
Joining the dots: Production, processing and targeting of U snRNP to nuclear bodies
8
Review Joining the dots: Production, processing and targeting of U snRNP to nuclear bodies Debra J. Shaw a , Paul Eggleton b , Philip J. Young a,b, ⁎ a Clinical Neurobiology, Institute of Biomedical and Clinical Science, St Luke's Campus, Peninsula Medical School, Exeter, EX1 2LU, UK b Inflammation and Musculoskeletal Disease Research, Institute of Biomedical and Clinical Science, St Luke's Campus, Peninsula Medical School, Exeter, EX1 2LU, UK abstract article info Article history: Received 8 May 2008 Received in revised form 22 July 2008 Accepted 23 July 2008 Available online 6 August 2008 Keywords: U snRNP Nuclear export Sm core proteins Nuclear import sequences Survival motor neuron (SMN) Cajal bodies Nucleoli Interchromatin granule clusters (IGC) The spliceosome is the RNP complex than catalyses the removal of introns from the Uridine-rich small nuclear ribonucleoproteins (U snRNPs) that make up the main components of this complex. The production of these RNPs is an intricate process, involving several key stages. These include: 1) the transcription of the U snRNAs; 2) their nuclear export; 3) the cytoplasmic assembly of the U snRNP; 4) their nuclear import; 5) their processing within Cajal bodies and the nucleolus; and 6) their storage in interchromatin granule clusters (IGCs). This review focuses on each of these stages, discussing the key complexes involved as well as the trafficking and targeting mechanisms involved. © 2008 Elsevier B.V. All rights reserved. RNA splicing is the process of removing introns from a transcribed heterogeneous nuclear RNA (hnRNA) (pre-mRNA) molecule [1]. This process is accomplished by the spliceosome, a large (40s) RNA/protein macromolecular machine [1]. The spliceo- some is made up of 5 major building units termed U1, U2, U4, U5 and U6 small nuclear ribonucleoproteins (snRNP) [1]. These U snRNPs are made up to a complex of RNA and proteins, containing U snRNA (Uridine-rich small nuclear RNA), Sm proteins (7 core proteins; Sm B or B′, D1, D2, D3, E, F and G) that are common to all snRNPs and several other proteins that are specific for each U snRNP [1]. The assembly of the spliceosome is a complex process that requires shuttling between the nucleus and the cytoplasm (Fig. 1) [1]. Initially the U snRNAs are synthesised in the nucleus, then are required to move to the cytoplasm for assembly into U snRNPs before returning to the nucleus where they mature in Cajal bodies and nucleoli and then transfer to splicing interchromatin granule clusters (IGCs) and perichromatin fibrils (PFs) to participate in splicing (Fig. 1) [2]. In this review we will discuss the production of these U snRNPs, including their transcription, nuclear export, cytoplasmic assembly, nuclear import and sub-nuclear targeting. 1. U snRNA transcription U1, U2, U4, and U5 snRNAs are synthesised in the nucleus by RNA polymerase II. 7-methylguanosine (monomethyl-cap; mG) cap struc- tures are added co-transcriptionally to all RNA polymerase II transcripts, which acts as one of the signals for nuclear export [3]. U3 snRNA is also transcribed by RNA polymerase II and participates in rRNA processing in the nucleus [4] and belongs to a group of three snoRNAs (U3, U8 and U13) [5]. U6 is transcribed by RNA polymerase III [6]. The mG cap structures are involved in several processes including RNA stability [7–10], pre-mRNA splicing [3,11–15], translation initia- tion [16,17], and RNA export from the nucleus [18–20]. U1, U2, U4 and U5 snRNA are all transcribed by RNA polymerase II and so contain this cap structure, which is required for their export from the nucleus to the cytoplasm across the nuclear membrane where they are required for U snRNP assembly. 2. Architecture of the nuclear membrane The nuclear membrane represents the interface between the nucleus and cytoplasm, and is essential for maintaining the unique identity of each compartment [21,22]. The membrane is composed of three main components, the lamina, the nuclear membranes and the nuclear pore complexes (NPCs). Each NPC is composed of building blocks of ∼ 30 different proteins called nucleoporins (nups), which are assembled to form sub-complexes within the NPC [23,21]. Analysis of Biochimica et Biophysica Acta 1783 (2008) 2137–2144 ⁎ Corresponding author. Fax: +44 1392 262926. E-mail address: [email protected] (P.J. Young). 0167-4889/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.bbamcr.2008.07.025 Contents lists available at ScienceDirect Biochimica et Biophysica Acta journal homepage: www.elsevier.com/locate/bbamcr
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Joining the dots: Production, processing and targeting of U snRNP to nuclear bodies

Biochimica et Biophysica Acta 1783 (2008) 2137–2144
Contents lists available at ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r.com/ locate /bbamcr
Review
Joining the dots: Production, processing and targeting of U snRNP to nuclear bodies
Debra J. Shaw a, Paul Eggleton b, Philip J. Young a,b,⁎a Clinical Neurobiology, Institute of Biomedical and Clinical Science, St Luke's Campus, Peninsula Medical School, Exeter, EX1 2LU, UKb Inflammation and Musculoskeletal Disease Research, Institute of Biomedical and Clinical Science, St Luke's Campus, Peninsula Medical School, Exeter, EX1 2LU, UK
⁎ Corresponding author. Fax: +44 1392 262926.E-mail address: [email protected] (P.J. Young)
0167-4889/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.bbamcr.2008.07.025
a b s t r a c t
a r t i c l e i n f oArticle history:
The spliceosome is the RN Received 8 May 2008Received in revised form 22 July 2008Accepted 23 July 2008Available online 6 August 2008Keywords:U snRNPNuclear exportSm core proteinsNuclear import sequencesSurvival motor neuron (SMN)Cajal bodiesNucleoliInterchromatin granule clusters (IGC)
P complex than catalyses the removal of introns from the Uridine-rich smallnuclear ribonucleoproteins (U snRNPs) that make up the main components of this complex. The productionof these RNPs is an intricate process, involving several key stages. These include: 1) the transcription of the UsnRNAs; 2) their nuclear export; 3) the cytoplasmic assembly of the U snRNP; 4) their nuclear import; 5) theirprocessing within Cajal bodies and the nucleolus; and 6) their storage in interchromatin granule clusters(IGCs). This review focuses on each of these stages, discussing the key complexes involved as well as thetrafficking and targeting mechanisms involved.
© 2008 Elsevier B.V. All rights reserved.
RNA splicing is the process of removing introns from atranscribed heterogeneous nuclear RNA (hnRNA) (pre-mRNA)molecule [1]. This process is accomplished by the spliceosome, alarge (40s) RNA/protein macromolecular machine [1]. The spliceo-some is made up of 5 major building units termed U1, U2, U4, U5and U6 small nuclear ribonucleoproteins (snRNP) [1]. These UsnRNPs are made up to a complex of RNA and proteins, containingU snRNA (Uridine-rich small nuclear RNA), Sm proteins (7 coreproteins; Sm B or B′, D1, D2, D3, E, F and G) that are common to allsnRNPs and several other proteins that are specific for each UsnRNP [1]. The assembly of the spliceosome is a complex processthat requires shuttling between the nucleus and the cytoplasm (Fig.1) [1]. Initially the U snRNAs are synthesised in the nucleus, thenare required to move to the cytoplasm for assembly into U snRNPsbefore returning to the nucleus where they mature in Cajal bodiesand nucleoli and then transfer to splicing interchromatin granuleclusters (IGCs) and perichromatin fibrils (PFs) to participate insplicing (Fig. 1) [2]. In this review we will discuss the production ofthese U snRNPs, including their transcription, nuclear export,cytoplasmic assembly, nuclear import and sub-nuclear targeting.
.
l rights reserved.
1. U snRNA transcription
U1, U2, U4, and U5 snRNAs are synthesised in the nucleus by RNApolymerase II. 7-methylguanosine (monomethyl-cap; mG) cap struc-tures are added co-transcriptionally to all RNA polymerase IItranscripts, which acts as one of the signals for nuclear export [3].U3 snRNA is also transcribed by RNA polymerase II and participates inrRNA processing in the nucleus [4] and belongs to a group of threesnoRNAs (U3, U8 and U13) [5]. U6 is transcribed by RNA polymerase III[6]. The mG cap structures are involved in several processes includingRNA stability [7–10], pre-mRNA splicing [3,11–15], translation initia-tion [16,17], and RNA export from the nucleus [18–20]. U1, U2, U4 andU5 snRNA are all transcribed by RNA polymerase II and so contain thiscap structure, which is required for their export from the nucleus tothe cytoplasm across the nuclear membrane where they are requiredfor U snRNP assembly.
2. Architecture of the nuclear membrane
The nuclear membrane represents the interface between thenucleus and cytoplasm, and is essential for maintaining the uniqueidentity of each compartment [21,22]. The membrane is composed ofthree main components, the lamina, the nuclear membranes and thenuclear pore complexes (NPCs). Each NPC is composed of buildingblocks of ∼30 different proteins called nucleoporins (nups), which areassembled to form sub-complexes within the NPC [23,21]. Analysis of

Fig. 1. Schematic overview of the production and processing of U snRNPs. Newly transcribed U snRNA containing a 5′ monomethyl-guanosine (mG) cap (1) is actively transportedthrough the nuclear pore complex (2) into the cytoplasm. Here the Sm core proteins are assembled onto the U snRNA (3) to form the U snRNP. Once the Sm core assembly has takenplace the 5′ guanosine cap is methylated to form a trimethylated guanosine cap (m3G) (4). The m3G and Sm core complex function as a bipartite signal and mediate the nuclearimport of the U snRNP (5).
2138 D.J. Shaw et al. / Biochimica et Biophysica Acta 1783 (2008) 2137–2144
NPC's and nucleocytoplasmic transport, at the molecular level,remains difficult due to the complex nature of the NPC and limitationsin current experimental approaches [23].
The nuclear lamina is essential for nuclear integrity and iscomposed of intermediate filaments made up of lamins and lamin-associated proteins. These proteins include the lamin-associatedprotein (LAP) 1 and 2 families, the lamin B receptor (LBR), andEmerin [24,21]. In vertebrate somatic cells, this is a 10–20 nm thickprotein meshwork, underlying the INM, and is composed primarilyof A and B type lamins [25]. Lamins are members of the IntermediateFilament (IF) family, and feature a coiled coil flanked by non-helicaltail and head domains [22]. There are a group of genetic disordersassociated with mutations in lamin proteins and lamin-bindingproteins termed laminopathies or the more generic term nuclearenvelopathies. The most common nuclear envelopathy with thehighest frequency in human populations is Emery–Dreifuss musculardystrophy, caused by an X-linked mutation in the EMD gene codingfor emerin, a lamin-binding protein and affects an estimated 1 in100,000 people [26].
The nuclear membranes are two morphologically different, butinterconnected domains named the outer nuclear membrane (ONM)and the inner nuclear membrane (INM). The ONM is continuouswith the rough endoplasmic reticulum (rER), has ribosomesattached [21,27,28], and provides attachment sites for structuralelements of the cytoplasm [21,29,30]. The INM contains a unique setof proteins that are involved in binding for lamina and chromatin[21,31–33]. The ONM and INM are separated by the perinuclearspace (PNS), which is a regular gap of 30–50 nm, and an extension
of the ER lumen [22]. Junctions between the ONM and INM formchannels capable of mediating selective exchange of macromo-lecules. These proteins are called the nuclear pore complexes (NPC)[22].
3. The nuclear pore complex (NPC)
The NPC are pore-like discontinuities within the nuclear envelope,and are the siteswhere nucleocytoplasmic exchange occurs [34,35,28].NPCs are made up of large proteinaceous assemblies (nucleoporins) ofapproximately 120 MDa and are able to selectively transport cargoesacross the nuclear envelope [23]. The NPC acts almost like a sieve,allowing smallmolecules and ions todiffuse through theNPC, howeverthe transport of large macromolecular assemblies (e.g. proteins, RNAsand RNPs) is dependent on their recognition by transport factors,which are able to interact with the NPC and carry their cargos acrossthe nuclear envelope [22,36–38]. Fig. 2 shows a schematic representa-tion of the NPC. The vertebrate NPC has a tripartite architecture,consisting of a central framework that is decorated by cytoplasmicfilaments and a nuclear basket (Fig. 2) [39]. The three dimensionalstructure of the NPC represents an eightfold rotational symmetryperpendicular to the nuclear membrane [40].
To pass through the NPC, transport receptors are required to bindto FG nucleoporin (nups) docking sites to trigger channel gating [41].FG nups contain repeating amino acid sequences, predominantly FG,GLFG or FxFG, separated by hydrophilic and charged amino acidlinker sequences [42]. FG nups are highly disordered and flexiblewith little secondary structure, and form a mesh work of random
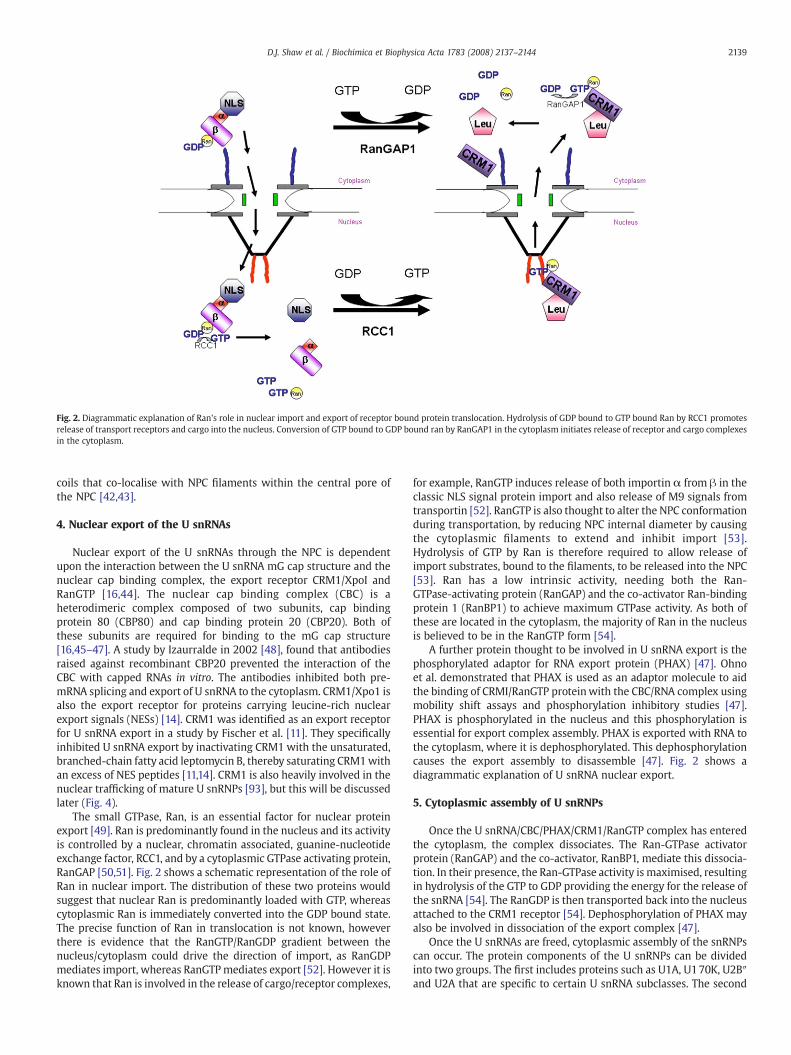
Fig. 2. Diagrammatic explanation of Ran's role in nuclear import and export of receptor bound protein translocation. Hydrolysis of GDP bound to GTP bound Ran by RCC1 promotesrelease of transport receptors and cargo into the nucleus. Conversion of GTP bound to GDP bound ran by RanGAP1 in the cytoplasm initiates release of receptor and cargo complexesin the cytoplasm.
2139D.J. Shaw et al. / Biochimica et Biophysica Acta 1783 (2008) 2137–2144
coils that co-localise with NPC filaments within the central pore ofthe NPC [42,43].
4. Nuclear export of the U snRNAs
Nuclear export of the U snRNAs through the NPC is dependentupon the interaction between the U snRNA mG cap structure and thenuclear cap binding complex, the export receptor CRM1/XpoI andRanGTP [16,44]. The nuclear cap binding complex (CBC) is aheterodimeric complex composed of two subunits, cap bindingprotein 80 (CBP80) and cap binding protein 20 (CBP20). Both ofthese subunits are required for binding to the mG cap structure[16,45–47]. A study by Izaurralde in 2002 [48], found that antibodiesraised against recombinant CBP20 prevented the interaction of theCBC with capped RNAs in vitro. The antibodies inhibited both pre-mRNA splicing and export of U snRNA to the cytoplasm. CRM1/Xpo1 isalso the export receptor for proteins carrying leucine-rich nuclearexport signals (NESs) [14]. CRM1 was identified as an export receptorfor U snRNA export in a study by Fischer et al. [11]. They specificallyinhibited U snRNA export by inactivating CRM1 with the unsaturated,branched-chain fatty acid leptomycin B, thereby saturating CRM1withan excess of NES peptides [11,14]. CRM1 is also heavily involved in thenuclear trafficking of mature U snRNPs [93], but this will be discussedlater (Fig. 4).
The small GTPase, Ran, is an essential factor for nuclear proteinexport [49]. Ran is predominantly found in the nucleus and its activityis controlled by a nuclear, chromatin associated, guanine-nucleotideexchange factor, RCC1, and by a cytoplasmic GTPase activating protein,RanGAP [50,51]. Fig. 2 shows a schematic representation of the role ofRan in nuclear import. The distribution of these two proteins wouldsuggest that nuclear Ran is predominantly loaded with GTP, whereascytoplasmic Ran is immediately converted into the GDP bound state.The precise function of Ran in translocation is not known, howeverthere is evidence that the RanGTP/RanGDP gradient between thenucleus/cytoplasm could drive the direction of import, as RanGDPmediates import, whereas RanGTPmediates export [52]. However it isknown that Ran is involved in the release of cargo/receptor complexes,
for example, RanGTP induces release of both importin α from β in theclassic NLS signal protein import and also release of M9 signals fromtransportin [52]. RanGTP is also thought to alter the NPC conformationduring transportation, by reducing NPC internal diameter by causingthe cytoplasmic filaments to extend and inhibit import [53].Hydrolysis of GTP by Ran is therefore required to allow release ofimport substrates, bound to the filaments, to be released into the NPC[53]. Ran has a low intrinsic activity, needing both the Ran-GTPase-activating protein (RanGAP) and the co-activator Ran-bindingprotein 1 (RanBP1) to achieve maximum GTPase activity. As both ofthese are located in the cytoplasm, the majority of Ran in the nucleusis believed to be in the RanGTP form [54].
A further protein thought to be involved in U snRNA export is thephosphorylated adaptor for RNA export protein (PHAX) [47]. Ohnoet al. demonstrated that PHAX is used as an adaptor molecule to aidthe binding of CRMI/RanGTP proteinwith the CBC/RNA complex usingmobility shift assays and phosphorylation inhibitory studies [47].PHAX is phosphorylated in the nucleus and this phosphorylation isessential for export complex assembly. PHAX is exported with RNA tothe cytoplasm, where it is dephosphorylated. This dephosphorylationcauses the export assembly to disassemble [47]. Fig. 2 shows adiagrammatic explanation of U snRNA nuclear export.
5. Cytoplasmic assembly of U snRNPs
Once the U snRNA/CBC/PHAX/CRM1/RanGTP complex has enteredthe cytoplasm, the complex dissociates. The Ran-GTPase activatorprotein (RanGAP) and the co-activator, RanBP1, mediate this dissocia-tion. In their presence, the Ran-GTPase activity is maximised, resultingin hydrolysis of the GTP to GDP providing the energy for the release ofthe snRNA [54]. The RanGDP is then transported back into the nucleusattached to the CRM1 receptor [54]. Dephosphorylation of PHAX mayalso be involved in dissociation of the export complex [47].
Once the U snRNAs are freed, cytoplasmic assembly of the snRNPscan occur. The protein components of the U snRNPs can be dividedinto two groups. The first includes proteins such as U1A, U170K, U2B″and U2A that are specific to certain U snRNA subclasses. The second

Fig. 3. Schematic representation of the U snRNP bipartite import complex. The bipartitesignal involves trimethylated guanosine (m3G) cap associated with snurportin1 andimportin β, and the proposed SMN dependent Sm core protein import, whichassociated either directly or indirectly with importin β and Ran. Although thisschematic shows SMN interacting directly with importin β, this proposed import signalmay involve an as yet unidentified import adaptor.
2140 D.J. Shaw et al. / Biochimica et Biophysica Acta 1783 (2008) 2137–2144
consists of the common Sm core proteins, named after Smith, a patientwith systemic lupus erythematosus who produced autoantibodiesagainst the proteins [55,56]. In the majority of adult mammalian celltypes there are eight common core proteins, B, B′, D1, D2, D3, E, F and G.The eight proteins are encoded by seven different gene; B and B′proteins are alternatively spliced products of a single gene, differingonly in their carboxyl-terminal part [57]. The Sm binding motif(5′-Pu-A-U3–6-G-Pu-3′, where Pu represents a purine nucleotide) ishighly conserved across species, with the adenosine residue inposition 2 present in 95% of characterised animal species [58]. Inadult neuronal cells and to a lesser extent in myocardial cells, the B/B′proteins are replaced by a ninth core protein, the N protein [59,60]. Itis believed that an ancestral gene duplication gave rise to the N proteinas B′/B share 98% homology to the N protein [59].
Newly synthesised Sm proteins are stored in four partiallyassembled cytoplasmic complexes that are snRNA-free [61,56].These are: (1) a (E.F.G)2 hexameric complex; (2) a 20S complexconsisting of D3.B and an uncharacterised 70 kDa protein; (3) acomplex of 2–6S consisting of (B)n; and (4) a D1.D2 complex[56,61–64]. An additional 6S complex consisting of D1.D2. (E.F.G)2,has also been identified in the cytoplasm, although this complex is lessstable than either the (E.F.G)2 hexameric complex or the D1.D2
complex [61,63]. All Sm core proteins contain two conserved motifs,Sm1 and Sm2. Sm1 contains strands β1–3, whilst Sm2 contains β4 andβ5 [65,66]. Kambach et al. found that an extended anti-parallelβ-sheet formed between strands β4 of the B protein and β5 of the D3
protein mediates the formation of the D3B heterodimer. Similarβ-sheet interactions were observed between D1and D2, where β4 ofthe D2 protein pairs with β5 of the D1 protein [66].
The assembly of the Sm core complex is believed to be a step-wiseordered process [67]. The initial step involves the E.G.F and D1D2
complexes interacting with the conserved 5′-Pu-A-U3–6-G-Pu-3′ Smbinding motif of the U snRNA to form a stable intermediate moleculecalled the subcore snRNP [63]. Once the subcore has been formed theD3.B/B′ complex binds to form the complete U snRNP molecule.
6. The role of the SMN complex in U snRNP
The survival motor neuron (SMN) protein has been implicated inthe cytoplasmic assembly of the U snRNPs [68]. The SMN protein is aubiquitously expressed RNA processing factor. Reduced levels of theSMN protein trigger the neurodegenerative disorder, spinal muscularatrophy (SMA) [69]. The SMN protein is part of a largemacromolecularcomplex, termed the Gemin complex [68]. This complex consists ofSMN, Gemin2, Gemin3, Gemin4, Gemin5, Gemin6, Gemin7 andGemin8. The Gemins predominantly co-localise with SMN in nuclearCajal bodies and also in the cytoplasm, where they also associate withthe Sm core proteins [70], and with the U snRNAs [71]. There is agrowing body of evidence demonstrating the SMN complex plays anintegral role in the cytoplasmic assembly of the U snRNPs. Earlyexperiments on oocytes demonstrated that functionally blocking theSMN protein with microinjected antibodies inhibited core proteinassembly on U1 snRNA [72]. In addition, reducing the levels of SMN,Gemin2, Gemin3, and Gemin4 proteins using interfering RNA (RNAi)all independently inhibit U snRNP assembly to some degree [73]. It isknown that the core proteins interact directly with several membersof the SMN complex, and that the U snRNA interacts directly withGemin5. This may suggest that the complex forms a scaffold thatfacilitates the assembly process. In addition, mutational analysisstudies on U snRNA demonstrate that the SMN complex may functionin a “quality control” role, by ensuring assembly only occurs onfunctional U snRNA [71].
Once the core domain has been assembled the 5′ mG cap ishypermethylated, forming a 2,2,7-trimethyl-guanosine (m2,2,7G; m3G)cap. This modification can only occur after the core complex has beenassembled [74]. The mature Sm core domain provides a binding site
for an RNA methyltransferase, which catalyses formation of the tri-methyl-guanosine (m3G) cap of the U snRNA via hypermethylation.The assembled particle is then transported actively to the nucleus[75,76]. Following assembly of the U snRNP they are required in thenucleus for splicing.
The import is controlled by a bipartite import signal consisting ofthe assembled Sm core complex and the m3G cap. By definition,independently neither of these signals is sufficient tomediate efficientimport. The more extensively studied pathway is the use of them3G cap as a nuclear targeting signal. This pathway utilises snurportin1 (SPN1) as an adaptor molecule [19]. The SPN1 molecule binds toboth the m3G cap and the importin βmolecule. The importin β is ableto mediate the interaction with the NPC and translocates the U snRNPimport complex through the NPC [19,77]. Studies by Humber et al. andRollenhagen et al. found that the m3G cap dependent import requiredimportin β but not Ran or energy [76,78].
The second pathway for U snRNP import is the use of the Sm coredomain of U snRNPs formed by Sm proteins. This needs to occur forthe hypermethylation of the (m3G) cap. This pathway again utilisesimportin β as a receptor for import, but also requires an unidentifiedadaptor molecule. Potentially, SMN could function as the adaptor(Fig. 3). In keeping with this, the nuclear import of both SMN and UsnRNPs are co-dependent, withmutations in either Sm core protein orSMN preventing the efficient import of both [77].
7. Maturation of U snRNPs
Once in the nucleus, binding of RanGTP to the import complextriggers dissociation and release of the U snRNP [79]. The absence ofthe m3G cap on the U snRNP ensures nuclear retention andsnurportin1 is transported back into the cytoplasm in associationwith CRM1 [80]. Unlike the cytoplasm, the nucleus is not compart-mentalised into membrane bound organelles. However, many nuclearantigens are localised in different sub-nuclear structures, distinct fromthe surrounding nucleoplasm [81,82]. Nuclear bodies include nucleoli,sites of ribosome production and RNA metabolism and transport[83], Cajal bodies (coiled bodies), Gemini of Cajal bodies (gems),promyelocytic leukaemia (PML) bodies, IGCs, interchromatin-granule-associated zones and PFs [84] and references therein.Antibody staining of Sm proteins demonstrates that snRNPs have acomplex nuclear localisation [85]. Once imported to the nucleus,snRNP particles are seen to interact with different sub-nuclearstructures, including IGCs, PFs and Cajal bodies [85]. Targeting tothese bodies is not a random event, but follows a clear step-wisepathway. Initially, newly imported U snRNPs are targeted to Cajalbodies (Fig. 4). From here, they then pass through the nucleolus, thenback to the Cajal bodies and finally on to the IGCs (Fig. 4). The central

Fig. 4. Schematic representation of the factors mediating the sub-nuclear targeting of U snRNPs. Newly imported U snRNPs are initially targeted to Cajal bodies (1). This process ismediated through an associationwith SMN, p80-coilin, ZPR1 and CRM1. The U snRNPs are then processed through the nucleolus (2), before returning to the Cajal bodies (3). Targetingto the nucleolus is depended on SMN, p80-coilin and Nopp140, while returning to Cajal bodies is controlled by SMN, p80-coilin, Nopp140 and protein phosphotase 4 (PP4). Finally,U snRNPs are targeted to the interchromatin granule clusters (IGCs; 4) through an association with SMN, CRM1 and p80-colin.
2141D.J. Shaw et al. / Biochimica et Biophysica Acta 1783 (2008) 2137–2144
role of the Cajal body in this process has lead to suggestions that theirprimary cellular role is the sorting and processing of snRNPs.
8. Mature U snRNPs accumulate in Cajal bodies
Unlike the mG cap associated with the immature U snRNA, boththe m3G cap and the Sm core proteins (B, B′, D1, D2, D3, E, F and G)have been detected in coiled bodies [86–88]. This suggests that UsnRNPs, which have passed through the cytoplasmic maturationstage, accumulate in Cajal bodies, but nascent snRNA transcripts donot. Targeting of the spliceosomal U snRNPs to coiled bodies appearsto be mediated through a coiled body marker protein, coilin p80 (Fig.4) [89]. Coilin p80 is an 80 kDa protein that is ubiquitously expressedin mammalian cells [90]. Coilin p80 contains two nuclear localisationsignals (NLS). However, mutational analysis studies show that ontheir own these NLSs are not sufficient to target coilin p80 to Cajalbodies [90].
Cajal bodies still form in the absence of coilin p80, but they do notcontain either the Sm core proteins or mature U snRNPs [89]. In cellsimmunodepleted for the Sm core proteins, coilin p80 levels in Cajalbodies are also reduced [89]. This suggests that coilin p80 and theU snRNPs are co-dependent on each other for Cajal body targeting.Although coilin p80 contains two nuclear localisation signals, it doesnot co-localise with U snRNAs or U snRNPs in the cytoplasm [89]. Thissuggests that coilin p80 has no role in either cytoplasmic maturationor nuclear import of the U snRNPs. However, it is possible that coilinp80 and the snRNPs form a complex once the snRNPs have beenreleased from the import complex and it is this nuclear complex thatmediates sub-nuclear distributions. Coilin p80 is known to be an SMNbinding partner [91,92]. Furthermore, symmetrical dimethylation of
the RGG-motifs of coilin p80 trigger the merging of Cajal bodies withtheir closely associated SMN-positive loci, Gemini of coiled bodies[91,92]. This process occurs during development in mammalian cells,with gems and Cajal bodies appearing as distinct sub-nuclear bodiesin fetal tissue, while the two are almost completely merged in adultcells [93]. This suggests that SMN not only mediates nuclear import ofthe U snRNP, but may also be involved in Cajal body targeting throughan associationwith coilin p80 (Fig. 4). In keepingwith this, a truncatedSMN protein lacking the first 28 amino acids inhibits Cajal bodytargeting of the U snRNPs [94]. This targeting also appears to involvethe nuclear export adaptor, CRM1 (Fig. 4), which interacts with bothSMN and the Sm core proteins [2], and the zinc-finger binding protein,ZPR1 (Fig. 4) [95].
Within the SMN protein several core internal targeting sequenceshave recently been identified. These include the domains encodedby exons 2b, 3 and 6 [96]. Removal of any of these domainsprevented Cajal body targeting of the SMN protein. Exons 2b and 6encode self-association domains, while exon 3 encodes the Tudordomain, suggesting that self-association and RGG-motif binding playan integral role in Cajal targeting of SMN [96]. As several Sm coreproteins contain RGG-domains, these observations suggest that anassociation with U snRNP may in part mediate SMN targeting toCajal bodies.
9. Nucleolar processing of U snRNPs
Antibodies against the m3G cap of mature snRNPs and coilin p80showed that low levels of U snRNPs are present in the nucleolus ofHeLa cells [97] and yeast cells [98]. This led to the suggestion thatalthough U snRNPs and/or coilin p80 do not accumulate in the

2142 D.J. Shaw et al. / Biochimica et Biophysica Acta 1783 (2008) 2137–2144
nucleolus, they may migrate through it as part of a nuclear matu-ration stage [97,99]. U6 snRNA localises to nucleoli where it undergoes2′-O-methylation and pseudouridylation of defined nucleotides byguide snoRNA's (as that with scaRNAs in Cajal bodies) [99]. U2, U4 andU5 snRNA are also modified in the nucleoli [100]. Mechanismmediating nucleoli localisation of U6 snRNA is through its 3′ nucleolarlocalisation element [100].
Distribution of U snRNPs is also altered by the phosphorylationstate of coilin p80 [97]. Coilin p80 contains several serine phospho-rylation sites [101]. Inhibition of PP1 and PP2A serine/threonine(Ser/Thr) protein phosphatases [102] results in accumulation of coilinp80 and U snRNPs in the nucleolus in structures resembling coiledbodies, again highlighting their associated distribution pattern [97].The phosphorylation state of a serine residue at position 202 of coilinp80 has been shown to control nuclear distribution of both coilin p80and U snRNPs [97]. Conversion of the serine to an alanine (S202A) themethyl-R group of which mimics serine in a dephosphorylated statetargeted both molecules to the Cajal bodies. In contrast, conversion toan aspartate (S202D), the carboxyl-R group of which simulates serinein a phosphorylated state, resulted in a nucleolar localisation [97,103].In accordance with the phosphorylation state mediated distribution,a protein phosphatase 1 isoform (PP1δ) and two separate proteinkinases (PKC and PKR) have been identified in the nucleolus and coiledbodies. This is consistent with the dephosphorylation inhibitionstudies with PP1 inhibition preventing both coilin p80 and U snRNPsfrom leaving the nucleolus [97]. In addition protein phosphotase 4(PP4) has been identified as a member of the SMN complex [104]. PP4associates directly with Gemin3 and Gemin4, and directly modulatescellular targeting of the U snRNPs (Fig. 4) [101], again highlightingimportance phosphorylation states have on sub-nuclear targeting.
10. Dynamic interaction between the nucleolus and coiled bodies
A functional interaction between Cajal bodies and the nucleolusis evident as over-expression of mutant coilin p80 lacking theC-terminus disrupts nucleolar morphology [90]. Both organelles alsoshare several components, including fibrillarin, nucleolin, topoisome-rase I and Nopp140 [86,105]. The exact function of Nopp140 isunknown, but it has been shown to shuttle between the nucleolus andthe cytoplasm, with the N- and C-termini encoding nuclear export andimport signals respectively, leading to a suggested role in trafficking ofpre-ribosomal subunits [105]. However, as Nopp140 containsno characterised RNA-binding motifs, interactions with ribosomal-subunits must be with the ribonucleoproteins rather than the rRNAmolecules [105]. Nopp140 has been shown to interact directly withcoilin p80. It is therefore interesting that Nopp140, when transportedinto the nucleus from the cytoplasm, passes through the nucleolusbefore it localises in coiled bodies (Fig. 4).
In addition, SMN has been reported in the nucleolus [93,106]. LikeU snRNPs, SMN does not appear to accumulate in the nucleolus,suggesting dynamic association [93,106]. However, two regions in theSMN protein have been linked to nucleolar targeting. Expressionof the exon 2b encoded domain alone specifically targets a GFPconstruct to the nucleolus [107], while removal of SMN exon 3(SMNΔ3) also targets SMN to the nucleolus [96]. This suggests severalthings: 1) that SMN exon 2b may contain a nucleolar localisationdomain; 2) that SMN exon 3 may contain a nucleolar exportsequence; and 3) SMN nuclear export is dependent on its ability toassociate with the U snRNPs. It is possible that all three of these arecorrect. However, a detailed analysis of the Tudor domain highlightedthat there is no minimal domain that efficiently enable nucleolarexport, suggesting an intact Tudor domain is essential for nucleolartrafficking. This is not too much of a surprise, considering thatassociation with both U snRNPs and coilin p80 both mediate SMNnuclear targeting, as both of these associations are mediated by theTudor domain. Therefore, a Nopp140/U snRNP/coilin-p80/SMN com-
plex may theoretically migrate through the nucleolus prior to coiledbody accumulation (Fig. 4) [105].
Expression of a truncated Nopp140 containing only the C-terminalportion of the protein results in the dispersal of coiled bodies andrelocation of coiled body proteins, including fibrillarin, coilin p80 andSm core proteins/U snRNPs [105]. This suggests that the N-terminus ofNopp140 is essential for the structural integrity of coiled bodies [105]and that full-length Nopp140 may act as a molecular chaperonebetween the nucleolus and coiled bodies. If sub-nuclear distribution ismediated by the phosphorylation states of proteins, Nopp140 is a goodcandidate for a distribution mediator, as it is potentially highly-phosphorylated, with up to 80 Ser/Thr phosphorylation sites.
11. Interchromatin granules and interchromatingranule-associated zones
Mature U snRNPs are efficiently targeted to IGCs, which are themain nuclear storage sites of the splicing machinery. U snRNPs freelymove between the IGCs and Cajal bodies in a bi-directional manner,and this process appears to be dependent on both SMN and CRM1[2,94,108]. The IGCs are smaller in size than Cajal bodies and are morenumerous, with an average mammalian cell containing between 30and 50 IGCs [109]. The low levels of poly(A)+RNA detected in IGCs andthe lack of newly transcribed RNA suggests they are not the sites oftranscription or pre-mRNA splicing [110]. Instead it is suggested thatinterchromatin granule clusters are involved in both storage ofspliceosome components and spliceosome assembly [111,112]. Thiswould suggest that, as the spliceosome assembles on poly(A)+RNA,the level of poly(A)+RNA in the IGCs is the limiting factor and that onceassembled the spliceosome/RNA complexmigrates to the PFs. IGCs areassociated with fibrillar structures termed interchromatin granuleassociated zones [110]. These zones contain both U1 snRNP and coilinp80, leading to the suggestion that they may be involved in a final U1snRNP maturation step, prior to storage in the IGCs [113].
12. Protein phosphorylation mediates distribution and function ofsplicing factors
As well as controlling nucleolar/coiled body shuttling, a phospha-tase-kinase network is believed to control both localisation of splicingfactors to the IGCs and spliceosomal assembly/disassembly. U snRNPsand non-snRNP serine/arginine rich (SR) proteins must be depho-sphorylated to accumulate in the IGs. Inhibition of PP1 causeslocalisation of both factors in irregular nuclear speckles [109]. Thismay suggest that only U snRNPs that have progressed through thenucleolar-maturation stage, and thus have been dephosphorylated bythe PP1δ phosphatase, may translocate to the IGCs, a mechanism thatwould ensure that only mature U snRNPs are involved in spliceosomeformation.
13. U snRNP processing and spinal muscular atrophy
The integral role SMN plays in U snRNP assembly, processing andtrafficking, suggests that spinal muscular atrophy (SMA) may be dueto a problem with RNA processing. However, pathology in SMA ismainly restricted to the anterior horn cells of the spinal cord [114], andtherefore, is unlikely to be due to a systemic issue with processing ofthe U snRNPs. It is more likely that SMN's associationwith RNPs is notrestricted to the U snRNPs. For example, SMN interacts directly withhnRNP R, and this complex is involved in the axonal transport ofβ-actin mRNA [115]. As axonal transport is restricted to neuronal sub-types it is more likely that a breakdown in this process rather thaninefficient U snRNP assembly triggers the disease pathology. However,complete loss of SMN protein results in an embryonic lethalphenotype, which is more conducive with loss of U snRNP as-sembly [116]. With no SMN present, hypothetically there would be no

2143D.J. Shaw et al. / Biochimica et Biophysica Acta 1783 (2008) 2137–2144
U snRNP assembly, and consequently developing cells would beunable to splice pre-mRNA or produce protein, which is in keepingwith the lethality of this genotype, but not with the phenotypeassociated with SMA. However, as the U snRNP assembly pathway isthe most accepted and understood of SMN's assigned functions, morebasic scientific research is needed to truly understand the mechanisticcause of SMA.
14. Concluding remarks
The assembly and trafficking of the U snRNPs is a complex andrigorously controlled process. Studying this process has helpedunderstand the function of several key proteins. For example, CRM1,which until recently was seen simply a nuclear export mediator, isnow known to function as a nuclear trafficking protein. However,there remain several key areas that need addressing before we fullyunderstand the process. Firstly, we need to functionally characterisethe role of the Gemin complex in the cytoplasmic assembly process;does the complex form a scaffold aroundwhich the complex can form,or does it actively catalyse the interaction? Secondly, we need todetermine whether the Gemin complex is the second import adaptorprotein, or whether this role is performed by an as yet unidentifiedfactor. Thirdly, we need to better understand the nucleolar processing/maturation step. Fourthly, we need to determine why the U snRNPsare cycled between the Cajal bodies and IGCs. Addressing thesequestions will provide a comprehensive understanding of a systemiccellular function that is essential for cell viability. They will alsoexpand our understanding of the SMN protein, and could helpelucidate to cause of SMA and highlight novel therapeutic targets forthis, and other disorders that involve issues with RNP assembly.
Acknowledgements
The authors would like to thank Dr. Robert Morse for comments.P.J.Y. and D.J.S are supported by grants from Fight SMA, the SMA Trust,and the Vandervell Foundation.
References
[1] C.L. Will, R. Luhrmann, Spliceosomal UsnRNP biogenesis, structure and function,Curr. Opin. Cell Biol. 13 (2001) 290–301.
[2] J. Sleeman, A regulatory role for CRM1 in the multi-directional trafficking ofsplicing snRNPs in the mammalian nucleus, J. Cell Sci. 120 (2007) 1540–1550.
[3] J. Hamm, I.W. Mattaj, Monomethylated cap structures facilitate RNA export fromthe nucleus, Cell 63 (1990) 109–118.
[4] S. Kass, K. Tyc, J.A. Steitz, B. Sollner-Webb, The U3 small nucleolar ribonucleo-protein functions in the first step of preribosomal RNA processing, Cell 60 (1990)897–908.
[5] K. Tyc, J.A. Steitz, U3, U8 and U13 comprise a new class of mammalian snRNPslocalized in the cell nucleolus, Embo. J. 8 (1989) 3113–3119.
[6] N. Hernandez, Small nuclear RNA genes: a model system to study fundamentalmechanisms of transcription, J. Biol. Chem. 276 (2001) 26733–26736.
[7] S. Shuman, Capping enzyme in eukaryotic mRNA synthesis, Prog. Nucleic. AcidRes. Mol. Biol. 50 (1995) 101–129.
[8] F.R. Bischoff, D. Gorlich, RanBP1 is crucial for the release of RanGTP from importinbeta-related nuclear transport factors, FEBS Lett. 419 (1997) 249–254.
[9] H.P. Bogerd, A. Echarri, T.M. Ross, B.R. Cullen, Inhibition of human immunode-ficiency virus Rev and human T-cell leukemia virus Rex function, but notMason–Pfizer monkey virus constitutive transport element activity, by a mutanthuman nucleoporin targeted to Crm1, J. Virol. 72 (1998) 8627–8635.
[10] C.N. Cole, C.M. Hammell, Nucleocytoplasmic transport: driving and directingtransport, Curr. Biol. 8 (1998) R368–R372.
[11] U. Fischer, J. Huber, W.C. Boelens, I.W. Mattaj, R. Luhrmann, The HIV-1 Revactivation domain is a nuclear export signal that accesses an export pathwayused by specific cellular RNAs, Cell 82 (1995) 475–483.
[12] D. Gorlich, R. Kraft, S. Kostka, F. Vogel, E. Hartmann, R.A. Laskey, I.W. Mattaj, E.Izaurralde, Importin provides a link between nuclear protein import and UsnRNA export, Cell 87 (1996) 21–32.
[13] M. Floer, G. Blobel, M. Rexach, Disassembly of RanGTP–karyopherin betacomplex, an intermediate in nuclear protein import, J. Biol. Chem. 272 (1997)19538–19546.
[14] M. Fornerod, M. Ohno, M. Yoshida, I.W. Mattaj, CRM1 is an export receptor forleucine-rich nuclear export signals, Cell 90 (1997) 1051–1060.
[15] D. Gorlich, Transport into andout of the cell nucleus, Embo. J.17 (1998) 2721–2727.
[16] E. Izaurralde, J. Lewis, C. McGuigan, M. Jankowska, E. Darzynkiewicz, I.W. Mattaj,A nuclear cap binding protein complex involved in pre-mRNA splicing, Cell 78(1994) 657–668.
[17] E. Izaurralde, S. Adam, Transport of macromolecules between the nucleus andthe cytoplasm, Rna. 4 (1998) 351–364.
[18] K. Hellmuth, D.M. Lau, F.R. Bischoff, M. Kunzler, E. Hurt, G. Simos, Yeast Los1p hasproperties of an exportin-like nucleocytoplasmic transport factor for tRNA, Mol.Cell Biol. 18 (1998) 6374–6386.
[19] J. Huber, U. Cronshagen, M. Kadokura, C. Marshallsay, T. Wada, M. Sekine, R.Luhrmann, Snurportin1, an m3G-cap-specific nuclear import receptor with anovel domain structure, Embo. J. 17 (1998) 4114–4126.
[20] J.K. Hood, P.A. Silver, In or out? Regulating nuclear transport, Curr. Opin. Cell Biol.11 (1999) 241–247.
[21] M.A. D'Angelo, M.W. Hetzer, The role of the nuclear envelope in cellularorganization, Cell Mol. Life Sci. 63 (2006) 316–332.
[22] M. Stewart, Molecular mechanism of the nuclear protein import cycle, Nat. Rev.Mol. Cell Biol. 8 (2007) 195–208.
[23] R.Y. Lim, B. Fahrenkrog, The nuclear pore complex up close, Curr. Opin. Cell Biol.18 (2006) 342–347.
[24] W.H. Raharjo, P. Enarson, T. Sullivan, C.L. Stewart, B. Burke, Nuclear envelopedefects associated with LMNA mutations cause dilated cardiomyopathy andEmery–Dreifuss muscular dystrophy, J. Cell Sci. 114 (2001) 4447–4457.
[25] H. Herrmann, U. Aebi, Intermediate filaments: molecular structure, assemblymechanism, and integration into functionally distinct intracellular Scaffolds,Annu. Rev. Biochem. 73 (2004) 749–789.
[26] A.E. Emery, Emery–Dreifuss syndrome, J. Med. Genet. 26 (1989) 637–641.[27] M.L. Watson, The nuclear envelope; its structure and relation to cytoplasmic
membranes, J. Biophys. Biochem. Cytol. 1 (1955) 257–270.[28] W.W. Franke, U. Scheer, G. Krohne, E.D. Jarasch, The nuclear envelope and the
architecture of the nuclear periphery, J. Cell Biol. 91 (1981) 39s–50s.[29] V.C. Padmakumar, S. Abraham, S. Braune, A.A. Noegel, B. Tunggal, I.
Karakesisoglou, E. Korenbaum, Enaptin, a giant actin-binding protein, is anelement of the nuclear membrane and the actin cytoskeleton, Exp. Cell Res. 295(2004) 330–339.
[30] Y.Y. Zhen, T. Libotte, M.Munck, A.A. Noegel, E. Korenbaum, NUANCE, a giant proteinconnecting the nucleus and actin cytoskeleton, J. Cell Sci. 115 (2002) 3207–3222.
[31] H.J. Worman, J.C. Courvalin, The inner nuclear membrane, J. Membr. Biol. 177(2000) 1–11.
[32] B. Burke, L.C. Mounkes, C.L. Stewart, The nuclear envelope in muscular dystrophyand cardiovascular diseases, Traffic 2 (2001) 675–683.
[33] J.M. Holaska, K.L. Wilson, M. Mansharamani, The nuclear envelope, lamins andnuclear assembly, Curr. Opin. Cell Biol. 14 (2002) 357–364.
[34] R.W. Merriam, On the fine structure and composition of the nuclear envelope,J. Biophys. Biochem. Cytol. 11 (1961) 559–570.
[35] W.W. Franke, Isolated nuclear membranes, J. Cell Biol. 31 (1966) 619–623.[36] I.G. Macara, Transport into and out of the nucleus, Microbiol. Mol. Biol. Rev. 65
(2001) 570–594 table of contents.[37] K. Weis, Nucleocytoplasmic transport: cargo trafficking across the border, Curr.
Opin. Cell Biol. 14 (2002) 328–335.[38] H. Fried, U. Kutay, Nucleocytoplasmic transport: taking an inventory, Cell Mol.
Life Sci. 60 (2003) 1659–1688.[39] B. Fahrenkrog, U. Aebi, The nuclear pore complex: nucleocytoplasmic transport
and beyond, Nat. Rev. Mol. Cell Biol. 4 (2003) 757–766.[40] M.S. Rodriguez, C. Dargemont, F. Stutz, Nuclear export of RNA, Biol. Cell 96 (2004)
639–655.[41] N. Shulga, D.S. Goldfarb, Binding dynamics of structural nucleoporins govern
nuclear pore complex permeability and may mediate channel gating, Mol. CellBiol. 23 (2003) 534–542.
[42] T.A. Isgro, K. Schulten, Cse1p-binding dynamics reveal a binding pattern forFG-repeat nucleoporins on transport receptors, Structure 15 (2007) 977–991.
[43] D.P. Denning, S.S. Patel, V. Uversky, A.L. Fink, M. Rexach, Disorder in the nuclearpore complex: the FG repeat regions of nucleoporins are natively unfolded, Proc.Natl. Acad. Sci. U. S. A. 100 (2003) 2450–2455.
[44] E. Izaurralde, J. Lewis, C. Gamberi, A. Jarmolowski, C.McGuigan, I.W.Mattaj, A cap-binding protein complex mediating U snRNA export, Nature 376 (1995) 709–712.
[45] N. Kataoka, M. Ohno, K. Kangawa, Y. Tokoro, Y. Shimura, Cloning of acomplementary DNA encoding an 80 kilodalton nuclear cap binding protein,Nucleic Acids Res. 22 (1994) 3861–3865.
[46] E. Izaurralde, C. McGuigan, I.W. Mattaj, Nuclear localization of a cap-bindingprotein complex, Cold Spring Harb. Symp. Quant. Biol. 60 (1995) 669–675.
[47] M. Ohno, A. Segref, A. Bachi, M. Wilm, I.W. Mattaj, PHAX, a mediator of U snRNAnuclear export whose activity is regulated by phosphorylation, Cell 101 (2000)187–198.
[48] E. Izaurralde, A novel family of nuclear transport receptorsmediates the export ofmessenger RNA to the cytoplasm, Eur. J. Cell Biol. 81 (2002) 577–584.
[49] F. Melchior, B. Paschal, J. Evans, L. Gerace, Inhibition of nuclear protein import bynonhydrolyzable analogues of GTP and identification of the small GTPaseRan/TC4 as an essential transport factor, J. Cell Biol. 123 (1993) 1649–1659.
[50] F.R. Bischoff, H. Ponstingl, Catalysis of guanine nucleotide exchange on Ran by themitotic regulator RCC1, Nature 354 (1991) 80–82.
[51] F.R. Bischoff, C. Klebe, J. Kretschmer, A. Wittinghofer, H. Ponstingl, RanGAP1induces GTPase activity of nuclear Ras-related Ran, Proc. Natl. Acad. Sci. U. S. A. 91(1994) 2587–2591.
[52] E. Izaurralde, U. Kutay, C. von Kobbe, I.W. Mattaj, D. Gorlich, The asymmetricdistribution of the constituents of the Ran system is essential for transport intoand out of the nucleus, Embo. J. 16 (1997) 6535–6547.

2144 D.J. Shaw et al. / Biochimica et Biophysica Acta 1783 (2008) 2137–2144
[53] M.W. Goldberg, S.A. Rutherford, M. Hughes, L.A. Cotter, S. Bagley, E. Kiseleva, T.D.Allen, P.R. Clarke, Ran alters nuclear pore complex conformation, J. Mol. Biol. 300(2000) 519–529.
[54] S. Nakielny, G. Dreyfuss, Transport of proteins and RNAs in and out of the nucleus,Cell 99 (1999) 677–690.
[55] E.M. Tan, Antinuclear antibodies: diagnostic markers for autoimmune diseasesand probes for cell biology, Adv. Immunol. 44 (1989) 93–151.
[56] R. Luhrmann, B. Kastner, M. Bach, Structure of spliceosomal snRNPs and their rolein pre-mRNA splicing, Biochim. Biophys. Acta 1087 (1990) 265–292.
[57] A. van Dam, I. Winkel, J. Zijlstra-Baalbergen, R. Smeenk, H.T. Cuypers, Clonedhuman snRNP proteins B and B′ differ only in their carboxy-terminal part,Embo. J. 8 (1989) 3853–3860.
[58] C.Guthrie, B. Patterson, Spliceosomal snRNAs, Annu. Rev. Genet. 22 (1988) 387–419.[59] J.D. Huntriss, D.S. Latchman, D.G. Williams, The snRNP core protein SmB and
tissue-specific SmN protein are differentially distributed between snRNPparticles, Nucleic Acids Res. 21 (1993) 4047–4053.
[60] T.A. Gray, M.J. Smithwick, M.A. Schaldach, D.L. Martone, J.A. Graves, J.R. McCarrey,R.D. Nicholls, Concerted regulation and molecular evolution of the duplicatedSNRPB′/B and SNRPN loci, Nucleic Acids Res. 27 (1999) 4577–4584.
[61] D.E. Fisher, G.E. Conner, W.H. Reeves, R. Wisniewolski, G. Blobel, Small nuclearribonucleoprotein particle assembly in vivo: demonstration of a 6S RNA-free coreprecursor and posttranslational modification, Cell 42 (1985) 751–758.
[62] C.L. Will, S.E. Behrens, R. Luhrmann, Protein composition of mammalianspliceosomal snRNPs, Mol. Biol. Rep. 18 (1993) 121–126.
[63] V.A. Raker, G. Plessel, R. Luhrmann, The snRNP core assembly pathway:identification of stable core protein heteromeric complexes and an snRNPsubcore particle in vitro, Embo. J. 15 (1996) 2256–2269.
[64] M. Fury, J. Andersen, P. Ponda, R. Aimes, G.W. Zieve, Thirteen anti-Smmonoclonalantibodies immunoprecipitate the three cytoplasmic snRNP core proteinprecursors in six distinct subsets, J. Autoimmun. 12 (1999) 91–100.
[65] H. Hermann, P. Fabrizio, V.A. Raker, K. Foulaki, H. Hornig, H. Brahms, R. Luhrmann,snRNP Sm proteins share two evolutionarily conserved sequence motifs whichare involved in Sm protein–protein interactions, Embo. J. 14 (1995) 2076–2088.
[66] C. Kambach, S.Walke, R. Young, J.M. Avis, E. de la Fortelle, V.A. Raker, R. Luhrmann,J. Li, K. Nagai, Crystal structures of two Sm protein complexes and theirimplications for the assembly of the spliceosomal snRNPs, Cell 96 (1999) 375–387.
[67] G. Plessel, R. Luhrmann, B. Kastner, Electron microscopy of assembly inter-mediates of the snRNP core: morphological similarities between the RNA-free(E.F.G) proteinheteromerand the intact snRNP core, J.Mol. Biol. 265 (1997) 87–94.
[68] S.J. Kolb, D.J. Battle, G. Dreyfuss, Molecular functions of the SMN complex, J. Child.Neurol. 22 (2007) 990–994.
[69] S. Lefebvre, L. Burglen, S. Reboullet, O. Clermont, P. Burlet, L. Viollet, B. Benichou,C. Cruaud, P. Millasseau, M. Zeviani, et al., Identification and characterization of aspinal muscular atrophy-determining gene, Cell 80 (1995) 155–165.
[70] Q. Liu, U. Fischer, F. Wang, G. Dreyfuss, The spinal muscular atrophy disease geneproduct, SMN, and its associated protein SIP1 are in a complex with spliceosomalsnRNP proteins, Cell 90 (1997) 1013–1021.
[71] T.J. Golembe, J. Yong, G. Dreyfuss, Specific sequence features, recognized by theSMN complex, identify snRNAs and determine their fate as snRNPs, Mol. Cell Biol.25 (2005) 10989–11004.
[72] U. Fischer, Q. Liu, G. Dreyfuss, The SMN–SIP1 complex has an essential role inspliceosomal snRNP biogenesis, Cell 90 (1997) 1023–1029.
[73] K.B. Shpargel, A.G. Matera, Gemin proteins are required for efficient assembly ofSm-class ribonucleoproteins, Proc. Natl. Acad. Sci. U. S. A. 102 (2005)17372–17377.
[74] I.W. Mattaj, Cap trimethylation of U snRNA is cytoplasmic and dependent onU snRNP protein binding, Cell 46 (1986) 905–911.
[75] J. Yong, T.J. Golembe, D.J. Battle, L. Pellizzoni, G. Dreyfuss, snRNAs contain specificSMN-binding domains that are essential for snRNP assembly, Mol. Cell Biol. 24(2004) 2747–2756.
[76] C. Rollenhagen, N. Pante, Nuclear import of spliceosomal snRNPs, Can. J. Physiol.Pharmacol. 84 (2006) 367–376.
[77] U. Narayanan, T. Achsel, R. Luhrmann, A.G. Matera, Coupled in vitro import ofUsnRNPsandSMN, the spinalmuscular atrophyprotein,Mol. Cell 16 (2004)223–234.
[78] J. Huber, A. Dickmanns, R. Luhrmann, The importin-beta binding domain ofsnurportin1 is responsible for the Ran- and energy-independent nuclear importof spliceosomal U snRNPs in vitro, J. Cell Biol. 156 (2002) 467–479.
[79] M. Rexach, G. Blobel, Protein import into nuclei: association and dissociationreactions involving transport substrate, transport factors, and nucleoporins, Cell83 (1995) 683–692.
[80] E. Paraskeva, E. Izaurralde, F.R. Bischoff, J. Huber, U. Kutay, E. Hartmann, R.Luhrmann, D. Gorlich, CRM1-mediated recycling of snurportin 1 to the cyto-plasm, J. Cell Biol. 145 (1999) 255–264.
[81] T. Misteli, D.L. Spector, The cellular organization of gene expression, Curr. Opin.Cell Biol. 10 (1998) 323–331.
[82] A.I. Lamond, W.C. Earnshaw, Structure and function in the nucleus, Science 280(1998) 547–553.
[83] T. Pederson, The plurifunctional nucleolus,Nucleic Acids Res. 26 (1998) 3871–3876.[84] J.E. Sleeman, A.I. Lamond, Newly assembled snRNPs associate with coiled bodies
before speckles, suggesting a nuclear snRNP maturation pathway, Curr. Biol. 9(1999) 1065–1074.
[85] A.I. Lamond, M. Carmo-Fonseca, Localisation of splicing snRNPs in mammaliancells, Mol. Biol. Rep. 18 (1993) 127–133.
[86] I. Raska, L.E. Andrade, R.L. Ochs, E.K. Chan, C.M. Chang, G. Roos, E.M. Tan,Immunological and ultrastructural studies of the nuclear coiled body withautoimmune antibodies, Exp. Cell Res. 195 (1991) 27–37.
[87] S. Fakan, G. Leser, T.E. Martin, Ultrastructural distribution of nuclear ribonucleo-proteins as visualized by immunocytochemistry on thin sections, J. Cell Biol. 98(1984) 358–363.
[88] G.L. Eliceiri, J.S. Ryerse, Detection of intranuclear clusters of Sm antigens withmonoclonal anti-Sm antibodies by immunoelectron microscopy, J. Cell Physiol.121 (1984) 449–451.
[89] D.W. Bauer, J.G. Gall, Coiled bodies without coilin, Mol. Biol. Cell 8 (1997) 73–82.[90] K. Bohmann, J.A. Ferreira, A.I. Lamond, Mutational analysis of p80 coilin indicates
a functional interaction between coiled bodies and the nucleolus, J. Cell Biol. 131(1995) 817–831.
[91] F.M. Boisvert, J. Cote, M.C. Boulanger, P. Cleroux, F. Bachand, C. Autexier, S. Richard,Symmetrical dimethylarginine methylation is required for the localization ofSMN in Cajal bodies and pre-mRNA splicing, J. Cell Biol. 159 (2002) 957–969.
[92] M.D. Hebert, P.W. Szymczyk, K.B. Shpargel, A.G. Matera, Coilin forms the bridgebetween Cajal bodies and SMN, the spinal muscular atrophy protein, Genes Dev.15 (2001) 2720–2729.
[93] P.J. Young, T.T. Le, M. Dunckley, T.M. Nguyen, A.H. Burghes, G.E. Morris, Nucleargems and Cajal (coiled) bodies in fetal tissues: nucleolar distribution of the spinalmuscular atrophy protein, SMN, Exp. Cell Res. 265 (2001) 252–261.
[94] L. Pellizzoni, N. Kataoka, B. Charroux, G. Dreyfuss, A novel function for SMN, thespinal muscular atrophy disease gene product, in pre-mRNA splicing, Cell 95(1998) 615–624.
[95] L. Gangwani, R.A. Flavell, R.J. Davis, ZPR1 is essential for survival and is requiredfor localization of the survival motor neurons (SMN) protein to Cajal bodies, Mol.Cell Biol. 25 (2005) 2744–2756.
[96] R. Morse, D.J. Shaw, A.G. Todd, P.J. Young, Targeting of SMN to Cajal bodies ismediated by self-association, Hum. Mol. Genet. 16 (2007) 2349–2358.
[97] C.E. Lyon, K. Bohmann, J. Sleeman, A.I. Lamond, Inhibition of protein depho-sphorylation results in the accumulation of splicing snRNPs and coiled bodieswithin the nucleolus, Exp. Cell Res. 230 (1997) 84–93.
[98] J.A. Potashkin, R.J. Derby, D.L. Spector, Differential distribution of factors involved inpre-mRNAprocessing in theyeast cell nucleus,Mol. Cell Biol.10 (1990) 3524–3534.
[99] T.S. Lange, S.A. Gerbi, Transient nucleolar localization Of U6 small nuclear RNA inXenopus Laevis oocytes, Mol. Biol. Cell 11 (2000) 2419–2428.
[100] S.A. Gerbi, T.S. Lange, All small nuclear RNAs (snRNAs) of the [U4/U6.U5]Tri-snRNP localize to nucleoli; identification of the nucleolar localizationelement of U6 snRNA, Mol. Biol. Cell 13 (2002) 3123–3137.
[101] M. Carmo-Fonseca, J. Ferreira, A.I. Lamond, Assembly of snRNP-containing coiledbodies is regulated in interphase and mitosis-evidence that the coiled body is akinetic nuclear structure, J. Cell Biol. 120 (1993) 841–852.
[102] P.T. Cohen, N.D. Brewis, V. Hughes, D.J. Mann, Protein serine/threoninephosphatases; an expanding family, FEBS. Lett. 268 (1990) 355–359.
[103] J. Sleeman, C.E. Lyon, M. Platani, J.P. Kreivi, A.I. Lamond, Dynamic interactionsbetween splicing snRNPs, coiled bodies and nucleoli revealed using snRNPprotein fusions to the green fluorescent protein, Exp. Cell Res. 243 (1998)290–304.
[104] G.K. Carnegie, J.E. Sleeman, N. Morrice, C.J. Hastie, M.W. Peggie, A. Philp, A.I.Lamond, P.T. Cohen, Protein phosphatase 4 interacts with the survival of motorneurons complex and enhances the temporal localisation of snRNPs, J. Cell Sci.116 (2003) 1905–1913.
[105] C. Isaac, Y. Yang, U.T. Meier, Nopp140 functions as a molecular link between thenucleolus and the coiled bodies, J. Cell Biol. 142 (1998) 319–329.
[106] K.A. Wehner, L. Ayala, Y. Kim, P.J. Young, B.A. Hosler, C.L. Lorson, S.J. Baserga, J.W.Francis, Survival motor neuron protein in the nucleolus of mammalian neurons,Brain Res. 945 (2002) 160–173.
[107] B. Renvoise, K. Khoobarry, M.C. Gendron, C. Cibert, L. Viollet, S. Lefebvre, Distinctdomains of the spinal muscular atrophy protein SMN are required for targeting toCajal bodies in mammalian cells, J. Cell Sci. 119 (2006) 680–692.
[108] J.E. Sleeman, P. Ajuh, A.I. Lamond, snRNP protein expression enhances theformation of Cajal bodies containing p80-coilin and SMN, J. Cell Sci. 114 (2001)4407–4419.
[109] T. Misteli, D.L. Spector, Protein phosphorylation and the nuclear organization ofpre-mRNA splicing, Trends Cell Biol. 7 (1997) 135–138.
[110] E. Puvion, F. Puvion-Dutilleul, Ultrastructure of the nucleus in relation totranscription and splicing: roles of perichromatin fibrils and interchromatingranules, Exp. Cell Res. 229 (1996) 217–225.
[111] N. Visa, F. Puvion-Dutilleul, J.P. Bachellerie, E. Puvion, Intranuclear distribution ofU1 and U2 snRNAs visualized by high resolution in situ hybridization: revelationof a novel compartment containing U1 but not U2 snRNA in HeLa cells, Eur. J. CellBiol. 60 (1993) 308–321.
[112] N. Visa, F. Puvion-Dutilleul, F. Harper, J.P. Bachellerie, E. Puvion, Intranucleardistribution of poly(A) RNA determined by electron microscope in situhybridization, Exp. Cell Res. 208 (1993) 19–34.
[113] F. Puvion-Dutilleul, S. Besse, E.K. Chan, E.M. Tan, E. Puvion, p80-coilin: acomponent of coiled bodies and interchromatin granule-associated zones, J. CellSci. 108 (Pt 3) (1995) 1143–1153.
[114] T.O. Crawford, C.A. Pardo, The neurobiology of childhood spinal muscularatrophy, Neurobiol. Dis. 3 (1996) 97–110.
[115] W. Rossoll, A.K. Kroning, U.M. Ohndorf, C. Steegborn, S. Jablonka, M. Sendtner,Specific interaction of Smn, the spinal muscular atrophy determining geneproduct, with hnRNP-R and gry-rbp/hnRNP-Q: a role for Smn in RNA processingin motor axons? Hum. Mol. Genet. 11 (2002) 93–105.
[116] B. Schrank, R. Gotz, J.M. Gunnersen, J.M. Ure, K.V. Toyka, A.G. Smith, M. Sendtner,Inactivation of the survival motor neuron gene, a candidate gene for humanspinal muscular atrophy, leads to massive cell death in early mouse embryos,Proc. Natl. Acad. Sci. U. S. A. 94 (1997) 9920–9925.




















