
Recent Advances in Polyamine Metabolism and Abiotic Stress Tolerance
Upload
independentCategory
view
1download
0
Jurassic lagoonal environments and quasi-abiotic platylimestone accumulation: microbial interventions
NICOLAS TRIBOVILLARD*, JEAN TRICHET , CHRISTIAN DE FARGE andALAIN TRENTESAUX**Universite Lille 1, Laboratoire de SeÂdimentologie et GeÂodynamique, UMR CNRS 8577, BaÃtiment SN5,F-59655 Villeneuve d'Ascq cedex, France (E-mail: [email protected];[email protected]) Universite d'OrleÂans, UMR CNRS 6531, Laboratoire de GeÂologie de la MatieÁre Organique, F-45067OrleÂans cedex 2, France (E-mail: [email protected]; [email protected])
ABSTRACT
Within the regressive sequence (eustatic sea-level fall) that characterizes the
French Southern Jura carbonate platform during the Kimmeridgian, a
succession of three facies (called here units 1±3) may be observed in a
palaeolagoon near Orbagnoux. Unit 1 is a shallow-water, intensely burrowed,
carbonate deposit; unit 2 (consisting of laminated, organic matter-rich
carbonate) represents a shallow lagoonal environment, prone to stromatolite
development and organic matter accumulation; unit 3 consists of platy
limestones almost devoid of identi®able biota. Trace element distributions
indicate that depositional and early diagenetic conditions must have been
reducing during deposition of unit 1 and, especially, unit 2, whereas they were
much less reducing during deposition of unit 3. However, the latter does not
contain body or trace fossils, with the exception of structures interpreted as
calci®ed bacteria. The reasons why the apparently normal marine environment
of unit 3 did not allow a normal biota to develop are explored and lead us to
propose that: (i) unit 2 may be a Jurassic counterpart of modern kopara deposits
(stromatolites of French Polynesian atolls); and (ii) unit 3 may be the result of
palaeokopara reworking. This model invokes the extensive intervention of
microbes in the formation of these carbonate deposits.
Keywords Bacteria, Jura, Kimmeridgian, kopara, platy limestone,
stromatolites.
INTRODUCTION
During the Late Jurassic, the area that is now theFrench Southern Jura was a carbonate platform,which underwent a shallowing trend from pelagicto paralic deposition (Bernier, 1984). During thisrelative sea-level fall, a barrier reef developed andproduced an isolated shallow lagoon. Whencirculation was most reduced, bituminous lam-inites formed in this lagoon. Such organic-richsediments are encountered in the vicinity of thesmall hamlet of Orbagnoux (Fig. 1). This potentialpetroleum source rock has been studied exten-sively by several authors (Riche, 1904; Gubler &Louis, 1956; Bernier & Courtinat, 1979; Bernier,1984; Courtinat, 1989; Gorin et al., 1989; Bernier
et al., 1991; Tribovillard et al., 1991, 1992, 1994;Mongenot et al., 1997; Tribovillard, 1998), whohave concluded that the organic-rich facies wasdeposited as a result of: enhanced phytoplanktonproduction in a very quiet depositional environ-ment; the presence of cyanobacterial activity,which developed bio®lms at the sediment sur-face; and a pronounced scarcity of reactive ironthat allowed the early vulcanization of organicmatter. The aim of the present paper is to examinethe sediments that lie immediately above andbelow the bituminous laminites. A comparisonbetween the facies of these strata and that of thebituminous laminites will help to determinewhich factors allowed or precluded the deposi-tion of the organic-rich facies.
Sedimentology (1999) 46, 1183±1197
Ó 1999 International Association of Sedimentologists 1183

GEOLOGICAL SETTING
The most recent and complete stratigraphicreconstruction of the French Southern JuraMountains was published by Bernier (1984).The very simpli®ed scheme that follows derivesmainly from this work. During Early Kimme-ridgian times, the study area (Fig. 1) belonged toa pelagic environment in which ammonite-richlimestones formed, re¯ecting open-marine condi-tions (`Couches aÁ CeÂphalopodes' Formation). Arelative sea-level fall and platform progradationled to the deposition of neritic limestones (`Calc-aires de Tabalcon' Formation or unit 1 of thisstudy; Fig. 2). These massive beds contain plank-tonic organisms (globigerinid foraminifera andradiolaria) and ammonites in the lower part of theformation, which are progressively replaced bygastropods, bivalves and sponges. Rare calcareousalgae (Salpingoporella, Clypeina) were observedby Bernier (1984). During the H. beckeri ammonitezone of the Kimmeridgian, the development of areef barrier isolated a shallow lagoon in whichbituminous laminites were deposited under a
water column that was most probably strati®ed(the `Laminites Bitumineuses' Member of the`Calcaires en Plaquettes' Formation or unit 2;Fig. 2). The environment was sometimes con-®ned: levels rich in gypsum pseudomorphs areobserved. During the Late Kimmeridgian, accord-ing to Bernier (1984), the depositional environ-ment deepened temporarily (a result of possiblelocal subsidence), which led to the deposition ofplaty limestones (the `Calcaires en Plaquettes'Formation sensu stricto or unit 3; Fig. 2). Alter-native interpretations are proposed below. Thismudstone unit contains very few fossils: nococcoliths have been observed, and only rarebenthonic foraminifera have been identi®ed. Onlyin the uppermost levels, ophiurids and bivalves(with the valves still articulated) occur togetherwith evidence of emersion (Enay & Hess, 1970;Bernier, 1984). Bernier (1984) was puzzled by theabsence of benthonic life and suggested that theepisodes of mud deposition were too sudden toallow benthonic populations to develop. DuringPortlandian times, the area was covered with veryshallow carbonate facies, representing tidal ¯at
Fig. 1. Geological setting of thestudy area. Redrawn from Bernier(1984).
1184 N. Tribovillard et al.
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

environments (`Couches de Chailley' and `Tidal-ites de Vouglans' Formations). By the end of theJurassic, the area corresponded to palustrine andperitidal environments (Purbeckian facies; Stras-ser, 1988, 1991).
This study focuses on stratigraphic units 1 and 3that frame the bituminous laminites (unit 2) in theOrbagnoux palaeolagoon, and emphasis is laidupon unit 3. The results of this study are valid for
the palaeolagoon of Orbagnoux, but cannot nec-essarily be extended to the entire Jura lagoon.
PREVIOUS WORK ON THE `LAMINITESBITUMINEUSES' (UNIT 2)
This Bituminous Laminites unit has been studiedextensively (Riche, 1904; Bernier & Courtinat,
Fig. 2. Schematic lithological col-umn of the three units studied,based on observations from theDorche Torrent, at Orbagnoux(French map IGN, Seyssel sheet,1:50 000; X � 866á91, Y � 2115á65).
Jurassic lagoonal environments 11851
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

1979; Bernier, 1984; Courtinat, 1989; Bernieret al., 1991), particularly in the Orbagnoux pal-aeolagoon (Gubler & Louis, 1956; Gorin et al.,1989; Tribovillard et al., 1991, 1992, 1994; Mon-genot et al., 1997; Tribovillard, 1998) and isdescribed in much detail elsewhere (Tribovillardet al., 1991, 1992; Mongenot et al., 1997; Tribovil-lard, 1998). A few aspects are reviewed here.Basically, the Orbagnoux bituminous laminitesare made up of two components: carbonate andorganic matter, whatever their respective natureand origin. The carbonate content ¯uctuatesbetween around 75% and 96%. The organiccontent is higher in the dark laminae than in thelight ones [total organic carbon, (TOC) � 4á47±8á55% compared with 0á65±8á03%]. Values of thehydrogen index of the kerogens are always veryhigh (755±966 mg of hydrocarbons g)1 TOC; Esp-italie et al., 1985). Elemental analyses performedon the isolated kerogens indicate that organicsulphur is unusually abundant (12±17á6% of thekerogens).
Within the `Laminites Bitumineuses', the fol-lowing two subfacies alternate.
Parallel laminated beds
These are made of alternating, submillimetre-scale, lighter or darker coloured laminae. Theseparallel, continuous, individual layers areclustered into millimetre- or centimetre-scalelaminae, the depth of colour of which dependson the dominant component in individual layers.Various macro- and microfossils are present inthe parallel laminae, including ammonites,benthonic foraminifera and ostracods. Someammonite mass mortality may be observed: verythin accumulations of juvenile forms of the genusAspidoceras, which do not disturb the laminatedstructure. Scanning electron microscopy (SEM)indicates that the light-coloured laminae arecomposed exclusively of coccospheres andcoccoliths. The dark-coloured laminae containfewer coccoliths and coccospheres, embeddedwithin a matrix of structureless, gel-like, organicmatter. Acid-etched samples allow the distribu-tion of the thin organic seams, which stand out inrelief, to be observed. The organic laminae buildan anastomosed mesh-like network.
The parallel laminated facies seems toderive mostly from settling mechanisms: peri-ods of ¯ourishing coccolithophorids led to theaccumulation of lighter coloured laminae, con-sisting exclusively of coccoliths. These periodsalternated with episodes of organic matter accu-
mulation derived from planktonic algae (Tribovil-lard et al., 1992; Mongenot et al., 1997, 1999).The exceptional quality of lamina-scale correla-tion testi®es to a remarkably calm depositionalenvironment.
Undulating lamina beds
This facies comprises irregularly undulated,alternating light- or dark-coloured, laminae,grossly parallel to bedding. The thickness ofthese laminae is the same as those in the parallellaminated facies. The light-coloured undulatinglaminae may exhibit bird's eyes and ¯at fenestrae.Thin-section observations show an alternation ofdark laminae, consisting of bundles of very thinseams, and light-coloured, thicker, carbonatelaminae, occasionally containing abundantpeloids. These peloids are interpreted as beingof cyanobacterial origin (Tribovillard, 1998). Inthe undulating laminae, no coccoliths have beenobserved, but SEM observations reveal that car-bonate is present as micrite grains, which arealmost equidimensional and have grain sizes of1±5 mm.
The undulating lamina beds must have beenless affected by or, more likely, unaffected bysettling mechanisms. As concluded previously byTribovillard et al. (1992), the biolaminationsmust have resulted from self-burial processes,i.e. mat-by-mat overgrowth related to ecologicalchanges possibly affecting water salinity, lightintensity and temperature in the absence ofparticle settling (Gerdes et al., 1991).
Throughout the laminites, some rare, authi-genic quartz grains occur. A few dedolomitizedrhombs are also interspersed within the sedi-ments. No gypsum crystals are preserved, butnumerous pseudomorphs (composed of calcite)occur in the various facies. The common presenceof oxygen-demanding planktonic and benthonicorganisms supports the conclusion that the watercolumn must have remained (almost?) continu-ously oxic, whereas the underlying sedimentswere constantly characterized by anoxic pore-waters. The sharp chemical interface was causedby the presence of cyanobacterial bio®lms actingas a barrier between the two contrasting environ-ments.
The simultaneous occurrence of abundantorganic matter and reducing conditions favouredintense sulphate reduction. Hence, large amountsof sulphide were released and trapped within thesediment below the cyanobacterial barrier. Asiron was limited, sulphide reacted with organic
1186 N. Tribovillard et al.
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

molecules; this early vulcanization of organicmatter is an ef®cient agent of organic matterpreservation (Sinninghe Damste et al., 1989;Tegelaar et al., 1989).
SAMPLES AND METHODS
The section studied is located alongside theDorche Torrent, in the small hamlet of Orb-agnoux, on the right bank of the RhoÃne River(French map IGN, Seyssel sheet, 1:50 000;X � 866á91, Y � 2115á65). The samples werestudied using light microscopy and scanningelectron microscopy (SEM) of gold-coated mate-rials (Folk & Lynch, 1997). Some samples wereetched with dilute HCl (0á2 M) before observationwith a binocular microscope, in order to revealpossible sedimentary features or structures, asetching proved successful in revealing peloids inthe undulated laminae (Tribovillard, 1998). Pal-ynofacies were isolated using classical methods(Combaz, 1980). Bitumen-free samples weredemineralized in order to obtain a kerogen.Initial decarbonatation was achieved using HCl(1 M). The decarbonated samples were thentreated with HCl (6 M) and HF (24 M) for furtherdemineralization. Clay mineral data were deter-mined by X-ray diffraction analysis of thecarbonate-free clay fractions (<2 lm). Sampleswere decalci®ed with 0á2 M HCl, and the <2 lmfraction was separated by settling. Orientedmounts were prepared on glass slides andscanned three times (untreated samples, glycola-ted and heated for 2 h at 490 °C). Analyticaltechniques, determination and semi-quantitativeestimation of clay minerals are detailed inHoltzapfel (1985). Total carbon, total organiccarbon (TOC) and total sulphur contents weredetermined by combustion using a Leco CS 125carbon±sulphur analyser. Elemental analyseswere carried out by ICP-AES (major and minorelements) or ICP-MS (trace elements) at thespectrochemical laboratory of the Centre deRecherches en PeÂtrographie et GeÂochimie ofVandúuvre-les-Nancy (geochemistry facility ofthe French Centre National de la RechercheScienti®que). The samples were prepared byLiBO2 fusion and HNO3 dissolution. The analyt-ical accuracy and precision were both found tobe better than 1% (mean 0á5%) for major andminor elements, or 5% for trace elements.This was checked using international referencematerial and analysing replicate samples respec-tively.
RESULTS
Lithologies of units 1 and 3
Massive limestones of unit 1
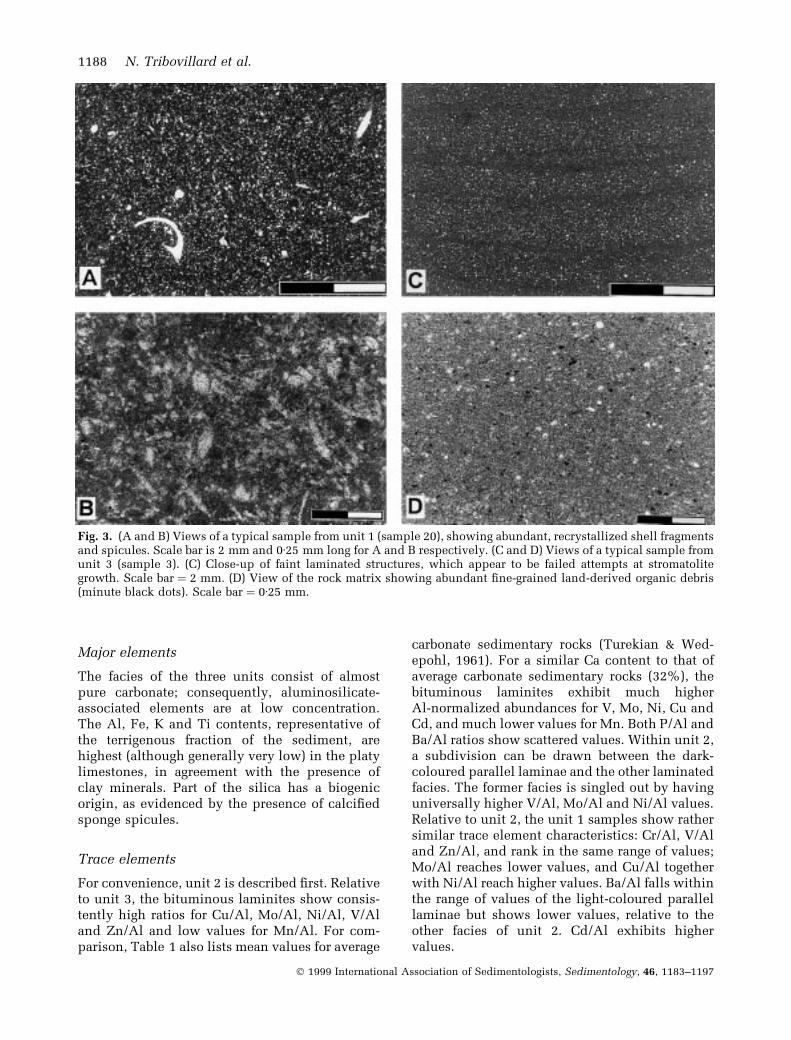
This unit is made of decimetre-thick, beige-coloured limestone beds with sharply de®nedboundaries. The ®ne-grained carbonate containsnumerous, entire or broken bivalve and gastropodshells. No sedimentary structures, such as lami-nation, are visible. The microfacies is a wacke-stone biopelmicrite with abundant tiny shellfragments and sponge spicules (Fig. 3A and B).SEM reveals the presence of abundant coccoliths.
Platy limestones of unit 3
This unit is made of thin limestone beds (a fewcentimetres thick), occasionally splitting intomillimetre-thick laminae. The dark beige-colouredmicrite shows a rather homogeneous texture withoccasional lamination (individual laminae orbundles of less than ®ve laminae; Fig. 3C and D).These unobtrusive laminae resemble individualcryptalgal laminae, as if better development of a(cyanobacterial) bio®lm had been hampered orprevented. No fossils, not even coccoliths, havebeen observed, whatever the scale of observation.However, SEM reveals abundant carbonate bodiesof <1 mm length, which may be interpreted ascalci®ed bacteria (Fig. 4A and B). These seem tohave been the only life form present, together withcysts of dino¯agellates (see below). Acid etchingdid not reveal additional features.
Mineralogy
Units 1 and 2 do not contain any mineral phasesother than calcite (iron sulphide, clay minerals andterrigenous quartz grains are all absent). Unit 2occasionally contains calcite pseudomorphs aftergypsum. The platy limestones of unit 3 are pre-dominantly carbonate (91±96%), but contain smallquantities of clay minerals: illite, illite±smectitemixed layers and kaolinite. The diversity of theclay mineral assemblage indicates a detrital ratherthan a diagenetic origin. Thin-section observationsalso demonstrate the presence of scattered, tinypyrite specks.
Geochemistry
Table 1 summarizes the chemical composition ofthe samples studied. The data concerning unit 2come from Mongenot (1998).
Jurassic lagoonal environments 11871
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197
Major elements
The facies of the three units consist of almostpure carbonate; consequently, aluminosilicate-associated elements are at low concentration.The Al, Fe, K and Ti contents, representative ofthe terrigenous fraction of the sediment, arehighest (although generally very low) in the platylimestones, in agreement with the presence ofclay minerals. Part of the silica has a biogenicorigin, as evidenced by the presence of calci®edsponge spicules.
Trace elements
For convenience, unit 2 is described ®rst. Relativeto unit 3, the bituminous laminites show consis-tently high ratios for Cu/Al, Mo/Al, Ni/Al, V/Aland Zn/Al and low values for Mn/Al. For com-parison, Table 1 also lists mean values for average
carbonate sedimentary rocks (Turekian & Wed-epohl, 1961). For a similar Ca content to that ofaverage carbonate sedimentary rocks (32%), thebituminous laminites exhibit much higherAl-normalized abundances for V, Mo, Ni, Cu andCd, and much lower values for Mn. Both P/Al andBa/Al ratios show scattered values. Within unit 2,a subdivision can be drawn between the dark-coloured parallel laminae and the other laminatedfacies. The former facies is singled out by havinguniversally higher V/Al, Mo/Al and Ni/Al values.Relative to unit 2, the unit 1 samples show rathersimilar trace element characteristics: Cr/Al, V/Aland Zn/Al, and rank in the same range of values;Mo/Al reaches lower values, and Cu/Al togetherwith Ni/Al reach higher values. Ba/Al falls withinthe range of values of the light-coloured parallellaminae but shows lower values, relative to theother facies of unit 2. Cd/Al exhibits highervalues.
Fig. 3. (A and B) Views of a typical sample from unit 1 (sample 20), showing abundant, recrystallized shell fragmentsand spicules. Scale bar is 2 mm and 0á25 mm long for A and B respectively. (C and D) Views of a typical sample fromunit 3 (sample 3). (C) Close-up of faint laminated structures, which appear to be failed attempts at stromatolitegrowth. Scale bar � 2 mm. (D) View of the rock matrix showing abundant ®ne-grained land-derived organic debris(minute black dots). Scale bar � 0á25 mm.
1188 N. Tribovillard et al.
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

The platy limestones of unit 3 are drasticallydifferent, with much lower values for Ba/Al,Cd/Al, Co/Al, Cu/Al, Mo/Al, Ni/Al, V/Al andZn/Al ratios. When compared with unit 1, unit 3also shows lower U/Al and Cr/Al, but higher Sr/Al (approximate double) and Zr/Al (relativelymore abundant lithogenic supply). Unit 3 has thehighest Mn content (128±149 lg g)1 comparedwith 9±36 lg g)1 for unit 2 and 36±37 lg g)1 forunit 1).
Organic content
The bituminous laminites contain abundantorganic matter (TOC reaching 12%; Tribovillardet al., 1991), which is overwhelmingly amorphous
under light microscopy and has a gel-like texture(SEM imaging; see Mongenot et al., 1997 for adetailed study of the organic matter in unit 2).
The massive limestones of unit 1 have a loworganic content (TOC <0á2%). Pyrolyses perform-ed upon the HCl-insoluble residue indicate amean TOC of around 19%. The mean sulphurcontent of the bulk rock and the HCl-insolubleresidue is <0á1% and 2á9% respectively. Palyno-facies observations reveal a dominance of marineamorphous organic matter (plus dinocysts andabundant foraminifera linings), together withland-derived organic material: spores and pollen,woody fragments (black or brown) and cuticles.
The platy limestones (unit 3) also have loworganic contents: TOC is below detection (<0á2%)in the bulk rock and 3á6% for the insolubleresidue. Sulphur contents are 0á1% and 0á4%respectively. The palynofacies are made of amor-phous organic matter and dinocysts, as well aswoody fragments, spores and pollen. Foraminif-era linings were not observed. The terrestrialorganic material is ®ner grained in this facies, andbisaccate pollen seems to be more abundant thanin the other units. All ligneous fragments areblack (inertinite-like).
INTERPRETATION
The massive limestones of unit 1 contain abund-ant body fossils, although the faunal diversity isrelatively low, consisting of sponges, gastropodsand bivalves. Intense bioturbation prevented thepreservation of sedimentary structures. The sur-face waters were favourable to the developmentof coccolithophorids. Nevertheless, at this time,the palaeolagoon already showed signs ofrestricted environmental conditions, as indicatedby the trace element content: relatively highV/Al, Mo/Al, Cd/Al, Ni/Al, Co/Al, Cu/Al, Zn/Al,U/Al, Cr/Al as well as low Mn/Al ratios, indicatethe occurrence of reducing conditions withinbottom waters or, more probably, below thewater±sediment interface, as a benthonic faunawas present (Brumsack, 1986; Breit & Wanty,1991; Klinkhammer & Palmer, 1991; Wanty &Goldhaber, 1992; Calvert & Pedersen, 1993, 1996;Jones & Manning, 1994; Piper & Isaacs, 1995;Crusius et al., 1996; Tuttle et al., 1996). How-ever, if the relatively low-diversity fauna, coupledwith the trace element distribution, suggestssomewhat restricted conditions, no indicatorsof hypersalinity or con®nement were observed:neither evaporite crystals nor their pseudomorphs
Fig. 4. SEM images from unit 3 samples, illustratingcalci®ed bodies, interpreted to be fossil bacteria. Image(A) resembles a dividing bacterial cell (arrow); image(B) shows how numerous the bacterial bodies can be inthis facies. The white scale bar in the upper lefthandcorner of both pictures is 1 lm long.
Jurassic lagoonal environments 11891
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

Table 1. Geochemical contents of the samples studied.
Units Facies Sample
Si
(%)
Al
(%)
Fe
(%)
Mn
(lg g)1)
Ca
(%)
K
(%)
P
(lg g)1) Sr/Ca Cr/Al Mo/Al V/Al U/Al Ni/Al Co/Al Cu/Al Zn/Al Cd/Al Ba/Al Th/Al Zr/Al
3 Platy 1 1á93 0á61 0á43 149 36á33 0á20 188 16 23 1 25 1 26 3 5 37 0 29 1 24
limestones 2 2á14 0á79 0á51 128 35á54 0á32 223 13 19 3 26 1 24 2 4 19 0 22 1 17
3 2á08 0á65 0á44 131 35á78 0á26 201 15 21 2 26 1 26 3 5 22 0 28 2 20
2 Dark-coloured 4 ND 0á31 0á18 18 32á55 0á16 173 ND ND 77 203 ND 94 ND 19 45 2 187 ND ND
parallel 5 ND 0á18 0á11 13 30á53 0á11 131 ND ND 194 378 ND 206 ND 33 83 4 294 ND ND
laminae 6 ND 0á26 0á15 22 30á03 0á14 198 ND ND 112 254 ND 115 ND 27 100 4 492 ND ND
7 ND 0á17 0á09 19 31á34 0á10 199 ND ND 106 394 ND 182 ND 41 118 5 747 ND ND
8 ND 0á17 0á10 19 31á66 0á10 151 ND ND 94 359 ND 188 ND 47 100 4 541 ND ND
9 ND 0á51 0á34 30 32á69 0á26 158 ND ND 41 100 ND 67 ND 22 41 1 84 ND ND
Light-coloured 10 ND 0á37 0á18 36 33á95 0á18 209 ND ND 41 122 ND 43 ND 14 49 3 124 ND ND
parallel 11 ND 0á32 0á16 32 34á22 0á15 228 ND ND 50 141 ND 50 ND 16 34 BD 141 ND ND
laminae 12 ND 0á16 0á08 25 36á73 0á09 141 ND ND 38 200 ND 106 ND 38 75 BD 388 ND ND
13 ND 0á23 0á09 20 35á95 0á10 226 ND ND 17 178 ND 65 ND 26 70 3 152 ND ND
14 ND 0á32 0á18 24 35á40 0á16 93 ND ND 44 131 ND 63 ND 19 44 2 97 ND ND
Dark coloured 15 ND 0á34 0á18 33 34á92 0á18 264 ND ND 32 121 ND 74 ND 24 38 BD 324 ND ND
stromatolites 16 ND 0á23 0á12 15 34á59 0á12 118 ND ND 43 170 ND 96 ND 22 91 2 261 ND ND
17 ND 0á36 0á21 23 35á58 0á17 196 ND ND 42 178 ND 72 ND 14 50 1 233 ND ND
Light coloured 18 ND 0á02 0á01 9 38á52 0á01 162 ND ND 50 1800 ND BD ND BD 300 BD 3200 ND ND
stromatolites 19 ND 0á07 0á02 8 19á81 0á02 76 ND ND 14 186 ND 29 ND 29 129 9 443 ND ND
1 Massive 20 0á69 0á06 0á04 37 38á19 0á05 94 8 105 7 139 26 304 13 52 60 10 185 1 20
limestone 21 0á76 0á06 0á02 36 37á93 0á03 119 9 98 9 191 28 272 12 69 90 2 173 1 17
22 0á71 0á07 0á02 36 38á02 0á03 101 8 99 8 143 26 289 12 61 66 4 181 1 17
Average carbonate 2á4 0á42 0á38 1100 30á23 0á27 400 20 26 1 48 5 48 0 10 48 0 24 4 45
All trace element ratios quoted ´104.ND, not determined; BD, below detection threshold.Average carbonate from Turekian & Wedepohl (1961).
1190
N.
Trib
ovilla
rdet
al.
Ó1999
Inte
rnatio
nal
Asso
cia
tion
of
Sed
imen
tolo
gists,
Sed
imen
tolo
gy,
46,
1183±1197

were developed, nor were palaeoecological con-ditions favourable for organic matter preservationin large amounts.
After this episode of massive limestone deposi-tion, the lagoon was clearly isolated behind the reefbarrier. This led to the peculiar ecological condi-tions that controlled the development of theOrbagnoux Bituminous Laminites. This aspectwas discussed by Bernier (1984), Tribovillard et al.(1991, 1992), Mongenot et al. (1997) and Tribovil-lard (1998). New information about these laminitesconcerns their geochemical content. The traceelement distribution (high V/Al, Mo/Al, Cd/Al,Ni/Al, Co/Al, Cu/Al, Zn/Al, U/Al, Cr/Al and lowMn/Al; Table 1) attests to the fact that stronglyreducing conditions developed within the sedi-ment, probably below bio®lms that acted as abarrier between oxygenated bottom water aboveand euxinic sediments below (Tribovillard et al.,1992).
With the transition to unit 3, an episode ofrelatively more open marine conditions followed(possibly induced by subsidence and relativesea-level rise at the local/regional scale; Bernier,1984). Reducing conditions vanished, as shownby the trace element distribution (lowest V/Al,Mo/Al, Cd/Al, Ni/Al, Co/Al, Cu/Al, Zn/Al, U/Al,Cr/Al, and highest Mn/Al; Table 1) and theabsence of signi®cant amounts of organic matter.In addition, neither evaporite minerals norpseudomorphs were observed, in contrast tothe bituminous laminites. These apparentlymore open conditions were accompanied bythe supply of limited amounts of organic andinorganic terrestrially derived material (silicic-lastic minerals, spores and pollen, ligneousdebris). The small mass and ®ne grain size ofthe land-derived material suggest that the terres-trial supply may have been aeolian, rather than¯uvial.
However, what seems to correspond to there-establishment of normal marine conditionsduring the deposition of unit 3 was apparentlynot accompanied by any evidence of life, exceptdinocysts (which may have been brought in bywind or storm waves) and calci®ed bacteria. Inthe bituminous laminites below, many traces ofcyanobacterial activity were observed (endolithic®laments, sheath remains), and a cyanobacterialorigin has been proposed for the small-sized,equidimensional micritic grains of the carbonatematrix (Tribovillard, 1998). Nevertheless, indi-vidual bacterial bodies were not observed in thisfacies, whereas they are abundant in the platylimestones above. The apparent opening of
the depositional environment seems to be con-tradicted by the absence of larger life forms. Itmust therefore be postulated that somethingrendered the water column toxic. Unlike thebituminous laminites, it apparently cannot behypersalinity, as evaporite minerals or theirpseudomorphs are absent.
DISCUSSION
To explain the puzzling quasi-absence of life inthe lagoon during the deposition of unit 3, at leastthree, not necessarily mutually exclusive, hypo-theses may be formulated:
The ®rst hypothesis is that a bloom of dino¯a-gellates may have poisoned the lagoon water, as isobserved at the present day when the toxinsreleased during such blooms cause mass mortal-ities in the surface waters of some shelfal envi-ronments (`red tides'; Seibold & Berger, 1993).The organic parts of the dino¯agellates would nothave been preserved because of their labilitywhen no cysts are formed (indeed, even cysts arenot always fossilized).
A second hypothesis is that the chemistry ofthe water column was not favourable to life. Ourchemical data indicate an increase in the Sr/Caratio in unit 3 relative to units 1 and 2. This mighthave been linked with an evolution of the lagoonwater chemistry, but Sr distribution may bein¯uenced by other factors as well: the Sr budgetin the lagoon might have been modi®ed by thedecrease in Sr incorporation into aragonite shells,falling temperatures or a salinity increase (Morse& Mackenzie, 1990; Stoll & Schrag, 1998; Baudinet al., 19992 ). A marked rise in water alkalinity (i.e.the sum of the charges of all anions of weak acids,HCO3
) + CO32) + H(BO3))+ OH)) may have had
two consequences: ®rst, it may have preventedthe development of most forms of life, as observedtoday in hyperalkaline environments (Kempeet al., 1991; Kempe & Kazmierczak, 1994; Braith-waite & Zedef, 1996); secondly, it may havetriggered early calci®cation of bacteria, in vivo orshortly post mortem (Awramik, 1984; Kempe &Kazmierczak, 1990; Kempe, 1990; Riding, 1991;Kazmierczak et al., 1996), if the environmentbecame supersaturated with respect to calcite(early in situ calci®cation can operate only inconjunction with carbonate supersaturation andalkalinity, but is prevented by high PCO2).Although the hypothesis of the role of hyper-alkalinity is attractive, its development duringunit 3 deposition must be explained.
Jurassic lagoonal environments 11911
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

The conditions that led to the formation of theplaty limestones apparently did not exist previ-ously during the accumulation of the bituminouslaminites, which show relatively more diverseand abundant fossils, except within the undulat-ing laminae facies. Nevertheless, the platy lime-stones always lie just above the bituminouslaminites. The two main points that distinguishthe laminites, with respect to the platylimestones, are the presence of abundant, well-preserved (vulcanized) organic matter and theprominent in¯uence of cyanobacterial bio®lmsupon sedimentation (see above). These may beclues that help to decipher the chemical condi-tions that affected the palaeolagoon. In the case ofthe bituminous laminites, the bio®lms acted asbarriers and retained the chemical products ofsulphate reduction reaction in sediment pore-waters:
2CH2O� SO2ÿ4 ! 2HCOÿ3 �H� �HSÿ �1�
Moreover, in this case, sulphate reduction wasestablished in Fe-depleted sediments (Tribovil-lard et al., 1994), leading to a rise in the HCO3
)
concentration and a release of H+. These chemicalconditions were unfavourable to carbonate pre-cipitation (Curtis, 1987). Additionally, the abund-ant HS) induced the early vulcanization oforganic matter that shielded it against furtherdegradation (Tribovillard et al., 1994). Theammonite mass extinction layers recorded withinthe bituminous laminites may have resulted fromH2S and/or excess alkalinity intermittentlyescaping from the sediment into the shallowwater column.
In contrast, during the deposition of the platylimestones, no such bio®lm barrier existed, andiron was present, supplied with the land-derivedmaterial. In this case, sulphate reduction wasaccompanied by iron reduction (tiny iron sul-phide blebs are abundant in the limestone) andpyrite formation:
CH2O� 2Fe2O3 � 3H2O!HCOÿ3 � 4Fe2� � 7 OHÿ �2�
9CH2O� 4SO2ÿ4 � 4FeOOH!
9HCOÿ3 �H� � 4FeS2 � 6H2O �3�
15CH2O� 8SO2ÿ4 � 2Fe2O3 !
15HCOÿ3 �OHÿ � 4FeS2 � 7H2O �4�
This resulted in the waters being enriched inHCO3
) and their pH being buffered to highervalues, relative to the conditions that developedwith sulphate reduction alone. Additionally,ammonia released during organic matter degra-dation would have helped to raise the pH. Hence,the waters were more prone to carbonateprecipitation (Curtis, 1987; Morse & Mackenzie,1990). Additionally, HS) evolved as a sulphatereduction by-product would have been capturedby iron and formed iron sulphide, whichwould have prevented early organic matter vul-canization. As the organic matter was no longerprotected against bacterial degradation, it wasextensively mineralized. Thus, it may be hypoth-esized that the presence of iron had a double role:(1) it prevented vulcanization and indirectlycaused organic matter degradation; (2) it inducedthe porewater to be more alkaline and favouredcarbonate precipitation. Diffusion of reactionproducts upwards in the absence of a cyanobac-terial barrier increased the alkalinity, which may,in turn, have rendered the water column toxic forlarger organisms and triggered early calci®cationof bacteria (in vivo or shortly post mortem).
The ®rst two hypotheses are problematic. In the®rst case, no evidence of dino¯agellate bloomshas be seen; only frequent cysts are observed. Inthe second case, the hypothesis is based upon theassumption that sulphate and iron reductionreactions were intense, which cannot be con-®rmed: iron sulphide contents are not very high,and the trace element contents do not indicateabnormally reducing conditions.
A third hypothesis arises from a comparisonbetween the bituminous laminites of the palaeo-lagoon of Orbagnoux and French Polynesian atolllakes containing modern lacustrine to brackishstromatolites: the so-called kopara (Fig. 5; DeÂfargeet al., 1994, 1996).
French Polynesian stromatolites are ¯at, severaltens of centimetres thick and cover the bottom ofmost shallow (<2 m deep) lakes and ponds on therims of atolls in the Tuamotu Archipelago and theSociety Islands, as described extensively byDeÂfarge et al. (1994). The salinity of the water¯uctuates between fresh and fully marine.A millimetre-scale lamination develops in thesur®cial sediments, produced by an alternation oforganic-rich and CaCO3-rich laminae. Microbesare principally ®lamentous cyanobacteria, livingwithin the ®rst 3 cm below the sediment±waterinterface. They are accompanied by a few coccoidspecies and are underlain by living purple photo-synthetic bacteria (DeÂfarge et al., 1994). The
1192 N. Tribovillard et al.
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

underlying sediment, the organic fraction ofwhich consists mainly of the remains of extracel-lular polymer secretions of cyanobacteria andbacteria (i.e. sheaths and dispersed mucilage),hosts a vertical succession of living anaerobicbacteria. Sulphate reducers are abundant in thelayers underlying the purple layers, while metha-nogens occur at deeper levels. In most lakes, thesuper®cial stromatolite layers lie upon olderones, which have a higher CaCO3 content (71±86%). The carbonates forming the horizontalmineral laminae of the stromatolites may besubdivided on the basis of their shape and origin.Most are precipitated in situ and are associatedwith minor reef-derived grains and other bioclasts
(molluscan shells, foraminifera tests, echinodermdebris, etc.), deposited during sea-water incur-sions or carried by winds. The precipitates can besubdivided into true biominerals (formed throughencapsulation of living bacteria or sheath impreg-nation of living cyanobacteria), minerals formedby post mortem mineralization of cyanobacterialsheaths, organominerals (i.e. minerals precipi-tated under the in¯uence of sedimentary organicmatter) and peloids (DeÂfarge et al., 1996; Trichet& DeÂfarge, 1995, 1997).
A detailed microbiological comparisonbetween the palaeolagoon of Orbagnoux and thekopara lakes is outside the scope of this paper andwill be presented elsewhere. Nevertheless, it isalready possible to propose that the bituminouslaminites are a Jurassic counterpart of present-daykopara laminated deposits. As a consequence oftheir particular ecological situation at the surfaceof oceanic atoll rims, kopara lakes are almostdevoid of fauna. Shrimps and crabs, probablyfeeding on non-cyanobacterial organic remains,are present at the lake margin, but cause onlylimited bioturbation of the aerobic upper fewmillimetres of the deposits (DeÂfarge et al., 1994).Very few ®sh species (the `bone ®sh' Chanoschanos and Tilapia) can live in kopara lakes,feeding exclusively on the top green layer of thesediment. These ®sh produce 0á5- to 1-cm-sizedballs through the reworking of the top sedimentwhile grazing. However, the presence of ®sh inpresent-day lakes is not universal and resultsessentially from their introduction by man.
Thus, larger organisms may be absent, althoughthe waters are not poisoned or hyperalkaline; thefrequently hot temperature of the pond water mayalso play a role in their absence. This could alsoexplain the low abundance of life in the bitumi-nous laminites of Orbagnoux. A kopara is a site ofintense bio- and organomineralization of microbi-ally derived organic matter (DeÂfarge et al., 1996;Trichet & DeÂfarge, 1997). If the kopara is stirred andresuspended by a storm, the bio- or organocrystalsof calcium carbonate are separated from theorganic phase and settle again as a white, verypure and apparently abiotic micrite, which looksvery much like the facies of the platy limestones(abiotic must be taken in the sense that noidenti®able fossils are observed, apart frombacterial remains). Furthermore, the physicaldispersion of the organic fraction facilitates itsmineralization, as a result of the presence ofdiverse and abundant (cyano-) bacterial popula-tions that lead to organic matter consumption.Relatively thick micrite layers, with grains in the
Fig. 5. Section of a typical kopara deposit (in two coretubes of different diameter), Rangiroa atoll, lake R2.The thickness of the sedimentary pro®le is 30 cm.Three units can be distinguished: (1) at the very top ofthe sedimentary column, a 1- to 2-mm-thick greenlayer, hosting living cyanobacteria; (2) at about 18 cmdepth from the top, a greyish layer composed of storm-derived remobilized material (arrows); (3) reddish towhitish, more or less regularly laminated layers (whitewell-laminated layers are visible under the greyishlayer). The red colour is the result of abundant carot-enoidic pigments, produced by both the cyanobacteriaand the sulphur-oxidizing bacteria that develop withinthe organic matter deriving from cyanobacterial popu-lations. The whitish layers correspond to initially redlayers in which authigenic (low-Mg) calcite has pre-cipitated diagenetically within the sediment.
Jurassic lagoonal environments 11931
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

size range of 1 lm, may build up almost instanta-neously: the thickness of the carbonate deposit istotally unconnected with time. Lastly, because themicrite grains are minute and very well sorted, nosedimentary structures are observed in theremobilized kopara.
This or a similar process may have beenresponsible for the deposition of the platylimestones, which would then correspond toremobilized Jurassic koparas (or `palaeokopara').However, at Orbagnoux, the platy limestones areseveral metres thick and do not show therecurrence of laminites or `palaeokopara'.Furthermore, the faint lamination sometimesobserved within the platy limestones may corres-pond to attempts by (cyano-) bacteria to resettleafter the deposition of mobilized kopara micrite.This would indicate short pauses in the course ofplaty limestone accumulation. Consequently, theplaty limestone at Orbagnoux must haveaccumulated during repeated events, withoutresettlement of the `factory' of micritic raw mate-rial. In other words, places at which the palaeoko-para could have developed steadily would havesupplied resuspended material to the place wherethe platy limestones of Orbagnoux built up. Time-equivalent laminated carbonate facies identical tothe palaeokopara of Orbagnoux are common onthe Southern Jura platform. In many places, thisfacies occurs with limited vertical extension, butit is well developed at Armailles and Saint-Champ, close to Belley, and at Forens, close toBellegarde (Fig. 1), although outcrop conditionsare poor (Bernier, 1984). Beyond the reef, theslope was rather steep, and hemi-pelagic condi-tions were rapidly developed to the south-east.The time-equivalent deposits cropping out to theeast of Lake Annecy are hemi-pelagic alternationsintercalated with reworked bioclastic carbonatescoming down from the reef barrier (Doudouxet al., 1992). This strongly suggests that the reefbarrier was exposed to storms, the in¯uences ofwhich could have been felt behind the reef,within the lagoon. The Orbagnoux site waslocated less than 1 km from the reef, and it couldhave been supplied with storm-generated resus-pended palaeokopara material.
To summarize, at Orbagnoux, the palaeokoparais overlain by platy limestones that are interpretedas reworked palaeokopara deposits. Nevertheless,the development of palaeokopara in other pondsof the lagoon (Fig. 6) continued, being occasion-ally interrupted by storms remobilizing the pal-aeokopara. The pond of Orbagnoux must havebeen supplied with the resuspended micritic
material, recurrent in¯uxes of which preventedthe development of in situ palaeokopara.
Finally, the demise of the bituminous laminitesmay have been caused by a simple event: amoderate increase in the hydrodynamic energy ofthe local depositional environment at Orbagnoux.This local rise in energy may have originated fromvarious mechanisms, including change in lagoonmorphology or a decrease in the protectionoffered by the reef barrier, as a result of eitherrelative sea-level rise or a decline in the coral reef.
CONCLUSIONS
In this paper, emphasis has been placed on aparticular platy limestone facies belonging to theglobally regressive sequence of the Southern Jura,developed during Late Jurassic times. This faciescorresponds to a temporary, limited increase inwater depth (perhaps) and/or a return to lesscon®ned marine conditions (more likely). Thefacies is almost abiotic (i.e. largely devoid ofidenti®able body fossils). Three hypothesis areput forward to explain this peculiar feature. Thewater column of the palaeolagoon might havebeen poisoned by (i) toxic dino¯agellate recurrentblooms; or (ii) by hyperalkalinity, also favourableto early calci®cation of the bacteria; (iii) the Platy
Fig. 6. Palaeogeographic sketch illustrating in section(A, see approximate location on Fig. 1) and in per-spective (B) what the palaeokopara environment mayhave looked like. Open marine conditions (1) wereseparated by a reef barrier (2) from a very shallowplatform (3) with numerous individual ponds wherekopara could develop. To the NW, the platform wasagain somewhat deeper (4).
1194 N. Tribovillard et al.
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

Limestone Formation may result from the remo-bilization of kopara-like sediments, i.e. a peculiartype of ¯at stromatolites covering the bottom oflakes and ponds on atolls in French Polynesia.We favour the third hypothesis, as this demandsthe fewest possible number of assumptionsunsupported by our observations. Nevertheless,the hypotheses are not mutually exclusive. Ourpreferred model envisages microbes (cyanobacte-ria and bacteria) playing a very important role inthe direct or indirect precipitation of carbonates.Bacterially induced bio- or organomineralizationprocesses operating within the palaeokoparawould have supplied the carbonate mud for theformation of the platy limestone. The remobiliza-tion of the laminated material of the palaeoko-para, devoid of metazoans, would have resultedin the accumulation of abiotic platy limestones.This new genetic scheme for micritic, quasi-abiotic limestone formation underlines theimportance of microbes in sedimentary processes.
ACKNOWLEDGEMENTS
We thank M. Boquet, J. Carpentier, J.-P. Dorn, D.Malengro and P. Recourt for their technicalassistance. We are indebted to G. Gerdes,A. Strasser and I. Jarvis for their careful andconstructive review and editing, which has im-proved the manuscript considerably. We thankthem all.
REFERENCES
Awramik, S.M. (1984) Ancient stromatolites and micro-bial mats. In: Microbial Mats: Stromatolites (Ed. byY. Cohen, R.W. Castenholz and H.O. Halvorson), pp.1±22. Alan R. Liss, New York.
Baudin, F., Tribovillard, N., Laggoun-Defarge, F.,Lichtfouse, E., Monod, O. and Gardin, S. (1999)Depositional environment of a Kimmeridgiancarbonate `black band' (Akkuyu Fm., SW Turkey).Sedimentology, 46, 589±602.
Bernier, P. (1984) Les formations carbonateÂes du Kim-meÂridgien et du Portlandien dans le Jura meÂridional.Stratigraphie, micropaleÂontologie et seÂdimentologie.Doc. Lab. GeÂol. Lyon, 92. 803 pp.3
Bernier, P. and Courtinat, B. (1979) La matieÁre organi-que des calcaires d'arrieÁre-reÂcifs du KimmeÂridgiensupeÂrieur dans le Jura meÂridional. SysteÂmatique,conditions de geneÁse et d'environnement. Doc. Lab.GeÂol. Lyon, 75, 95±117.
Bernier, P., Gaillard, C., Barale, G., Bourseau, J.-P.,Buffetaud, E. and Wenz, S. (1991) Morphogenetic
impact of microbiological mats on surface structuresof Kimmeridgian micritic limestone (Cerin, France).Sedimentology, 38, 127±136.
Braithwaite, C.J.R. and Zedef, V. (1996)Hydromagnesite stromatolites and sediments in analkaline lake, Salda GoÈluÈ , Turkey. J. Sedim. Res.,66, 991±1002.
Breit, G.N. and Wanty, R.B. (1991) Vanadium accu-mulation in carbonaceous rocks: a review of geo-chemical controls during deposition and diagenesis.Chem. Geol., 91, 83±97.
Brumsack, H.J. (1986) The inorganic geochemistry ofCretaceous black shales (DSDP leg 41) in comparisonto modern upwelling sediments from the Gulf ofCalifornia. In: North Atlantic Palaeoceanography(Ed. by C.P. Summerhayes and N.J. Shackleton),Geol. Soc. London Spec. Publ., 21, 447±462.
Calvert, S.E. and Pedersen, T.F. (1993) Geochemistry ofRecent oxic and anoxic sediments: implications forthe geological record. Mar. Geol., 113, 67±88.
Calvert, S. and Pedersen, T. (1996) Sedimentary geo-chemistry of Mn: implications for the environment offormation of manganiferous black shales. Econ. Geol.,91, 36±47.
Combaz, A. (1980) Les keÂrogeÁnes vus au microscope.In: Kerogen (Ed. by B. Durand), pp. 55±111. Technip,Paris.
Courtinat, B. (1989) Les organoclastes des formationslithologiques du Malm dans le Jura meÂridional.SysteÂmatique, biostratigraphie and eÂleÂments d'inter-preÂtation paleÂoeÂcologique. Doc. Lab. GeÂol. Lyon, 1054 .361 pp.
Crusius, J., Calvert, S., Pedersen, T. and Sage, D. (1996)Rhenium and molybdenum enrichments in sedimentsas indicators of oxic, suboxic and sul®dic conditionsof deposition. Earth Planet. Sci. Lett., 145, 65±78.
Curtis, C. (1987) Mineralogical consequences of organicmatter degradation in sediments: inorganic-organicdiagenesis. In: Marine Clastic Sedimentology (Ed. byJ.K. Leggett and G.G. Zuffa), pp. 108±123. Grahamand Trotman, London.
DeÂfarge, C., Trichet, J., Maurin, A. and Hucher, M.(1994) Kopara in Polynesian atolls: early stages offormation of calcareous stromatolites. Sedim. Geol.,89, 9±23.
DeÂfarge, C., Trichet, J., Jaunet, A.-M., Robert, M., Trib-ble, J. and Sansone, F. (1996) Texture of microbialsediments revealed by cryo-scanning electronmicroscopy. J. Sedim. Res., 66, 935±947.
Doudoux, B., BarfeÂty, J.C., Carfantan, J.C., Tardy, M.and Nicoud, G. (1992) Notice explicative, feuilleAnnecy-Ugine (702) Carte geÂol. France (1:50 000),BRGM, OrleÂans.
Enay, R. and Hess, H. (1970) Nouveaux gisements aÁStelleroõÈdes dans le KimmeÂridgien supeÂrieur (Calc-aires en plaquettes) du Jura meÂridional, Ain, France.Eclogae Geol. Helv., 63, 1093±1107.
EspitalieÂ, J., Deroo, G. and Marquis, F. (1985) La pyr-olyse Rock Eval et ses applications. Rev. Inst. FrancË.PeÂtrol., 40, 563±579.
Folk, R.L. and Lynch, F.L. (1997) The possible role ofnannobacteria in clay-mineral diagenesis and the
Jurassic lagoonal environments 11951
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

importance of careful sample preparation in high-magni®cation study. J. Sedim. Res., 67, 583±589.
Gerdes, G., Krumbein, W.E. and Reineck, H.-E. (1991)Biolaminations ± Ecological versus depositionaldynamics. In: Cycles and Events in Stratigraphy(Ed. by G. Einsele, W. Ricken and A. Seilacher), pp.592±607. Springer, Berlin.
Gorin, G., GuÈ lacËar, F. and Cornioley, Y. (1989) Organicgeochemistry, maturity, palynofacies and paleoenvi-ronment of Upper Kimmeridgian and Lower Tertiaryorganic-rich samples in the southern Jura (Ain,France) and Subalpine massifs (Haute-Savoie,France). Eclogae Geol. Helv., 82, 491±515.
Gubler, Y. and Louis, M. (1956) Etude d'un certainmilieu du KimmeÂridgien bitumineux de l'est de laFrance. Rev. Inst. FrancË. PeÂtrol., 11, 1536±1543.
Holtzapfel, T. (1985) Les mineÂraux argileux: preÂpara-tion, analyse diffractomeÂtrique et deÂtermination.Ann. Soc. GeÂol. Nord, 125 . 199 pp.
Jones, B. and Manning, D.A.C. (1994) Comparison ofgeochemical indices used for the interpretation ofpaleoredox conditions in ancient mudstones. Chem.Geol., 114, 111±129.
Kazmierczak, J., Coleman, M.L., Gruszczynski, M. andKempe, S. (1996) Cyanobacterial key to the genesis ofmicritic and peloidal limestones in ancient seas.Acta Palaeont. Pol., 41, 319±338.
Kempe, S. (1990) Alkalinity: the link between anaero-bic basins and shallow water carbonates? Nat-urwissenschaften, 77, 426±427.
Kempe, S. and Kazmierczak, J. (1990) Calcium car-bonate supersaturation and the formation of in situcalci®ed stromatolites. In: Facets of Modern Biogeo-chemistry (Ed. by V. Ittekot, S. Kempe, W. Michaelisand A. Spitz), pp. 225±278. Springer, Berlin.
Kempe, S. and Kazmierczak, J. (1994) The role ofalkalinity in the evolution of ocean chemistry, orga-nization of living systems and biocalci®cation pro-cesses. In: Past and Present BiomineralisationProcesses. Considerations About Carbonate Cycle(Ed. by F. Doumenge), Bull. Inst. Ocean., Monaco,Spec. Publ., 13, 61±117.
Kempe, S., Kazmierczak, J., Landmann, G., Konuk, T.,Reiner, A. and Lipp, A. (1991) Largest knownmicrobialites discovered in Lake Van, Turkey.Nature, 349, 605±608.
Klinkhammer, G.P. and Palmer, M.R. (1991) Uraniumin the oceans: where it goes and why. Geochim.Cosmochim. Acta, 55, 1799±1806.
Mongenot, T. (1998) EÂtude PeÂtrographique et GeÂoc-himique D'un DeÂpoÃt SeÂdimentaire TreÁs Riche EnSoufre Organique (Orbagnoux, KimmeÂridgien Sup-eÂrieur). PhD Thesis, Orleans University.
Mongenot, T., Boussa®r, M., Derenne, S., Lallier-Verg-es, E.6 , Largeau, Cl. and Tribovillard, N.-P. (1997)Sulphur-rich organic matter from Bituminous Lam-inites of Orbagnoux (France, upper Kimmeridgian) ±the role of early vulcanization. Bull. Soc. GeÂol. Fr.,168, 331±341.
Mongenot, T., Derenne, S., Largeau, Cl., Tribovillard,N.-P., Lallier-Verges, E., Dessort, D. and Connan, J.(1999) Spectroscopic and pyrolytic studies of an
extremely sulphur-rich kerogen: implications forsource organism determination. Org. Geochem., 30,39±56.
Morse, J.W. and Mackenzie, F.T. (1990) Geochemistryof Sedimentary Carbonates. Developments in Sedi-mentology, 48, Elsevier, Amsterdam.
Piper, D.Z. and Isaacs, C.M. (1995) Geochemistry ofminor elements in the Monterey Fm, Cal. seawaterchemistry of deposition. US Geol. Surv. Prof. Papers,15667 . 41 pp.
Riche, A. (1904) Feuille de Lyon au 320 000eÁme. Bull.Carte geÂol. France, 13, 1±7.
Riding, R. (1991) Classi®cation of microbial carbonates.In: Calcareous Algae and Stromatolites (Ed. by R.Riding), pp. 21±51. Springer, Berlin.
Seibold, E. and Berger, W.H. (1993) The Sea Floor. AnIntroduction to Marine Geology, 2nd edn. Springer,Berlin.
Sinninghe DamsteÂ, J.S., Rijpstra, W.I.C., Kock-Van Da-len, A.C., Leeuw, J.W. and Schenck, P.A. (1989)Quenching of labile functionalisable lipids by inor-ganic sulfur species: evidence for the formation ofsedimentary organic sulfur compounds at the earlystages of diagenesis. Geochim. Cosmochim. Acta, 53,1343±1356.
Stoll, H.M. and Schrag, D.P. (1998) Effects of Quater-nary sea level cycles on strontium in seawater.Geochim. Cosmochim. Acta, 62, 1107±1118.
Strasser, A. (1988) Shallowing-upward sequences inPurbeckian peritidal carbonates. Sedimentology, 35,369±383.
Strasser, A. (1991) Lagoonal-peritidal sequences incarbonate environments: autocyclic and allocyclicprocesses. In: Cycles and Event Stratigraphy (Ed. byG. Einsele, W. Ricken and A. Seilacher), pp. 709±720.Springer, Berlin.
Tegelaar, E.W., de Leeuw, J.W., Derenne, S. andLargeau, C. (1989) A reappraisal of kerogen forma-tion. Geochim. Cosmochim. Acta, 53, 3103±3106.
Tribovillard, N. (1998) Mise en eÂvidence de peÂlod'origine cyanobacteÂrienne dans des seÂdiments car-bonateÂs lamineÂs riches en matieÁre organique duKimmeÂridgien du Jura. Ann. Soc. GeÂol. Nord, 6,35±42.
Tribovillard, N.P., Gorin, G., Belin, S., Hopfgartner, G.and Pichon, R. (1992) Organic-rich biolaminatedfacies from a Kimmeridgian lagoonal environment inFrench Southern Jura Mountains ± A way of esti-mating accumulation rate variations, Palaeogeogr.,Palaeoclimatol., Palaeoecol., 99, 163±177.
Tribovillard, N., Desprairies, A., Lallier-Verges, L. andBertrand, Ph. (1994) Sulfur incorporation of lipidicorganic matter in reactive-iron de®cient environ-ments: a possible enhancement for the storage ofhydrogen-rich organic matter. C.R. Acad. Sci. Paris,319, 1199±1206.
Trichet, J. and DeÂfarge, C. (1995) Non-biologicallysupported organomineralization. Bull. Inst. OceÂa-nogr. Monaco Spec. Publ., 14, 203±236.
Trichet, J. and DeÂfarge, C. (1997) High resolution cryo-scanning electron microscopy of modern stromato-lites (Lake R2, Rangiroa Atoll, Tuamotu Archipelago.
1196 N. Tribovillard et al.
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197

In: Biosedimentology of Microbial Buildups, IGCPProject No. 380 (Ed. by F. Neuweiler, J. Reitner andC. Monty), Facies, 36, 195±284.
Turekian, K.K. and Wedepohl, K.H. (1961) Distributionof the elements in some major units of the Earth'scrust. Geol. Soc. Am. Bull., 72, 175±191.
Tuttle, M.L., Klett, T.R., Richardson, M. and Breit, G.N.(1996) Geochemistry of two interbeds in the Pensyl-vanian Paradox Fm., Utah & Colorado Ð A record ofdeposition and diagenesis of repetitive cycles in amarine basin. US Geol. Surv. Bull., 2000 N8 . 86 pp.
Wanty, R.B. and Goldhaber, G. (1992) Thermodynamicsand kinetics of reactions involving vanadium innatural systems: accumulation of vanadium insedimentary rock. Geochim. Cosmochim. Acta, 56,1471±1183.
Manuscript received 12 October 1998;revision accepted 13 April 1999.
Jurassic lagoonal environments 11971
Ó 1999 International Association of Sedimentologists, Sedimentology, 46, 1183±1197
Copyright © 2022 FDOKUMEN




















