus; (f) same in winglessTimarcha tenebricosa; (g) same in wingless Lethrus apterus. Not to scale.
4. Discussion
4.1. Reduction of wings and transformation of the pretothorax
Short wings per se do not indicate flightlessness of a beetle:perfectly flying jewel beetles (Buprestidae) have wings which areshorter than the elytra and devoid of transverse folds (Fedorenko,2004). But such wings are rare among beetles, most flying formshave foldable macropterous wings which are much longer thancorresponding elytra. The initial step of wing reduction directlyaffects only wingmuscles during the entire or the second half of theimaginal lifetime, whereas wings presumably remain intact.
A “refuse” to invest resources into the potent hind wingmusculature rendered unnecessary also hind wings themselves, aswell as vast sclerotized surfaces for support of indirect wing mus-cles. Loss of the flight ability and respective mophological changesin the metathorax originated independently in diverse primarilyflying insect taxa (see references in Section 1). Correspondingtransformations of elytra also occur in flightless beetles of diversetaxa.
Transformations in themetathorax of flightless beetles emergednot abruptly, but rather continuously with intermediate stages re-ported in different taxonomically related recent species. The sameholds true with respect to transformations of the elytra, theirarticulation, locks, elytral muscles and sensory organs. Grades ofreduction may be different in different organs.
Even reduced, hind wings are not devoid of some functionalimportance. Jackson (1956) noticed that a sclerotized frictionalplate in the cubital area of the wing, used for attachment betweenthe folded wing and the elytron (catastigma, after Heberdey, 1938),is still present in its full size in the strongly reduced wing ofGaurodites bifarius. Indeed, dark-brown catastigmae are seen on thewings of all three species of flying or not flying dytiscids (Fig. S2 inSupplementary materials).
The loss of the flying ability, as long as elytra are able to split,does not exclude active movements of the elytra. For example,some short-legged flying beetles open their elytra for righting:elevating elytra (with respect to body-fixed reference) and usethem to push the posterior part of the overturned body upwards,the anterior part downwards. Thus, during this action, fore andmiddle legs have a better chance to grasp the ground. The over-head roll was described by Nachtigall (1961) in Acilius sulcatus(Adephaga, Dytiscidae), by Frantsevich (2004, Fig. 8) in a leafbeetle Cassida nebulosa (Polyphaga, Chrysomelidae). One of theauthors (L.F.) also observed righting using the same mechanism in

L. Frantsevich et al. / Arthropod Structure & Development 44 (2015) 1e98
lady birds Coccinella septempunctata, Harmonia axyridis (Poly-phaga, Coccinellidae) and in the leaf beetle Galeruca tanaceti(unpublished data). Much earlier, this righting behavior wasdescribed by Rüschkampf (1927) in leaf beetles Chrysomela (Mel-asoma auct.) populi, Ch. tremulae and even in non-flying Chrysolinaspp. This residual locomotory function is hardly probable for themajority of non-flying beetles and can be excluded for all beetleswith permanent elytral locks.
4.2. Reduction of CFS in flightless beetles
The impact of the hind wing reduction on the presence anddistribution of hind wing mechanoreceptors, including fields ofCFS on wing veins, still remains unclear. There are also no reportsin the literature concerning other sensory organs, except suchconspicuous ones as compound eyes. The effect of wing reductionon the morphology of compound eyes was noticed in dwarfbeetles of two families. Smith (1964) found that the number ofommatidia in the eyes of macropterous Ptinidae was 150e350, inbrachypterous e 100e150, in apterous e 25e50. Dybas (1978)inspected dimorphic species of featherwing beetles of the genusPinellodes and stated that the eyes in wingless morphs werereduced or absent. Eyes are presumably involved in the reductionprocess due to the adaptation to the changed way of life. Bouchardand Gorb (2000) stated absence of mechanosensory setae, whichcontrol the contact between co-opted frictional fields of micro-trichia on the metanotum and the elytra in flying tenebrionids, inthe flightless darkling beetle Tabarus montanus with permanentlylinked elytra.
We suggest that the loss of the flight ability and especially theacquisition of immobile elytra make strain measurements in theelytral articulation unnecessary and lead to the extinction of thefield of campaniform sensilla in this articulation. Our observationsrevealed that the grade of the wing reduction, the grade of elytrontransformation and the grade of Lehr's field reduction are differentin diverse taxa (Table 1). Future observations on Lehr's fields indimorphic species may elucidate the morphogenetic backgroundfor those linked events. The main tendencies and discrepanciesconcerning the field of CFS are as follows:
The decrease in number of CFS on elytral roots correlates withthe shortening of the hind wings (Figs. 1 and 4). Mesopterousspecies bear 50e30% of CFS, when compared with macropterousspecies. The reduction of Lehr's fields is even more profound inbrachyptrerous beetles.
Sensory fields are present in insects with strongly reducedwings (Gaurodites bifarius, Carabus coriaceus, Car. hungaricus,Chrysolina coerulea, Ch. olivacea, Sitophilus granarius, Liparus glab-rirostris). As long as elytra are separable (excluding Melo€e), theLehr's field is present.
The complete loss of CFS on the root of the elytron is obligatorycoinciding with the presence of the permanent sutural lock. Thesign of such a lock is a thickened rim of the ventral ridge, which isbulb-like in cross-section. Certainly, permanent locks appearedindependently in different taxa of beetles.
The inverse coincidence between the presence of permanentlocks and the loss of CFS does not hold true. Carabids with suchlocks, Carabus spp. (Fig. 7) or Zabrus tenebrioides, exhibit a residualset of sensilla.
4.3. Reduction of sensilla in feather-wing beetles
In the first part of this study (Frantsevich et al., 2014), weinspected several dwarf flying beetles: a lady bird Stethoruspunctillum, a weevil Sitophilus oryzae, and a rove beetleEusphalerium alpinum. All these species had elytra with an area
in the range of 0.63e0.73 mm2 and the number of CFS in theirLehr's fieldsein the range of 17e26. The elytron in P. anodontawas about 20 times smaller (0.031 mm2). According to thegeneral allometric relationship in Lehr's field, the number ofsensilla could be predicted to be about √20 times lower thanthat observed in dwarf beetles, i.e. about 4e5 CFS. We found noCFS either at the base of the elytron or at the base of the hindwing, where they are abundant in bigger flying beetles (Lehr,1914; Pfau and Honomichl, 1979), as well as in insects ofother orders (Pringle, 1957, Table 4).
Aerodynamic conditions for flight in Ptiliidae and other minia-ture insects with fringed wings (thrips Thysanoptera, fairy waspsMymaridae) are different, if compared to the insects with mem-branous wings. An important functional principle of membranouswings is their ability to create a difference in air pressure on bothsides of the wing and thus generate aerodynamic forces (Bennett,1972). It was previously reported that miniature insects, evenwith membranous wings, fly at very low Reynolds numbers (Re10e20 and less, Horridge, 1956; Walker, 2002). Weis-Fogh (1973)recorded membranous wing kinematics in a miniature waspEncarsia formosa. The value of Re was assessed as 17 during hov-ering, at a high stroke frequency of 400 s�1. Weis-Fogh alsoobserved a structure on the forewings, which he recognized as agroup of CFS.
Branching elements (setae) of a fringed wing are slim, whichcauses even further diminution of Re by several orders of magni-tude. Miniature insects may float in the air like the seeds of adandelion. According to the model computed by Cheer and Koehl(1987), a meshwork of slim cylinders is adjoined with a boundarylayer of fluid or air which moves together with the cylinders. Thefan of unfolded setae is able to produce sufficient aerodynamicforce during the power stroke, whereas passive bending of bend-able stem segments and setae presumably changes the shape, re-duces wing area, and diminishes the drag of the fringed wingduring the return stroke.We suppose that automatic change of dragmay occur in the feather-wing beetle, and that such a mode ofpropulsion needs minimal sensory control.
We suppose that automatic change of drag may occur in thefeather-wing beetle, and that such a mode of propulsion needsminimal sensory control.
Our general conclusion is that reduction or simplification of thehind wing structure and function in beetles correlates with partialor complete reduction of Lehr's field of CFS.
Acknowledgments
We acknowledge generous donations of beetle specimens byVassily Dyadichko, Boris Korotyaev, Vitaliy Nazarenko, AlexanderParkhomenko, Alexander Puchkov, Bogdan Vas'ko, Taras Yanicki.Our cordial thanks are due to Joachim Oesert and Angela Vee-nendaal for their skilled technical assistance. We are thankful toanonymous reviewers for their valuable suggestions and advice.Vicky Kastner and one of the reviewers provided linguisticcorrections of the manuscript. Visits of L.F. to the University ofKiel were supported by the project C10 of the German ScienceFoundation (DFG SFB 677). Studies on featherwing beetles werefunded by Russian Scientific Foundation (grant 14-14-00208 toA.P).
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.asd.2014.10.003.

L. Frantsevich et al. / Arthropod Structure & Development 44 (2015) 1e9 9
References
Bennett, L., 1972. Effectiveness and flight of small insects. Ann. Entomol. Soc. Am.66, 1187e1190.
Blum, P., 1979. Zur Phylogenie und Oekologischen Bedeutung der Elytrenreduktionund Abdomenbeweglichkeit der Staphylinidae (Coleoptera). Vergleichend- undfunktionmor phologische Untersuchungen. Zoologische Jahrbücher. Abt. Anat.Ontog. Tiere 102, 533e582.
Bouchard, P., Gorb, S., 2000. The elytra-to-body binding mechanism of the flightlessrainforest species Tabarus montanus Kaszab (Coleoptera: Tenebrionidae).Arthropod Struct. Dev. 29, 323e331.
Bowker, J.-D., 2002. A mark and recapture study of water beetles (Coleoptera:Dytiscidae) in a group of semi-permanent and temporary ponds. Aquat. Ecol.36, 435e446.
Brovdiy, V.M., 1973. Leaf beetles. Galerucinae. In: Fauna of Ukraine (in Ukrainian),vol. 19, Iss. 17. Naukova dumka, Kiev.
Brovdiy, V.M., 1977. Leaf beetles. Chrysomelinae. In: Fauna of Ukraine (in Ukrainian),vol. 19, Iss. 16. Naukova dumka, Kiev.
Cheer, A.Y.L., Koehl, M.A.R., 1987. Paddles and rakes: fluid flow through bristledappendages of small organisms. J. Theor. Biol. 129, 17e39.
Cherney, L.S., 2005. Darkling beetles (Coleoptera, Tenebrionidae). In: Fauna ofUkraine (in Russ.), vol. 19, Iss. 10. Naukova dumka, Kiev.
Darlington, P.J., 1936. Variation and atrophy of flying wings of some carabid beetles.Ann. Entomol. Soc. Am. 29, 136e175.
Davydova, E.D., 1966. Wing muscles of fire-flies (Coleoptera, Lampyridae) withdifferent grade of wing reduction (in Russ.). Zool. Zhurnal 45, 1023e1032.
Duncan, F.D., 2002. The role of the subelytral cavity in water loss in the flightlessdung beetle, Circellium bacchus (Coleoptera: Scarabaeinae). Eur. J. Entomol. 99,253e258.
Dybas, H.S., 1978. Polymorphism in featherwing beetles, with a revision of thegenus Pinellodes (Coleoptera, Ptiliidae). Ann. Entomol. Soc. Am. 71, 695e714.
Fedorenko, D.N., 2004. Folding system in beetles' wings and its main types (inRuss.). Zool. Zhurnal 83, 693e700 and 795e808.
Fiori, G., 1975. La 'sutura' elitrale dei coleotteri. Atti del Congresso Naz. Ital. Entomol.10, 91e111.
Frantsevich, L., 2004. Righting kinematics in beetles (Insecta: Coleoptera).Arthropod Struct. Dev. 33, 221e235.
Frantsevich, L., Gorb, S., Radchenko, V., Gladun, D., Polilov, A., 2014. Lehr's fields ofcampaniform sensilla in beetles (Coleoptera): functional morphology. I. Generalpart and allometry. Arthropod Struct. Dev. 43, 523e535.
Furth, D.G., 1980. Wing polymorphism, host plant ecology, and biogeography ofLongitarsus in Israel (Coleoptera: Chrysomelidae). Israel J. Entomol. 13,125e148.
Hammond, P.M., 1985. Dimorphism of wings, wing-folding and wing-toiletry de-vices in the ladybird, Rhyzobius litura (F.) (Coleoptera: Coccinellidae), with adiscussion of inter-population variation in this and other wing-dimorphicbeetle species. Biol. J. Linn. Soc. 24, 15e33.
Heberdey, R.F., 1938. Beitr€age zum Bau des Subelytralraumes und zur Atmung derColeopteren. Z. für Morphol. €Okol. Tiere 33, 667e734.
Horridge, A., 1956. The flight of very small insects. Nat. Lond. 178, 1334e1335.Jackson, D.J., 1928. The inheritance of long and short wings in the weevil (Sitona
hispidula), with a discussion of wing reduction among beetles. Trans. R. Soc.Edinb. 55, 665e735.
Jackson, D.J., 1956. Observations on flying and flightless water beetles. J. Linn. Soc.Lond. 43, 18e42.
Kolbe, H., 1920. Über Mutationen und Aberrationen deutscher und ausw€artigerColeopteren und anderer Insekten. Berl. Entomol. Z. (3/4), 386e408.
Leech, H.B., 1942. Dimorphism in the flying wings of a species of water beetle,Agabus bifarius (Kirby). Ann. Entomol. Soc. Am. 35, 76e80.
Lehr, R., 1914. Die sinnesorgane der beiden Flügelpaare von Dytiscus marginalis.Z. für Wiss. Zool. 110, 87e150.
Lindroth, C.H., 1946. Inheritance of wing dimorphism in Pterostichus anthracinus Ill.Hereditas 32, 37e40.
Linnaeus, C.N., 1758. Systema Naturae per Regna Tria Naturae. Vol. 1. LaurentisSalvii, Holmiae impensis direct.
Miguelez, D., Valladares, L.F., 2008. Seasonal dispersal of water beetles (Coleoptera)in an agricultural landscape: a study using Moericke traps in northwest Spain.Ann. la Soc. Entomol. Fr. 44, 317e326.
Munray, F.V., Tiegs, O.W., 1935. The metamorphosis of Calandra oryzae. Quat. J.Microsc. Sci. 77, 405e495.
Nachtigall, W., 1961. Einige Beobachtungen über die Fortbewegung der Dytiscidenaußerhalb des Wassers. Zool. Anz. 166, 105e108.
Nazarenko, V.Y., 1996. On the structure of rudimentary wings in weevils of thegenus Liparus Olivier (Coleoptera, Curculionidae) (in Russ.). Entomol. Obozr. 75,113e116.
Nikolaev, G.V., 1987. Lamellicornian Beetles (Coleoptera, Scarabaeidae) of theKazakhstan and Middle Asia (in Russ.). Nauka, Alma-Ata.
Oertel, R., 1924. Studien üeber Rudimentation bei der Gattung Carabus. Z. fürMorphol. €Okologie Tiere 1, 38e120.
Parkhomenko, O.V., 2002. Carrion Beetles (Coleoptera, Silphidae) of the UkrainianFauna (in Ukrainian) (Thesis). National Pedagogical Dragomanov-University,Kiev.
Pfau, H.K., Honomichl, K., 1979. Die campaniformen Sensillen des Flügels vonCetonia aurata L. und Geotrupes silvaticus Panz. (Insecta, Coleoptera) in ihrerBeziehung zur Flügelmechanik und Flugfunktion. Zoologische Jahrbücher. Abt.Anat. Ontog. 102, 583e613.
Plavilschikov, N.N., 1936. Long-horn beetles (part 1). In: Fauna of the USSR. InsectaColeoptera (in Russ.). Volume 23. Academy of Sciences of the USSR, Moscow-Leningrad.
Plavilschikov, N.N., 1958. Long-horn beetles (part 3). In: Fauna of the USSR. InsectaColeoptera (in Russ.). Volume 23. Academy of Sciences of the USSR, Moscow-Leningrad.
Polilov, A.A., 2008. Anatomy of the smallest of the Coleoptera, feather-wingedbeetles from tribe Nanosellini (Coleoptera, Ptiliidae) and limits to insect mini-aturization. Entomol. Rev. 88, 26e33.
Polilov, A.A., Beutel, R.G., 2009. Miniaturization effects in larvae and adults of Mi-kado sp. (Coleoptera: Ptiliidae), one of the smallest free-living insects.Arthropod Struct. Dev. 38, 247e270.
Pringle, J.W.S., 1957. Insect Flight. Cambridge University Press.Rüschkampf, F., 1927. Der Flugapparat der K€aefer. Vorbedingung, Verlauf und
Ursache seiner Rüeckbildung. Zoologica 75, 1e88.Samuelson, G.A., 1996. Binding sites: elytron-to-body meshing structures of
possible significance in the higher classification of Chrysomelidae. In:Joliet, P.H.A., Cox, M.L. (Eds.), Chrysomelidae Biology, Vol. 1: the Classification,Phylogeny, and Genetics. SPP Academic Publishing, Amsterdam, pp. 267e290.
Schneider, P., 1978. Die Flug- und Faltungstypen der K€afer (Coleoptera). ZoologischeJahrbücher. Abt. Anat. Ontog. Tiere 99, 174e210.
Schneider, P., 1986. Studies about the flight of Dytiscus marginalis. 1. (Coleoptera,Dytiscidae). Entomol. Basil. 11, 451e460.
Schulze, P., 1912. Die Flüegelrudimente der Gattung Carabus. Zool. Anz. 40, 188e194.Smith, D.S., 1964. The structure and development of flightless Coleoptera: a light
and electron microscopic study of the wings, thoracic exoskeleton and rudi-mentary flight musculature. J. Morphol. 114, 107e184.
Suzuki, S., Kobayashi, N., Kato, T., Katakura, H., 2001. Flight muscle dimorphism inGeotrupes laevistriatus (Geotrupidae: Coleoptera). Entomol. Sci. 4, 291e294.
Taylor, V.A., 1981. The adaptive and evolutionary significance of wing poly-morphism and parthenogenesis in Ptinella Motschulsky (Coleoptera, Ptiliiidae).Ecol. Entomol. 6, 89e98.
Tietze, F., 1963. Untersuchungen über die Beziehungen zwischen Flügelreduktionund Ausbildung des Metathorax bei Carabiden unter besonderer Ber-ücksichtigung der Flugmuskulatur. Beitr. zur Entomol. 13, 88e167.
Walker, J.A., 2002. Functional morphology and virtual models: physical constraintson the design of oscillating wings, fins, legs, and feet at intermediate Reynoldsnumbers. Integr. Comp. Biol. 42, 232e242.
Weis-Fogh, T., 1973. Quick estimates of flight fitness in hovering animals, includingnovel mechanisms for lift production. J. Exp. Biol. 59, 169e230.
Zaitsev, F.A., 1953. Diving and whirligig beetles (in Russ.). In: Fauna of the USSR.Insecta Coleoptera. Volume 4. Academy of Sciences of the USSR, Moscow-Leningrad.
Copyright © 2022 FDOKUMEN




















