
Acid azo dye degradation by free and immobilized horseradish peroxidase (HRP) catalyzed process

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Journal of Membrane Science 339 (2009) 215–223
Contents lists available at ScienceDirect
Journal of Membrane Science
journa l homepage: www.e lsev ier .com/ locate /memsci
Kinetic study of a biocatalytic membrane reactor containing immobilized�-glucosidase for the hydrolysis of oleuropein
R. Mazzei a,b, L. Giorno a,∗, E. Piacentini a,c, S. Mazzuca b, E. Drioli a,c
a Institute on Membrane Technology, CNR-ITM, C/o University of Calabria, Via P. Bucci 17/C, 87036 Rende (CS), Italyb Department of Ecology, University of Calabria, Via P. Bucci 6/B, 87036 Rende (CS), Italyc Department of Chemical Engineering and Materials, University of Calabria, Via P. Bucci 42/A, 87030, Rende (CS), Italy
a r t i c l e i n f o
Article history:Received 30 December 2008Received in revised form 23 March 2009Accepted 25 April 2009Available online 3 May 2009
Keywords:Biocatalytic membrane reactorCapillary membraneImmobilized enzymeOleuropeinKinetics�-Glucosidase
a b s t r a c t
Kinetic parameters of �-glucosidase from almond immobilized in a continuous membrane reactor havebeen studied and compared with free enzyme used in a stirred tank reactor. The enzyme has been immo-bilized by cross-flow ultrafiltration in asymmetric capillary membranes made of polysulphone havingnominal molecular weight cut-off of 30 kDa. The hydrolysis of oleuropein into aglycon and glucose hasbeen used as a reaction system to demonstrate the efficiency of biocatalytic membrane reactors usingimmobilized �-glucosidase. The membrane allowed to support the enzyme in a defined microenviron-ment, the microporous matrix, as well as to control substrate feeding to and product removal from thereaction environment in a single unit operation.
Results showed that kinetic parameters of immobilized enzyme are the same as for the free enzyme,meaning that the enzyme immobilized within the membrane did not undergo decrease of activity whilemaintaining higher stability compared to the free enzyme. This confirms that the common observationof inverse relationship between activity decrease and stability increase for immobilized enzymes is nota general rule. In particular, the catalytic activity of immobilized enzyme can be optimized by the use ofhigh throughput microstructured membrane systems.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Phenolic compounds are a complex but important group of nat-urally occurring compounds in plant. Although the main phenoliccompounds in olive fruit are secoiridoid derivates, leaves also con-tain phenyl acids, phenyl alcohols, and flavonoids. The increasinginterest towards these compounds is due to the fact that they havepharmacological properties, are natural antioxidants, and inhibitGram positive micro-organisms involved in fermentation of olivefruit [1].
Oleuropein is a heterosidic ester of elenoic acid and 3,4-dihydroxy-phenylethanol (hydroxytytrosol) [2] that producesbitterness in unripe olives. This is the major phenolic compoundin the olive fruit and leaves, and it is completely absent from oliveoil, because of its high water solubility [3]. Relevant hypotensivedose-dependent effects and protection against induced arrhyth-mias, shown in rats, by various extract of Olea europaea werecorrelated with oleuropein content in bud and leaf preparation. It isalso the high concentration of oleuropein and its hydrolysis prod-ucts that confers to Olea europaea part to its resistance to microbial
∗ Corresponding author. Tel.: +39 0984 492050/2134; fax: +39 0984 402103.E-mail addresses: [email protected], [email protected] (L. Giorno).
attack [4]. Other studies demonstrated that the hydrolysis prod-ucts of oleuropein, thanks to �-glucosidase action, have a muchmore marked inhibitory effect than oleuropein. In particular, theaglycon, produced by oleuropein hydrolysis, is well known as phar-macologically active molecule. In fact, it is described for its potentialapplication as an antimicrobial agent in some fairly common dis-eases of olive trees [5] and also for its strong active antioxidantproperty [6–10].
�-Glucosidases, the enzymes responsible for the oleuropeinhydrolysis, belong to the group of hydrolases widely existing in var-ious sources, such as fungi, bacteria, plant and animal tissue [11].Almost all these kind of enzymes in plant showed best pH stabilityvalues between 5 and 7 [12–14].
The broad optimum pH and temperature range makes thisenzyme particularly useful for application in biotechnology pro-cesses. The �-glucosidase from different source has been studied invarious systems, as free or attached to different supports [12,14–19].Enzymatic hydrolysis of oleuropein, in a NMR tube (a thin glasswalled tube used to contain samples in nuclear magnetic resonancespectroscopy) by a �-glucosidase at 25 ◦C giving two diastereoiso-meric aglycons was described [20].
The �-glucosidase activity was also tested on biological in vivosystems like Lactobacillus plantarum [21] and using recombinantenzymes from hyperthermophilic archeon.
0376-7388/$ – see front matter © 2009 Elsevier B.V. All rights reserved.doi:10.1016/j.memsci.2009.04.053

Author's personal copy
216 R. Mazzei et al. / Journal of Membrane Science 339 (2009) 215–223
The enzymatic hydrolysis of oleuropein, extracted from oliveplant, by free �-glucosidase from almond was studied by Capassoet al. [22].
Homogeneous recombinant �-glycosidase from hyperther-mophilic archaeon Sulfolobus solfactaricus expressed in E. coli(EcS�Gly), was immobilized on chitosan support and used to pro-duce hydroxytyrosol from commercially available oleuropein [1]. Inaddition the antioxidant properties of the main products obtainedwere purified and characterized [23] until a production of hydrox-ytyrosol of 91–94% in weight was achieved [24].
As recently described by Guiso and Marra [25] the first step ofoleuropein hydrolysis in the reaction catalyzed by �-glucosidasefrom almond, gives two products: glucose and aglycon.
The aglycon is interesting due to the strong antioxidant activity.Oleuropein is abundant in nature, easily available and consideredas a waste material [22]. The production of aglycon from oleuropeinhas glucose as co-product. The development of bioprocesses basedon oleuropein–�-glucosidase system for the production of aglyconand glucose represents a sustainable production strategy able tovalorise wastes while approaching to zero discharge.
In line with this strategy, biocatalytic membrane reactors areable to integrate bioconversions with selective membrane separa-tions leading to continuous clean, safe and low energy consumptionproduction systems [26–33]. Enzyme immobilization in biocat-alytic membrane reactors for production of high added valuecomponents in pharmaceutical, food and biotechnology has beenwidely studied. Examples of systems developed at industrial pro-duction level have been recently reviewed [28]. Although theirpotentialities have not been fulfilled yet, the current needs and chal-lenges in satisfying the increasing consumer demand of safe goodsand the limited resources availability will force towards selectiveand efficient technologies such as biocatalytic membrane reactors.
In this work, the development of a continuous reactor systembased on �-glucosidase-loaded membrane for the conversion ofcommercial oleuropein into glucose and aglycon was studied. Inparticular, the work aimed at studying the kinetic properties of �-glucosidase immobilized in a continuous membrane reactor and itscomparison with free enzyme used in a conventional batch reactor.�-Glucosidase from almond was used as enzymatic model systemto prove the viability of the proposed continuous reaction processfor oleuropein hydrolysis compounds production.
2. Materials and methods
The reaction equation of the oleuropein (1) hydrolysis to agly-con (2) and glucose (3) catalyzed by �-glucosidase from almond isillustrated in Fig. 1.
The �-glucosidase from almond (chromatographically purifiedlyophilized powder, Cod G4511, one unit will liberate 1.0 �moleof glucose from salicin per min at pH 5.0 at 37 ◦C) and glucosewere obtained from Sigma–Aldrich, oleuropein from Extrasyn-these (France). Acetic acid (Carlo Erba) and sodium acetate(Sigma–Aldrich) were used to prepare 50 mM acetate buffer pH6.5. Acetonitrile and o-phosphoric acid for HPLC mobile phasepreparation were purchased from Carlo Erba and Sigma–Aldrich,respectively.
2.1. Analytical methods
Protein concentration was measured by Bicinchoninic Acid Kit(BCA) (Sigma–Aldrich).
A method adapted from Ranalli et al. [34] was used to measureoleuropein by HPLC. A reverse silica LiChoCART Superspher RP8 col-umn, 250–4 mm, 5 �m (Merck) was used. The mobile phase was amixture of acetonitrile/water (21:79) acidified with o-phosphoric
acid (up to pH 3). The flow rate and pressure were 0.8 ml/min and210 (±4) bar, respectively. The volume of sample injection was20 �l. The oleuropein was detected at 280 nm wavelength.
Glucose was measured using a reagent kit (glucose HK assay kit,Sigma–Aldrich) by means of a spectrophotometric method basedon the absorbance of NADH at 340 nm.
Aglycon was calculated as equivalent moles of glucose on thebasis of the reaction stoichiometry. For precise thorough kineticmeasurement, it is mandatory to instantaneously stop the reactionin the collected samples.
For reactions carried out in the biocatalytic membrane reactor,the enzyme is immobilized in the membrane and samples are col-lected as permeate solution, therefore enzyme is not present inthese samples, the reaction cannot occur anymore and they do notrequire any further treatment for enzyme deactivation and separa-tion from other components.
Samples withdrawn from reactions carried out with free enzymeneed fast deactivation of the enzyme, in order to stop the reaction,and its separation from the other components, in order to avoidinterferences with HPLC and spectrophotometric analyses.
Among the various chemical physical methods to deactivateenzyme, the thermal deactivation was identified as the most suit-able method due to the possibility to avoid the use of chemicalsthat may cause interference with HPLC analyses. The sample pre-treatment method was optimized by identifying the temperatureand minimum incubation time necessary to deactivate the enzyme.Enzyme solution samples were immersed in a bath at 95 ◦C for 30,60, 120, 180 s and after that the enzyme activity was measured.After the denaturizing step, the enzyme was separated from theother compounds present in the sample solution by ultrafiltration.Polysulphone (PS) (Romicon, Germany), polyethersulphone (PES)(PBI international) and regenerated cellulose (PBI international) of30 kDa molecular weight cut-off (MWCO) were used. The most suit-able membrane was selected on the basis of its inertness towardsthe other reaction components commercially available and testedto monitor the reaction rate, i.e. oleuropein and glucose (aglycon isnot commercially available yet). Therefore, to verify that no adsorp-tion of these molecules on polymeric membranes was occurringduring sample treatment, ultrafiltration tests with standard solu-tions through the different polymers were carried out. Adsorptionwas calculated by mass balance between initial and permeate solu-tions.
2.2. Equipments
Hydrolysis reaction catalyzed by free �-glucosidase was carriedout in a stirred tank reactor (STR), which is characterized by timevariant conditions (Fig. 2a). The reaction catalyzed by immobilizedenzyme was performed in a continuous biocatalytic membranereactor (BMR), which is characterized by time invariant conditionsat the steady state (Fig. 2b).
2.2.1. Stirred tank reactor setupThe reaction took place in a tank with 50 ml total volume of reac-
tion mixture, 45 ml of substrate solution plus 5 ml (0.035 mg/ml).The enzyme solution was prepared dissolving 3.5 mg in 10 mlof buffer solution at pH 6.5, to obtain a final concentration of0.35 mg/ml. 5 ml of this enzyme solution, were then added to45 ml of oleuropein solution to obtain a final concentration of0.035 mg/ml. The concentration obtained was also confirmed byBCA test. The reaction tank was kept at 25 ◦C in a thermostatic bath.Complete mixing was achieved by using a magnetic stirrer set at50 rpm. Samples were taken as function of time to measure theamount of oleuropein converted and corresponding glucose pro-duced.

Author's personal copy
R. Mazzei et al. / Journal of Membrane Science 339 (2009) 215–223 217
Fig. 1. Scheme of the first step of the reaction catalyzed from �-glucosidase from almond having oleuropein as substrate.
2.2.2. Biocatalytic membrane reactor setupThe equipment used to carry out the hydrolysis reaction is illus-
trated in Fig. 2b. It consists of a membrane module made of a pyrexcylinder as shell containing four polysulphone capillary mem-branes of 30 kDa nominal molecular weight cut-off (NMWCO) (PS30 kDa) and internal/external diameter of 1.08/1.75 mm. The mem-branes have asymmetric structure with the selective layer on thelumen side and the sponge layer on the shell side. The membranemodule effective length was 10.5 (±0.5) cm and the inter-nal/external membrane surface area was 1.46 × 10−3/2.36 × 10−3
(±4 × 10−4)m2. The membrane void volume was 0.61 (±0.02)cm3.This represents the membrane reactor volume. The membrane void
volume was calculated as difference between the volume of themembrane cylinder (0.69 cm3) (which includes polymer and voidvolume) and the volume occupied by the polymer (0.08 cm3). Thevolume of the membrane cylinder was calculated on the basis ofthe fiber internal/external diameter and length. The volume of thepolymer was calculated from the polymer density (1.24 g/ml) andthe mass of the dried fibers.
The system is constituted also by a tank containing the oleu-ropein solution, a peristaltic pump (P) to feed the reagent solutionto the biocatalytic membrane reactor, pressure gauges (PG) to mea-sure inlet and outlet pressure and a graduated cylinder to collectpermeate solution containing reaction product.
Fig. 2. Experimental reactors setup. (a) Stirred tank reactor and corresponding concentration profile of substrate and product. (b) Experimental apparatus used to carry outreaction in the continuous biocatalytic membrane reactor and corresponding concentration profile of substrate and product. (P: peristaltic pump, PG: pressure gauges).

Author's personal copy
218 R. Mazzei et al. / Journal of Membrane Science 339 (2009) 215–223
2.3. Enzyme immobilization by cross-flow ultrafiltration
After module preparation, the pure water permeability of virginnative membranes was measured from Darcy’s equation
J = Lp �P (1)
as the linear coefficient (Lp) between flux and applied transmem-brane pressure as driving force.
This was done in order to state initial membrane performance.The �-glucosidase was entrapped within the membrane asymmet-ric sponge layer by cross-flow ultrafiltration from shell to lumen[26]. In fact, the 65 kDa MW enzyme macromolecules could enterthe macrovoids of the sponge layer but could not pass through the30 kDa cut-off thin layer. During the immobilization process, per-meate flux was measured as a function of time and when foulingdue to enzyme entrapment reached steady state the process wasstopped. The membrane system was rinsed to remove all reversiblyadsorbed protein and pure water permeability was measured againto evaluate hydraulic resistance promoted by immobilized enzyme.The membrane was rinsed with buffer using a pressure of 0.1 barand an axial velocity of 1.03 m/s.The overall characteristics of fluxreduction during ultrafiltration can be described by [35]:
J = TMP�R
(2)
where J (l h−1 m−2) is the flux, TMP is the transmembrane pressure(bar), � is the viscosity (Pa s) and R (m−1) is the hydraulic resistance.From Eqs. (1) and (2) the hydraulic resistance is
R = 1�Lp
(3)
The total hydraulic resistance is also given by:
RTOT = Rm + Rrev + Rirr (4)
where Rm is the resistance due to the membrane, calculated con-sidering the pure water permeability of virgin membranes; Rrev isthe resistance due to reversible fouling; Rirr is the resistance due toirreversible fouling calculated considering the pure water perme-ability after washing steps. The sum between Rrev and Rirr gives theresistance due to the overall fouling Rf.
The amount of immobilized enzyme was calculated by mass bal-ance and its contribution to hydraulic resistance was evaluated byEqs. (3) and (4) as Rirr.
2.4. Reaction operation mode in the biocatalytic membranereactor
After the enzyme was immobilized within the membrane andthe system rinsed to remove the reversible fouling, the shell cir-cuit was recirculated with oleuropein solution, in buffer at pH 6.5,which permeated through the membrane thickness (where theenzyme was loaded) (Fig. 2b) with an applied transmembrane pres-sure of 0.1 bar, an axial velocity of 0.35 m/s, and a permeate flowrate of 0.30 (±0.05)ml/min. A thermostatic bath was used to keepthe temperature constant at 25 ◦C. As oleuropein (1) (540.23 Da)passed through the enzyme-loaded membrane it was convertedinto the corresponding reaction products, aglycon (2) and glucose(3).
In order to confirm that changes in substrate and productconcentration were only due to the biocatalyst action and nointerference with the membrane materials was occurring, blankexperiments were carried out, where no enzyme or deactivatedenzyme was present in the membrane.
2.5. Evaluation of reaction rate, kinetic parameters, and enzymestability
The initial reaction rate (V0) for stirred tank reactor and bio-catalytic membrane reactor obtained as a function of differentsubstrate concentrations [S] were used to fit Michaelis–Mentenequation thus obtaining estimates of Km and Vmax. More accu-rate graphical evaluation of Km and Vmax was obtained by theLineweaver–Burk equation [36]:
1V0
= Km
Vmax· 1
S+ 1
Vmax(5)
The value of the kinetic constant k+2 was also evaluated usingthe following equation:
Vmax = k+2[E]tot (6)
where [E]tot is the total concentration of enzyme solution used inthe reaction.
Equations for calculation of initial reaction rate (V0) in the usedreactor systems were derived from balance equation:
IN − OUT + PROD = ACC (7)
(FC)IN − (FC)OUT + (vrV) = dVC
dt(8)
where F is the flow rate (volume/time), C is the concentration(mass/volume); vr is the volumetric reaction rate (mass/time vol-ume); V is the reaction volume; t is the time.
For the batch stirred tank reactor the flow term is zero and pro-duction is equal to accumulation:
vr = dC
dt(9)
Therefore, the reaction rate is calculated as time variant concen-tration (C) from the linear section of the curve C vs t (Fig. 2a).
For the biocatalytic membrane reactor, the reaction takes placeat the asymmetric porous membrane level, through which INand OUT flow terms occur mainly by convective flow. At steadystate, each membrane micropore works as a continuous stirredmicroreactor. Therefore, the overall membrane matrix forms a highthroughput reactor, whose volume is the membrane void volumewhich works as a continuous stirred tank reactor (CSTR).
For a steady state CSTR, the term ACC = 0:
(FC)IN − (FC)OUT + (vrV) = 0 (10)
The reaction rate is then derived as:
vr = F(Cf − Cp)V
(11)
In this equation vr is the reaction rate (mmol/cm3 min), F is thepermeate flow rate (cm3/min), V is the reactor volume (cm3). Thereactor volume is represented by the membrane void volume.
Another important parameter in membrane bioreactor is theresidence time as it influences the conversion degree. High perme-ate flow rate means low residence time of substrate solution withinthe pores. The residence time (�) was obtained from the followingequation:
� = V
F(12)
where V is the membrane reactor volume (cm3) and F is the volu-metric flow rate (cm3/s)
Enzyme stability is also a crucial parameter. Pre-screening testswere carried out to verify the stability of immobilized enzyme inorder to make sure that kinetic tests were not affected by stabilityproblems. Tests were performed as follows: the reaction was carriedout through the enzyme-loaded membrane for 2.5 h, using 2 mM

Author's personal copy
R. Mazzei et al. / Journal of Membrane Science 339 (2009) 215–223 219
oleuropein concentration. The system was then rinsed and a newreaction was carried out with a different substrate concentration.Afterwards the test with the initial substrate concentration was car-ried out to verify that the enzyme kept constant performance. Thereactor was preserved at −80 ◦C overnight. The subsequent daystests were repeated as mentioned before for a total of about 30 hoperation. The reactor was then preserved at −80 ◦C and after twomonths the activity was tested again.
3. Results and discussion
Initially, the sample pre-treatment procedure was standard-ized, then the kinetic performance of �-glucosidase from almondimmobilized in the UF-capillary membrane reactor was studied andcompared with the one of free enzyme. Results are reported in thefollowing sections.
3.1. Development of sample pre-treatment procedure
Experiments to set up the appropriate sample pre-treatmentprocedure for reaction rate evaluation as a function of time havebeen performed. In particular, the work focused on the identifica-tion of conditions for instantaneously stopping the reaction rateby deactivating the enzyme in samples withdrawn from stirredtank reactor and for separating the deactivated enzyme from otherreaction components (e.g. oleuropein, glucose and aglycon). Resultsshowed that 30 s of sample treatment at 95 ◦C were enough to dena-turize the enzyme and that the high temperature did not causeany interference on the analysis of oleuropein and glucose. In fact,analytical tests of standard samples before and after treatment athigh temperature, confirmed that concentration of both compo-nents remained unchanged. Aglycon was not tested as it is notcommercially available.
The denaturized enzyme was then separated from the samplesolution by ultrafiltration. Membranes with 30 kDa cut-off weresuitable to completely remove the enzyme. Tests were performedin order to select a membrane material inert towards the reactioncomponents, i.e. no adsorption of both substrate and product wasoccurring. Results are summarized in Table 1, which shows thevalues of oleuropein and glucose concentration before and afterfiltration through 30 kDa PES, PS and RC membrane. The amountof oleuropein after the filtration with PES membrane decreasedof about 9% respect to the initial solution, due to absorption phe-nomena of oleuropein to the polymer. The mass adsorbed wascalculated as mass balance between feed and permeate solution.Different behaviour was observed when the oleuropein was fil-tered through PS and RC membranes, where the concentrationof oleuropein remained unchanged between feed and permeatesolution. The glucose concentration remained the same after fil-tration through the three types of membranes. Therefore, bothPS and RC 30 kDa membranes were suitable for the sampletreatment. In subsequent experiments RC membranes have beenused.
The procedure to pre-treat samples withdrawn from STR wasthen identified as follows: (1) stopping the reaction by heating sam-ple at 95 ◦C under agitation for 30 s; (2) cooling down the solutionby immersing test tube in ice for few minutes; (3) separation ofdeactivated enzyme by ultrafiltration through RC membranes; (4)collect permeate solution for further HPLC and spectrophotometricanalyses.
It is worth noting that the removal of enzyme by ultrafiltra-tion, without first deactivating it by thermal treatment, did notresult a suitable method as it did not perform a fast and repro-ducible stopping time of the reaction. For samples withdrawn fromthe continuous biocatalytic membrane reactor, this procedure was
not necessary, as samples are collected as permeate from PS capil-lary membrane through which the reaction is occurring and whichsimultaneously support the enzyme and remove unconverted sub-strate and obtained products.
3.2. Reaction kinetics with ˇ-glucosidase free in the stirred tankreactor
Experiments with free �-glucosidase from almond were car-ried out in order to compare the performance of native enzymewith the immobilized one, at the same operating conditions (tem-perature, pH, buffer concentration, and substrate). Experimentswere carried out using 2, 2.5, 5, 7.5, 10, 15 and 20 mM of oleu-ropein at pH 6.5, temperature of 25 ◦C and enzyme concentration of0.035 mg/ml. Fig. 3 illustrates an example (5 mM) of the behaviourof oleuropein and glucose concentration as function of time. Bothsubstrate decrease and product formation were measured in orderto verify that results were congruent with the reaction stoichiom-etry (Fig. 1). As evidenced in the upper part of Fig. 3, the moles ofoleuropein disappeared were equivalent to the moles of glucose andthen of aglycon produced. From this type of graph V0 was calculatedas the tangent to the curve in the linear range when the conver-sion is less than 5%, so that product concentration was negligiblecompared to the substrate concentration.
An overall conversion degree of 30 (±3)% was reached after9000 s (2.5 h) of residence time and the initial reaction rate wasabout 1.80 × 10−2 (±0.001)mmol/cm3 h for the mentioned exam-ple.
Km and Vmax were calculated from the Lineweaver–Burk graphillustrated in Fig. 4. Their values are summarized in Table 2, wherethey are also compared with literature data. Results show that theKm value was comparable with the one from Capasso et al. [22],who used the oleuropein as substrate, even thought experimentswere carried out at different pH and temperature conditions.
The higher value for Vmax obtained in the present work com-pared to [22] might be due to a higher purity of the enzyme used.
The value of k+2, calculated according to Eq. (6), is also reportedin Table 2. The total amount of enzyme used in the STR was5.38 × 10−7 mmol/cm3.
Fig. 3. Behaviour of unreacted oleuropein ( ) and glucose concentration ( ) asfunction of time in the stirred tank reactor and relative correspondence to the reac-tion stoichiometry between mol of converted oleuropein ( ) and mol of glucoseproduced ( ).
Author's personal copy
220 R. Mazzei et al. / Journal of Membrane Science 339 (2009) 215–223
Table 1Concentration of oleuropein and glucose before and after the ultrafiltration through polyethersulphone (PES), regenerated cellulose (RC), andpolysulphone (PS) membranes of 30 kDa MWCO.
PES RC PS
Oleuropein initial solution (mg/ml) 1.35 (±0.02) 1.35 (±0.05) 1.35 (±0.03)Oleuropein after filtration (mg/ml) 1.23 (±0.02) 1.35 (±0.05) 1.35 (±0.03)Oleuropein adsorbed to the membrane (%) 9 – –Glucose initial solution (mg/ml) 0.23 (±0.02) 0.23 (±0.02) 0.23 (±0.02)Glucose after filtration (mg/ml) 0.23 (±0.02) 0.23 (±0.02) 0.23 (±0.02)Glucose adsorbed to the membrane (%) – – –
3.3. Reaction kinetics with ˇ-glucosidase immobilized in thebiocatalytic membrane reactor
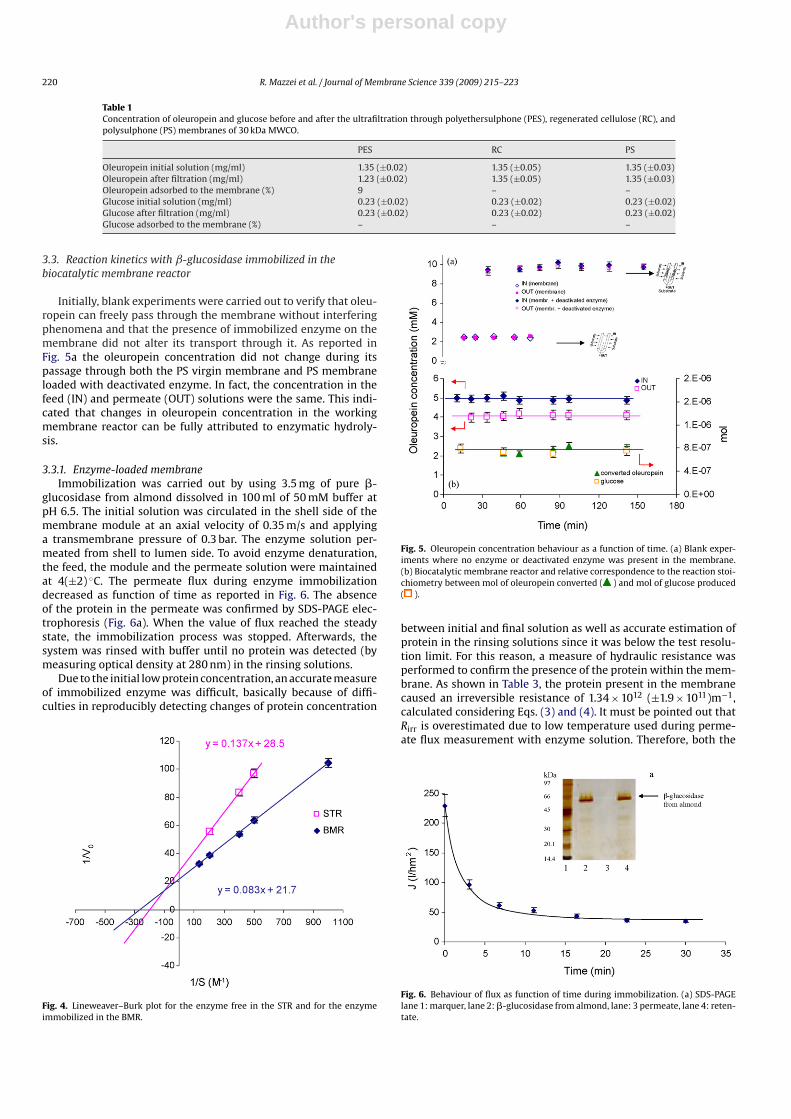
Initially, blank experiments were carried out to verify that oleu-ropein can freely pass through the membrane without interferingphenomena and that the presence of immobilized enzyme on themembrane did not alter its transport through it. As reported inFig. 5a the oleuropein concentration did not change during itspassage through both the PS virgin membrane and PS membraneloaded with deactivated enzyme. In fact, the concentration in thefeed (IN) and permeate (OUT) solutions were the same. This indi-cated that changes in oleuropein concentration in the workingmembrane reactor can be fully attributed to enzymatic hydroly-sis.
3.3.1. Enzyme-loaded membraneImmobilization was carried out by using 3.5 mg of pure �-
glucosidase from almond dissolved in 100 ml of 50 mM buffer atpH 6.5. The initial solution was circulated in the shell side of themembrane module at an axial velocity of 0.35 m/s and applyinga transmembrane pressure of 0.3 bar. The enzyme solution per-meated from shell to lumen side. To avoid enzyme denaturation,the feed, the module and the permeate solution were maintainedat 4(±2) ◦C. The permeate flux during enzyme immobilizationdecreased as function of time as reported in Fig. 6. The absenceof the protein in the permeate was confirmed by SDS-PAGE elec-trophoresis (Fig. 6a). When the value of flux reached the steadystate, the immobilization process was stopped. Afterwards, thesystem was rinsed with buffer until no protein was detected (bymeasuring optical density at 280 nm) in the rinsing solutions.
Due to the initial low protein concentration, an accurate measureof immobilized enzyme was difficult, basically because of diffi-culties in reproducibly detecting changes of protein concentration
Fig. 4. Lineweaver–Burk plot for the enzyme free in the STR and for the enzymeimmobilized in the BMR.
Fig. 5. Oleuropein concentration behaviour as a function of time. (a) Blank exper-iments where no enzyme or deactivated enzyme was present in the membrane.(b) Biocatalytic membrane reactor and relative correspondence to the reaction stoi-chiometry between mol of oleuropein converted ( ) and mol of glucose produced( ).
between initial and final solution as well as accurate estimation ofprotein in the rinsing solutions since it was below the test resolu-tion limit. For this reason, a measure of hydraulic resistance wasperformed to confirm the presence of the protein within the mem-brane. As shown in Table 3, the protein present in the membranecaused an irreversible resistance of 1.34 × 1012 (±1.9 × 1011)m−1,calculated considering Eqs. (3) and (4). It must be pointed out thatRirr is overestimated due to low temperature used during perme-ate flux measurement with enzyme solution. Therefore, both the
Fig. 6. Behaviour of flux as function of time during immobilization. (a) SDS-PAGElane 1: marquer, lane 2: �-glucosidase from almond, lane: 3 permeate, lane 4: reten-tate.

Author's personal copy
R. Mazzei et al. / Journal of Membrane Science 339 (2009) 215–223 221
Tab
le2
Kin
etic
par
amet
ers
of�
-glu
cosi
das
efr
omal
mon
dob
tain
edin
the
pre
sen
tw
ork
and
com
par
ison
wit
hli
tera
ture
dat
a.
Enzy
me
stat
us
Enzy
me
sup
por
tSu
bstr
ate
Km
(mM
)V
max
ak +
2(s
−1)
k +2/K
m(m
M−1
s−1)
pH
T(◦ C
)R
efer
ence
Free
–O
leu
rop
ein
4.80
(±1.
1×
10−1
)3.
5×
10−1
(±4
×10
−3)
(Mh
−1)
18(±
9×
10−1
)3.
756.
525
Pres
ent
wor
k1.
9×
10−2
(±9.
5×
10−4
)(U
/mg)
b
Imm
obil
ized
(en
trap
ped
)Po
lysu
lph
one
cap
illa
rym
embr
anes
Ole
uro
pei
n3.
80(±
1.4
×10
−1)
4.6
×10
−2(±
2×
10−3
)(M
h−1
)18
c4.
736.
525
Pres
ent
wor
k
Free
–O
leu
rop
ein
4.66
3.6
×10
−3(U
/mg)
––
5.5
37[2
2]
aU
nit
isin
dic
ated
per
each
valu
eas
auth
ors
use
dd
iffe
ren
tu
nit
s.b
We
stro
ngl
yre
com
men
dth
eu
seof
“mol
arco
nce
ntr
atio
n/t
ime”
asre
acti
onra
teu
nit
.How
ever
,sin
ceth
eu
nit
U/m
g(o
rm
mol
min
−1m
g−1)
isco
mm
only
use
d,w
eal
sore
por
tth
eV
max
acco
rdin
gto
this
un
it.T
his
Vm
axw
asca
lcu
late
dfr
omLi
new
eave
r–B
urk
plo
tu
sin
gV
0ex
pre
ssed
inte
rms
ofU
/mg.
cA
ssu
med
equ
ival
ent
toth
eon
eof
the
free
enzy
me
onth
eba
sis
ofsi
mil
arK
man
dV
max
valu
es.
amount of immobilized enzyme and promoted hydraulic resistanceare rough estimations.
Anyway, the hydraulic resistance due to irreversible foulingpromoted by immobilized enzyme resulted roughly as high asthe hydraulic resistance due to the polymeric membrane itself.Although this does not give the mass of immobilized enzyme, itindicates that the enzyme remained stably entrapped within themembrane.
The entrapment immobilization method provides a random dis-tribution and orientation of the protein within the membrane, as itwas confirmed by in situ detection of the enzyme using an adaptedimmunoassay method reported in our previous work [37].
Reaction rates were measured with both new prepared andre-used enzyme-loaded membranes. In the latter case, after eachexperiment, the efficiency of the enzyme-loaded membrane wasverified by carrying out a reaction in the initial operating conditions.
The catalytic activity of the immobilized enzyme as a function oftime was also evaluated, as described in Section 2. As shown in Fig. 7the catalytic activity did not show any significant decay for about30 h operation time. Test carried out after two months preservationat −80 ◦C confirmed that the enzyme was still active.
3.3.2. Reaction kinetics with immobilized ˇ-glucosidaseThe enzymatic hydrolysis in the enzyme-loaded membrane
reactor was carried out using 100 ml of oleuropein solution at pH6.5 and 25 ◦C. An example of time course of substrate concentra-tion in the feed (IN) and permeate (OUT) solution through thebiocatalytic membrane for an oleuropein initial concentration of5 mM is reported in Fig. 5b. The figure also shows the stoichio-metric correspondence between moles of oleuropein convertedand glucose (and equivalent moles of aglycon) produced. The con-version was about 20 (±3)% for a residence time of about 126 s.During the experiment the reaction rate was about 2.5 × 10−02
(±1 × 10−3)mmol/cm3 h (calculated with Eq. (11)).The different conversion degree in STR (30%) and MBR (20%) was
due to the lower residence time of the substrate in the biocatalyticmembrane reactor, 126 s, compared with 9000 for STR.
Experiments at different substrate concentration (1, 2, 2.5, 5,7.5, 10 and 15 mM) were carried out to measure Km and Vmax forimmobilized enzyme. Lineweaver–Burk plot is reported in Fig. 4,and values of kinetic parameters are reported in Table 2. Obtainedresults are comparable to the one of the free native enzyme.The common observation in the literature was that immobilizedenzymes increase their stability while decreasing their catalyticactivity, which is one of the major drawback that limits the appli-cation of the technology on a large scale. This study shows thatthe inverse relationship between stability and activity of immobi-lized enzymes is not a general rule. It is worth nothing that thedemonstration has been made on the basis of kinetic parameters.
Results confirmed that the immobilization procedure did notalter the catalytic properties of the enzyme and that the system issuitable for further optimization and development at larger scale.
Fig. 7. Enzyme activity of immobilized �-glucosidase as a function of time. T = 25 ◦C,pH = 6.5, oleuropein concentration = 2 mM.

Author's personal copy
222 R. Mazzei et al. / Journal of Membrane Science 339 (2009) 215–223
Table 3Hydraulic resistance of membrane and fouling components for 30 kDa polysulphone capillary membrane.
Membrane Solution Permeability (m/s Pa) Resistance
R m−1
New membrane Ultrapure water 6.39 × 10−10 Rm 1.75 × 1012 (±7.6 × 1014)Membrane duringa immobilization �-Glucosidase solution 2.97 × 10−10 a RTOT 3.78 × 1012 (±2.5 × 1015)Membrane after immobilization and rinsing step Ultrapure water 3.64 × 10−10 Ri 1.34 × 1012 (±1.9 × 1011)– – – Rf 2.03 × 1012 (±2.0 × 1011)– – – Rr 6.9 × 1011 (±6.9 × 1010)
Rm = hydraulic resistance due to the membrane; RTOT = hydraulic resistance due to membrane and fouling, Rf = hydraulic resistance due the overall fouling; Ri = hydraulicresistance due to the irreversible fouling; Rr = hydraulic resistance due to reversible fouling.
a This value was calculated from the constant flux value as a function of time (see Fig. 6).
For what concerns k+2 a thorough evaluation of its value was notpossible, due to the difficulties in measuring the precise immobi-lized enzyme amount. On the other hand, on the basis of similarvalues of Km and Vmax between free and immobilized enzymewe can state that the immobilized enzyme is working with sameperformance, or even slightly better (low Km and higher Vmax) com-pared to the free enzyme. Then k+2 for the immobilized enzymeshould be also of the same order as the one of the free enzyme.Assuming k+2 equal to the one for free enzyme the working immo-bilized enzyme can be calculated. In order to have a k+2 similar tothe free enzyme, the working immobilized enzyme concentrationshould be around 7.05 × 10−7 mmol/cm3.
Table 2 also reports the k+2/Km ratio. This ratio expresses theenzyme efficiency. Either a large value of k+2 (rapid turnover) or asmall value of Km (high affinity for substrate) makes k+2/Km large. Ifwe consider the k+2 value similar to the one of the free enzyme, theratio between k+2/Km becomes larger for the biocatalytic membranereactor, suggesting that the enzyme may have higher efficiencywhen immobilized.
4. Conclusions
In this work, the catalytic performance of a biocatalytic mem-brane reactor using �-glucosidase immobilized in polysulfonecapillary membranes was investigated. The membrane reactorwas used for the continuous hydrolysis of oleuropein (an abun-dant vegetable raw material) into aglycon (a powerful naturalantimicrobial agent) and glucose. In addition, the membrane reac-tor system simultaneously performed their separation from thereaction microenvironment. Kinetic parameters of �-glucosidaseimmobilized in the membrane reactor have been evaluated andcompared with those of the ones of the free enzyme. Resultsevidenced that intrinsic kinetics of enzyme immobilized inthe polysulfone membrane reactor was not negatively affected(Km = 3.80 ± 0.14 mM; Vmax = 0.046 ± 0.002 M h−1) compared to thefree enzyme [Km = 4.80 (±0.11)mM, Vmax = 0.035 (±0.004)M h−1;k+2 = 18 (±0.9)s−1]. On the contrary it seemed slightly improved,i.e. lower Km value indicates higher enzyme–substrate affin-ity. This shows that lower catalytic activity of immobilizedenzyme, commonly observed in the literature, is not an intrinsictechnological drawback and that appropriate control of bio-hybrid microstructured systems, microenvironment conditionsand transport properties can even improve their catalytic effi-ciency.
Acknowledgements
The Italian “Ministero dell’Istruzione dell’Università e dellaRicerca” (MIUR), project FIRB-RBNE03JCR5, is gratefully acknowl-edged for the financial support.
The “NanoMemPro European Network of Excellence onNanoscale-Based Membrane Technologies, NMP3-CT-2004-
500623” is acknowledged for the support in the critical analysis ofthe state of the art in the field.
References
[1] R. Briante, F. La Cara, F. Febbraio, R. Baroni, G. Piccialli, R. Carolla, P. Mainolfi, L. DeNapoli, M. Patumi, G. Fontanazza, R. Nucci, Hydrolysis of oleuropein by recom-binant �-glycosidase from hyperthermophilic archaeon Sulfolobus solfataricusimmobilized on chitosan matrix, J. Biotechnol. 77 (2000) 275–286.
[2] M. Esti, L. Cinquanta, E. La Notte, Phenolic compounds in different olive varieties,J. Agric. Food Chem. 46 (1998) 32–35.
[3] F. Paiva-Martins, M.H. Gordon, Isolation and characterization of the antioxidantcomponent 3,4-dihydroxyphenylethyl 4-formyl-3-formyl-3-formylmethyl-4-hexenoate from olive (Olea europaea) leaves, J. Agric. Food Chem. 49 (2001)4214–4219.
[4] I. Kubo, A. Matsumoto, I. Takase, A multichemical defense mechanism of bitterolive Olea europaea (oleaceae), J. Chem. Ecol. 11 (1985) 251–263.
[5] M.J. Amiot, A. Fleuriet, J.J. Macheix, Accumulation of oleuropein derivativesduring olive maturation, Phytochemistry 28 (1989) 67–69.
[6] F. Visioli, G. Bellomo, G. Montedoro, C. Galli, Antioxidant and other biologicalactivities of olive mill waste waters, Atherosclerosis 117 (1995) 25–32.
[7] M.H. Gordon, F. Paiva-Martins, F. Almeida, Antioxidant activity of hydroxyty-rosol acetate compared with that of other olive oil polyphenols, J. Agric. FoodChem. 49 (2001) 2480–2485.
[8] P.S. Rodis, V.T. Karathanos, A. Mantzavinou, Partitioning of olive oil antioxidantsbetween oil and water phases, J. Agric. Food Chem. 50 (2002) 596–601.
[9] F. Paiva-Martins, M.H. Gordon, P. Gameiro, Activity and location of olive oilphenolic antioxidants in liposomes, Chem. Phys. Lipids 124 (2003) 23–36.
[10] M.N. Vissers, P.L. Zock, M.B. Katan, Bioavailability and antioxidant. Effectsof olive oil phenols in humans: a review, Eur. J. Clin. Nutr. 58 (2004) 955–965.
[11] M. Scigelova, S. Singh, D.H.G. Crout, Glycosidases—a great synthetic tool, J. Mol.Catal. B: Enzymol. 6 (1999) 483–494.
[12] H. Yeoh, Kinetic properties of Cassava ß-glucosidase immobilized in photo-crosslinkable resins, Biotechnol. Lett. 13 (1991) 697–700.
[13] A. Esen, �-Glucosidases: Biochemistry and Molecular Biology, American Chem-ical Society, Washington, 1993.
[14] M.A. Montero, A. Romeu, Properties of almond �-glucosidase immobilizedon concanavalin A–sepharose, Biochem. Mol. Biol. Int. 30 (1993) 685–689.
[15] Y. Hsuanyu, K.J. Laidler, Flow kinetics of immobilized �-glucosidase, Biochem.Cell Biol. 64 (1986) 139–145.
[16] S.A. Backer, S.H. Doss, C.J. Gray, J.F. Kennedy, M. Stacy, T.H. Teo, �-Glucosidasechemically bound to microcrystalline cellulose, Carbohydr. Res. 20 (1971)1–7.
[17] V.R. Srinivasan, M.W. Bumm, Isolation and immobilization of �-d-glucosidasefrom Alcaligenes faecalis, Biotechnol. Bioeng. 16 (1974) 1413–1418.
[18] J.L. Iborra, M.R. Castellar, M. Cànovas, A. Manjòn, Picrocicrin hydrolysis byimmobilized �-glucosidase, Biotechol. Lett. 14 (1992) 475–480.
[19] H. Nagamoto, Y. Matsushita, K. Sugamoto, T. Matsui, Preparartion and propertiesof gelatin-immobilized �-glucosidase from Pirococcus furious, Biosci. Biotech-nol. 69 (2005) 128–136.
[20] R. Limiroli, R. Consonni, G. Ottolina, V. Marsilio, G. Bianchi, L. Zetta, J. Chem. Soc.Perkin Trans. 5 (1995) 1519–1523.
[21] V. Marsilio, B. Lanza, N. Pozzi, Progress in table olive debittering: degradationin vitro of oleuropein and its derivatives by Lactobacillus plantarum, J. Am. OilChem. Soc. 73 (1996) 593–597.
[22] R. Capasso, A. Evidente, C. Visca, L. Gianfreda, M. Maremoti, G. Greco, Produc-tion of glucose and bioactive aglycone by chemical and enzymatic hydrolysis ofpurified oleuropein from Olea europaea, Appl. Biochem. Biotechnol. 61 (1997)365–377.
[23] R. Briante, F. La Cara, M.P. Tonziello, F. Febbraio, R. Nucci, Antioxidant activity ofthe main bioactive derivates from oleuropein hydrolysis by hyperthermophilic�-glycosidase, J. Agric. Food Chem. 49 (2001) 3198–3203.
[24] R. Briante, M. Patumi, F. Febbraio, R. Nucci, Production of highly purified hydrox-ytyrosol from Olea europaea leaf extract biotransformed by hyperthermophilic�-glycosidase, J. Biotechnol. 111 (2004) 67–77.

Author's personal copy
R. Mazzei et al. / Journal of Membrane Science 339 (2009) 215–223 223
[25] M. Guiso, C. Marra, Highlights in oleuropein aglycone structure, Nat. Prod. Res.19 (2005) 105–109.
[26] E. Drioli, L. Giorno, Biocatalytic Membrane Reactors: Applications in theBiotechnology and Pharmaceutical Industry, Taylor & Francis, London, 1999,p. 51.
[27] L. Giorno, E. Drioli, Biocatalytic membrane reactors: applications and perspec-tives, Trends Biotechnol. 18 (2000) 339–349.
[28] L. Giorno, R. Mazzei, E. Drioli, Biochemical membrane reactors in industrial pro-cesses in Membranes Operations: Innovative separations and transformations,Eds. E. Drioli, L. Giorno, Wiley-VCH, Weinheim, 2009, p. 397.
[29] G.M. Rios, M. Belleville, D. Paolucci-Jeanjean, Membrane engineering in biotech-nology: quo vamus? Trends Biotechnol. 25 (2007) 242–246.
[30] V.M. Balcão, A.L. Paiva, F.X. Malcata, Bioreactors with immobilized lipase: stateof the art, Enzyme Microb. Technol. 18 (1996) 392–416.
[31] R. Valadez-Blanco, F. Castelo Ferreira, R. Ferreira Jorge, A.G. Livingston, Mem-brane bioreactor for biotransformations of hydrophobic molecules usingorganic solvent nanofiltration (OSN) membranes, J. Membr. Sci. 317 (2008)50–64.
[32] H.A. Sousa, C. Rodrigues, E. Klein, C.A.M. Afonso, J.G. Crespo, Immobilisationof pig liver esterase in hollow fibre membranes, Enzyme Microb. Technol. 29(2001) 625.
[33] Z.M. Liu, J. Dubremez, V. Richard, Q. Yang, Z.K. Xu, P. Seta, Useful method forthe spatial localization determination of enzyme (peroxidase) distribution onmicrofiltration membrane, J. Membr. Sci. 267 (2005) 2.
[34] A. Ranalli, S. Contento, L. Lucera, M. Di Febo, D. Marchigiani, V. Di Fonzo, Factorsaffecting the contents of irinoids oleuropein in olive leaves (Olea europaea L.),J. Agric. Food Chem. 54 (2006) 434–440.
[35] P. Aimar, J. Howell, Effect of concentration polarization boundary layer devel-opment on the flux limitations in ultrafiltration, Chem. Eng. Res. Des. 67 (1989)225–261.
[36] J.E. Bailey, D.F. Ollis, Biochemical Engineering Fundamentals, McGraw-Hill, NewYork, 1986, p. 97.
[37] S. Mazzuca, L. Giorno, A. Spadafora, R. Mazzei, E. Drioli, Immunolocalizationof �-glucosidase immobilized within polysulphone capillary membrane andevaluation of its activity in situ, J. Membr. Sci. 285 (2006) 152–158.
Copyright © 2022 FDOKUMEN




















