
Sublingual immunotherapy for allergic rhinitis: systematic review and meta-analysis
Upload
independentCategory
view
5download
0
Clinical and Experimental Allergy, 1992, Volume 22, pages 734-740
REVIEW
Kinins and rhinitis
K. RAJAKULASINGAM, R. POLOSA, M. K. CHURCH, S. T. HOLGATE andP. H. HOWARTH
Medicine 1, Southampton General Hospital, Tremona Road. Southampton
Introduction
Allergic rhinitis is characterized by cell accumulation andcell activation within the nasal mucosa along withenhanced plasma vascular permeability. This disorderedpathophysiology is associated with sneezing, rhinorrhoeaand nasal congestion, symptoms which are common toboth allergic and non-allergic rhinitis. The introductionof H|-antihistamines in the i940s differentiated theseaetiologies, in that treatment benefit could be identifiedwith this mode of therapy in allergic but not infectiousrhinitis. Even in allergic rhinitis, however. Hi-antihista-mines are not fully effective, especially with respect torelief of nasal blockage [1]. It is thus apparent thatmediators other than histamine contribute to the patho-physiology of rhinitis. In this respect prostaglandins.leukotrienes and kinins have all been identified in associa-tion with experimental exacerbations of disease ex-pression [t]. This review will consider the possible role ofkinins in rhinitis.
Kinin structure
Kinins are potent vasoactive peptides formed de novo inbody fluids and tissues during inflammation. They arederived from the 0L2 globulins, high and low molecularweight kininogens. through proteolytic cleavage by avariety of enzymes, most important of which are tissueand plasma kallikreins [2]. Three distinct kinins have beenisolated in man, bradykinin, kallidin (lys-bradykinin) andmet-lys-bradykinin (see structures below) [3]. All areoligopeptides which have in common the nine amino acidC-terminal sequence of the nonapeptide bradykinin.
H-Arg-Pro-Pro-Gly-Phc-Ser-Pro-Phe-Arg-OilH-Lys-Arg-Pro-Pro-Gly-Phc-Ser-Pro-Phe-Arg-OHH-Mei-Lys-Arg-Pro-Pro-Gly-Phe-SerPro Phe Arg-
BradykininKallidin
Mcl-lys-bradykinin
Kinin generation
Kinin generation in inflammation is known to occur viathree different pathways: a plasma pathway, a tissue
Correspondence: Dr K. Rajakulasiiigam, Medicine 1, Centre Block,Southampton General Hospital, Tremona Road, Southampton SO94XY.
pathway and a plasma/tissue independent pathway (Fig.1). Kinin production by the plasma pathway, also knownas the Hagemun factor (HF) dependent pathway, isinitiated by the interaction of factor XII (HF) [4] with thecirculating, loosely-adherent, complex of prekallikrein [5]and high molecular weight kininogen [6]. induced byexposure to negatively charged surfaces, e.g. sulphatedpolysaccharides and acidic groups. Activated Hagemanfactor (factor Xlla) initiates the conversion of prekalli-krein to kallikrein. which in a positive feed back mechan-ism rapidly activates the further conversion of factor XIIto Xlla. Kallikrein has been identified as the majorprotease capable of activating HF [7], an action aug-mented by high molecular weight kininogen (HMWK).While enzymes other than kallikrein are also able tocleave and activate HF, including activated factor XI(XIa) and plasmin [7.8]. these stimuli are less efficacious.The active kallikrein released by this sequence of eventscleaves HMWK to release the biologically active peptidebradykinin (Fig. 1).
The tissue pathway for bradykinin synthesis involvestissue kallikreins. which are single chain, acidic glycopro-teins, with molecular weights ranging from 25 to 43 kD,that are physicochemically and immunologically distinctfrom plasma kallikrein [9]. These kallikreins are widelydistributed throughout the body tissues, with particularlyhigh levels in pancreas [10]. prostate gland [11], kidneys[12]. salivary glands [ 13], pituitary gland [14] and the lungs[15]. Although this indicates a potential for kinin genera-tion at multiple discrete sites its specific role at these sites isunclear. While HMWK is clearly tbe preferred substratefor plasma kallikrein. tissue kallikreins readily releasekinins from both high and low molecular weight kinino-gens (LM WK). the latter comprising approximately 70'̂ )of the total human kininogen pool [16.17]. Thus the abilityof tissue kallikreins to react equally well with HMWK andLMWK provides these enzymes with a larger pool ofavailable substrate than is available to plasma kallikrein[16]. Once activated, tissue kallikreins are less susceptibleto inhibition by protease inhibitors than plasma kallikreinand they are the only enzymes known to hydrolyse twodissimilar bonds in LMWK lo generate the bradykininprecursor kallidin. Kallidin may be converted into brady-
734

Kinins and rhinitis 735
PLASMA
PREKALLIKREIN-I HwlWK
KALLIKREIN
JI-MWK
I TISSUE I
PRO-KALLIKREIN
PlasminTissue kallikrein
1
TISSUE KALLIKREIN
LMWK
-HBRADYKININ
PUSMA/riSSUE INDEPENDENT
HMWK & LMWK
Non-kallikfeinKjninogenases
LYS-BRADYKiNINAmino-peptjdase
[DES-ARG9J-BRADYKININ-*-CPN
BRADYKININ -*-
ACEproteases
INACTIVE PEPTIDE, AMINO ACIDS
Fig. I. Mechanisms of hradykinin formalion by plasma, tissue and plasma/tissue independent pathways. HMWK, high molecularwcighl kininogen. LMWK, low molecular wcighl kininogen. CPN, carboxy peplidase/V. ACE, angiotensin converting enzyme. HF,Hageman factor. HFA. activated Hageman factor.
kinin by the removal of 7V-terminal lysine by the actions ofaininopeptidases and trypsin (Fig. 1).
The third pathway, the plasma/tissue independentpathway, involves the actions of kininogenases other ihankallikreins in the cleavage of HMWK and LMWK toform bradykinin. Elastase released from activated neu-trophils possesses kininogenase activity [18] and may beinvolved in the production of met-lys-bradykinin. Ofrelevance to allergic airways disease are the observationsthai both mast cells [19] and basophils [20] possesskininogenase-like activity and may thus be involved inkinin generation in allergic inflammation.
Kinin metabolism
Bradykinin and kallidin have a circulating half-life of lessthan 15 and 25 sec respectively [2]. The degradation ofbradykinin is initiated by the action of carboxypeplidase/V, which cleaves the terminal phe-arg bond to yield theinactive octapeptidc [dcs-ar'']-bradykinin [21]. The [des-urg'']-bradykinin thus formed is inactive within the nose[22] although it has been shown to have activity in severalIn vitro vascular models [23]. Further degradation of [des-arg.,]-bradykinin is achieved by the actions of angioten-sin-converting enzyme and plasma proteases to produceinactive pentapeptide, tripeptide and single amino acidfragments [24]. Endopeptidases may also play a role in themetabolism of bradykinin. Endopeptidase24.15, ametal-
loendopeptidase, is present in rat lung and is also capableof degrading bradykinin [25]. The role of endopeptidasesin human kinin metabolism is, however, not clearlydetermined.
Evidence for involvement in rhinitis
To implicate kinins in the pathogenesis of rhinitis it isnecessary to satisfy three criteria. First, kinin generationoccurs in association with disease expression. Second,when administered in physiological quantities to the nose,kinins reproduce at least some of the symptoms orpathophysiological abnormalities of the disease. Third,specific receptor antagonism or inhibition of kinin syn-thesis prevents those symptoms produced by exogenouskinins, At present only two of these criteria can be met. asthere are no highly selective and specific antagonists orkallikrein inhibitors that have acceptance for use inclinical practice. Thus the role of kinins in rhinitis mustrely on circumstantial evidence which strongly suggeststhat they are axially involved in the pathogenesis of thiscommon disease.
Kinin generation in relationship to disease expression
Nasal lavage has been used widely to sample the nasallining fluid in order to measure levels of inflammatorymediators. Using this technique, increased levels of kininshave been demonstrated in naturally occurring seasonal

736 K. Rajakulasingam et al.
allergic rhinitis and during naturally occurring upperrespiratory tract infections with rbinovirus [26,27]. In thelatter condition, the levels of kinins in secretions arcsignificantly greater when infection and symptoms arepresent than when they are not [27]. Experimental viralnasal inoculation [28] and allergen challenge [29] hasconfirmed these findings.
Nasal challenge with allergen has been used in anattempt to dissect out the mechanism of kinin formationand its relationship to disease expression in allergicrhinitis. Kinin generation has been identified in both theearly [30.31] and late [32] nasal responses to allergenchallenge. During the early response, kinin formationcorrelates well with the increase in nasal lavage fluid ofother inflammatory mediators including histamine,prostaglandin D^ and the sulphidopeptide leukotrienes[29,31,33]. Further analysis of this kinin pool has shown itto be a mixture of bradykinin and kallidin [31] andconsistent with this, enzyme and precursor pools containboth plasma [34] and tissue [35] kallikreins together withhigh and low molecular weight kininogens [36]. Theincrease in plasma kallikrein activity in nasal lavage isparalleled by increased recovery of kinins, histamine and7V-a-tosyl-L-arginine methyl esterase (TAME-esterase)activity and with the onset of nasal symptoms, suggestinga relationship between kinin formation and plasmaextravasation [34]. TAME-esterase activity has beenshown in several studies to correlate with immediateallergic response and the majority of ihc TAME-esteraseactivity detected in nasal secretions during the allergicresponse is due to activities consistent with a plasmakallikrein/a:>-macroglobulin complex and mast cell tryp-tase [34]. Purified human lung mast cells have also beenshown to contain a kininogenase that is released in a dose-dependent fashion by anti-IgE [ 19]. As mast cell heparin isnegatively charged, it has been proposed that the co-release of vasopermeable mediators (histamine andleukotrienes) along with heparin following immunologi-cal mast cell degranulation Ciin account lor the enhancedvascular permeability and the subsequent extravascularconversion of prekallikrein to kallikrein and the genera-lion of kinins. Contrary to this, however, tissue kallikreinhas been shown to be responsible for most of tbekininogenase activity in post-allergen challenge lavage[35] and at least 45 "/n of the kinin detected during the earlyallergic response comprises kallidin [31]. Concordantwith this, elevated levels of aminopeptidase, capable ofconverting kallidin to bradykinin, huve been demon-strated in post-allcrgcn challenge nasal tavage [37].
The metabolism of bradykinin and kallidin during theearly response is considered to be regulated by carboxy-peptidase-A' (CPN) followed by additional cleavage byangiotensin converting enzyme (ACE) [21,24]. Increased
activity of carboxypeptidase-A'^has been demonstrated inhuman nasal secretions following nasal challenge withmcthacholine. histatnine and allergen. Using itninunohis-tochemical techniques, the localization of this enzyme hasbeen shown to correspond to the distribution of brady-kinin receptors in the nasal mucosa [38]. Nasal lavagescontain low levels of angiotensin converting enzyme [39].
The late phase reaction following nasal allergen chal-lenge has also been shown to be associated witb thegeneration of bradykinin and kallidin [32]. Although themechanism of kinin generation during late phase re-sponses has not been investigated, several differenceshave been observed from the early responses. Analysis ofthe profile of infiammatory mediators show that whilehistamine is present in both responses, PGD^ is onlydetectable during early response [29.30,32]. Since thebasophils secrete histaminc but not PGD: [40], this hasbeen interpreted as the basophil playing a role in the latephase response and the kinin generation at this juncturemay be related to basophil degranulation or tbe partici-pation of other cell types which demonstrate a delayedaccumulation in the nasal mucosa following allergenchallenge [4!]. Although kinin levels are found to beelevated during late response, they are significantly lowerwhen compared to early response but the levels of bothbistamine and TAME esterase activity are similar to thosemeasured during the early and late phase nasal allergenresponses [30.32]. Kinins generation has also beendemonstrated following nasal challenge with cold, dry air[42]. In this instance the release of kinins is associated withthe reporting of symptoms and the release of othermediators [42] similar to that seen during immediate nasalallergic response. In contrast the lack of any increase inhistamine during symptomatic rhinovirus-induced rhini-tis strongly suggest tbat. in contrast to allergic rhinitis,mast cells and basophils play little, if any. role in thepathogenesis of this type of rhinitis [27].
Action of kinins in the nose
The second criterion which must be met to demonstrate arole for kinins in rhinitis is that exogenous kinins shouldreproduce at least some of the symptoms of the clinicalcondition. Many laboratories have now shown that nasalbradykinin challenge in humans induces many of thesymptoms of rhinitis, including nasal congestion, rhinorr-hoea. sore throat and increase plasma protein leakage[22,43-47]. These studies have clearly shown that brady-kinin induces a dose-dependent increase in nasal symp-toms of rhinitis (Fig. 2) and an increase in plasma vascularleakage in normal and atopic subjects (Fig. 3) [43,44].These nasal challenge experiments with bradykinin haveproduced consistent results from several different labora-
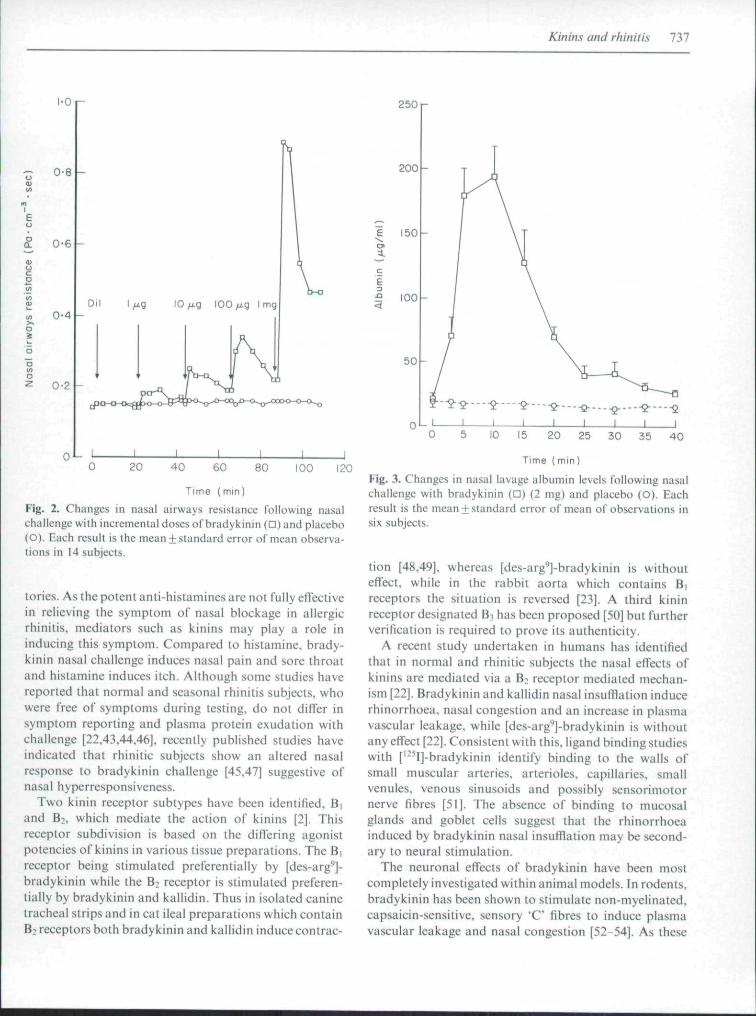
Kinins and rhinitis
1-0 I-
0-8
0-6
0-4
0-2
0 * -0 20 40 60 80 100 120
Time (mm)
Fig. 2. Changes in nasal airways resistance following nasalL-hallenge with incremental doses of bradykinin (n) and placebo(O). Each result is the mean + standard error of mean observa-tions in 14 subjects.
tories. As the potent anti-histamines are not fully effectivein relieving the symptom of nasal blockage in allergicrhinitis, mediators such as kinins may play a role ininducing this symptom. Compared to histamine, brady-kinin nasal challenge induces nasal pain and sore throatand histamine induces itch. Although some studies havereported that normal and seasonal rhinitis subjects, whowere free of symptoms during testing, do not differ insymptom reporting and plasma protein exudation withchallenge [22.43.44,46], recently published studies haveindicated that rhinitic subjects show an altered nasalresponse to bradykinin challenge [45,47] suggestive ofnasal hyperresponsiveness.
Two kinin receptor subtypes have been identified, B(and B:, which mediate the action of kinins [2]. Thisreceptor subdivision is based on the differing agonistpotencies of kinins in various tissue preparations. The B|receptor being stimulated preferentially by [des-arg^]-bradykinin while the B̂ receptor is stimulated preferen-tially by bradykinin and kallidin. Thus in isolated caninetracheal strips and in cat ileal preparations which containB: receptors both bradykinin and kallidin induce contrac-
2501-
200
150
:9 100
50
Time (min)
Fig. 3. Changes in nasal lavage albumin levels following nasalchallenge with bradykinin (D) (2 mg) and placebo (O). Eachresult is the mean + standard error of mean of observations insix subjects.
tion [48,49]. whereas [des-arg^]-bradykinin is withouteffect, while in the rabbit aorta which contains B|receptors the situation is reversed [23]. A third kininreceptor designated B;, has been proposed [50] but furtherverification is required to prove its authenticity.
A recent study undertaken in humans has identifiedthat in normal and rhinitic subjects the nasal effects ofkinins are mediated via a B; receptor mediated mechan-ism [22]. Bradykinin and kallidin nasal insufflation inducerhinorrhoea. nasal congestion and an increase in plasmavascular leakage, while [des-arg'']-bradykinin is withoutany effect [22]. Consistent with this, ligand binding studieswith ['-^l]-bradykinin identify binding to the walls ofsmall muscular arteries, arterioles, capillaries, smallvenules, venous sinusoids and possibly sensorimotornerve fibres [51]. The absence of binding to mucosalglands and goblet cells suggest that the rhinorrhoeainduced by bradykinin nasal insufflation may be second-ary to neural stimulation.
The neuronal effects of bradykinin have been mostcompletely investigated within animal models. In rodents,bradykinin has been shown to stimulate non-myclinated,capsaicin-sensitive. sensory 'C" fibres to induce plasmavascular leakage and nasal congestion [52-54]. As these

738 K. Rajakulasingam et al.
effects diminish after repeated capsaicin challenge, indica-tive of tachyphylaxis. it has been suggested that theseeffects of bradykinin are mediated by local axonalreflexes, producing neuropeptide release [55]. There are.however, difficulties in translating these findings to thehuman airways as a comparative study of the nasal effectsof bradykinin and capsaicin, although confirming thepreviously identified nasal effects of bradykinin failed toidentify any influence of capsaicin on nasal blockage orplasma vascular leakage [56]. Several alternative indirectmechanisms have, however, also been suggested fromanimal experiments. Firstly that bradykinin stimulationof vascular or other structures may lead to prostaglandinrelease, which could in turn stimulate glandular secretion[57]. Consistent with this is the observation in rodents thatibuprofen, a cyclo-o.xygenase inhibitor, reduces the secre-tory response to bradykinin suggestive of an intermedia-tory production of prostanoids [57]. Secondly bradykininhas been shown to be a secretogogue for rodent mast cells[58] and any nasa! secretory response to kinins could thusbe secondary to mast cell degranulation. However, it hasnot been possible to substantiate either of these mechan-isms in humans as bradykinin nasal challenge is notassociated with increased recovery of histamine [43,45]and neither H|-antihistamines nor cyclo-oxygenase in-hibitors have been shown to influence either bradykinin-induced weal response in human skin [59] or bradykinin-induced bronchoconstriction [60,61], The nasal vasculareffects of bradykinin (nasal blockage + permeabilitychanges) are thus likely to be a direct effect of kinin onvascular B; receptors. These vascular effects are likely tobe distinct, the blockage relating to engorgement ofvenous sinusoids while the increased recovery of albuminfollowing challenge results from increased fenestrationand vasodilation in the superficial mucosal vasculature.
Comparative relevance of kinins
There have been few studies directly comparing the nasaleffects of kinins with other mediators identified in associa-tion with rhinitis. In rhinovirus induced exacerbationskinins are the only mediator so far identified in nasallavage [27], suggestive of its potential importance ininfective rhinitis, while co-release of histamine, prosta-glundin D2, leukotriene C4 and D4 is identified in allergicrhinitis [29,31,33], One comparative study investigatingthe nasal effects of bradykinin. histamine. PGD2 andPGF2;, identified that subjeclively both histamine andPGD2 have a greater effect on nasal blockage [44].However, an objective study cotnparing histamine andbradykinin on a molar basis, rather than a mg/ml basis,produced a reverse order of potency with objectivemonitoring of nasal airways resistance and plasma pro-
tein leakage [62]. Of relevance to these considerations isalso the relative proportions of histamine, kinins andeicosanoids released during allergic exacerbations ofrhinitis. Measurement of nasal lavage level in allergenindticed rhinitis has identified similar release of kinins andhistamine with respective mean maximum levels of up to 8ng/ml and 5 ng/ml [31]. These studies therefore suggestthat histamine and kinins may be comparable in theirnasal effects but the future development of selective andspecific kinin receptor antagonists is required beforerelevant contribution of kinins to different forms ofrhinitis can be fully determined.
Conclusions
Kinin generation occurs during rhinitis as evidenced byits recovery in association with allergen and viral-inoculation induced rhinitis and the elevated levels innaturally occurring seasonal allergic rhinitis and rhino-viral upper respiratory tract infections. Investigativestudies have further identified the potential relevance ofkinins to rhinitis as nasal insufflation with the kininsbradykinin and kallidin but not the metabolite [des-arg*^]-bradykinin induce nasal blockage, rhinorrhoea and nasalpain along with an increase in plasma protein recovery inlavage suggestive of enhanced vascular permeability.These studies identify that these kinin effects are Bireceptor mediated. The future availability of a specificand potent Bi-receptor antagonist will permit the identifi-cation of the relative contribution of kinins (o thepathogenesis of rhinitis in its different forms.
Acknowledgment
This work was supported by a grant from the FerringPeptide Research Partnership, Malmo, Sweden.
References
1 Howarth PH. Allergic rhinitis; a rational choice of treat-tncnt. Rcsp Med 1989: 83:179-88.
2 Regoli D, Barabe J. Pharmacology of bradykinin andrelated peptides. Pharmacol Rev 1980: 32:1 46.
3 Bhoola KD, Collier HOJ, Schachter M, Shorlcy PG.Actions ofsome peptides on bronchial muscle. Br J Pharma-col 1962; 19:190-7.
4 RatnofTOD, Rosenblum JM. Role ofHageman factor in theinitiation of clotting by glass: evidence that glass freesHageman factor from inhibition. Am J Med 1958; 25:160-8.
5 Weiss AS, Gallin Jl, Kaplan AP. Fletcher factor deficiency.Abnormalities of coagulation, fibrinolysis, chemotacticactivity and kinin generation attributable to absence ofprekallikrein. J Clin Invest 1974; 53:622-33.
6 Colman RW. Bagdasarian A. Talamo RC. Scott CF. Seavcy

Kinins and rhinitis 739
M, Guimaraes JA et cd. William trait: human kininogendeficiency wilh diminished levels of plasminogen activatorand prekallikrein associated with abnormalities of theHageman factor dependent pathways. J Clin Invest 1975;56:1650-62.
7 Cochrane CG, Revak SD. Wucpper KD. Activation ofHageman factor in solid and fluid phase. J Exp Med 1973;138:1564 83,
8 Kaplan AP, Austen KF, A prealbumin activator of prekal-likrein [I. Derivation of activators of prekallikrein fromactivated Hageman factor with plasmin. J Exp Med 1971;138:696-712.
9 Nustad K, Orstavik TB, Gautvik KM. Pierce JV. Glandularkallikreins. Gen Pharmacol 1978; 9:1-9.
10 Pinkus GS, Maier M. Seldin DC, Ole-Moiyoi O. Austen KF,Spragg J. Immunohistochemical localization of glandularkallikrein in the endocrine and exocrine human pancreas. JHistochcm Cytochem 1983; 31:1279-88.
11 Lazure C. Leduc R. Seidah NG. Chretien M. Dube J.Chapdelaine P et al. The major androgen-dependant pro-tease in dog prostate belongs lo the kallikrein family:confirmation by partial amino acid sequencing. FEBS Lett1984; 175:1-7.
12 Figueroa CD. Maclver AG, Mackenzie JC, Bhoola KD.Localization of immunoreactive kininogen and ti.ssue kaiUk-rein in the human nephron. Histochemistry 1988: 89:437-42.
13 Geiger R. Kellermann J, Duetzmann R, Lottspeich F.Biochemistry of human tissue (urinary) kallikrein. In:Vasopressor hormones. Basel: BirkhauserVerlag, 1987:223-37.
14 Powers CA. Nasjlelti A. A novel kinin-generating protease(kininogenase) in the porcine anterior pituitary. J Bio Chem1982; 257:5594-600.
15 Christiansen SC. Proud D, Cochrane CG. Detection oftissue kallikrein in the bronchoalveolar lavage fluids ofasthmatic subjects. J Clin invest 1987; 79:188-97.
16 Pierce JV. Guimaraes JA. Further characterization of highlypurified human plasma kininogens. In: Pisano JJ. AustenKF, eds. Chemistry and biology of the kallikrein-kininsystem in health and disease. DHEW Publ, No. (NIH)76-791, Washington DC. 1976:121-7,
17 Proud D, Kaplan AP, Kinin formation: mechanisms androle in inflammatory disorders, Ann Rev Immunol 1988;6:49-83.
IX Meier HL. Flowers B. Silverberg M. Kaplan AP. MacGlas-han DW Jr, Shulman ES, Newball HH. Localization andcharacterization of lung Hageman factor activator. FedProc 1982; 41:377,
19 Proud D. MacGlashan DW Jr. Newball HH, Shulman ES,Lichtenstein. LM. Immunoglobulin E-mediated release of akininogenase from purified human lung mast cells. Am RevDis 1985; 132:405-8.
20 Newball HH. Talamo RC. Lichtenstein LM. Anaphylacticrelease of a basophil kalUkrein-like activity, I. Purificationand characterization. J Clin Invest 1979; 64:457-65.
21 Sheikh lA. Kaplan AP, Studies of the digestion of brady-kinin, lysyl bradykinin, and kinin degradation products by
carboxypeplidase. A, B and N. Biochem Pharmacol 1986;35:1957-63,
22 Rajakulasingam K, Polosa R. Holgate ST, Howarth PH.Comparative nasal effects of bradykinin, [dcs-arg**!-brady-kinin in atopic rhinitic and normal volunteers. J Physiol1991; 437:577-87.
23 Regoli D, Maraceu F, Barabe J. De novo formation ofvascular receptors for bradykinin. Can J Physiol Pharmacol1978; 56:674-7.
24 Sheikh lA, Kaplan AP. Studies of the digestion of brady-kinin. lysyl-bradykinin and [des-arg'']-bradykinin by angio-tensin converting enzyme. Bio Chem Pharmacol 1986;35:1951-6.
25 Choi HSH, Lesser M. Cardozo C, Orlowski M. Immunocy-tochemical localization of endopeptidase 24.15 in rat lungs.Am Rev Respir Dis 1990: i4l:A727.
26 Svensson C. Andersson M. Persson CGA. Venge P, AlknerU. Pipkorn U. Albumin, bradykinin and eosinophil cationicprotein on the nasal mucosal surface in patients with hay-fever during natural allergen exposure, J Allergy ClinImmunol 1990; 85:828-33.
27 Proud D. Naclerio RM. Gwaltney JM. Hendley JO, Kininsare generated in nasal secretions during natural rhinoviruscolds. J Infect Dis 1990; 161:120-3,
28 Naclerio RM, Proud D, Lichtenstein LM, Sobotka AK.Hendley JO. Sorrentino J. Gwaltney JM. Kinins are genera-ted during experimental rhinovirus colds. J Infect Dis 1988;157:133-42.
29 Naelerio RM. Meier HL. Sobotka AK, Adkinson NF Jr,Meyers DA, Norman PS ei al. Mediator release after nasalairway challenge with allergen. Am Rev Respir Dis 1983;128:597-602,
30 Naclerio RM. Proud D. Peters SP. Silber G. Sobotka AK.Adkinson NF et at. Inflammatory mediators in nasalsecretions during indueed rhinitis. Clin Allergy 1986;16:101-10.
31 Proud D, Togias A. Naclerio RM, Crush SA. Norman PS.Lichtenstein LM. Kinins are generated in vivo followingnasal airway challenge of allergic individuals with allergen. JClin invest 1983; 72:1678-85.
32 Naclerio RM. Proud D. Togias AG, Adkinson NF Jr.Meyers DA. Sobotka AK et al. Inflammatory mediators inlate antigen-induced rhinitis. N EnglJ Med 1985;3I3:65-7O.
33 Creticos PS. Peters SP, Adkinson NF Jr, Naclerio RM.Hayes EC. Norman PS et at, Peptide leukotriene releaseafter antigen challenge in patients sensitive to ragweed,N Engl J Med 1984; 310:1626-30,
34 Baumgarten CR, Nicols RC. Naclerio RM, LichtensteinLM. Norman PS, Proud D, Plasma kallikrein duringexperimentally-induced allergic rhinitis: role in kinin forma-tion and contribution to TAME-esterase activity in nasalsecretions. J Immunol 1986; 137:977-82.
35 Baumgarten CR, Nichols RC. Naclerio RM. Proud D,Concentrations of glandular kallikrein in human nasalsecretions increase during experimentally-induced allergicrhinitis. J Immunol l986;l37:1323-8,
36 Baumgarten CR, Togias AG, Naclerio RM, Lichtenstein

740 K. Rajakulasingam et al.
LM, Norman PS. Proud D. Influx of kininogcns into nasalsecretions after antigen challenge of allergic individuals. JClin invest 1985; 76:191-7.
37 Proud D. Baumgarlen CR. Naclerio RM, Ward PE. Kininmetabolism in nasal secretions during experimentally-induced allergic rhinitis. J Immunol 1987; 138:428-34.
38 Ohkubo K. Kaulbach H. Mcrida M, Hausfeld. Okada H,Erdos EG et al. Carboxypeptidase N (CPN) in the humannasal mucosa and secretions. Am Rev Respir Dis 1991;143:A616.
39 Proud D. Baumgarten CR, Naclerio RM. Ward PE. Kininmetabolism in nasal secretions during experimentally-induced allergic rhinitis. J Immunol 1987; 138:428-34.
40 MacGlashan DW Jr, Sehleimer RP, Peters SP, SchulmanES. Adams GK III, Sobotka AK ei al. Comparative studiesofhunian basophilsand mast cells. Fed Proc 1983: 42:2504-9.
41 Bascom R. Pipkorn U. Gleich GJ. Lichtenstein LM,Naclerio RM. Effect of systemic steroids on eosinophils andmajor basic protein during nasal antigen challenge. J AllergyClin Immunol 1986; 77:246.
42 Togias AG. Naclerio RM. Proud D. Fish JE. Adkinson NFJr. Sobotka AK et al. Nasal challenge with cold, dry airresults in release of inflammatory mediators. Possible mastcell involvement. J Clin Invest 1985; 76:1374-81.
43 Proud D. Reynolds CJ. LaCapra S. Sobotka AK. Lichten-stein LM, Naclerio RM. Nasal provocation with bradykinininduces symptoms of rhinitis and a sore throat. Am RevRespir Dis 1988; 137:613-16.
44 Doyle WJ. Boehm BS. Skoner DP. Physiologic responses lointranasal dose-response challenges with histamine. metha-eholine, bradykinin and prostaglandin in adult volunteerswith and without nasal allergy. J Allergy Clin Immunol1990; 86:924-35.
45 Brunnee T, Nigam S, Kunke! G, Baumgarten CR. Nasalchallenge studies with bradykinin: influence upon mediatorgeneration. Clin Exp Allergy 1991; 21:425-31.
46 Holmberg K, Blake B, Pipkorn U. Vaseular effects oftopically applied bradykinin on the human nasal mueosa.Eur J Pharmacol 1990; 175:35-41.
47 Baraniuk JN. Brayton P, Kaliner M. Barnes PJ. Perennialrhinitic subjects have altered vascular, glandular and neuralresponses to bradykinin. Am Rev Dis 1991; 143;A617.
48 Rangachari PK, McWade D. Donoff J. Luminal receptorsfor bradykinin on the canine traeheal epithelium: functionalsubtyping. Reg Peptides 1988; 21:237-44.
49 DrouinJN.St-PierreSA. RegoliD. Receptor for bradykininand kallidin. Can J Physiol Pharmacol 1979; 57:357-9.
50 Farmer SG, Burch RM. Meeker SA, Wilkins DE. Evidencefor a pulmonary Bi bradykinin receptor. Mol Pharmacol1989; 36:1-8.
5! Baraniuk J, Lundgren JD, Mizoguchi H. Peden D. Gawin A.Merida M ei al. Bradykinin and respiratory mucous mem-branes. Am Rev Dis 1990; 141:706-14.
52 Kaufman MP. Coleridge HM, Coleridge JCG. Baker DG.Bradykinin stimulates afferent vagal C-fibres in intrapul-monary airways in dogs. J Appl Physiol 1980; 48:511-7.
53 Davis B, Roberts AM, Coleridge HM. Coleridge JCG.Reflex tracheal gland secretion evoked by stimulation ofbroEichial C-fibrcs in dogs. J Appl Physiol 1982; 53:985-91.
54 Lundblad L. Saria A, Lundbcrg JM, Anggard A. Increasedvaseular permeability in rat nasal mucosa induced bysubstance P and stimulation of capsaicin-sensitive trigemi-nal neurons. Acta Otolaryngologica 1983; 96;479-84.
55 Lundblad L. Brodin E. LundbergJM. Anggard A. Effects ofnasal capsaicin pretreatment and cryosurgery on sneezingreflexes, neurogenie plasma extravasation, sensory andsympathetic neurons. Acta Otolaryngologica (Stockh) 1985;100:117-27.
56 Rajakulasingam K. Polosa R. Lau LCK. Holgate ST.Howarth PH. The nasal eflfects of bradykinin and capsaicin:role of ' C sensory nerve fibres. J Appl Physiol 1992;72:1418-24.
57 Leikauf GD, Ueki IF, Nadel JA. Widdicombe JH. Brady-kinin stimulates Cl secretion and prostaglandin E: release bycanine tracheal epithelium. Am J Physio! 1985; 248:F48-F55.
58 Ishizaka T. Iwata M. Ishizaka K. Release of histamine andaraehidonates from mouse mast cells induced by glycosyla-tion enhancing factor and bradykinin. J Immunol 1985;134:1880-7.
59 Crossman D, Fuller RW. Bradykinin induced wheal andflare is not mediated hy histamine release or cyclooxygenaseproducts. Br J Pharmacol 1988; 26:113-15.
60 Fuller RW, Dixon CMS. Cuss FMC. Barnes PJ. Brady-kinin-indueed bronchoconstriction in man: mode of action.Am Rev Res Dis 1987; 135:176 81.
61 Polosa R. Phillips G, Lai CKW. Holgate ST. Contributionof histamine and prostanoids to bronchoconstriction pro-voked by inhaled bradykinin in atopic asthma. Allergy 1990;45:174-82.
62 Rajakulasingam K, Polosa R, Lau LCK. Church MK.Holgate ST. Howarth PH. Comparative nasal eflects ofbradykinin and histamine: influence on nasal airways resis-tance and plasma vascular leakage. Thorax 1992; 47:232.

Copyright © 2022 FDOKUMEN




















