
Prevalence of widespread pain and associations with work status: a population study
Upload
independentCategory
view
0download
0
1
Helmut Kruckenberg, Alexander Kondratyev, Johan H. Mooij, Christoph Zöckler & Elmira Zaynagutdinova
White-fronted Goose Flyway Population StatusInterim Report of a preliminary study in 2006

2
Impressum
H.Kruckenberg, A.Kondratyev, J.H. Mooij, C.Zöckler & E. Zaynagutdinova (2008): White-fron-ted Goose Flyway Population Status. - Interim Report of a preliminary study in 2006. - Angewandte Feld-biologie Band II: 1-63. ISSN 1861-227X
This scientitic project was financially supported by Vogelschutz-Komitee VsK e.V. (Hamburg, Göttingen) and GEF/ UNEP-ECORA-project (Moscow)

3
1. Introduction
Wild geese migrating each year long distances from their breeding grounds to wintering grounds in wes-tern, southeastern and southern Europe. Although the populations of Whitefronted (Anser albifrons), Bean (A. fabalis) and Barnacle Geese (Branta leucopsis) decreased in the early 20th century, their numbers began recovering in the 1970s, thanks to hunting restrictions and intensified agriculture. By the end of the 20th century, the large goose populations wintering in western Europe caused more and more discussion about damage to agricultural farmland and pastures, and flyway countries often tried to end this through a reduction in hunting restrictions. Barnacle geese winter exclusivly in Western Europe, mainly in the Wadden Sea region, where population estimates with a high level of reliability are taken regularly. In contrast to the Barnacle Goose popula-tion estimates,the those for wintering Whitefronted and Bean Geese populations are far less reliable, since both species winter over a large range, from Kazakstan to France. Often numbers at staging sites are hard to count within a short number of days, and the individuals are constantly changing. So, over the length of the flyway, there are large gaps in populati-on numbers and often great uncertiainties even with the numbers we do have. However, valid data on population numbers, status and trends are the foun-dation upon which all protection and management tasks are built. So, alternative ways to obtain reliable knowledge of the present situation of all Arctic goo-se populations are urgently required.
The flyway approach as an important cornerstone for the study and conservation of goose populationsBy using neckbanded birds marked on theTaimyr, MOOIJ et al. (1996, 1996a) proposed a closely connected network of migration routes used by populations of Anser spp. covering the whole western Palaearctic flyway. Since the European Whitefronted Research Project was started in 1998, a lot of data on Whitefronts have been gathered by using neck-bands, mainly in The Netherlands, Germany and Belgium (KRUCKENBERG 2002). But east of the river Oder the number of observations declines marked-ly, and we and are lacking data for the last phase of spring migration through the north of Russia and Si-beria. There may be highly important stepping-stone
sites , but they remain quite unknown. How species of Anser, such as Whitefronts, migrate in spring and autumn is even less understood. In order to accomplish this, addi-tional methods to study bird migration, such as telemetry and inventory expeditions, are required to collect more data in the eastern part of the flyway.
Stopover sites – key points for successful reproduction and for human impactThe importance of spring staging areas for successful reproduction of Arctic-breeding migrating avian spe-cies has received thorough attention in recent decades, so that several theoretic models have been proposed to understand the spring migration strategy of Arctic geese via the network of staging sites, where the energy re-sources, necessary for reproduction are gained (DRENT & DAAN 1980, VAN DEN GRAAF 2006, EICHHORN 2008, EBBIN-GE & SPAANS 1995). At the same time, the relationships between natural causes („Green-wave” hypothesis) and human impact are still poorly studied. Nonetheless, it is clear enough that human disturbance may play an impor-tant role in shifting geese between feeding habitats during intensive feeding in spring (PRINS & YDENBERG 1985). The majority of Arctic geese breeding in the Russian Arctic stage nowadays on agricultural lands, where they depend both on the habitat structure of the agricultural fields and on disturbance factors caused by different types of human activities. Thus the urgent need for such kind of a stopo-ver-study has become more and more clear. Agricultural fields around the town of Olonets on the eastern side of Lake Ladoga were among the first sites where detailed populational studies of staging Arctic-breeding geese were carried out, and have been ongoing since 1994 (ZI-MIN et al 2007). Understanding the role of this seasonally protected area in terms of the flyway approach to the conservation of goose populations, and of human impact on staging flocks of geese, were therefore the goals of the current studies. Such studies had the possible benefit of linking with investigations of the population ecology of Western Palaearctic geese on their most important bree-ding grounds in the Russian Arctic.
Breeding areas: the importance of Kolguyev, owing to its position and history Being known as an important breeding ground of Western Palaearctic geese, Kolguev Island for a long time remai-ned one of the most poorly studied and mysterious places

4
in the Russian Arctic. Due to its wet climate and comple-te absence of lemmings and other small mammals, this island supported great numbers and the highest nesting densities of species of Anser geese ever recorded in the northern hemisphere. Despite this, in the 100-year histo-ry of the exploration of the island, one can count barely more than 10 studies devoted to any kind of population estimates of geese living there, and such population parameters as total numbers, patterns of distribution, and breeding success remained unknown. At the same time it appeared clear, that the total numbers of geese that nest on Kolguev might be high enough to play an important role in the entire Western Palaearctic population of at least three goose species.
Tasks and objectives for the base-line phase.In order to fill these wide gaps we had several first and most important tasks and objectives to do during the in-ventory phase of our studies on Kolguev, where the first answers to the following questions were to be received: 1. How many geese and of what species nest nowadays on Kolguev? And what exactly is the importance of this breeding site?2. What factors influence nesting density and breeding success in Arctic areas? 3. Are there behavioural or ecological differences bet-ween goose species that explain the diverse development of population numbers?4. Which goose species do best under modern conditi-ons, and 5. What are the future needs for study?
Finally Dr. Eberhard Schneider, president of the Bird Protection Comitee (Vogelschutz-Komitee e.V. VsK, Hamburg), committed financial help for these issues of goose-migration studies in the Autumn of 2005, thereby enabling a lot of activitiy and scientific work in subse-quent years. During the preparations of the expeditions, the UNEP-GRID ECORA programme was also willing to support our studies on Kolguev, one of the focus areas of the programme. This report presents an overview of the results of 2006, the first study season within this frame-work.
1.1 Breeding distributionIn spite of the fact that this preliminary study focus-es on the White-fronted Goose (Anser albifrons), two other Arctic goose species are encountered at both main research sites: the Western Tundra Bean Goose (Anser fabalis rossicus) and the Barnacle Goose (Branta leucopsis). To obtain a more complete picture, informa-tion on both species will be included in this report.The Tundra Bean Goose breeds on the Arctic tundra from Chukotka in the east to the Kola Peninsula in the west. Within this breeding range we find two subspecies: the Eastern Tundra Bean Goose (Anser fabalis serrirostris), which overwinters in southeastern China and the Western Tundra Bean Goose (Anser fabalis rossicus), which overwinters in Europe (MADSEN et al. 1999, MOOIJ & ZÖCKLER 1999). The boundary between the subspecies seems to be situated on the western part of the Taimyr Peninsula near the Piasina and Pura rivers (BORSHONOV 1986, MOOIJ & ZÖCKLER 1999, ROGACHEVA 1992, see Fig. 1).
The (Greater) White-fronted Goose Anser albifrons is a species of the Arctic tundra and has an almost circumpolar breeding range. The species is divided into a number of subspecies, of which the nominate subspecies, Anser albifrons albifrons, breeds on the Arctic tundra between the Kanin Peninsula and the Chatan-ga and Popingay rivers east of the Taimyr Peninsula, and winters in Europe and southwestern Asia (KEAR 2005, MADSEN et al. 1999, MOOIJ & ZÖCKLER 2000, ROGACHEVA 1992).
The Barnacle Goose Branta leucopsis breeds on the Arctic tundra between Greenland and Vaygach Island and is divided into a number of subpopulations, of which the Russian-Baltic subpopulation is with the one encountered in this study. The Russian-Baltic subpopulation was known originally to breed on the islands of Novaya Zemlya and Vaygach, but nowa-days also nests from the Kola and Kanin Peninsulas in the west to Vaygach and Novaya Zemlya in the east, both on islands, including Kolguev, and on the Russian mainland along the southern shore of the Barents Sea. Since the 1970s the species has also been breeding in Sweden and Finland, as well as along the Estonian coast, on some Danish islands, and in The Netherlands and Germany (HAGEMEIJER & BLAIR 1997, GANTER et al. 1999, MADSEN ET AL. 1999, HUSTINGS & VERGEER 2002). The species is expanding its breeding

5
range to the west at an astonishing rate, but the cur-rent status of the breeding populations of Vaygach and Novaya Zemlya is unknown (K.E. LITVIN, pers.comm.).
As far as we know, the Tundra Bean Goose Anser fabalis rossicus nests from the Timan-Tundra to eastern Taimyr. While breeding numbers increased in the western part of this range (Timan Tundra to Yamal) until the 1990s, the numbers decreased further east (TOMKOVICH et al. 1994). Wintering numbers of Beans seemed to be more or less stable in the last 20 years (MADSEN et al. 1999), although there are some hints for a slight decline since the mid-1990s. Un-fortunately, there is no clear trend, because of gaps in the current count network and confusion between subspecies in some areas (MADSEN et al. 1999, KEAR 2005).During the past century we established a rather good understanding of the breeding range of several goose species (MADSEN et al. 1999). But these breeding ran-ges seem to be fairly dynamic, and a number of sites, known to host a high number of breeding pairs of a particular goose species decades ago, are deserted today, whereas other sites show an almost explosive expansion of a breeding goose species, unknown for the site a few years previously. Especially given the scope of the enormous ecological changes expected in connection with the warming up of Arctic regions in the coming decades, further changes in the bree-ding range of Arctic species are inevitable. A number of Arctic goose species could lose considerable areas
of their present breeding range (ZÖCKLER & LYSENKO 2000). Therefore, for the conservation of Arctic species, it is of extremely great importance to put in place a permanent monitoring scheme to record all changes, in order to be able to react in time to develop effective conservation measures.
1.2 Stopover sites – key points for successful reproduc-tion and for human impactThe importance of spring staging areas for successful reproduction of Arctic-breeding migrating avian species has received thorough attention in recent decades, so that several theoretic models have been proposed to under-stand the spring migration strategy of Arctic geese via the network of staging sites, where the energy resources, necessary for reproduction are gained (DRENT & DAAN 1980, VAN DER GRAAF 2006, EICHHORN 2008, EBBINGE & SPAANS 1995). At the same time, the relationships between natural causes („Green-wave” hypothesis, VAN DER GRAAF et al. 2006) and human impact are still poorly studied. Nonetheless, it is clear enough that human disturbance may play an important role in shifting geese between fee-ding habitats during intensive feeding in spring (PRINS & YDENBERG 1985). The majority of Arctic geese breeding in the Russian Arctic stage nowadays on agricultural lands, where they depend both on the habitat structure of the agricultural fields and on disturbance factors caused by different types of human activities. Thus the urgent need for such kind of a stopover-study has become more and more clear.one can hardly count more than 10 studies devoted to any kind of population estimates of geese living there,
Figure. 1: Breeding distribution and wintering grounds of White-fronted Geese (all subspecies, from: UNEP-WCMC)

6
birds as well as from birds banded in The Nether-lands and Germany, we learned that White-fronted Geese regularly shift between different wintering sites (KRUCKENBERG 2002, KRUCKENBERG & WILLE 2005). The recordings of neck-banded birds also sho-wed that these individual birds used a wide range of migratory routes between the breeding and wintering grounds and vice versa, which did not always corres-pond with the shortest connection between the two areas (KRUCKENBERG 2002, MADSEN et al. 1999, MOOIJ 1993,1997).Barnacle Geese colour-marked (two coloured leg-rings with a digital code) and satellite tracked (EICHHORN 2008) in Western Europe have shown for the Russian-Baltic population a migratory route over the Baltic and White Seas to breeding grounds on the Kanin Peninsula, Kolguev Island, the southern Barents Seashore, and Novaya Zemlya and Vaygach Islands. All these birds seem to have their wintering grounds along the Wadden Sea coast of The Nether-lands, Germany and Denmark (MADSEN et al. 1999).In spite of these findings, there is a considerable lack of knowledge about the migratory routes, especially of White-fronted and Bean Geese, as well as about the way the different wintering grounds are used by different goose species and how important a certain wintering site is for a particular species.
and such population parameters as total numbers, patterns of distribution, and breeding success remai-ned unknown. At the same time it appeared clear, that the total numbers of geese that nest on Kolguev might be high enough to play an important role in the entire western Palaearctic population of at least three goose species.
1.3 Wintering distributionBased on the results of previous goose-marking projects (e.g., BORSHONOV 1986, BURGERS et al. 1991, EBBINGE 1991, EBBINGE & ST.JOSEPH 1992, KRUCKEN-BERG 2002, KRUCKENBERG & WILLE 2004, MADSEN et al. 1999, MOOIJ 1993, 1997), it is known that Arctic geese breeding in the Western Palearctic winter in Europe; Branta species along the sea coasts of Western Europe and Anser species at a number of inland sites.Tundra Bean Geese ringed with metal rings in The Netherlands were recovered from nesting grounds on the northern Russian tundra (BURGERS et al 1991), whereas some Tundra Beans ringed on the Taimyr Peninsula were recovered at wintering sites in Eas-tern Europe (BORSHONOV 1986).White-fronted Geese ringed in The Netherlands were recovered from most Arctic breeding sites of the Russian Tundra west of Khatanga River, whereas birds ringed with leg and neck-bands on the Taimyr Peninsula were recovered on most European winte-ring grounds (GANTER et al. 1999, MOOIJ 1993, 1997, DEKKERS & EBBINGE 2004). From these neck-banded
Figure 2: To ob-tain more precise waterfowl data, more than 10 000 volunteers count birds simultane-ously

7
1.4 Stop-over sitesDuring migration between the nesting and wintering grounds. geese use in autumn and spring a variable number of stop-over sites. The birds need these areas for intensive feeding, in order to replenish the fat reserves used up by the preceding flight, as well as to store energy for the subsequent flight. At most stop-over sites, individual birds stay as little as a few days or as long as several weeks. It seems that stop-over sites near the nesting areas are extremely important for breeding birds to fatten-up and to reach an optimal condition for nesting. Although a considerable number of stop-over sites is known, there is a considerable lack of knowledge about the importance of each of these sites for the different goose species. Furthermore, the duration of stay of individual geese at important sites, the time needed for crucial fattening-up and the need of protection of each of these sites remain unknown.
1. 5 Population estimatesBased on the results of international midwinter goose counts, every three years an estimate of the winter-population size of all goose species is made by the experts of Wetlands International (MADSEN et al. 1999, MOOIJ & ZÖCKLER 1999, 2000, WETLANDS INTERNATIONAL 1994,1996, 1997, 2006, Fig.3).For the Western Tundra Bean Goose, the popula-tion is estimated to be stable at a level of 500,000 – 600,000 individuals, which would mean a breeding population of 100,000 – 150,000 pairs.For the Greater White-fronted Goose, during the last 10 years the size of the Western Palaearctic population in winter has been estimated at between 1.0 and 1.2 million birds (EBBINGE in comm.), which would mean a breeding population of about 300,000 - 400,000 pairs. WETLANDS INTERNATIONAL (2006) considers the population size to be stable, but a decline of breeding success since mid of the 1990th suggest declining population as well (KRUCKENBERG & MOOIj 2008). For the Russian-Baltic population of the Barnacle Goose, a clear increase has been documented and, according to the latest estimate in 2006, is now 420,000, which would mean 80,000 – 100,000 breeding pairs. In spite of these triennual population-size and trend estimates, there seem to be considerable discrepancies between these estima-tes and our population estimates from the different breeding grounds.
2. Flyway approach to assess population status and recent trends
In the scope of AEWA and the EU-Birds Directive for all goose species, Species Management Plans should be developed to keep the species in a favourable conditi-on and to organize sustainable management for each of these populations. Before such plans can be developed it is crucial to have better data on the size of the bree-ding and wintering populations of each species, as well as on population trends, migratory routes, the situation on the breeding grounds, stop-over sites, and wintering grounds (protection status, current and potential threats, hunting pressure, etc.). Based on these data, it would be possible to develop Species Management Plans effective in maintaining favourable population levels. For most species, this basic information is still poor and surely not enough to make sustainable management of these popu-lations possible. The most effective way to fill in these knowledge gaps is to collect the data for each species on a flyway basis.As part of the flyway approach for the White-fronted Goose in the first and preliminary phase of the project, this study is divided into three areas of studiey for field research:
1) Spring migration using neckbands and satellite transmitters2) Stop-over site examination (Olonets fields in Karelia, Russia)3) Breeding-ground studies (Kolguev Island in the Nenets Autonomous Region, Russia)
This study is part of the GEF ECORA project (Integra-ted management approach, area of special interest for biodiversity) and part of the agenda of the Vogelschutz Committee Germany to supply research on long-distance migratory birds for better protection of the species and their staging sites.
Case Studies1) Spring migration To study the migration of wild birds, a methodology had to be developed to recognize important staging sites as well as to make it possible to follow some of these birds individually. During the past 30 years several methods were developed, but the best results may be found by combining all three methods commonly in use today.

8
MethodologyA. Local observations (bird counts)Since 1956 regular counts of waterfowl have been estab-lished in Western Europe. Today a network of volunteer observers covers most staging sites in wintering areas. During internationally coordinated synchronous censu-ses, waterfowl counters cover the entire wintering area. The results are in a central database to estimate the popu-lation size of all migratory waterfowl species. With this method it is possible to record waterfowl concentrations, but without any information about where they come from or where they go to afterwards. A further problem of the current system is that more-or-less complete wa-terfowl counts are carried out in most Western European nations, but there are still enormous gaps in southern and eastern Europe. Because of the lack of observers, tech-nical infrastructure and funding in these countries, the counts are stretched over weeks instead of days, and the results are thus extremely questionable. Because of these problems, the results of the Internatio-nal Waterfowl Census of Wetlands International despite being the best we have ever had, are still very questio-nable and a poor tool for the sustainable management of these populations.
B. Banding (neckband, metal)Since the 1930s, an increasing number of birds have been individually marked with metal rings on the leg. The different combinations of digits on these rings make it possible to re-encounter individual birds, e.g, when they are recaptured, found dead or shot. With high quality optics, in some cases it is even possible to read such rings on living birds. But from most birds this marking method only produces data about the moment they were ringed as well as the moment they died or were recaptured, but does not give any infor-mation about the whereabouts of these birds between the two events. This gap can be filled by the use of neckbands, with an alphanumeric code, which can be read at a dis-tance with the help of a spotting scope or binoculars. Such neck bands enable the gathering of a lot of in-formation on the whereabouts of marked birds thoug-hout their lifetime, provided there are people looking for these birds and reporting their observations. With the help of regular intensive recording of neckbands at a given site, it is also possible to calculate how long the marked birds use that specific site (see Fig. 3).The weakness of this method is that a network of re-
Figure 3: Neckbands used on White-fronted Geese since 1986 (1, 2 & 3 = EWRP, 4 Kostin & Mooij Taimyr, 5 E. Rutschke Lake Gülpe, Brandenburg)

9
gular observers currently only exists in some Western European countries. In most of southern and Eastern Europe, such a network still has to be developed. Furthermore, in most Eastern European countries the birds stay in such remote areas that they can hardly be expected to be observed, even if an extremely dense network of observers could be build up.
C. Tracking (satellite, GPS)To fill in the gaps in knowledge about the move-ments and migration of animals even in remote areas, the method of tracking animals with transmitters was developed. In this method a transmitter is fixed to the animal and the signals from the transmitter are tracked by receivers on the ground or by satelli-tes. At first these transmitters were too heavy to be attached to birds, but in the 1970s transmitters were made light enough to be used on big birds. Since the 1980s the weight of these devices has been further reduced so that since the 1990s they could be fixed on medium-sized birds like geese.
At the beginning of their technical development, trans-mitters could be followed only on the ground and be tracked at a distance of up to 10 km by means of a special receiver. The position of the transmitter was determined by cross-tracking with two receivers at different positions. Since the development of the satellite technique, trans-mitters have been developed whose signals are received by satellites. With this technique, the position of the transmitter-bearing animal is calculated from the time difference between the recording of the transmitter signal by different satellites. Only when the signal is received by more than two satellites is it possible to fix a more-or-less reliable position of the transmitter, but never to within less than about 10 km.Recently transmitters using GPS were developed. These devices can fix the position of the transmitter-bearing animal to within as little as 15 metres. The relatively light weight (35-45 grams) of these high-tech devices makes it possible to use them on medium-sized birds like geese (Fig. 4). The use of this kind of transmitter enables scientists to get exact location information on migratory
Figure 4: A Whitefronted Goose with neckband and satellite transmitter

10
routes, and staging and breeding areas, as well as on the use and period of use of a particular staging area by the transmittered birds. Such information is crucial for the development of effective conservation strategies. Additionally, Platform Transmitter Terminals (PTTs) have been equipped with an FM-transmitter to allow fieldworkers to find it on the ground. This option is used regularly to search for birds on the nest, for example. In our case, we used this ground-tracking (GT) only in the
Season start date end date start hour end hour hour step TX duty GT on?1 Feb 16 Jun 15 5 24 2 3 no2 Jun 16 Aug 15 4 22 6 7 no3 Aug 16 Nov 15 6 21 2 6 no4 Nov 16 Feb 15 6 21 2 3 no
Table 1: The programmed schedule of PTTs in 2006
TX = intervall between recordings, GT = Groundtrack
Figure 4: Population sizes and trends in Whitefronted (A) and Barnacle Geese (B)
A
B
case of death; the ground tracking was turned on by a mortality switch. If the PTT hadn’t moved for more than 30 hours, the mortality switch was activated. This also appeared in the satellite-transmitted data. So, we`d know bird had died or lost its transmitter. Transmitters were programmed with the transmission schedules shown in Table 1.
2.1 Results: population estimates and trendsa) International concurrent counts White-fronted Geese are the most numerous species of goose wintering in the Western Palaearctic. The current population size is estimated at about one million birds wintering in Western Europe, and a further 10.000 – 40.000 in the Pannonian, 350.000 – 700.000 in the Pontic-Anatolian and 15.000 in the Caspian parts of the wintering area (WETLANDS INTERNATIONAL 2006). So, altogether a maximum of 1.36 – 1.75 million Whitefronts are breeding in the Western Palaearctic at present. While the population status of most of the wintering regions is stable, the trend in the Pannonian region shows a clear decrease. On the other hand more detailed analyses of the current data show a lot of gaps in the internatio-nal counts and very nonconcurrent dates for these counts, so some additional methods have to be used to check the validity of the international estimates (see fig. 4).
b) Results of banding with neckbandsSince 1998 more than 9,000 Whitefronts have been marked individually by using black- and lime-colou-red neckbands. Fig. 5 shows the distributon of repor-ted sightings of these marked birds. Obviously most of these sightings were made in The Netherlands, Belgium and Germany, where most of the active bird-watchers live. However, some remarkable observati-ons were made in England (the most western part of

11
Figure 6: Neckbanded Whitefronts reported as shot (1998 - 2005)
Figure 5: Reported observations of neckbanded White-fronted geese (2004/05)

12
wintering area), the Baltics and Russia. Furthermore, a lot of marked birds were seen in Hungary and a few in Kaz-akhstan and Turkey. So, the reports of marked birds show some emigration ringed in The Netherlands and changed to the southeastern European wintering areas, but most of the birds wintering in the western part of Europe and returning there year after year. It seems noteworthy to remark that observation density decreases the more the birds migrate to the northeast. As a consequence, neck-banding is not the optimal method for investigation of goose migration over the last stretch before the northern nesting grounds. The high value of colour marking in geese comes from the large number of individuals that can be marked. Spe-cial analyses to calculate mortality, survival rates, bree-ding success and individual body condition in relation to breeding success or disturbances can be done for co-lour-marked birds. To find out detailed migration routes, however, marked birds can give us only some hints when reported as dead (shot) birds (Fig. 6). Density of obser-vers decreases markedly east of the German border, and the infrastructure is often too poor to allow goose ob-servers numerous sightings of marked birds. The further north the bird migrates, the less probable that it will be sighted. So, additional methods are required to study the routes of the migratory geese.
c) Results of tracking Starting at the end of January 2006, five male Whitefron-ted Geese were caught in The Netherlands and fitted with Solar-GPS-Transmitters (Microwave 100PTT 45g) and released again. These birds were named after the Dutch goose catchers who did the catching work using an old, traditional method with living decoys and clapping nets (EBBINGE 2000). Because of the very cold spring tempe-ratures, the geese started migration very late in 2006: at the end of March and the beginning of April. Most of them stayed very close to the release sites. Only one bird (“Bouke”) moved from De Wielen near Leeuwarden further to the Afsluitdijk and De Petten (Noord-Holland). In early April, transmission from the first bird (“Alco”) stopped. This bird was staging in the Lauwersmeer area and was seen several times. No technical problems had been visible, so different reasons are possible: predation, or illegal hunting, for example. For sure, the bird wasn`t shot within the official crop-protection-programme area.Three birds started migration at the end of March (“Har-ry”, “Adri” and “Bouke”: see Fig. 7). All of them went to the eastern part of Germany first, staging in well known
areas at Elbe, the fishponds of Linum and the Oder lowlands. After some days of roosting, Harry and Bouke went to the Rusne area (Lithuania) while Adri flew to East Poland, then turned southeast to cross Belarus and reach the Russian-Ukrianian border area 90km from Kiev. In May, Harry and Bouke went north through the Baltic states and Karelia and reached the Archangelsk region, where Bouke was shot by a local hunter on 14-05-2006. The signal from Harry stopped on 28-05-2006 near the Mezen River, east of Archangelsk. This PTT was able to be retrieved in August and it was shown that this bird had also been shot, but had not been collected by the hunter. PTT “Evert” was roosting with a flock of Greylag Geese on the Eempolder and turned to Oostvaarders-plassen later on. Maybe because of a breeding White-tailed Eagle is this area, Evert (and probably the other Greylags, too) migrated to Brandenburg, where Evert was staging there in the nature reserve “Koblentzer See” near Parsewalk. In June the signal of this PTT deteriorated, and finally stopped on 17-06-2006. Jochen Bellebaum visited the Koblentzer See with Winfried Krämer (of the nature conservation admi-nistration) in July. They found a suitable site for wild geese on flooded grasslands of a re-created wetland. We assume that the bird was moulting there. The PTT might just as well have stopped due to technical reasons as by death of the bird (probably killed by a fox or an eagle). The most successful tracking was that of PTT “Adri”. This bird migrated via Brandenburg and Poland to Ukraine at the end of April. In mid-May it migrated straight through the north of Russia and came to southern edge of the Kanin Penninsula on 27-05-2006. On 9-06-2006 Adri was located during migra-tion on theVaygach Peninsula and reached the Yamal Peninsula on the same day. There the bird stayed for more than three weeks. Probably it tried to nest there or just fed well for the next migration step. On 2-07-2006, Adri started to migrate eastwards again and was tracked over the Taimyr Peninsula on 3-07-2006 at 12 o`clock (noon). On 8th July Adri reached East Taimyr Lake, where he was moulting until 28 August. Then Adri started autum migration, during which he flew further inland to south of theYamal Penin-sula (13-09-2006) and reached the Ob River on 26 September. Then Adri migrated south along the Ob River, turned southwest on 29 September and went

13
PTT ID Start date End date Route Length No. datasetsHarry 62348 25-2-2006 21-7-2006 3 990 790Alco 62349 19-1-2006 5-3-2006 391 211Adri 62350 8-2-2006 9-11-2006 13 311 1 586Bouke 62351 23-1-2006 14-5-2006 3 966 697Sasha 62351(2) 3-8-2006 5-8-2006 4.2 124Evert 62352 5-2-2006 21-6-2006 1 001 677
Table 2: Migration dates and distances of satellite-tracked White-Fronted Geese in 2006
to the Volga River (21-10-2006). Then he migrated in a westerly direction on 31-10-2006 and flew via Ukraine and Belarus to Poland, where the signal was lost on 17-11-2006 near Lodz. In the days just prior to this, the data quality of PTT deteriorated signi-ficantly, so the reason for loosing the PTT is quite unknown. The bird wasn`t seen during the following winter season, so it might be dead. By re-using the PTT of “Bouke”, another bird was fitted with a transmitter at the beginning of August during a scientific expedition to Kolguev. This was the first time we used a standard teflon harness in-stead of a harness made from leather like the Dutch
Figure 7: Spring migration of satellite-tracked Whitefronted Geese in 2006 (yellow = Adri, magenta = Harry, blue = Bouke)
goose catcher use. The bird was tracked for 10 days, after which the signal remained at one position and sent out the GT-signal. Two actions were undertaken to rescue the PTT in 2006, but both were unsuccessful. The first time, the helicopter hadn`t enough fuel to reach the Pechanka River valley and the second time the NW part of Kolguev was covered with snow. Also two further actions to get the PTT in 2007 and 2008 failed. The bird itself (“Sasha”) was encountered six times during the subsequent winter season (2006/07) by volunteer neckband observers. So, in fact, the goose itself had damaged the harness and kicked off the transmitter. Because of the very interesting results of the first satellite

14
tracking of Whitefronts, Vogelschutz-Komitee e.V. and the Dutch Ministry of Agriculture and Environment gave financial support to continue the satellite tracking in 2006/07.By using Whitefronted-Extension (JENESS 2000) for Arc-View 3.2, step-by-step distances of the migration of each PTT could be calculated as well as the total length of the migration. Table 2 (page 15) shows the overall length of the routes along which the birds were tracked in 2006.By using Microwave GPS-transmitters, we gain some additional information about the PTTs by satellite. The PTT also takes measurements of actual altitude and speed during the fix. So, Fig. 8 shows the recorded altitudes only of flying birds in relation to speed In all categories, speeds of more than 70 km/h were reached, but there is a trend to higher speed at heigher altitudes. So, we think that two different categories of flight are shown: those made during stays in staging areas, maybe morning flights or changes of feeding grounds, and those which are migration flights. Most local flight are made for altitudes of less than 100m and at speeds of 10-50 km/h, whereas the others are above 100 -1000 m altitude and at greater than 70km/h. Speeds greater than 100 km/h were only found at altitudes above 250 m; the maximum speed was 110 km/h. Maximum altitude was fixed at 928 m.
Figure 8: Flight height and speed of satellite-tracked Whitefronted Geese
Conclusions As a result of the preliminary tracking of the White-fronted Geese, we have found migration to consist of a series of brief but long-distance flights interspersed with long breaks of 10-30 days in which the birds to replenish their fat reserves. For this process, step-ping stones are needed at distances of 800-1,300km. There seems to be a large variety of actual routes of migration for the birds. We see this in the results of the observations of neckbanded geese during the wintering period (KRUCKENBERG 2002). More than one strategy seems to exist to cope with the hunting pressure in Russia. While Harry and Bouke tried to move through Russia right away and very fast, Adri lingered in a remote part of Ukraine until the end of the hunting season, and then moved north very quickly (KRUCKENBERG et al. 2007, 2008). In all three cases the social status of the bird is fairly unknown and we don`t know the exact energetic limits of the individuals. In fact, birds preparing to breed the next season will have rather different restrictions in their time-and-energy budgets than non-breeders. The latter group has lots of time to migrate to the moul-ting sites, whereas breeding birds are very closely confined by their biological schedule as well as that of the climate.

15
3) Stop-over site (Olonets fields)
Olonets is the administrative centre of the Olonets District and the oldest documented city in Russi-an Karelia. It is situated on the Olonka River, east of Lake Ladoga and has about 11,000 inhabitants. Around the city lies an extended agricultural area of about 46,000 ha, which can be described as an agricultural island in a sea of taiga forest and bogs. Although small numbers of geese use the traditio-nal habitat of the bogs as a feeding site, most geese concentrate on the grasslands and fields for spring feeding. After the break-up of the Soviet Union at the end of the 1980s, the agricultural system col-lapsed. Agricultural use of the area was reduced and an increasing part of the grasslands was not used at all for agriculture, or used much less intensively. As a result, areas of the fields and grassland became overgrown by shrubs, and the numbers of staging geese decreased. As part of a nature conservation project of WWF-Sweden to restore goose-feeding sites, local farms were given support to return to former farming practices. In its second phase the project also was supported by the Baltic Fund for Nature, based in St. Petersburg. As a result of this project, agricultural production has almost doubled, income of the farm employees has increased considerably and goose numbers have increased again. Since the middle of 1980th, the staging geese have become very popular in the Olonets region, even with the farmers, and the city of Olonets organizes a goose festival every spring. A temporary hunting free goose reserve has been founded to protect the geese against the extre-mely intensive goose hunting in spring. Wild geese only visit the Olonets region in spring, when they use the grasslands for fattening-up before flying to the breeding grounds. The area is a stop-over site for migratory White-fronted Geese (Anser albifrons), Bean Geese (Anser fabalis) and Barnacle Geese (Branta leucopsis). The main staging period is from the middle of April until the end of May each year. For a number of years the number of staging geese in the reserve were regularly counted by Russi-an scientists from the Karelian Centre of the Russian Academy of Sciences. Because of limited mobility, technical equipment and financial budget, these scientists were not able to do an intensive census for neckbands nor to collect data about turn-over rates,
staging periods, proportions of immature bird (yearlings), etc. But these data are extremely important for the deve-lopment of an effective conservation strategy.In the scope of this preliminary study a first effort was made to close these gaps. For this reason a group of four scientists from Germany as well three from the University of St. Petersburg carried out an intensive goose census in the Olonets region between the 24th of April and the 12th of May in 2006. A freelance reporter from German radio accompanied the German scientists to report live about their work in the Olonets fields.
MethodologyIn order to have optimal mobility in the field, the Ger-man team (H. KRUCKENBERG, J. MOOIJ, M. FANCK, H.-H.BERGMANN, K. TOSS and C. ZÖCKLER) travelled to Olonets with two 4-wheel-drive vehicles and the Russi-an crew (A. KONDRATYEV and E. ZAYNAGUTDINOVA) also brought one 4-wheel-drive car from St. Petersburg. With these vehicles, the team was able to carry out a daily cen-sus of neckbands and estimate of the number of staging geese inside and outside the reserve, as well as to collect data about hunting pressure, hunting practices and hun-ting bags outside the reserve. Because of the need to visit hunters in the field, and because of the poor condition of secondary and lower-classified roads in Russia, 4-wheel-drive vehicles were absolutely necessary.To record the numbers of neckbanded geese, the regi-on was divided into three zones, each covered by one crew with a 4-wheel-drive car. Each morning and eve-ning, when the conditions for finding marked birds were optimal, all zones were searched for neck-banded geese. Once a week, in co-operation with the scientists of the Karelian Academy of Sciences, all geese in the region were counted. During the hunting season, 30 April th-rough 9 May, the team counted shots and visited hunting groups to check hunting bags and to interview the hun-ters about hunting practices. Between these activities, the team met with the project coordinators of WWF-Sweden and the Baltic Fund for Nature (BFN), as well as with the head of the administration of the Olonets Region, to discuss the future development of the goose reserve.
3.2 Numbers and distribution The total numbers of geese using the Olonets fields were estimated from the total counts made on a daily basis by car surveys. This way of counting is a routine research method of the study team of the Karelian Biological Research Institute. We joined their surveys three times, counting also in fields outside the hunt-free zone, parti-

16
cularly in the vicinity of the villages of Tuksa, Jurgelitsa and Megrega. Maximum numbers were registered on 29 April: more than 17,000 geese altogether (90% White-fronted and 10% Bean Geese).Outside the hunt-free area, the geese were found on 27 April on fields east of Tuksa village (ca 500 White-fronts), and also on 28 April on ploughed fields near the villages of Jurgelitsa (ca 4,000 geese, 50% of them being Bean Geese) and Megrega (ca 600 geese). These 4,600 geese must be added to the 10,000 found in the hunt-free zone (Fig. 9).But the following day, 29 April, as the hunting season approached and hunters started to gather around the fields of Juregiltsa and Megrega, these fields were deser-ted by geese, despite the fact that the hunting season was still closed. This may explain the drastic increase in geese numbers in the hunt-free zone observed during total counts on 29 April (Fig. 9). On all other days after 29 April, geese were observed only within the hunt-free zone. Intensive transit migration was observed on 5 and 7 May, when flocks of 70-100 geese flew over the Olonets fields. Newly arrived geese were distributing themselves
in different ways to former flocks, occupying new areas within the sanctuary. Some geese also departed during that time. Therefore the numbers sometimes changed during the day. Thus, on 7 May, during the morning counts, when the geese had arrived from their roosting places on Lake Ladoga, where all geese spend the night, about 19,000 geese were counted. But during the evening car survey, only 13,000 geese were counted on the fields. This could be explai-ned by the departure of some geese with the transit flocks.
3.3 Duration, turnover rate, and percentage of immatures of White-fronted GeeseDuration and turn-over rate were estimated based on the records of marked birds. Over a period of 15 days between April 26 and May 13, a total of 38 marked geese were observed at least once. Not surprisingly, most of the geese were only observed once. Howe-ver, six individuals were observed over a period of at least 10 days and one pair over the entire period. Our sighting data enable us to calculate that birds
Figure 9: Goose numbers in the hunt-free zone (Olonets Fields Bird Sanctuary)
Figure 10: Percentage of White-fronted geese in the Olonets fields that were immature (Whisker-Box-Plot, n=68)

17
stayed for an average of 4.8 days at the Olonets Fields. By using biostatistics programme MARK, we calculated a 9.4% (± 2.1%) exchange of individuals per day. Through analysing the data, we found that the likelihood of a neckband being seen was 35,2% (±4,1%). We estimated that more than 300 000 birds might use the Olonets Field during spring migrati-on. Complex estimates are possible using program-me MARK to estimate turnover-rates, numbers of staging birds changing daily, migration density, etc. Such calculations will be made during the next step of analysis. On four days between 29 April and 9 May, the percentage of immature birds in the goose flocks was recorded (Fig. 10). During this period this percenta-ge varied between mean of 19.54% on 29 April and 20.7% on May to 18.0% on 9 May. The average was 19.4%. This percentage is considerably lower as the percentage of about 26 % found in the Netherlands, Flanders and Germany in during winter 2005 / 06 (K. KOFFIJBERG, pers.comm.).
3.4 Human impact on staging geese Different kinds of human activity influence geese staging on the Olonets fields. The most important factors are hunting and agriculture. Whereas geese more-or-less benefit from agricultural activities, goo-se hunting scares geese from feeding and roosting sites (see below). In the agricultural area, geese bene-fit from the open grasslands used by local farms for cattle grazing as well as for fodder. The geese mainly incorporate the protein fraction of the vegetation during metabolism. Therefore young grass with a high protein content is more valuable for geese than older grass, which has low levels of indigesti-ve proteins. To breed successfully and migrate far distances geese are forced to optimize their intake rates and the protein content of food to fat up very fast. So, for the replenishment of fat resources during migration, the availability of high quality food and a low level of disturbance are important factors. Inside the Olonets goose sanctuary the geese are fairly well protected against human disturbances because human access to the sanctuary is limited during the hunting period. After arrival the geese mainly feed on young grass between the stubble of the previous year.
3.5 Impact and intensity of hunting3.5.1 Impact of hunting Goose hunting is practised only in spring in the Olonets region because it is only then that considerable numbers of geese staged here. In spring 2006, hunting occurred during the morning and evening flights, at the feeding sites and even at the roosts, which not only caused enor-mous disturbance, and made the geese more shy, but also considerably influenced other behaviours (WILLE 2000) and had a negative impact on the overall condition of the birds (MADSEN 1995) . Hunting during spring has the greatest impact on hunted populations not only because the birds are heavily distur-bed during the phase of fattening-up to make them fit for migrating and breeding, but also because stable breeding pairs may be separated temporarily or even permanently right before breeding starts. Geese regularly establish new pair bonds in autumn and winter. Birds losing their partner in spring may be unable to find a new partner immediately and will be lost to the reproductive effort of that year. Furthermore, they likely will have a lower reproductive success with a new partner in the first years of the new partnership (BERGMANN et al. 1994, OWEN & BLACK 1990). Birds crippled and wounded during hunting, of which there are a high number (see NOER & MADSEN 1996), will not be in optimal condition to breed, which, without doubt, will also reduce the overall reproductive success of the hunted population. Therefore it can be sta-ted that spring hunting not only influences the population through direct withdrawal of individuals, but through indirect impacts that reduce reproductive success. These indirect impacts are clearly stronger from spring hunting than from autumn hunting and can result in the consi-derable decrease of a population in only a few years (G. GAUTHIER, pers.comm 2002).
3.5.2 Intensity of huntingFor the Olonets region in spring 2006, an average of 300 shots per hour during the morning flight were counted from the base camp in Tuksa. Shooting started early during this flight, under rather poor light conditions, and proceeded for several hours. After morning hunting most hunters ate breakfast, and then slept. The rest of the day until evening irregular shooting could be heard. In the evening hours a second phase of intensive shooting started, and lasted up to two hours. Around Tuksa at least 10 hunting parties with up to 10 hunters in each were active. These hunters were all resi-dents of St. Petersburg and Moscow and legally rented a

18
hunting area for 5,000 Russian roubles per person. The greatest hunting pressure was reached during the week-end of 1 May (30 April to 2 May). During this holiday weekend, the highest number of hunters and the highest number of shots were counted. A second, but conside-rably lower, peak was recorded during the last weekend of the spring hunting season (6-9 May). On the 18 days between these weekends, most hunters left the area and the intensity of hunting was low.
3.5.3 Temporal and spatial dimensions of hunting During the first three days of the hunting season, as el-sewhere all over the globe, hunters at all known hunting sites were very active. In addition to known (rented) hunting places in the Tuksa fields, many local hunters were using additional places outside the vicinity of these fields. So, for example, hunting was recorded near the Tuksa forest. During beginning of the hunting season we counted shots during the morning flight. As a result we found a really high hunting pressure with more than 300 shots per hour in the Tuksa fields only. By using a range-finder Leica Geovid we measured flight altitute during the morning flight. Most of the geese flew higher than 100m. Nevertheless, hunters in fact killed many geese by using special gun pellets. After three days, hunting pres-sure decreased rapidly, owing to the dramatic decrease in the number of geese in the morning flight. In fact, after the initial days of the hunting period, geese were no lon-ger found outside the sanctuary. So, most hunters broke camp and went home for the rest of season. We also encountered illegal goose hunting: close to the nest of a White-tailed Eagle (Haliaeetus albicilla), and, during a night excursion, hunting on a night roost (bog). So, the geese are hunted the whole day and then intensi-vely on their night roost. Illegal hunting was also repor-
ted by different local birders. During checks of the remains of hunting at the hunting sites, we found the remains (for example, feathers and bills) of many protected species, such as Whimbrel (Numenius phaeopus), Curlew (Numenius arquata), and Snipe (Gallinago gallinago, Fig 12).
3.5.4 Behavioural response to hunting pressureAll along the flyway staging areas for geese often attract recreational activities such as boating and hunting. A number of studies have revealed that dis-turbance due to human activity may result in incre-ased energy expenditure or loss of feeding habitats in geese and other birds (MADSEN 1985, RIDDINGTON 1996, HÜPPOP & GABRIELSEN 1999). Escape or reaction distance has been widely used to assess the impact of human disturbance, especially hunting activities, on geese and other waterfowl (e. g. OWENS 1977, MADSEN 1985, LAURSEN et al. 2005). Geese react rapidly to hunting. So, for example, du-ring the morning flight in the current study, once the hunting season opened, the geese flew much higher to feeding areas than they had in the preceding days. Furthermore, the geese then fed in the sanctuary only and showed significantly higher flushing distances on hunting days. Flushing distances inside the bird sanc-tuary (see Fig. 11) decreased with decreasing hun-ting activity outside the reserve during the hunting
Figure 11. Flushing distances of geese measured inside the bird reserve Olonets Fields
Figure 12. Illegally shot Common Snipe (Gallinago gallinago) found near a hunting camp in the Tuksa fields

19
period., No geese were found outside the sanctuary after the start of the hunting season. However, the number of geese staging within the sanctuary did not increase dramatically, so we assume that many geese left the area and flew northward to avoid hunting.
3.5.5 Hunting practice and methods During our stay in the Olonets region we inter-viewed a number of hunters about their hunting practices. We observed hunting during the morning. The exact positions of inspected hunting camps in vicinity of the village of Tuksa are marked on the following map by red dots (see Fig. 13). All hunters arrived at their camps by evening of 29 April, prior to the opening of the hunting season the following day, and most of them stayed until 2 May, then returned home before coming back to the fields for
the closing of the season (8-9 May). Only a few camps re-mained inhabited during the whole period. Only the first two mornings were successful for the hunting camps. On being asked about their hunting technique and success, most hunters reported willingly and even showed us their harvest. They said they on average took 10-15 shots per morning, and on average bagged one goose per hunter per day.The overall result was, that most hunters showed a rather poor species knowledge, just being able to separate “gee-se” from other species groups, which did not hinder them from shooting ducks and waders, which is not allowed on the Olonets fields. Furthermore they started to shoot at geese flying at heights between 60 and >120 meters with lead shot (pellet size 4.5-5.0 mm), which makes it very likely that a high number of birds were injured, rather than killed outright. One group of hunters related that in addition to bagging eight birds, they crippled four geese. Considering the long shooting distance, this is certainly is a heavy underestimation! The average number of shots was said to be 25-35 per hunter per day. To attract geese in front of their hides most hunters used decoys. Besides plastic and metal decoys, which are officially allowed, some hunters used acoustic or living decoys, which are forbidden. There was also some use of automatic wea-pons, which is also illegal.We did not meet with any kind of hunting inspection in the hunting areas. Only the bird sanctuary was controlled by police and municipal hunting inspectors, in order to ban illegal hunting there. An investigation of the hunting bags of five hunting parties with 27 hunters on 30 April, as well as 1 and 3 May, showed that these hunters bagged a total of eight unspecified geese, 17 Bean Geese, and 16 White-fronted Geese, as well as at least four Black
Figure 13. Hunting sites in the Tuksa fields found by expedition partici-pants during the spring hunt of 2006
Figure 14. Hunters waiting close to plastic decoys for wild geese to shoot
20
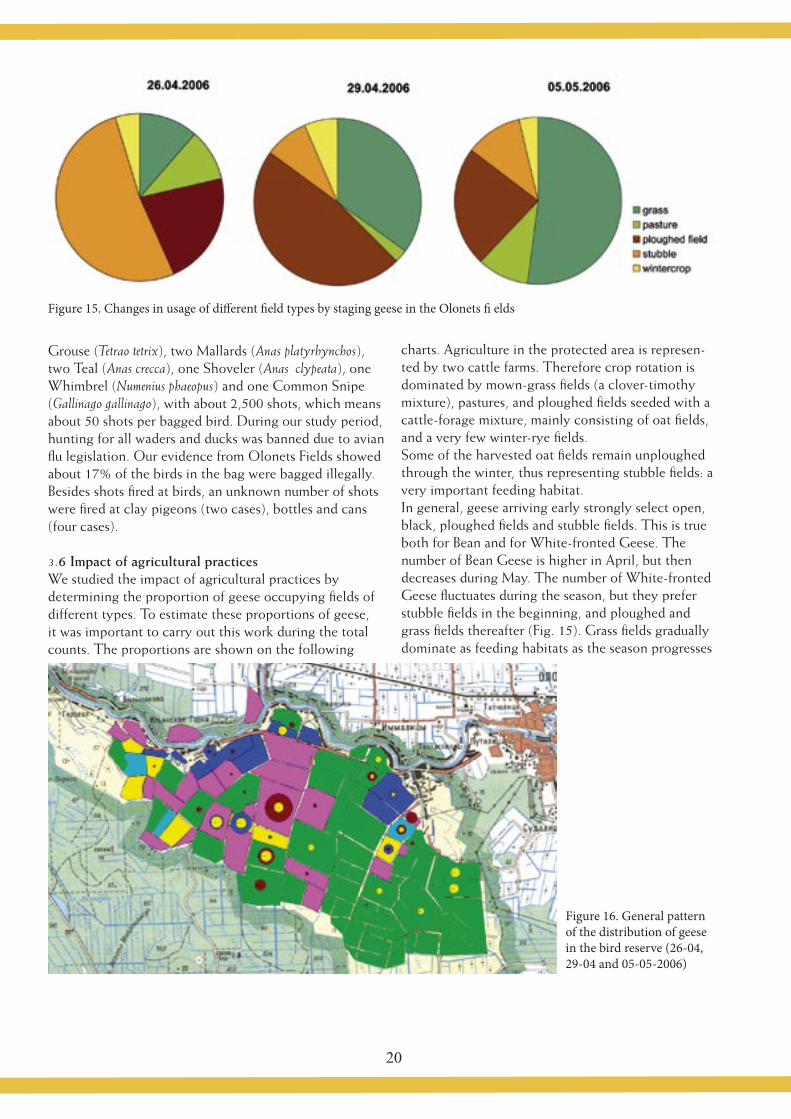
Figure 15. Changes in usage of diff erent fi eld types by staging geese in the Olonets fi elds
Figure 16. General pattern of the distribution of geese in the bird reserve (26-04, 29-04 and 05-05-2006)
Grouse (Tetrao tetrix), two Mallards (Anas platyrhynchos), two Teal (Anas crecca), one Shoveler (Anas clypeata), one Whimbrel (Numenius phaeopus) and one Common Snipe (Gallinago gallinago), with about 2,500 shots, which means about 50 shots per bagged bird. During our study period, hunting for all waders and ducks was banned due to avian fl u legislation. Our evidence from Olonets Fields showed about 17% of the birds in the bag were bagged illegally. Besides shots fi red at birds, an unknown number of shots were fi red at clay pigeons (two cases), bottles and cans (four cases).
3.6 Impact of agricultural practices We studied the impact of agricultural practices by determining the proportion of geese occupying fi elds of different types. To estimate these proportions of geese, it was important to carry out this work during the total counts. The proportions are shown on the following
charts. Agriculture in the protected area is represen-ted by two cattle farms. Therefore crop rotation is dominated by mown-grass fi elds (a clover-timothy mixture), pastures, and ploughed fi elds seeded with a cattle-forage mixture, mainly consisting of oat fi elds, and a very few winter-rye fi elds. Some of the harvested oat fi elds remain unploughed through the winter, thus representing stubble fi elds: a very important feeding habitat.In general, geese arriving early strongly select open, black, ploughed fi elds and stubble fi elds. This is true both for Bean and for White-fronted Geese. The number of Bean Geese is higher in April, but then decreases during May. The number of White-fronted Geese fl uctuates during the season, but they prefer stubble fi elds in the beginning, and ploughed and grass fi elds thereafter (Fig. 15). Grass fi elds gradually dominate as feeding habitats as the season progresses

21
and these fields and pastures turn green.The map (Fig. 16) shows the general patterns of spatial distribution of geese in the hunting-free zone on 26 April, 29 April and 5 May (colours the same as in Fig. 15). The group of fields in the central part of the area was used more often then the other fields. The same is true for some pastures near the high-way. Grass fields in the eastern part of the area were used only after 4 May. It is worth noticing that all the above-mentioned important areas had also been important in previous years, so they could be called the traditional important goose-feeding fields of the hunting-free area. Fields in the central part, where the majority of the geese were observed, represent a mosaic of intermingled fields of different types (ploughed fields, stubble fields, and grass fields).
3.6.2 Impact of spring burning Because the current agriculture in the Olonets Fields is rather extensive, the leftover grass from the second growth after the first cut or after grazing remains on the fields over winter. This delays the growth of fresh grass in the spring and reduces the quality of fodder considerably; the farmers disli-ke the old grass. The cheapest way to solve the problem is to burn the previous year’s grass in the spring. The main spring-burning period is between the end of April and the first weeks of May, which means during the staging period of the geese and the egg-laying period of ground-nesting grassland birds, such as the Short-eared Owl (Asio flammeus), Curlews (Numenius arquata), the Lapwing (Vanellus vanellus) and other waders. During burning and for a period of up to 10 days after burning, the burned grasslands cannot be used by the geese as a feeding site. Therefore, because of spring burning within the limits of the hunting-free goose sanctuary during the hunting season, some of the geese are forced to leave the protected area and to search for food in the area where goose hunting is allowed. All-in-all, spring burning in the goose sanctuary during the hunting season is completely in contradiction with the aims of the sanctuary. On the other hand, when the green grass starts growing 10 days after the burning, it is shorter and grows more slowly in comparison to grass on the unburned fields. However, the unburned fields re-main yellow because of the predominance of over-lying dead grass. Therefore, although green grass is
taller on such fields, it is less accessible and scarcer than on the burned fields. This all makes the burned fields no less attractive then the unburned. In fact, for the later arriving geese, good feeding habitats are represented only by either burned fields (regardless of whether or not they have been grazed) or by unburned fields that have been grazed by geese or cattle. Therefore spring burning causes only a temporary delay in their usage by the geese. Spring burning is a usual practice in the Olonets fields, but besides disturbance of the staging geese it has further negative aspects.
3.6.2.1 Impacts on other wildlifeThe large-scale burning of fields in the main setting and nesting period harms a huge number of ground-nesting birds, who often lose their first clutch and also shelter for nesting, as in the case of the Short-eared Owl, which is still present in this area in good numbers. Furthermore the burning has an adverse impact on certain wintering insects and mammals, such as hares and rodents.
3.6.2.2 Impact on local populationDuring spring burning in the vicinity of Olonets and neighbouring villages, about three houses per week catch fire. In 2006 five houses caught fire. In most cases this means a total loss and, besides the personal tragedy for the house owner concerned, a considerable economic and social problem for the region. In addition, the local population suffers from the smoke as well as the soil and air pollution that may result from the grass burning.
3.7 Conclusions 3.7.1 Of the preliminary resultsThe Olonets fields are an important stop-over site for Arctic-breeding geese on their way north. A high number of Whitefronts uses the Olonets fields during spring mig-ration. The average duration of use in 2006 was 4.8 days, so the mean of all geese using Olonets Fields was less than one week. The probability of finding a marked bird back in the in the field the next day is rather low (35%), so lots of birds staging in the area might have migrated through without having been seen. This result underlines the need of a large number of observation turns or obser-vers in the field to look for marked birds. More detailed analyses have to be done with these data to learn more about the concrete role the Olonets fields play in the migration system of White-fronted Geese, so the results of neckband observations should be combined with daily goose counts ZIMIN et al. (2007) The Abdominal Profile

22
Index (API) estimated for each bird must be analysed with regard to time and to social status of the bird; more data are needed from the Olonets fields. Research at this important spring staging site must be continued in order, too, to verify our results and explanations. We do not know a lot about the function of staging sites. Migration routes of satellite-tracked Whitefronts have indicated several stop-over sites at which the individuals stayed for a long time, presumably to replenish their fat reserves. So, migration could be understood as a series of quite long and fast movements interspersed with long pauses. Unfortunately only one of five PTTs reached its summer area, while at least two birds were shot on migration north. However, PTT Adri showed a specta-cular and unexpected migration route through Poland, Belarus and Ukraine, thence north to Taimyr in July. On his return south in the autumn, Adri followed the same route as some transmittered Lesser White-fronted Geese to Kazakhstan (see www.piskula.org). South of the Ural Mountains, PTT Adri left this route and flew once more to Ukraine. This situation could be viewed as an additio-nal hint of an old traditional network of migration routes connecting the large wintering regions of the Western Palaearctic, as MOOIJ (1996) has proposed.
3.7.2 RecommendationsOur preliminary results from the neckbanding and tracking of Whitefronts show an intensely used flyway along the Baltic states then across Karelia to northern Russia. Nearly all sites our tracked geese used were
agricultural fields. So open landscape and agricultural management is essential for migratory geese at the moment. This implies changes in agriculture have to be monitored carefully.Olonets Fields is an important stop-over site for wild geese during spring migration. The sanctuary itself is well protected, especially during the hunting period. On the other hand, we think the size of the sanctu-ary might be too small to provide enough space for the number of migratory geese using the site. As a result, there is high pressure on the sanctuary, which might increase foraging damage to its fields. We recommend verification of this. Furthermore, areas between the night roost and the feeding sites should be protected. The effect of spring burning lies in the huge per-centage of grasslands being burned within a small number of days. Additionally, a lot of ground-bree-ding meadow birds are affected by spring burning in early May, when some species are in the process of building nests and laying eggs.It is recommended to look into alternative agricultu-ral management schemes to spring burning. A second or late cut or grazing in late summer on the fields might achieve the same effect of clearing away old growth that prevents the early growth of fresh green grass the following spring. Spring burning not only has adverse effects on goose-staging areas, breeding grassland birds and mammals, but also jeopardizes the livelihood of local people and contributes consi-
Figure 17. Spring burning in Olonets Fields Bird Sanctuary

23

24
derably to air pollution and the release of carbon dioxide into the atmosphere. Local authorities, with the support of the Baltic Fund for Nature, are encouraged to look into alternatives to spring burning, for the benefit of geese, people and wildlife in general. Agricultural land acts as a stopover site for many geese in many different areas outside Russia and Poland, where practices such as spring burning are no longer used. Such areas remain agricultu-rally productive, while still providing extensive habitat for the roosting and feeding of goose flocks.
3.7.3 For future research Further research on the function and importance of stepping stones along the routes to breeding grounds is necessary. By using the data from satellite-tracking and neckband studies, the key staging sites could be determi-ned, and the gap in knowledge of their importance and roles in migration could be filled. Key areas should be investigated intensely. By using standardized counts in combination with sightings of neckbands, turnover rates and number of individuals using the sites during their migration has to be investigated in order to assess the importance of each site. Threats to these sites must be examined (e.g., the decrease of area of land use, change in intensity and of land-use management, increase of hunting and other human disturbance). Ways to reduce threats to the key areas should be discussed and imple-mented as soon as possible. In the case of Olonets Fields, a full season of investigati-on should be carried out: neckband reading, and measu-ring flushing distances, hunting pressure and percentage of immatures in staging flocks. The results of the long-term studies of ZIMIN et al. (2007) should be incorporated and the studies should be connected with international scientific work.
Preliminary results from satellite transmitters have de-monstrated that the Olonets fields are one important site but not the only one along the flyway. A thorough ana-lysis of the satellite-transmitter data no doubt will reveal other important stop over sites requiring further research.

25
4. Research on a nesting ground (Kolguev Island)
Kolguev Island is an island of 4,850 sq.km in the southeastern part of the Barents Sea and separated from the mainland by the mere 70-80 km of the Pomor Strait..The island is approximately round in shape and stretches about 83 km east to west and about 92 km from north to south. Although most of the island has a hilly relief ranging from 20 to 60 m in height, elevations of 140 - 173 m are attained in the central part of the island. From the inland, rain and meltwater are transported to the sea by many rivers of varying sizes, which radiate from the centre to the sea. With a length of about 150 km, the biggest river is the Peschanka, characterized by sandy shores and sandbanks. The island shows a mosaic of swampy tundra in the river valleys and dry and swampy tundra in the higher parts. Permafrost is found at a depth of about one meter and more in summer. In the southern and eastern parts of the is-land there are extended lowland plains, at elevations of 1.5 – 3 m above sea level in the east and 5 and 12 m. in the south (southern lapta). These rill-cut (laidy) and gullied (lapti) plains are flooded by the sea during storms. Along these low terraces lie sandy spits and bars (koshki) that form large lagoons. The
largest spit, with a length of about 22 km and a width of about 7 km, is found at the mouth of the Peschanka River on the eastern coast of Kolguev.
On the southeastern coast of the island, at the mouth of the Bugryanka River, lies the village of Bugrino, which is inhabited by about 450 individuals, most of them are Nenets people, who mainly earn their living from the fishery and reindeer herding. At present the herd of rein-deer numbers about 7,800 animals. Bugrino has a wooden landing pad for helicopters and up to one flight per week. In the eastern part of the island, two Russian oil and gas companies have each established a settlement of about 120 people on the banks of Peschan Lake, where they exploit the fossil fuels. The people of these two villages go fishing along the upper and lower Peschanka River. On the eastern spit not far from the mouth of the Peschanka River, an airstrip (Peschanka Airport) has been built where planes up to the size of an AN 26 - a light, twin-engine cargo turboprop, land at irregular times. At some times, such as the exchange crews of the oil companies, airpla-ne frequency is one per day, landing and taking off. In between these flurries of flights, there are periods of up to 50 days without any airplane at all. Sandy roads for buses,
Figure 19. Flight route to Kol-guev by German participants in the goose project in the summer of 2006

26
trucks and all-terrain vehicles link the company settle-ments, several exploration sites, and the airport.
In the north of the island are a meteorological station and a nautical station, both with crews of two, as well as an unmanned lighthouse. A few times per year there are transports by special tundra vehicles between these stati-ons and the company settlements. In the neighbourhood of the meteorological station is a deserted former military camp for border troops as well as a hunting camp for rich oil-company officials. In the hunting season, selected executives of the oil companies are flown into the hun-ting camp by helicopter, spread out over the island for
Figure 20. ECORA zones of the Kolguev Island study area
hunting, re-gather in the camp to celebrate hunting success, and leave again by helicopter at the end of the hunting season. Except for the hunting, most more-or-less intensive human activities are concen-trated along the east coast of the island, whereas cen-tral parts of the inland are used for reindeer herding.
Kolguev is one of the model areas of the ECORA project (Integrated Ecosystem Approach to Conserve Biodiversity and Minimize Habitat Fragmentation in the Russian Arctic). Fig. 20 shows the zonation of Kolguev used for this project. While oil- and gas exploitation is still allowed on the eastern part of

27
Kolguev Island, nature conservation and the tradi-tional lifestyle of the Nenets people has priority in the ethno-ecological zone on the western side of the island. One of the ECORA tasks is to improve the maintenance and integrity and functionality of this zonation and to collect basic data in order to optimize the concept of ethno-ecological areas. So, during our three-year study, we shall investigate the whole island and examine the distribution and den-sity of breeding birds (geese as well as waders, gulls and ducks). We shall present our results in 2009 giving evaluations of the importance for migratory bird species of specific parts of the island, as well as for Kolguev as a whole.
4.1 Breeding biology
Geese arrive at Kolguev during the first third of May (MOROZOV & SYROECHKOVSKY 2004), but mass arrival takes place during last third of May (25-27 May). When we arrived on Kolguev on 29 May, the geese had just arrived en masse (with the peak on 27 May), but were not all the-re. Newcomers were still arriving as late as 4 June, when we observed arriving flock of 80 birds. On the wetlands around our camp, situated on the lower reaches of the Peschanka River, we observed three pairs on 31 May, se-ven pairs on 2 June and 13 pairs on 4 June. All these pairs were continually observed within that area, and no doubt represented the local nesting population.
The number of geese on upland tundra ran-ges from 16 to 65 birds per sq.km, perhaps including mainly non-breeding birds, as the majority of birds encountered during the incu-bation period are families with young from the previous year. The geese feed extensively on Aquatic Sedge, Carex aquatilis, which is growing not only along the shores of lakes and ponds, but also on upland, moss-grassy tundra.
The first nests of White-fronted Geese were found on 3 June, after the first nests of Bean Geese and Barnacle Geese had already been found, on 30 May and 2 June, respectively The geese started staying at their nests after laying the first egg, and incubation constancy was extremely high. We twice carried out 24-hour observations of seven incubating pairs, in order to determine the nesting constancy of the males and of the females. Male constancy and behaviour differed drastically between the pairs, and these differences were not related to the incubation stage, but were more probably explained by the individualsocial status of the birds. In contrast, all observed females had only one recess per day, usually for 4 – 8 mi-nutes, during which time they eagerly fed on sedges, drank, bathed and made short flights.
The nest-initiation period is long on Kolguev and, consequently, the hatching period is also prolonged, lasting almost a month. We first observed hatching on 28 June, although through calculation of hatching dates based on incubation stage and gosling size, we estimated
Figure 21. White-fronted Goose hatching dates, Kolguev 2006
Figure 22. Bean Goose hatching dates, Kolguev 2006

28
the earliest hatching to have taken place on 23 June. The majority of White-fronted Goose nests hatched during 1-9 July, with an observed peak on 6 July (Fig. 21). Bean Goose hatching dates were several days earlier (Fig. 22).
Calculating back from hatching date, we can estimate clutch-initiation date, based on an incubation period of 26 days, and an additional day for every egg in the clutch, plus one extra day for clutches with more than four eggs (ELY & RAVELING 1984). Then we can look at the relationship of clutch size and initiation date. This relationship is shown on the following graphs, on which Day #1 corresponds to 1 June, Day #-1 to 30 May, Day #31 to1 July, and so on (Fig 23).
It should be noted that in the case of the Bean Goose, the period of nest initiation is shorter, the negative correlati-on stronger, and the regression line steeper than for the White-fronted Goose. This situation might be explained by fact that the “strategy” of the Bean Goose more stron-gly corresponds to that of a “Capital-Breeder”, whereas that of the White-fronted Goose is more a mix of “Capi-tal Breeder” and “Income Breeder”. We collected remains of eggs and food samples during the expedition to analyse them for heavy isotopes. This will give us more informations about egg proteins build up from carried or just taken food, analyses are still going on. All this derives from the fact that Bean Geese arrive earlier, when no food resources are available yet on the tundra. Therefore, clutch size is decreased drastically by any delay in nest initiation. On the other hand, White-fronted Geese arrive later, but have longer pre-laying and nest-initiation periods. These geese have the pos-sibility of better feeding on the nesting grounds during
the pre-laying and egg-laying periods than do the Bean Geese. Therefore, in White-fronted Geese the decrease of clutch size concomitant with delay in egg-laying is not as drastic and clear-cut as in Bean Geese (Fig. 23).By the end of June the majority of non-breeders and failed breeders began a pre-moult migration. We star-ted observing flocks of White-fronted Geese flying from Kolguev Island in a northeasterly direction after 20 June. Very intensive migration was observed on 24 June, 3 July, 4 July, and 8 July. During these days, no fewer than 10 flocks of 8–44 birds were observed from the camp area alone as transit migrants.
4.1.1 Habitat distribution during nesting and brood rearing periodWhite-fronted Geese, Bean Geese and Barnacle Gee-se on Kolguev are all found all over the island, but species-specific differences exist in their distribution patterns.In general, landscape types on Kolguev are presented by:a) wet former sea terraces of different levels with numerous thermokarst lakes and bogs, b) glacial landscapes in the central part of the island with elevated hills, occupied by numerous small lakes of glacial origin and drained by steep-sloped ravines,c) gentle wet and boggy fluvioglacial slopes between the second and first landscape typesd) drained, elevated, dry sandy hills in the eastern part of the island west of Peshanoye Lake, ande) modern sea terrace with coastal sedge marshes, spits and mudflats partly flooded by autumn storms and high tides. In all these landscape types different habitats are
Figure 23: Regression of initiation date and clutch size of White-fronted and Bean Geese (Kolguev 2006)

29
intermingled, varying from dry-lichen flat tundra to hummocky bogs with willow bushes.
White-fronted Geese are the most widespread and opportunistic in their habitat selection. They are especially numerous in habitats with sedge wet-lands, low bushes and hummocks. Other important breeding-habitat types are the slopes and bottoms of steep ravines. In general there is a tendency to build nests along edge habitats, such as the upper or lower slopes of ravines, the bushy edges of creeks and on hummocks with bushy vegetation along the lake shores or water courses on upland tundra . All these habitats combine both good shelter and good sight-lines from the nesting site. Very common nesting habitats are ravine sources in upland habitats, where water courses are overgrown with scrub willow. These kinds of habitats are widespread throughout the island, thus making all territory very suitable for
nesting. Coastal sedge marshes and drained uplands are also used for nesting, but with markedly less density.
Bean Goose nesting distribution differs from that of the White-fronts; Bean Geese select habitats that are more well-drained and more strongly associated with glacial lakes and the upper slopes of ravines, where the habitat is more open, with less cover but better views. These kind of habitats are more common in the central part of the island, where relief is more mixed and mosaic, so Bean Geese are only common not less than 10 km from the wet and flat coastal zone. Barnacle Geese, on the contrary, mostly breed on the flat, wet plains of the Peshanka River valley, occupying the lower 25x5 km of its delta. Besides the Peshanka River valley and coastal sedge marshes in the valleys and mouths of other small rivers, Barnacle Geese were also found nesting in the middle sources of several rivers, where they were strongly associated with Peregrine Falcon nesting sites. Near one of the Peregrine
Figure 25: Time budget of males 13 and 14 JuneFigure 24: Time budget of females 13 and 14 June
Figure 26: Time budget of females 28 and 29 June Figure 27: Time budget of males 28 and 29 June

30
Falcon nests, we found a very dense colony of geese: 32 nests of White-fronted geese, 15 nests of Barnacle Geese and 5 nests of Bean Geese, all within an area of 0.09 sq km.The habitat distribution of White-fronted Geese du-ring the brood-rearing period differs not much from the situation described above. During the first week after hatching, White-fronted Geese remain in close proximity to their nests, occupying areas dense with Willow bushes, where they seek both shelter and food supply in the form of Equisetum and young shoots of the sedge Carex aquatilis. So, after hatch the families are usually found within the ravine system, on glacial lakes and within the Peschanka River valley. When the goslings have grown somewhat, the families move slowly downstream from the ravines and select vast sedge marshes in vicinity of lake systems. At the same time, a large number of families remains on upland tundra in the area of extensive willow scrub not connected with water bodies. In contrast to the above situation, Barnacle Geese after hatching change their distribution drastically. This change could be partly exp-lained by the fact that Barnacles, both females and males, feed a lot during incubation, so their food resources are severely depleted. On the other hand, coastal sedge marshes of low Carex subspathacea, which were not at all suitable for nesting, appear to be extremely good brood-rearing habitats. On the other hand, whereas Barnacle Geese were only very seldom found nesting in the central part of island, especially in the area of glacial lakes, after hatching they become extremely common in this type of landscape. They arrive here both in family flocks and also in groups of non-breeders consisting of up to 200 indivi-duals. It all leads to a situation that after hatching Barnac-le Geese start to become widespread all over the island, occupying it more evenly in comparison to during the nesting period. The size of the family groups occupying the central parts of island varies from 2-4 broods on small glacial lakes, to 60 – 100 broods (up to 650 – 700 birds in mixed flock) on the rivers.Adult White-fronts start their wing moult during the last ten days of July, and the first flying families were observed on 10 August. Barnacle geese generally moulted during
the same period, but non-breeders started their wing moult on 4 July. Between 2 and 4 August, it was recorded that most of the Barnacle Geese, including young birds, had started flying
4.1.2 Incubation behaviourIn all pairs studied, we found that in the second half of the incubation period both the females and the males started to devote more time to feeding, but in females the total duration of such feeding bouts was not more than 5 – 15 minutes a day. Females devoted much of their time to sleeping and resting, while males rested and slept only about 50% time of the time, and they only were noted sleeping when their females were sleeping, and never when the females were resting or doing something else (the example of one pair is shown in Fig. 24 & 25). Nest-building activity was observed only during the first days of incubation. In two pairs we recorded that males were absent from the nest for quite a long time. Mostly they were feeding or sleeping 80 – 100 metres from their nests. In five pairs, however, the male’s attendance was ex-tremely high; they did not go farther than 10 metres distance from their mates to feed. On the following charts we can see time budgets for 24-hour observations of females (left) and males (right) at the beginning (Fig. 24 & 25) and at the end of incubation (Fig. 26 & 27). The figures along the x-axis correspond to the nest-code numbers; the letters F and M for the corresponding sex.In the pairs under observation, clutch size varied from 2 to 6, and incubation stage varied from days 1 to 10 on 13-14 June and from days 15 to 24 on 28-29 June (Tab. 3).
Nest Code 25 26 27 36 50 51 71Clutch Size 4 3 6 3 2 5 2incubation stage 13-6-06 10 4 7 3 1 2incubation stage 28-6-06 24 19 22 18 16 18 15
Table 3: Clutch size and incubation stage of observed breeding pairs

31
Plot ID habitat plot size (ha) no of nests nest density (per ha)1 upland tundra 0.2 11 552 upland tundra 0.16 11 693 river valley 0.135 7 524 upland tundra 0.12 10 835 river valley 0.26 2 86 creek valley 0.41 17 417 upland hill 0.28 17 618 coastal terrace 0.24 17 71
Table 5: Nest density (White-fronted Goose) in study plots 2006
Site (year) Average Density Source
Kolguev (2006) 63.4 nests / sq.km Kruckenberg et al. (2007)
Kolguev (2007) 30.0 nests / sq.km unpubl. data
Kolguev (1995) 49.3 nests / sq.km Syroechkovsky & Morozov (2004)
Tobseda (2005) 28 nests / sq.km Litvin et al. in Soloviev & Tomkovich 2006
Mongocheyevaya River, Gydan (2005)
0.9 nests / sq.km Emelchenko in Soloviev & Tomkovich 2006
Omulyavaya River, Gydan 0.86 nests / sq.km Emelchenko in Soloviev & Tomkovich 2006
Cape Sterlegova, Taimyr (1994) 0.7 nests / sq.km Tulp et al. 1998
East Taimyr Lake (1989) 0.2 - 0.8 nests / sq.km Mooij in Hötker et al. 1995
Malaya Lopgata, Taimyr (1990-1992)
0.1-1.0 nests / sq.km Mooij et al. (1995)
Kyttyk Peninsula, Chukotka (2005)
0.12 - 1.0 nests / sq.km Solovieva et al. (2006)
Table 4: Breeding density of Whitefronted Geese
4.2 Breeding distribution
4.2.1 Breeding density, clutch size, hatching success, chick survival, brood sizeWe estimated breeding density on the basis of square plots of 400x400m, 500x400m, 500x500m, and 500x1000 m, in which complete nest surveys were carried out. Such plots were established in up-land tundra (three plots), in the Peschanka River val-ley (two plots), on the hill among the sedge valley (one plot) and on the southwestern coast on former sea terrace (one plot), (see Fig.28). Besides that we used transect nest counts during our route surveys in the central part of the island. Taking into account the transect width of 20 m during such censuses, we made additional calculations of breeding density for some remote areas, where complete nest surveys on plots were not carried out because of time cons-
traints. Based on these study plots and transect counts, we could calculate nesting density, presented in Tab. 5.
Thus, if we only take into consideration upland habitats, we obtain an average breeding density of 63.4 nests per sq.km, which is 1.3 times higher than that recorded for 1995 (MOROZOV & SYROECHKOVSKY 2004). Compared to other breeding areas, density is extraordinarily high (see Table 4). The line transects performed on upland tundra gave the comparable figures of 58 (1995) and 85 (2006) nests per sq. km in the middle of June, and 29 and 39 nests per sq. km, respectively, at the beginning of the nesting period, when only 50% of the breeding population had started incubation (5-6 June).These figures are the highest in the world among known data for this species (MOROZOV & SYROECHKOVSKY 2004).The breeding density of Bean Geese was much lower than

32
Figure 28: Nest distribution of White-fronted Geese (Anser albifrons) in 2006 and study plots in Peschanka camp area.
Figure 29: Structure of Barnacle Goose (Branta leucopsis) colony in 2006: the lowest density shown pale yellow was 1,100, and the highest shown deep brown was 3,000 nests per sq.km.

33
that of Barnacle Geese, and the data were only ob-tained from the transect-line census in upland tund-ra, because only two nests were found in the vicinity of the Peshanka camp area. In the central part of the island the calculated breeding density was 10 nests per sq. km at the beginning of the nesting period (5-6 June) and 33 nests per sq km in the middle of June (16 nests were found along a 16-km transect of 30 metres width). Breeding density of Barnacle Geese varied in different parts of the colony, from 1,100 to 3,000 nests per sq. km (Fig. 28), and the total number of nests in the main colony on the Peschan-ka River was estimated to be no less than 60,000. Clutch size in the White-fronted Geese varied from 1 to 9, being 3,52±0,07 (N=364) on average (Fig. 30). In the nests of Bean Geese clutches varied from 1 to 6, with an average of 3,70±0,23 (n=44) (Fig. 31) Breeding success of both species depended upon predation by Arctic Foxes, Glaucous Gulls and Arctic Skuas. All these predators were responsible for complete clutch loss and also for partial clutch loss (egg loss). Due to complete clutch loss, the overall hatching success of both species was de-termined to be 78% (per cent of nests in which at least one gosling hatched). With the White-fronted Geese we also carried out a more detailed study of complete clutch loss in relation to clutch size and breeding chronology. It appeared that big and early clutches were more successful than small and late clutches (especially unsuccessful were clutches of
Figure 30: White-fronted Goose clutch size (2006)
Figure 31: Bean Goose clutch size (2006)
Figure. 32: Clutch predation in (A) early White-fronts nests, (B) late Whitefronts nest and (C) Bean Goose nests

34
two eggs initiated after peak initiation – Fig. 33). For early clutches the success rate was 80%, while for later clutches (initiated after 3 June) hatching success was only 68%. Another cause of decreased nesting success was partial clutch loss (or egg loss) due to the predation of some eggs by the same three main predators (Arctic Skuas were more important avian predators compared to Glaucous Gulls at this stage of breeding). Contrary to the greater rate of complete clutch loss for smaller clutches, egg loss was markedly higher in bigger clutches in both the Bean Goose and the White-fronted Goose (Fig. 32A - C). The level of clutch loss (partial and total) was markedly higher in Bean Geese, in which only 16% of nests were not predated at all, whereas for White-fronted Geese nearly 30% of all nests remained untouched (Fig. 34-36). For White-fronted Geese, clutches of 4 appeared to be the most productive. For Bean Geese, clutches of 4 were modal, but overall production was maximal in clutches of 6 (Fig. 30). As a result of egg loss, average brood size at hatch was markedly smaller than average clutch size at the beginning of incubation; it was highest in the early nests of White-fronted Geese (2,95±0,09, n=126), and lowest in the late nests (2,69±0,08, n=143), with an average of 2,92±0,08 for both groups combi-ned (n=269). For Bean Geese, brood size at hatch was 2,5±0,22 (n=50). The spatial distributions of hatched (red dots) and predated (blue crosses) nests of White-fronted Geese are shown on the map (Fig. 38). Arctic Fox dens are shown as purple circles. The majority of predated nests are situated in close proximity to the camp area, where anthropogenic nest disturbance was highest. In remote areas, nest predation was low regardless of proximity to fox dens. This also means that in areas with no anthropogenic disturbance, complete clutch loss is a rare event, and overall hatching success is therefore even higher than 78%, and may reach 85-95%. (see Tab. 6).
Figure 33: Egg loss in nest of Bean Goose (top) and White-fronts (below) depending on clutch size
Position disturbance factor No of nests overall success„falcon“ colony 2 visits during nesting 32 0.9375„gull“ colony 2 visits during nesting 16 0.75upland tundra route survey 1 visit 25 0.92upland tundra route survey 1 visits 22 0.863636Study plot #6 1 visits 17 0.941176Study plot #4 1 visits 10 0.90Study plot #1 visited oft en 11 0.454Study plot #2 visited oft en 10 0.70Study plot #3 4 visits 17 0.647Study plot #4 visited oft en 7 0.5714
Table 6: Nesting success in diff erent study plots
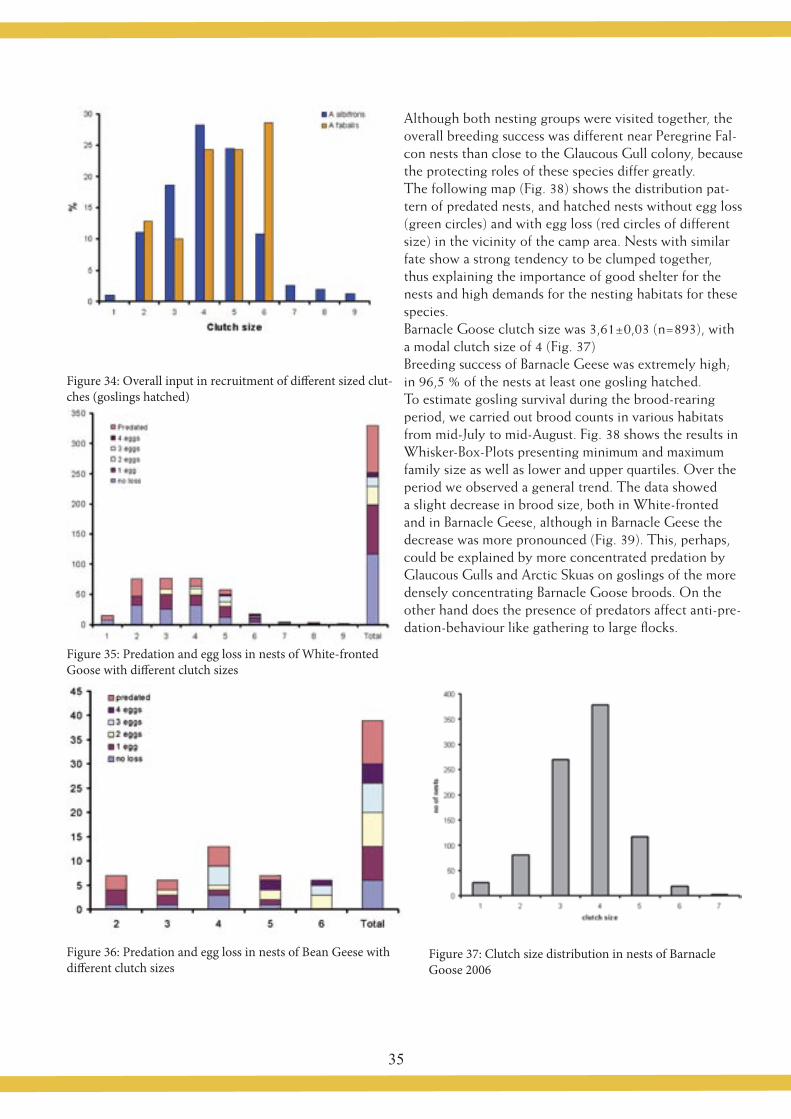
35
Figure 34: Overall input in recruitment of diff erent sized clut-ches (goslings hatched)
Figure 35: Predation and egg loss in nests of White-fronted Goose with diff erent clutch sizes
Figure 36: Predation and egg loss in nests of Bean Geese with diff erent clutch sizes
Although both nesting groups were visited together, the overall breeding success was different near Peregrine Fal-con nests than close to the Glaucous Gull colony, because the protecting roles of these species differ greatly. The following map (Fig. 38) shows the distribution pat-tern of predated nests, and hatched nests without egg loss (green circles) and with egg loss (red circles of different size) in the vicinity of the camp area. Nests with similar fate show a strong tendency to be clumped together, thus explaining the importance of good shelter for the nests and high demands for the nesting habitats for these species.Barnacle Goose clutch size was 3,61±0,03 (n=893), with a modal clutch size of 4 (Fig. 37)Breeding success of Barnacle Geese was extremely high; in 96,5 % of the nests at least one gosling hatched.To estimate gosling survival during the brood-rearing period, we carried out brood counts in various habitats from mid-July to mid-August. Fig. 38 shows the results in Whisker-Box-Plots presenting minimum and maximum family size as well as lower and upper quartiles. Over the period we observed a general trend. The data showed a slight decrease in brood size, both in White-fronted and in Barnacle Geese, although in Barnacle Geese the decrease was more pronounced (Fig. 39). This, perhaps, could be explained by more concentrated predation by Glaucous Gulls and Arctic Skuas on goslings of the more densely concentrating Barnacle Goose broods. On the other hand does the presence of predators affect anti-pre-dation-behaviour like gathering to large fl ocks.
Figure 37: Clutch size distribution in nests of Barnacle Goose 2006

36
Figure 38: Distribution pattern of pre-dated nests with egg loss (red dots) and without egg loss (green dots) / insert of distribution map - nests with similar fate clumps together
fig. 39: Daily family size during the latest brood rearing period with minimax, median and 25%-75% quartiles

37
4.2.2 Synchronous countsOnce a week we organised synchronous counts of the whole saltmarsh area, as well as the upland-tund-ra area right around the camp site. During the bree-ding season, a portion of the non-breeding Barnacle Geese gathered in the salt marshes at the mouth of the Peshanka River. After hatch, an increasing number of Barnacle families moved from the inland tundra and river valleys to the coastal salt marshes. To record these movements between 22 July and 11 August, counts of the numbers of both goose species in the salt marshes and on hill slopes were underta-ken. Despite the growth of the vegetation, which obstructed the counts, the synchronous count data showed a clear decrease in the number of Barnacle Geese, starting at the end of July. Probably some of
Figure 40: Results of syncronous counts in study area July and August 2006
the geese left the Kolguev salt marshes immediately after moulting. The number of White-fronted Geese in the salt marshes was low throughout the period and did not change signifi cantly. The number was much lower than was the number of successful breeding pairs in the study area. So we suspect that most of the White-fronted Goo-se families went inland toward central Kolguev and reared their goslings around the glacial lakes there.
4.2.3 API-Index and family strcture surveyWe censused family size in groups of non-breeders during our route surveys across the island. To get an impression of and categorise the body condition of geese, OWEN (1981) established the use of the shape of the abdomen. The better the goose subsist, the larger the abdomen. OWEN (1981) used this Abdominal Profile Index (API) for the fi rst time in Barnacle Geese. This system was adopted
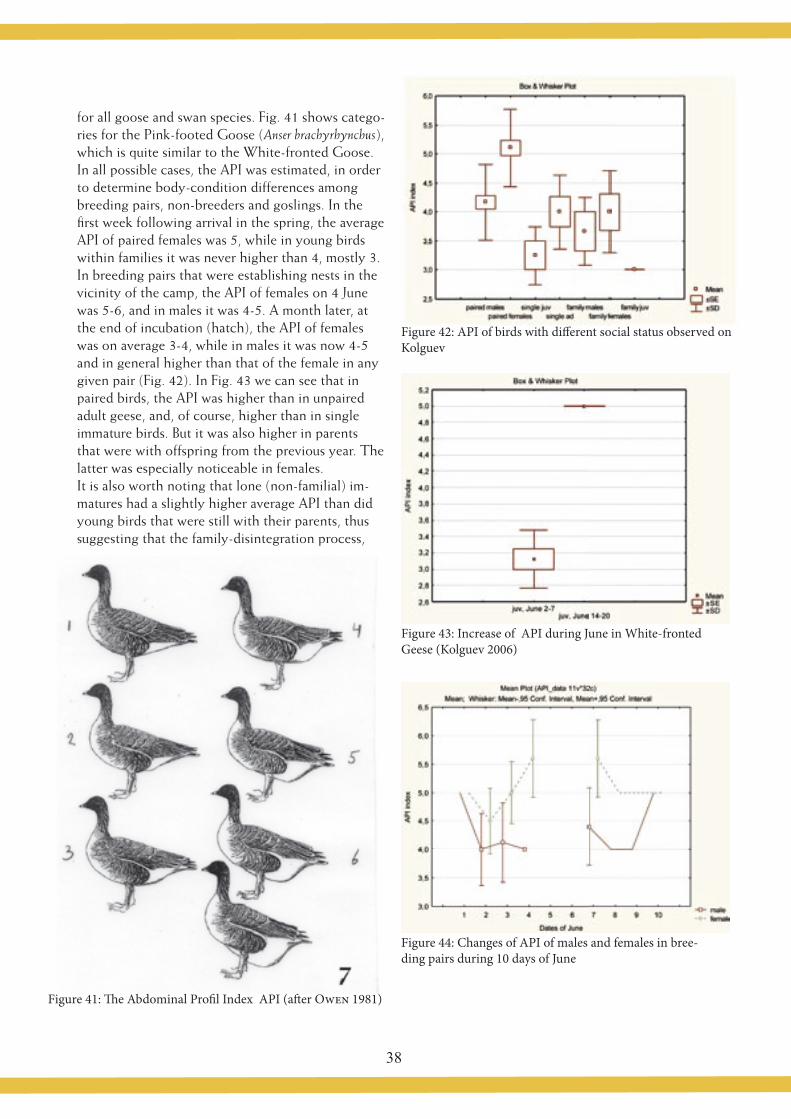
38
for all goose and swan species. Fig. 41 shows catego-ries for the Pink-footed Goose (Anser brachyrhynchus), which is quite similar to the White-fronted Goose. In all possible cases, the API was estimated, in order to determine body-condition differences among breeding pairs, non-breeders and goslings. In the fi rst week following arrival in the spring, the average API of paired females was 5, while in young birds within families it was never higher than 4, mostly 3. In breeding pairs that were establishing nests in the vicinity of the camp, the API of females on 4 June was 5-6, and in males it was 4-5. A month later, at the end of incubation (hatch), the API of females was on average 3-4, while in males it was now 4-5 and in general higher than that of the female in any given pair (Fig. 42). In Fig. 43 we can see that in paired birds, the API was higher than in unpaired adult geese, and, of course, higher than in single immature birds. But it was also higher in parents that were with offspring from the previous year. The latter was especially noticeable in females. It is also worth noting that lone (non-familial) im-matures had a slightly higher average API than did young birds that were still with their parents, thus suggesting that the family-disintegration process,
Figure 42: API of birds with diff erent social status observed on Kolguev
Figure 43: Increase of API during June in White-fronted Geese (Kolguev 2006)
Figure 44: Changes of API of males and females in bree-ding pairs during 10 days of June
Figure 41: Th e Abdominal Profi l Index API (aft er Owen 1981)

39
which occurs in June on the nesting grounds, may begin with high-ranking yearlings, who are ready to leave the family fi rst. During June these young indi-viduals markedly increased their API from 3 to 5. In paired adult females, but not in males, the API mar-kedly increased during the peak egg-laying period (fi rst 5 days of June), thus confi rming the tendency of White fronted Goose females to rely on the food resources of their nesting grounds, as well as confi r-ming a shift to the “income” strategy, as revealed by the very gradual decrease in clutch size with time (Fig. 46). In breeding pairs, the API of both the male and the female decreased during the incubation period, but then during the brood-rearing period increased again, almost to its former level (Fig. 45).
In females, the API was maximal at the beginning of incu-bation, and its decrease throughout incubation was more prominent than in males.
We carried out route surveys of non-breeding White-fronted Geese in upland habitats, in ravines and in the Peschanka River valley. Their density (birds per sq. km) slightly decreased as the season progressed (Fig. 47). Density variations were highest in ravines; in upland habitats density was quite even. In general, the density of non-breeders was between 20 and 40 birds per sq. km, i.e., three times less than the density of breeding pairs. The distribution of White-fronted Geese of different social status varied between habitats, so, for example, families with yearlings were encountered only in upland
Figure 46: Changes in API of focus breedig pair
Figure 47: Bird density in diff erent habitats
Figure 48: Distribution of social status for diff erent habitats in (White-fronted Geese)
Figure 49: Family sizes in three time segments (2006)

40
habitats, while in ravines we met only pairs without yearlings, single adult birds and lone yearlings. In river valleys and around lakes, we mostly encountered geese in pairs; perhaps some of them were breeding birds, espe-cially during the first half of June. Family size was not constant in June, and during the nest initiation stage (up to 10 June) we met some single adult birds, some lone yearlings, and some families with yearlings – mostly with 1 or 2 (P1 and P2 Fig. 47). A smaller number of families included 3 or 4 yearlings. By the middle of June we no longer saw families with 3 or 4 yearlings, while the majority of families had only one young bird. By the end of June, the proportion of lone yearlings had increased drastically, and no more families with yearlings were encountered. This allowed us to con-clude that the disintegration of most families occurred during June.
4.2.4 PredationOn Kolguev, the most important prey-animals of the Eurasian tundra - lemmings - are lacking, but still the typical tundra predators, such as the Arctic Fox (Alopex
lagopus), Glaucous Gull (Larus hyperboreus), Arctic Skua (Stercorarius parasiticus) and Snowy Owl (Nyctea scandi-aca), were present in considerable numbers. Within the study area of approximately 80 sq.km there were at least four fox dens, each with 5-7 pups, and at least 30 broods of the Glaucous Gull, each with 2-3 chicks. These species have been shown to be the main predators of the geese on Kolguev, Shortly after hatching, especially, most families reacted with panic to the appearance of one of these predators and started to run, which left the goslings largely unpro-tected. This behaviour made it easy for the predators to catch small goslings. Furthermore, because of the panic reaction of their parents, a considerable number of goslings lost their parents and became easy prey. Later in the season, an increasing number of parents reacted not with panic, but with defensive beha-viours, and defended their goslings successfully. Ho-wever, we observed a distinct difference in the pre-dator defence behaviour of the two species of geese. Whereas Barnacle Geese often gathered in big flocks, with a few adult birds defending the entire flock of
Figure 50: Upland tundra of northeastern Kolguev (August 2006 )

41
geese, the White-fronted Geese remained largely by themselves or only in loose contact with neigh-bouring families. Their defence strategy relied much more on hiding in the vegetation at the advance of a predator. They behaved rather secretively, hiding in the vegetation and actively sought out the taller sedges and rushes of the salt marsh. Barnacle Geese flocked into increasingly larger groups heading for the open but fertile salt marshes, where together they could fight off attacks from avian predators such as Glaucous Gulls, but also Snowy Owls, by creating defence circles, with young and vulnerable members of the group placed in the middle. We observed several times this strategy used successfully when each of these predators attempted to attack the goose families. However, Glaucous Gulls deve-loped a strategy of attacking the wandering geese at their most vulnerable sites. These were channel crossings and slope descents. The gulls often acted in groups of two to five birds, which could include Heuglin’s Gulls (Larus heugli-ni), and rather more obstructively and at a distance Arctic Skuas joined in. Increasingly Arctic Skuas be-came a nuisance for the Glaucous Gull and while the skuas were attacking viciously the gulls, the goslings were given another chance to catch up with their families. In many cases the gulls were successful in separating members from the group and subsequent-ly attacked the lost goslings without difficulty, even in dense vegetation, since the goslings were not in-conspicuous, noisily calling to their families. Only in a very few, exceptional cases could we observe that adult geese made the effort to return and guide the lost gosling back to the group. The goslings were also seemingly without guidance on how to catch up and often we found that the goslings, as soon as they lost sight of their families, were easily distracted and lost their way, wandering randomly, loudly calling, over the tundra. In fact, on many occasions we witnessed goslings coming straight to us intruders, without any shyness, assuming we might be the parents or some friendly member of the group. This behaviour of actively approaching intruders has also been observed in the case of attacking gulls. The gulls were in no hurry to kill the goslings and often sat near the gosling, which again approached the gull, what looked like confusing the attacker with a member of the family. In one case we observed a gosling actively jumping
at the gull, which could only be interpreted as a counter-attack, as goslings do not beg for food. We observed, too, flocks of up to six goslings, separated from their adults, keeping together while searching for their families. All the above observations almost exclu-sively refer to the Barnacle Goose, because of its more secretive behaviour. During the study period, the food supply for the pre-dators was so large that at least some of the Glaucous Gulls, after killing a gosling, only ate the liver (ZÖCKLER et al. in press). Quite a few freshly killed goslings were found around the fox dens.Counts of the number of goslings in families showed that Barnacle Goose and White-front families had an average of 2.5 and 2.3 goslings, respectively. Failed breeders were not included in these counts, which means that these data cannot be used to calculate reproductive success.4.2.5 Interspecific relations with other breeding species Since the first study of the geese breeding on Kolguev (MOROZOV & SYROECHKOVSKI 2004), the number of breeding Barnacle geese on the island has increased considerably. Therefore it was feared that because of interspecific competition for brood and feeding sites the breeding popula-tion of the White-fronted Goose could have come under pressure and even could have declined. During the current study, nesting densities of 30-50 bree-ding pairs per sq.km were found, which is comparable with the densities of found in the 1990s by MOROZOV & SYROECHKOVSKI (2004). During their study, considerable differences were found between the Barnacle and White-fronted Geese in nesting and feeding habitats .
4.2.6 Nesting habitatFor feeding, Barnacle Goose families mainly used the riverbanks and salt marshes, whereas the White-fronted Goose and the Bean Goose families mainly were feeding in the inland marshes and marshy valleys. During July and August big and small groups of Barnacle Goose families were observed daily walking through the uplands and along the rivers from the inland to the coastal salt mars-hes. The same behaviour was not observed in the other goose species, and right up until the final days of the expedition only minor numbers of the Anser species were observed in the salt marshes.Based on these observations, there does not seem to be direct competition between the species of Anser geese and the Barnacle Goose for nesting or for feeding habitats. Nonetheless, the question remains as to the extent to which the high number of predators is influenced by the

42
Figure 51: Families of Whitefronted Geese - moulting adults on a lake
additional food source the strong increase of breeding Barnacle geese on the island represents. This increased food supply could have triggered an increase of predation pressure on the Anser species as well. This possible nega-tive effect on the reproductive success of these species needs to be studied more intensively in the future.
4.3 Moulting habitat
We observed moulting flocks of non-breeding individuals of the three main goose species starting in early (Barnac-le Geese) and mid-July (Bean Geese and White-fronted Geese). In contrast to White-fronted Geese, Bean Geese did not leave Kolguev on a moult migration in late June. On the contrary, we observed flocks of Bean Geese flying over the island in a variety of directions. Moulting groups of Bean Geese were recorded in different parts of the island interior. They were most often encountered in the glacial landscape, occupying small lakes, but also were re-corded on the lakes in the lower reaches of the Dorozh-kina River and on some other rivers in the northern part of the island. The moulting-group size varied from four to 750 individuals. At the same time, the biggest moul-ting flock of non-breeding White-fronted Geese was only 50 birds. If moulting flocks of non-breeding White-fronts were extremely rare on Kolguev, perhaps they consisted only of late, failed breeders.
Barnacle Geese were the most abundant moulting species, and flocks of up to 250 individuals were observed on the Peschanka River, with distances of 2 – 3 km between flocks. Such moulting flocks were also recorded on another big river – the Elgov-Tarka, and also on glacial lakes in the interior of the island. The biggest flocks outside the coastal-marsh zone were observed on 21 July on a system of glacial lakes on the upper Dorozhkina River. These flocks were comprised of 1,140, 3,150 and 700 individuals.
No less common were moulting flocks of Barnacle Geese north of the Velikaya River. But, of course, the majority of non-breeders were associated with the coastal marshes of the Peschanka River delta, where their numbers were difficult to estimate correctly because they were commonly intermingled with families.
4.4 Bird ringing / neckbanding
During this expedition we banded 19 White-fronts, 15 Barnacles and three Bean Geese with colour markers. Although we used leg-rings for the Barnacle Geese, we used neckbands for the Bean and White-fronted Geese. Three times we prepared the large goose trap, formed by a long net, to catch the geese.

43
Figure 52: Distribution of observed colour-marked Whitefronts banded on Kolguev 2006
grass swards there, which make reading of leg-rings quite difficult.
4.5 Human impact 4.5.1 Oil industryOn the eastern part of Kolguev, two Russian companies are prospecting and drilling for oil and gas. These com-panies each employ about 120 people, who live in two settlements in the centre of the exploitation area. Every two months there is a complete turnover of crews. On the eastern spit not far from the mouth of the Peschanka River, an airstrip has been built (Peschanka Airport), whe-re planes (up to the size of an AN 24) land at irregular times. There are some periods (crew-exchange of the oil companies) when airplane frequency is one plane per day landing and taking off. In between there are long periods without any airplane at all. Sandy roads have been built for lorry-buses, trucks and all-terrain vehicles (vesdekhods) between these company settlements and several exploration sites as well as the airport. To build these roads, a number of hills were dug
Additionally, we caught geese on open tundra, when they appeared in our front. Fig. 52 shows the dis-tribution of banded White-fronts in the following winter season (2006/07). More than two-thirds of the birds were reported. Most of the reported birds were seen in The Netherlands and Belgium; one of them was also reported from the United Kingdom (Kent, England). Birds were also reported from the Oder lowlands (Brandenburg, Germany) and central Poland. Because one bird was reported only once on spring migration in Poland, we suggest that some of the missing birds might have flown to Hungary or Southeastern Europe, where observation density is quite low and marked birds often not reported. Fur-thermore, numerous observers send their sightings to their national ringing centres and it takes some time for all the data of a winter season to be tallied. One of three Beans was seen on Rügen Island in autumn, and only two of 15 Barnacles were seen at all during the following winter season. This may be a result of mild winter temperature and of the high

44
up and several square kilometres of tundra disturbed, lakes drained and rivers dammed up. Furthermore, there are a considerable number of former drilling sites. At these sites and in their vicinity, the tundra vegetation is disturbed The people of the oil villages do not hunt on the is-land, but do fish along the upper and lower parts of the Peschanka River.
4.5.2 Hunting (indigenous and hunting tourists)Indigenous people of the village of Bugrino traditionally live from reindeer-herding, fishing and hunting. Besides the usual disturbance around settlements, their influence through hunting will be highest in the vicinity of Bugri-no. Because of the relatively low number of people living in the village, the influence of indigenous hunting might not be too high. On average, the number of hunters in Bugrino village fluctuates between 120 and 150 people, and an average hunter uses about 400 – 600 cartridges per year, mostly hunting only geese in the spring and Willow (Ptarmigan) Grouse (Lagopus mutus) in the autumn. Their average harvest might roughly be estimated at 20-100 geese per person per season, with a total bag of 2,000 – 3,000 geese for the village per year. This total includes the summer hunt of moulting geese on some lakes (not as widespread a practice as spring hunting). Another way villagers use this natural resource is through egg collection, which mainly takes place in the colony of Barnacle Geese situated on the nearest sand spit. The total bag for the village could be estimated as up to 3,000 eggs, with an average of 50 – 100 eggs collected per family.The influence of hunting by the people of the settlements of oil-and-gas workers may be low; officially they do not go hunting at all and if it occurs, the level of illegal hunting seems to be low.
A totally different problem is that of disturbances caused by sport-hunting tourists. In the north of the island there is a hunting camp for rich oil officials. During the hun-ting season, selected oil executives are flown into the hunting camp by helicopter. These hunters are equipped with the most modern equipment and are very mobile. By means of helicopter they can reach every part of the island and are transported by helicopter to the best hun-ting sites with high numbers of geese. These sites are ex-plored by special scouts before the arrival of the hunters. After hunting, the tourist hunters are collected again and brought back to the comfortable camp to celebrate and
relax. At the end of the hunting season these hunters leave the island again by helicopter. This kind of hunting not only brings disturbance to considerable parts of the island, but also is very effective from a hunting point of view, which means that it is easy for these hunters to shoot a large number of geese almost everywhere on Kolguev. Most of the birds bagged are buried. There are no data to estimate the hunting bag of such tourists, and the level of such hunting tourism has decreased. In recent years the number of such tourists has decreased in the vicinity of Bugrino village. Formerly, their total bag was estimated at 2,000 – 3,000 geese per spring – i.e., nearly the same as the subsistence hunt by the inhabitants of Bugrino village.4.5.3 Scientific researchScientific research itself also should be taken into account in calculating overall human impact. This is especially important when estimating breeding success on the study plots for White-fronted Goose nests. Four plots, situated in close proximity to the camp, were visited quite often, as they had to be crossed on the way to remote areas. As a result, they appeared to have the lowest breeding success (see the table in 3.2.1) - only 45 – 64%, while on the plots visited only once the breeding success was 86-95%. This result demonstrates that even a relatively low level of human activity can have a great impact on Arctic-breeding birds.
4.6 Scientific Outlook
There is a high priority to continue the marking and tracking of White-fronted Geese. Unfortunately only one of the five transmittered birds reached the nesting grounds, while two were shot north of the Arctic Circle and two birds failed for unknown re-asons. To get an idea of how the migration system of White-fronts works - general directions, flyways and stepping stones, a greater number of tracked birds is necessary. The spring migration of 2006 showed that the system of solar-GPS transmitters works and that we can set up a very popular web tool for the general public to follow the geese „live“ on the Internet. We shall need 15-30 more transmitters to find out the real “hot spots” of stepping stones in the migration system. These key sites ought to be studied in the future. So, regular geese counts should be done there as well as investigations to find out important night

45
Figure 53: Dunlin (Calidris alpina) declined on Kolguev as well as in the waddensea
Palaearctic. That`s why Kolguev is predestined as a model region and key area for future studies.
All ongoing changes should be identified on this island. During the following years, one of the most important tasks is to understand more concerning the pro-cesses of the breeding biology that are responsible for this decline (e.g., density dependence, gosling survival, predation, hu-man impact, influence of weather and climate). Only once we understand the cause of this development can nature-conservation organisations and administrations respond appropriately and successfully. Kolguev is also an important breeding area for wader species that migrate via the Waddensea to their wintering areas. Some of these species have shown a sharp decline over the last 15 years. Special studies should be done especially on the breeding grounds to explain the decline in wader, geese and swan species. Kolguev could be a pro-per place for such examination because of the absence of rodents; complicated cross-links with the lemming cycle are excluded on this island.
roosts and potentially key feeding sites. In these sites in particular there should be an intensive look at threats and at conservation options.
In the Olonets Fields it might be useful to collect more data on flushing distance relative to hunting intensi-ty. In particular, more data from the non-hunting period would be needed. These data - in combination with those from the German and Dutch wintering grounds - will give us results of high value to the optimization of the conservational require-ments at all goose staging sites, especially within the European Union, where managers need valid tools to guarantee success in bird protection. Since the beginning of the 1990s, the breeding success of White-fronts and Beans has declined continuously, while the breeding success of the Barnacles has been constantly high. During this period, hunting pressure increased remarkably. As far as we know, low breeding success must be a result of processes based on the nesting grounds. Kolguev is the hotspot of Arctic-nesting geese in the Western

46
especially by the Arctic Fox, has been primarily focused on the Bean Goose because of its preference for rather open habitats, which are more vulnerable to fox predation.
Egg loss during incubation (due to predation by Arctic Skuas, Glaucous Gulls and Arctic Foxes) was an important factor decreasing overall productivity and affecting the majority of nests, so that clutch size at hatching was 3.1 goslings at average. Nesting success was greater in earlier and bigger clutches; egg loss in later nests was higher. These patterns of reproductive success correspond to those for White-fronted Geese in other parts of its range.
Almost all non-breeders and early failed breeders left eastern Kolguev by the end of June, migrating to the northeast in flocks, so that during the brood-rearing period only families with goslings and some small groups of non-or-failed breeders remained on the eastern part of the island. Brood-size decrease due to heavy predation, observed in Lower-Peschanka habitats during brood-rearing led to a brood size at fledging time (mid-August) of just slightly more than two goslings per family (complete brood loss exclu-ded). We may speculate that this heavy predation is partly explained by the proximity of an extraordina-rily large colony and brood / moulting geese concen-trations of Barnacle Geese in the Peschanka delta, which, as was ob-served, was an important attractor for numerous avian and mammalian predators.
There is great diversity in habitat, both nesting and brood-rearing, and geese occupy almost all types of boggy valleys, bushy uplands and glacial lakes and creeks with rather a homogenous nesting density.
The suitable nesting area for White-fronted Geese on Kolguev is about 4,500 sq.km and, with an average breeding density 40 nests per sq km (exactly the same as it was 1995; see MOROZOV & SYROECHKOVSKI 2004), we calculated entire breeding population to be 180,000 breeding pairs, which, together with a 20% portion of non-breeders, gives us approximately 450,000 White fronted Geese arriving on Kolguev in spring. This comprises almost one third of the total Western Palaearctic population of this species (MAD-
5. Discussion
The nesting density and total number of White-fronted Geese on Kolguev have no comparable data anywhe-re within the breeding range of this species (MADSEN et al. 1999, ELY & RAVELING 1984). The nesting density was found to be about 40 - 50 nests per sq.km in 1995 (MOROZOV & SYROECHKOVSKY 2004), which corresponds well to our data, so we may conclude that it has remained rather stable during past 10 years.The phenology of nest initiation, and the pattern of decreasing clutch size as the season progresses show marked differences in the Bean and the White-fron-ted geese. The former species is earlier, and this corresponds well to the breeding phenology of this species in other Arctic regions (LITVIN & SYROECHKOVSKY 1996). The stee-per decline of clutch size with time found in Bean Geese may illustrates the more “capital” breeding strategy of this species (DRENT & DAAN 1980), while White-fronted Geese seem to rely much more heavily on the rich food resources on the nesting grounds (flooded wetlands in river valleys), thus representing more of an “income” stra-tegy on the “capital-income” axis. This also corresponds to a marked increase in the API of females observed in breeding pairs during the pre-laying stage on Kolguev, while the API of males remained more or less constant. A similar pattern of increase in body mass for female geese and lack of increase for males between arrival and incubation onset has been described in Alaska (BUDEAU et al. 1991). In the following years we`ll intensify studies of “capital”- vs “income-breeder” strategy in arctic breeding geese in cooperation with Moscow Bird Ringing Center and Moscow University.
Because of the low predation pressure and good wea-ther conditions, nesting success is very high on Kolguev Island (78% on average overall, 90% on non-disturbed plots), which is rather high for this species, in which nes-ting success in different areas varies from 35,9 to 90,9% (ELY & DZUBIN 1994).
We found that Bean Geese tend to nest in more open habitats, and suffer more markedly from complete clutch loss. This is most remarkable if we point out that all Bean Goose nests were found outside the camp area, i.e. in areas with low human-induced disturbance, where the nesting success of White-fronted Geese was markedly higher. We can only explain this by the fact that the increase over the past 10 years in predation pressure,

47
SEN et al 1999). We may conclude that the vast wet plains of Kolguev serve as the core breeding area for the entire population.For White-fronted Geese and for Bean Geese, inves-tigations are urgently needed into nesting density and breeding success in the interior of the island, far from the main Barnacle colony and from its associa-ted predators.
Egg loss during incubation (due to predation of Arctic Skuas, Glaucous Gulls and Arctic Foxes) was important factor decreasing overall productivity, influencing the majority of nests, so clutch size at hatching was 3.1 goslings at average. Nesting success was greater in earlier and bigger clutches, as well as egg loss in later nests was also higher. These parameters correspond to average pattern of bree-
Figure 54: chicks of White-fronted goose during measurements
ding success of White-fronted Geese in other parts of its species range.Almost all non-breeders and early failed breeders left eastern part of Kolguev by the end of June, migrating in flocks north-eastward, so during brood-rearing period only families with goslings and some small groups of non-or-failed breeders remained on the eastern part of the island. Brood-size decrease due to heavy predation, observed at lower Peschanka habitats during brood-re-aring lead to brood size at fledging time to be slightly more than 2 goslings per family by mid-August (complete brood loss excluded). We may speculate that this heavy predation is partly explained by the neighbourhood of extraordinary big colony and brood / moulting geese con-centrations of Barnacle Geese at Peschanka delta, which, as it was observed, was an important attractor for nume-rous avian and mammalian predators.

48
Both nesting habitats and brood-rearing habitats are very diverse, and geese occupy almost all kind of boggy valleys, bushy upland and glacial lakes and creeks with rather homogenous nesting density. Having the suitable nesting area for White-fronted Geese on Kolguev about 4,500 sq.km and with average breeding density 40 nests / sq km (that remained exactly the same as it was 1995, see MOROZOV & SYROECHKOVS-KY 2004), we calculate entire breeding population as 180,000 breeding pairs, that together with 20% of non-breeders gives us approximately 450,000 White fronted geese arriving to Kolguev at spring. This comprises al-most one third of the total Western Palearctic Population of this species (MADSEN et al. 1999). We may conclude that vast wet plains of Kolguev serve as core breeding area for entire population with all ecological and conser-vational conclusions from that fact.Future investigations of White-fronted Geese and Bean Geese breeding density and breeding success in central parts of the island, far from the main Barnacle colony and associated predators is urgently needed.
6. Overall Summary
White-fronted Geese are the most numerous Arctic geese in the Western Palaearctic. Their breeding ran-ge is confined to the Russian Arctic tundra, between the Kanin and Taimyr peninsulas, and their wintering range is in Hungary, Bulgaria, Austria, Poland, Ger-many, The Netherlands, Belgium and England. Becau-se of the extended breeding range and the spread-out nature of the wintering areas, it is extremely difficult to collect reliable data on population size.
By using neckbands, satellite transmitters and expe-ditions to selected sites, we studied population size as well as the migration and breeding biology of this important species. The present report gives an overview of the activities and results of our studies in 2006, when we worked at the Olonets Fields (Lake Ladoga, Republic of Karelia, Russia) and on Kolguev Island (Nenets Autonomous Okrug, Russia), which is probably one of the most important nesting areas in the Western Palaearctic.
To analyse different techniques to assess the func-tion of a stepping-stone site within the flyway system, we selected the Olonets Fields, a seasonal bird sanctu-ary in Karelia that is well known from the literature as a goose staging site. During the second half of April and the first half of May 2006 we coun-ted and mapped the staging geese, studied the intensity and effects of goose hunting and read neckbands to cal-culate length of the staging period as well as turnover rates at this specific site.
We found a maximum of 16,800 White-fronts roosting at Olonets Fields. By using individually marked geese we calculated a 9,6% daily exchange rate in the area. This means the stock of staging geese is completely changed every 10 days and an estima-ted maximum of 100,000-120,000 White-fronts may use Olonets Fields as a step-ping stone to the bree-ding grounds during spring migration. By using the abdominal profile index (API) as a measure of body condition, we found out that the geese fatten up very quickly at Olonets Fields. During a single week they regain sufficient body condition to allow them to continue migration.

49
Whereas the geese spread out over the whole area in late April, they concentrated within the temporal hunting sanctuary after the 1st of May, when the hunting season opened. Inside the sanctuary we found mainly White--fronts. The Bean Geese had left the area with the start of the hunting season. We measured flushing distances in the sanctuary and found this to be strongly correlated with goose hunting and its intensity.
To look for further important staging sites on the migration route, we used five solar-GPS satellite transmitters, which were put on male White-fronted Geese in the Netherlands to gain technical and sci-entific experience with this new tool. Whereas two of the birds flew the expected route via the Baltic States and Karelia to the Archangelsk region, one of the geese took an unexpected alternative route, crossing Belarus and staging in Ukraine for three weeks before it turned north to the Kanin Peninsula. The Nemunas Delta (Lithuania), which was used by two of the three birds, probably is one of the most important staging sites in Eastern Europe. One of the birds was roosting near the Kaliningrad region. The importance of the Nemunas Delta, especially, should be studied intensively in the future.
On Kolguev Island we studied the breeding biology of White-fronted, Bean and Barnacle geese around our base camp on the northeastern part of the island. All goose nests were mapped and nest positions fixed with GPS, egg weight and size were measured, and hatching date calculated. We found very high densities of nesting White-fronts, Beans and Barnacle Geese and made a conservative estimate of the per-centage of the total breeding population nesting on the island. According to the counts of 2006, it seems that up to 20-30% of the White-fronted Goose, 30-50% of the Barnacle Goose and 10-15% of the Bean Goose populations may breed on this relatively small island. The breeding density is more then 40-times higher than on the Taimyr Peninsula, and more than 3-4 times higher than on the mainland around the Pechora Delta. So, based on the results from 2006, Kolguev is the most important nesting ground of the Western Palaearctic for the three goose species studied. Nesting conditions on this island have an extremely strong effect on the total populations of White-fronts, Beans and Barnacles.
Because all work on Kolguev Island has to be done on foot, we have been able to investigate only a part of the whole island. In the coming two seasons we want to investigate the nesting distribution and density of the-se Arctic-nesting geese on the entire island, in order to be able to make more reliable extrapolations from the Kolguev breeding populations of the three studied goose species.
The results of these Arctic studies should have a strong impact on the discussion of goose-flyway concepts as well as on the protection of these species. Such high nesting concentrations on such a relatively small island should result in specific protection measures.

50
die Olonets-Fields als Zwischenrastgebiet. Innerhalb nur einer Woche ist es den Gänsen im Gebiet mög-lich, ihre Körperkondition (d.h. ihre Fettreserven) soweit wieder aufzubauen, dass sie den Zug fortset-zen konnten.
Während sich die Gänse Ende April über ein weites Areal verteilten, konzentrierten sich die Gänse mit Beginn der Gänsejagd am 1. Mai ausschließlich im Vogelschutzgebiet. Im Schutzgebiet wurden überwie-gend Blessgänse angetroffen, während die Saatgänse das Rastgebiet nach Norden verließen. Wir maßen die Fluchtdistanzen im Vogelschutzgebiet und fanden einen engen Zusammenhang mit der einsetzenden Gänsejagd.
Im Januar 2006 wurden fünf Blessgansmännchen mit Satellitensendern in den Niederlanden versehen, um diese neue Technik (GPS Solar Transmitter) sowohl in wissenschaftlicher wie auch technischer Hinsicht zu erproben. Die Sender sollen ebenfalls dazu dienen, weitere wichtige Zwischenrastplätze im russischen Raum ausfindig zu machen, wo Beobachtungen mar-kierter Vögel außergewöhnlich selten gelingen. Während zwei Vögel durch Polen, das Baltikum an das Weiße Meer flogen, nahm ein Vogel eine andere Route: er flog vom Nemounas-Delta aus nonstop über Weißrussland hinweg und rastete für fast einen Mo-nat in der Nordukraine. Von hier aus flog der Vogel zur Kanin-Halbinsel. Das Nemounas-Delta wurde von zwei der drei Vögel als Zwischenrastgebiet ge-nutzt. Angesichts dessen, dass der dritte Sendervogel in der Kalingradregion rastete, darf festgestellt wer-den, dass das Gebiet um Nemounas-Delta vermutlich eines der wichtigsten Zwischenrastgebiete auf dem Zugweg in Nordosteuropa ist. Besonders die Bedeu-tung des Nemounas-Delta sollte in den kommenden Jahren intensiv untersucht werden.
Auf Kolguyev, einer Insel in der Barentssee, wur-den intensive Untersuchungen zur Brutbiologie der Bless-, Saat- und Nonnengänse durchgeführt. Die Nester aller Gänse wurden kartiert und georeferen-ziert, mittels von Eigewicht und –größe wurde das Schlupfdatum errechnet. Es wurden die höchsten Nestdichten von Bless-, Saat- und Nonnengänsen für die gesamte Westpaläarktis festgestellt. Auf Basis der Wintervogelzählungen von 2006 bedeuten diese Brutbestände, dass 20-30% aller Blessgänse, 30-50%
Zusammenfassung
Die Blässgansesind die individuenstärkste Gänseart der Westpaläarktis. Ihr Brutverbreitungsgebiet erstreckt sich in den arktischen Tundren zwischen der Kanin-Halbinsel und der Osthälfte Taimyrs. Den Winter verbringen die Blässgänse in einem Gürtel gemäßigter Temperaturen, der von Kasachstan über die Bulgarien, Rumänien, die Tür-kei, Ungarn, Österreich im Südosten sowie über Polen, Deutschland, die Niederlande und Belgien bis nach En-gland und Frankreich reicht. Aufgrund dieser weiträumi-gen Verbreitung ist es außergewöhnlich problematisch, genaue Daten über die Populationsgröße dieser Art zu sammeln. Durch die Anwendung moderner Forschungs-methoden wie der Markierung mit Halsmanschetten oder Satellitentelemetrie sowie durch direkte Untersuchungen an ausgewählten Rast- und Brutplätzen wurden Unter-suchungen zum Zugverhalten und der Brutbiologie der Blässgans als wichtiger Zugvogelart durchgeführt.
Durch die finanzielle Förderung des Vogelschutz-Komi-tee (VsK) e.V. konnte eine Reihe von Studien begonnen werden, die Aufschluß über wichtige Fragen zum Verhal-ten und letztlich zum Schutz der arktischen Gänsearten bringen sollen. Dieser Zwischenbericht gibt einen Über-blick über die Forschungsaktivitäten des Jahres 2006, in dem Untersuchungen mittels Satellitentelemetrie und am Zwischenrastplatz Olonets (Karelien, Russland) sowie im Brutgebiet (Kolguyev, Russland), den möglicherweise bedeutsamsten Brutplatz der Blessgänse, durchgeführt wurden.
Um mittels markierter Vögel, Bestandserfassungen und Köperkondition Bewertungsmethoden für Trittstein-gebiete zu entwickeln, wurde das Vogelschutzgebiete der Olonets Fields am Ladoga-See (Russisch-Karelien) ausgewählt, da dies aus der Literatur gut bekannt und zudem zahlreiche Erfassungsdaten vorlagen. In der zweiten Aprilhälfte und der ersten Maidekade wurde das Rastgeschehen sowie der Einfluss der Gänsejagd intensiv untersucht. Durch die Beobachtung markierter Gänse sollte die Aufenthaltsdauer sowie die Durchzugsraten im Gebiet analysiert werden. Maximal 16.800 Blessgänse rasteten in den Olonets Fields. Aufgrund der markier-ten Tiere errechneten wir eine tägliche Austauschrate von 9,6%. Das bedeutet, dass der Gesamtbestand aller rastender Gänse im Durchschnitt alle 10 Tage vollständig ausgetauscht wird. Auf die Durchzugszeit berechnet, nutzen also 100.000-120.000 verschiedene Individuen

51
aller Nonnengänse und 10-15% aller Saatgänse auf dieser relativ kleinen Insel brüten. Die Nestdichte der Blessgänse liegt 40mal höher als auf Taimyr und mehr als 3-4mal höher als auf dem Festland am Pechorafluss. Kolguyev ist demnach das wichtigste Brutgebiet für Gänse in der westlichen Paläarktis. Die Umweltbedingungen auf Kolguyev beeinflussen daher maßgeblich die gesamte Population von Bless-, Saat- und Nonnengans. Da die Untersuchungen auf Kolguyev ausschießlich zu Fuß durchgeführt wurden, konnte nur ein Teil der Insel untersucht werden. In den kommenden zwei Sommern soll daher sowohl der nordwestliche Teil der Insel sowie das Zentrum und der Süden gründli-cher untersucht werden.
Die Gänsepopulationen sind aufgrund dieser Kon-zentrationen der Brutplätze auf kleinem Raum empfindlicher für Lebensraum- oder Klimaverän-derungen als bisher angenommen. Die Ergebnisse dieser Arktisstudien müssen daher in der Diskussion um ein Zugwegkonzept der Bless- (und Saat-)gänse internationale Beachtung finden.
Белолобый гусь – наиболее многочисленный вид гусей в Западной Палеарктике. Его гнездовой ареал простирается в арктических тундрах между Канином и Таймыром, а районы зимовок расположены в Венгрии, Болгарии, Австрии, Польше, Германии, Нидерландах, Бельгии и Англии. В связи c протяженным гнездовым ареалом и обширным районом зимовок получить надежные данные относительной численности популяции очень трудно.Мы оценивали популяционную численность, а также изучали биологию этого важного вида в период миграций и размножения, используя мечение цветными ошейниками, спутниковыми передатчиками, а также в ходе экспедиционных исследований.В отчете содержится обзор наших активностей и результатов наших работ в 2006 году, когда мы проводили исследования белолобых гусей на Олонецких полях (побережье Ладожского озера, республика Карелия, Россия) и на острове Колгуеве (Ненецкий автономный округ, Россия), по всей вероятности – ключевом районе воспроизводства этого вида во всей Западной Палеарктике.С целью анализа различных методик оценки роли весенних миграционных стоянок в системе всего пролетного пути мы выбрали Олонецкие поля – сезонную охраняемую территорию со статусом «зоны покоя дичи», весьма известную из литературы как важное место долговременной стоянки гусей в период весеннего пролета. Во второй половине апреля и первой половине мая 2006 года мы оценивали численность и картировали распределение гусиных стай на полях, изучали интенсивность и влияние весенней охоты на гусей и наблюдали за индивидуально помеченными гусями с целью оценки длительности пребывания и сменяемости отдельных птиц в этом конкретном месте.По нашим оценкам максимальная единовременная численность белолобых гусей на Олонецких полях достигала 16800 птиц. На основе данных о продолжительности пребывания индивидуально помеченных птиц ежедневная

52
сменяемость особей может быть оценена в 9,6%. Это означает, что весь состав полностью сменяется в среднем в течение 10 дней, и примерно 100 000 – 120 000 гусей использует весной Олонецкие поля в качестве миграционной стоянки на пути к местам гнездования. Используя индексы абдоминального профиля (API) для оценки упитанности мы обнаружили, что набор жира у гусей на Олонецких полях происходит очень быстро. В течение одной недели они восстанавливают жировые запасы до уровня, позволяющего продолжить миграцию. В то время как в конце апреля гуси были распределены в пределах достаточно обширной территории, после 1 мая, когда начинался сезон весенней охоты, они концентрировались в пределах охраняемой «зоны покоя дичи». Гуменники к началу сезона охоты практически покинули Олонецкие поля. Мы измеряли дистанцию вспугивания у гусей в пределах охраняемой зоны и выявили сильное влияние охоты на степень доверчивости гусей к автотранспорту.Для того чтобы изучить другие важные места весенних стоянок в пределах миграционного пути и получить технический и научный опыт использования новых технологий мы использовали 5 спутниковых передатчиков с системой ГПС на солнечных батареях, установленных в Нидерландах на спинах самцов белолобых гусей. В то время как две птицы использовали вполне ожидаемый маршрут в район Архангельска через страны Балтии и Карелию, один из гусей использовал довольной неожиданный альтернативный маршрут, перелетев через Беларусь, он в течение трех недель находился на севере Украины, прежде чем продолжил маршрут на полуостров Канин. Дельта Немана в Литве, где останавливались две из трех птиц, является, по всей видимости, одним из важнейших районов весенних стоянок гусей во всей Восточной Европе. Одна из птиц останавливалась поблизости от Калининградской области. Важность дельты Немана требует интенсивного изучения в
будущем.На острове Колгуеве мы изучали гнездовую биологию белолобых гусей, гуменников и белощеких казарок в окрестностях временного палаточного лагеря на северо-востоке острова. Все найденные гнезда были закартированы при помощи ГПС, на основе промеров и взвешивания яиц были определены даты начала откладки яиц и вылупления птенцов. На основе полученных данных были сделаны выводы о чрезвычайно высокой плотности гнездования всех трех видов гусей на острове и проведены предварительные оценки доли популяции гнездящихся на Колгуеве гусей от общей численности вида. На основе данных 2006 года на этом сравнительно небольшом острове гнездится до 20-30% западно-палеарктических белолобых гусей, 30-50% белощеких казарок и 10-15% гуменников. Плотность гнездования этих видов более чем в 40 раз превышает плотность гнездования гусей на Таймыре и в 3-4 раза выше, чем в дельте Печоры. Таким образом, на основе данных 2006 года, остров Колгуев является ключевым районом воспроизводства для западно-палеарктических популяций этих трех видов гусей, и условия гнездования гусей на острове оказывают высокое влияние на всю европейскую популяцию этих трех видов гусей. Результаты данных исследований в Арктике будут крайне важны в плане обсуждения концепции пролетного пути при изучении и охране этих видов гусей. Столь высокие концентрации в период гнездования также заслуживают серьезного внимания при разработке природоохранных мер.

53
9. Acknowledgements
We would like to thank Vogelschutz-Komitee e.V., and UNEP/GEF ECORA project for the financial help to the expeditions, Murmanks oil company “Arcticmorneftegazrazvedka” helped in solving logistical problems in getting to Kolguev and cont-ributed to technical support of the work on Kolguev 2006. We‘d like to thank especially Marjourie Bousfield for highlighting the english texts. Our Russian colleagues Vladimir Zimin, Nikolai Lap-sin, and Alexander Artemjev shared their knowledge and experience in collecting data in Olonets regi-on. Data collection could not be possible without personal input of participants of the expeditons to Olonets and Kolguev: Hans-Heiner Bergmann, Valery Buzun, Matthias Fanck, Petr Glazov, Kai Toss, Juri Anisimov. For their technical help we are grateful to Konstantin Litvin (Bird Ringing centre, Moscow), Bart Ebbinge & Gerhard Müskens (Alterra institute, Wageningen), Albert Ardeev (Bugrino village of Kolguev), Society of Nederlandse Ganzenflappers, Microwave Ltd. and more than 2000 observers of colour-marked geese.Friedrich-Löffler-Institute (Riems) supported expedi-tions with necessary scientific equipment and cove-red costs for transport of equipment and samples.
PhotographsHelmut Kruckenberg: front top, 6, 8, 9, 12, 13,22, 23 (4) , 37, 45Alexander Kondratyev: front buttom, 42, Matthias Fanck: 23 (1)Kai Toss: 23 (2),Alena Dolgova : 47
10. Addresses of authors
DR. HELMUT KRUCKENBERG
Deutsche Ornithologen-Gesellschaft (DO-G), Pro-jektgruppe „Gänseökologie“European Whitefronted Research ProgrammeAm Steigbügel 3D-27283 VerdenGermanyemail: [email protected]
DR. JOHAN H. MOOIJ Biologische Station im Kreis Wesel e.V.Freybergweg 9D-46483 WeselGermanyemail: [email protected]
DR. CHRISTOPH ZÖCKLER
ArcCona Consulting 30 Eachard RoadUK-CB3 0HY CambridgeUnited Kingdomemail: [email protected]
DR. ALEXANDER KONDRATYEV
Faculty of Biology and Soil SciencesSt-Petersburg State UniversityUniversitetskaya emb 7/9RUS-199034 St-PeterburgRussia
ELMIRA ZAYNAGUTDINOVA
Faculty of Biology and Soil SciencesSt-Petersburg State UniversityUniversitetskaya emb 7/9RUS-199034 St-PeterburgRussia

54
11. Literature
BORZHONOV, B.B. (1986): On the morphological features of Bean Goose of Taimyr. In: Morphology, ecology and use of mam-mals and birds of Taimyr, - Sci.Techn.Bull., Siberian Sektion of the Acad.Agri.Sci. Novosibirsk 33: 9-12.
BUDEAU D.A., RATTI J., ELY C.R. (1991): Energy dynamics, foraging ecology, and behavior of prenesting Greater White-fron-ted geese. - J. Wildl. Manage. 55: 556-563
BURGERS, J., J.J. SMIT & H. VAN DER VOET (1991): Origins and systematics of two types of the Bean Goose Anser fabalis (Latham, 1787) wintering in the Netherlands. Ardea 79: 307-316.
DEKKERS, H. & B.S. EBBINGE (2004): Geese and swans – arctic migratory birds as guest at agricultural fields. – Fontaine, s’Graveland (in Dutch).
DRENT, R.H. & S. DAAN (1980): The prudent parent: energetic adjustments in avian breeeding. - Ardea 68: 225-252.EBBINGE, B.S. (1991). The impact of hunting on mortality rates and spatial distribution of geese wintering in the Western
Palearctic. - Ardea 79: 197-210.EBBINGE, B.S. (2000): Goose catching for science. – Alterra report 155, Wageningen (In Dutch).EBBINGE, B.S. & A.K.M. ST. JOSEPH (1991): The Brent Goose colour-ringing scheme: unraveling annual migratory movements
from high arctic Sibiria to the coasts of western Europe. – in: Proefschrijft B.S. EBBINGE (1992), Rijksuniversiteit Groningen.
EBBINGE, B.S. & B. SPAANS (1995): The importance of body reserves accumulated in spring staging areas in the temperate zone for breeding in Dark-bellied Brent Geese Branta b. bernicla in the high Artic. - J. Avian Biol. 26: 105-113.
EICHHORN, G. (2008): Travels in a chaging world. - PhD Rijksuniversiteit Groningen. ELY C.R. & A.X. DZUBIN (1994): Greater White-fronted Goose Anser albifrons. - The Birds of North America, No 131.ELY, C.R. & D.G. RAVELING (1984): Breeding biology of Pacific White-fronted Geese. - J. Wildl. Manage. 48: 823-837.GANTER, B., K. LARSSON, E.E. SYROECHKOVSKY, K.E. LITVIN, A.LEITO & J. MADSEN (1999): Barnacle Goose Branta leucopsis: Rus-
sia/Baltic. In: MADSEN J., G. CRACKNELL & A. D. FOX (eds) 1999: Goose populations of the Western Palearctic. A review of status and distribution. Wetlands International Publication No.48, Wetlands International, Wageningen, The Netherlands & National Environmental Research Institute, Rönde, Denmark: 270-283.
HAGEMEIJER, W.J.M. & M.J. BLAIR (eds, 1997): The EBCC Atlas of European Breeding Birds: their distribution and abundance. T&AD Poyser, London.
HUSTINGS, F. & J.-W. VERGEER (2002): Atlas of Dutch breeding birds. – SOVON Vogelonderzoek Nederland, Beek-Ubbergen (in Dutch).
JENESS, J. (2000): Whitefronted Geese – Extension Tool for ArcView 3.3KEAR, J. (2005): Ducks, Geese and Swans Vol. 1. - Oxford University Press, Oxford. KRUCKENBERG, H. & V. WILLE (2005): Wanderrouten und Wiederkehrraten am Unteren Niederrhein (Nordrhein-Westfalen)
markierter Blessgänse Anser alb albifrons 1999/00 und 2000/01. – Charadrius 39: 122-132. KRUCKENBERG, H. (2002): Muster der Raumnutzung markierter Blessgänse (Anser alb albifrons) in West- und Mitteleuropa unter
Berücksichtigung sozialer Aspekte. – Diss. University Osnabrück. KRUCKENBERG, H., B.S. EBBINGE & G. MÜSKENS (2007): Satellitentelemetrie von Blässgänsen Anser albifrons albifrons auf zudem
Frühjahrszug 2006 und 2007. – Vogelwarte 45: 330-331.KRUCKENBERG, H. & J.H. MOOIJ (2007): Warum Wissenschaft und Vogelschutz die Gänsejagd in Deutschland ablehnen. - Ber.
Vogelschutz 44: 107-119KRUCKENBERG, H. B.S. EBBINGE & G. MÜSKENS (2008): Reise der Blessgänse - Ergebnisse der Satellitentelemetrie. - Vogelwarte
46: 294-295.LITVIN, K.E. & SYROECHKOVSKY, E.E. (1996): Breeding biology of Tundra Bean Goose (Anser fabalis rossicus) in the North-east of
European Russia. - Casarca 2: 138-167. (in Russian) MADSEN, J. (1995): Impact of disturbance on migratory waterfowl. - Ibis 137: 67-74. MADSEN, J., G. CRACKNELL & A. D. FOX (eds) 1999: Goose populations of the Western Palearctic. A review of status and
distribution. Wetlands International Publication No.48, Wetlands International, Wageningen, The Netherlands & National Environmental Research Institute, Rönde, Denmark.
MOOIJ, J.H. & C. ZÖCKLER (1999): Reflections on the Systematics, Distribution and Status of Anser fabalis. - Casarca 5: 103-120.

55
MOOIJ, J.H. (1993): Development and management of wintering geese in the Lower Rhine area of North Rhine-Westfalia/Germany. - Die Vogelwarte 37: 55-77.
MOOIJ, J.H., I.O. KOSTIN & R. BRÄSECKE (1995): Deutsch-Russische Gänseforschung auf der Halbinsel Taiymr, Erste Ergebnis-se. - Beitr. Jagd- u. Wildforsch. 20: 275-301.
MOOIJ, J.H. (1997): The status of White-fronted Goose (Anser a. albifrons) in the Western Palearctic. - Vogelwarte 39: 61-81.MOROZOV, V.V. & E.E. SYROECHKOVSKY-JR. (2004): Materials to our knowledge of bird fauna of Kolguev. - Ornitologia, 31:
9-50 (in Russian).NOER, H. & J. MADSEN (1996): Shotgun pellet loads and infliction rates in pink-footed geese Anser brachyrhynchus. - Wildl. Biol.
2: 65-73. PRINS , H.H.TH. & R.C. YDENBERG , R.C. (1985). Vegetation growth and a seasonal habitat shift of the Barnacle Goose (Branta
leucopsis). - Oecologia 66: 122-125.ROGACHEVA, H. (1992): The Birds of Central Siberia. – Husum Verlag, HusumTOMKOVICH, P.S., M.Y. SOLOVIWV & E.E. SYROECHKOVSKY JR (1994): Bird of the Artic tundra of northern Taimyr (Knipovich
Bay area). In: Arctic tundras of Taimyr and Kara Sea Islands: nature, fauna and convervation problems, Vol. 1: 41-107.
VAN DER GRAAF, A. (2006): Geese on a green wave: flexible migrants in a changing world. - PhD Rijksuniversiteit Groningen. VAN DER GRAAF, A. J., J. STAHL, A. KLIMKOWSKA , J.P. BAKKER & R.H. DRENT (2006): Surfing on a green wave – how plant growth
drives spring migration in the Barnacle Goose Branta leucopsis. – Ardea 94: 567-577.WETLANDS INTERNATIONAL (1994): Waterfowl Population Estimates. - IWRB Publication 29.WETLANDS INTERNATIONAL (1997): Waterfowl Population Estimates. Second Edition. - Wetlands International Publication 44. WETLANDS INTERNATIONAL (1996): Atlas of Anatidae Populations in Africa and Western Eurasia. Wetlands International Publi-
cation 41.WETLANDS INTERNATIONAL (2002): Waterbird Population Estimates. Third Edition – Wetlands International Global Series No.
12.WETLANDS INTERNATIONAL (2006): Waterbird Population Estimates. Forth Edition – Wetlands International, Wageningen,
NetherlandsWILLE, V. (2000): Grenzen der Anpassungsfähigkeit überwinternder Wildgänse an anthropogene Nutzungen. - PhD universi-
ty of Osnabrueck. (in German). ZIMIN, V.B., A.V. ARTEMEV, N.V. LAPSCHIN & A.R. TJURLIN (2007): Olonetkie vesennnie skorleinia ptits - Gusi, - Nauka, Petro-
savosk (in Russian)ZÖCKLER, C. & I. LYSENKO (2000): Water birds on the edge. First circumpolar assessment of climate change impact on Arctic
Breeding Water Birds. WCMC Biodiversity Series No.11. 20p plus AnnexZÖCKLER, C., M. GAVRILO, V. BUZUN, A. KONDRATYEV, H. KRUCKENBERG (in press): Status and ecology of the Glaucous Gull
(Larus hyperboreus) on the ‘Goose- Island’ Kolguev, Barents Sea, Russia. - Arctic

56
Russian English Latin GermanКраснозобая гагара Red-throated Diver Gavia stellata SterntaucherЧернозобая гагара Black-throated Diver Gavia arctica PrachttaucherЧомга Great Crested Grebe Podiceps cristatus HaubentaucherБольшой баклан Great Cormorant Phalacrocorax carbo. KormoranБелый аист White Stork Ciconia ciconia WeißstorchЛебедь-кликун Whooper Swan Cygnus cygnus SingschwanМалый лебедь Bewick Swan Cygnus bewickii ZwergschwanБелолобый гусь White-fronted Goose Anser albifrons BlessgansГуменник Bean Goose Anser fabalis SaatgansСерый гусь Greyleg Goose Anser anser GraugansКанадская казарка Canada Goose Branta canadensis KanadagansБелощекая казарка Barnacle Goose Branta leucopsis NonnengansКряква Mallard Anas plathyrynchos StockenteСвиязь Wigeon Anas penelope PfeifenteЧирок-свистунок Teal Anas crecca KrickenteШилохвость Pintail Anas acuta SpießenteЧирок-трескунок Garganey Anas querquedula KnäkenteКрасноголовый нырок Pochard Aythia ferina TafelenteХохлатая чернеть Tufted Duck Aythia fuligula ReiherenteМорянка Long-tailed Duck Clangula hyemalis EisenteГоголь Goldeneye Bucephala clangula SchellenteЛуток Smew Mergellus albellus ZwergsägerБольшой крохаль Goosander Mergus merganser GänsesägerДлинноносый крохаль Merganser Mergus serrator MittelsägerОрлан-белохвост White-tailed Eagle Haliaethus albicilla SeeadlerБеркут Golden Eagle Aquila chrysaetos SteinadlerМалый подорлик Pomeranian Eagle Aquila pomarina SchreiadlerПолевой лунь Hen Harrier Circus cyaneus KornweiheБолотный лунь Marsh Harrier Circus aeruginosus RohrweiheЛуговой лунь Montagu’s Harrier Circus pygargus WiesenweiheСтепной лунь Pallid Harrier Circus macrourus SteppenweiheПерепелятник Sparrowhawk Accipiter nisus SperberТетеревятник Goshawk Accipiter gentiles HabichtКанюк Buzzard Buteo buteo MäusebussardЗимняк Rogh-legged Buzzard Buteo lagopus RaufussbussardСкопа Osprey Pandion haliaetus FischadlerПустельга Kestrel Falco tinnunculus TurmfalkeСапсан Peregrine Falcon Falco peregrinus Wanderfake
Table 1 : Bird list of Olonets Fields Expedition 2006
Appendix I

57
Дербник Merlin Falco columbarius MerlinБелая куропатка Willow Grouse Lagopus lagopus MoorschneehuhnГлухарь Capercailie Tetrao urugallus AuerhuhnТетерев Back Grouse Tetrao tetrix BirkhuhnРябчик Hazelhen Bonasia bonasia HaselhuhnЛысуха Common Coot Fulica atra BlesshuhnСерый журавль Common Crane Grus grus KranichЗолотистая ржанка Golden Plover Pluvialis apricaria GoldregenpfeiferЧибис Lapwing Vanellus vanellus KiebitzЧерныш Green Sandpiper Tringa ochropus WaldwasserläuferПеревозчик Common Sandpiper Actitis hyperleucos FlußuferläuferТравник Redshank Tringa totanus Rotschenkelщеголь Spottet Redshank Tringa erythropus DunkelwasserläuferБольшой улит Greenshank Tringa nebularia Grünschenkelфифи Wood Sandpiper Tringa glareola BruchwasserläuferТурухтан Ruff Philomachus pugnax KampfläuferБольшой кроншнеп Curlew Numenius arquatus Großer BrachvogelСредний кроншнеп Whimbrel Numenius phaeobus RegenbrachvogelБольшой веретенник Black-tailed Godwit Limosa limosa UferschnepfeВальдшнеп Woodcock Scolopax rusticola WaldschnepfeДупель Great Snipe Gallinago media DoppelschnepfeБекас Snipe Gallinago gallinago BekasineГаршнеп Jack Snipe Lymnocryptes minimus ZwergschnepfeКороткохвостый поморник Arctic Skua Stercorarius parasiticus SchmarotzerraubmöweОзерная чайка Black-headed Gull Larus ridibundus LachmöweМалая чайка Little Gull Larus minutus ZwergmöweСизая чайка Common Gull Larus canus SturmmöweКлуша Lesser Black-backed Gull Larus fuscus HeringsmöweСеребристая чайка Herring Gull. Larus argentatus Silbermöwe Речная крачка Common Tern Sterna hirunda FlussseschwalbeВяхирь Common Wood Pigeon Columba palumbus RingeltaubeУшастая сова Long-eared Owl Asio otus WaldohreuleБолотная сова Short-eared Owl Asio flammeus SumpfohreuleВертишейка Wryneck Jynx torquilla WendehalsБольшой пестрый дятел Great Spotted Woodpecker Dendrocopus major BuntspechtБелоспинный дятел White-backed Woodpecker Dendrocopus leucopus WeissrückenspechtЖелна Black Woodpecker Dryocopus martius SchwarzspechtПолевой жаворонок Sky Lark Alauda arvensis FeldlercheМалый жаворонок Short-toed Lark Calendrella brachydac-
tylaKurzzehenlerche
Рогатый жаворонок Shore Lark Eremophila alpestris OhrenlercheДеревенская ласточка Barn Swallow Hirundo rustica Rauchschwalbe

58
Луговой конек Meadow Pipit Anthus pratensis WiesenpieperЛесной конек Tree Pipit Anthus trivialis BaumpieperЖелтая трясогузка Yellow Wagtail Motacilla flava SchafstelzeБелая трясогузка White Wagtail Motacilla alba BachstelzeКрапивник (Winter-)Wren Troglodytes troglodytes ZaunkönigСвиристель Waxwing Bombycilla garrulus SeidenschwanzЛесная завирушка Dunnock Prunella modularis HeckenbraunelleЗарянка Robin Erithacus rubetra RotkehlchenВаракушка Bluethroat Luscinia svecica BlaukehlchenГорихвостка Redstart Phoenicurus phoenicu-
rusGartenrotschwanz
Каменка Wheatear Oenanthe oenante SteinschmätzerЛуговой чекан Whinchat Saxicola rubetra BraunkehlchenЧерный дрозд Blackbird Turdus merula AmselПевчий дрозд Song Thrush Turdus philomelus SingdrosselБелобровик Redwing Turdus iliacus RotdrosselРябинник Fieldfare Turdus pilaris WacholderdrosselДеряба Mistle Thrush Turdus viscivorus MisteldrosselПеночка-весничка Willow Warbler Philoscopus trochilus FitisПеочка-теньковка Chiffchaff Phylloscopus collybita ZilpzalpКоролек Goldcrest Regulus regulus WntergoldhähnchenМухоловка-пеструшка Pied Flycatcher Ficedula hypoleuca TrauerschnäpperБольшая синица Great Tit Parus major KohlmeiseЛазоревка Blue Tit Parus caeruleus BlaumeiseХохлатая синица Crested Tit Parus cristatus HaubenmeiseПухляк Willow Tit Parus montanus WeidenmeiseДлиннохвостая синица Long-tailed Tit Aegithalos caudatus SchwanzmeiseПищуха Short-toed Treecreeper Certhia familiaris WaldbaumläuferСерый сорокопут Great grey Shrike Lanius excubitor RaubwürgerСорока Magpie Pica pica Elster Сойка Jay Garrulus glandarius Eichelhäher Галка Jackdaw Corvus monedula Dohle Грач Rook Corvus frugilegus SaatkräheСерая ворона Carrion Crow Corvus corone cornix AaskräheВорон Raven Corvus corax KolkrabeСкворец Common Starling Sturnus vulgaris StarДомовый воробей House Sparrow Passer domesticus HaussperlingЗяблик Chaffinch Fringilla coelebs BuchfinkЮрок Brambling Fringilla montifringilla BergfinkЩегол Goldfinch Carduelis carduelis StieglitzЧечетка Redpoll Carduelis flammea BirkenzeisigЧиж Siskin Carduelis spinus Zeisig
59
Коноплянка Linnet Carduelis cannabina HänflingСнегирь Bullfinch Pyrrhula pyrrhula DompfaffКлест-еловик Common Crossbill Loxia curvirostra FichtenkreuzschnabelКамышовая овсянка Reed Bunting Emberiza schoeniclus RohrammerОбыкновенная овсянка Yellowhammer Emberiza citrinella GoldammerЛапландский подорожник Lapland Bunting Calcarius lapponicus SpornammerПуночка Snow Bunting Plectrophenax nivalis Schneeammer
60
Russian English Latin StatusКраснозобая гагара
Red-throated Diver Gavia stellata b common breeding species
Чернозобая гагара Black-throated Diver Gavia arctica b common breeding speciesБелоклювая гагара Yellow-billed Diver Gavia adamsi v not seen in 2006Глупыш Northern Fulmar Fulmarus glacialis v not seen in 2006Лебедь-кликун Whooper Swan Cygnus cygnus v seen once on 29.07.2006 in Peschanka
deltaМалый лебедь Bewick’s Swan Cygnus bewickii b common breeding speciesСерый гусь Greylag Goose Anser anser v not seen in 2006Белолобый гусь White-fronted Goose Anser albifrons b common breeding speciesПискулька Lesser white-fronted Goose Anser erythropus ? 2 birds seen 31.06.2006 at middle
PeschankaГуменник Bean Goose Anser fabalis b common breeding speciesГорный гусь Bar-headed Goose Anser indicus v seen once on 8.08.2006 in Peschanka
deltaГусь Росса Ross goose Anser rossi v male recorded breeding in pair with
Branta leucopsis in Peschanka deltaБелощекая казарка Barnacle Goose Branta leucopsis b numerous breeding speciesЧерная казарка Brent / Black brant Branta bernicla v recorded once on 26.06.2006 in coastal
marshes southeast of Peschanoye lakeКраснозобая казарка
Red-breasted Goose Branta ruficollis v not seen in 2006
Свиязь Wigeon Anas penelope v seen in 2006 on Pervaya river by Anufriev
Чирок-свистунок Common Teal Anas crecca b common breeding speciesКряква Mallard Anas platyrhynchos v seen once on 7.08.2006 on spits near
Light houseШилохвость Pintail Anas acuta b common breeding speciesХохлатая чернеть Tufted Duck Aythya fuligula v not seen in 2006Морская чернеть Greater Scaup Aythya marila b common breeding speciesГага Eider Somateria mollis-
sima b recorded near camp site, recorded at
see in lagoon entranceГага-гребенушка King Eider Somateria specta-
bilis b breeding species of coastal, rarely
inland habitatsМорянка Long-tailed Duck Clangula hyemalis b common breeding speciesСиньга Common Scoter Melanitta nigra b rare breeding speciesТурпан Velvet Scoter Melanitta fusca b breeds in Kolguev interiorГоголь Common Goldeneye Bucephala clangula mt moulting flocks recorded in lagoon
and at Peschanoye Lake
Table 2: Bird list of Northeastern Kolguev summer 2006
Appendix II

61
Длинноносый крохаль
Red-breasted Merganser Mergus serrator b uncommon, probably breeding species
Большой крохаль Common Merganser Mergus merganser mt moulting flocks recorded in lagoon 4.07 and 28.07.2006
Зимняк Rough-legged Buzzard Buteo lagopus b breeding speciesОрлан-белохвост White-tailed Eagle Haliaeetus albicilla v not seen in 2006Сапсан Peregrine Falcon Falco peregrinus b common breeding speciesДербник Merlin Falco columbarius b seen once on 17 & 19.06.2006Белая куропатка Willow Grouse Lagopus lagopus b common breeding speciesСерый журавль Common Crane Grus grus v 2 birds seen near Bugrino 18.06.2006Тулес Black-bellied Plover Squatarola squa-
tarola b common breeding species
Золотистая ржанка
Golden Plover Pluvialis apricaria b common breeding species
Бурокрылая ржанка
Pacific Golden Plover Pluvialis fulva v seen once near Bugrino at spring in flocks of P.apricaria
Галстучник Ringed Plover Charadrius hiat-icula
b common breeding species
Хрустан Dotterel Eudromias mori-nellus
m not seen in 2006
Камнешарка Ruddy Turnstone Arenaria interpres b rare breeding speciesКулик-сорока Oystercatcher Haematopus os-
tralegus b rare breeding species
Фифи Wood Sandpiper Tringa glareola b rare breeding speciesЩеголь Spotted Redshank Tringa erythropus b not seen in 2006Мородунка Terek Sandpiper Xenus cinereus v not seen in 2006Плосконосый плавунчик
Red Phalarope Phalaropus fuli-carius
v 2 birds seen in lagoon 28.07.2006
Круглоносый плавунчик
Red-necked Phalarope Phalaropus lobatus b common breeding species
Турухтан Ruff Philomachus pugnax
b common breeding species
Кулик-воробей Little Stint Calidris minutus b common breeding speciesБелохвостый песочник
Temminck’s Stint Calidris tem-minckii
b common breeding species
Краснозобик Curlew Sandpiper Calidris ferruginea m rare migrant, 4 birds seen 15.07.2006Чернозобик Dunlin Calidris alpina b common breeding speciesМорской песочник Purple Sandpiper Calidris maritima m seen 9.06.2006 at Pervaya river and at
spits 28.07.2006Исландский песочник
Red Knot Calidris canutus m seen at coastal marshes 27-29.07.2006
Песчанка Sanderling Calidris alba m seen at spits 28.07.2006Гаршнеп Jack Snipe Lymnocryptes
minimus b breeding species
Бекас Common Snipe Gallinago gallinago b breeding species

62
Средний кроншнеп
Whimbrel Numenius phaeo-pus
v not seen in 2006
Большой поморник
Skua Stercorarius skua b breeding recorded in coastal marshes in 2006
Средний поморник
Pomarine Skua / Jaeger Stercorarius poma-rinus
s nonbreeders seen on migration in spring and at sea in summer
Короткохвостый поморник
Arctic Skua / Parasitic Jaeger Stercorarius para-siticus
b common breeding species
Длиннохвостый поморник
Long-tailed Skua / Jaeger Stercorarius longi-caudus
m several records made in spring and summer
Халей Heuglin’s Gull Larus heuglini b common breeding speciesБургомистр Glaucous Gull Larus hyperboreus b common breeding speciesМорская чайка Great Black-backed Gull Larus marinus v rare visitor, seen at spring near Bu-
grino, 21.07 at Peschanka and 25,29.07 at sea coast
Моевка Black-legged Kittiwake Rissa tridactyla b breeds at “Ob’” ship near BugrinoБелая чайка Ivory Gull Pagophila eburnea v not seen in 2006Полярная крачка Arctic Tern Sterna paradisaea b breeds at spitsТонкоклювая кайра
Common Murre Uria aalge v not seen in 2006
Толстоклювая кайра
Thick-billed Murre Uria lomvia v not seen in 2006
Тупик Puffin Fratercula arctica v not seen in 2006Белая сова Snowy Owl Nyctea scandiaca b seen at spring 2.06, male stayed at
Peschanka for summerБолотная сова Short-eared Owl Asio flammeus m seen on spring migration 31.05 and
3.06.2006 and 4.06 near BugrinoБольшой пестрый дятел
Great Spotted Woodpecker Dendrocopos major v not seen in 2006
Береговая ласточка
Bank Swallow Riparia riparia v not seen in 2006
Деревенская ласточка
Barn Swallow Hirundo rustica v not seen in 2006
Городская ласточка
House Martin Delichon urbica v not seen in 2006
Рогатый жаворонок
Horned Lark, Shorelark Eremophila alp-estris
b breeding species
Луговой конек Meadow Pipit Anthus pratensis b common breeding speciesКраснозобый конек
Red-throated Pipit Anthus cervinus b common breeding species
Желтая трясогузка Yellow Wagtail Motacilla flava v not seen in 2006Желтоголовая трясогузка
Citrine Wagtail Motacilla citreola v not seen in 2006
Белая трясогузка White Wagtail Motacilla alba b breeding speciesСкворец Starling Sturnus vulgaris v not seen in 2006Серая ворона Hooded Crow Corvus cornix v 2 birds stayed near Bugrino in June

63
Ворон Raven Corvus corax v seen 20.07, 25.07 and 4.08Свиристель Waxwing Bombycilla gar-
rulus v 6 birds seen in Bugrino 20.05.2006
Камышевка-барсучок
Sedge Warbler Acrocephalus schoenobaenus
b breeding species
Пеночка-весничка Willow Warbler Phylloscopus tro-chilus
b breeding species
Пеочка-теньковка Chiffchaff Phylloscopus col-lybita
v not seen in 2006
Каменка Northern Wheatear Oenanthe oenanthe b breeding speciesВаракушка Bluethroat Luscinia svecica b breeding speciesРябинник Fieldfare Turdus pilaris v 2 birds seen in central part of island
5.07.2006Белобровик Redwing Turdus iliacus b breeding speciesДомовый воробей House Sparrow Passer domesticus v not seen in 2006Чечетка Common Redpoll Acanthis flammea b breeding speciesКлест-сосновик Parrot crossbill Loxia pityopsit-
tacus v not seen in 2006
Камышовая овсянка
Reed Bunting Emberiza schoe-niclus
b rare breeding species
Овсянка-крошка Little Bunting Emberiza pusilla b breeding speciesЛапландский подорожник
Lapland Longspur Calcarius lapponi-cus
b common breeding species
Пуночка Snow Bunting Plectrophenax nivalis
b breeding species
b - breedingm - migratant mt - moultingv - vagrants - spends summer? - status unknown

64
EURING Species engl. Name Moscow Ring No colour-marking Date sex age01591 Anser albifrons Whitefronted Goose Moscow CS-006501 ANB black 01.07.2006 female adult01591 Anser albifrons Whitefronted Goose Moscow CS-006502 GGU black 02.07.2006 female adult01591 Anser albifrons Whitefronted Goose Moscow CS-006503 23.07.2006 male pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006504 19.07.2006 male pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006505 19.07.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006506 19.07.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006507 23.07.2006 unknown pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006509 20.07.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006511 03.08.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006512 ZPA black 03.08.2006 male adult01591 Anser albifrons Whitefronted Goose Moscow CS-006513 03.08.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006515 ZTK black 03.08.2006 male pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006516 03.08.2006 male pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006518 03.08.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006519 F75 lime 03.08.2006 female adult01591 Anser albifrons Whitefronted Goose Moscow CS-006520 F93 lime 06.08.2006 male adult01591 Anser albifrons Whitefronted Goose Moscow CS-006521 20.07.2006 unknown pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006522 30.07.2006 male pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006523 30.07.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006524 01.08.3006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006525 01.08.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006526 02.08.2006 unknown pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006527 02.08.2006 unknown pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006530 ZPR black 06.08.2006 male adult01591 Anser albifrons Whitefronted Goose Moscow CS-006531 06.08.2006 male pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006532 06.08.2006 male pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006533 06.08.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006534 F88 lime 06.08.2006 male adult01591 Anser albifrons Whitefronted Goose Moscow CS-006535 ZTY black 06.08.2006 female adult01591 Anser albifrons Whitefronted Goose Moscow CS-006536 ZPK black 06.08.2006 male adult01591 Anser albifrons Whitefronted Goose Moscow CS-006537 F81 lime 06.08.2006 female adult01591 Anser albifrons Whitefronted Goose Moscow CS-006538 ZPT black 06.08.2006 female adult01591 Anser albifrons Whitefronted Goose Moscow CS-006539 ZTB black 06.08.2006 female adult01591 Anser albifrons Whitefronted Goose Moscow CS-006540 F80 lime 06.08.2006 male adult01591 Anser albifrons Whitefronted Goose Moscow CS-006545 06.08.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006546 ZPZ black 06.08.2006 male adult01591 Anser albifrons Whitefronted Goose Moscow CS-006547 06.08.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006548 06.08.2006 male pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006549 06.08.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006552 11.08.2006 female pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006564 09.08.2006 unknown pulli01591 Anser albifrons Whitefronted Goose Moscow CS-006565 F76 lime 09.08.2006 female adult01591 Anser albifrons Whitefronted Goose Moscow CS-006566 ZPC black 09.08.2006 male adult01591 Anser albifrons Whitefronted Goose Moscow CS-006567 F77 lime 09.08.2006 female adult01591 Anser albifrons Whitefronted Goose Moscow CS-006568 ZPS black 09.08.2006 male adult01574 Anser fab.rossicus Tundra Bean Goose Moscow CS-006510 03.08.2006 unknown pulli01574 Anser fab.rossicus Tundra Bean Goose Moscow CS-006514 03.08.2006 unknown pulli01574 Anser fab.rossicus Tundra Bean Goose Moscow CS-006517 P01 yellow 03.08.2006 male adult01574 Anser fab.rossicus Tundra Bean Goose Moscow CS-006561 08.08.2006 unknown pulli01574 Anser fab.rossicus Tundra Bean Goose Moscow CS-006562 P02 yellow 09.08.2006 female adult01574 Anser fab.rossicus Tundra Bean Goose Moscow CS-006563 P03 yellow 09.08.2006 unknown pulli10110 Anthus pratensis Meadow Pipit Moscow XY01807 08.07.2006 unknown pulli10110 Anthus pratensis Meadow Pipit Moscow XY01832 01.08.2006 unknown pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010604 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010605 LKO7 27.07.2006 female adult
Appendix III
Table 3: Birds ringed on Kolguev in 2006

65
01670 Branta leucopsis Barnacle Goose Moscow DS-010606 LJO5 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010607 LJO6 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010608 LNO3 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010609 LNO8 27.07.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010610 LNO9 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010611 LTO8 27.07.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010612 LTO9 27.07.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010613 LTOA 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010614 LTOC 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010615 LTOD 27.07.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010616 LYO6 27.07.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010617 LYOA 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010618 LYOC 27.07.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010619 LYOD 27.07.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010620 27.07.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010621 27.07.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010622 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010623 27.07.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010624 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010625 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010626 27.07.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010627 27.07.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010628 27.07.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010629 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010630 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010631 27.07.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010632 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010633 27.07.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010634 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010635 27.07.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010636 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010637 27.07.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010638 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010639 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010640 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010641 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010642 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010643 27.07.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010644 27.07.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010645 03.08.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010646 03.08.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010647 03.08.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010648 03.08.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010649 03.08.2006 male juv01670 Branta leucopsis Barnacle Goose Moscow DS-010650 06.08.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010651 06.08.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010652 06.08.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010653 06.08.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010654 06.08.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010655 06.08.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010656 06.08.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010657 06.08.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010658 06.08.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010659 06.08.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010660 06.08.2006 female pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010661 06.08.2006 male pulli01670 Branta leucopsis Barnacle Goose Moscow DS-010662 06.08.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010663 06.08.2006 female adult01670 Branta leucopsis Barnacle Goose Moscow DS-010664 06.08.2006 male adult01670 Branta leucopsis Barnacle Goose Moscow DS-010665 06.08.2006 female adult02900 Buteo lagopus Rough-legged Buzzard Moscow CS-006528 02.08.2006 unknown pulli02900 Buteo lagopus Rough-legged Buzzard Moscow CS-006529 02.08.2006 unknown pulli02900 Buteo lagopus Rough-legged Buzzard Moscow CS-006550 08.08.2006 unknown pulli

66
18470 Calcarius lapponicus Lappland Bunting Moscow XY01801 01.07.2006 unknown juv18470 Calcarius lapponicus Lappland Bunting Moscow XY01802 01.07.2006 unknown juv18470 Calcarius lapponicus Lappland Bunting Moscow XY01816 20.07.2006 unknown pulli18470 Calcarius lapponicus Lappland Bunting Moscow XY01831 01.08.2006 unknown pulli18470 Calcarius lapponicus Lappland Bunting Moscow XY01846 30.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13501 02.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13502 02.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13503 02.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13504 02.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13505 02.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13506 02.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13507 02.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13508 02.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13509 08.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13510 08.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13511 03.07.2006 unknown pulli05120 Calidris alpina Dunlin Moscow KS 13512 03.07.2006 unknown 05120 Calidris alpina Dunlin Moscow KS 13513 12.07.2006 unknown 05120 Calidris alpina Dunlin Moscow KS 13514 12.07.2006 unknown 05120 Calidris alpina Dunlin Moscow KS 13521 22.07.2006 unknown pulli05010 Calidris minuta Little Stint Moscow XY01825 25.07.2006 unknown pulli05010 Calidris minuta Little Stint Moscow XY01826 25.07.2006 unknown pulli05010 Calidris minuta Little Stint Moscow XY01827 25.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01804 04.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01805 04.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01806 06.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01808 15.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01810 02.08.2006 unknown juv05020 Calidris temmincki Temminck Stint Moscow XY01812 22.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01813 22.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01818 15.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01819 15.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01828 25.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01829 25.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01830 25.07.2006 unknown pulli05020 Calidris temmincki Temminck Stint Moscow XY01837 11.08.2006 unknown pulli04700 Charadrius hiaticula Ringed Plover Moscow XY01820 08.08.2006 unknown pulli04700 Charadrius hiaticula Ringed Plover Moscow XY01821 08.08.2006 unknown pulli04700 Charadrius hiaticula Ringed Plover Moscow KS13515 26.07.2006 unknown pulli09780 Eremophila alpestris Shorelark Moscow XY01803 02.07.2007 unknown juv09780 Eremophila alpestris Shorelark Moscow XY01817 22.07.2006 unknown juv03200 Falco peregrinus Peregrine Falcon Moscow CS-006541 28.07.2006 unknown pulli03200 Falco peregrinus Peregrine Falcon Moscow CS-006542 28.07.2006 unknown pulli03200 Falco peregrinus Peregrine Falcon Moscow CS-006543 28.07.2006 unknown pulli03200 Falco peregrinus Peregrine Falcon Moscow CS-006544 28.07.2006 unknown pulli03200 Falco peregrinus Peregrine Falcon Moscow DS-010601 23.07.2006 female pulli03200 Falco peregrinus Peregrine Falcon Moscow DS-010602 23.07.2006 male pulli03200 Falco peregrinus Peregrine Falcon Moscow DS-010603 23.07.2006 male pulli05990 Larus hyperboreus Glaucous Gull Moscow DS-010666 06.08.2006 unknown pulli05990 Larus hyperboreus Glaucous Gull Moscow DS-010667 06.08.2006 unknown pulli05990 Larus hyperboreus Glaucous Gull Moscow DS-010668 06.08.2006 unknown pulli11460 Oenanthe oenanthe Wheatear Moscow XY01811 21.07.2007 unknown juv05640 Phalaropus lobatus Red necked Phalarope Moscow XY01809 19.07.2006 unknown pulli05170 Philomachus pugnax Ruff Moscow TA-35802 22.07.2006 unknown 13120 Phylloscopus trochilus Willow Warbler Moscow XY01814 24.07.2006 unknown juv13120 Phylloscopus trochilus Willow Warbler Moscow XY01815 24.07.2006 unknown juv04850 Pluvialis apricaria Golden Plover Moscow TA-35806 28.07.2006 unknown 04860 Pluvialis squatorola Grey Plover Moscow TA-35801 21.07.2006 unknown 02060 Somateria mollissima Common Eider Moscow CS-006503 04.07.2006 male adult05690 Stercorarius skua Great Skua Moscow DS-010669 11.08.2006 unknown pulli

67

68
page
1 Introduction 3
2 Flyway approach 7
3 Stop-over site Olonets Fields
15
4 Research on nesting grounds: Kolguev
25
5 Discussion 46
8 Overall summary 48
Zusammenfassung 50
Белолобый гусь 51
9 Acknowlegdement 53
10 Addresses of authors 53
11 Literature 54
Appendix I Bird list of Olonets Fields 2006 56
Appendix II Bird list of NE Kolguev 2006 60
Appendix III Birds ringed on Kolguev 2006 64
Content Inhaltsverzeichnis
Copyright © 2022 FDOKUMEN




















