
Isotopic evidence for zooplankton as an important food source for the mysid Paramysis lacustris in...
8
Isotopic evidence for zooplankton as an important food source for the mysid Paramysis lacustris in the Curonian Lagoon, the South-Eastern Baltic Sea J urat _ e Lesutien _ e a, * , Elena Gorokhova b , Zita R. Gasi unait _ e a , Art uras Razinkovas a a Coastal Research and Planning Institute, Klaip _ eda University, Manto 84, LT-92294 Klaip _ eda, Lithuania b Department of Systems Ecology, Stockholm University, SE 10691 Stockholm, Sweden Received 20 September 2006; accepted 18 December 2006 Available online 7 February 2007 Abstract To evaluate the contribution of mesozooplankton to the diet of a coastal mysid Paramysis lacustris in the Curonian Lagoon, stable isotope analysis of mysids and mesozooplankton was conducted from July to October 2004. Nitrogen isotopic composition (d 15 N) of both mysid bodies and dissected stomachs was analyzed and their d 15 N values were compared to those in ambient mesozooplankton. We also evaluated changes in phyto- and mesozooplankton assemblages during the study period and related the community dominance structure to the isotopic composition of the mesozooplankton and mysids. Stable isotope data suggest that during the ontogenetic development, mysid diet changes to include a larger proportion of mesozooplankton and that mysids need to attain a threshold size of 8.7 0.7 mm to feed on mesozooplankton. In adult mysids, d 15 N values of the dissected stomachs and stomach content were indistinguishable from those of ambient mesozooplankton, indicating nearly complete reliance on mesozooplankton prey. A significant depletion of 15 N in both mesozooplankton and mysids occurred during the study period, correlating with cyanobacteria abundance in the lagoon. These findings contribute to the understanding of P. lacustris ’ feeding biology, and the pelagic food web structure and functioning in the coastal environment. Ó 2007 Elsevier Ltd. All rights reserved. Keywords: nitrogen isotopes; trophic relationships; cyanobacteria; Lithuania; South-Eastern Baltic Sea; Curonian Lagoon 1. Introduction Mysids are generally considered to be omnivorous crusta- ceans feeding on phytoplankton, detritus, zooplankton and small benthic animals in lakes and estuaries (Mauchline, 1980). There are also numerous observations that the compo- sition of mysid diet changes during their ontogeny (life-history omnivory), with herbivory dominating among juveniles and carnivory among adults (Branstrator et al., 2000; Viherluoto et al., 2000; Viherluoto and Viitasalo, 2001). The broad diet implies a variety of possible functional roles in ecosystem food webs and points to difficulties in predicting their trophic linkages and in extrapolating from one ecosystem to another. In particular, mysids are important in consumption of sus- pended matter in the detritus-based estuarine food webs (Fock- edey and Mees, 1999), bentho-pelagic coupling by diel vertical migrations (Lasenby and Shi, 2004) and regulation of zooplankton community structure (Rudstam et al., 1989; Jane et al., 1996; Ketelaars et al., 1999; Spencer et al., 1999). To support a commercial fishery, the Ponto-Caspian mysids of Paramysis genus were extensively introduced into many inland water reservoirs of Russia (Komarova, 1991 and the ref- erences therein) as well as of former Soviet countries, includ- ing Lithuania (Arba ciauskas, 2002). In feeding experiments conducted prior to their introduction, Paramysis lacustris showed preferential feeding on detritus and phytoplankton * Corresponding author. E-mail address: [email protected] (J. Lesutien _ e). 0272-7714/$ - see front matter Ó 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.ecss.2006.12.010 Estuarine, Coastal and Shelf Science 73 (2007) 73e80 www.elsevier.com/locate/ecss
Transcript of Isotopic evidence for zooplankton as an important food source for the mysid Paramysis lacustris in...

Estuarine, Coastal and Shelf Science 73 (2007) 73e80www.elsevier.com/locate/ecss
Isotopic evidence for zooplankton as an important food sourcefor the mysid Paramysis lacustris in the Curonian Lagoon,
the South-Eastern Baltic Sea
J�urat _e Lesutien _e a,*, Elena Gorokhova b, Zita R. Gasi�unait _e a, Art�uras Razinkovas a
a Coastal Research and Planning Institute, Klaip_eda University, Manto 84, LT-92294 Klaip_eda, Lithuaniab Department of Systems Ecology, Stockholm University, SE 10691 Stockholm, Sweden
Received 20 September 2006; accepted 18 December 2006
Available online 7 February 2007
Abstract
To evaluate the contribution of mesozooplankton to the diet of a coastal mysid Paramysis lacustris in the Curonian Lagoon, stable isotopeanalysis of mysids and mesozooplankton was conducted from July to October 2004. Nitrogen isotopic composition (d15N) of both mysid bodiesand dissected stomachs was analyzed and their d15N values were compared to those in ambient mesozooplankton. We also evaluated changes inphyto- and mesozooplankton assemblages during the study period and related the community dominance structure to the isotopic composition ofthe mesozooplankton and mysids. Stable isotope data suggest that during the ontogenetic development, mysid diet changes to include a largerproportion of mesozooplankton and that mysids need to attain a threshold size of 8.7� 0.7 mm to feed on mesozooplankton. In adult mysids,d15N values of the dissected stomachs and stomach content were indistinguishable from those of ambient mesozooplankton, indicating nearlycomplete reliance on mesozooplankton prey. A significant depletion of 15N in both mesozooplankton and mysids occurred during the studyperiod, correlating with cyanobacteria abundance in the lagoon. These findings contribute to the understanding of P. lacustris’ feeding biology,and the pelagic food web structure and functioning in the coastal environment.� 2007 Elsevier Ltd. All rights reserved.
Keywords: nitrogen isotopes; trophic relationships; cyanobacteria; Lithuania; South-Eastern Baltic Sea; Curonian Lagoon
1. Introduction
Mysids are generally considered to be omnivorous crusta-ceans feeding on phytoplankton, detritus, zooplankton andsmall benthic animals in lakes and estuaries (Mauchline,1980). There are also numerous observations that the compo-sition of mysid diet changes during their ontogeny (life-historyomnivory), with herbivory dominating among juveniles andcarnivory among adults (Branstrator et al., 2000; Viherluotoet al., 2000; Viherluoto and Viitasalo, 2001). The broad dietimplies a variety of possible functional roles in ecosystem
* Corresponding author.
E-mail address: [email protected] (J. Lesutien_e).
0272-7714/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.ecss.2006.12.010
food webs and points to difficulties in predicting their trophiclinkages and in extrapolating from one ecosystem to another.In particular, mysids are important in consumption of sus-pended matter in the detritus-based estuarine food webs (Fock-edey and Mees, 1999), bentho-pelagic coupling by dielvertical migrations (Lasenby and Shi, 2004) and regulationof zooplankton community structure (Rudstam et al., 1989;Jane et al., 1996; Ketelaars et al., 1999; Spencer et al., 1999).
To support a commercial fishery, the Ponto-Caspian mysidsof Paramysis genus were extensively introduced into manyinland water reservoirs of Russia (Komarova, 1991 and the ref-erences therein) as well as of former Soviet countries, includ-ing Lithuania (Arba�ciauskas, 2002). In feeding experimentsconducted prior to their introduction, Paramysis lacustrisshowed preferential feeding on detritus and phytoplankton

74 J. Lesutien_e et al. / Estuarine, Coastal and Shelf Science 73 (2007) 73e80
(Komarova, 1991) and it was therefore expected to improvethe efficiency of detritus and primary production utilizationin the eutrophic Curonian Lagoon. Contrary to these expecta-tions, however, the stomach content analysis of field-collectedP. lacustris has revealed a significant proportion of planktoniccrustaceans, especially during nocturnal vertical migrations,suggesting that mysids feed actively on mesozooplankton(Jankauskien _e, 2003).
Assessing the contribution of a particular food source tomysid diet is a difficult task, because mysids macerate theirprey beyond recognition and traditional gut content analysisoften fails to reliably quantify prey types in the stomach con-tent (Mauchline, 1980). In such cases, stable isotope analysis(SIA) can provide a useful alternative and give insights intothe feeding relationships between the organisms within a givenfood web (Minagawa and Wada, 1984; Post, 2002). Thisapproach has been successfully applied to study trophic inter-actions in a variety of organisms, including mysids and zoo-plankton (Toda and Wada, 1990; Hansson et al., 1997;Gorokhova and Hansson, 1999; Branstrator et al., 2000; Jo-hansson et al., 2001).
This study examined changes in nitrogen isotopic composi-tion (d15N) of different size classes of Paramysis lacustris andcrustacean mesozooplankton in the Curonian Lagoon duringthe growth season. The primary objectives were to evaluateontogenetic and seasonal variations of d15N in mysids, andto ascertain the contribution of mesozooplankton to the dietof P. lacustris. We also evaluated the usefulness of analyzingisotopic composition of dissected mysid stomachs andcompared bolus d15N values with those of ambient mesozoo-plankton. When evaluating seasonal pattern in mysid and mes-ozooplankton d15N, we expected to observe depletion relatedto the development of cyanobacteria, a phenomenon previ-ously reported from other estuaries, including the Baltic Sea(Rolff, 2000).
2. Methods
2.1. Study site and species
The central part of the Curonian Lagoon is greatly influ-enced by the Nemunas River inflow, which together withwind-induced re-suspension of bottom sediments causes highwater turbidity (Zaromskis, 1996). Paramysis lacustris domi-nates mysid assemblages (Razinkovas, 1996), while mesozoo-plankton community is composed of Daphnia, Bosmina,Diaphanosoma, Chydorus, Cyclops and Eudiaptomus species(Gasi�unait _e, 2000). The detailed composition and seasonaldynamic of mesozooplankton community are described else-where (Gasi�unait _e and Razinkovas, 2004).
2.2. Sampling
Mysids, zooplankton and phytoplankton samples werecollected from July to October 2004, in the central part ofthe Curonian Lagoon (55�N 200 5600 21�E 100 4600; bottomdepth 2e3 m; Fig. 1). A sledge net with a 60� 20 cm mouth
and 1000-mm mesh size was used to collect mysids. On eachsampling occasion, 3 replicate tows were performed toreduce variation due to the patchy distribution of the mysids.Zooplankton samples were collected using a 10-L waterbottle. Samples were taken from 0.5, 1, and 2 m depths, con-centrated by filtering through a 70-mm net and then pooledtogether. Mysid and zooplankton samples were fixed with4% formaldehyde solution and stored for 6 months atroom temperature until the analyses. The phytoplanktonwere collected weekly or bi-weekly with a 10-L water bottlefrom the middle part of the water column; for microscopicanalysis, a 300 mL sample was preserved with acid Lugol’ssolution.
2.3. Microscopic analysis
Cell density of cyanobacteria (each 100 mm of filamentsand colonies of coccoids considered as a counting unit) wascounted using a MEIJI inverted microscope at 300x, 600xmagnifications according to the HELCOM recommendations(HELCOM, 1988). The zooplankton samples were analyzedusing an Olympus SZ40 stereomicroscope equipped with anocular micrometer. Mesozooplankton (0.2e2 mm; HELCOM,1988) were identified to genus, counted, and length, measured.Daphnia individuals were divided into two size classes: small(carapace length< 1.25 mm) and large (>1.25 mm). The indi-vidual weight of each specimen was calculated from its bodylength using allometric equations (Rumohr et al., 1987; Jor-gensen et al., 1995); total biomass expressed as wet weight
Fig. 1. Map of sampling stations; filled triangle e sampling station of this
study, open triangle e monitoring station of Marine Research Center (MRC).

75J. Lesutien_e et al. / Estuarine, Coastal and Shelf Science 73 (2007) 73e80
(WW, mg) was calculated from abundance data and individualweights.
2.4. Sample preparation for SIA (stable isotope analysis)
For SIA, formaldehyde-preserved mysid and mesozoo-plankton samples were used as no significant effects of form-aldehyde on d15N signatures were previously found for smallcrustaceans (Gorokhova et al., 2005). In preparation for SIA,mysids were rinsed with distilled water, and their body lengthswere measured under a dissecting microscope with an ocularmicrometer. To derive isotopic composition of the bolus atthe time of collection, stomachs of adult mysids were dis-sected from the dorsal side, transferred to tin capsules (2e3analytical replicates per size group, 6e7 stomachs perreplicate), and processed separately from the bodies. Thebody length (BL, mm) was measured from the tip of rostrumto the end of telson. All specimens were classified into 4 sizegroups: juveniles (<5 mm; grand mean� SD: 4.2� 0.5 mm),subadults (5e7 mm; 6.3� 0.4 mm), small adults (7e10 mm;8.7� 0.7 mm), and large adults (>10 mm; 11.2� 0.8 mm);see Table 1 for BL variation on each sampling occasion.From each collection, 10e20 randomly selected individualsper group were used, depending on availability. Wholeshrimps (juveniles and subadults) or abdominal tissue (adults)were used to derive body isotopic signatures; mysid abdomenwas assumed to represent muscle tissue of the animal. Allsamples were dried at 60 �C for 48 h, grinded in an agate mor-tar, and assigned to 5 analytical replicates, 3e4 individuals perreplicate. The dry mass of the samples was determined to thenearest microgram using a Sartorius M3P microbalance.
Zooplankton samples were filtered through a 200-mm nylonmesh and rinsed with tap water. Using a dissecting micro-scope, specimens of Leptodora kindtii, visible debris occasion-ally found in zooplankton samples as well as large filaments ofcyanobacteria were removed with forceps. Residual water wasremoved from underneath the sieve with paper tissue. Bulksamples of crustaceans were then transferred to pre-weighedtin capsules (1e5 replicates for each sampling occasion), driedat 60 �C for 24 h and stored desiccated until the analysis.When present in sufficient numbers, L. kindtii, which is knownas an obligate zooplanktivore (Morduchai-Boltovskoi and Riv-ier, 1987), was analyzed separately; the samples containingabout 40 individuals were processed following the same pro-cedure as zooplankton samples.
2.5. SIA (stable isotope analysis)
Concentrations of 15N, and 14N in the samples were deter-mined using continuous-flow isotope mass spectrometry pro-vided in Automated NC Analysis (ANCA) SL 20-20, PDZEuropa at the Stable Isotope Facility, UC Davis, USA. Thestandard reference material was atmospheric N2. Isotope ratioswere expressed as parts-per-thousand (&) differences from thestandard reference material (Peterson and Fry, 1987).Repeated analyses of homogeneous material yielded standarddeviations of less than 0.05%.
2.6. Statistics and calculations
Unless specified otherwise, data are presented as meansalong with standard deviations. The Pearson correlation coef-ficient was calculated for untransformed d15N mesozooplank-ton values and square root transformed cyanobacteria densitydata. A one-way ANOVA was used to test the effect of sam-pling date on d15N values; the non-replicated measurement(October 13) was excluded from the analysis. A two-wayANOVA was applied to test effects of sampling date and mysidBL on the d15N values. The homogeneity of variance wastested with the Levene’s test and normality was checked byvisual exploration of probability plots. The lack of 2 replicatesof juvenile mysids on July 17 (Table 1) and 3 replicates ofmesozooplankton on September 26 was considered to havelittle effect on the ANOVA balance (Underwood, 1997). TheFisher’s least significant difference (LSD) procedure wasapplied as an ANOVA post hoc test.
For adults, we calculated d15N of the bolus using isotopicsignatures of mysid stomachs and muscle tissue assumingstomach tissue has the same isotopic composition as muscletissue from the abdomen. The mass of the stomach tissue inthe full mysid stomach was assumed to be 25% of the stomachmass as was experimentally determined for Mysis mixta in thesame size range (Gorokhova, unpubl.). As the stomach full-ness would affect the proportion of the stomach tissue Nin the bulk (i.e., stomachþ bolus), we investigated two possi-ble scenarios: (1) d15Nbolus¼ (d15Nstomach� 0.25� d15Ntissue)/0.75, assuming the 100% gut fullness and (2) d15Nbolus¼(d15Nstomach� 0.4� d15Ntissue)/0.6, assuming 50% gut full-ness. In these calculations, SE was determined by first-ordererror propagation of uncertainties. To compare bolus d15Nvalues with those of mesozooplankton as well as bolus d15Nvalues in different size groups, we used a paired t test withsampling date as a matching factor.
3. Results
3.1. The seasonal dynamic of cyanobacteria
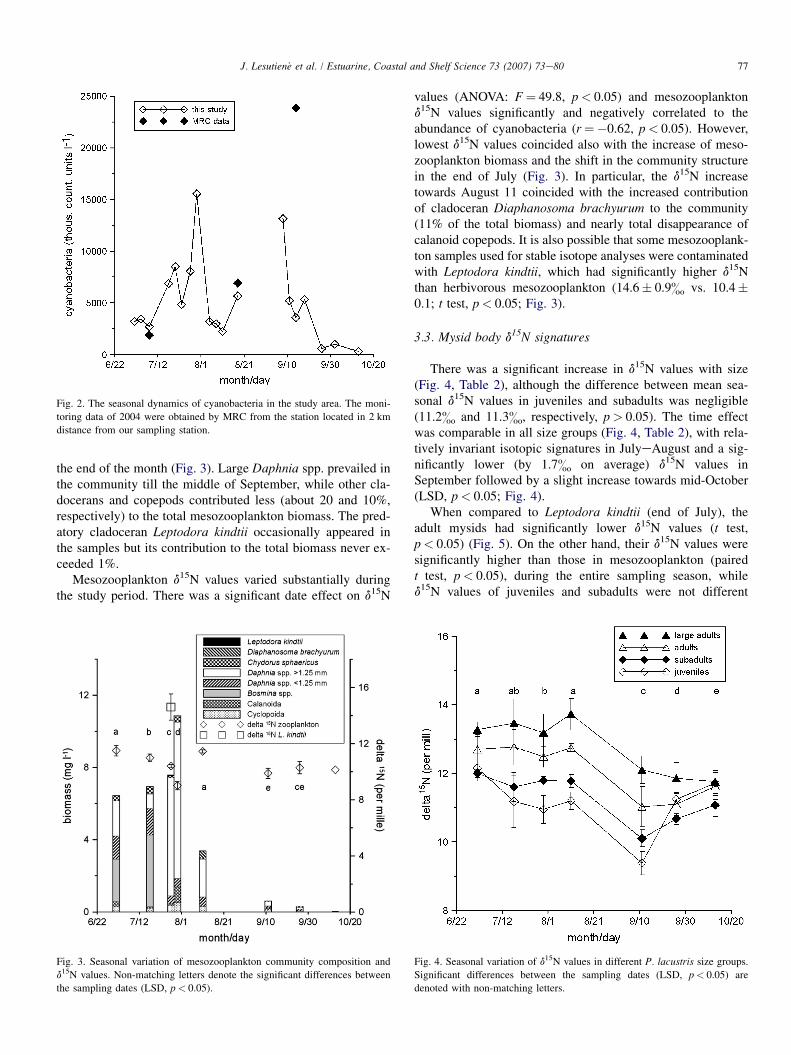
The assemblages of cyanobacteria were dominated byAphanizomenon flos-aquae, Microcystis aeruginosa and Ana-baena flos-aquae. The cyanobacteria abundance peaked twice ein the end of July and beginning of September (Fig. 2). FromAugust 18 to September 8, the sampling was interrupted, whichmight have resulted in the underestimated magnitude of the sec-ond peak. Indeed, the national monitoring in the area reportedthe cyanobacteria density as being considerably higher thanthat observed in late September (Fig. 2).
3.2. Mesozooplankton composition and d15N signatures
Mesozooplankton biomass peaked at the end of July(10.87 mg L�1) and dropped to less than 0.02 mg L�1 in Octo-ber. The mesozooplankton community was dominated by Bos-mina spp. and small Daphnia spp. in the first half of July andswitched to dominance by large-bodied Daphnia spp. towards
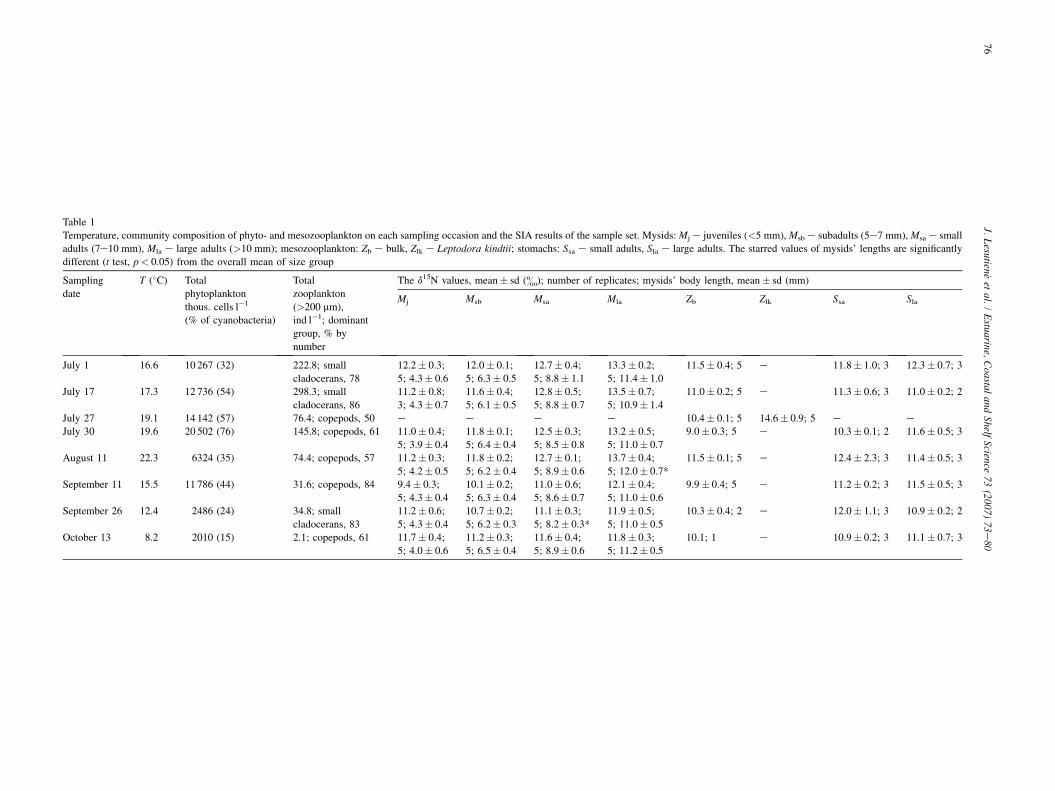
Table 1
Temperature, community ample set. Mysids: Mj e juveniles (<5 mm), Msb e subadults (5e7 mm), Msa e small
adults (7e10 mm), Mla e all adults, Sla e large adults. The starred values of mysids’ lengths are significantly
different (t test, p< 0.05
Sampling
date
T (�C) replicates; mysids’ body length, mean� sd (mm)
Mla Zb Zlk Ssa Sla
July 1 16.6 13.3� 0.2;
5; 11.4� 1.0
11.5� 0.4; 5 e 11.8� 1.0; 3 12.3� 0.7; 3
July 17 17.3 13.5� 0.7;
5; 10.9� 1.4
11.0� 0.2; 5 e 11.3� 0.6; 3 11.0� 0.2; 2
July 27 19.1 e 10.4� 0.1; 5 14.6� 0.9; 5 e eJuly 30 19.6 13.2� 0.5;
5; 11.0� 0.7
9.0� 0.3; 5 e 10.3� 0.1; 2 11.6� 0.5; 3
August 11 22.3 13.7� 0.4;
5; 12.0� 0.7*
11.5� 0.1; 5 e 12.4� 2.3; 3 11.4� 0.5; 3
September 11 15.5 12.1� 0.4;
5; 11.0� 0.6
9.9� 0.4; 5 e 11.2� 0.2; 3 11.5� 0.5; 3
September 26 12.4 11.9� 0.5;
5; 11.0� 0.5
10.3� 0.4; 2 e 12.0� 1.1; 3 10.9� 0.2; 2
October 13 8.2 11.8� 0.3;
5; 11.2� 0.5
10.1; 1 e 10.9� 0.2; 3 11.1� 0.7; 3
76
J.L
esutien_e
etal.
/E
stuarine,C
oastaland
ShelfScience
73(2007)
73e80
composition of phyto- and mesozooplankton on each sampling occasion and the SIA results of the s
large adults (>10 mm); mesozooplankton: Zb e bulk, Zlk e Leptodora kindtii; stomachs: Ssa e sm
) from the overall mean of size group
Total
phytoplankton
thous. cells l�1
(% of cyanobacteria)
Total
zooplankton
(>200 mm),
ind l�1; dominant
group, % by
number
The d15N values, mean� sd (&); number of
Mj Msb Msa
10 267 (32) 222.8; small
cladocerans, 78
12.2� 0.3;
5; 4.3� 0.6
12.0� 0.1;
5; 6.3� 0.5
12.7� 0.4;
5; 8.8� 1.1
12 736 (54) 298.3; small
cladocerans, 86
11.2� 0.8;
3; 4.3� 0.7
11.6� 0.4;
5; 6.1� 0.5
12.8� 0.5;
5; 8.8� 0.7
14 142 (57) 76.4; copepods, 50 e e e20 502 (76) 145.8; copepods, 61 11.0� 0.4;
5; 3.9� 0.4
11.8� 0.1;
5; 6.4� 0.4
12.5� 0.3;
5; 8.5� 0.8
6324 (35) 74.4; copepods, 57 11.2� 0.3;
5; 4.2� 0.5
11.8� 0.2;
5; 6.2� 0.4
12.7� 0.1;
5; 8.9� 0.6
11 786 (44) 31.6; copepods, 84 9.4� 0.3;
5; 4.3� 0.4
10.1� 0.2;
5; 6.3� 0.4
11.0� 0.6;
5; 8.6� 0.7
2486 (24) 34.8; small
cladocerans, 83
11.2� 0.6;
5; 4.3� 0.4
10.7� 0.2;
5; 6.2� 0.3
11.1� 0.3;
5; 8.2� 0.3*
2010 (15) 2.1; copepods, 61 11.7� 0.4;
5; 4.0� 0.6
11.2� 0.3;
5; 6.5� 0.4
11.6� 0.4;
5; 8.9� 0.6
77J. Lesutien_e et al. / Estuarine, Coastal and Shelf Science 73 (2007) 73e80
the end of the month (Fig. 3). Large Daphnia spp. prevailed inthe community till the middle of September, while other cla-docerans and copepods contributed less (about 20 and 10%,respectively) to the total mesozooplankton biomass. The pred-atory cladoceran Leptodora kindtii occasionally appeared inthe samples but its contribution to the total biomass never ex-ceeded 1%.
Mesozooplankton d15N values varied substantially duringthe study period. There was a significant date effect on d15N
Fig. 2. The seasonal dynamics of cyanobacteria in the study area. The moni-
toring data of 2004 were obtained by MRC from the station located in 2 km
distance from our sampling station.
Fig. 3. Seasonal variation of mesozooplankton community composition and
d15N values. Non-matching letters denote the significant differences between
the sampling dates (LSD, p< 0.05).
values (ANOVA: F¼ 49.8, p< 0.05) and mesozooplanktond15N values significantly and negatively correlated to theabundance of cyanobacteria (r¼�0.62, p< 0.05). However,lowest d15N values coincided also with the increase of meso-zooplankton biomass and the shift in the community structurein the end of July (Fig. 3). In particular, the d15N increasetowards August 11 coincided with the increased contributionof cladoceran Diaphanosoma brachyurum to the community(11% of the total biomass) and nearly total disappearance ofcalanoid copepods. It is also possible that some mesozooplank-ton samples used for stable isotope analyses were contaminatedwith Leptodora kindtii, which had significantly higher d15Nthan herbivorous mesozooplankton (14.6� 0.9& vs. 10.4�0.1; t test, p< 0.05; Fig. 3).
3.3. Mysid body d15N signatures
There was a significant increase in d15N values with size(Fig. 4, Table 2), although the difference between mean sea-sonal d15N values in juveniles and subadults was negligible(11.2& and 11.3&, respectively, p> 0.05). The time effectwas comparable in all size groups (Fig. 4, Table 2), with rela-tively invariant isotopic signatures in JulyeAugust and a sig-nificantly lower (by 1.7& on average) d15N values inSeptember followed by a slight increase towards mid-October(LSD, p< 0.05; Fig. 4).
When compared to Leptodora kindtii (end of July), theadult mysids had significantly lower d15N values (t test,p< 0.05) (Fig. 5). On the other hand, their d15N values weresignificantly higher than those in mesozooplankton (pairedt test, p< 0.05), during the entire sampling season, whiled15N values of juveniles and subadults were not different
Fig. 4. Seasonal variation of d15N values in different P. lacustris size groups.
Significant differences between the sampling dates (LSD, p< 0.05) are
denoted with non-matching letters.
78 J. Lesutien_e et al. / Estuarine, Coastal and Shelf Science 73 (2007) 73e80
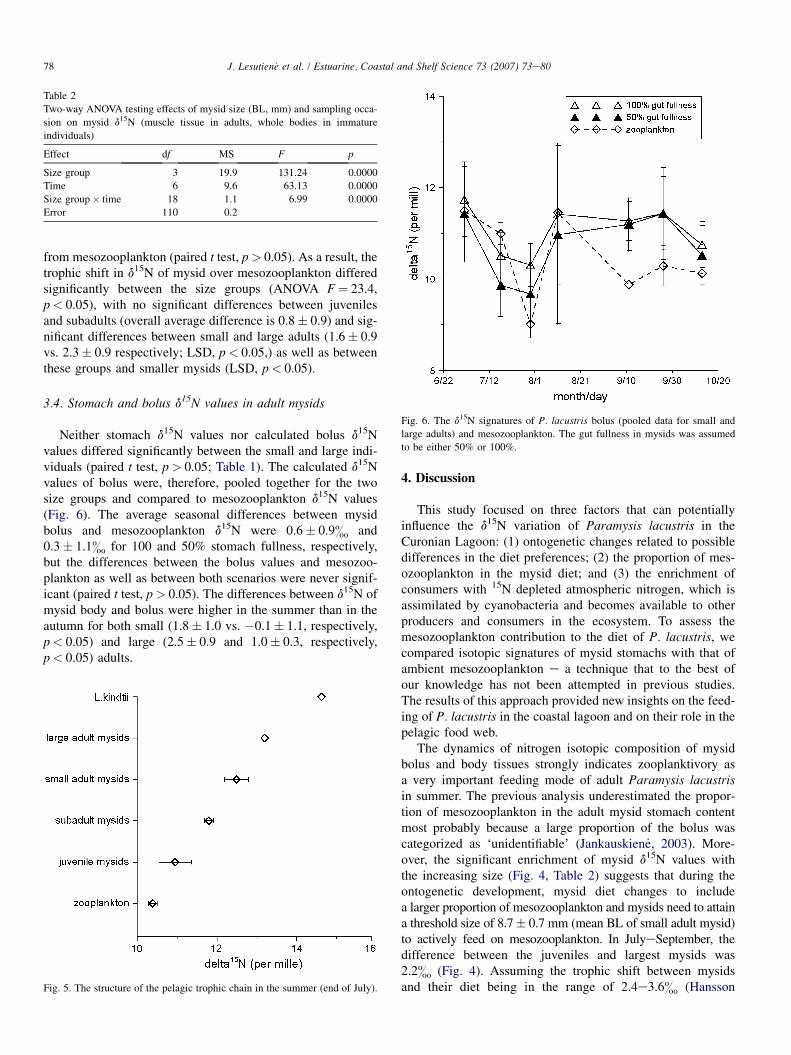
from mesozooplankton (paired t test, p> 0.05). As a result, thetrophic shift in d15N of mysid over mesozooplankton differedsignificantly between the size groups (ANOVA F¼ 23.4,p< 0.05), with no significant differences between juvenilesand subadults (overall average difference is 0.8� 0.9) and sig-nificant differences between small and large adults (1.6� 0.9vs. 2.3� 0.9 respectively; LSD, p< 0.05,) as well as betweenthese groups and smaller mysids (LSD, p< 0.05).
3.4. Stomach and bolus d15N values in adult mysids
Neither stomach d15N values nor calculated bolus d15Nvalues differed significantly between the small and large indi-viduals (paired t test, p> 0.05; Table 1). The calculated d15Nvalues of bolus were, therefore, pooled together for the twosize groups and compared to mesozooplankton d15N values(Fig. 6). The average seasonal differences between mysidbolus and mesozooplankton d15N were 0.6� 0.9& and0.3� 1.1& for 100 and 50% stomach fullness, respectively,but the differences between the bolus values and mesozoo-plankton as well as between both scenarios were never signif-icant (paired t test, p> 0.05). The differences between d15N ofmysid body and bolus were higher in the summer than in theautumn for both small (1.8� 1.0 vs. �0.1� 1.1, respectively,p< 0.05) and large (2.5� 0.9 and 1.0� 0.3, respectively,p< 0.05) adults.
Table 2
Two-way ANOVA testing effects of mysid size (BL, mm) and sampling occa-
sion on mysid d15N (muscle tissue in adults, whole bodies in immature
individuals)
Effect df MS F p
Size group 3 19.9 131.24 0.0000
Time 6 9.6 63.13 0.0000
Size group� time 18 1.1 6.99 0.0000
Error 110 0.2
Fig. 5. The structure of the pelagic trophic chain in the summer (end of July).
4. Discussion
This study focused on three factors that can potentiallyinfluence the d15N variation of Paramysis lacustris in theCuronian Lagoon: (1) ontogenetic changes related to possibledifferences in the diet preferences; (2) the proportion of mes-ozooplankton in the mysid diet; and (3) the enrichment ofconsumers with 15N depleted atmospheric nitrogen, which isassimilated by cyanobacteria and becomes available to otherproducers and consumers in the ecosystem. To assess themesozooplankton contribution to the diet of P. lacustris, wecompared isotopic signatures of mysid stomachs with that ofambient mesozooplankton e a technique that to the best ofour knowledge has not been attempted in previous studies.The results of this approach provided new insights on the feed-ing of P. lacustris in the coastal lagoon and on their role in thepelagic food web.
The dynamics of nitrogen isotopic composition of mysidbolus and body tissues strongly indicates zooplanktivory asa very important feeding mode of adult Paramysis lacustrisin summer. The previous analysis underestimated the propor-tion of mesozooplankton in the adult mysid stomach contentmost probably because a large proportion of the bolus wascategorized as ‘unidentifiable’ (Jankauskien _e, 2003). More-over, the significant enrichment of mysid d15N values withthe increasing size (Fig. 4, Table 2) suggests that during theontogenetic development, mysid diet changes to includea larger proportion of mesozooplankton and mysids need to attaina threshold size of 8.7� 0.7 mm (mean BL of small adult mysid)to actively feed on mesozooplankton. In JulyeSeptember, thedifference between the juveniles and largest mysids was2.2& (Fig. 4). Assuming the trophic shift between mysidsand their diet being in the range of 2.4e3.6& (Hansson
Fig. 6. The d15N signatures of P. lacustris bolus (pooled data for small and
large adults) and mesozooplankton. The gut fullness in mysids was assumed
to be either 50% or 100%.

79J. Lesutien_e et al. / Estuarine, Coastal and Shelf Science 73 (2007) 73e80
et al., 1997; Gorokhova and Hansson, 1999), the trophic distancebetween the diet of the smallest and largest individuals of P. la-custris is nearly as high as one trophic level. This suggests thatmysid diet is composed largely by phytoplankton in juvenilesand by mesozooplankton in adults, whereas immature or sub-adult individuals have mixed diets (Froneman, 2001, this study).The similar ontogenetic changes of d15N were documented fortwo other Baltic mysids, Neomysis integer and Mysis mixta(Gorokhova and Hansson, 1999), as well as for Mysis relictain the North American Great Lakes (Branstrator et al., 2000).In the North American lakes, M. relicta was found to becomemore carnivorous in conjunction with maturation (>10 mm),however, the time of the shift varied from one lake to another,which was suggested to be related to the size spectra ofphytoplankton (and perhaps mesozooplankton) in these lakes(Branstrator et al., 2000). The lower threshold size for zooplank-tivory observed in the Baltic P. lacustris could be explained bythe high mesozooplankton prey availability in the eutrophicCuronian Lagoon and high relative abundance of microzoo-plankton with rotifers, nauplii and small cladocerans. Asthe d15N values did not differ significantly between juvenilesand subadults, the immature P. lacustris (BL< 8.7� 0.7 mm)may be considered as a homogenous, largely herbivorous groupin feeding studies.
Our approach to investigate mysid stomachs separatelyfrom the body provides a new tool to assess immediate dietisotopic composition and to compare it with that of potentialprey. Indeed, analysis of dissected stomachs of Paramysislacustris revealed a good correspondence of their isotopiccomposition to the ambient mesozooplankton (Fig. 6); this in-dicates a very high contribution of mesozooplankton to thediet of adult mysids and lower contribution of other sources.Moreover, the correspondence was not distorted by variationsin the stomach fullness degree (50 vs. 100%; Fig. 6). A moreprecise estimation of stomach tissue proportion as a functionof the stomach fullness coupled with an assessment of thestomach fullness in concert with stable isotope analysis islikely to improve the accuracy of the method, particularly,when multiple food sources are considered.
Interestingly, the bolus d15N values differed more fromthose of mesozooplankton in autumn than during summer(Fig. 6). This was likely related to seasonal changes in preyavailability, which is considered to be as important a factorfor mysid feeding as their body size (Viherluoto et al.,2000). The opportunistic feeding causes the typical positivecorrelation between the proportion of the prey in mysid stom-achs and its densities in the environment (Fockedey and Mees,1999). In our study, the mesozooplankton abundance and bio-mass were highest in the end of July and lowest in October, thedifference being 70- and 580-fold, respectively (Table 1,Fig. 3). Moreover, the structure of the summer mesozooplank-ton community shifted from the dominance of small cladoc-erans in July to more abundant copepods and appearance ofDiaphanosoma brachyurum in August. Although Daphniaspp. did not dominate the community by number, it contrib-uted greatly to the total mesozooplankton biomass, and hence,influenced the d15N values of bulk mesozooplankton (Fig. 3).
The similarity of bolus d15N values with that of mesozoo-plankton suggests Daphnia spp. as the preferred mysid preyin summer. In autumn, when mesozooplankton becomes lessabundant, mysids might shift to other food sources (e.g., ben-thic invertebrates), resulting in the greater discrepancy be-tween bolus isotopic composition of mysids and that ofmesozooplankton.
The seasonal development of cyanobacteria communitymight also have influenced the d15N values of both the mysidsand mesozooplankton. The moderately strong negativecorrelation between cyanobacteria abundance and meso-zooplankton d15N suggests the direct consumption or use ofimmediately recycled cyanobacterial nitrogen by mesozoo-plankton. It has been shown that 15N depleted nitrogen ofcyanobacteria can reach the higher plankton trophic levelsvia direct consumption of cyanobacteria by cladocerans orsecondary consumption of dissolved organic nitrogen by otherplankton groups (Rolff, 2000). The significant drop of d15N inSeptember, pronounced in all size groups of mysids, could beattributed to the second, probably more intense, cyanobacteriabloom in the end of Augustefirst half of September (Fig. 2).As muscle tissue in similarly sized mysids integrates the iso-topic signal over 6e8 weeks (Gorokhova and Hansson,1999), the body d15N observed in SeptembereOctober islikely to reflect mysid diet in August, i.e., the time of thebloom. In SeptembereOctober, 15N depleted nitrogen wassedimented or washed out from the ecosystem, thus explainingthe observed levels of d15N in bolus, mesozooplankton andnewly hatched juveniles in the end of September and October(Figs. 3, 4, and 6).
A trophic structure of the summer pelagic food web couldbe outlined as follows. Since Leptodora kindtii is a true zoo-planktivore mainly feeding on cladocerans (Morduchai-Boltovskoi and Rivier, 1987), it can serve as a referencefor characterizing the diet of mysids in the Curonian Lagoon.Its tissues ought to theoretically reflect the signature of a pri-mary carnivore in the system. In our study, immature mysids(BL< 8.7� 0.7 mm) were generally not different from mes-ozooplankton, which implies that mysids within this sizerange are likely to be largely herbivorous. On the otherhand, adults (BL� 8.7� 0.7 mm) were less enriched thanthe primary carnivore L. kindtii (Fig. 5), indicating that, inaddition to mesozooplankton, large mysids feed on otherfood sources (e.g., sediment, benthic invertebrates, etc.).Moreover, the contribution of these sources appears to in-crease towards the autumn. These estimates of the trophicposition are supported by the similarity between mysid bolusd15N and that of mesozooplankton in summer and increaseddifference between the bolus and mesozooplankton d15N inthe end of the season.
The ontogenetic and seasonal changes in Paramysis lacust-ris’ feeding must be taken into account when predicting theirtrophic linkages in the food web. The strongest influence ofmysids on mesozooplankton community is expected in sum-mer, when the mesozooplankton is most available, and alsomost valuable as a prey for fish and other zooplanktivores.What other food sources mysids use in the autumn should

80 J. Lesutien_e et al. / Estuarine, Coastal and Shelf Science 73 (2007) 73e80
be investigated further as the population of P. lacustris reachesits highest abundance during that time and becomes the keyspecies in the food web of the lagoon.
Acknowledgements
We thank Lithuanian Marine Research Center for thecyanobacteria and hydrometeorological monitoring data ofthe year 2004; A. Saskov, J. Macien _e and R. Karickait _e forfield and laboratory assistance, A. Sjosten (Systems Ecology,Stockholm University) for his kind permission to use labora-tory facilities, and Dr. David Harris (University of Californiaat Davis, Stable Isotope Facility) for stable isotope analyses.This research was funded by EU FW6 505446 MARBEF(Marine Biodiversity and Ecosystem Functioning) project.
References
Arba�ciauskas, K., 2002. Ponto-Caspian amphipods and mysids in the inland
waters of Lithuania: history of introduction, current distribution and rela-
tions with native malacostracans. In: Leppakoski, E., Gollasch, S.,
Olenin, S. (Eds.), Invasive Aquatic Species of Europe e Distribution,
Impacts and Management. Kluwer Academic Publishers, Dordrecht, pp.
104e115.
Branstrator, D.K., Cabana, G., Mazumder, A., Rasmussen, J.B., 2000. Measur-
ing life-history omnivory in the opossum shrimp, Mysis relicta, with stable
nitrogen isotopes. Limnology and Oceanography 45, 463e467.
Fockedey, N., Mees, J., 1999. Feeding of the hyperbenthic mysid Neomysis
integer in the maximum turbidity zone of the Elbe, Westerschelde and
Gironde estuaries. Journal of Marine Systems 22, 207e228.
Froneman, P.W., 2001. Feeding ecology of the mysid, Mesopodopsis
wooldridgei, in a temperate estuary along the eastern seaboard of South
Africa. Journal of Plankton Research vol. 23 (No. 9), 999e1008.
Gasi�unait _e, Z.R., 2000. Coupling of the limnetic and brackish water plankton
crustaceans in the Curonian Lagoon (Baltic Sea). International Revue of
Hydrobiology 5e6, 653e662.
Gasi�unait _e, Z.R., Razinkovas, A., 2004. Temporal and spatial patterns of
crustacean zooplankton dynamics in transitional lagoon ecosystem. Hydro-
biologia 514, 139e149.
Gorokhova, E., Hansson, S., 1999. An experimental study on variations in
stable carbon and nitrogen isotope fractionation during growth of Mysis
mixta and Neomysis integer. Canadian Journal of Fishery and Aquatic Sci-
ences 56, 2203e2210.
Gorokhova, E., Hansson, S., Hoglander, H., Andersen, C.M., 2005. Stable
isotopes show food web changes after invasion by the predatory cladoceran
Cercopagis pengoi in a Baltic Sea bay. Oecologia 143, 251e259.
Hansson, S., Hobbie, J.E., Elmgren, R., Larsson, U., Fry, B., Johansson, S.,
1997. The stable nitrogen isotope ratio as a marker of food-web interac-
tions and fish migration. Ecology 78, 2249e2257.
HELCOM, 1988. Guidelines for the Baltic Monitoring Programme for the
Third Stage. Baltic Sea Environment Proceedings No. 27D: 23e86.
Jane, M., Almond, R., Bentzen, E., Taylor, W.D., 1996. Size structure and
species composition of plankton communities in deep Ontario lakes with
and without Mysis relicta and planktivorous fish. Canadian Journal of Fish-
ery and Aquatic Sciences 53, 315e325.
Jankauskien _e, R., 2003. Selective feeding of Ponto-Caspian higher crustaceans
and fish larvae in the littoral zone of the Curonian Lagoon. Ekologija 2,
19e27.
Johansson, O.E., Leggett, M.F., Rudstam, L.G., Servos, M.R.,
Mohammadian, M.L., Gal, G., Dermott, R.M., Hesslein, R.H., 2001. Diet
of Mysis relicta in Lake Ontario as revealed by stable isotope and gut content
analysis. Canadian Journal of Fishery and Aquatic Sciences 58, 1975e1986.
Jorgensen, S.E., Nielsen, S.N., Jorgensen, L.A., 1995. Handbook of Ecological
Parameters and Ecotoxicology. Elsevier, AmsterdameLondone
New YorkeTokyo, pp. 1e1264.
Ketelaars, H.A.M., Lambregts-van de Clundert, F.E., Carpentier, C.J.,
Wagenvoort, A.J., Hoogenboezem, W., 1999. Ecological effects of the
mass occurrence of the Ponto-Caspian invader, Hemimysis anomala
G.O.Sars., 1907 (Crustacea: Mysidacea), in a freshwater storage reservoir
in the Netherlands, with notes on its autecology and new records. Hydro-
biologia 394, 233e248.
Komarova, T.I., 1991. Fauna of the Ukraine, Mysidacea. Fauna of the Ukraine
26, 1e104 (in Russian).
Lasenby, D., Shi, Y., 2004. Changes in the elemental composition of the
stomach contents of the opossum shrimp Mysis relicta during diel vertical
migration. Canadian Journal of Zoology 82, 525e528.
Mauchline, J., 1980. The biology of mysids and euphausids. Advances in
Marine Biology 18, 81e97.
Minagawa, M., Wada, E., 1984. Stepwise enrichment of 15N along food
chains: further evidence and the relation between d15N and animal age.
Geochimica et Cosmochimica Acta 48, 1135e1140.
Morduchai-Boltovskoi and Rivier, 1987. The Predativorous Cladocerans
Podonidae, Polyphemidae, Cercopagidae and Leptodoridae of the World
Fauna. The Scientific Academy USSR, Leningrad, pp. 97e106 (in Russian).
Peterson, B.J., Fry, B., 1987. Stable isotopes in ecosystem studies. Annual
Review of Ecological Systems 18, 293e320.
Post, D., 2002. Using stable isotopes to estimate trophic position: models,
methods, and assumptions. Ecology 83, 703e718.
Razinkovas, A., 1996. Spatial distribution and migration patterns of the mysids
in the Curonian Lagoon. In: Andrushaitis, A. (Ed.), Proceedings of the
13th Baltic Marine Biologists Symposium, pp. 117e120.
Rolff, C., 2000. Seasonal variation in d13C and d15N of size-fractionated
plankton at a coastal station in the northern Baltic proper. Marine Ecology
Progress Series 203, 47e65.
Rudstam, L.G., Danielsson, K., Hansson, S., Johansson, S., 1989. Diel vertical
migration and feeding patterns of Mysis mixta (Crustacea, Mysidacea) in
the Baltic Sea. Marine Biology 101, 43e52.
Rumohr, H., Brey, T., Ankar, S., 1987. A compilation of biometrics conversion
factors for benthic invertebrates of the Baltic Sea. The Baltic Marine
Biologists Publication No. 9.
Spencer, C.N., Potter, D.S., Bukantis, R.T., Stanford, J.A., 1999. Impact of
predation by Mysis relicta on zooplankton in Flathead Lake, Montana,
USA. Journal of Plankton Research 21, 51e64.
Toda, H., Wada, E., 1990. Use of 15N/14N rations to evaluate the food source of
the mysid, Neomysis intermedia Czerniawsky, in a eutrophic lake in Japan.
Hydrobiologia 194, 85e90.
Underwood, A.J., 1997. Experiments in Ecology. Cambridge University Press,
1e504.
Viherluoto, M., Kuosa, H., Flinkman, J., Viitasalo, M., 2000. Food utilisation
of pelagic mysids, Mysis mixta and M. relicta, during their growing season
in the northern Baltic Sea. Marine Biology 136, 553e559.
Viherluoto, M., Viitasalo, M., 2001. Temporal variability in functional
responses and prey selectivity of the pelagic mysid, Mysis mixta, in natural
prey assemblages. Marine Biology 138, 575e583.
Zaromskis, R., 1996. Oceans, Seas, Estuaries. Debesija Vilnius, 1e293.




















