
Late postglacial paleoenvironmental change in the northeastern Mediterranean region: Combined...
10
Late postglacial paleoenvironmental change in the northeastern Mediterranean region: Combined palynological and molecular biomarker evidence K. Kouli a, * , A. Gogou b , I. Bouloubassi c , M.V. Triantaphyllou a , Chr. Ioakim d , G. Katsouras b , G. Roussakis b , V. Lykousis b a University of Athens, Faculty of Geology & Geoenvironment, Dept. of Historical Geology-Paleontology, Panepistimiopolis, 15784 Athens, Greece b Hellenic Centre for Marine Research, Institute of Oceanography,19013 Anavyssos, Greece c LOCEAN-IPSL, UMR 7159 CNRS/IRD/UPMC/MNHN, Université P. et M. Curie, 4 Place Jussieu, 75252 Paris Cedex 05, France d Institute of Geology and Mineral Exploration, Olympic Village, Acharnae-Attiki 13677, Greece article info Article history: Available online xxx abstract Three gravity cores collected from the NE Mediterranean (NEMR) across a transect from the northern Aegean Sea (North Skyros basin) to the south Cretan margin (SCM), were investigated for pollen and terrestrial biomarkers derived from epicuticular waxes of vascular plants during the last w20 ky. Pollen data show diversified mixed temperate forest in the northern borderlands and enhanced Mediterranean vegetation in the southern areas, documenting an NeS climatic trend. Terrestrial plant biomarkers and their diagnostic geochemical indices exhibit latitudinal patterns which are interpreted in terms of the different delivery pathways (fluvial/runoff vs. atmospheric transport), resulting from the climate conditions during different periods. During the Late Glacial and early deglaciation periods (20e14 ka BP) relatively increased humidity (H-index) is recorded in the north Aegean Sea, while in the South drier climate was the limiting factor for vegetation development. During this interval, terrestrial n-alkanes showed increased accumulation rates, suggesting massive transport of terrestrial organic matter through runoffs and rivers, followed by weaker input after 14 ka BP. After w11 ka BP a major expansion of forest cover is evidenced in the NEMR, accompanied by a higher H-index because of the climatic amelioration. The forest vegetation exhibited regionally different characteristics, with cool temperate taxa being more abundant in the Aegean cores, while the SCM record is being featured by Mediterranean elements. At the onset of the Holocene and throughout the Holocene Climatic Optimum the delivery of terrestrial biomarkers increased and became more significant in the Aegean sites compared to the SCM site. Within the Holocene, the average chain length (ACL) of long chain n-alkanes exhibits lower values in the northern Aegean than in the southeastern Aegean and SCM, indicating the predominance of warmer species southwards. Finally, the H-index records a conspicuous humidity increase between 5.4 and 4.3 ka BP in the south Aegean that coincides with an increase in the terrestrial biomarker supply and the deposition of a distinct sapropel-like layer, SMH (Sapropel Mid Holocene). Similar trends in T (temper- ature) and H indices are slightly delayed and attenuated in the northern Aegean and are accompanied by an increase in the ACL index. A noticeable increase in the accumulation rates (ARs) of terrestrial biomarkers and the HPA index values during this period are clearly recorded in all three cores, indicative of enhanced terrigenous inputs of organic matter along with higher in-situ preservation. Ó 2011 Elsevier Ltd and INQUA. 1. Introduction The northeastern Mediterranean region (NEMR, Fig. 1) covers an area between 38 and 45 N latitude and 20 e30 E longitude, including continental Greece and the Aegean Sea, western Turkey and the southern shelf of Crete (south Cretan margin, SCM). NEMR resides in a climatological transition zone that is under the influ- ence of both tropical and mid-latitude regimes, large topographic contrasts that act as specific boundary conditions and global atmospheric teleconnections (Lionello et al., 2006). A temperate humid climate prevails in its northern part, while drier conditions characterize the south. Within this region, the Aegean Sea has been recognized as highly sensitive area to regional and global climate * Corresponding author. E-mail address: [email protected] (K. Kouli). Contents lists available at SciVerse ScienceDirect Quaternary International journal homepage: www.elsevier.com/locate/quaint 1040-6182/$ e see front matter Ó 2011 Elsevier Ltd and INQUA. doi:10.1016/j.quaint.2011.10.036 Quaternary International xxx (2011) 1e10 Please cite this article in press as: Kouli, K., et al., Late postglacial paleoenvironmental change in the northeastern Mediterranean region: Combined palynological and molecular biomarker evidence, Quaternary International (2011), doi:10.1016/j.quaint.2011.10.036
-
Upload
sorbonne-fr -
Category
Documents
-
view
0 -
download
0
Transcript of Late postglacial paleoenvironmental change in the northeastern Mediterranean region: Combined...

at SciVerse ScienceDirect
Quaternary International xxx (2011) 1e10
Contents lists available
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
Late postglacial paleoenvironmental change in the northeastern Mediterraneanregion: Combined palynological and molecular biomarker evidence
K. Kouli a,*, A. Gogou b, I. Bouloubassi c, M.V. Triantaphyllou a, Chr. Ioakim d, G. Katsouras b, G. Roussakis b,V. Lykousis b
aUniversity of Athens, Faculty of Geology & Geoenvironment, Dept. of Historical Geology-Paleontology, Panepistimiopolis, 15784 Athens, GreecebHellenic Centre for Marine Research, Institute of Oceanography, 19013 Anavyssos, Greecec LOCEAN-IPSL, UMR 7159 CNRS/IRD/UPMC/MNHN, Université P. et M. Curie, 4 Place Jussieu, 75252 Paris Cedex 05, Franced Institute of Geology and Mineral Exploration, Olympic Village, Acharnae-Attiki 13677, Greece
a r t i c l e i n f o
Article history:Available online xxx
* Corresponding author.E-mail address: [email protected] (K. Kouli).
1040-6182/$ e see front matter � 2011 Elsevier Ltd adoi:10.1016/j.quaint.2011.10.036
Please cite this article in press as: Kouli, KCombined palynological and molecular biom
a b s t r a c t
Three gravity cores collected from the NE Mediterranean (NEMR) across a transect from the northernAegean Sea (North Skyros basin) to the south Cretan margin (SCM), were investigated for pollen andterrestrial biomarkers derived from epicuticular waxes of vascular plants during the last w20 ky. Pollendata show diversified mixed temperate forest in the northern borderlands and enhanced Mediterraneanvegetation in the southern areas, documenting an NeS climatic trend. Terrestrial plant biomarkers andtheir diagnostic geochemical indices exhibit latitudinal patterns which are interpreted in terms of thedifferent delivery pathways (fluvial/runoff vs. atmospheric transport), resulting from the climateconditions during different periods. During the Late Glacial and early deglaciation periods (20e14 ka BP)relatively increased humidity (H-index) is recorded in the north Aegean Sea, while in the South drierclimate was the limiting factor for vegetation development. During this interval, terrestrial n-alkanesshowed increased accumulation rates, suggesting massive transport of terrestrial organic matter throughrunoffs and rivers, followed by weaker input after 14 ka BP. After w11 ka BP a major expansion of forestcover is evidenced in the NEMR, accompanied by a higher H-index because of the climatic amelioration.The forest vegetation exhibited regionally different characteristics, with cool temperate taxa being moreabundant in the Aegean cores, while the SCM record is being featured by Mediterranean elements. At theonset of the Holocene and throughout the Holocene Climatic Optimum the delivery of terrestrialbiomarkers increased and became more significant in the Aegean sites compared to the SCM site. Withinthe Holocene, the average chain length (ACL) of long chain n-alkanes exhibits lower values in thenorthern Aegean than in the southeastern Aegean and SCM, indicating the predominance of warmerspecies southwards. Finally, the H-index records a conspicuous humidity increase between 5.4 and 4.3 kaBP in the south Aegean that coincides with an increase in the terrestrial biomarker supply and thedeposition of a distinct sapropel-like layer, SMH (Sapropel Mid Holocene). Similar trends in T (temper-ature) and H indices are slightly delayed and attenuated in the northern Aegean and are accompanied byan increase in the ACL index. A noticeable increase in the accumulation rates (ARs) of terrestrialbiomarkers and the HPA index values during this period are clearly recorded in all three cores, indicativeof enhanced terrigenous inputs of organic matter along with higher in-situ preservation.
� 2011 Elsevier Ltd and INQUA.
1. Introduction
The northeasternMediterranean region (NEMR, Fig. 1) covers anarea between 38� and 45�N latitude and 20�e30�E longitude,including continental Greece and the Aegean Sea, western Turkey
nd INQUA.
., et al., Late postglacial palearker evidence, Quaternary
and the southern shelf of Crete (south Cretan margin, SCM). NEMRresides in a climatological transition zone that is under the influ-ence of both tropical and mid-latitude regimes, large topographiccontrasts that act as specific boundary conditions and globalatmospheric teleconnections (Lionello et al., 2006). A temperatehumid climate prevails in its northern part, while drier conditionscharacterize the south. Within this region, the Aegean Sea has beenrecognized as highly sensitive area to regional and global climate
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036

Fig. 1. Location map of the NEMR, showing the locations of the studied sites, thebathymetry and the rivers.
K. Kouli et al. / Quaternary International xxx (2011) 1e102
change and is hence ideal for recording past climate variability dueto its higher sedimentation rates compared to the more oceanicsites of the Mediterranean Sea (Aksu et al., 1995; Geraga et al.,2000; 2005; Casford et al., 2003; Roussakis et al., 2004; Gogouet al., 2007; Ehrmann et al., 2007a; 2007b; Hamann et al., 2008;Marino et al., 2009; Triantaphyllou et al., 2009a, 2009b, 2010a;2010b).
Since the Last Glacial Maximum (w20 ka BP), the marine realmof the NEMR has undergone several important, climate-drivenphases of reorganization of its hydrological and biogeochemicalfunctioning that are recorded in its sediment archives (Myers et al.,1998; Stratford et al., 2000; Casford et al., 2003; Emeis et al., 2003;Gogou et al., 2007; Triantaphyllou et al., 2009a, 2009b; 2010b;Schmiedl et al., 2010). At the same time, terrestrial ecosystemsappear highly variable according to the local conditions (latitude,relief, hydrological regime etc) and record centennial climaticoscillations (e.g. Kotthoff et al., 2008b; Triantaphyllou et al., 2009b;Geraga et al., 2010). Paleovegetation changes have been identifiedin pollen abundances of the Mediterranean region both in terres-trial (Tzedakis, 1999; Tzedakis et al., 2002; Magny et al., 2003;Lawson et al., 2004; Triantaphyllou et al., 2010a) and in marinedeposits (Aksu et al., 1995; 1999; Mudie et al., 2002a; 2002b;Geraga et al., 2005; 2010; Kotthoff et al., 2008a; Kholeif andMudie, 2009; Triantaphyllou et al., 2009a; 2009b). Quantitativeclimate reconstructions across the Mediterranean based on marinepollen data document the increased sensibility of Aegean Searecords to climatic oscillations compared to western Mediterra-nean sites, suggesting a West/East precipitation gradient mostlyuntil the 8.2 ka event (Combourieu Nebout et al., 2009; Dormoyet al., 2009).
Biomarkers have become an increasingly common tool in thereconstruction of past climate and environmental conditions.Biomarkers from higher (terrestrial) plants are common in marinesediments and can be used to reconstruct changes in past conti-nental vegetation, from which climatic change can be inferred(Pancost and Boot, 2004; Sicre et al., 2006; Eglinton and Eglinton,2008). Among the biomarkers, long chain n-alkyl compounds (n-alkanes, n-alcohols and n-alkanoic acids) are major components ofepicuticular waxes from vascular plant leaves (Eglinton and
Please cite this article in press as: Kouli, K., et al., Late postglacial paleCombined palynological and molecular biomarker evidence, Quaternary
Hamilton, 1967) and degradation products of terrestrial plantdetritus in general (Aksu et al., 1999). Based on patterns of longchain n-alkanes and n-alkanols, present and past fluctuations inthe supply of terrigenous organic matter in different marine sitesin the Mediterranean have been documented for various time-windows [Late Glacial, Holocene Climate Optimum (HCO), 8.2 kacooling event, Mid-Late Holocene] and have provided insight intocontinental environmental/climatic conditions (Aksu et al., 1995;1999; Bouloubassi et al., 1997; 1999; Cacho et al., 2000; Gogouand Stephanou, 2004; Gogou et al., 2007; Triantaphyllou et al.,2009b). However, the integrative approach of using vascularplant biomarkers and pollen/spores is rare in high resolutionmarine sedimentary sequences (Aksu et al., 1999; Abrajano et al.,2002).
The purpose of this study is to assess the Late Glacial to Holo-cene paleoenvironmental changes in NERM based on the combinedinvestigation of terrestrial palynological proxies and biomarkersthat yield information on climate-related parameters e.g. vegeta-tion type, precipitation/humidity, temperature, supply of terrestrialmaterial to the sea. We focus on three cores along a transect fromthe northern Aegean Sea (North Skyros basin) to the south Cretanmargin (SCM), in order to examine possible environmental andclimatic trends in this region. Our proxies identify distributionpatterns of pollen taxa, forestation, accumulation rates of terrestrialbiomarkers as well as humidity (H), temperature (T) and biomarkerindices that provide important information on continental climateand vegetation dynamics and gradients and their delivery path-ways to the selected marine sites.
2. Regional and climatic setting
The Aegean Sea is located in the northern part of the NEMR(Fig. 1). It is linked to the Black Sea through the Dardanelles andBosporus Straits and to the eastern Mediterranean through theCretan Straits. It exhibits a complicated physical and geographicconfiguration, displaying complicated seabed morphology andnumerous island complexes (Lykousis et al., 2002). Numerousmajor rivers (catchments > 1000 km2) from the surrounding areasof the Balkans and Turkey flow into the Aegean Sea. Evros, Nestos,Strymon, Axios and Aliakmon Rivers, discharging along the northcoast, and Bakircay, Gediz, Büyük and KücükMenderes draining thewestern Turkey are the most important sediment contributorsproviding 75% of the Aegean sediment influx (Lykousis et al., 2002;Roussakis et al., 2004; Ehrmann et al., 2007a; Poulos, 2009). Theserivers collectively constitute an important source of land-derivedorganic matter to the northern study site (Fig. 1). In contrast,river influx in the southeastern Aegean and SCM sites is signifi-cantly lower. There aeolian transport is of major importance for thedelivery of land-derived material (Gogou et al., 1996; 2000; Gogouand Stephanou, 2004; Ehrmann et al., 2007b).
The present-day climate in the NEMR is characterized bya strong seasonality, with warm to hot, dry summers and mild, wetwinters (Lionello et al., 2006). The wind field of the Aegean Sea isdominated by northwinds, while south-southwesterly winds occurduring spring and diurnal sea-breezes occur along the Greek andTurkish coasts during summer (Poulos et al., 1997). Seasonality is ofgreat importance in interpreting pollen records in the NEMR:winter temperature and summer drought are the bioclimaticfactors that controlled the vegetation dynamics through the lastclimatic cycles (Rossignol-Strick, 1999). Additionally, physiographyplays an important role in the distribution of temperature andprecipitation. For example, temperature progressively increasesfrom north to south during all seasons, and differences betweenplains and mountains, interior and coastal areas are observed(Tzedakis, 2000).
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036

K. Kouli et al. / Quaternary International xxx (2011) 1e10 3
3. Materials and methods
3.1. Sampling sites
Three gravity cores along an NeS transect were used in thepresent study (Fig.1), two from theAegeanSea andone further southfrom the SCM. Core MNB-3 (39�15.4300N, 25�00. 0000E, length 4 m),was retrieved fromNorth Skyros Basin at awater depth of 800mandcovers the last 24 ky (Gogou et al., 2007). North Skyros basin islocated in the southern part of the North Aegean Sea (e.g. Aksu et al.,1995; Papanikolaou et al., 2002) and is characterized by the “conti-nental margin” sedimentation processes of the North Aegean(Lykousis et al., 2002; Poulos, 2009). Core NS-14 (36�38,9190N,27�02,8750E, length 400 cm) was collected from the western Kosbasin (southeastern Aegean) at a water depth of 505 m, and coversabout the last 13 ky (Triantaphyllou et al., 2009a; 2009b). Finally coreHCM2/22 (34�33.9680N, 24�53.7700E, length 175 cm) was collectedoffshore of south Crete (SCM) at a water depth of 2211 m. The upper60 cm of HCM2/22 covers the last 20 kyr (Katsouras et al., 2010). Allthree cores contain sapropel S1, consisting of two discrete intervals,the S1a and S1b sections, along with the S1 interruption. S1 depo-sition onset shows an SeN trend (Katsouras et al., 2010) startingearlier in the SCM (HCM2/22: 10.8 ka cal BP) and later in the Aegean(NS-14: 10 cal ka BP; MNB-3: 9.6 ka BP).
3.2. Age Models
The chronostratigraphy of the cores studied is based on sixaccelerator mass spectroscopy (AMS) radiocarbon (14C) datesavailable for MNB-3 (Gogou et al., 2007), seven for core NS-14(Triantaphyllou et al., 2009b) and three for core HCM2/22 (Ioakimet al., 2009; Katsouras et al., 2010). For the age model of MNB-3additional dating based on planktonic fauna variations in accor-dance to the model proposed by Casford et al. (2002) have beenused (Geraga et al., 2010). The age model of NS-14 (Triantaphyllouet al., 2009b) has been tested against the multi-proxy chronologicalframework proposed by Casford et al. (2007). The dates of coreHCMR2/22 (Ioakim et al., 2009) have been calibrated usinga marine reservoir correction, R 58 � 85 (Stuiver et al., 1998). Allages in this study refer to calibrated 14C ages.
3.3. Pollen analysis
Pollen samples were collected at intervals of 0.5e3 cm (repre-senting average time resolution of 160 yrs in MNB-3, 90 yrs in NS-14 and 400 yrs in HCM2/22), oven-dried, weighed, spiked withLycopodium clavatum spores, treated with 10% HCl and 38% HF andeventually sieved over a 10 mm sieve. Pinus, pteridophyte sporesand aquatic taxa were excluded from pollen sums. Detailed paly-nological diagrams of MNB-3 and NS-14 cores have been presentedin Geraga et al. (2010) and Triantaphyllou et al. (2009a) respec-tively. For the present study pollen data are summarized in thefollowing groups: Cool temperate evergreen taxa include Abies,Cedrus, Picea and Taxus. Cool temperate summergreen taxa includeAlnus, Betula, Corylus, Fagus, Quercus robur and Tilia. Warmtemperate summergreen taxa include Acer, Fraxinus, Juglans, Ostrya,Platanus, Populus and Ulmus. Mediterranean taxa include Cistus,Quercus ilex, Phillyrea, Pistacia and Olea. In addition the total curveof Non Arboreal Pollen (NAP) is used. Two different pollen ratioshave been used as qualitative paleoenvironmental proxies. Theratio H ¼ AP/St (AP: Arboreal taxa excluding Pinus; St: Steppic taxaincluding Artemisia, Chenopodiaceae, Asteraceae and Poaceae), hasbeen used as a humidity index (H-index; Triantaphyllou et al.,2009b). As a relative temperature index the ratio of cool to warmtemperate broadleaved taxa (T-index ¼ cool temperate/(sum of
Please cite this article in press as: Kouli, K., et al., Late postglacial paleCombined palynological and molecular biomarker evidence, Quaternary
temperate andMediterranean) has been used, while the forestationcover index has been calculated based on the ratio of broadleavedtaxa (Acer, Alnus, Betula, Cistus, Corylus, Fagus, Fraxinus, Juglans, Olea,Phillyrea, Pistacia, Platanus, Populus, Ostrya, Quercus, Tilia, Ulmus)versus pollen sum excluding bisaccates (Kotthoff et al., 2008b).
3.4. Terrestrial biomarkers
Sediment samples were collected with 0.5e2 cm resolution(representing average time resolution of 300 y inMNB-3, 200 yrs inNS-14 and 400 yrs in HCM2/22) and freeze-dried. Lipids wereextractedwith amixture of dichloromethane/methanol (4:1) undersonication (three times). The extracts were concentrated ona rotary evaporator under vacuum and evaporated to dryness undera nitrogen stream. Aliphatic hydrocarbons and alcohols were iso-lated using silica gel column chromatography and analysed on gaschromatography and gas chromatography/mass spectrometry (GC/MS) (Gogou et al., 2007). The compounds were identified on thebasis of their mass spectra, GC retention times, and comparisonwith literature mass spectra.
Odd-carbon number long chain n-alkanes and even-carbonnumber long chain n-alkanols derive from vascular plant leafwaxes (Eglinton and Hamilton, 1967). The sum of the most abun-dant terrestrial n-alkanes (n-C27, n-C29 and n-C31) and n-alkanols(n-C26, n-C28 and n-C30) are referred to as Ter-alkanes and Ter-alkanols respectively.
Accumulation rates (ARs) of Ter-alkanes were calculated usingabsolute biomarker amounts (expressed in ng g�1), as the productof linear sedimentation rates (LSR) and the dry bulk density of thesediment, according to the equation
AR�g=m2kyr�1
�¼ LSRðm=kyrÞ � Dry Bulk Density
�g=m3
�
Chain length variations in land plant biomarker are commonlyrelated to changes in the temperature and humidity/aridity in thegrowing environment of source vegetation, since plants tend tosynthesize longer chain length waxy components in response toelevated temperatures (Poynter et al., 1989; Poynter and Eglinton,1990). Thus, changes in the vegetation cover during differentclimatic periods could be inferred by changes in the average chainlength of terrestrial n-alkanes (ACL index). Furthermore, the rela-tion of the abundances of long chain n-alkanes and n-alkanols (HPAIndex; Poynter and Eglinton, 1990), are used to evaluate theproportions of labile and refractory organic matter (OM) deliveredin the marine environment, as also as the in situ preservation vs.degradation trends under different redox conditions.
ACL and HPA indices used in this study were calculatedaccording to the following calculations:
ACL ¼ fð27� ½n� C27�Þ þ ð29� ½n� C29�Þ þ ð31� ½n� C31�Þþ ð33� ½n� C33�Þg=
X½n� C27þ n� C29þ n� C31
þ n� C33�
HPA ¼ ½Ter� alkanols�=ð½Ter� alkanols� þ ½Ter� alkanes�Þ
4. Results
4.1. Northern Aegean Sea (core MNB 3)
4.1.1. PollenThe high abundances of NAP pollen is the main feature of the
period between 20 and 14 ka BP (Fig. 2), in spite the modest
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036
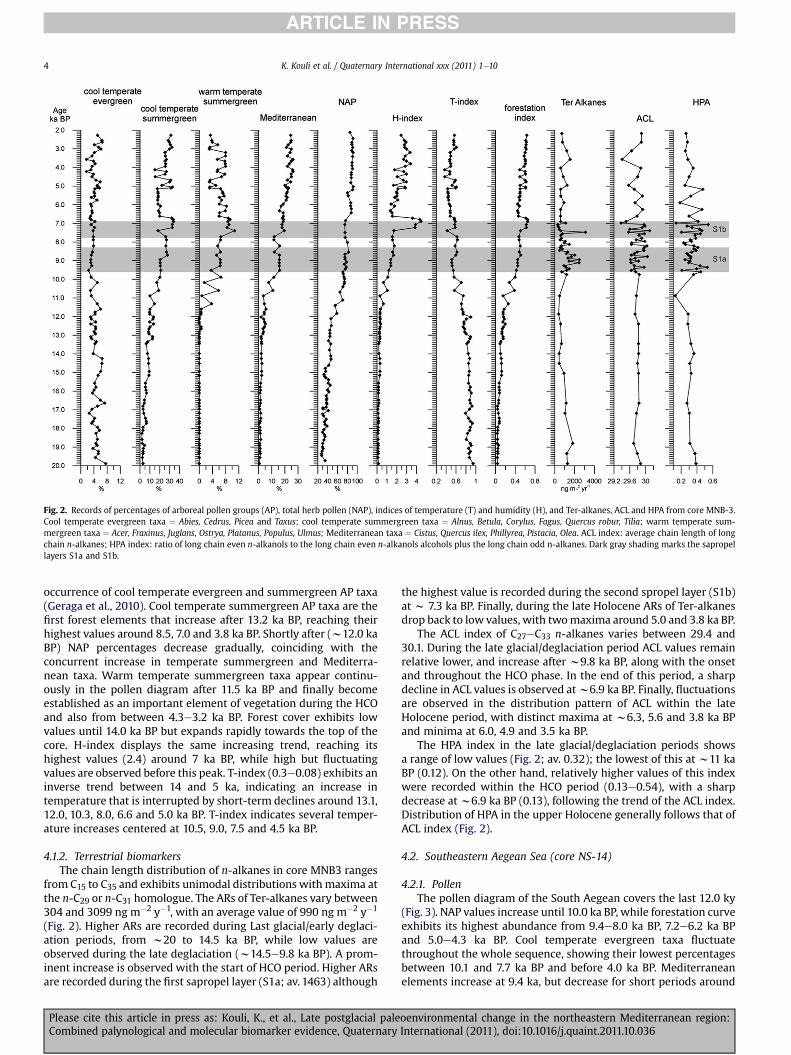
Fig. 2. Records of percentages of arboreal pollen groups (AP), total herb pollen (NAP), indices of temperature (T) and humidity (H), and Ter-alkanes, ACL and HPA from core MNB-3.Cool temperate evergreen taxa ¼ Abies, Cedrus, Picea and Taxus; cool temperate summergreen taxa ¼ Alnus, Betula, Corylus, Fagus, Quercus robur, Tilia; warm temperate sum-mergreen taxa ¼ Acer, Fraxinus, Juglans, Ostrya, Platanus, Populus, Ulmus; Mediterranean taxa ¼ Cistus, Quercus ilex, Phillyrea, Pistacia, Olea. ACL index: average chain length of longchain n-alkanes; HPA index: ratio of long chain even n-alkanols to the long chain even n-alkanols alcohols plus the long chain odd n-alkanes. Dark gray shading marks the sapropellayers S1a and S1b.
K. Kouli et al. / Quaternary International xxx (2011) 1e104
occurrence of cool temperate evergreen and summergreen AP taxa(Geraga et al., 2010). Cool temperate summergreen AP taxa are thefirst forest elements that increase after 13.2 ka BP, reaching theirhighest values around 8.5, 7.0 and 3.8 ka BP. Shortly after (w12.0 kaBP) NAP percentages decrease gradually, coinciding with theconcurrent increase in temperate summergreen and Mediterra-nean taxa. Warm temperate summergreen taxa appear continu-ously in the pollen diagram after 11.5 ka BP and finally becomeestablished as an important element of vegetation during the HCOand also from between 4.3e3.2 ka BP. Forest cover exhibits lowvalues until 14.0 ka BP but expands rapidly towards the top of thecore. H-index displays the same increasing trend, reaching itshighest values (2.4) around 7 ka BP, while high but fluctuatingvalues are observed before this peak. T-index (0.3e0.08) exhibits aninverse trend between 14 and 5 ka, indicating an increase intemperature that is interrupted by short-term declines around 13.1,12.0, 10.3, 8.0, 6.6 and 5.0 ka BP. T-index indicates several temper-ature increases centered at 10.5, 9.0, 7.5 and 4.5 ka BP.
4.1.2. Terrestrial biomarkersThe chain length distribution of n-alkanes in core MNB3 ranges
from C15 to C35 and exhibits unimodal distributions with maxima atthe n-C29 or n-C31 homologue. The ARs of Ter-alkanes vary between304 and 3099 ng m�2 y�1, with an average value of 990 ng m�2 y�1
(Fig. 2). Higher ARs are recorded during Last glacial/early deglaci-ation periods, from w20 to 14.5 ka BP, while low values areobserved during the late deglaciation (w14.5e9.8 ka BP). A prom-inent increase is observed with the start of HCO period. Higher ARsare recorded during the first sapropel layer (S1a; av. 1463) although
Please cite this article in press as: Kouli, K., et al., Late postglacial paleCombined palynological and molecular biomarker evidence, Quaternary
the highest value is recorded during the second spropel layer (S1b)at w 7.3 ka BP. Finally, during the late Holocene ARs of Ter-alkanesdrop back to low values, with twomaxima around 5.0 and 3.8 ka BP.
The ACL index of C27eC33 n-alkanes varies between 29.4 and30.1. During the late glacial/deglaciation period ACL values remainrelative lower, and increase after w9.8 ka BP, along with the onsetand throughout the HCO phase. In the end of this period, a sharpdecline in ACL values is observed atw6.9 ka BP. Finally, fluctuationsare observed in the distribution pattern of ACL within the lateHolocene period, with distinct maxima at w6.3, 5.6 and 3.8 ka BPand minima at 6.0, 4.9 and 3.5 ka BP.
The HPA index in the late glacial/deglaciation periods showsa range of low values (Fig. 2; av. 0.32); the lowest of this at w11 kaBP (0.12). On the other hand, relatively higher values of this indexwere recorded within the HCO period (0.13e0.54), with a sharpdecrease atw6.9 ka BP (0.13), following the trend of the ACL index.Distribution of HPA in the upper Holocene generally follows that ofACL index (Fig. 2).
4.2. Southeastern Aegean Sea (core NS-14)
4.2.1. PollenThe pollen diagram of the South Aegean covers the last 12.0 ky
(Fig. 3). NAP values increase until 10.0 ka BP, while forestation curveexhibits its highest abundance from 9.4e8.0 ka BP, 7.2e6.2 ka BPand 5.0e4.3 ka BP. Cool temperate evergreen taxa fluctuatethroughout the whole sequence, showing their lowest percentagesbetween 10.1 and 7.7 ka BP and before 4.0 ka BP. Mediterraneanelements increase at 9.4 ka, but decrease for short periods around
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036

Fig. 3. Records of percentages of arboreal pollen groups (AP), total herb pollen (NAP), indices of temperature (T) and humidity (H), and Ter-alkanes, ACL and HPA from core NS-14.Cool temperate evergreen taxa ¼ Abies, Cedrus, Picea and Taxus; cool temperate summergreen taxa ¼ Alnus, Betula, Corylus, Fagus, Quercus robur, Tilia; warm temperate sum-mergreen taxa ¼ Acer, Fraxinus, Juglans, Ostrya, Platanus, Populus, Ulmus; Mediterranean taxa ¼ Cistus, Quercus ilex, Phillyrea, Pistacia, Olea. ACL index: average chain length of longchain n-alkanes; HPA index: ratio of long chain even n-alkanols to the long chain even n-alkanols alcohols plus the long chain odd n-alkanes Dark gray shading marks the sapropellayers S1a and S1b and the SMH sapropelic layer.
K. Kouli et al. / Quaternary International xxx (2011) 1e10 5
7.8 ka BP (S1 interruption) and before 5.5 ka BP. The calculated H-index, ranging from 0.1 to 4.5, reflects repeated wet phasescentered atw9 ka BP,w6.8 ka BP andw4.8 ka BP corresponding toS1a, S1b, SMH depositional intervals (Triantaphyllou et al., 2009b).T-index values (0.3e0.8) record the occurrence of increasedtemperature during S1a deposition (10.0e7.8 ka BP), followed byfluctuations which indicate climatic deterioration and instability(Triantaphyllou et al., 2009b).
4.2.2. Terrestrial biomarkersTer-alkanols exhibit low ARs from 12 to 10 ka BP (av.
253 ng m�2 yr�1).A marked increase is recorded within the sap-ropel layer S1a with lower values within S1b and SMH (average:862, 101 and 180 ng m�2 y�1 respectively; Fig. 3).
Values of the ACL and HPA indices range from 29.14 to 30.69, andfrom 0.13 to 0.83, respectively (Fig. 3). ACL shows an increasingtrend from the late deglaciation to the Holocene in the southAegean record. The highest values for this index appear within thesapropel units S1a and S1b and the SMH (up to 30.69, 30.24 and30.22, respectively), whereas lower values occur at w 11 (29.59),8.4 (29.81), 6.8 (29.8), 5.8 (29.7) and 3.5 (29.96) ka BP. Furthermore,HPA index exhibits a pronounced increase during the S1a periodand less important ones during S1b and SMH periods (up to 0.83,0.64 and 0.53, respectively).
4.3. South Cretan margin (core HCM2/22)
4.3.1. PollenPollen patterns reveal the dominance of open vegetation
elements before 11.0 ka BP, with a short-term decline around11.5 ka BP. NAP values decrease during the deposition of S1a. Areverse trend is observed in H-index that exhibits increased valuesaround 11.5 ka BP (0.2) and between 11.0 and 5.5 ka BP (2.5),although lower in respect to the Aegean records. Mediterranean
Please cite this article in press as: Kouli, K., et al., Late postglacial paleCombined palynological and molecular biomarker evidence, Quaternary
and warm temperate summergreen taxa are absent in the lowerpart of the core with the exception of a minor peak centered at12.5 ka BP. Both groups are well represented in pollen spectra from10.8 ka BP and upwards. Forestation follows the same trend, whileT-index exhibits in the lower part of the sequence the highestvalues among all studied records.
4.3.2. Terrestrial biomarkersTer-alkanes ARs range from 170 to 3175 ngm�2 y�1 (Fig. 4). They
exhibit low values in sediments deposited during the last glacialand deglaciation periods, (av. 428 ngm�2 y�1), with a large increaseduring S1a (av. 2573 ng m�2 y�1) and S1b (1568 ng m�2 y�1) alongwith a slight drop during the S1 interruption atw8.4 to 8 ka BP (av.1325 ng m�2 y�1). The same trend is observed for the ACL and HPAindices, with higher values within the S1a and S1b and lower onesin the intervals below and above the sapropel (Fig. 4). Fluctuationsin the of ACL and HPA values were observed from 18e16 and 13e11ka BP and appear as lower ACL values along with an increase in theHPA values (Fig. 4).
5. Discussion
The majority (75%) land-delivered material in the Aegean isprovided by its watershed with the remaining being attributed toaeolian transport and to inflow from the Black and Levantine Seasthrough the Dardanelles Strait and the eastern Cretan passages(Poulos, 2009). Black Sea outflow, althoughminor comparing to theriver discharging to the North Aegean Sea (Ehrmann et al., 2007b),started between 8 and 7 ka BP, and reached its maximum at around5e4 ka BP (Sperling et al., 2003), Therefore, terrestrial paly-nomorphs and vascular plant biomarkers are transported to thenorthern Aegean (North Skyros Basin), mainly by fluvial transportand are expected to closely reflect the vegetation in the northernborderlands of the Aegean Sea, including Thessaly, Macedonia,
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036
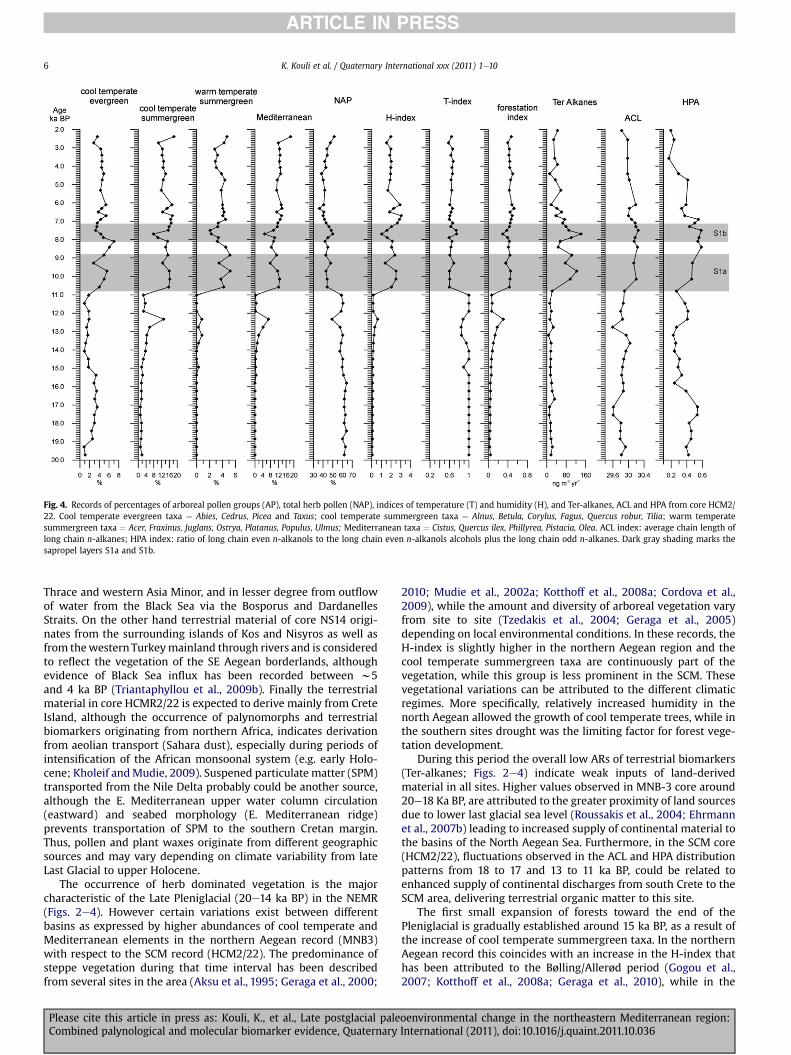
Fig. 4. Records of percentages of arboreal pollen groups (AP), total herb pollen (NAP), indices of temperature (T) and humidity (H), and Ter-alkanes, ACL and HPA from core HCM2/22. Cool temperate evergreen taxa ¼ Abies, Cedrus, Picea and Taxus; cool temperate summergreen taxa ¼ Alnus, Betula, Corylus, Fagus, Quercus robur, Tilia; warm temperatesummergreen taxa ¼ Acer, Fraxinus, Juglans, Ostrya, Platanus, Populus, Ulmus; Mediterranean taxa ¼ Cistus, Quercus ilex, Phillyrea, Pistacia, Olea. ACL index: average chain length oflong chain n-alkanes; HPA index: ratio of long chain even n-alkanols to the long chain even n-alkanols alcohols plus the long chain odd n-alkanes. Dark gray shading marks thesapropel layers S1a and S1b.
K. Kouli et al. / Quaternary International xxx (2011) 1e106
Thrace and western Asia Minor, and in lesser degree from outflowof water from the Black Sea via the Bosporus and DardanellesStraits. On the other hand terrestrial material of core NS14 origi-nates from the surrounding islands of Kos and Nisyros as well asfrom thewestern Turkeymainland through rivers and is consideredto reflect the vegetation of the SE Aegean borderlands, althoughevidence of Black Sea influx has been recorded between w5and 4 ka BP (Triantaphyllou et al., 2009b). Finally the terrestrialmaterial in core HCMR2/22 is expected to derive mainly from CreteIsland, although the occurrence of palynomorphs and terrestrialbiomarkers originating from northern Africa, indicates derivationfrom aeolian transport (Sahara dust), especially during periods ofintensification of the African monsoonal system (e.g. early Holo-cene; Kholeif andMudie, 2009). Suspened particulate matter (SPM)transported from the Nile Delta probably could be another source,although the E. Mediterranean upper water column circulation(eastward) and seabed morphology (E. Mediterranean ridge)prevents transportation of SPM to the southern Cretan margin.Thus, pollen and plant waxes originate from different geographicsources and may vary depending on climate variability from lateLast Glacial to upper Holocene.
The occurrence of herb dominated vegetation is the majorcharacteristic of the Late Pleniglacial (20e14 ka BP) in the NEMR(Figs. 2e4). However certain variations exist between differentbasins as expressed by higher abundances of cool temperate andMediterranean elements in the northern Aegean record (MNB3)with respect to the SCM record (HCM2/22). The predominance ofsteppe vegetation during that time interval has been describedfrom several sites in the area (Aksu et al., 1995; Geraga et al., 2000;
Please cite this article in press as: Kouli, K., et al., Late postglacial paleCombined palynological and molecular biomarker evidence, Quaternary
2010; Mudie et al., 2002a; Kotthoff et al., 2008a; Cordova et al.,2009), while the amount and diversity of arboreal vegetation varyfrom site to site (Tzedakis et al., 2004; Geraga et al., 2005)depending on local environmental conditions. In these records, theH-index is slightly higher in the northern Aegean region and thecool temperate summergreen taxa are continuously part of thevegetation, while this group is less prominent in the SCM. Thesevegetational variations can be attributed to the different climaticregimes. More specifically, relatively increased humidity in thenorth Aegean allowed the growth of cool temperate trees, while inthe southern sites drought was the limiting factor for forest vege-tation development.
During this period the overall low ARs of terrestrial biomarkers(Ter-alkanes; Figs. 2e4) indicate weak inputs of land-derivedmaterial in all sites. Higher values observed in MNB-3 core around20e18 Ka BP, are attributed to the greater proximity of land sourcesdue to lower last glacial sea level (Roussakis et al., 2004; Ehrmannet al., 2007b) leading to increased supply of continental material tothe basins of the North Aegean Sea. Furthermore, in the SCM core(HCM2/22), fluctuations observed in the ACL and HPA distributionpatterns from 18 to 17 and 13 to 11 ka BP, could be related toenhanced supply of continental discharges from south Crete to theSCM area, delivering terrestrial organic matter to this site.
The first small expansion of forests toward the end of thePleniglacial is gradually established around 15 ka BP, as a result ofthe increase of cool temperate summergreen taxa. In the northernAegean record this coincides with an increase in the H-index thathas been attributed to the Bølling/Allerød period (Gogou et al.,2007; Kotthoff et al., 2008a; Geraga et al., 2010), while in the
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036

K. Kouli et al. / Quaternary International xxx (2011) 1e10 7
SCM it is documented by the first appearance of Mediterraneantaxa and the contemporaneous decrease in T-index. The increasingtrend in forestation and humidity is interrupted by several episodesindicating the climatic instability of the Late Glacial/deglaciationperiod. Around 13.2 and 12.1 ka BP the peaks in T-index recorded inthe northern Aegean indicate rapid drops in temperature linkedwith the H1 and YD events (Geraga et al., 2010), that are also re-flected in the pronounced decline of sea surface temperatures (SST;Gogou et al., 2007).
In the SCM (Fig. 4), the short-term disappearance of Mediter-ranean elements at w11.5 ka BP coincides with the temporaryretreat of temperate summergreen taxa, marked decrease in H-index and the shift in T-index, corresponding to the PreborealOscillation (PBO; Bjorck et al., 1997; Magny et al., 2001). A similartrend is observed also in the Aegean regionwhere a short retreat oftemperate summergreen taxa and a reduced H-index are evidencedaround 11 ka BP. In core NS-14 the PBO cooling event has beenrecognized also in coccolithophore and foraminifera assemblages(Triantaphyllou et al., 2009a) and in the Uk
37 SST record(Triantaphyllou et al., 2009b). Quantitative climate reconstructionbased on pollen records from the north Aegean showed that a shortlived cooling event, related to the PBO, interrupts the early Holo-cene warming trend between 11.4 and 10.9 ka BP (Dormoy et al.,2009). This is consistent with the lower values of the ACL indexof n-alkanes.
From w11 ka BP, a major expansion of forest cover is evident inthe NEMR, accompanied with the increase of H-index as a result ofthe climatic amelioration at the onset of the Holocene. Theobserved expansion of temperate summergreen and Mediterra-nean vegetation identifies the shift towards summer wetter andwarmer climatic conditions. Forest expansion appears earlier butestablishes progressively in north Aegean pollen spectra (w11.4 kaBP), in contrast to the delayed and abrupt development in the SCMand the south Aegean (w10 ka BP and w9.5 ka BP respectively).Furthermore forest vegetation type exhibits different characteris-tics with cool temperate taxa being more abundant in the Aegeancores, while in the SCM forestation is less pronounced and mostlyfeatured by Mediterranean elements.
This paleoclimate change follows the Holocene precession-related insolation maximum in Northern hemisphere summerand coincides with the monsoon intensification that resulted ina widespread increase in humidity over the Mediterranean regionand concomitant increase of freshwater input to the MediterraneanSea (e.g. Rossignol-Strick, 1985; Rohling and Hilgen, 1991; Rohling,1999; Kholeif and Mudie, 2009). Concurrently, alkenone-based SSTreconstructions in the investigated cores (Gogou et al., 2007;Triantaphyllou et al., 2009b; Katsouras et al., 2010) reveal a warm-ing trend that reflects the changing regional climatic conditions inwhich the Eastern Mediterranean became also warmer into theHolocene Climatic Optimum. This warm/humid period is associatedwith increased stratification and the development of Deep Chlo-rophyll Maximum in the marine water column (e.g. Castradori,1993; Negri and Giunta, 2001; Triantaphyllou et al., 2009a;2009b, 2010a) and corresponds to the deposition of the S1 sapro-pel (Gogou et al., 2007; Triantaphyllou et al., 2009a, 2009b; 2010b;Katsouras et al., 2010).
The freshening of sea surface waters during S1 deposition,driven by enhanced riverine discharges and land runoff due tointensified rainfall and the contribution of Black Sea outflow after8e7 ka BP (Aksu et al., 1995; 1999; Sperling et al., 2003; Gogouet al., 2007; Ehrmann et al., 2007b), resulted in the increaseddelivery of land-derived material, as indicated by the high accu-mulation of terrestrial biomarkers (Figs. 2e4). Furthermore, shiftsin the ACL index across the late glacial/deglaciation to Holoceneinterglacial transition, are in accordance with the climate-induced
Please cite this article in press as: Kouli, K., et al., Late postglacial paleCombined palynological and molecular biomarker evidence, Quaternary
vegetation change previously deduced from palynological anal-yses. Consistent with the pollen data, a concomitant increase of theACL index in all sites (Figs. 2e4) mark a vegetation shift towards“warmer” species. This trend is more obvious in the southern sites(NS14 and HCM2/22; Figs. 3 and 4) while in the northern Aegeansite (MNB3; Fig. 2), a smoother shift towards higher values for thisindex is observed. The North Aegean Sea receives higher levels ofplant waxes transported by Etesian northerly winds and riverinedischarges from southeastern Europe/Balkans, while in the south-eastern Aegean and the SCM, predominant inputs reflect warmerand drier conditions from local sources and/or North Africa (Gogouet al., 1996). Finally, higher HPA values within the S1 period typifyterrestrial material that has undergone little degradation, due toenhanced riverine inputs that deliver ‘fresh’ land plantmaterial, butalso due to the enhanced stratification/oxygen depletion in themarine water column that promotes in-situ preservation(Versteegh et al., 2010).
During the Holocene, decreases in temperate summergreen andMediterranean taxa and shifts in H and T indices indicate briefvegetation setbacks centered at w8.0, w6.5 and w4.3 ka BP. Thefirst time interval corresponds to the interruption of S1 that hasbeen observed in several records from the Adriatic, Ionian andAegean Seas (e.g. Rohling et al., 2002; Gogou et al., 2007; Geragaet al., 2008; Triantaphyllou et al., 2009a; 2009b). In the northernAegean record (North Skyros basin), alkenone-based SSTs showa pronounced centennial-scale cooling between 8.2 and 7.8 kaduring the interruption of S1 (Gogou et al., 2007). This temperaturedecline described as the expression of the 8.2 ka northern Hemi-sphere global cooling event (Mayewski et al., 1997), has beenassociated with outbreaks of cold and dry polar/continental airmasses from higher latitudes (Rohling et al., 2002; Casford et al.,2003; Rimbu et al., 2004). It has been also concluded froma massive terrestrial ecosystem turnover (Pross et al., 2009) andfluctuations in pollen and d18O values of planktonic foraminifera(Geraga et al., 2010). In the southeastern Aegean, the interval from7.9 to 7.3 ka is characterized by lower humidity values and high seasurface temperatures, therefore it is considered to correlate withaweakening in the Africanmonsoon intensity, rather than to reflecta very cold and dry spell as recorded in the higher latitude locations(Triantaphyllou et al., 2009b). In support of this interpretation,declines of ACL and HPA indices observed during this period arestronger in the north Aegean compared to the south Aegean andthe SCM (Figs. 2e4). Decrease in the ACL values during this timeinstead reflect a shift in biosynthesis of chain lengths to lowercarbon atom numbers due to cooler conditions. Furthermore, thedecrease in the HPA index could be attributed to a shift in thepredominant source of terrestrial inputs related to less intensefreshwater supply and/or an enhancement of mixing conditions inthe water column that led to deeper oxygenation and theconcomitant degradation of organic matter (Gogou et al., 2007).
In addition, at w6.5 ka BP a retreat in temperate summergreenvegetation and reduced H-index values is recorded in both Aegeansites by this study. This event is also characterized by a drop of SSTand marks the termination of the S1 sapropel (Gogou et al., 2007)that is associated with the decline of stratified conditions in thewater column (Triantaphyllou et al., 2009a; b). It also is coeval withmarked coolings described in marine cores from the south Adriaticand south Aegean Seas (Geraga et al., 2000; Sangiorgi et al., 2003;Triantaphyllou et al., 2010a), although not recorded further south,off the Nile Delta (Kholeif andMudie, 2009). Reduced accumulationof terrestrial biomarkers and a shift to lower ACL values in theinvestigated records reflect less humid conditions and “cooler”vegetation type, as also seen in the pollen records. In the SCM thissignal is less pronounced in the pollen record and is expressed asa small decrease of the H-index and the Mediterranean elements.
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036

K. Kouli et al. / Quaternary International xxx (2011) 1e108
However, both terrestrial biomarkers and ACL index show markedshifts that suggest cooler and less humid conditions on land. Thehigh temporal resolution of the southeastern Aegean corecombinedwith the proximity of the coring site to the land results ina more complex and detailed terrestrial signal. The fluctuations inall forest elements and the T-index suggest short-term episodes ofclimatic instability during this time interval associated withenhanced runoff conditions during a wet and cold period from 6.6to 6.4 ka BP (Triantaphyllou et al., 2009b).
The H-index records a conspicuous humidity increase between5.4 and 4.3 ka BP in the southeastern Aegean that coincides with anincrease in the terrestrial biomarker supply. During this interval SSTfluctuates, however it exhibits a sharp positive shift at w4.8 ka BPin associationwith the deposition of a sapropel-like layer (SapropelMid Holocene layer, SMH; Triantaphyllou et al., 2009b; Katsouraset al., 2010). Similar to SST records, T-index is featured bya number of fluctuations that can be attributed to relative increaseof Picea, transported by north wind periodical outbursts (Mudieet al., 2002a), during a generally warm-wet, although instableclimate period (Triantaphyllou et al., 2009a; 2009b). Similar trendsin T and H indices, although a bit delayed and attenuated, areobserved in the northern Aegean MNB-3 record (centered atw4.5 ka BP) and are accompanied by an increase in the ACL index(warmer vegetation type). The low sedimentation rate in the SCMprobably prevents the detection of this event in core HCM2/22.However, a noticeable increase in the accumulation rates ofterrestrial biomarkers and the HPA index values are clearly recor-ded (Fig. 4). The mid-Holocene warm and wet phase providesevidence of on-going, albeit weak, mid Holocene African monsoonforcing in the southeastern Aegean (Triantaphyllou et al., 2009b)that apparently was even more weakened at the north part of theAegean. The end of this humid phase is associated with the 4.2 kaBP Northern Hemisphere mega drought event (Triantaphyllouet al., 2009b), coinciding with a significant Northern Hemisphererapid climate cooling (Mayewski et al., 2004; Migowski et al.,2006), the concomitant reduction of Black Sea surface wateroutflow (Sperling et al., 2003), and the termination of the AfricanHumid Period at 3.8 ka BP (Jung et al., 2004).
6. Conclusions
Changes in the vegetation and vascular plant biomarkers duringdifferent climatic periods within the last 20 ky (Late Glacial anddeglaciation, Holocene Climate Optimum, Mid-Late Holocene)were investigated in three sediment gravity cores collected alonga transect from the north Aegean Sea to the south Cretan margin inthe Northeastern Mediterranean region.
Pollen records show diversified and mixed temperate forest inthe northern borderlands and enhanced Mediterranean vegetationin the southern areas document an NeS climatic trend. This isfurther supported by the increase in the average chain length ofterrestrial n-alkanes (ACL) from the northern Aegean (av. 29.79) tosoutheastern Aegean and south Cretan margin sites (av. 30.06 and30.10, respectively), suggesting a shift in the vegetation cover towarmer-climate species eastwards and southwards.
Relatively increased humidity is recorded during 20e14 ka BP inthe northern Aegean, enabling the growth of cool temperate andMediterranean trees. In contrast, drier climate was the limitingfactor for vegetation development in the southern sites. Thedelivery of land-derived material was found to be most significantin the northern site most likely due to the greater proximity of landsources due to lower last glacial sea level that increased supplyfrom the continental shelf to the basins of the North Aegean Sea.
The first small expansion of forest occurred gradually around15 ka BP, as marked by the increase in pollen of cool temperate
Please cite this article in press as: Kouli, K., et al., Late postglacial paleCombined palynological and molecular biomarker evidence, Quaternary
summergreen taxa. In the northern Aegean record this coincideswith an increase in H-index that has been attributed to the Bølling/Allerød period. Around 13.2 and 12.1 ka BP peaks in T-index indi-cate rapid drops in temperature linked with the H1 and YD events.In the SCM, the short-termed disappearance of Mediterraneanelements at w11.5 ka BP coincides with temporal retreat oftemperate summergreen taxa and marked decrease in H-index,corresponding to the Preboreal Oscillation. A simultaneous reduc-tion in the ACL index in all sites could be indicative of variations inthe amount of terrestrial organic matter delivered and/or changesin the delivery pathways.
After w11 ka BP, a major expansion of forest cover is evidencedin the NEMR, accompanied by the increase of the H-index asa result of the climatic amelioration at the onset of the Holoceneand throughout the Holocene Climatic Optimum. Furthermoreforest vegetation type exhibits different characteristics with cooltemperate taxa being more abundant in the Aegean cores, while inthe SCM forest pollen production is lower and mostly includesMediterranean elements. Enhanced riverine discharges, land runoffand partially Black Sea outflow resulted in the delivery of increasedland-derived material, evidenced by the high accumulations ofterrestrial biomarkers. The transition to higher values of the n-alkane ACL index in the early Holocene indicates changes in thecontinental vegetation towards warm-climate species.
Decrease in the abundances of temperate summergreen andMediterranean taxa and shifts in H and T indices indicate short-term vegetation setbacks, corresponding to a major cooling eventrecognized in all cores between w8.5 to 7.5 ka BP. ACL and HPAindices decrease significantly at this time, especially at the northernAegean site, reflecting cooler prevailing conditions, a shift in thepredominant source of terrestrial inputs and/or enhanced degra-dation of organic matter.
The H-index records a conspicuous humidity increase between5.4 and 4.3 ka BP in the southeastern Aegean that coincides with anincrease in the terrestrial biomarker supply andwith the depositionof a distinct sapropel-like layer (Sapropel Mid Holocene, SMH).Similar trends in T and H indices are observed in the northernAegean record (centered atw4.5 ka BP) and are accompanied by anincrease in the ACL index. In all three cores, warmer/wetterconditions and increased supply of land-derived material isevidenced.
Acknowledgments
We acknowledge financial support by the EC funded HERMES(GOCE-CT-2005-511234-5) and HERMIONE (FP7-ENV-2008-226354) projects, by the ESF funded EUROCORES Euro-CLIMATE(contract No: ERAS-CT-2003-980409), as well as the Pythagoras Iproject (EU and Greek Ministry of Education, EPEAEK II), andPENED 2003 Project (GSRT, Greece; Contract No. 669). The officersand the crew the R/V Aegaeo are gratefully acknowledged for theirimportant and effective contribution to the field work andsampling. Philip Meyers and an anonymous reviewer providedthoughtful reviews which greatly improved the manuscript.
References
Abrajano, T., Aksu, A.E., Hiscott, R.N., Mudie, P.J., 2002. Aspects of carbon isotopebiogeochemistry of late Quaternary sediments from the Marmara Sea and BlackSea. Marine Geology 190 (1), 151e164.
Aksu, A.E., Abrajano, T., Mudie, P.J., Yasar, D., 1999. Organic geochemical and paly-nological evidence for terrigenous origin of the organic matter in the AegeanSea sapropel S1. Marine Geology 153, 303e318.
Aksu, A.E., Yasar, D., Mudie, P.J., Gillespie, H., 1995. Late glacial-Holocene paleo-climatic and paleoceanographic evolution of the Aegean Sea: micropaleonto-logical and stable isotopic evidence. Marine Micropaleontology 25, 1e28.
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036

K. Kouli et al. / Quaternary International xxx (2011) 1e10 9
Bjorck, S., Rundgren, M., Ingolfsson, O., Funder, S., 1997. The Preboreal oscillationaround the Nordic Seas: terrestrial and lacustrine responses. Journal ofQuaternary Science 12, 455e465.
Bouloubassi, I., Lipiatou, E., Saliot, A., Tolosa, I., Bayona, J.M., Albaiges, J., 1997.Carbon sources and cycle in the Western Mediterranean: II. The use ofmolecular markers to determine the origin of organic matter. Deep-SeaResearch II 44, 781e799.
Bouloubassi, I., Rullkotter, J., Meyers, P.A., 1999. Origin and transformation oforganic matter in PlioceneePleistocene Mediterranean sapropels: organicgeochemical evidence reviewed. Marine Geology 153, 177e197.
Cacho, I., Grimalt, J.O., Sierro, F.J., Shackleton, N.J., Canals, M., 2000. Evidence forenhanced Mediterranean thermohaline circulation during rapid climatic cool-ings. Earth and Planetary Science Letters 183, 417e429.
Casford, J.S.L., Abu-Zied, R., Rohling, E.J., Cooke, S., Fontanier, Ch., Leng, M.,Millard, A., Thomson, J., 2007. A stratigraphically controlled multi-proxy chro-nostratigraphy for the eastern Mediterranean. Paleoceanography 22, PA4215.doi:10.1029/2007PA001422.
Casford, J.S.L., Rohling, E.J., Abu-Zied, R., Cooke, S., Fontanier, C., Leng, M.,Lycousis, V., 2002. Circulation changes and nutrient concentrations in the lateQuaternary Aegean Sea: a non-steady state concept for sapropel formation.Paleoceanography 17 (2), 1024e1034.
Casford, J.S.L., Rohling, E.J., Abu-Zied, R.H., Fontanier, C., Jorissen, F.J., Leng, M.J.,Schmiedl, G., Thomson, J., 2003. A dynamic concept for eastern Mediterraneancirculation and oxygenation during sapropel formation. Palaeogeography,Palaeoclimatology, Palaeoecology 190, 103e119.
Castradori, D., 1993. Calcareous nannofossil and the origin of Eastern Mediterraneansapropels. Paleoceanography 8, 459e471.
Combourieu Nebout, N., Peyron, O., Dormoy, I., Desprat, S., Beaudouin, C.,Kotthoff, U., Marret, F., 2009. Rapid climatic variability in the west Mediterra-nean during the last 25,000 years from high resolution pollen data. Climate ofthe Past 5, 503e521.
Cordova, C., Harrison, S.P., Mudie, P.J., Riehl, S., Leroy, S.A.G., Ortiz, N., 2009. Pollen,plant macrofossil and charcoal records for palaeovegetation reconstruction inthe Mediterranean-BlackSea Corridor since the Last Glacial Maximum.Quaternary International 197, 12e26.
Dormoy, I., Peyron, O., Combourieu-Nebut, N., Goring, S., Kotthoff, U., Magny, M.,Pross, J., 2009. Terrestrial climate variability and seasonality changes in theMediterranean region between 15,000 and 4000 BP deduced from marinepollen records. Climate of the Past 5, 615e632.
Eglinton, T.I., Eglinton, G., 2008. Molecular proxies for paleoclimatology. Earth andPlanetary Science Letters 275, 1e16.
Eglinton, G., Hamilton, R.J., 1967. Leaf epicutilar waxes. Science 256, 1322e1335.Ehrmann, W., Schmiedl, G., Hamann, Y., Kuhnt, T., 2007a. Distribution of clay
minerals in surface sediments of the Aegean Sea: a compilation. InternationalJournal of Earth Sciences 96, 769e780.
Ehrmann, W., Schmiedl, G., Hamann, Y., Kuhnt, T., Hemleben, C., Siebel, W., 2007b.Clay minerals in late glacial and Holocene sediments of the northern andsouthern Aegean Sea. Palaeogeography, Palaeoclimatology, Palaeoecology 249,36e57.
Emeis, K.-C., Schulz, H., Struck, U., Rossignol-Strick, M., Erlenkeuser, H.,Howell, M.W., Kroon, D., Mackensen, A., Ishizuka, S., Oba, T., Sakamoto, T.,Koizumi, I., 2003. Eastern Mediterranean surface water temperatures and d18Ocomposition during deposition of sapropels in the late Quaternary. Paleo-ceanography 18, 5-1e5-18.
Geraga, M., Mylona, G., Tsaila-Monopoli, St., Papatheodorou, G., Ferentinos, G.,2008. Northeastern Ionian Sea: palaeoceanographic variability over the last 22ka. Journal of Marine Systems 74, 623e638.
Geraga, M., Chr., Ioakim, Lykousis, V., Tsaila Monopolis StMylona, G., 2010. The high-resolution palaeoclimatic and palaeoceanographic history of the last 24,000years in the central Aegean Sea, Greece. Palaeogeography, Palaeoclimatology,Palaeoecology 287, 101e115.
Geraga, M., Tsaila-Monopoli, St., Ioakim, Ch., Papatheodorou, G., Ferentinos, G.,2000. An evaluation of paleoenvironmental changes during the last 18000 yr BPin the Myrtoon Basin, S.W. Aegean Sea. Palaeogeography, Palaeoclimatology,Palaeoecology 156, 1e17.
Geraga, M., Tsaila-Monopolis, S., Ioakim, C., Papatheodorou, G., Ferentinos, G., 2005.Short-term climate changes in the southern Aegean Sea over the last 48,000years. Palaeogeography, Palaeoclimatology, Palaeoecology 220, 311e332.
Gogou, A., Bouloubassi, I., Lykousis, V., Arnaboldi, M., Gaitani, P., Meyers, P.A., 2007.Organic geochemical evidence of late glacial-Holocene climate instability in thenorth Aegean sea. Palaeogeography, Palaeoclimatology, Palaeoecology 256,1e20.
Gogou, A., Bouloubassi, I., Stephanou, E.G., 2000. Marine organic geochemistry ofthe Eastern Mediterranean: 1. Aliphatic and polyaromatic hydrocarbons inCretan Sea surficial sediments. Marine Chemistry 68, 265e282.
Gogou, A., Stratigakis, N., Kanakidou, M., Stephanou, E.G., 1996. Organic aerosols inEastern Mediterranean: components source reconciliation by using molecularmarkers and atmospheric back trajectories. Organic Geochemistry 25, 79e96.
Gogou, A., Stephanou, E.G., 2004. Marine organic geochemistry of the EasternMediterranean: 2. Polar biomarkers in Cretan Sea surficial sediments. MarineChemistry 85, 1e25.
Hamann, Y., Ehrlmann, W., Schmiedl, G., Krüger, S., Stuut, J.B., Kuhnt, T., 2008.Sedimentation processes in the Eastern Mediterranean Sea during the lateGlacial and Holocene revealed by end-member modelling of the terrigenousfraction in marine sediments. Marine Geology 248, 97e114.
Please cite this article in press as: Kouli, K., et al., Late postglacial paleCombined palynological and molecular biomarker evidence, Quaternary
Ioakim, C., Triantaphyllou, Tsaila-Monopolis S., Geraga, M., Dimiza, M., Lykousis, V.,2009. New micropalaeontological records of eastern Mediterranean marinesequences recovered offshore of Crete, during HERMES cruise and theirpalaeoclimatic-palaeoceanographic significance. In: Abstract in: 13th RCMNSCongress, Naples, Italy 2e6 September 2009.
Jung, S.J.A., Davies, G.R., Ganssen, G.M., Kroom, D., 2004. Stepwise Holocene aridi-fication in NE Africa deduced from dust-borne radiogenic isotope records. Earthand Planetary Science Letters 7031, 1e11.
Katsouras, G., Gogou, A., Bouloubassi, I., Emeis, K.-C., Triantaphyllou, M.,Roussakis, G., Lykousis, V., 2010. Organic carbon distribution and isotopiccomposition in three records from the eastern Mediterranean Sea during theHolocene. Organic Geochemistry 41, 935e939.
Kholeif, S.E.A., Mudie, P.J., 2009. Palynological records of climate and oceanicconditions in the late Pleistocene and Holocene of the Nile Cone, southeasternMediterranean, Egypt. Palynology 33, 1e24.
Kotthoff, U., Müller, U.C., Pross, J., Schmiedl, G., Lawson, I.T., Van DeSchootbrugge, B., Schulz, H., 2008a. Lateglacial and Holocene vegetationdynamics in the Aegean region: an integrated view based on pollen data frommarine and terrestrial archives. The Holocene 18, 1019e1032.
Kotthoff, U., Pross, J., Müller, U.C., Peyron, O., Schmiedl, G., Schulz, H., Bordon, A.,2008b. Climate dynamics in the borderlands of the Aegean Sea during forma-tion of sapropel S1 deduced from a marine pollen record. Quaternary ScienceReviews 27, 832e845.
Lawson, I.T., Frogley, M.R., Bryant, C., Preece, R.C., Tzedakis, P.C., 2004. The Late-glacial and Holocene environmental history of the Ioannina basin, north-westGreece. Quaternary Science Reviews 23, 1599e1625.
Lionello, P., Malanotte-Rizzoli, P., Boscolo, R., Alpert, P., Artale, V., Li, L.,Luterbacher, J., May, W., Trigo, R., Simplis, M.T., Ulbrich, U., Xoplaki, E., 2006. TheMediterranean climate: an overview of the main characteristics and issues. In:Lionello, P., Malanotte-Rizzoli, P., Boscolo, R. (Eds.), Mediterranean ClimateVariability. Elsevier, Amsterdam, pp. 1e26.
Lykousis, V., Chronis, G., Tselepides, A., Price, N.B., Theocharis, A., Siokou-Fragou, I.,van Wambeke, F., Danovaro, R., Stavrakakis, S., Duineveld, G., Georgopoulos, D.,Ignatiades, L., Souvermezoglou, A., Voutsinou-Taliadouri, F., 2002. Majoroutputs of the recent multidisciplinary biogeochemical researches undertakenin the Aegean Sea. Journal of Marine Systems 33e34, 313e334.
Magny, M., Bégeot, C., Guiot, J., Marguet, A., Billaud, Y., 2003. Reconstruction andpalaeoclimatic interpretation of mid-Holocene vegetation and lake-level changesat Saint-Jorioz, Lake Annecy, French Pre-Alps. The Holocene 13, 265e275.
Magny, M., Guiot, J., Schoellammer, P., 2001. Quantitative reconstruction of YoungerDryas to mid-Holocene paleoclimates at Le Locle, Swiss Jura, using pollen andlake-level data. Quaternary Research 56, 170e180.
Marino, G., Rohling, E.J., Sangiorgi, F., Hayes, A., Casford, J.L., Lotter, A.F., Kucera, M.,Brinkhuis, H., 2009. Early and middle Holocene in the Aegean Sea: interplaybetween high and low latitude climate variability. Quaternary Science Reviews28, 3246e3262.
Mayewski, P.A., Meeker, L.D., Twickler, M.S., Whitlow, S., Yang, Q., Lyons, W.B.,Prentice, M., 1997. Major features and forcing of high-latitude northern hemi-sphere atmospheric circulation using a 110,000-year long glaciochemical series.Journal of Geophysical Research 102, 26345e26366.
Mayewski, P.A., Rohling, E., Stager, C.J., Karlen, W., Maasch, K.A., Meeker, L.D.,Meyerson, E.A., Gasse, F., van Kreveld, S., Holmgren, K., Lee-Thorp, J.,Rosqvist, G., Rack, F., Staubwasser, M., Schneider, R.R., Steig, E., 2004. Holoceneclimate variability. Quaternary Research 62, 243e255.
Myers, P.G., Haines, K., Rohling, E.J., 1998. Modeling the paleocirculation of theMediterranean: the last glacial maximum and the Holocene with emphasis onthe formation of sapropel S1. Paleoceanography 13, 586e606.
Migowski, C., Mordechai, S., Prasad, S., Negendank, J.F.W., Agnon, A., 2006. Holoceneclimate variability and cultural evolution in the Near East from the Dead Seasedimentary record. Quaternary Research 66, 421e431.
Mudie, P.J., Rochon, A., Aksu, A.E., Gillespie, H., 2002a. Pollen stratigraphy of LateQuaternary cores from Marmara Sea: land-sea correlation and paleoclimatichistory. Marine Geology 190, 233e260.
Mudie, P.J., Rochon, A., Aksu, A.E., Gillespie, H., 2002b. Dinoflagellate cysts, fresh-water algae and fungal spores as salinity indicators in Late Quaternary coresfrom Marmara and Black seas. Marine Geology 190, 203e231.
Negri, A., Giunta, S., 2001. Calcareous nannofossil paleoecology in the sapropel S1 ofthe Eastern Ionian sea: paleoceanographic implications. Palaeogeography,Palaeoclimatology, Palaeoecology 169, 101e112.
Pancost, R.D., Boot, C.S., 2004. The palaeoclimatic utility of terrestrial biomarkers inmarine sediments. Marine Chemistry 92, 239e261.
Papanikolaou, D., Alexandri, M., Nomikou, P., Ballas, D., 2002. Morphotectonicstructure of the North Aegean Basin based on swath bathymetry. MarineGeology 190, 465e492.
Poulos, S.E., 2009. Origin and distribution of theterrigenous component of theunconsolidated surface sediment of the Aegean floor: a synthesis. ContinentalShelf Research 29, 2045e2060.
Poulos, S.E., Drakopoulos, P.G., Collins, M.B., 1997. Seasonal variability in sea surfaceoceanographic conditions in the Aegean Sea (Eastern Mediterranean): anoverview. Journal of Marine Systems 13 (1e4), 225e244.
Poynter, J.G., Eglinton, G., 1990. Molecular composition of three sediments fromHole 717C: the Bengal Fan. Proceedings of the Ocean Drilling Program, ScientificResults 116, 155e161.
Poynter, J.G., Farrimond, P., Robinson, N., Eglinton, G., 1989. Aeolian derived higherplants in the marine sedimentary record: links with palaeoclimate. In:
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036

K. Kouli et al. / Quaternary International xxx (2011) 1e1010
Leinen, M., Sarntheim, M. (Eds.), Palaeoclimatology and Palaeometeorology:Modern and Past Patterns of Global Atmospheric Transport, pp. 435e462.
Pross, J., Kotthoff, U., Muller, U.C., Peyron, O., Dormoy, I., Schmiedl, G., Kalaitzidis, S.,Smith, A.M., 2009. Massive perturbation in terrestrial ecosystems of the EasternMediterranean region associated with the 8.2 kyr B.P. climatic event. Geology37 (10), 887e890.
Rimbu, N., Lohmann, G., Lorenze, S.J., Kim, J.H., Schneider, R.R., 2004. Holoceneclimate variability as derived from alkenone sea surface temperature andcoupled oceaneatmosphere model experiments. Climate Dynamics 23,215e227.
Rohling, E.J., 1999. Environmental control on Mediterranean salinity and d18O.Paleoceanography 14, 706e715.
Rohling, E.J., Hilgen, F.J., 1991. The eastern Mediterranean climate at times of sap-ropel formation: a review. Geologie en Mijnbouw 70, 253e264.
Rohling, E.J., Mayewski, P.A., Abu-Zied, R.H., Casford, J.S.L., Hayes, A., 2002. Holoceneatmosphere-ocean interactions: records from Greenland and the Aegean Sea.Climate Dynamics 18, 587e593.
Rossignol-Strick, M., 1985. Mediterranean Quaternary sapropels, an immediateresponse of the African monsoon to variation of insolation. Palaeogeography,Palaeoclimatology, Palaeoecology 49, 237e263.
Rossignol-Strick, M., 1999. The Holocene climatic optimum and pollen records ofsapropel 1 in the eastern Mediterranean, 9000e6000 BP. Quaternary ScienceReviews 18, 515e530.
Roussakis, G., Karageorgis, A.P., Conispoliatis, N., Lykousis, V., 2004. Last glacial-Holocene sediment sequences in N. Aegean basins: structure, accumulationrates and clay mineral distribution. Geo-Marine Letters 24, 97e111.
Sangiorgi, F., Capotondi, L., Nebout, N.C., Vigkiotti, L., Brinkhuis, H., Giounta, S.,Lotter, A.F., Morigi, C., Negri, A., Reichart, G.-J., 2003. Holocene seasonal sea-surface temperature variation in the southern Adriatic Sea inferred froma multiproxy approach. Journal of Quaternary Science 18, 723e732.
Schmiedl, G., Kuhnt, T., Ehrmann, W., Emeis, K.-C., Hamann, Y., Kotthoff, U.,Dulski, P., Pross, J., 2010. Climatic forcing of eastern Mediterranean deep-waterformation and benthic ecosystems during the past 22,000 years. QuaternaryScience Reviews 29, 3006e3020.
Sicre, M.-A., Labeyrie, L., Ezat, U., Mazaud, A., Turon, J.-L., 2006. A 27 kyr terrestrialbiomarker record in the southern Indian Ocean. Geochemistry, Geophysics,Geosystems 7, Q07014. doi:10.1029/2005GC001234.
Sperling, M., Schmiedl, G., Hemleben, Ch., Emeis, K.C., Erlenkeuser, H., Grootes, P.M.,2003. Black Sea impact on the formation of eastern Mediterranean sapropel S1?
Please cite this article in press as: Kouli, K., et al., Late postglacial paleCombined palynological and molecular biomarker evidence, Quaternary
Evidence from the Marmara sea. Palaeogeography, Palaeoclimatology, Palae-oecology 190, 9e21.
Stratford, K., Williams, R.G., Myers, P.G., 2000. Impact of the circulation on sapropelformation in the eastern Mediterranean. Global Biogeochemical Cycles 14,683e695.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen, K.A., Kromer, B.,McCormac, G., Van Der Plicht, J., Spurk, M., 1998. INTCAL98 radiocarbon agecalibration, 24,000e0 cal BP. Radiocarbon 40, 1041e1083.
Triantaphyllou, M.V., Antonarakou, A., Dimiza, M., Anagnostou, Ch, 2010a. Calcar-eous nannofossil and planktonic foraminiferal distributional patterns duringdeposition of sapropels S6, S5 and S1 in the Libyan Sea (Eastern Mediterra-nean). Geo-Marine Letters 30, 1e13.
Triantaphyllou, M.V., Antonarakou, A., Kouli, K., Dimiza, M., Kontakiotis, G.,Papanikolaou, M., Ziveri, P., Mortyn, G., Lianou, V., Lykousis, V.,Dermitzakis, M.D., 2009a. Comparing late glacial e Holocene Plankton ecozonesand pollen assemblage zones: basis for a multi-proxy ecostratigraphy in thesouth - eastern Aegean Sea (E. Mediterranean). Geo-Marine Letters 29,249e267.
Triantaphyllou, M.V., Kouli, K., Tsourou, T., Koukousioura, O., Pavlopoulos, K.,Dermitzakis, M.D., 2010b. Paleoenvironmental changes since 3000 BC in thecoastal marsh of Vravron (Attiki, SE Greece). Quaternary International 216,14e22.
Triantaphyllou, M.V., Ziveri, P., Gogou, A., Marino, G., Lykousis, V., Bouloubassi, I.,Emeis, K.C., Kouli, K., Dimiza, M., Rosell-Mele, A., Papanikolaou, M.,Katsouras, G., Nunez, N., 2009b. Late Glacial-Holocene climate variability at thesouth-eastern margin of the Aegean Sea. Marine Geology 266 (1e4), 182e197.
Tzedakis, P.C., 1999. The last climatic cycle at Kopais, central Greece. Journal of theGeological Society, London 156, 425e434.
Tzedakis, P.C., 2000. Vegetation variability in Greece during the last interglacial.Geologie en Mijnbouw/Netherlands. Journal of Geosciences 79 (2/3), 355e367.
Tzedakis, P.C., Frogley, M.R., Heaton, T.H.E., 2002. Duration of last interglacialconditions in Northwest Greece. Quaternary Research 58, 53e55.
Tzedakis, P.C., Frogley, M.R., Lawson, I.T., Preece, R.C., Cacho, I., De Abreu, L., 2004.Ecological thresholds and patterns of millennial-scale climate variability: theresponse of vegetation in Greece during the last glacial period. Geology 32 (2),109e112.
Versteegh, G.J.M., Zonneveld, K.A.F., de Lange, G.J., 2010. Selective aerobic andanaerobic degradation of lipids and palynomorphs in the Eastern Mediterra-nean since the onset of sapropel S1 deposition. Marine Geology 278, 177e192.
oenvironmental change in the northeastern Mediterranean region:International (2011), doi:10.1016/j.quaint.2011.10.036















![Palynological survey in honeys of Bahia (Brazil) [Original title: Prospecção palinológica em méis da Bahia]](https://static.fdokumen.com/doc/165x107/6341f6b0e8d6d6284d07e1db/palynological-survey-in-honeys-of-bahia-brazil-original-title-prospeccao-palinologica.jpg)




