
The first Pliocene mammals from the northeast (Mesopotamia) of Argentina: Biostratigraphic and...
8
ARTICLE THE FIRST PLIOCENE MAMMALS FROM THE NORTHEAST (MESOPOTAMIA) OF ARGENTINA: BIOSTRATIGRAPHIC AND PALEOENVIRONMENTAL SIGNIFICANCE ADRIANA M. CANDELA, *,1 JORGE I. NORIEGA, 2 and MARCELO A. REGUERO 1 1 Departamento Científico Paleontología Vertebrados, Museo de La Plata, Paseo del Bosque s/n, (1900) La Plata, Argentina; CONICET; [email protected]; 2 Laboratorio de Paleontología de Vertebrados, CICYTTP- CONICET, Matteri y España, (3105) Diamante, Argentina; [email protected] ABSTRACT—The record of Neogene faunas from the northeast of Argentina (Mesopotamia) is sporadic, whereas the late Miocene vertebrate-bearing horizons in this area contain a very abundant and diverse fauna, the Pliocene deposits are poorly known and have been unfossiliferous until now. This situation makes the understanding of the biogeographic and evolutionary history of the Mesopotamian taxa difficult. In this contribution, the first mammals are described that were recovered from the most basal levels of the Punta Gorda Group ( Alvear Formation) at Entre Ríos Province, assigned to the late Pliocene (Lower Matuyama) by previous paleomagnetic studies. These beds are paleomagnetically bracketed between 1.95 and 2.6 Ma (late Pliocene) in the Alvear section and help to refine the biostratigraphy of these species elsewhere in northeastern Argentina. The echimyids Paramyocastor diligens (Ameghino, 1888) and Eumysops sp., and the hegetotheriid Paedotherium cf. typicum herein reported support a Pliocene age for the sediments from which they were recovered. These mammals suggest a paleoenvironment characterized by semiarid and open lowlands, markedly different than that inferred for the Late Miocene in this area. INTRODUCTION The continental Neogene and Quaternary vertebrates of the Mesopotamia region in Northeast of Argentina (Fig. 1) are very diverse and abundant. They come mainly from the “Con- glomerado osífero” ( “Mesopotamiense”, lower levels of the Ituzaingó Formation, Late Miocene; Cione et al., 2000), and the Arroyo Feliciano Formation (Late Pleistocene, Noriega et al., 2004). Conversely, Pliocene vertebrates were unknown in this area until now, making the reconstruction of the biogeographic and evolutionary history of the Mesopotamian taxa difficult. In this contribution, we describe the first mammals from the Pliocene of Mesopotamia coming from the most basal levels of the Punta Gorda Group (Alvear Formation; Iriondo, 1980), at Diamante Department, Entre Ríos Province (Fig. 1). They are represented by the echimyid rodents Paramyocastor diligens (Ameghino, 1888) and Eumysops sp., and the hegetotheriid notoungulate Paedotherium cf. typicum. These taxa indicate a Pliocene age for the horizons bearing them. This appears in agreement with the previous assignment of the Alvear Forma- tion to the Lower Matuyama (Late Pliocene) period (Bidegain, 1991). In addition, these mammals suggest a paleoenvironmental scenario of open lowlands under dry and arid climatic conditions, different from the very humid habitats proposed for the Late Miocene in this area (Cione et al., 2000). Institutional Abbreviations—CICYTTP, Centro de Inves- tigaciones Científicas y Transferencia de Tecnología a la Pro- ducción de Diamante, Argentina; MLP, Museo de La Plata, Argentina; MACN, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia,” Argentina. Anatomical Abbreviations—DAP, anteroposterior diameter; DTA, transverse anterior diameter; DTP, transverse posterior diameter. Dental terminology follows Candela (2002). GEOLOGY AND CHRONOLOGY The fossils were collected in sediments of units 2 and 3 of the profile described by Vucetich and colleagues (2005), which is summarized here (Fig. 2). 1. Grey and light green clays, with abundant reddish limonite spots, separate from the overlying greenish clayey-sandy silts by calcareous levels. They belong to the Paraná Formation; 6.5–7 m thick. 2. Light grayish green silts with high calcium carbonate content in discontinuous horizontal and vertical veins, forming a net- work, with abundant MnO 2 nodules. They correspond to the lower part of the Punta Gorda Group ( Alvear Formation; although with this name, this unit is more typically repre- sented somewhat farther north of Diamante); 2–3 m. 3. Conglomerate of calcium carbonate pebbles derived from the erosion of the preceding unit, cemented by a carbonate matrix ( Alvear Formation). This well-defined and strong carbon- ate floor forms a discomformity as a ridge on the profile of the cliff; 0.6–1 m. 4. Clayey silt, partly sandy, light brown to pink and reddish brown; with loessoid and massive sections, forming a slope of approximately 50 to 60° of rounded or somewhat conic ero- sion microforms. It has MnO 2 and gypsum nodules. It inter- calates with other sections with more vertical slope composed of lenses with cross-bedding or poorly defined conglomeratic levels of small calcium carbonate clasts in coarse sand matrix. This unit corresponds to the upper part of the Punta Gorda * Corresponding author. Journal of Vertebrate Paleontology 27(2):476–483, June 2007 © 2007 by the Society of Vertebrate Paleontology 476
Transcript of The first Pliocene mammals from the northeast (Mesopotamia) of Argentina: Biostratigraphic and...

ARTICLE
THE FIRST PLIOCENE MAMMALS FROM THE NORTHEAST (MESOPOTAMIA) OFARGENTINA: BIOSTRATIGRAPHIC AND PALEOENVIRONMENTAL SIGNIFICANCE
ADRIANA M. CANDELA,*,1 JORGE I. NORIEGA,2 and MARCELO A. REGUERO1
1Departamento Científico Paleontología Vertebrados, Museo de La Plata, Paseo del Bosque s/n, (1900) La Plata, Argentina;CONICET; [email protected];
2Laboratorio de Paleontología de Vertebrados, CICYTTP- CONICET, Matteri y España, (3105) Diamante, Argentina;[email protected]
ABSTRACT—The record of Neogene faunas from the northeast of Argentina (Mesopotamia) is sporadic, whereas thelate Miocene vertebrate-bearing horizons in this area contain a very abundant and diverse fauna, the Pliocene depositsare poorly known and have been unfossiliferous until now. This situation makes the understanding of the biogeographicand evolutionary history of the Mesopotamian taxa difficult. In this contribution, the first mammals are described thatwere recovered from the most basal levels of the Punta Gorda Group (� Alvear Formation) at Entre Ríos Province,assigned to the late Pliocene (Lower Matuyama) by previous paleomagnetic studies. These beds are paleomagneticallybracketed between 1.95 and 2.6 Ma (late Pliocene) in the Alvear section and help to refine the biostratigraphy of thesespecies elsewhere in northeastern Argentina. The echimyids Paramyocastor diligens (Ameghino, 1888) and Eumysops sp.,and the hegetotheriid Paedotherium cf. typicum herein reported support a Pliocene age for the sediments from which theywere recovered. These mammals suggest a paleoenvironment characterized by semiarid and open lowlands, markedlydifferent than that inferred for the Late Miocene in this area.
INTRODUCTION
The continental Neogene and Quaternary vertebrates of theMesopotamia region in Northeast of Argentina (Fig. 1) arevery diverse and abundant. They come mainly from the “Con-glomerado osífero” (� “Mesopotamiense”, lower levels of theItuzaingó Formation, Late Miocene; Cione et al., 2000), and theArroyo Feliciano Formation (Late Pleistocene, Noriega et al.,2004). Conversely, Pliocene vertebrates were unknown in thisarea until now, making the reconstruction of the biogeographicand evolutionary history of the Mesopotamian taxa difficult.
In this contribution, we describe the first mammals from thePliocene of Mesopotamia coming from the most basal levels ofthe Punta Gorda Group (�Alvear Formation; Iriondo, 1980), atDiamante Department, Entre Ríos Province (Fig. 1). They arerepresented by the echimyid rodents Paramyocastor diligens(Ameghino, 1888) and Eumysops sp., and the hegetotheriidnotoungulate Paedotherium cf. typicum. These taxa indicate aPliocene age for the horizons bearing them. This appears inagreement with the previous assignment of the Alvear Forma-tion to the Lower Matuyama (Late Pliocene) period (Bidegain,1991). In addition, these mammals suggest a paleoenvironmentalscenario of open lowlands under dry and arid climatic conditions,different from the very humid habitats proposed for the LateMiocene in this area (Cione et al., 2000).
Institutional Abbreviations—CICYTTP, Centro de Inves-tigaciones Científicas y Transferencia de Tecnología a la Pro-ducción de Diamante, Argentina; MLP, Museo de La Plata,Argentina; MACN, Museo Argentino de Ciencias Naturales“Bernardino Rivadavia,” Argentina.
Anatomical Abbreviations—DAP, anteroposterior diameter;DTA, transverse anterior diameter; DTP, transverse posteriordiameter. Dental terminology follows Candela (2002).
GEOLOGY AND CHRONOLOGY
The fossils were collected in sediments of units 2 and 3 of theprofile described by Vucetich and colleagues (2005), which issummarized here (Fig. 2).
1. Grey and light green clays, with abundant reddish limonitespots, separate from the overlying greenish clayey-sandy siltsby calcareous levels. They belong to the Paraná Formation;6.5–7 m thick.
2. Light grayish green silts with high calcium carbonate contentin discontinuous horizontal and vertical veins, forming a net-work, with abundant MnO2 nodules. They correspond to thelower part of the Punta Gorda Group (� Alvear Formation;although with this name, this unit is more typically repre-sented somewhat farther north of Diamante); 2–3 m.
3. Conglomerate of calcium carbonate pebbles derived from theerosion of the preceding unit, cemented by a carbonate matrix(� Alvear Formation). This well-defined and strong carbon-ate floor forms a discomformity as a ridge on the profile of thecliff; 0.6–1 m.
4. Clayey silt, partly sandy, light brown to pink and reddishbrown; with loessoid and massive sections, forming a slope ofapproximately 50 to 60° of rounded or somewhat conic ero-sion microforms. It has MnO2 and gypsum nodules. It inter-calates with other sections with more vertical slope composedof lenses with cross-bedding or poorly defined conglomeraticlevels of small calcium carbonate clasts in coarse sand matrix.This unit corresponds to the upper part of the Punta Gorda*Corresponding author.
Journal of Vertebrate Paleontology 27(2):476–483, June 2007© 2007 by the Society of Vertebrate Paleontology
476
JVP Reprint Policy
SVP policy on distribution of the author's pdf file gives the following rights to the author:(1) Subject to payment of the required, one-time, non-refundable fee as determined by the Society, the right to post the unaltered pdf file as provided by JVP on a single, author-controlled, private or institutional web site for public access.(2) Without fee payment, the right to include on the author's Web page (a) a link to the JVP web site where members and subscribers may download a pdf copy of the article; (b) a link whereby interested persons may email the author and request a pdf copy for their own personal use but not for re-distribution.(3) Without fee payment, the right to send a copy of the article in pdf form to persons on their reprint mailing list, and to persons who request a copy as per (2) above, provided that the pdf is accompanied by the clear statement that: (a) Society of Vertebrate Paleontology is the copyright holder of the article and pdf; (b) the pdf is provided to the recipient for her/his personal use only and is not to be redistributed or disseminated, except for educational use within a school, college, or university setting, and then only if accompanied by a clear statement of these conditions. Such an author-distributed version must be identical to the final published version.

Group (� Hernandarias Formation, which is more typicallyrecognized north of Paraná City); 4.5–6 m.
5. Overlying discontinuously, clayey loessoid silts, massive anddusty, very light brown to yellowish, with abundant sub-spheric or ramified calcium carbonate scattered in the matrix(Tezanos Pinto Formation); 1.5–2 m.
6. Developed soil in unit 5; 1 m.
The stratigraphic sequence in the area of Diamante Depart-ment begins at the marine and/or estuarine levels of the ParanáFormation, regarded as Tortonian in age (Aceñolaza, 2000;Cione et al., 2000). Over them, the oldest “Pampean” formationscropping out at the SW of the Entre Ríos province overlie themdiscontinuously (Iriondo, 1980, 1996; Bidegain, 1991). They aremainly composed of loessoid silts and calcretes that are consid-ered part of the Punta Gorda Group, divisible in major units ofregional significance as the Alvear and the Hernandarias For-mations (Iriondo 1980: 130); nevertheless, Iriondo (1996) distin-guished the Hernandarias Formation as a sedimentary unit de-posited later than the Punta Gorda Group, around 1.3-0.8 Ma.Later, Iriondo (1998) assigned the Alvear Formation to the earlyPleistocene based on its stratigraphic position. The deposition ofthe latter would have been mainly aeolian in lacustrine or pal-ustrine environments, probably in a semiarid climate (Iriondo,1980: 139).
On the other hand, Bidegain (1991) stated that the Hernan-darias Formation was deposited during the Olduvai Event(ca.1.77-1.95 Ma; Fig. 3), and that the levels regarded by Iriondo(1980) as Alvear Formation would represent the zone recognizedas Lower Matuyama (around 1.95-2.6 Ma; Fig. 3). Thus, theAlvear Formation and the correlated units 2 and 3 of the PuntaGorda Group (see description of the profile; Fig. 2) may beregarded as late Pliocene in age.
Paleomagnetic studies of one section (Alvear) to the north ofPunta Gorda (Bidegain, 1991) suggest a correlation of ChronC2n-C2r for the lowest levels. This places the correlated units 2
and 3 of the Punta Gorda Group at approximately 1.95–2.6 Main the late Pliocene.
SYSTEMATIC PALEONTOLOGY
Order RODENTIA Bowdich, 1821Suborder HYSTRICOGNATHI Tullberg, 1899
Superfamily OCTODONTOIDEA Waterhouse, 1839Family ECHIMYIDAE Gray, 1825
Genus PARAMYOCASTOR Ameghino, 1904PARAMYOCASTOR DILIGENS (Ameghino, 1888)
Material—CICYTTP-PV-M-1-96, right m3; CICYTTP-PV-M-1-97, right M1?.
Geographical and Stratigraphic Occurrence—Materialscome from the upper cliff in the south margin of the large valleyof La Ensenada stream, 300 m north of its outlet into the Paranáriver, Diamante Department, Entre Ríos Province, Argentina(32°40’S- 60°38’W). This cliff rises along the coastal road fromDiamante harbor to the village of Strobel (Fig. 1). Fossil-bearingsediments correspond to the basal levels of the Punta GordaGroup, Late Pliocene.
Measurements—CICYTTP-PV-M-1-96, right m3, DAP: 5.55mm, DTA: 4.05 mm, DTP: 4.01 mm. CICYTTP-PV-M-1-97, rightM1?, DAP: 4. 21 mm, TD: 4.62 mm.
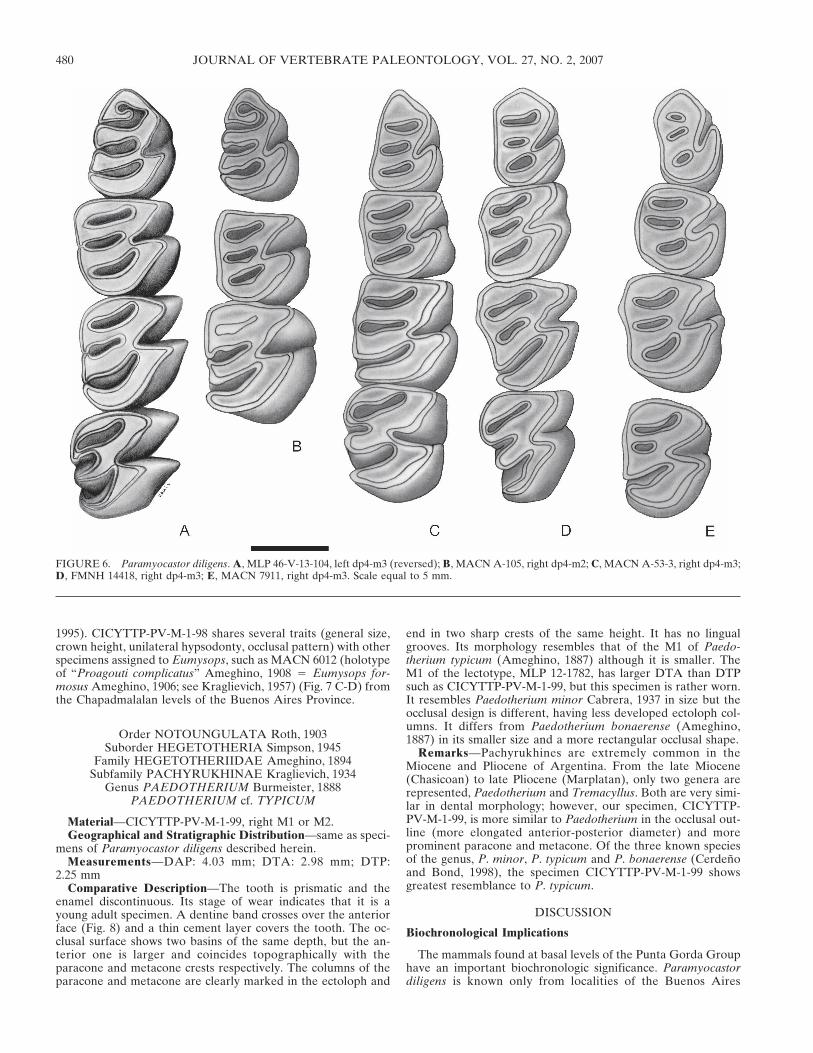
Comparative Description—The upper molar (CICYTTP-PV-M-1-97, Fig. 4 A–B) is tetralophodont, protohypsodont, with uni-lateral hypsodonty. Its stage of wear is heavier than the M1 ofFMNHN 14418 but less than the M1 of the specimens MACN7912 and MACN 7344 of this species (Fig. 4 C-E). Labial flexi
FIGURE 1. Location map of the study area, Diamante Department,Entre Ríos Province; shaded area shows the Mesopotamia region inArgentina, asterisk indicates fossiliferous locality.
FIGURE 2. Stratigraphic profile of the Punta Gorda Group at thefossiliferous locality. Modified from Vucetich et al. (2005).
CANDELA ET AL.—FIRST PLIOCENE MAMMALS FROM THE NORTHEAST OF ARGENTINA 477

have become fossettes; the lingual end of the hypoflexus, al-though almost closed, maintains continuity with the protoconeand hypocone. Thus, the hypoflexus is not yet transformed intoa hypofossette, such as in MACN 7912 and MACN 7344. Thebottom of the hypoflexus is oriented towards the bottom of theparaflexus. The metafossette is larger than the parafossette, atypical pattern found in M1 of this species, in which the hypo-
fossette is already evident. Based on the general size of the molarand on the position and relative size of the hypoflexi and labialfossettes, it is likely that CICYTTP-PV-M-1-97 is an M1.
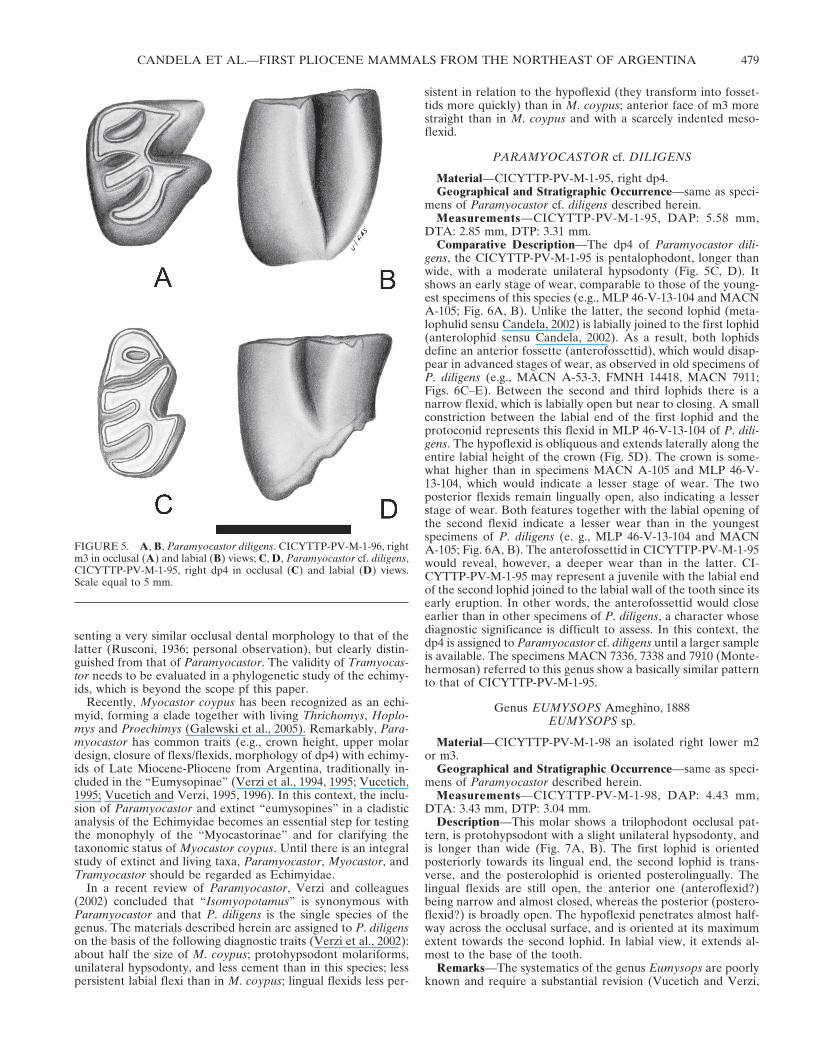
The m3 (CICYTTP-PV-M-1-96) is tetralophodont (Fig. 5A,B), protohypsodont, with similar length and width (see measure-ments). It shows an early stage of wear, comparable to that of m3of MLP 46-V-13-104 of this species (Verzi et al., 2002) (Fig. 6A).The hypoflexid extends laterally along the entire height of thecrown (Fig. 5B), and its bottom is oriented towards the lingualend of the hypolophid. The mesoflexid is poorly penetrating,although more than in MLP 46-V-13-104. The anterior flexid hastransformed into a fossettid, while the second and third flexidsare widely opened lingually, which indicates little wear. In lin-gual view, the most posterior of these flexids is much deeper thanthe anterior one, such as in MLP 46-V-13-104.
Remarks—The systematic status of Myocastor and its closelyrelated extinct genera, like Paramyocastor and TramyocastorRusconi, 1936, is controversial (e.g., Woods, 1982; McKenna andBell, 1997; Verzi et al., 2002). Tramyocastor is a poorly knownextinct genus, traditionally included among the “Myocastori-dae”, and represented by two species. T. andiai (Rusconi, 1936)and T. majus (Rusconi, 1936) are defined on very scarce andfragmentary remains with a dubious stratigraphic provenance.Both species are smaller than those of Myocastor, although pre-
FIGURE 3. South American chronostratigraphic Late Cenozoic con-tinental units (modified from Cione et al., 2000), the Magnetic PolarityTime Scale (MPTS), and the stratigraphic distribution of Paedotherium,Paramyocastor and Eumysops.
FIGURE 4. Paramyocastor diligens. A, B, CICYTTP-PV-M-1-97, rightM1? in occlusal (A), and lingual (B) views; C, FMNHN 14418, rightDP4-M3 (modified from Verzi et al., 2002); D, MACN 7912, right DP4-M3 (modified from Verzi et al., 2002); E, MACN 7344, left DP4-M3(reversed). Scale equal to 5 mm.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 2, 2007478
senting a very similar occlusal dental morphology to that of thelatter (Rusconi, 1936; personal observation), but clearly distin-guished from that of Paramyocastor. The validity of Tramyocas-tor needs to be evaluated in a phylogenetic study of the echimy-ids, which is beyond the scope pf this paper.
Recently, Myocastor coypus has been recognized as an echi-myid, forming a clade together with living Thrichomys, Hoplo-mys and Proechimys (Galewski et al., 2005). Remarkably, Para-myocastor has common traits (e.g., crown height, upper molardesign, closure of flexs/flexids, morphology of dp4) with echimy-ids of Late Miocene-Pliocene from Argentina, traditionally in-cluded in the “Eumysopinae” (Verzi et al., 1994, 1995; Vucetich,1995; Vucetich and Verzi, 1995, 1996). In this context, the inclu-sion of Paramyocastor and extinct “eumysopines” in a cladisticanalysis of the Echimyidae becomes an essential step for testingthe monophyly of the “Myocastorinae” and for clarifying thetaxonomic status of Myocastor coypus. Until there is an integralstudy of extinct and living taxa, Paramyocastor, Myocastor, andTramyocastor should be regarded as Echimyidae.
In a recent review of Paramyocastor, Verzi and colleagues(2002) concluded that “Isomyopotamus” is synonymous withParamyocastor and that P. diligens is the single species of thegenus. The materials described herein are assigned to P. diligenson the basis of the following diagnostic traits (Verzi et al., 2002):about half the size of M. coypus; protohypsodont molariforms,unilateral hypsodonty, and less cement than in this species; lesspersistent labial flexi than in M. coypus; lingual flexids less per-
sistent in relation to the hypoflexid (they transform into fosset-tids more quickly) than in M. coypus; anterior face of m3 morestraight than in M. coypus and with a scarcely indented meso-flexid.
PARAMYOCASTOR cf. DILIGENS
Material—CICYTTP-PV-M-1-95, right dp4.Geographical and Stratigraphic Occurrence—same as speci-
mens of Paramyocastor cf. diligens described herein.Measurements—CICYTTP-PV-M-1-95, DAP: 5.58 mm,
DTA: 2.85 mm, DTP: 3.31 mm.Comparative Description—The dp4 of Paramyocastor dili-
gens, the CICYTTP-PV-M-1-95 is pentalophodont, longer thanwide, with a moderate unilateral hypsodonty (Fig. 5C, D). Itshows an early stage of wear, comparable to those of the young-est specimens of this species (e.g., MLP 46-V-13-104 and MACNA-105; Fig. 6A, B). Unlike the latter, the second lophid (meta-lophulid sensu Candela, 2002) is labially joined to the first lophid(anterolophid sensu Candela, 2002). As a result, both lophidsdefine an anterior fossette (anterofossettid), which would disap-pear in advanced stages of wear, as observed in old specimens ofP. diligens (e.g., MACN A-53-3, FMNH 14418, MACN 7911;Figs. 6C–E). Between the second and third lophids there is anarrow flexid, which is labially open but near to closing. A smallconstriction between the labial end of the first lophid and theprotoconid represents this flexid in MLP 46-V-13-104 of P. dili-gens. The hypoflexid is obliquous and extends laterally along theentire labial height of the crown (Fig. 5D). The crown is some-what higher than in specimens MACN A-105 and MLP 46-V-13-104, which would indicate a lesser stage of wear. The twoposterior flexids remain lingually open, also indicating a lesserstage of wear. Both features together with the labial opening ofthe second flexid indicate a lesser wear than in the youngestspecimens of P. diligens (e. g., MLP 46-V-13-104 and MACNA-105; Fig. 6A, B). The anterofossettid in CICYTTP-PV-M-1-95would reveal, however, a deeper wear than in the latter. CI-CYTTP-PV-M-1-95 may represent a juvenile with the labial endof the second lophid joined to the labial wall of the tooth since itsearly eruption. In other words, the anterofossettid would closeearlier than in other specimens of P. diligens, a character whosediagnostic significance is difficult to assess. In this context, thedp4 is assigned to Paramyocastor cf. diligens until a larger sampleis available. The specimens MACN 7336, 7338 and 7910 (Monte-hermosan) referred to this genus show a basically similar patternto that of CICYTTP-PV-M-1-95.
Genus EUMYSOPS Ameghino, 1888EUMYSOPS sp.
Material—CICYTTP-PV-M-1-98 an isolated right lower m2or m3.
Geographical and Stratigraphic Occurrence—same as speci-mens of Paramyocastor described herein.
Measurements—CICYTTP-PV-M-1-98, DAP: 4.43 mm,DTA: 3.43 mm, DTP: 3.04 mm.
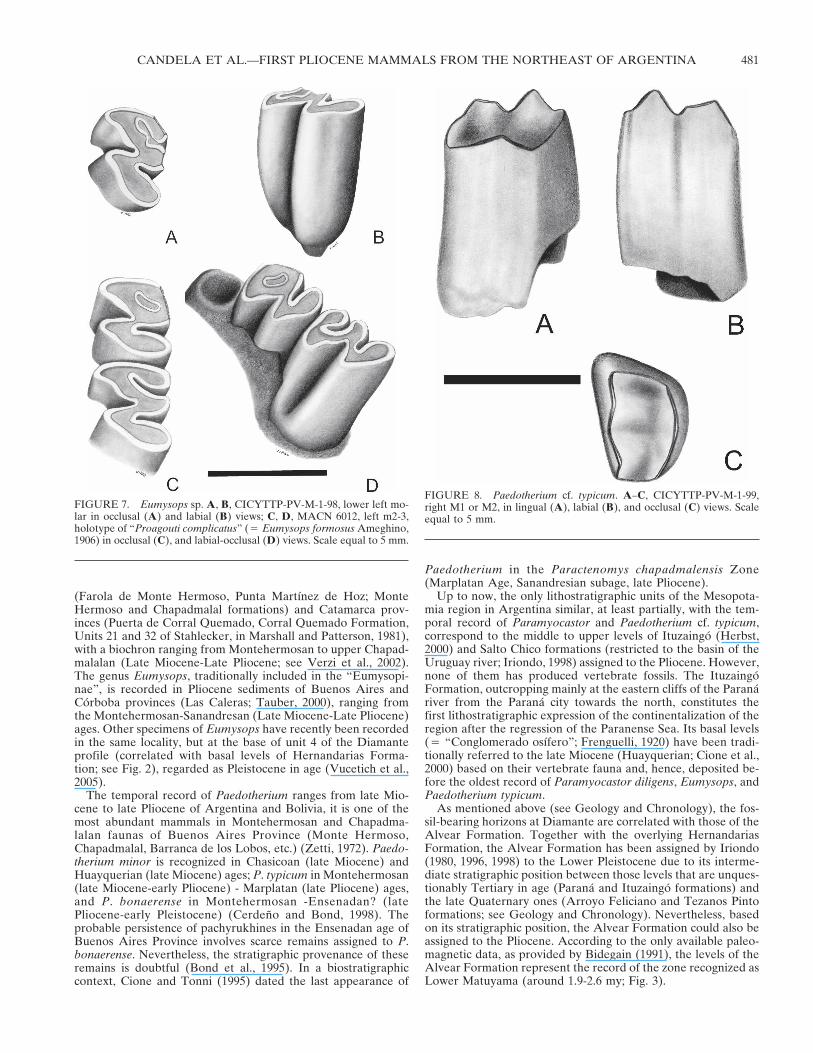
Description—This molar shows a trilophodont occlusal pat-tern, is protohypsodont with a slight unilateral hypsodonty, andis longer than wide (Fig. 7A, B). The first lophid is orientedposteriorly towards its lingual end, the second lophid is trans-verse, and the posterolophid is oriented posterolingually. Thelingual flexids are still open, the anterior one (anteroflexid?)being narrow and almost closed, whereas the posterior (postero-flexid?) is broadly open. The hypoflexid penetrates almost half-way across the occlusal surface, and is oriented at its maximumextent towards the second lophid. In labial view, it extends al-most to the base of the tooth.
Remarks—The systematics of the genus Eumysops are poorlyknown and require a substantial revision (Vucetich and Verzi,
FIGURE 5. A, B, Paramyocastor diligens. CICYTTP-PV-M-1-96, rightm3 in occlusal (A) and labial (B) views; C, D, Paramyocastor cf. diligens,CICYTTP-PV-M-1-95, right dp4 in occlusal (C) and labial (D) views.Scale equal to 5 mm.
CANDELA ET AL.—FIRST PLIOCENE MAMMALS FROM THE NORTHEAST OF ARGENTINA 479
1995). CICYTTP-PV-M-1-98 shares several traits (general size,crown height, unilateral hypsodonty, occlusal pattern) with otherspecimens assigned to Eumysops, such as MACN 6012 (holotypeof “Proagouti complicatus” Ameghino, 1908 � Eumysops for-mosus Ameghino, 1906; see Kraglievich, 1957) (Fig. 7 C-D) fromthe Chapadmalalan levels of the Buenos Aires Province.
Order NOTOUNGULATA Roth, 1903Suborder HEGETOTHERIA Simpson, 1945
Family HEGETOTHERIIDAE Ameghino, 1894Subfamily PACHYRUKHINAE Kraglievich, 1934
Genus PAEDOTHERIUM Burmeister, 1888PAEDOTHERIUM cf. TYPICUM
Material—CICYTTP-PV-M-1-99, right M1 or M2.Geographical and Stratigraphic Distribution—same as speci-
mens of Paramyocastor diligens described herein.Measurements—DAP: 4.03 mm; DTA: 2.98 mm; DTP:
2.25 mmComparative Description—The tooth is prismatic and the
enamel discontinuous. Its stage of wear indicates that it is ayoung adult specimen. A dentine band crosses over the anteriorface (Fig. 8) and a thin cement layer covers the tooth. The oc-clusal surface shows two basins of the same depth, but the an-terior one is larger and coincides topographically with theparacone and metacone crests respectively. The columns of theparacone and metacone are clearly marked in the ectoloph and
end in two sharp crests of the same height. It has no lingualgrooves. Its morphology resembles that of the M1 of Paedo-therium typicum (Ameghino, 1887) although it is smaller. TheM1 of the lectotype, MLP 12-1782, has larger DTA than DTPsuch as CICYTTP-PV-M-1-99, but this specimen is rather worn.It resembles Paedotherium minor Cabrera, 1937 in size but theocclusal design is different, having less developed ectoloph col-umns. It differs from Paedotherium bonaerense (Ameghino,1887) in its smaller size and a more rectangular occlusal shape.
Remarks—Pachyrukhines are extremely common in theMiocene and Pliocene of Argentina. From the late Miocene(Chasicoan) to late Pliocene (Marplatan), only two genera arerepresented, Paedotherium and Tremacyllus. Both are very simi-lar in dental morphology; however, our specimen, CICYTTP-PV-M-1-99, is more similar to Paedotherium in the occlusal out-line (more elongated anterior-posterior diameter) and moreprominent paracone and metacone. Of the three known speciesof the genus, P. minor, P. typicum and P. bonaerense (Cerdeñoand Bond, 1998), the specimen CICYTTP-PV-M-1-99 showsgreatest resemblance to P. typicum.
DISCUSSION
Biochronological Implications
The mammals found at basal levels of the Punta Gorda Grouphave an important biochronologic significance. Paramyocastordiligens is known only from localities of the Buenos Aires
FIGURE 6. Paramyocastor diligens. A, MLP 46-V-13-104, left dp4-m3 (reversed); B, MACN A-105, right dp4-m2; C, MACN A-53-3, right dp4-m3;D, FMNH 14418, right dp4-m3; E, MACN 7911, right dp4-m3. Scale equal to 5 mm.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 2, 2007480
(Farola de Monte Hermoso, Punta Martínez de Hoz; MonteHermoso and Chapadmalal formations) and Catamarca prov-inces (Puerta de Corral Quemado, Corral Quemado Formation,Units 21 and 32 of Stahlecker, in Marshall and Patterson, 1981),with a biochron ranging from Montehermosan to upper Chapad-malalan (Late Miocene-Late Pliocene; see Verzi et al., 2002).The genus Eumysops, traditionally included in the “Eumysopi-nae”, is recorded in Pliocene sediments of Buenos Aires andCórboba provinces (Las Caleras; Tauber, 2000), ranging fromthe Montehermosan-Sanandresan (Late Miocene-Late Pliocene)ages. Other specimens of Eumysops have recently been recordedin the same locality, but at the base of unit 4 of the Diamanteprofile (correlated with basal levels of Hernandarias Forma-tion; see Fig. 2), regarded as Pleistocene in age (Vucetich et al.,2005).
The temporal record of Paedotherium ranges from late Mio-cene to late Pliocene of Argentina and Bolivia, it is one of themost abundant mammals in Montehermosan and Chapadma-lalan faunas of Buenos Aires Province (Monte Hermoso,Chapadmalal, Barranca de los Lobos, etc.) (Zetti, 1972). Paedo-therium minor is recognized in Chasicoan (late Miocene) andHuayquerian (late Miocene) ages; P. typicum in Montehermosan(late Miocene-early Pliocene) - Marplatan (late Pliocene) ages,and P. bonaerense in Montehermosan -Ensenadan? (latePliocene-early Pleistocene) (Cerdeño and Bond, 1998). Theprobable persistence of pachyrukhines in the Ensenadan age ofBuenos Aires Province involves scarce remains assigned to P.bonaerense. Nevertheless, the stratigraphic provenance of theseremains is doubtful (Bond et al., 1995). In a biostratigraphiccontext, Cione and Tonni (1995) dated the last appearance of
Paedotherium in the Paractenomys chapadmalensis Zone(Marplatan Age, Sanandresian subage, late Pliocene).
Up to now, the only lithostratigraphic units of the Mesopota-mia region in Argentina similar, at least partially, with the tem-poral record of Paramyocastor and Paedotherium cf. typicum,correspond to the middle to upper levels of Ituzaingó (Herbst,2000) and Salto Chico formations (restricted to the basin of theUruguay river; Iriondo, 1998) assigned to the Pliocene. However,none of them has produced vertebrate fossils. The ItuzaingóFormation, outcropping mainly at the eastern cliffs of the Paranáriver from the Paraná city towards the north, constitutes thefirst lithostratigraphic expression of the continentalization of theregion after the regression of the Paranense Sea. Its basal levels(� “Conglomerado osífero”; Frenguelli, 1920) have been tradi-tionally referred to the late Miocene (Huayquerian; Cione et al.,2000) based on their vertebrate fauna and, hence, deposited be-fore the oldest record of Paramyocastor diligens, Eumysops, andPaedotherium typicum.
As mentioned above (see Geology and Chronology), the fos-sil-bearing horizons at Diamante are correlated with those of theAlvear Formation. Together with the overlying HernandariasFormation, the Alvear Formation has been assigned by Iriondo(1980, 1996, 1998) to the Lower Pleistocene due to its interme-diate stratigraphic position between those levels that are unques-tionably Tertiary in age (Paraná and Ituzaingó formations) andthe late Quaternary ones (Arroyo Feliciano and Tezanos Pintoformations; see Geology and Chronology). Nevertheless, basedon its stratigraphic position, the Alvear Formation could also beassigned to the Pliocene. According to the only available paleo-magnetic data, as provided by Bidegain (1991), the levels of theAlvear Formation represent the record of the zone recognized asLower Matuyama (around 1.9-2.6 my; Fig. 3).
FIGURE 7. Eumysops sp. A, B, CICYTTP-PV-M-1-98, lower left mo-lar in occlusal (A) and labial (B) views; C, D, MACN 6012, left m2-3,holotype of “Proagouti complicatus” (� Eumysops formosus Ameghino,1906) in occlusal (C), and labial-occlusal (D) views. Scale equal to 5 mm.
FIGURE 8. Paedotherium cf. typicum. A–C, CICYTTP-PV-M-1-99,right M1 or M2, in lingual (A), labial (B), and occlusal (C) views. Scaleequal to 5 mm.
CANDELA ET AL.—FIRST PLIOCENE MAMMALS FROM THE NORTHEAST OF ARGENTINA 481

Paleontological evidence indicates an age extending from thelate Miocene (Montehermosan) to the late Pliocene (Marplatan)for the vertebrate-bearing sediments of the lowest part of thePunta Gorda Group based on the taxon ranges (Fig. 3). On theother hand, geologic evidence does not contradict a Pliocene agefor this unit and the paleomagnetic data, not only reinforces thisassignment, but adjusts the age to the late Pliocene. The inter-pretation extends the temporal record of Paramyocastor to theMarplatan age. These are the first vertebrate-bearing levels rec-ognized for this period in the Argentinean Mesopotamia.
Evolutionary and Paleoenvironmental Implications
In a recent molecular study, Galewski and colleagues (2005)pointed out that most of the extant echimyid genera appeared tohave diverged during the middle Miocene. Interestingly, Candelaand Noriega (2004) have confirmed the presence of Myocastor inthe late Miocene of the Mesopotamia (Huayquerian SALMA9.8-9.0 Ma; Cione et al., 2000). With the exception of Myocastor,living echimyid genera are known since the Pleistocene (Vuce-tich and Verzi, 1995). Therefore, Myocastor is undoubtedly themost ancient living genus of South American Hystricognathi(Candela and Noriega, 2004), and the late Miocene could be astrict minimum age for the origin of this genus, as it has beenproposed for other living echimyids (Galewski et al., 2005). Thus,the fossil record of Myocastor appears in agreement with mo-lecular data. Assuming the close relationship between Paramyo-castor and Myocastor, it is possible that they diverged very early,at least during the late Miocene. The oldest representatives ofMyocastor (Ameghino, 1885, 1986, 1989; Rusconi, 1929, 1945;Candela and Noriega, 2004) seem to have been confined to thePampean and Mesopotamian regions. They are not recorded inlate Miocene-Pliocene sediments of the northwestern Argentina(e.g., Andalhualá Formation, Chiquimil Formation, Corral Que-mado Formation). The conservative morphology of extinct spe-cies of Myocastor (Candela and Noriega, 2004) suggests a higherdegree of habitat specificity in wetlands and the development ofa semiaquatic lifestyle since the earliest records of the genus(Galewski et al., 2005). Paramyocastor could have represented adifferent ecomorphological type. Unlike Myocastor, it was dis-tributed on both Northwest and Pampean areas, showing a geo-graphic distribution similar to that of the “eumysopines” fromthe late Miocene–Pliocene of Argentina, which could have beenadapted to open areas with dry climates (Verzi et al., 1994, 1995;Vucetich and Verzi, 1995). Probably as result of climatic andenvironmental changes produced by the Diaguita diastrophicphase of Andean orogeny, more arid conditions were establishedduring the Pliocene (Pascual et al., 1996). Paramyocastor wouldhave become extinct during the Pliocene, probably at the end ofthe Marplatan Stage (late Pliocene), together with many nativemammals, during a change towards more humid conditions in-ferred for this time (Cione and Tonni, 1995). The distributionalpatterns of Paramyocastor, Tramyocastor, and Myocastor seemto support an austral biogeographical history developed in thesouthern plains of the continent for the “myocastorines.”
Paedotherium is one of the most abundant mammals in theMontehermosan and Chapadmalalan faunas from the BuenosAires Province (Monte Hermoso, Chapadmalal, Barranca de losLobos, etc.). The presence of Paedotherium cf. typicum in EntreRíos represents the most northern distribution of the subfamilyPachyrukhinae in the Pliocene, and suggests that the environ-ment where it lived could have been similar to that of the Pam-pean region for the same time. Except for the records of Paedo-therium minor in the late Miocene (Chasicoan-Huayquerian)from the Guandacay Formation in Bolivia (Marshall and Sem-pere, 1991), and middle Miocene from Cura Mallín Formation inChile (Wyss et al., 2003), the genus Paedotherium had a strictlyPampean distribution.
Pachyrukhines are the most hypsodont notoungulates andshow cranio-dental and pedal adaptations very similar to rodents(Scott, 1937) or lagomorphs (Cifelli, 1985). Paedotherium and itssister taxon Tremacyllus (Cerdeño and Bond, 1998) strongly ex-hibit those rodent-like characters. According to Cerdeño andBond (1998:810) Paedotherium would have been “. . . a cursorialmammal and open habitat dweller, possibly with semi-fossorialand maybe nocturnal habits”. Based on a morphometric analysis,Elissamburu (2004) pointed out that Paedotherium is better pre-pared to develop force (humerus robustness, olecranon propor-tion and ulna robustness) rather than speed, and a fossorial habitcan be inferred for this genus. The cranio-dental morphologyreveals morphofunctional aspects which resemble those of cer-tain caviomorph rodents, i.e., Dolichotis and Pediolagus. Theparticular shape of the central superior incisor (curvilinear withoblique implantation), the shape and proportion of the snout, thegreatly developed masseteric fossa, its high hypsodonty index(HI > 4) and the small size suggests a dietary specialization forhighly abrasive food with hard substrate (dust and ash). In thissense, the high abundance of Paedotherium during the Pliocenein the Pampean region and its absence in faunas from the high-lands of the Puna and Cordillera Oriental in northwestern Ar-gentina and Bolivia suggests that it would have been adapted tosemiarid regions of plains and lowlands.
In sum, the Pliocene mammals herein reported suggest morearid and open lowlands than those proposed for the continentallate Miocene (“Conglomerado osífero”) of the Mesopotamia re-gion, characterized by humid and forested habitats (Cione et al.,2000). This paleonvironmental scenario agrees with the sedimen-tological evidence, indicative of semiarid and dry climatic con-ditions during the deposition of the Alvear Formation (Iriondo,1980: 139; Bidegain, 1991).
ACKNOWLEDGMENTS
We are grateful to A. Kramarz for giving permission to accessMACN collections; to Juan and D. Diederle for their coopera-tion in field work and for finding the fossil material. To G. Vuce-tich for her advice. D. Croft, R. Reisz, and one anonymousreviewer significantly improved this paper with their criticalreviews. This paper is a contribution to the “Programa de Biodi-versidad del Litoral” (SECYT) and to PICT 11928 (ANPCYT).
LITERATURE CITED
Aceñolaza, F. G. 2000. La Formación Paraná (Mioceno Medio): estrati-grafía, distribución regional y unidades equivalentes; pp. 9–27 in F.Aceñolaza, and R. Herbst (eds.), El Mio-Plioceno Argentino. Insu-geo, Serie de Correlación Geológica 14.
Ameghino, F. 1885. Nuevos restos de mamíferos fósiles oligocenosrecogidos por el profesor Pedro Scalabrini y pertenecientes al Mu-seo Provincial de la Ciudad de Paraná. Boletín de la AcademiaNacional de Ciencias de Córdoba 8:5–207. Buenos Aires.
Ameghino, F. 1886. Contribución al conocimiento de los mamíferosfósiles de los terrenos terciarios antiguos de Paraná. Boletín de laAcademia Nacional de Ciencias de Córdoba 9:5–228.
Ameghino, F. 1887. Enumeración sistemática de las especies demamíferos fósiles coleccionados por Carlos Ameghino en terrenoseocenos de la Patagonia austral y depositados en el Museo de LaPlata. Boletín del Museo La Plata 1:1–26.
Ameghino, F. 1888. Lista de las especies de mamíferos fósiles del Mio-ceno superior de Monte Hermoso, hasta ahora conocidas. E. Coni,Buenos Aires. pp. 1–21.
Ameghino, F. 1889. Contribución al conocimiento de los mamíferosfósiles de la República Argentina. Boletín de la Academia Nacionalde Ciencias de Córdoba 6:1–127.
Ameghino, F. 1904. Nuevas especies de mamíferos cretácicos y terciariosde la República Argentina. Anales de la Sociedad Científica Argen-tina, 56, 57, 58: 103.
Ameghino, F. 1908. Las Formaciones sedimentarias de la región litoral
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 2, 2007482

de Mar del Plata y Chapadmalal. Anales del Museo Nacional deBuenos Aires 10:342–428.
Bidegain, J. C. 1991. Sedimentary development, magnetostratigraphyand sequence of events of the Late Cenozoic in Entre Ríos andsurrounding areas in Argentina. Ph.D. dissertation, Stockholm Uni-versity, Stockholm, Sweden, 128 pp.
Bond, M., E. Cerdeño, and G. López. 1995. Los ungulados nativos deAmérica del Sur; pp. 259–275 in M. T. Alberdi, G. Leone, and E. P.Tonni (eds.), Evolución Biológica y Climática de la Región Pam-peana durante los últimos cinco millones de años. Monografías delMuseo Nacional de Ciencias Naturales, Madrid.
Burmeister, C. V. 1888. Relación de un viaje a la Gobernación deChubut. Anales del Museo Nacional de Buenos Aires 3:175–252.
Cabrera, A. 1937. Notas sobre el suborden Typotheria. Notas del Museode La Plata 2(8):17–43.
Candela, A. M. 2002. Lower deciduous tooth homologies in Erethizon-tidae (Rodentia, Hystricognathi): Evolutionary significance. ActaPalaeontologica Polonica 47:717–723.
Candela, A. M., and J. I. Noriega. 2004. Los coipos (Rodentia, Cavio-morpha, Myocastoridae) del “Mesopotamiense” (Mioceno tardío;Formación Ituzaingó) de la provincia de Entre Ríos, Argentina; pp.5–12 in F. G. Aceñolaza (ed.), Temas de la Biodiversidad del LitoralFluvial Argentino, serie Insugeo, Miscelánea 12.
Cerdeño, E., and M. Bond. 1999. Taxonomic revision and phylogeny ofPaedotherium and Tremacyllus (Pachyrukhinae, Hegetotheriidae,Notoungulata) from the late Miocene to the Pleistocene of Argen-tina. Journal of Vertebrate Paleontology 18:799–811.
Cifelli, R. L. 1985. South American ungulate evolution and extinction;pp. 249–266 in F. Stehli, and S. D. Webb (eds.), The Great AmericanBiotic Interchange Plenum Press, New York and London.
Cione, A. L., and E. P. Tonni. 1995. Chronostratigraphy and “Land-mammal Ages” in the Cenozoic of southern South America: Prin-ciples, Practices, and the “Uquian” Problem. Journal of Paleontol-ogy 69:135–159.
Cione, A. L., M. Azpelicueta, M. Bond, A. A. Carlini, J. R. Casciotta,M. A. Cozzuol, M. de la Fuente, Z. Gasparini, F. J. Goin, J. Noriega,G. J. Scillato-Yané, L. Soibelzon, E. P. Tonni, D. Verzi, and M. G.Vucetich. 2000. Miocene vertebrates from Entre Ríos province,eastern Argentina; pp. 191–237 in F. G. Aceñolaza, and R. Herbst(eds.), El Mio-neógeno Argentino. Insugeo, Serie CorrelaciónGeológica 14.
Elissamburu, A. 2004. Análisis morfométrico y morfofuncional del es-queleto apendicular de Paedotherium (Mammalia, Notoungulata).Ameghiniana 4:363–380.
Frenguelli, J. 1920. Contribución al conocimiento de la geología de EntreRíos. Boletín de la Academia Nacional de Ciencias de Córdoba24:55–256.
Galewski, T., J. F. Mauffrey, Y.L. R. Leite, J. L. Patton, and E. J. P.Douzery. 2005. Ecomorphological diversification among SouthAmerican spiny rats (Rodentia; Echimyidae): a Phylogenetic andchronological approach. Molecular Phylogenetic and Evolution 34:601–615.
Herbst, R., L. M. Anzótegui, G. Esteban, L. R. Mautino, S. Morton, andN. Nasif. 2000. Síntesis paleontológica del Mioceno de los vallesCalchaquíes, noroeste argentino. Miocene vertebrates from EntreRíos province, eastern Argentina; pp. 263–268 in F. G. Aceñolaza,and R. Herbst (eds.), El Mio-Plioceno Argentino. Insugeo, Serie deCorrelación Geológica 14.
Iriondo, M. H. 1980. El Cuaternario de Entre Ríos. Revista AsociaciónCiencias Naturales del Litoral 1:125–141.
Iriondo, M. H. 1996. Estratigrafía del Cuaternario de la cuenca del ríoUruguay. Actas IV XIII Congreso Geológico Argentino y III Con-greso de Exploración de Hidrocarburos, 15–25.
Iriondo, M. 1998. Loess in Argentina: Temperate and Tropical. Excur-sion Guide N° 3, Province of Entre Ríos. International Joint FieldMeeting, INQUA, 16 pp.
Kraglievich, J. L. 1957. Revisión de los roedores extinguidos del géneroEumysops Ameghino 1888. (Nota preliminar). Ameghiniana 1:37–39.
Marshall, L. G., and B. Patterson. 1981. Geology and geochronology of
the mammal-bearing Tertiary of the Valle de Santa María and RíoCorral Quemado, Catamarca Province, Argentina. Fieldiana Geol-ogy 9:1–80.
Marshall, L. G., and T. Sempere. 1991. The Eocene to Pleistocene ver-tebrates of Bolivia and their stratigraphic context: a review; pp.631–652 in R. Súarez (ed.), Fósiles y facies de Bolivia, Revista Téc-nica de Yacimientos Petrolíferos Fiscales Bolivianos Volume 1, 12.
McKenna, M. C., and S. K. Bell. 1998. Classification of Mammals Abovethe Species Level. Columbia University Press, New York, 631 pp.
Noriega, J. I., A. A. Carlini, and E. P. Tonni. 2004. Vertebrados delPleistoceno tardío de la cuenca del Arroyo Ensenada (Departa-mento Diamante, Provincia de Entre Ríos); pp. 71–76 in F. G.Aceñolaza (ed.), Temas de la Biodiversidad del Litoral fluvial ar-gentino. Insugeo, Miscelánea 12.
Pascual, R., E. Ortiz Jaureguizar, and J. L. Prado. 1996. Land Mammals:Paradigm for Cenozoic South American Geobiotic Evolution; pp.265–319 in G. Arratia (ed.), Contribution of Southern SouthAmerica to Vertebrate Paleontology. Munchner Geowissenschaft-liche Abhandlungen 30.
Rusconi, C. 1929. Revisión de las especies fósiles argentinas del géneroMyocastor, con descripción de nuevas especies. Anales de la Socie-dad Argentina de Estudios Geográficos “G.E.A.” Tomo 3, Nº 2,504–518.
Rusconi, C. 1936. Nuevo género de roedores del Puelchense de VillaBallester. Boletín paleontológico de Buenos Aires 7:1–4.
Rusconi, C. 1945. Varias especies de roedores del Puelchense de BuenosAires. Anales de la Sociedad Científica Argentina 140:369–376.
Scott, W. B. 1937. A History of Land Mammals in the Western Hemi-sphere. The Mac Millan Company, New York, 786 pp.
Tauber, A. A. 2000. Las Caleras, Provincia de Córdoba, primeryacimiento fosilífero de origen cárstico de Argentina. Revista Es-pañola de Paleontología 15:233–244.
Verzi, D. H., M. G. Vucetich, and C. Montalvo. 1994. Octodontid-likeEchimyhidae (Rodentia); an Upper Miocene episode in the radia-tion of the Family. Palaeovertebrata 23:199–210.
Verzi, D. H., M. G. Vucetich, and C. Montalvo. 1995. Un nuevo Eumy-sopinae (Rodentia, Echimyidae) del Mioceno tardío de la provinciade La Pampa y consideraciones sobre la historia de la subfamilia.Ameghiniana 32:191–195.
Verzi, H. D., C. M. Deschamps, and M. G. Vucetich. 2002. Sistemática yantigüedad de Paramyocastor diligens (Ameghino, 1888) (Rodentia,Caviomorpha, Myocastoridae). Ameghiniana 39:193–200.
Vucetich, M. G. 1995. Theridomysops parvulus (Rovereto, 1914), unprimitivo Eumysopinae (Rodentia, Echimyidae) del Mioceno tardíode Argentina. Mastozoología Neotropical 2:167–172.
Vucetich, M. G., and D. H. Verzi. 1995. Los Roedores Caviomorpha;211–225 in: M. T. Alberdi, G. Leone, and E. P. Tonni (eds.), Evo-lución Biológica y Climática de la Región Pampeana durante losúltimos Cinco Millones de años. Monografías del Museo de CienciasNaturales de Madrid 12.
Vucetich, M. G., and D. H. Verzi. 1996. Un nuevo Eumysopinae (Ro-dentia, Echimyidae) de la “Formación“ Irene (Chapadmalalenseinferior?) y la diversidad de la Subfamilia. Actas IV Jornadas deGeología y Geofísica Bonaerenses 1:159–165.
Vucetich, M. G., E. C. Vieytes, D. H. Verzi, J. I. Noriega, and E. P.Tonni. 2005. Unexpected primitive rodents in the Quaternary ofArgentina. Journal of South American Earth Sciences 20:57–64.
Woods, C. A. 1982. The history and classification of South AmericanHystricognathi rodents: reflections on the far away and long ago. pp.377–392 in M. A. Mares, and J. H. Genoways (eds.), MammalianBiology in South America. University of Pittsburgh, Pittsburgh.
Wyss, A., R. Charrier, D. Croft, J. Flynn, and J. Wertheim. 2003. Newmiddle Cenozoic mammals from the Laguna del Laja region (CuraMallín Formation, South Central Chile). Journal of Vertebrate Pa-leontology 23(Supplement 3):113A.
Zetti, J. 1972. Observaciones sobre los Pachyrukhinae (Notoungulata)del Plioceno argentino. Publicaciones del Museo Municipal de Cien-cias Naturales de Mar del Plata 2: 41–52.
Submitted August 9, 2006; accepted January 16, 2007.
CANDELA ET AL.—FIRST PLIOCENE MAMMALS FROM THE NORTHEAST OF ARGENTINA 483




















