
Evaluation and Modelling of Water Absorption Characteristics of Paddy
Upload
independentCategory
view
1download
0
Vol. 39, No. 5APPLIED AND ENVIRONMENTAL MICROBIOLOGY, May 1980, p. 1010-10180099-2240/80/05-1010/09$02.00/0
Isolation, Enumeration, and Characterization of Aeromonasfrom Polluted Waters Encountered in Diving Operations
RAMON J. SEIDLER,'t D. A. ALLEN,' H. LOCKMAN,' R. R. COLWELL,' S. W. JOSEPH,2 AND 0. P.DAILY2
Department ofMicrobiology, University ofMaryland, College Park, Maryland 20742,' and Department ofMicrobiology, Naval Medical Research Institute, Bethesda, Maryland 200142
Counts of total viable, aerobic, heterotrophic bacteria, indicator organisms, andAeromonas spp. were made at a diver training site on the Anacostia River inWashington, D.C. The numbers ofAeromonas cells in Anacostia River sedimentand water increased during periods of elevated water temperature, to maxima of4 x 105 cells per g of sediment and 300 cells per ml of water. Correspondingly,Aeromonas counts dropped 2 to 4 logs as the water temperature decreased to 0to 0.5°C. Cultures taken by sterile swabs from the ears and face masks of diversafter a 30-min swim in the Anacostia River yielded bacterial types and numberssimilar to those found in the river. The nasal passages of the divers apparentlydid not become contaminated by swimming, possibly because of the protectiveeffect of the face masks used by the divers. Properties associated with virulencein Aeromonas hydrophila and Aeromonas sobria strains isolated from the river,sediment, and divers were investigated. Nearly 40% of the strains of both speciescollected during the study produced cytotoxic activity for mouse Y-1 adrenalcells, as well as elastase. Enterotoxin activity, as detected by the Y-1 assay, wasobserved in 3% (1 of 35) of the strains of A. sobria and in 6% (19 of 330) of the A.hydrophila strains. Fluid accumulation in rabbit ileal loops induced by bothspecies of Aeromonas varied greatly among the 17 strains examined. Fluidaccumulation of at least 0.4 ml/cm was correlated with positive cytotoxin- orenterotoxin-like response in the Y-1 tissue culture assay.
It is clear that waste discharges into aquaticenvironments can have harmful effects on hu-man health. During the past 10 years, a varietyof research programs have been established tostudy the effects of pollution in freshwater, es-tuarine, and deep ocean environments (15, 19,20). A hitherto overlooked effect of pollution isthe health risk to aquatic sportsmen, as well asmilitary and commercial diving personnel whomust work in water contaminated by microor-ganisms potentially pathogenic for humans.
It has been known for many years that Aero-monas species can cause infections in fish andreptiles, as well as in warm-blooded animals.These organisms are ubiquitous in aquatic en-vironments and in warmer climates and warmwater conditions can contribute to epizootics infish (4, 11, 15). The symptomatology of a varietyof human Aeromonas infections is becomingincreasingly clear (2, 7, 8, 10-14, 21, 26, 29). Oncebelieved to be an opportunistic pathogen of lowvirulence, Aeromonas is now recognized as aprimary pathogen (8, 13, 21). Compromisedhosts may experience serious and even fatal
t On sabbatical leave from Oregon State University, Cor-vallis, OR 97331.
infections (8). Clinical strains from various infec-tions elaborate a variety of extracellular viru-lence factors, including hemolysin, cytotoxic en-zymes, and enterotoxin (2, 3, 9, 22).Human gastrointestinal and soft tissue Aero-
monas infections are often a result of water-related traumas and/or consumption of contam-inated water supplies (7, 8, 13, 26). Human in-fections associated with diving activities in pol-luted waters have been described (12, 14, 26),and indeed, during the study reported here adiver on a training exercise at the samplingstation sustained a leg wound infection causedby two Aeromonas species (17).This study documents the occurrence and sea-
sonal fluctuations of Aeromonas in the Anacos-tia River in Washington, D.C., where increasednumbers of cases of gastroenteritis and woundinfections have occurred in personnel undergo-ing diver training operations throughout theyear. Results of the work reported here establishthat a significant number of the Aeromonasisolates in the microbial population of a pollutedenvironment demonstrate multiple resistance toantibiotics, are capable of colonizing humans,and produce cytotoxic and enterotoxic sub-stances. Thus, new information concerning the
1010
on July 31, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEROMONAS IN POLLUTED WATERS 1011
potential infectious disease risk of Aeromonasspecies in aquatic environments is provided.
MATERIALS AND METHODSMeasurement ofphysical and chemical param-
eters. Dissolved oxygen and temperature were re-corded at stations located in the Anacostia River witha YSI 51B oxygen meter (Yellow Springs InstrumentCo., Inc., Yellow Springs, Ohio). Temperature andsalinity at the New York Bight stations were recordedby using an Interocean vertical probe aboard the R/VGeorge B. Kelez.
Collection of samples. Samples for study werecollected over a 10-month period during 1978 and 1979.Samples from the New York Bight, an area of highsalinity (27 to 32%o), were collected at three stations;these were near a sewage dump site (longitude, 40025.4' N; latitude, 730 48.0' W), at a dredge spoils site(longitude, 400 23.4' N; latitude, 730 51.9' W), and ata location northwest of the other two stations near themouth of New York harbor (longitude, 400 29.4' N;latitude, 730 59.2' W). Samples from the AnacostiaRiver, a freshwater site of low salinity (0 to 2%o), werecollected at the U.S. Naval Yard, Washington, D.C.Water samples were collected at three depths at
each station. A surface water sample was collected byusing a sterile carboy held 2 to 8 cm below the surface.Samples from the top of the water column (ca. 1 mbelow the surface) and at the bottom of the watercolumn (ca. 1 m above the bottom) were collected witha Niskin sampler (General Oceanics, Inc., Miami, Fla.).Sediment samples were collected by using a PetitePonar grab sampler (19).
Bacterial enumeration. At the Anacostia Riverstation, total viable, aerobic, heterotrophic counts onplate count agar (Difco) were obtained by the spreadplate technique, total viable counts of samples col-lected at the New York Bight were obtained on marinesalt yeast extract agar (30). All plates were incubatedat 25°C after inoculation, and colonies were countedat 7 days. Sediment samples were diluted with phos-phate-buffered saline (7.2 g of NaCl per liter, 1.48 g ofNa2HPO4 per liter, 0.43 g of K2HPO4 per liter, pH 7.2)before the treatment described above for water sam-ples. Direct microscopic counts were made by epiflu-orescent microscopy (16).The total and fecal coliforms in the Anacostia River
were enumerated by membrane filtration, using mediaand methods described in detail elsewhere (1). Thetotal and fecal coliforms in the samples collected inthe New York Bight were enumerated by using thethree-tube, most-probable number method (1). NewYork Bight water samples of known volume wereconcentrated by filtration through nitrocellulose filters(pore size, 0.45 pm; Gelman). Triplicate samples (300,30, and 3 ml) were filtered, and the filters were placedin tubes of lactose broth. Positive tubes (i.e., thosetubes showing acid and gas production after incuba-tion for 48 h at 35°C) were processed through thecompleted coliform test, inoculated into EC broth at44.5°C, and incubated for 24 h for the detection offecal coliforms. Sediment samples (10, 1.0, 0.1, and0.01 g of each sample) were examined for total andfecal coliforms. The 10-g samples were pipetted into
10-ml volumes of double-strength lactose broth. Di-luted sediment suspensions, which were prepared inphosphate-buffered saline, were processed as de-scribed above for water samples.
Presumptive Aeromonas species were enumeratedby pour plating samples in duplicate on PBG agar (23)for those samples for which counts were expected toexceed 10 cells per ml, and, if necessary, samples werediluted with phosphate-buffered saline. After incuba-tion for 24 h at 37°C, 100 to 200 presumptive Aero-monas colonies appearing on PBG agar were trans-ferred to Aeromonas screening medium (J. B. Kaper,R. J. Seidler, H. Lockman, and R. R. Colwell, submit-ted for publication). Strains yielding an alkaline sur-face and acid butt and demonstrating motility weretransferred from the screening medium to gelatin agar(32). Oxidase-positive, gelatin-hydrolyzing strainsgrowing in the absence of added sodium chloride wereconsidered to be confirmed Aeromonas spp. Repre-sentative strains of confirmed Aeromonas spp. werefurther tested by using the API 20E identificationsystem (Analytab Products, Inc., Plainview, N.Y.); theresults of this testing permitted identification of Aero-monas hydrophila. For those samples yielding Aero-monas counts of fewer than 10 cells per ml, enumera-tion was accomplished by a most-probable numberprocedure in which a modified Rimler-Shotts mediumwas used for initial enrichment (31). The broth me-dium did not contain L-lysine hydrochloride, L-orni-thine hydrochloride, sodium thiosulfate, or agar. Ap-propriate volumes of water and diluted sediment sam-ples were filtered by using membrane filters (pore size,0.45 pm; Millipore Corp., Bedford, Mass.), which wereplaced directly into tubes of modified Rimler-Shottsmedium after filtration was completed. The Rimler-Shotts broth tubes containing the filters were incu-bated for 24 h at 37°C. Inoculated tubes showing anacid reaction at 24 h were streaked onto MacConkeyagar plates and incubated at 37°C for 24 h. Approxi-mately 5 to 10 lactose-negative colonies were pickedfrom each plate; these colonies were transferred togelatin agar and subsequently to Aeromonas screeningmedium, as described above. The species A. hydro-phila and Aeromonas sobria were identified by usingthe scheme of Popoff and Veron (25) and tests whichhave been described previously (17). A. sobria is anew species described after publication of Bergey'sManual of Determinative Bacteriology, 8th ed. (6).
Salmonella spp. were isolated by using a three-tube, most-probable number, primary, nonselectiveenrichment involving dulcitol broth (20). Water sam-ples (300, 30, and 3 ml) were concentrated by filtrationthrough 0.45-pum membrane filters. The filters wereplaced in tubes containing 10 ml of dulcitol broth.Sediment samples (30, 3, and 0.3 g) were processedsimilarly; the 30-g sample was transferred to 100 ml ofbroth. All samples were incubated for 24 h at 37°C.Broth tubes showing growth were transferred to sele-nite cysteine broth (Difco), XLD agar (Difco), andKligler iron agar (Difco). Salmonella isolates wereconfirmed with the API 20E system (Analytab Prod-ucts, Inc.).
Vibrio cholerae and Vibrio parahaemolyticus were
isolated by using alkaline peptone and ethyl violetbroth enrichments, respectively (19). Confirmation of
VOL. 39, 1980
on July 31, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from
1012 SEIDLER ET AL.
the isolation of these species was by the API 20Esystem.
Virulence assays. Aeromonas strains were prop-agated in 10 ml of tryptic soy broth incubated staticallyat 30°C. Tests for cytotoxin and enterotoxin wereperformed with culture supernatants added to Y-1adrenal cells, as previously described (17, 19, 28). Areaction was read as enterotoxic if the morphologicalappearance after exposure to a supernatant heated at560C for 10 min resembled that observed with choleraenterotoxin. Cytotoxic activity was revealed by round-ing, shrinking, granule formation, and vacuolization ofthe cells.Rabbit ileal loop tests. Aeromonas strains were
grown for 24 h at 37°C on brain heart infusion agar(Difco). Cells were harvested and suspended in 5 ml ofbrain heart infusion broth, and 1 ml of the suspension,containing 108 to 109 cells, was inoculated into eachloop, as previously described (17,19). Rabbits weighing1 kg were used and sacrificed 8 h after loop inoculation.
Statistical analyses. Physical, chemical, and mi-crobiological data were entered in an IBM 370 com-puter, and multiple linear correlation coefficients werecomputed by using statistical program packages (19).
Antibiotic susceptibility assays. Antibiotic sus-ceptibilities were determined by the disk diffusiontechnique, as previously described (17).Sampling ofdivers. Diver training at the sampling
station included a timed 1,500-yard (ca. 1,375-tn) sur-face swim as a measure of physical fitness and swim-ming ability. Two classes of trainees were sampled forthe presence of Aeromonas spp. and other bacteria intheir noses, ears, and throats and on their masks beforeand after a timed surface swim. This permitted deter-minations of the degree to which exposure to knownconcentrations of bacteria resulted in transient and/orlong-term colonizations. Sterile swabs were used totake samples before and after the timed swim. Theswabs were placed in 2 ml of sterile normal saline andtransported on ice to our laboratory immediately aftercollection of the samples. Samples (1 and 0.1 ml) ofthe saline solutions were placed in petri dishes, and 15ml of PBG agar was added to each dish. After incu-bation for 24 h at 37°C, glycogen-fermenting colonieswere picked and identified.
RESULTSThe Anacostia River sampling site is located
in a tidal basin at the U.S. Naval Yard, Wash-ington, D.C. The depth of water at the samplingstation in the Anacostia River is ca. 20 feet (6.1m), and the width of the river in the general areaof the study is ca. 1,000 feet (305 m). Throughoutthe year, the water in the river is turbid (i.e.,Secchi disk transparency of 0.34 to 0.17 m), andthe temperature of the water fluctuates widelyon a seasonal basis, with a range of 0.5 to 28°C.Microbiological enumeration data for water andsediment samples are summarized in Fig. 1 and2. Aeromonas spp. and total coliforms exhibitedsimilar cell densities and similar fluctuationsthroughout the sampling period. The number of
EPIFLUORESCENTCOUNT (I 103)
104
103_
-i
fr12
-J-Jw
i lo,a.0
V
FCLCLIFORMS
JUL AUG SEPT OCT NOV DEC JAN FES MAR APR MAY JMMONTH
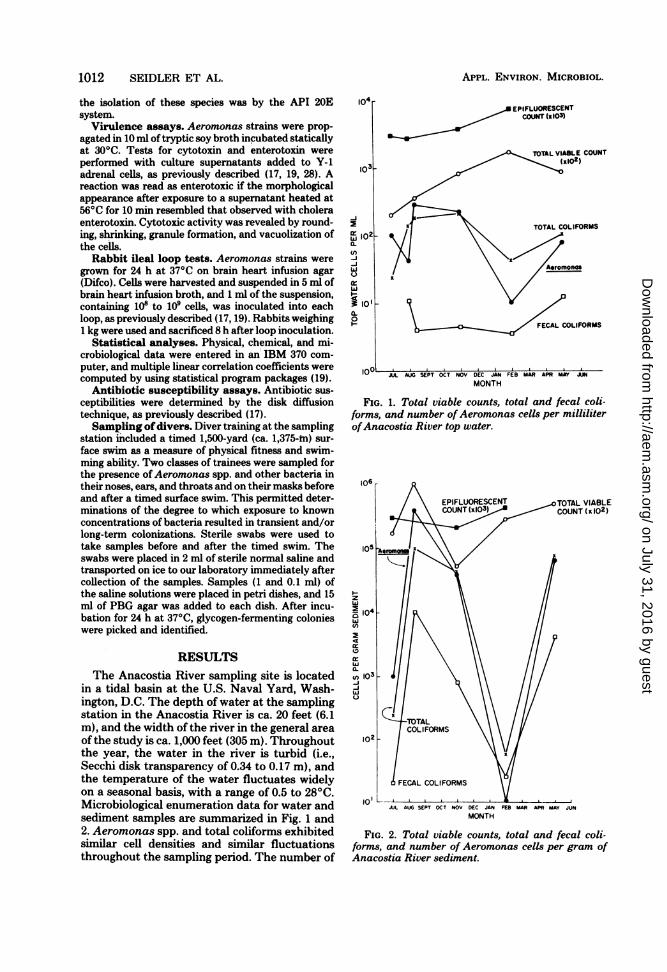
FIG. 1. Total viable counts, total and fecal coli-forms, and number ofAeromonas cells per milliliterofAnacostia River top water.
106
EPIFLUORESCENT TOTAL VIABLECOUNT(:x103) COUNT lx 102)
105 -AeM \ \:/
zw
w
CDSEw0.
-j-jw
JUL AUG SEPT OCT NOV DEC JAN FEB MAR AR MAY JUNMONTH
FIG. 2. Total viable counts, total and fecal coli-forms, and number of Aeromonas cells per gram ofAnacostia River sediment.
APPL. ENVIRON. MICROBIOL.
E
on July 31, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEROMONAS IN POLLUTED WATERS
Aeromonas spp. in the top water samplesreached 300 cells per ml in August and declinedto 10 cells per ml in February. In May, when thewater temperature was 22.50C, the Aeromonascounts began to rise.
Fecal coliform counts in the Anacostia Riverchanged very little throughout the study,whereas total viable counts increased fivefold forsamples collected in February compared withthe samples collected in August. The tempera-ture of the water was 27 to 280C during thesummer; this decreased to 10°C in Novemberand 0.50C in February, at which time there wasa 0.5-cm-thick cover of ice on the water.
Total and fecal coliforms and numbers ofAeromonas cells in the sediment exhibited dra-matic changes as the water temperaturedropped; maximum numbers were observed inAugust, when counts of Aeromonas spp. ex-ceeded 105 cells per g of sediment. The numbersof Aeromonas and indicator species declinedsignificantly; there was a 103- to 104-fold decreasefrom August 1978 to February 1979. In contrast,the total viable counts showed little or no de-tectable change, remaining at ca. 105 to 106 cellsper g of sediment. The ratio of Aeromonas tototal coliforms to fecal coliforms changed signif-icantly with decreasing water temperature. Dur-ing July and August, water samples yielded rel-ative propo;'tions averaging 100:75:5, respec-tively, wheteas in November the ratio was 100:100:2 and in February it was 100:400:30. In Maythe ratio was intermediate between the ratiosrecorded for November and February (100:120:16).The strongest correlations among the study
parameters were recorded for the bottom waterand sediment data. The correlation coefficientbetween number of Aeromonas cells and tem-perature was ca. 0.66 and, very likely, was nothigher because of the lag in the temperature-Aeromonas response observed during the Julyto November sampling period (Fig. 1 and 2).
Aeromonas counts increased from 100 to 300cells per ml during July and August, even thoughthe temperature of the river water remained at27 to 280C. The cyclic nature of the Aeromonascounts was most dramatic for the November,February, and May samples, when water tem-peratures ranging from 0.5 to 22.50C were re-corded. The correlation value for bottom watercounts of Aeromonas cells and temperature de-termined for samples collected during Augustthrough May was 0.91.Aeromonas counts for bottom water and sed-
iment yielded significant correlations with fecalcoliforms (0.90) and total coliforms (0.95 to 0.96);i.e., they were significant at the 1% level (df = n- 2 = 4; r = 0.92). Interestingly, Aeromonascounts for bottom water and sediment exhibitedan inverse correlation with concentration of dis-solved oxygen (r = -0.63 to -0.68).Large numbers of opportunistic pathogens
were recovered from the ears and diving masksof nearly all of the 15 divers after the timedswim (Table 1) when the divers were sampled60 to 80 min after leaving the water. The nosesof all divers except diver 12 appeared to bepartially protected from contamination by theface masks. Throat swabs of three divers (divers13, 14, and 15) yielded significant numbers oforganisms, but these divers yielded positive cul-tures before the swim. More than 70% of thecolonies picked from PBG agar and originatingfrom divers or their masks were identified asEnterobacter and Klebsiella spp. The other cul-tures included Aeromonas spp. (21%) and Esch-erichia coli (6%).The incidence of Aeromonas was monitored
for two separate diving groups, one group divingin August and the other diving in October. Aero-monas spp. were recovered from the ears ofmore than 90% of the divers of both groups, butwere less frequently recovered from the nosesand throats of divers participating in the Octoberswim. In October the water was colder, and
TABLE 1. Number ofglycogen-fernenting colonies appearing on PBG agar after inoculation with samplescollected from divers and their diving equipment at the Anacostia Rivera
Avg counts
Diver Before swim After 30-min swim
Mask Ear Nose Throat Mask Ear Nose Throat
1-11 0 0 0 0.4 45 136 2 512 0 0 0 0 36 200 100 213 0 0 0 25 30 100 0 10014 0 0 0 100 100 1 0 10015 0 0 0 20 100 100 0 30
a Isolates were identified as Enterobacter spp. (55%), Aeromonas spp. (21%), KlebsielUa spp. (18%), and E.coli (6%).
VOL. 39, 1980 1013
on July 31, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

1014 SEIDLER ET AL.
smaller numbers of Aeromonas were found inthe water (Table 2).Two strains of non-01 V. cholerae were iso-
lated from 250-ml samples of top and bottomwater collected on 9 August 1978. These twostrains corresponded to the Heiburg group Vphenotype (i.e., they were Voges-Proskauer andsucrose negative). Salmonella spp. were alsoisolated from sediment and water samples col-lected in the Anacostia River at the diving schoolstudy site during July, August, and May. V.cholerae was not isolated from samples collectedin the New York Bight, but Salmonella andAeromonas spp. were readily isolated from theseseawater samples. Aeromonas was present atconcentrations of approximately 1 to 10 cells perliter of water in the Bight.The biochemical properties of 48 strains of
Aeromonas isolated from Anacostia River waterand sediment and from divers after a trainingswim in the Anacostia River were determined.Although the Aeromonas strains were identifiedas A. hydrophila with the API 20E test system,different responses were exhibited among thestrains for the Voges-Proskauer test, lysine de-carboxylase, citrate utilization, and fermentationof rhamnose, sucrose, amygdalin, and arabinose.Biochemical reactions were recorded for 365
Aeromonas strains. Biotyping distinguished thespecies A. hydrophila and A. sobria. Of all Aero-monas strains recovered from the AnacostiaRiver water and sediment and the divers aftertheir timed swim, 84 to 85% were A. hydrophila.In comparison, 33 of 34 Aeromonas strains (97%)obtained from the New York Bight were iden-tified as A. hydrophila.During the summer sampling period, 190
Aeromonas strains were collected from Anacos-tia River water and sediment, and each strainwas tested for resistance to seven antibiotics.Most of the strains (183 of 190) were resistant toampicillin, 49 were resistant to erythromycin, 17were resistant to tetracycline, 3 were resistant tonalidixic acid, and 1 was resistant to kanamycin.None of the isolates was resistant to chloram-phenicol or gentamicin.Table 3 shows the results of the Y-1 mouse
APPL. ENVIRON. MICROBIOL.
adrenal cell assays for cytotoxin and enterotoxin.Approximately 39% (123 of 330) of the A. hydro-phila strains and 37% (13 of 35) of the A. sobriastrains produced cytotoxin, as judged by cellrounding, shrinking, and granule formation.When culture supernatants heated for 10 min at56°C were tested, 6% (20 of 330) of the A. hy-drophila strains and 3% (1 of 35) of the A. sobriastrains induced cell rounding without shrinkingand peripheral granule formation. Such changeswere conditionally recorded as resulting fromenterotoxin activity (21, 22).A total of 17 Aeromonas strains were exam-
ined for fluid accumulation response in the rab-bit ileal loop test (Table 4); positive controlresponses were recorded for accumulations offluid ranging from 0.6 to 0.8 ml/cm. Sterile cul-ture fluid, which served as a control for theexperiments, promoted no detectable fluid ac-cumulation. Five of the nine A. sobria strainstested induced accumulations of ca. 0.9 to 1.1ml/cm. Each of the A. sobria culture superna-tants containing a detectable enterotoxin or cy-totoxin in the Y-1 assay also induced fluid ac-cumulation in the rabbit ileal loop test.
Positive cytotoxin- or enterotoxin-induced Y-1 adrenal cell responses were noted for the fourstrains of A. hydrophila which stimulated thelargest amounts of fluid accumulation in rabbitloops (Table 4). Interestingly, three strains (2A1,F12, and HADSc) induced no morphologicalresponses in tissue culture and elicited little orno fluid accumulation in the rabbit loop test.Also, strain 9 did not produce detectable toxinsin either system. The latter strain was elastasenegative and was found to be A. hydrophilabiovar X2 (23), since it is Voges-Proskauer neg-ative and does not produce gas from glucose.
DISCUSSIONFrom a search of the primary literature, we
concluded that this is the first report to docu-ment the seasonal distribution of Aeromonasspp. in water and sediment at a location wherea human wound infection caused by Aeromonaswas sustained (17). In addition to enumeration
TABLE 2. Incidence ofAeromonas spp. recovered from divers and their diving equipment before and afterthey swam in the Anacostia River
No. of No. positive/no. testedAero- Water
Month monas temp Before swim After swimcells per (oC)b
mWa Mask Ear Nose Throat Mask Ear Nose ThroatAugust 300 28 0/15 0/15 0/15 3/15 13/15 14/15 1/15 4/15October 50 13 0/10 0/10 0/10 0/10 2/10 9/10 0/10 2/10
aCount in Anacostia River water at time of sampling.b Temperature of Anacostia River water at time of sampling.
on July 31, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEROMONAS IN POLLUTED WATERS
TABLE 3. Results of Y-1 adrenal cell assays forcytotoxin and enterotoxin production by Aeromonas
spp. isolated from samples collected at theAnacostia Rivera
% Showing positive re-No. of sponse
Species strainstested Cytotoxin Entero-
toxin
A. hydrophila 330 39 (128)b 6 (19)A. sobria 35 37 (13) 3 (1)
a A cytotoxic response was scored when the tissueculture cells exhibited shrinking, rounding, and gran-ulation. An enterotoxic response was scored whenheated (560C for 10 min) culture supernatants causedcell rounding without granule formation (18, 19, 21,22).
b Numbers in parentheses indicate the numbers ofstrains exhibiting a positive response.
of Aeromonas spp., microbiological and physi-cochemical parameters were also monitored.
Total coliform and Aeromonas populations ofthe top and bottom water and sediment ex-hibited seasonal fluctuations; counts rose duringperiods when the water temperature was ele-vated and reached a maximum when the tem-perature of the water exceeded 200C. The countsdecreased significantly in February, when thetemperature of the water dropped to 0.50C. Fe-cal coliform counts for top water samples alsoexhibited some seasonal fluctuation, but thechanges were smaller (from 400 cells per 100 mlto 1,000 cells per 100 ml). In contrast, the Aero-monas and total coliform counts ranged from2,000 cells per 100 ml to 20,000 cells per 100 ml.Fecal coliform populations in the sediment fluc-tuated nearly 1,000-fold over the study period.
It should be pointed out that the minimumtemperature for growth of Aeromonas undercontrolled laboratory conditions is reported torange from 5 to 150C, depending on the strain(27), whereas that of total and fecal coliforms isgenerally lower, ranging from 5 to 100C (6, 24).Thus, it is not surprising that counts were lowerin the natural environment when water temper-atures below 200C were recorded.The relationship between water temperature
and incidence of Aeromonas and other indica-tors observed in this study is similar to thatreported by Fliermans et al. (11) and consistentwith the results of a recent study reported byRippey and Cabelli (S. R. Rippey and V. J.Cabelli, Abstr. Annu. Meet. Am. Soc. Microbiol.1979, p. 179), who found that the concentrationsof A. hydrophila were not correlated with thetotal number of heterotrophic bacteria. How-ever, Rippey and Cabelli suggested that ambientwater temperatures below 150C precluded de-
termination ofA. hydrophila numbers, a conclu-sion not consistent with the results of the studyreported here (i.e., Aeromonas counts were low,but still detectable by our enumeration tech-nique).
In another recent study involving a nation-wide survey, the number ofAeromonas cells was
not correlated with temperature, pH, turbidity,or salinity, but a correlation with conductivitywas reported (15). In the nationwide survey,
each of the many stations that were sampledpossessed a unique set of physical, chemical, andbiological parameters, as well as demonstratingseasonal diversity. Thus, the microbial commun-ities at each station could not be identical.Therefore, it is not surprising that few correla-tions were detected for Aeromonas and themeasured parameters when only a single samplefrom each station was collected for analysis.
Results from the study reported here, in whichcultures were collected by swabbing divers be-fore and after they swam in the Anacostia River,illustrated dramatically a quantifiable effect ofexposure to polluted water when the skin micro-flora of the swimmers was analyzed. A 30-minexposure to river water containing as few as 50to 300 Aeromonas cells per ml, 200 total coli-
TABLE 4. Rabbit ileal loop fluid accumulation andY-l tissue culture response to A. sobria and A.
hydrophilaRabbit ileal
Strain loop fluid ac- Y-1 re-cumulation sponse'(nl/cm)
A. sobriaA3 1.10 C3A36 1.10 CI2b 1.00 CG9 0.90 CIla 0.35 ED8 ob 0G22 ob 0K6f 0 0HA Hosp.c 0.90 C
A. hydrophilaBAll 0.93 CE10 0.74 ED9 0.67 CG29 0.55 E9 0.33 02A2 0.29 0F12 0.20 0HADScc 0 Ea Responses in the Y-1 culture assay: C, cytotoxin;
E, enterotoxin; 0, no response.b A small (<0.1 ml/cm) amount of blood accumu-
lated in these loops.'Isolate obtained from a leg wound sustained by a
diver at the Anacostia River sampling station (datapreviously reported [17]).
1015VOL. 39, 1980
on July 31, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

1016 SEIDLER ET AL.
forms per ml, and 5 fecal coliforms per ml alteredthe skin microfloral composition sufficiently toreflect the kinds of bacteria present in the water.The number of Aeromonas cells and coliformsrecovered from ear swabs suspended in 1 ml ofsaline immediately after sampling and held for1 to 3 h before culture averaged 130 cells per ml.It is not known how many Aeromonas cells arerequired to establish a wound infection, but in-timate contact of the organism with an abrasionor wound could lead to an infection. A shorttime of exposure to river water may facilitatethe first step in pathogenesis, i.e., attachment ofthe organisms to the skin surface. It is interestingto note that one of the strains, which was ex-amined by electron microscopy, was found to bepiliated. It is known that pili promote adherence(5, 18). Hence, it can be speculated that adhesionand/or colonization of the skin by Aeromonasmay be significant under the circumstances re-
ported here.Popoff and Veron (25) concluded that A. sob-
ria is a separate and distinct species within thegenus Aeromonas and can be differentiated fromA. hydrophila, based on a set of biochemicalreactions, including esculin hydrolysis, fermen-tation of salicin, growth in KCN broth, andoverall deoxyribonucleic acid base composition.The majority of the Aeromonas strains isolatedin this study were identified as A. hydrophila.Seasonal influence on the incidence of the twospecies was similar; that is, when the total num-ber of Aeromonas cells rose or fell, the numberof both species fluctuated in unison.
Species identification for Aeromonas isolatedin this study proved useful in the interpretationof the antibiograms. With the exception of thegeneral resistance of both Aeromonas species toampicillin (183 of 190 strains resistant), only 1 ofthe 21 A. sobria strains was resistant to ampi-cillin, tetracycline, and kanamycin. A total of 16strains of A. hydrophila were resistant to tetra-cycline. The strain of A. hydrophila isolatedfrom the wound sustained by the diver was alsoresistant to tetracycline (17). Of the 16 tetracy-cline-resistant A. hydrophila strains, 4 weremultiply antibiotic resistant (3 resistant to tet-racycline, erythromycin, and ampicillin; 1 resist-ant to tetracycline, nalidixic acid, and erythro-mycin).
Strains ofA. sobria do not produce an elastase(25). Elastase production by the strains of A.hydrophila isolated in this study was variable;a positive response was observed in about one-
third of the strains. However, a good correlationwas noted between Y-1 tissue culture response
and elastase production. Indeed, in ca. 80% ofthe cases positive elastase production was asso-
ciated with positive cytotoxin production and
negative elastase reaction was associated with anegative tissue culture response. Unfortunately,elastase activity was lost after storage of isolatesfor 5 to 7 weeks at room temperature under oil.Therefore, the assay for elastase should be doneimmediately after freshly isolated strains havebeen purified and are ready for testing. Cyto-toxin activity proved to be more stable and wasnot readily lost, a result in agreement with datapublished by other workers (7). Based on theresults obtained in this study, it is concludedthat elastase production cannot be consideredcritical for eliciting cytotoxin activity in the Y-1cell assay system.
Discrepancies have been noted in the inci-dence of cytotoxin and/or enterotoxin produc-tion by Aeromonas strains isolated from clinicalspecimens. In fact, the clinical significance of thespecies A. hydrophila and A. sobria has beendisputed. Ljungh et al. (21) examined the etio-logical agents of diarrhea in 354 Ethiopian chil-dren: 11 of the A. hydrophila isolates recoveredfrom stools were determined to be enterotoxi-genic by the rabbit loop, rabbit skin, and Y-1adrenal cell assays. An additional 16 strains ofAeromonas originally isolated in India werefound to be nontoxigenic. However, no isolatesof A. sobria were obtained from stool specimensin the two surveys. Biochemical analyses re-vealed that A. hydrophila produces two cyto-lytic toxins (hemolysins) and a heat-labile enter-otoxin (22). Other workers have been more re-luctant to conclude that Aeromonas spp. areenterotoxigenic, since culture filtrates are notsteroidogenic and do not stimulate adenylatecyclase production (7, 9). Based on the ability ofsome of the strains isolated in this study, weconclude that Aeromonas can produce an enter-otoxin which induces fluid accumulation in rab-bits. Although the molecular mechanism causingfluid accumulation is not clear, our findings andthose of other workers (7) substantiate the en-terotoxic-like nature of the Aeromonas cultures.The fact thatAeromonas enterotoxin may differfrom E. coli and V. cholerae enterotoxins isconsistent with the immunological distinctive-ness of the Aeromonas enterotoxin (4, 9). Thestrongest and most relevant piece of evidence insupport of the contention that Aeromonas doesindeed produce an enterotoxin is the docu-mented association of Aeromonas with cases ofhuman diarrhea (2, 7, 13, 21, 29). However, thisdoes not exclude the possibility that Aeromonasmay be invasive, although this has not beenproven.An examination of the incidence and amount
of fluid accumulation in the rabbit loop testleads us to conclude that strains of both A.hydrophila and A. sobria are enterotoxigenic. A
APPL. ENVIRON. MICROBIOL.
on July 31, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEROMONAS IN POLLUTED WATERS 1017
good correlation was observed between amountof fluid accumulation and detectable response inthe tissue culture assay. It should be noted that6 of 10 strains inducing fluid accumulation in therabbit loop test were cytotoxic, but did not ex-hibit enterotoxin-like activity for Y-1 adrenalcells.The potential public health significance of
large numbers ofAeromonas in aquatic environ-ments cannot be ignored. Several reports havedemonstrated wound infections caused by Aero-monas, and such infections have been linkedcircumstantially with recreational and other ac-tivities in aquatic environments known to con-tain Aeromonas (12, 14, 17, 26). The results ofthis study underscore the significance of thispotential pathogen in polluted aquatic environ-ments in which the water temperature may riseto more than 150C during certain times of theyear (11, 12, 17). The numbers of Aeromonascells show a significant correlation with fecalcoliform counts for both sediment and water andan inverse correlation with dissolved oxygen.Indeed, we conclude that Aeromonas is a usefulindicator of pollution and of public health safetyoffreshwater environments (Rippey and Cabelli,Abstr. Annu. Meet. Am. Soc. Microbiol. 1979).The occurrence ofAeromonas strains producingcytolytic hemolysins, proteases, enterotoxins,and other virulence factors (3, 7, 9, 22) is muchgreater than the occurrence of fecal coliforms(9). Thus, Aeromonas spp. pose a highly signif-icant potential public health problem.
ACKNOWLEDGMENTSWe thank B. R. Merrell, J. Gilmore, J. Perry, C. Adkins, R.
Gates, J. W. Deming, and David Maneval for expert technicalassistance. We especially thank W. S. Hunt for his assistanceand advice.
This work was performed under National Oceanic andAtmospheric Administration grant 04-8-MO1-71, NationalOceanic and Atmospheric Administration contract 01-8-M01-2027, and Naval Medical Research and Development Com-mand Work Unit ZF51524009.0057. Support for microbialidentifications and classification was provided, in part, byNational Science Foundation grant DEB77-14646. Computertime was made available by the University of Maryland Com-puter Science Center.
LITERATURE CIMD1. American Public Health Association. 1975. Standard
methods for the examination of water and wastewater,14th ed. American Public Health Association, NewYork.
2. Annapurna, E., and S. C. Sanyal. 1977. Enterotoxicityof Aeromonas hydrophila. J. Med. Microbiol. 10:317-323.
3. Bernheimer, A. W., and L. S. Avigad. 1974. Partialcharacterization of aerolysin, a lytic exotoxin from Aero-monas hydrophila. Infect. Immun. 9:1016-1021.
4. Boulanger, Y., R. Lallier, and G. Consineau. 1977.Isolation of enterotoxigenic Aeromonas from fish. Can.J. Microbiol. 23:1161-1164.
5. Brinton, C. C., Jr. 1977. The piliation phase syndrome
and the uses of purified pili in disease control, p. 34-70.In Proceedings of the 13th Joint Conference on Cholera.U.S. Dept. of Health, Education and Welfare, Washing-ton, D.C..
6. Buchanan, R. E., and N. E. Gibbons (ed.). 1974. Ber-gey's manual of determinative bacteriology, 8th ed. TheWilliams & Wilkins Co., Baltimore.
7. Cumberbatch, N., M. J. Gurwith, C. Langston, R. B.Sack, and J. L. Brunton. 1979. Cytotoxic enterotoxinproduced by Aeromonas hydrophila: relationship oftoxigenic isolates to diarrheal disease. Infect. Immun.23:829-837.
8. Davis, W. A., II, J. G. Kane, and V. F. Garagusi. 1978.Human Aeromonas infections. A review of the litera-ture and a case report of endocarditis. Medicine 57:267-277.
9. Donta, S. T., and A. D. Haddow. 1978. Cytotoxic activityof Aeromonas hydrophila. Infect. Immun. 21:989-993.
10. Feaster, F. T., R. M. Nisbet, and J. C. Barber. 1978.Aeromonas hydrophila corneal ulcer. Am. J. Ophthal-mol. 85:114-117.
11. Fliermans, C. B., R. W. Gorden, T. C. Hazen, and G.W. Each. 1977. Aeromonas distribution and survival ina thermally altered lake. Appl. Environ. Microbiol. 33:112-114.
12. Fulghum, D. D., W. R. Linton, Jr., and D. Taplin.1978. Fatal Aeromonas hydrophila infection of the skin.South. Med. J. 71:739-741.
13. Gurwith, M., C. Bourque, E. Cameron, G. Forrest,and M. Green. 1977. Cholera-like diarrhea in Canada.Arch. Intern. Med. 137:1461-1464.
14. Hanson, P. G., J. Standridge, F. Jarett, and D. G.Maki. 1977. Freshwater wound infection due to Aero-monas hydrophila. J. Am. Med. Assoc. 238:1053-1054.
15. Hazen, T. C., C. B. Fliermanns, R. F. Hirsch, and G.W. Esch. 1978. Prevalence and distribution of Aero-monas hydrophila in the United States. Appl. Environ.Microbiol. 36:731-738.
16. Hobbie, J. E., R. J. Daley, and S. Jasper. 1977. Use ofNucleopore filters for counting bacteria by fluorescencemicroscopy. Appl. Environ. Microbiol. 33:1225-1228.
17. Joseph, S. W., 0. P. Daily, W. S. Hunt, R. J. Seidler,D. A. Allen, and R. R. Colwell. 1979. Aeromonasprimary wound infection of a diver in polluted waters.J. Clin. Microbiol. 10:46-49.
18. Joseph, S. W., B. R. Merrell, M. Sochard, and W. P.Brown. 1978. Visualization of the attachment of Vibrioparahaemolyticus in the rabbit ileum by electron mi-croscopy. Scanning Electron Microsc. 2:727-732.
19. Kaper, J. B., H. Lockman, R. R. Colwell, and S. W.Joseph. 1979. Ecology, serology, and enterotoxin pro-duction of Vibrio cholerae in Chesapeake Bay. Appl.Environ. Microbiol. 37:91-103.
20. Kaper, J. B., G. S. Sayler, M. M. Baldini, and R. R.Colwell. 1977. Ambient-temperature primary nonselec-tive enrichment for isolation of Salmonella spp. froman estuarine environment. Appl. Environ. Microbiol.33:829-835.
21. Ljungh, A., M. Popoff, and T. Wadstrom. 1977. Aero-monas hydrophila in acute diarrheal disease: detectionof enterotoxin and biotyping of strains. J. Clin. Micro-biol. 6:96-100.
22. Ljungh, A., B. Wietlind, and T. Wadstrom. 1978.Evidence for enterotoxin and two cytolytic toxins inhuman isolates of Aeromonas hydrophila, p. 947-960.In P. Rosenberg (ed.), Proceedings of the 5th Interna-tional Symposium on Toxins: Animal, Plant and Micro-bial. Pergamon Press, New York.
23. McCoy, R. H., and K. S. Pilcher. 1974. Peptone beefextract glycogen agar, a selective and differential Aero-monas medium. J. Fish. Res. Board Can. 31:1553-1555.
24. Naemura, L G., and R. J. Seidler. 1978. Significance oflow-temperature growth associated with the fecal coli-form response, indole production, and pectin liquefac-
VOL. 39, 1980
on July 31, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

1018 SEIDLER ET AL.
tion in Klebsiella. Appl. Environ. Microbiol. 35:392-396.
25. Popoff, M., and M. Veron. 1976. A taxonomic study ofthe Aeromonas hydrophila-Aeromonas punctatagroup. J. Gen. Microbiol. 94:11-22.
26. Rosenthal, S. G., H. E. Bernhardt, and J. A. Phillips.1974. Aeromonas hydrophila wound infection. Plast.Reconstr. Surg. 53:77-79.
27. Rouf, M. A., and M. M. Rigney. 1971. Growth temper-atures and temperature characteristics of Aeromonas.Appl. Microbiol. 22:503-506.
28. Sack, D. A., and R. B. Sack. 1975. Test for enterotoxi-genic Escherichia coli using Y1 adrenal cells in mini-culture. Infect. Immun. 11:334-336.
29. Sanyal, S. C., S. J. Singh, and P. C. Sen. 1975. Enter-
APPL. ENVIRON. MICROBIOL.
opathogenicity of Aeromonas hydrophila and Plesio-monas shigelloides. J. Med. Microbiol. 8:195-198.
30. Schwartz, J. R., and R. R. Colwell. 1974. Effect ofhydrostatic pressure on growth and viability of Vibrioparahaemolyticus. Appl. Microbiol. 28:977-981.
31. Shotts, E. B., Jr., and R. Rimler. 1973. Medium for theisolation of Aeromonas hydrophila. Appl. Microbiol.26:550-553.
32. Smith, H. L., Jr., and K. Goodner. 1958. Detection ofbacterial gelatinases by gelatin agar plate methods. J.Bacteriol. 76:662-665.
33. Tenney, J. H., T. F. Smith, and J. A. Washington II.1979. Sensitivity, precision, and accuracy of the Y1adrenal cell enterotoxin assay. J. Clin. Microbiol. 9:197-199.
on July 31, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















