
Immunomodulatory properties of stem cells from human exfoliated deciduous teeth
Upload
irsp-ouidahCategory
view
2download
0
Journal of Tropical Ecologyhttp://journals.cambridge.org/TRO
Additional services for Journal of Tropical Ecology:
Email alerts: Click hereSubscriptions: Click hereCommercial reprints: Click hereTerms of use : Click here
Tree roots can penetrate deeply in African semi-deciduous rain forests:evidence from two common soil types
Vincent Freycon, Christelle Wonkam, Adeline Fayolle, Jean-Paul Laclau, Eric Lucot, Christophe Jourdan, Guillaume Cornuand Sylvie Gourlet-Fleury
Journal of Tropical Ecology / Volume 31 / Issue 01 / January 2015, pp 13 - 23DOI: 10.1017/S0266467414000595, Published online: 08 December 2014
Link to this article: http://journals.cambridge.org/abstract_S0266467414000595
How to cite this article:Vincent Freycon, Christelle Wonkam, Adeline Fayolle, Jean-Paul Laclau, Eric Lucot, Christophe Jourdan, Guillaume Cornuand Sylvie Gourlet-Fleury (2015). Tree roots can penetrate deeply in African semi-deciduous rain forests: evidence from twocommon soil types. Journal of Tropical Ecology, 31, pp 13-23 doi:10.1017/S0266467414000595
Request Permissions : Click here
Downloaded from http://journals.cambridge.org/TRO, IP address: 193.51.118.94 on 09 Dec 2014

Journal of Tropical Ecology (2015) 31:13–23. © Cambridge University Press 2014doi:10.1017/S0266467414000595
Tree roots can penetrate deeply in African semi-deciduous rain forests:evidence from two common soil types
Vincent Freycon∗,1, Christelle Wonkam∗,†,‡, Adeline Fayolle§,#, Jean-Paul Laclau∗∗, Eric Lucot††,Christophe Jourdan∗∗, Guillaume Cornu∗ and Sylvie Gourlet-Fleury∗
∗ CIRAD, UPR BSEF, F-34398 Montpellier, France† ISEM, UMR 5554, F-34095 Montpellier, France‡ Faculty of Science, University of Yaounde, ENS, P.O. Box 047, Yaounde, Cameroon§ Ministere des Eaux, Forets, Chasse et Peche, P.O. Box 3314, Bangui, Central African Republic# Gembloux Agro-Bio Tech, FORTROP, B-5030 Gembloux, Belgique∗∗ CIRAD, UMR Eco&Sols, F-34060 Montpellier, France†† UMR 6249 Chrono-Environnement, F-25030 Besancon, France
(Received 17 February 2014; revised 26 September 2014; accepted 26 September 2014)
Abstract: Despite the important functional role of deep roots in withdrawing water during drought, directmeasurements of root distribution are very rare in tropical rain forests. The aim of this study was to investigatethe root distribution of Entandrophragma cylindricum, a common tree species in the Central African semi-deciduousrain forest, in Ferralsols and Arenosols. We dug two pits to a depth of 6 m in Ferralsols and two pits to a depth of3 m in Arenosols, close to E. cylindricum trees. The vertical soil profiles were divided into 10 × 10-cm grid cells andthe roots counted were distributed in three diameter classes. We fitted a root distribution model to our dataset. Wefound that vertical root distribution was shallower in Arenosols than in Ferralsols. Root penetration was not stoppedeven by a Ferralsol with high gravel content in its subsoil. Overall, our measurements showed that 95% of all rootswere distributed to depths of between 258 and 564 cm from the soil surface, which is much deeper than the 95 cmdepth previously reported in the literature for tropical rain forests. As sampling depth could explain this discrepancy,we recommend a sampling depth of at least 3–5 m to accurately estimate root distribution. The drier the dry season,the deeper the sampling depth should be. Our results are consistent with global models of root distribution in forestecosystems, which are driven by climate variables. We thus suggest that deep rooting could be common in rain forestswith a marked dry season.
Key Words: Africa, Entandrophragma cylindricum, forest, gravel, root mat, rooting depth, soil texture, tropical, verticalroot distribution
INTRODUCTION
Tropical forests strongly influence regional climate byacting as a biotic pump (Makarieva & Gorshkov 2007,Spracklen et al. 2012). The considerable amount of waterpumped from the soil to the atmosphere is of particularimportance in the dry season in Amazonia, when mosttranspiration by trees comes from water withdrawn fromdeep soil layers or from the water table (Bruno et al. 2006,Fan & Miguez-Macho 2010, Kleidon & Heimann 2000,Nepstad et al. 1994). Soil-vegetation-climate models arethus sensitive to the distribution of tree roots in deep
1 Corresponding author. Email: [email protected]
soil layers in tropical regions (Akkermans et al. 2012,Nepstad et al. 2004). Root distribution can be measureddirectly in the field, or estimated indirectly using primaryproduction data and soil-vegetation-climate models (Ichiiet al. 2007, Kleidon & Heimann 2000, Schenk & Jackson2005). Direct measurements of deep roots are still rarein natural tropical forests. Only in three studies out ofthe 26 listed by Schenk & Jackson (2002) were rootssampled a depth of at least 3 m (Cerri & Volkoff 1987,Nepstad et al. 1994, Sternberg et al. 1998). To ourknowledge, only one additional measurement of deeprooting (Ohnuki et al. 2008) has been made since then(Table 1). Many tropical root profiles are thus sampledtoo shallowly to enable accurate estimation of the 95%rooting depth, D95 (Schenk & Jackson 2002). The values

14 VINCENT FREYCON ET AL.
Table 1. Quantitative studies of vertical root distribution in tropicalforests with sampling soil depths � 3 m. Sternberg et al. (1998) studiedroot distribution with a root auger.
Sampling Number ofSource Geographic location depth (m) pits
Cerri & Volkoff 1987 Manaus, Brazil, Amazonia 5 1Nepstad et al. 1994 Para, Brazil, Amazonia 5.8 1Ohnuki et al. 2008 Cambodia, Asia 9.4 1Sternberg et al. 1998 Para, Brazil, Amazonia 4 0Our study CAR, Central Africa 3 or 6 4
of rooting depth and maximum rooting depth in tropicalforests are still the subject of debate. Schenk & Jackson(2002) reported that 50% and 95% of the roots of trees intropical semi-deciduous, deciduous and evergreen forestsare generally found in the upper 16 cm and 95 cm of thesoil, respectively. However, Ohnuki et al. (2008) showedthat roots can penetrate much deeper. The maximumrooting depth of trees in tropical semi-deciduous forests,which was deduced from a single study in China, was 3.7± 0.5 m (Canadell et al. 1996), but Nepstad et al. (1994)reported a maximum rooting depth of more than 10 m inan Amazonian rain forest.
Meta-analysis and global models predict that rootdistribution and maximum rooting depth are mainlyaffected by climate variables and soil characteristics (Ichiiet al. 2007, Schenk & Jackson 2002, 2005). Firstly,Schenk & Jackson (2005) predicted that tropical forestsare most likely to be deep-rooted in strongly seasonalsemi-deciduous or in evergreen forests. A soil waterbalance model confirmed that root distributions tend tobe as shallow as possible and as deep as needed to fulfilevapotranspiration demands (Schenk 2008). Ichii et al.(2007) estimated maximum rooting depth to be 1–3, 3–5 and 5–10 m in Amazonian regions with a 0–2-mo,3–4-mo and 5–6-mo dry season, respectively. Secondly,Schenk & Jackson (2002) showed that root distributionin tropical evergreen forests was shallower in sandy soilsthan in soils with a finer texture. Canadell et al. (1996)cited some examples in which roots reached great depthseven in rocky soils, and by penetrating through hardpans. However, overall, global models have been validatedusing only very few deep field measurements, none ofwhich were made in Central Africa which is home tolarge areas of semi-deciduous tropical forests.
Given the scarcity of data, some of which are biased andcontroversial, the aim of our study was to investigate rootdistributions of a common tree species Entandrophragmacylindricum Sprague (Meliaceae), in two common soiltypes, Ferralsols and Arenosols, in a semi-deciduousrain forest with a dry season in Central Africa. Wehypothesized that (1) root distribution is deeper thanthat frequently reported in the literature from fieldmeasurements, i.e. 50% rooting depth > 16 cm, 95%
rooting depth > 95 cm, and the maximum rooting depth> 3.7 m; (2) rooting depth is shallower in Arenosols thanin Ferrasols. Our study, which provides data from fieldmeasurements made in an understudied part of the world,can be used to validate global models which predict rootdistribution and maximum rooting depth.
METHODS
Study sites
The study was carried out in the Central African Republic(CAR), at two sites located 80 km apart, one close to thetown of Mbaiki (3°52′30′′N, 17°59′00′′E) and the otherclose to Ngotto (4°01′30′′N, 17°20′00′′E) (Figure 1).Mbaiki and Ngotto have a similar topographical position,top of a plateau, at an altitude of 560 m and 630 m asl,respectively. In CAR, this topographic position accountsfor a large part of the semi-deciduous rain forest (Boulvert1996). The climate at both sites is humid-tropical withan average annual rainfall of 1740 mm (1982–2007),a dry season lasting 3–5 mo (November/December–February/March, with less than 100 mm precipitation)and a mean temperature of 24.9 °C (1981–1989)(Boukoko meteorological station, A. Ougou, pers. comm.).The vegetation at both sites is semi-deciduous rain forest ofthe Guineo-Congolian region which covers about 43 000km2 in CAR. The floristic composition differed slightlybetween the two study sites: the number of evergreen taxawas higher in the Ngotto forest than in the Mbaiki forest(Gond et al. 2013, Rejou-Mechain et al. 2008). At Mbaiki,the canopy was dominated by species from the Malvaceae,Meliaceae, Ulmaceae and Sapotaceae (Boulvert 1986),whereas at Ngotto, the canopy was dominated by speciesfrom the Caesalpiniaceae, Sapotaceae, Meliaceae andMoraceae (Yongo 2003). Finally, at a global scale, theclimate and the vegetation at both sites belongs to thebioclimatic type that Blasco et al. (2000) called warmhumid lowland forests, i.e. 1000–2000 mm mean annualrainfall, 1–4 dry months and a mean temperature in thecoldest month > 20 °C. This bioclimatic type is foundin a large part of Central Africa, the south-eastern andthe southern part of Amazonia, and a small part of Asia.The soils and the geology also differed at the two sites(Table 2). At Mbaiki, the soils are Ferralsols developed onPrecambrian quartzite-sandstone, whereas at Ngotto thesoils are Arenosols developed on Cretaceous sandstone(Boulvert 1996; Figure 1). At Mbaiki, the Ferralsols maycontain a gravel horizon at a depth of between 1 m and 3m, followed by an iron duricrust and/or weathered rocks,whereas such horizons are not present in the Arenosolsof Ngotto down to a depth of at least 3 m (Boulvert1983). Ferralsols are very common in rain forests inCentral Africa and Amazonia, whereas Arenosols are only

Root distribution in African rain forests 15
Figure 1. Location of the two study sites, Mbaiki and Ngotto, in Central African Republic (CAR). The colours correspond to the soilmapping units of the Harmonized World Soil Database Viewer, version 1.2 (http://webarchive.iiasa.ac.at/Research/LUC/External-World-soil-database/HTML/): dark pink, Ferralsol; light pink, Arenosol.
Table 2. Characteristics of the study sites. Dry-season length was defined by the number of months with rainfall � 100 mm mo−1.The monthly rainfall data were taken from MeteoSat 2000–2008 and from Boukoko Meteorological Station (1982–2007). Soil typeWRB classification was determined according to Boulvert (1983) and the Harmonized World Soil Database Viewer, version 1.2(http://webarchive.iiasa.ac.at/Research/LUC/External-World-soil-database/HTML/).
Dry-season Geological substrate Soil type WRBSite Profile Coordinates length Biome (Boulvert 1996) classification
Mbaiki Mbaiki 1 3°52′39′ ′N 17°54′20′ ′E 3–5 Tropical semi-deciduous forest PrecambrianQuartzite-sandstone
Ferralsol
Mbaiki 2 3°53′08′ ′N 17°54′13′ ′ENgotto Ngotto 1 4°01′15′ ′N 17°10′19′ ′E 3–5 Tropical semi-deciduous forest with
many evergreen taxaCretaceous Sandstone Arenosol
Ngotto 2 4°01′40′ ′N 17°09′36′ ′E
common in rain forests in Central Africa (Driessen et al.2001).
Root sampling
We dug four pits (130–240 cm in length, 150 cm inwidth, 300–600 cm in depth), two per site, between Apriland June 2011. At each site, the two pits were located1–1.5 km apart, on top of a plateau. We studied one sideof each pit. The centre of this side was 50 cm away froman Entandrophragma cylindricum tree, a species of highcommercial value commonly found in the tropical semi-deciduous forest in the Guineo-Congolian zone, whichincludes the Central African rain forest. It is distributedfrom Ivory Coast to Uganda (Gerdat 1974, Lebrun & Stork2011). The location of each pit was carefully chosen
to make sure that, on the one hand, the selected E.cylindricum specimens had similar diameters at breastheight (44 ± 6 cm) and, on the other hand, that thesize of neighbouring trees was similar (i.e. the basal areawithin 3 m of the specimen selected was 7 ± 4 m2 ha−1).The two pits in the Ferralsols (Mbaiki 1, Mbaiki 2) weredug to a depth of 6 m. Athough we planned to dig the twopits in the Arenosols (Ngotto 1, Ngotto 2) down to thesame depth, we were obliged to stop digging at a depth of3 m, due to technical difficulties caused by the sandy soils(the pit collapsed). We determined the root distributionprofile inside each trench using the root intersect countingmethod (Laclau et al. 2013, Tardieu 1988). A 1.50 × 1-mgrid subdivided into grid cells measuring 10 × 10 cm wasplaced vertically against the pit wall and the root intersectswere counted individually. Roots with a diameter greaterthan 1 mm were measured with callipers. Root intersects

16 VINCENT FREYCON ET AL.
were counted at 1-m intervals as the pits was dug, so thatthe roots would not dry out. Roots of E. cylindricum treescan be identified by their particular scent (they smell likeroses). Although we did not distinguish each individualE. cylindricum root from the roots of neighbouring treespecies, observations showed that the majority of rootscounted belonged to E. cylindricum trees.
At Mbaiki 2 (Ferralsol) and Ngotto 2 (Arenosol), soilcores were sampled from the bottom of each pit using aroot auger. The soil layers sampled were 600–620 cm,630–650 cm, 680–700 cm, 700–720 cm, 730–750 cm,780–800 cm at Mbaiki 2, and 300–320 cm, 330–350cm, 380–400 cm, 400–420 cm, 430–450 cm, 480–500cm, 500–520 cm at Ngotto 2. All the roots were placedin a 550-μm-mesh sieve and washed free of soil with tapwater. The volume of soil sampled in each layer was 4710cm3. Rooting depth was observed down to a depth of 8 min the Ferralsol at Mbaiki 2 and to a depth of 5.20 m inthe Arenosol at Ngotto 2.
Each soil profile was characterized. Soil samplescollected in the topsoil, at depths of 0–20 cm or 10–20cm, and samples collected in deep layers, at depths of 100and 300 cm were the subject of standard physical andchemical analyses at Cirad laboratories in Montpellier,France.
Data analysis
We classified the roots according to their diameter (d): fineroots (d < 2 mm); medium roots (2 � d < 10 mm); andcoarse roots (d � 10 mm). In the coarse-root class, we didnot distinguish subclasses (e.g. structural roots) becauseonly one to three roots>50 mm in diameter were countedin each soil profile.
In the two topsoil profile at the Ngotto site, we observeda dense root mat, whose thickness ranged from 1 to8.5 cm. Since it was impossible to count these rootsindividually, for each grid cell of 10 × 10 cm, from the soilsurface to the depth of 10 cm, we estimated the number ofroot intersects in the root mat, Nmat, using the equation:
Nmat = Nbelow mat × Tmat/(10 − Tmat)
where Nbelow_mat is the number of root intersects below theroot mat and Tmat is the thickness of the root mat (cm).
We counted the number of fine, medium and coarseroots in each layer of the soil profiles we sampled (150cm wide and 10 cm deep). We calculated the absoluteroot intersect density (RID, expressed in roots m−2), therelative RID, which represents the percentage of rootsat the depth considered relative to the total number ofroots counted in the soil profile, and the cumulative rootfraction, which represents the percentage of roots fromthe soil surface to the depth considered relative to thetotal number of roots counted in the soil profile. RIDs were
calculated to the maximum depth of the soil profile (6 mfor the Ferralsol and 3 m for the Arenosol) and relativedensities were calculated down to a depth of 3 m, so thatwe were able to compare our two study sites with eachother, as well as with the meta-analysis as per Schenk &Jackson (2002).
For each profile, we estimated the two parameters, D50
and c, of a logistic model proposed by Schenk & Jackson(2002):
Y = 1/(1 + (D/D50)c) (1)
where Y is the cumulative root fraction, D is depth, D50 isthe depth above which 50% of the roots were present andc is a shape parameter. The value of D95, the depth abovewhich 95% of roots are present, was predicted a posteriori,after estimating the two parameters of the model, D50 andc, using the formula:
D95 = D50 × (0.05/0.95)1/c (2)
We also estimated the parameters of the modelproduced by Schenk & Jackson (2002) for the datasetpublished in previous studies carried out at least to 3 m indepth in evergreen tropical forests (Cerri & Volkov 1987,Nepstad et al. 1994, Ohnuki et al. 2008) or in a tropicalEucalyptus plantation (Christina et al. 2011).
The consequences of sampling depth for the estimationsof the parameters in Eq. (1) and Eq. (2) for fine rootswere assessed in the two Ferralsols, which were sampleddown to a depth of 6 m. The complete dataset in each soilprofile (i.e. down to 6 m) was compared with five datasetssimulating the observation of pit walls down to depths of1, 2, 3, 4 or 5 m.
RESULTS
Soil properties
The description of the four soil profiles and the physicaland chemical analyses confirmed that the two soil typeswere typical of Ferralsols and Arenosols. Both Ferralsolprofiles had a ferralic horizon below a depth of 40 cm,with a micro-aggregated structure, and a weathered rocklayer at a depth of 4 m (Figure 2). A Bsk skeletic horizon(i.e. 40% or more of the volume occupied by gravels)then a Bpp petroplinthic horizon (i.e. iron duricrust) wereencountered between the depths of 2 m and 4 m in theMbaiki 1 profile. The texture differed slightly between thetwo Ferralsol profiles. Mbaiki 1 had a sandy clay loam tovery gravelly sandy loam texture whereas Mbaiki 2 hada sandy clay to clay texture (Table 3). No root mats werefound in the two Ferralsols.
Both Arenosol profiles had a root mat made of high-density fine and medium roots in their topsoil layer. Themat was between 3 and 8.5 cm thick at Ngotto 1 and

Root distribution in African rain forests 17
Figure 2. Total root intersect density and diagram of four soil profiles in CAR, two Ferralsols, Mbaiki 1 and Mbaiki 2, and two Arenosols, Ngotto 1and Ngotto 2. The majority of roots belonged to Entandrophragma cylindricum. Designations for horizons: A, mineral topsoil horizon; B, mineral deepsoil horizon; C, weathered rock (i.e. saprolite); ar, arenic (i.e. having a texture class of sand); fl, ferralic; pp, petroplinthic (i.e. iron duricrust); sk,skeletic (i.e. 40% or more of the volume occupied by gravels).
Table 3. Physical and chemical properties of the four profiles. CEC, cation exchange capacity; EB, exchangeable bases ( = Ca2+ + Mg2+ + K+ +Na+); BS, base saturation ( = EB/CEC × 100). Designation for horizons: A, mineral topsoil horizon; B, mineral deep soil horizon; ar, arenic (i.e.having a texture class of sand); fl, ferralic; sk, skeletic (i.e. 40% or more of the volume occupied by gravels).
(cmolc kg−1)
Depth Clay Silt Sand C N P Bray2 BSProfile Horizon (cm) (%) (%) (%) pH (%) (‰) C/N (mg kg−1) CEC EB Al (%)
Mbaiki 1 A 10–20 28 10 62 4.5 1.20 0.89 13 2.8 6.4 0.99 1.4 16Bfl 100 33 10 57 4.7 0.14 0.11 13 1.3 3.9 0.07 1.1 2Bsk 300 18 13 69 5.0 0.41 0.35 12 0.6 5.3 0.10 0.9 2
Mbaiki 2 A 0–20 39 8 53 4.3 1.57 1.42 11 2.3 7.6 0.80 2.1 11Bfl 100 47 8 45 4.9 0.27 0.34 8 1.2 4.3 0.28 0.8 7Bfl 300 51 13 36 4.8 0.13 0.21 6 1.1 7.0 0.06 2.0 1
Ngotto 1 AB 10–20 8 1 91 4.3 0.66 0.54 12 4.6 2.1 0.25 0.3 12B1ar 100 8 0 92 4.6 0.20 0.19 11 4.2 0.9 0.04 0.4 4B2ar 280 9 0 91 4.7 0.04 0.08 6 4.2 0.5 0.05 0.3 10
Ngotto 2 AB 0–20 5 1 94 3.7 1.09 0.76 14 6.0 2.4 0.12 0.5 5B1ar 80 7 0 92 4.5 0.19 0.17 12 3.1 1.0 0.02 0.6 2B2ar 300 7 1 93 4.9 0.05 0.08 6 4.3 0.5 0.04 0.1 8
between 1 and 6 cm thick at Ngotto 2. Beneath thisroot mat, the soil horizons were characterized by a sandytexture and a weakly developed structure down to a depthof 50 cm and absent below this depth.
The two soil types also differed in their chemicalproperties (Table 3). The Ferralsols had higher CEC, sum
of exchangeable bases and exchangeable aluminiumconcentrations than the Arenosols. The Ferralsols alsohad a higher total N concentration at a depth of 300cm than the Arenosols – 0.29‰ on average versus0.08‰. Conversely, the Ferralsols had lower availableP concentrations than the Arenosols, regardless of depths
18 VINCENT FREYCON ET AL.
Table 4. Estimated values of c, D50 (Equation 1) and predicted values of D95 (Equation 2) in the Schenk & Jackson (2002) model for root distributionsin our study, compared with those computed from other studies, with a sampling depth fixed at 3 m. RID, root intersect density (unit: number ofroots per unit surface area).
Model parameters
Studies Biome Soil Root trait measured c D50 D95
Our study Semi-deciduous Ferralsol Mbaiki 1 RID −1.642 94 564Semi-deciduous Ferralsol Mbaiki 2 RID −1.390 58 482Semi-deciduous Arenosol Ngotto 1 RID −1.331 28 258Semi-deciduous Arenosol Ngotto 2 RID −1.237 37 397
Other studiesCerri & Volkoff 1987 Evergreen Ferralsol Biomass −0.927 7 172Nepstad et al. 1994 Evergreen Ferralsol Biomass −0.608 0.4 51Christina et al. 2011 Eucalyptus plants Ferralsol Biomass −0.882 9 249Ohnuki et al. 2008 Evergreen Acrisols RID −1.394 65 534Meta-analysisSchenk & Jackson 2002 Semi-deciduous and
deciduousArenosol, Cambisol,
Ferralsol, Mollisol, . . .Biomass (11 studies) or
RID (1 study)−1.681 16 95
Schenk & Jackson 2002 Evergreen Acrisol, Arenosol, Ferralsol,Inceptisol, Podzol, . . .
Biomass (18), length(4) or RID (15)
−1.632 15 91
– e.g. 0.9 mg kg−1 on average versus 4.3 mg kg−1 at adepth of 300 cm.
Vertical root distribution
In all the soil profiles, RIDs, including all root intersects,decreased with depth (Figure 2). The sharpest decreasewas found in the Ngotto 1 Arenosol, where root densitydecreased from 3427 roots m−2 in the 0–10 cm soillayer to 33 roots m−2 in the 290–300-cm soil layer. Thedecrease was smoother in the Mbaiki 1 Ferralsol, with adecrease from 1293 roots in the upper soil layer down to353 roots m−2 in the 290–300-cm soil layer. In this soilprofile, the RID remained high between the depths of 2 mand 4 m, which corresponded to the Bsk skeletic horizon.
We estimated D50 between a depth of 28 and 94 cm,and predicted D95 between a depth of 258 and 564 cm,depending on the soil profiles. The Ferralsols had higherD50 and D95 values than the Arenosols – on average 76cm versus 33 cm for D50, and 523 cm versus 328 cm forD95 (Table 4).
The vertical distribution of total roots was similar to thatof the fine roots since the latter represented the majorityof the roots we counted (Figure 3). Fine roots penetrateddeep layers whereas coarse roots were mainly superficial.We found no coarse roots at a depth below 430 cm, 230cm, 120 cm and 110 cm in Mbaiki 1, Mbaiki 2, Ngotto 1and Ngotto 2, respectively.
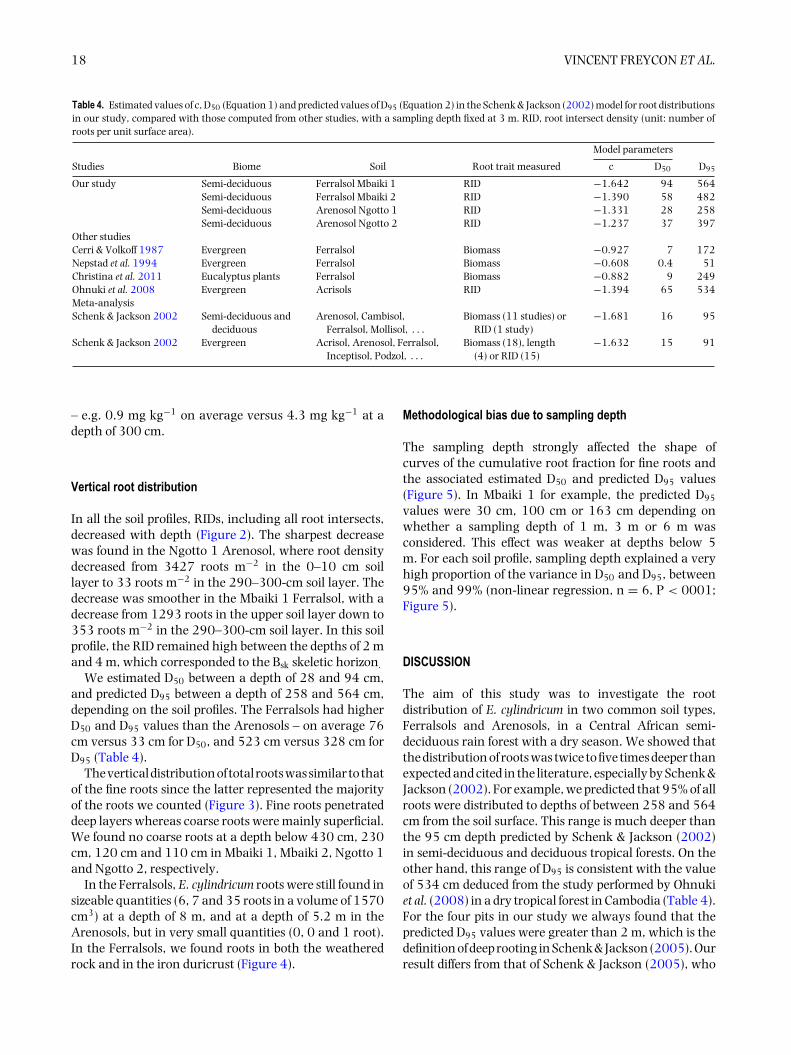
In the Ferralsols, E. cylindricum roots were still found insizeable quantities (6, 7 and 35 roots in a volume of 1570cm3) at a depth of 8 m, and at a depth of 5.2 m in theArenosols, but in very small quantities (0, 0 and 1 root).In the Ferralsols, we found roots in both the weatheredrock and in the iron duricrust (Figure 4).
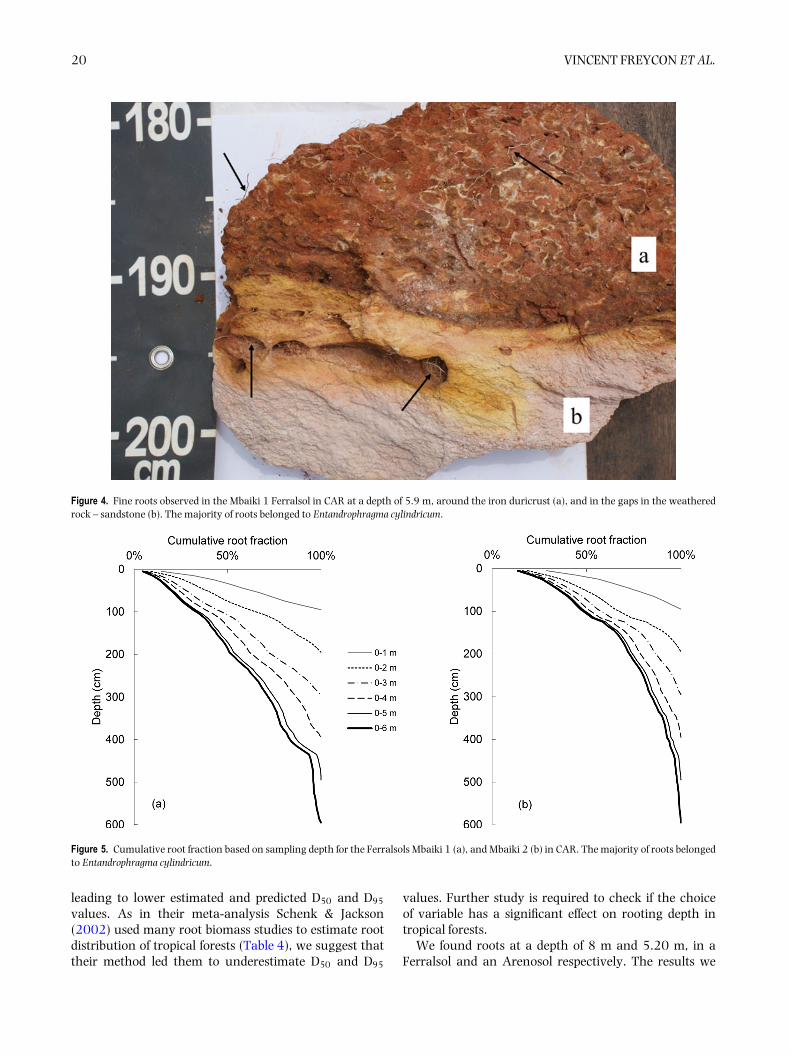
Methodological bias due to sampling depth
The sampling depth strongly affected the shape ofcurves of the cumulative root fraction for fine roots andthe associated estimated D50 and predicted D95 values(Figure 5). In Mbaiki 1 for example, the predicted D95
values were 30 cm, 100 cm or 163 cm depending onwhether a sampling depth of 1 m, 3 m or 6 m wasconsidered. This effect was weaker at depths below 5m. For each soil profile, sampling depth explained a veryhigh proportion of the variance in D50 and D95, between95% and 99% (non-linear regression, n = 6, P < 0001;Figure 5).
DISCUSSION
The aim of this study was to investigate the rootdistribution of E. cylindricum in two common soil types,Ferralsols and Arenosols, in a Central African semi-deciduous rain forest with a dry season. We showed thatthe distribution of roots was twice to five times deeper thanexpected and cited in the literature, especially by Schenk &Jackson (2002). For example, we predicted that 95% of allroots were distributed to depths of between 258 and 564cm from the soil surface. This range is much deeper thanthe 95 cm depth predicted by Schenk & Jackson (2002)in semi-deciduous and deciduous tropical forests. On theother hand, this range of D95 is consistent with the valueof 534 cm deduced from the study performed by Ohnukiet al. (2008) in a dry tropical forest in Cambodia (Table 4).For the four pits in our study we always found that thepredicted D95 values were greater than 2 m, which is thedefinition of deep rooting in Schenk & Jackson (2005). Ourresult differs from that of Schenk & Jackson (2005), who

Root distribution in African rain forests 19
Figure 3. Cumulative root fraction based on depth for total roots (a); fine roots, diameter < 2 mm (b); medium roots, 2 � diameter < 10 mm (c);and coarse roots, diameter � 10 mm (d) of four profile soils in CAR, two Ferralsols, Mbaiki 1 and Mbaiki 2, and two Arenosols, Ngotto 1 and Ngotto2. The majority of roots belonged to Entandrophragma cylindricum. The tropical deciduous forest curve is derived from the Schenk & Jackson (2002)model of semi-deciduous and deciduous forests.
predicted that the probability of deep rooting in our studyarea was only 0.27 in Ferralsols, and 0.18 in Arenosols.
A methodological bias and the choice of the roottraits considered could explain these differences betweenobserved and expected root distribution by Schenk &Jackson (2002, 2005). Firstly, the differences could havebeen the result of a methodological bias due to thesampling depth. Indeed, we showed that the values of D50
and D95 strongly depended on sampling depth (Figure 5).Our results confirmed in semi-deciduous tropical forestsa methodological bias suggested by Schenk & Jackson(2002) in the estimates of rooting depths in all typesof tropical forest because of too shallow sampling depth.Indeed, our results suggest that the values of D50 and D95
proposed by Schenk & Jackson (2002, 2005) for tropicalforests are greatly underestimated: their meta-analysisand empirical model was based on studies in which
the sampling depth was generally less than 1 m and amaximum depth of 3 m. We recommend a sampling depthof at least 5 m and 3 m to enable more reliable estimatesand prediction of D50 and D95 for semi-deciduous tropicalforests with 3–5-mo dry season, and other rain forestswith a less marked dry season, respectively.
Secondly, these differences could also be the resultof the variable we chose to quantify root distribution,i.e. root intersect density. Our results are in fact similarto those deduced in the study by Ohnuki et al. (2008)who also measured root intersect density. This variableis mainly affected by fine roots, the relative proportion ofwhich increases with depth (Schenk & Jackson 2002). Bycontrast, our results differ from those of Christina et al.(2011), Cerri & Volkoff (1987) and Nepstad et al. (1994),who measured root biomass. This variable is mainlyaffected by coarse roots, which are generally superficial,
20 VINCENT FREYCON ET AL.
Figure 4. Fine roots observed in the Mbaiki 1 Ferralsol in CAR at a depth of 5.9 m, around the iron duricrust (a), and in the gaps in the weatheredrock – sandstone (b). The majority of roots belonged to Entandrophragma cylindricum.
Figure 5. Cumulative root fraction based on sampling depth for the Ferralsols Mbaiki 1 (a), and Mbaiki 2 (b) in CAR. The majority of roots belongedto Entandrophragma cylindricum.
leading to lower estimated and predicted D50 and D95
values. As in their meta-analysis Schenk & Jackson(2002) used many root biomass studies to estimate rootdistribution of tropical forests (Table 4), we suggest thattheir method led them to underestimate D50 and D95
values. Further study is required to check if the choiceof variable has a significant effect on rooting depth intropical forests.
We found roots at a depth of 8 m and 5.20 m, in aFerralsol and an Arenosol respectively. The results we

Root distribution in African rain forests 21
obtained in an African semi-deciduous rain forest with a3–5-mo dry season, are consistent with those of Ichii et al.(2007), who estimated maximum rooting depth to be 3–5m and 5–10 m in Amazonian regions with a 3–4-mo anda 5–6-mo dry season, respectively. In the present study,we were unable to reach the maximum rooting depth and,given the number of fine roots still found in our deepestsamples, we suggest that, in Ferralsols, roots extend downto at least 10 m. Similar rooting depths were found byOhnuki et al. (2008) in a dry tropical evergreen foreston Acrisols in Cambodia, and in eucalypt plantations, byLaclau et al. (2001) on Arenosols in the Congo Republic.As all these results were obtained in the same bioclimatictype, i.e. warm humid lowland forests (Blasco et al. 2000),taken together, these results suggest that a maximumrooting depth around 10 m might be shared by tropicalforest trees of this bioclimatic type that cover a large partof Central Africa, part of Amazonia, and a small part ofAsia. At a local scale, it is likely that these results will needto be adjusted depending on the topographical position.
A rooting depth of 10 m could provide trees with alarge enough soil water reserve to survive the dry season,as already shown in Amazonia (Nepstad et al. 1994,2004). In the Ferralsols at Mbaiki, the roots could reachthe weathered rocks, giving them access to an additionalreserve of water stored in the pores of the rocks (Ohnukiet al. 2008, Ruiz et al. 2010). Deep rooting could alsoenable the roots to reach the water table, whose depth isestimated to range from 8 to 25 m (Bonsor & McDonald2011) in this region of the Central African Republic.Christina et al. (2011) showed in Brazil that eucalypt rootsreached the water table at a depth of about 16 m as earlyas 3.5 y after planting. Such rooting depths suggest thatforests growing on Ferralsols and Arenosols in CentralAfrica could be resilient to periods of drought.
We found that the vertical root distribution wasshallower in Arenosols than in Ferralsols. This isconsistent with the results of Schenk & Jackson (2002)in evergreen tropical forests, implying that their resultscan be extrapolated to semi-deciduous forests. This is alsoconsistent with the results of Schenk & Jackson (2005)who predicted that the probability of deep rooting islower in coarse-textured soils (e.g. Arenosols) than in fine-textured soils (e.g. the Ferralsols in our study). This can bepartly explained by the presence of a root mat in the topsoilof the Arenosols, which was not present in the Ferralsols.This type of root mat has also been observed in sandysoils elsewhere in the tropics, in Venezuela, Congo andMalaysia (Baillie et al. 2006, Kingsbury & Kellman 1997,Laclau et al. 2004). It has been hypothesized that the rootmat favours direct recycling of nutrients from organicmatter in chemically poor soils (Laclau et al. 2004, Sayeret al. 2006, Vitousek & Sanford 1986). However, thisresult was also partially derived from the large number ofroots found between depths of 2 m and 4 m in one of the
two Ferralsols, in a skeletic horizon, rich in gravels. Thisabundance of roots could be associated with the presenceof gravels, in agreement with the results of Magier &Ravina (1984), who observed the same patterns in peachtrees. This is also consistent with the results of Ohnukiet al. (2008) who observed a peak in the number of rootsat a depth of 8 m in a horizon rich in iron nodules. Theunderlying mechanism could be that root contact withan obstacle, such as a gravel or an iron nodule, favoursthe production of secondary lateral roots more frequentlythan when no obstacles are encountered (Lucot 1994,Riedacker 1978). The phenomenon is probably fairlycommon in the forests of the flat tropical areas of CentralAfrica and Amazonia since their soils often contain oneor more horizons that are rich in gravels and stones,sometimes known as stone-lines (Lucas et al. 1990,Segalen 1969).
Finally, we found that the roots were not stopped bythe gravelly or stony horizons present in the Ferralsol.We found roots even inside the weathered rock, after theyhad penetrated a horizon rich in gravel followed by an ironduricrust. These observations are consistent with those ofCanadell et al. (1996) showing that roots reached greatdepths, even in rocky soils, and penetrated hard pans.In a study conducted at the forest-savanna boundary inthe Central African Republic, Beauvais (2009) suggestedthat tree roots play a role in the disaggregation of the ironduricrust to create horizons rich in gravel.
Concluding remarks
This study showed that the distribution of Entandro-phragma cylindricum roots, a common tree in the semi-deciduous rain forest of Central Africa with 3–5-modry season, penetrates deeper than previously thought,whatever the soil type. Our results suggest that rootdistribution has been underestimated up to now, due toboth a sampling bias – the pits were not deep enough– and to a measurement bias – total biomass of theroots was measured rather than fine-root density. Werecommend that in future rooting studies dealing withroot distribution in rain forests, soils should be sampleddown to depths of at least 3–5 m for accurate predictionof D95. The drier the dry season, the deeper the samplingdepth should be. We also recommend that the climate-soil-vegetation models and models of soil water reserveuse D95 values of 2–5 m and maximum rooting depthsof 10 m for semi-deciduous tropical forests in CentralAfrica. Our results suggest that Central African forestsmay be resilient to current dry seasons partly becausethey have access to water reserves in the very deep layersof the soil. As our results are consistent with globalmodels of root distribution in forest ecosystems drivenby climate variables and especially the length of the dry

22 VINCENT FREYCON ET AL.
season, we suggest that deep rooting could be commonin rain forests with a marked dry season. At a localscale, other factors, like topographic position and soilproperties impeding root growth (e.g. superficial bedrock,water table) need to be taken into consideration. Thisstudy, which was performed on a common species in therain forest of Central Africa, now needs to be extendedto other species because tree species in the same biomecan differ in rooting depth (Holdo & Timberlake 2008).Simultaneous monitoring of soil water content up to theedge of the rooting system and tree transpiration wouldbe particularly relevant for assessing the role of deep soilhorizons in providing water to trees throughout the year.
ACKNOWLEDGEMENTS
This research is part of the CoForChange project(www.coforchange.eu). It was funded by the ERA-NetBiodivERsA, with the national funders ANR (France)and NERC (UK), part of the 2008 BiodivERsA call forresearch proposals. We thank the ARF Project (Appuia la Recherche Forestiere) and its seven partners:AFD (Agence Francaise de Developpement), CIRAD(Centre de Cooperation Internationale en RechercheAgronomique pour le Developpement), ICRA (InstitutCentrafricain de Recherche Agronomique), MEFCP(Ministere centrafricain des Eaux, Forets, Chasse andPeche), SCAC/MAE (Service de Cooperation and d’ActionsCulturelles), University of Bangui and SCAD (SocieteCentrafricaine de Deroulage) for providing access to theM’Baıki. We thank IFB (Industrie Forestiere de Batalimo)for providing access to the Ngotto site. Christelle Wonkamwas funded by AFD. We thank the two teams in thefield who dug the pits at Mbaiki and Ngotto. We thankS. Guitet, J. Schenk and another anonymous reviewerfor their insightful comments on a former version of themanuscript.
LITERATURE CITED
AKKERMANS, T., LAUWAET, D., DEMUZERE, M., VOGEL, G.,
NOUVELLON, Y., ARDO, J., CAQUET, B., DE GRANDCOURT, A.,
MERBOLD, L., KUTSCH, W. & VAN LIPZIG, N. 2012. Validation
and comparison of two soil-vegetation-atmosphere transfer models
for tropical Africa. Journal of Geophysical Research – Biogeosciences
117:1–15.
BAILLIE, I. C., ASHTON, P. S., CHIN, S. P., DAVIES, S. J., PALMIOTTO,
P. A., RUSSO, S. E. & TAN, S. 2006. Spatial associations of humus,
nutrients and soils in mixed dipterocarp forest at Lambir, Sarawak,
Malaysian Borneo. Journal of Tropical Ecology 22:543–553.
BEAUVAIS, A. 2009. Ferricrete biochemical degradation on the
rainforest-savannas boundary of Central African Republic. Geoderma
150:379–388.
BLASCO, F., WHITMORE, T. C. & GERS, C. 2000. A framework for the
worldwide comparison of tropical woody vegetation types. Biological
Conservation 95:175–189.
BONSOR, H. C. & MCDONALD, A. M. 2011. An initial estimate of depth
to groundwater across Africa. British Geological Survey, Nottingham.
26 pp.
BOULVERT, Y. 1983. Carte pedologique de la republique centrafricaine a
1 : 1 000 000. ORSTOM, Paris. 126 pp.
BOULVERT, Y. 1986. Carte phytogeographique de la Republique
Centrafricaine (Feuille Ouest-Feuille Est) a 1 : 1 000 000. ORSTOM,
Paris. 131 pp.
BOULVERT, Y. 1996. Etude geomorphologique de la republique
centrafricaine. Carte a 1 : 1 000 000 en deux feuilles Ouest et Est. Notice
explicative n°110. ORSTOM Editions, Paris. 258 pp.
BRUNO, R. D., DA ROCHA, H. R., DE FREITAS, H. C., GOULDEN, M. L. &
MILLER, S. D. 2006. Soil moisture dynamics in an eastern Amazonian
tropical forest. Hydrological Processes 20:2477–2489.
CANADELL, J., JACKSON, R. B., EHLERINGER, J. B., MOONEY, H. A.,
SALA, O. E. & SCHULZE, E. D. 1996. Maximum rooting depth of
vegetation types at the global scale. Oecologia 108:583–595.
CERRI, C. C. & VOLKOFF, B. 1987. Carbon content in a yellow latosol
of Central Amazon rain forest. Acta Oecologica–Oecologia Generalis
8:29–42.
CHRISTINA, M., LACLAU, J. P., GONCALVES, J. L. M., JOURDAN,
C., NOUVELLON, Y. & BOUILLET, J. P. 2011. Almost symmetrical
vertical growth rates above and below ground in one of the world’s
most productive forests. Ecosphere 2:1–10.
DRIESSEN, P., DECKERS, J., SPAARGAREN, O. & NACHTERGAELE, F.
2001. Lecture notes on the major soils of the world. Food and Agriculture
Organization, Rome. 334 pp.
FAN, Y. & MIGUEZ-MACHO, G. 2010. Potential groundwater
contribution to Amazon evapotranspiration. Hydrology and Earth
System Sciences 14:2039–2056.
GERDAT. 1974. Sapelli. Bois et Forets des Tropiques 154:27–40.
GOND, V., FAYOLLE, A., PENNEC, A., CORNU, G., MAYAUX, P.,
CAMBERLIN, P., DOUMENGE, C., FAUVET, N. & GOURLET-FLEURY,
S. 2013. Vegetation structure and greenness in Central Africa from
Modis multi-temporal data. Philosophical Transactions of the Royal
Society B 368: 20120309.
HOLDO, R. M. & TIMBERLAKE, J. 2008. Rooting depth and above-
ground community composition in Kalahari sand woodlands in
western Zimbabwe. Journal of Tropical Ecology 24:169–176.
ICHII, K., HASHIMOTO, H., WHITE, M. A., POTTERS, C., HUTYRA,
L. R., HUETE, A. R., MYNENI, R. B. & NEMANIS, R. R. 2007.
Constraining rooting depths in tropical rainforests using satellite data
and ecosystem modeling for accurate simulation of gross primary
production seasonality. Global Change Biology 13:67–77.
KINGSBURY, N. & KELLMAN, M. 1997. Root mat depths and surface
soil chemistry in southeastern Venezuela. Journal of Tropical Ecology
13:475–479.
KLEIDON, A. & HEIMANN, M. 2000. Assessing the role of
deep rooted vegetation in the climate system with model
simulations: mechanism, comparison to observations and
implications for Amazonian deforestation. Climate Dynamics 16:183–
199.

Root distribution in African rain forests 23
LACLAU, J. P., ARNAUD, M., BOUILLET, J. P. & RANGER, J. 2001.
Spatial distribution of Eucalyptus roots in a deep sandy soil in the
Congo: relationships with the ability of the stand to take up water
and nutrients. Tree Physiology 21:129–136.
LACLAU, J. P., TOUTAIN, F., M’BOU, A. T., ARNAUD, M., JOFFRE,
R. & RANGER, J. 2004. The function of the superficial root mat
in the biogeochemical cycles of nutrients in Congolese Eucalyptus
plantations. Annals of Botany 93:249–261.
LACLAU, J. P., SILVA, E. A., LAMBAIS, G., BERNOUX, M., LE MAIRE,
G., STAPE, J. L., BOUILLET, J. P., GONCALVES, J. L. M., JOURDAN, C.
& NOUVELLON, Y. 2013. Dynamics of soil exploration by fine roots
down to a depth of 10 m in Eucalyptus grandis plantations. Frontiers
in Plant Science 4:1–12.
LEBRUN, J.-P. & STORK, A. L. 2011. Tropical African flowering plants.
Ecology and distribution. Vol. 6. Burseraceae-Apiaceae. Addendum to
volumes 1–5 (Families A–C). Conservatoire et Jardin Botanique de la
ville de Geneve, Geneve. 398 pp.
LUCAS, Y., BOULET, R. & CHAUVEL, A. 1990. In situ genesis of stone
lines – Demonstrative example from a lateritic cover in Brazilian
Amazonia. Comptes Rendus Academie des Sciences, Serie II 311:713–
718.
LUCOT, E. 1994. Influence des caracteristiques de la pierrosite des sols sur la
prospection racinaire et l’alimentation hydrique des arbres. Application a
l’estimation de la valeur des sols forestiers. Universite de Franche-Comte,
Besancon. 101 pp.
MAGIER, J. & RAVINA, I. 1984. Rock fragments and soil depth as factors
in land evaluation of terra rossa. Pp. 99–103 in Nichols, J. D., Brown,
P. L. & Grant, W. J. (eds.). Erosion and productivity of soils containing rock
fragments. Special Publication 13. Soil Science Society of America,
Madison.
MAKARIEVA, A. M. & GORSHKOV, V. G. 2007. Biotic pump of
atmospheric moisture as driver of the hydrological cycle on land.
Hydrology and Earth System Sciences 11:1013–1033.
NEPSTAD, D. C., DE CARVALHO, C. R., DAVIDSON, E. A., JIPP, P. H.,
LEFEBVRE, P. A., NEGREIROS, G. H., DA SILVA, E. D., STONE, T. A.,
TRUMBORE, S. E. & VIEIRA, S. 1994. The role of deep roots in the
hydrological and carbon cycles of Amazonian forests and pastures.
Nature 372:666–669.
NEPSTAD, D., LEFEBVRE, P., DA SILVA, U. L., TOMASELLA, J.,
SCHLESINGER, P., SOLORZANO, L., MOUTINHO, P., RAY, D. &
BENITO, J. G. 2004. Amazon drought and its implications for forest
flammability and tree growth: a basin-wide analysis. Global Change
Biology 10:704–717.
OHNUKI, Y., SHIMIZU, A., CHANN, S., TORIYAMA, J., KIMHEAN, C.
& ARAKI, M. 2008. Seasonal change in thick regolith hardness and
water content in a dry evergreen forest in Kampong Thom Province,
Cambodia. Geoderma 146:94–101.
REJOU-MECHAIN, M., PELISSIER, R., GOURLET-FLEURY, S.,
COUTERON, P., NASI, R. & THOMPSON, J. D. 2008. Regional
variation in tropical forest tree species composition in the Central
African Republic: an assessment based on inventories by forest
companies. Journal of Tropical Ecology 24:663–674.
RIEDACKER, A. 1978. Study of deviation of horizontal or oblique roots of
poplar cutting encountering an obstacle – Applications on container
design. Annales des Sciences Forestieres 35:1–18.
RUIZ, L., VARMA, M. R. R., KUMAR, M. S. M., SEKHAR, M., MARECHAL,
J.-C., DESCLOITRES, M., RIOTTE, J., KUMAR, S., KUMAR, C. &
BRAUN, J.-J. 2010. Water balance modelling in a tropical watershed
under deciduous forest (Mule Hole, India): Regolith matric storage
buffers the groundwater recharge process. Journal of Hydrology
380:460–472.
SAYER, E. J., TANNER, E. V. J. & CHEESMAN, A. W. 2006. Increased
litterfall changes fine root distribution in a moist tropical forest. Plant
and Soil 281:5–13.
SCHENK, H. J. 2008. The shallowest possible water extraction profile: a
null model for global root distributions. Vadose Zone Journal 7:1119–
1124.
SCHENK, H. J. & JACKSON, R. B. 2002. The global biogeography of
roots. Ecological Monographs 72:311–328.
SCHENK, H. J. & JACKSON, R. B. 2005. Mapping the global distribution
of deep roots in relation to climate and soil characteristics. Geoderma
126:129–140.
SEGALEN, P. 1969. Le remaniement des sols et la mise en place de la
stone-line en Afrique. Cahiers ORSTOM, serie Pedolologique 7:113–
131.
SPRACKLEN, D. V., ARNOLD, S. R. & TAYLOR, C. M. 2012. Observations
of increased tropical rainfall preceded by air passage over forests.
Nature 489:282–286.
STERNBERG, L. D. L., GREEN, L., MOREIRA, M. Z., NEPSTAD, D.,
MARTINELLI, L. A. & VICTORIA, R. 1998. Root distribution in an
Amazonian seasonal forest as derived from delta C-13 profiles. Plant
and Soil 205:45–50.
TARDIEU, F. 1988. Analysis of the spatial variability of maize root
density. 1. Effect of wheel compaction on the spatial arrangement
of roots. Plant and Soil 107:259–266.
VITOUSEK, P. M. & SANFORD, R. L. 1986. Nutrient cycling in moist
tropical forest. Annual Review of Ecology and Systematics 17:137–167.
YONGO, O. 2003. Contribution aux etudes floristiques, phytogeogra-
phique et phytosociologiques de la foret de Ngotto (Republique
centrafricaine). Acta Botanica Gallica 150:119–124.
Copyright © 2022 FDOKUMEN




















