
Isolation and characterisation of dog uropathogenic Escherichia coli strains and their fimbriae
15
Antonie van Leeuwenhoek 54:149--163 (1988) 9 Kluwer Academic Publishers, Dordrecht - Printed in the Netherlands Isolation and characterisation of dog uropathogenic Escherichia coli strains and their fimbriae E. GARCIA I, H.E.N. BERGMANS 2, J.F. VAN DEN BOSCH 3, I. ORSKOV 4, B.A.M. VAN DER ZEIJST 1 & W. GAASTRA 1 1 Institute of Infectious Diseases and Immunology, Dept. of Bacteriology, Faculty of Veterinary Medicine, University of Utrecht, Yalelaan 1, P.O. Box 80171, 3508 TD Utrecht, The Netherlands; 2Dept. of Molecular Cell Biology, Transitorium 3, University of Utrecht, Padualaan 8, 3584 CH Utrecht, The Netherlands; Slntervet International B.V., Wire de Korverstraat 5, P.O. Box 31, 5830 AA Boxmeer, The Netherlands; 4International Escherichia coli Centre (WHO), Statens Seruminstitut, Copenhagen, Denmark Received 2 December 1987; accepted 3 December 1987 Key words: Escherichia coli, urinary tract infection, dogs, fimbriae Abstract. A number of Escherichia coli strains have been isolated from dogs with urinary tract infections, These strains have been characterised with respect to their O, K, H, and fimbrial antigens, colicin production, antibiotic resistance, plasmid content and their ability to hae- magglutinate erythrocytes from various species. Crossed immunoelectrophoresis of fimbrial extracts, as well as the reaction of partly purified fimbriae of a number of these strains with monoclonal antibodies revealed homology or a strong crossreaction with an F12 fimbrial subunit protein of human uropathogenic E. coli strains. Unlike human F12 fimbriae producing strains, the dog isolates did agglutinate dog erythrocytes in the presence of D-mannose but not human erythrocytes, indicating that the adhesin carried by these strains is different from the adhesin on fimbriae of human uropathogenic E. coli. Similar indications were obtained from experiments with latex beads coated with the receptor for P-fimbriae. These beads were agglutinated by Escherichia coli strains from human urinary tract infections, but not by the dog isolates described here. Preliminary adhesion experiments of human and dog Escherichia coli to human bladder epithelial and canine kidney epithelial cells also showed differences in adhesion depending on the origin of the strain tested. Introduction As in man, urinary tract infections in dogs are commonly caused by either one of two pathogenic organisms, i.e. Escherichia coli and Proteus mirabilis (Kivis- t6 et al. 1978). Whereas E. coli is the organism most commonly found in human urinary tract infections (Baselier 1983), E. coli and P. mirabilis are found with almost equal frequency in urine samples of dogs with urinary tract infections sent to the Microbial Diagnostics Centre of the Institute of In- fectious Diseases and Immunology of the University of Utrecht. Whether
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Isolation and characterisation of dog uropathogenic Escherichia coli strains and their fimbriae

Antonie van Leeuwenhoek 54:149--163 (1988) �9 Kluwer Academic Publishers, Dordrecht - Printed in the Netherlands
Isolation and characterisation of dog uropathogenic Escherichia coli strains and their fimbriae
E. GARCIA I, H.E.N. BERGMANS 2, J.F. VAN DEN BOSCH 3, I. O R S K O V 4, B.A.M. VAN DER ZEIJST 1 & W. GAASTRA 1 1 Institute of Infectious Diseases and Immunology, Dept. of Bacteriology, Faculty of Veterinary Medicine, University of Utrecht, Yalelaan 1, P.O. Box 80171, 3508 TD Utrecht, The Netherlands; 2 Dept. of Molecular Cell Biology, Transitorium 3, University of Utrecht, Padualaan 8, 3584 CH Utrecht, The Netherlands; S lntervet International B.V., Wire de Korverstraat 5, P.O. Box 31, 5830 AA Boxmeer, The Netherlands; 4International Escherichia coli Centre (WHO), Statens Seruminstitut, Copenhagen, Denmark
Received 2 December 1987; accepted 3 December 1987
Key words: Escherichia coli, urinary tract infection, dogs, fimbriae
Abstract. A number of Escherichia coli strains have been isolated from dogs with urinary tract infections, These strains have been characterised with respect to their O, K, H, and fimbrial antigens, colicin production, antibiotic resistance, plasmid content and their ability to hae- magglutinate erythrocytes from various species. Crossed immunoelectrophoresis of fimbrial extracts, as well as the reaction of partly purified fimbriae of a number of these strains with monoclonal antibodies revealed homology or a strong crossreaction with an F12 fimbrial subunit protein of human uropathogenic E. coli strains. Unlike human F12 fimbriae producing strains, the dog isolates did agglutinate dog erythrocytes in the presence of D-mannose but not human erythrocytes, indicating that the adhesin carried by these strains is different from the adhesin on fimbriae of human uropathogenic E. coli. Similar indications were obtained from experiments with latex beads coated with the receptor for P-fimbriae. These beads were agglutinated by Escherichia coli strains from human urinary tract infections, but not by the dog isolates described here. Preliminary adhesion experiments of human and dog Escherichia coli to human bladder epithelial and canine kidney epithelial cells also showed differences in adhesion depending on the origin of the strain tested.
Introduction
As in man, urinary tract infections in dogs are commonly caused by either one of two pathogenic organisms, i.e. Escherichia coli and Proteus mirabilis (Kivis- t6 et al. 1978). Whereas E. coli is the organism most commonly found in human urinary tract infections (Baselier 1983), E. coli and P. mirabilis are found with almost equal frequency in urine samples of dogs with urinary tract infections sent to the Microbial Diagnostics Centre of the Institute of In- fectious Diseases and Immunology of the University of Utrecht. Whether

150 E. Garcia et al.
these samples are representative for the frequencies observed in the general veterinary practices is not known. Also like in man, the majority of cases is observed in the female of the species. Human urinary tract infections have been studied extensively during the last decade. During these studies a number of virulence factors have been established in uropathogenic E. coli strains. Of the more than 160 O antigens described for E. coli, only a limited number is found on the majority of strains isolated from human urinary tract infections, i.e. 01, 02, 04, 06, 07,018, and 075 (r & Drskov 1985). Also of the more than 100 different K antigens described for E. coli only K1, K2, K3, K12 and K13 have been found on strains associated with human urinary tract infection (Kaijser et al. 1977, van den Bosch et al. 1982). No reports have been made on whether or not some of the approximately 50 known flagellar antigens of E. coli preferentially occur on strains associated with urinary tract infections. Other possible virulence factors of E. coli causing urinary tract infections are their ability to produce haemolysin (Minshew et al. 1978; van den Bosch et al. 1981, 1982; Hughes et al. 1983), the production of colicins and the production of siderophores like aerobactin (Carbonetti & Williams 1985). Adherence to uroepithelial cells is also an important factor in urinary tract infections (Svan- borg-Ed6n et al. 1976). This adherence is mediated by fimbriae, long thin, threadlike proteinaceous structures on the bacterial surface. Uropathogenic strains are almost invariably fimbriated (Svanborg-Ed6n et al. 1982; Korhonen et al. 1982; Drskov & Drskov 1983; Hagberg et al. 1981; Svanborg-Ed6n & Hansson 1978).
The majority of E. coli strains causing urinary tract infections are able to produce more than one type of fimbrial adhesion antigen (Klemm et al. 1982; Salit et al. 1983; Rhen et al. 1983). These fimbrial types can be divided in those that cause mannose-sensitive and those that cause mannose-resistant hae- magglutination. Mannose-sensitive fimbriae are referred to as type 1 fimbriae. Type 1 fimbriae are found on more than 85% of all E. coli strains and their role in urinary tract infections is still disputed (Hagberg et al. 1981; V/iis/inen-Rhen et al. 1984). A definite role in the adhesion to human uroepithelial cells has been assigned to the mannose-resistant fimbriae, especially to the socalled P fimbriae, binding to the P blood group antigen on human cells (Khllenius et al. 1980). The actual binding however, is not mediated by the fimbrial subunits, but by one of the minor fimbrial components, the adhesin (Normark et al. 1985; Van Die et al. 1986). Besides the P fimbriae, other mannose-resistant adhesion antigens, of which the role in the pathogenesis of urinary tract infections still is unclear are found on human uropathogenic E. coli. These adhesion antigens include the M. fimbriae, binding to the blood group M associated glycoprotein glycophorin A (V/iis/~nen et al. 1982), and the S fimbriae that bind to sialic acid (Korhonen et al 1984).
In addition there are X fimbriae for which the receptor substance is not yet

Dog uropathogenic Escherichia coli 151
known. Furthermore an adhesive protein capsule consisting of a fibrillar structure has been described (Orskov et al. 1985). A number of E. coli strains from the urine of dogs with urinary tract infections has been isolated and analysed for the presence of the virulence factors mentioned above. Special attention was paid to the production of fimbriae, which was demonstrated by electron microscopy, haemagglutination and purification of fimbriae. The presence of genes encoding fimbriae was demonstrated by Southern hybrid- isation. It was demonstrated that the fimbrial subunit proteins on some human and dog uropathogenic E. coli are homologous or at least strongly related, but that the actual adhesin possibly is different. It is speculated that the difference in the adhesin molecule might account for species specificity.
Materials and methods
Bacterial strains and growth media
The E. coli strains were isolated at the Microbiological Diagnostics Centre of the Institute of Infectious Diseases and Immunology. The strains were cultur- ed from the urine of infected dogs on selective media (Brilliant green agar, Oxoid). After streaking to single colonies, the cells were grown overnight in L Broth and stored at - 7 0 ~ C untill further use as a suspension in a 1 : 1 mixture of L- Broth and glycerol. The strains were grown on isosensitest agar (Oxoid) to test their sensitivity to various antibiotics. For the preparation of chromoso- mal and plasmid DNA and for the isolation of fimbriae the strains were grown in L- Broth. Bacteria used for the mannose-resistant haemagglutination test, were grown on bloodagar plates prepared with fresh horse blood. For the preparation of fimbrial extracts for F antigen analysis, the bacteria were grown on the solid medium described previously (Orskov et al. 1985).
Haemagglutination
Haemagglutination was performed with dog, human, rabbit, guinea pig, sheep, pig, horse and bovine erythrocytes. The red blood cells were washed in phosphate buffered saline (PBS) untill no haemoglobin was present in the supernatant. Agglutination was performed on glass slides on ice and read macroscopically. Bacteria from overnight plate cultures were suspended in 2ml PBS with a Drigalsky spatula, centrifuged, washed in PBS to remove components of the growth medium and resuspended in PBS with and without mannose (0.5% final concentration) at a concentration of 109 cells/ml. To 100 ul bacterial suspension, 10 ul of a 10% erythrocyte suspension was added.

152 E. Garcia et al.
Agglutination with Latex beads coated with P-fimbriae receptor
The Bach-test (KabiVitrum, Stockholm, Sweden), a Latex test kit for the identification of P-fimbriated Escherichia coli was kindly donated by Dr. C. Svanborg-Ed6n. The agglutination tests were performed according to the manufacturers description.
Serotyping
Serotyping of the strains was performed at the International E. coli Centre, Statens Serum Institut, Copenhagen, Denmark, (Orskov & Orskov 1984).
F typing offimbriae
Serological analysis of fimbriae containing bacterial extracts by crossed immu- noelectrophoresis (CIE) was performed against F71, F7~, F8-F16 and F1B, F1C antisera as described before (~rskov & ~rskov 1983). The presence of certain F types of fimbriae was also tested in a whole-bacteria ELISA, using monoclonal antibodies specific for F71, F72, F8, F9, Fll , F12, F12rel or F13 fimbriae (de Ree et al. 1986; de Ree 1986).
Haemolysin production
Haemolysin production was tested on washed - blood agar plates as described by Van den Bosch et al. (1981).
Isolation and purification of fimbriae
The cells from an 800 ml overnight culture were pelleted by centrifugation and resuspended in 20 ml PBS containing 4M urea. This suspension was heated for 30 rain at 60 ~ C and allowed to cool to room temperature. The cells were then removed by centrifugation and the supernatant containing the fimbriae was centrifuged for 18 h at 40,000 rpm in a Ti 50 rotor. The pellet of this centrifu- gation step was resuspended in 5 ml PBS, containing 0.4% sodium dodecyl sulfate (SDS) and heated for 30 min at 40 ~ C. The fimbriae were pelleted again from this solution by centrifugation at 40,000 rpm. The pellet was redissolved in one rnl PBS. The fimbriae were precipitated by the addition of polyethylene glycol (PEG) and sodium chloride to a final concentration of 4.0% (W/V) and

Dog uropathogenic Escherichia coli 153
0.5% (W/V), respectively. The latter precipitation was repeated if necessary, Finally the fimbriae were dissolved in 10mM Tris-HC1 buffer (pH = 8.0) containing 1 mM EDTA.
Sodium dodecyl sulfate polyacrylamide gelelectrophoresis (SDS-PA GE)
The purity of the fimbrial preparations was determined by SDS - PAGE at the various stages of the isolation procedure. SDS- PAGE was performed on 15% acrylamide gels as described by Laemmli (1970).
Isolation of chromosomal DNA and digestion with restriction enzymes
Chromosomal DNA was isolated from 40 ml overnight cultures by resuspend- ing the bacteria in 10 m120 mM Tris-HCl buffer (pH = 8.0), containing 50 mM glucose. The cells were incubated with lysozyme (2 mg/ml) for 10 min on ice and lysed by the addition of SDS to a final concentration of 0.5% (W/V). The DNA solution was then incubated with proteinase K (1 mg), at 65 ~ C for 18 h. The DNA was precipitated by the addition of an equal volume ice cold isopropanol. After gentle mixing of the two phases the DNA clump was removed from the solution with the bend tip of a sterile pasteur pipette and redissolved in 1 ml Tris-EDTA (TE) buffer (10 mM Tris, 0.1 mM EDTA, pH 8.0). The DNA was used as such if it could be digested with restriction enzymes. If this was not the case the proteinase K treatment was repeated and the solution was extracted with phenol. Incubations with restriction enzymes were performed according to the instructions of the manufacturer.
Plasmid isolation
Plasmid isolations were performed as described by Birnboim & Doly 1979.
Preparation of radioactive probe
The Hind III fragment of pPIL 110-76 (Van Die et al. 1985) and the EcoR I - Sph I fragment of pANN 801-11 (Hacker et al. 1985) containing parts of the fimbrial gene clusters of F71 and S fimbriae respectively were isolated from agarose gels by electroelution. One ug of DNA was labeled with 35S-ATP by nick translation according to Maniatis et al. 1982. Plasmid pANN 801-11 was kindly donated by Dr. J. Hacker.

154 E. Garcia et al.
Southern hybridisations
Chromosomal DNA was digested with various restriction endonucleases and fractionated by agarose gel electrophoresis. The DNA was transfered to Gene Screen filters (DuPont), as described by Southern (1975). Hybridisation with radioactively labelled probe DNA was performed in hybridisation solution containing 50% formamide, as described by Maniatis et al. 1982 during 16 h at 42 ~ C. The filters were washed twice in 5X SSC at 50~ for 30min and mounted for autoradiography.
Colicin production
The production of colicins by the bacterial strains was tested as described by Oudega et al. 1977.
Electron microscopy
Purified fimbriae preparations were negatively stained with a 1% solution of uranyl acetate for 2 min and viewed in a Philips 201 electron microscope.
Results
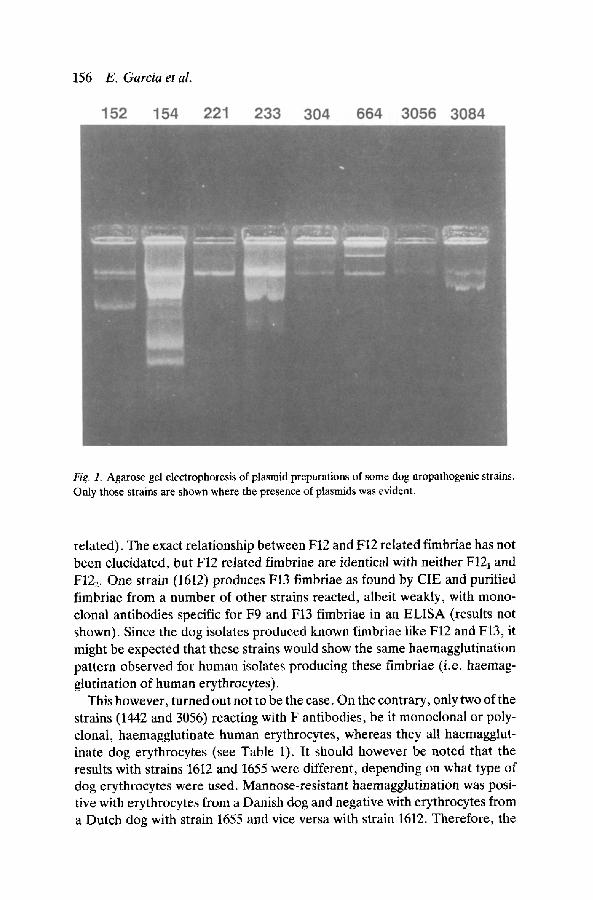
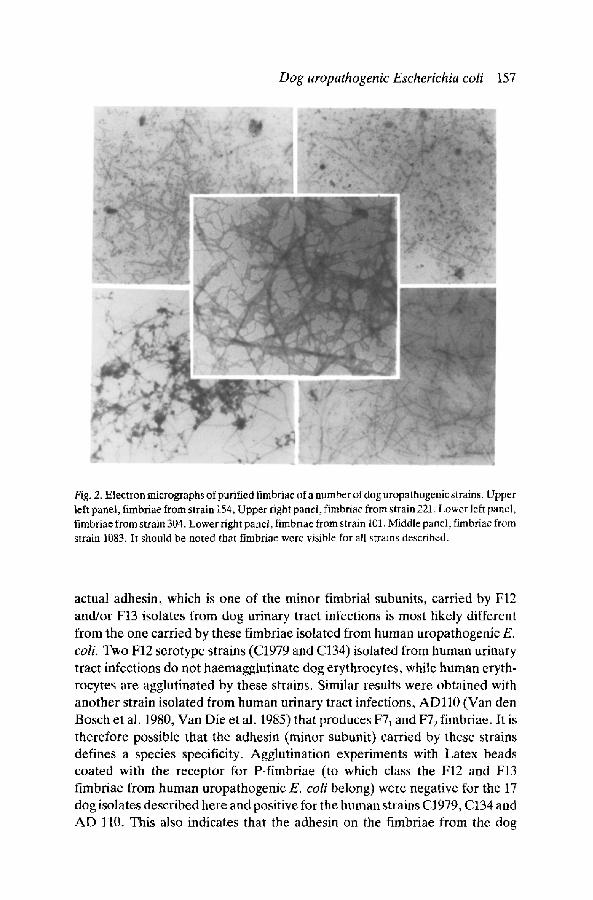
From November 1985 untill January 1987, 44 E. coli strains were isolated from urine samples, sent for diagnosis, by veterinarians from all over The Nether- lands. Here the characterisation of the first 17 E. coli strains isolated is reported. In Table 1, these strains have been listed together with a number of their characteristics, like serotype, mannose-resistant haemagglutination and haemolysin production. Of the seventeen strains described here none was sensitive to all of ten antibiotics tested. There are no antibiotics to which resistance was observed more frequently than to others. This is in agreement with the fact that the plasmid content varies from strain to strain, both with regard to the number and the size of the plasmids present (Fig. 1). The presence of fimbriae on the strains is also indicated in Table 1. The typable strains belonged to 8 different O groups. Nine strains belonged to O serotypes commonly associated with human urinary tract infections, namely 02, 04, 06 and 075. Twelve of the strains were haemolytic. Comparable results have been described by Westerlund et al. 1987. The production of fimbriae by the strains described here was demonstrated by various methods, directly by electron microscopy of purified fimbriae (Fig. 2) and indirectly by their ability to

Dog uropathogenic Escherichia coli 155
haemagglutinate red blood cells. The fimbriae were further characterised in two ways, in CIE, using fimbriae containing bacterial extracts and antisera raised in rabbits against reference strains (Orskov & Orskov 1983) and in ELISA using intact bacterial cells and monoclonal antibodies against fimbriae isolated from E. coli from human urinary tract infections (de Ree et al. 1985, 1986). Seven of the strains reacted with monoclonals against F12 fimbriae and four of these strains were also F122 positive in the crossed immunoelectropho- resis. On one strain the presence of antigen F121 was shown in CIE, but it did not react with monoclonal F12 antibodies. Klemm et al. (1983) and Orskov & Orskov (1983) have suggested that the F12 test strain C1979 produces two highly homologous fimbrial proteins, which recently have been defined F121 and F122 (Orskov & Orskov unpubl, results).
Likewise de Ree (1986) observed different reactions with monoclonal anti- bodies raised against fimbriae of strain C1979 (F12) and strain C134 (F12
Table 1. Characterisation of dog uropathogenic Escherichia coli strains.
Strain no. Serotype M R H A F typing Mabs Haemolysin
152 O?:H45 Hu, Do - - -
154 O8:K?:H12 . . . .
221 rough:H7 Do F1B/F1C F12 ( + + ) + + +
233 09:K32:H4 . . . .
304 075:K5/35:H5 Do - F12 ( + + + ) + + +
664 083:H31 Hu, Do - - +
1013 06:K53:H1 Do F122, F1C F12 ( + + + ) + + +
1083 06:K53:H1 Do F122, F1C F12 ( + + + ) + + +
1104 04:H5 Hu, Do - - + + +
1389 06:K13:H1 - F1C - + + +
1442 022:H1 Hu, Do F122, F1C F12 ( + + + ) + + +
1520 06:K13:H1 Do F122 F12 ( + + + ) + + +
1565 O?:H21 . . . .
1612 O2:Kl:H6 Do? F13 - + + +
1655 06:K14:H- Do? F121 - + +
3056 04:H5 Hu, Do F1B/F1C F12 ( + + ) + + +
3084 O9:K34:H . . . . .
Where no K antigen is given we consider the strain to be K -
K? = means the presence of a polysaccharide K antigen not reacting with our antisera.
M R H A = mannose-resistant haemagglutination.
Hu = positive haemagglutination with human erythrocytes.
Do = positive haemagglutination with dog erythrocytes.
Do? = positive haemagglutination with dog erythrocytes, depending on what type of erythrocytes
were used (see also text).
F typing stands for the crossed immuno electrophoresis experiments.
Mabs = reaction with monoclonal antibodies in whole cell ELISA.
156 E. Garcia et al.
Fig. 1. Agarose gel electrophoresis of plasmid preparations of some dog uropathogenic strains. Only those strains are shown where the presence of plasmids was evident.
related). The exact relationship between F12 and F12 related fimbriae has not been elucidated, but F12 related fimbriae are identical with neither F121 and F122. One strain (1612) produces F13 fimbriae as found by CIE and purified fimbriae from a number of other strains reacted, albeit weakly, with mono- clonal antibodies specific for F9 and F13 fimbriae in an ELISA (results not shown). Since the dog isolates produced known fimbriae like F12 and F13, it might be expected that these strains would show the same haemagglutination pattern observed for human isolates producing these fimbriae (i.e. haemag- glutination of human erythrocytes).
This however, turned out not to be the case. On the contrary, only two of the strains (1442 and 3056) reacting with F antibodies, be it monoclonal or poly- clonal, haemagglutinate human erythrocytes, whereas they all haemagglut- inate dog erythrocytes (see Table 1). It should however be noted that the results with strains 1612 and 1655 were different, depending on what type of dog erythrocytes were used. Mannose-resistant haemagglutination was posi- tive with erythrocytes from a Danish dog and negative with erythrocytes from a Dutch dog with strain 1655 and vice versa with strain 1612. Therefore, the
Dog uropathogenic Escherichia coli 157
Fig. 2. Electron micrographs of purified fimbriae of.a number of dog uropathogenic strains. Upper left panel, fimbriae from strain 154, Upper right panel, fimbriae from strain 221. Lower left panel, fimbriae from strain 304. Lower right panel, fimbriae from strain 101. Middle panel, fimbriae from strain 1083. It should be noted that fimbriae were visible for all strains described.
actual adhesin, which is one of the minor fimbrial subunits, carried by F12 and/or F13 isolates from dog urinary tract infections is most likely different from the one carried by these fimbriae isolated from human uropathogenic E. coli. Two F12 serotype strains (C1979 and C134) isolated from human urinary tract infections do not haemagglutinate dog erythrocytes, while human eryth- rocytes are agglutinated by these strains. Similar results were obtained with another strain isolated from human urinary tract infections, ADl l0 (Van den Bosch et al. 1980, Van Die et al. 1985) that produces F7~ and F72 fimbriae. It is therefore possible that the adhesin (minor subunit) carried by these strains defines a species specificity. Agglutination experiments with Latex beads coated with the receptor for P-fimbriae (to which class the F12 and F13 fimbriae from human uropathogenic E. coli belong) were negative for the 17 dog isolates described here and positive for the human strains C1979, C134 and AD 110. This also indicates that the adhesin on the fimbriae from the dog

158 E. Garcia et al.
isolates differs from the one on the fimbriae from human uropathogenic E. coli. The data from preliminary adhesion experiments, where human isolates seem to bind to human bladder uroepithelial cells (T24 cells) but not to dog kidney epithelial cells (MDCK cells) and vice versa also point in this direction.
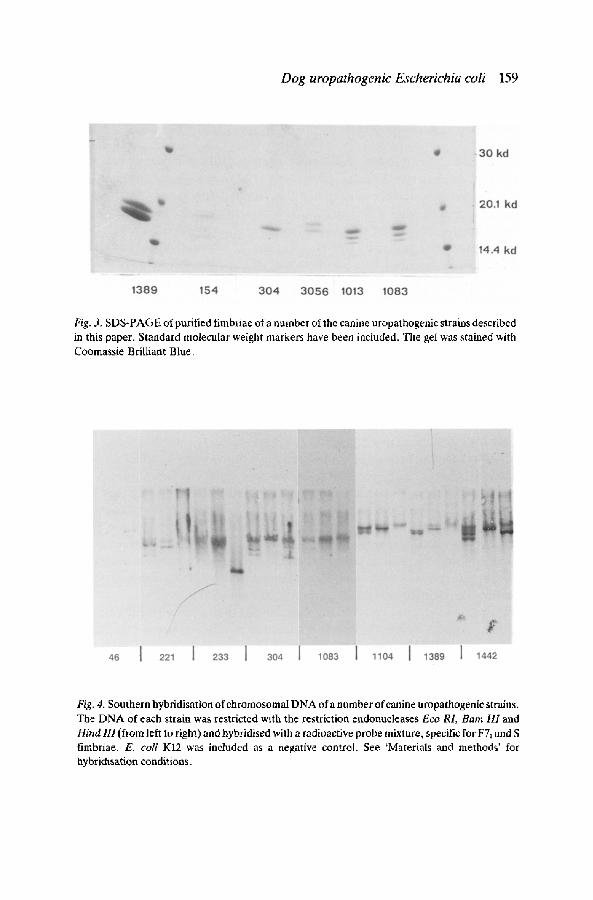
Moreover, it has recently been demonstrated that the human uropathogenic E. coli strain J96 produces both F13 fimbriae carrying an adhesin that specifies haemagglutination of human erythrocytes and F13 fimbriae that carry an adhesin, specific for haemagglutination of sheep erythrocytes (Lund 1987). Strains 221, 1013, 1083, 1104 and 3056 also show mannose-resistant hae- magglutination with sheep erythrocytes and the latter two also haemagglut- inated horse erythrocytes. Rabbit and bovine erythrocytes are haemagglut- inated by strains 152 and 664. The purified fimbriae preparations were analysed by SDS-polyacrylamide gel electrophoresis; from this it became clear that most strains produce more than one type of fimbriae, as judged from the presence of more than one band on the SDS-gels (Fig. 3). As can be seen in Fig. 3, the F12 positive strains have a band with an apparent molecular weight of 18kD, which is the molecular weight of the F12 subunit molecule. The purified fimbriae from strains 304 and 1104, that showed only one band on the SDS-gels and those of strain 3056, that gave two bands were submitted to automatic Edman degradation in a gas phase sequenator.
In all three cases it was clear that a mixture of at least three different proteins had been sequenced, however it could easily be deduced that those proteins were related to FIA and F12 fimbrial subunit proteins (sequences not shown here). FIA fimbrial subunits are not normally seen upon SDS-PAGE analysis of fimbriae, since they do not depolymerize in SDS at neutral pH. Southern hybridisation of chromosomal DNA, digested with the restriction enzymes Eco RI, Barn HI and Hind III with a mixture of probes containing sequences of the structural genes from F71 fimbriae and S fimbriae confirmed the presence of such genes in the strains investigated.
In some of the digests more than one DNA fragment lights up, which also could be an indication for the presence of more than one fimbrial gene in the strains concerned (Fig. 4). However, it was recently demonstrated that the amino acid sequence of S fimbriae and F1C fimbriae are very homologous (SchmoU et al. 1987). Therefore one of the bands that lights up in Fig. 4 might very well represent the F1C structural gene. The production of F1C fimbriae was also demonstrated in the crossed immuno electrophoresis experiments (Table 1). The production of colicins only be demonstrated in three of the strains studied.
Dog uropathogenic Escherichia coli 159
Fig. 3. SDS-PAGE of purified fimbriae of a number of the canine uropathogenic strains described in this paper. Standard molecular weight markers have been included. The gel was stained with Coomassie Brilliant Blue.
Fig. 4. Southern hybridisation of chromosomal DNA of a number of canine uropathogenic strains. The DNA of each strain was restricted with the restriction endonucleases Eco RL Barn HI and Hind 111 (from left to right) and hybridised with a radioactive probe mixture, specific for F7~ and S fimbriae. E. coli K12 was included as a negative control. See 'Materials and methods' for hybridisation conditions.

160 E. Garcia et al.
Discussion
As reported earlier (Orskov et al. 1982) human uropathogenic strains belong to a limited number of O groups, particularly O1, 02, 04, 06, 07, O16, O18 and 075. Investigation of a number of E. coli strains isolated from dog urinary tract infections has shown that some of these O groups are also present among the strains studied. The most frequently found O group was 06 (5 x). Similar results have been reported by Westerlund et al. 1987. The presence of fimbriae on the strains described here was demonstrated in a number of ways, hae- magglutination, electron microscopy, crossed immunoelectrophoresis, reac- tion with monoclonal antibodies and DNA-DNA hybridisation. It is thought that in P fimbriae, isolated from human uropathogenic E. coli, the actual receptor binding activity is not located on the structural fimbrial subunit, but on a socalled minor subunit associated with the fimbrial structure (Normark et al. 1985; Van Die et al. 1986; Lindberg et al. 1987). This protein specifies with what type of erythrocytes haemagglutination occurs in the presence of man- nose.
The fact that the F12 fimbriae produced by dog uropathogenic E. coli in contrast to F12 fimbriae produced by human uropathogenic E. coli do not haemagglutinate human erythrocytes but dog erythrocytes and vice versa, as well as the fact that the dog isolates in contrast to the human isolates do not agglutinate Latex beads coated with P-fimbriae receptor can be interpreted to mean that these strains produce fimbriae composed of a major subunit homo- logous to F12, but carry adhesins associated with thes e fimbriae that recognise different receptors. A similar situation has recently been found for the F13 fimbriae produced by the human uropathogenic strain J96. This strain pro- duces F13 fimbriae with an adhesin that is associated with mannose-resistant haemagglutination of human erythrocytes and F13 fimbriae associated with the haemagglutination of sheep erythrocytes (Lund 1987).
It can however, not be ruled out that for the strains described here, the adhesin associated with the haemagglutination of dog erythrocytes is inde- pendent of the F12 fimbriae, and is the representative of another not yet identified surface structure. Using CIE it was found that the dog uropathogen- ic strains can produce either one of two very homologous F12 fimbriae i.e. F121 and F122. In addition to the fimbriae specific for uropathogenic strains a number of strains also produce F1C fimbriae. Fimbriae with subunits homolo- gous to P-fimbriae are undoutedly an important virulence factor for canine uropathogenic strains, but other adhesion antigens must also be present. Many of the strains described here produce more than one type of fimbriae, a fact that is also observed in human uropathogenic strains.
It has been noted that uropathogenic strains that produce P fimbriae also are haemolytic (V~iisanen-Rhen et al. 1984). In the strains described here, produc-

Dog uropathogenic Escherichia coli 161
tion of fimbriae and strong haemolytic activity also occur simultaneously. Iron is an important growth limiting factor for human uropathogenic E. coli. Haemolysin production is thought to be a virulence factor, since it makes iron available to the bacterial cells. Aerobactin production which is often encoded for by genes located on large Col V plasmids, together with colicin V produc- tion (Krone et al. 1986) is also involved in iron scavaging. Colicin production was however only found in a few strains. The presence of large plasmids was also not frequently observed. Since all the strains tested were resistant to one or more antibiotics it is possible that plasmids observed in the various strains are encoding for antibiotic resistance genes. There is no common R factor in the various strains however.
In conclusion it can be said that canine uropathogenic E. coli strains re- semble human uropathogenic E. coli strains apart from their receptor specific- ity and that a detailed study of the receptor binding protein of the fimbriae on canine uropathogenic Escherichia coli might add to the prevention of urinary tract infections in dogs.
Acknowledgements
We thank Drs. E.G. Hartman and J.L. Cornelisse for collecting the strains described in this study and Dr. J. Hacker for the gift of plasmid pANN 801-11. Dr. C. Svanborg-Ed6n is thanked for kindly providing us with the P-fimbriae receptor test kit.
References
Baselier, P.J.A.M. (1983) Acute bacteriele urineweg-infecties in de huisartspraktijk. PhD. thesis. Katholieke Universiteit Nijmegen, The Netherlands
Birnboim, M.C. & F.G. Doly (1979) A rapid alkaline extraction procedure for screening recombi- nant plasmid DNA. Nucleic Acids Res. 7:1513-1523
Carbonetti, N.H. & P.H. Williams (1984) A cluster of five genes specifying the aerobactin iron uptake system of plasmid Col V-K30. Infect. Immun. 45:7-12
De Ree, J.M., P. Schwillens & J.F. van denBosch (1985) Monoclonal antibodies that recognize the P-fimbriae F71, F72, F9 and F l l from uropathogenic E. coli. Infect. Immun. 50:900-904
De Ree, J.M., P. Schwillens & J.F. van den Bosch (1986) Monoclonal antibodies for serotyping the P-fimbriae of uropathogenic E. coli. J. of Clin. Microbiol. 24:121-125
De Ree, J.M. (1986) Molecular and serological characterization of P-fimbriae from uropathogenic Escherichia coli. PhD. thesis, University of Limburg, Maastricht, The Netherlands
Hacker, J., G. Smidt, C. Hughes, S. Knapp, M. Marget & W. Goebel (1985) Cloning and characterization of genes involved in production of mannose - resistant, neurammidase - susceptible (X) fimbriae from a uropathogenic 06:K15:H31 Escherichia coli strain. Infect. Immun. 47:434--440

162 E. Garcia et al.
Hagberg, I., U. Jodal, T.K. Korhonen, G. Lidin-Janson, U. Lindberg & C. Svanborg-Ed6n (1981) Adhesion, haemagglutination and virulence of E. coli causing urinary tract infection. Infect. Immun. 31:564-570
Hughes, C., J. Hacker, A. Roberts & W. Goebel (1983) Haemolysin production as a virulence marker in urinary tract infections caused by E. coll. Infect. Immun. 39:545-551
Kaijser, B., L.A. Hanson, U. Jodal, G. Lidin-Janson & J.B. Robbins (1977)Frequency orE. coli K antigens in urinary tract infections in children. Lancet i: 663-664
K~illenius, G., R. M611by, S.B. Svenson, J. Winberg, A. Lundblad, S. Svensson & B. Cedergren (1980) The pk antigen as receptor for the haemagglutinin of pyelonephritic Escherichia coli FEMS Microbiol. Letters 7:297-302
Kivist6, A-K, H. Vasenius, L-A. Lindberg & M. Sandholm (1978) Desorption assay. A functional in vitro test for measuring the ahesion of E. coli on the urinary tract epithelium. Invest. Urol. 15: 412-415
Klemm, P., I. Orskov & F. Orskov (1982) F7 and type 1 like fimbriae from three Escherichia coli strains isolated from urinary tract infections; protein chemical and immunological aspects. Infect. Immun. 36:462-468
Korhonen, T.K., V. V/iis/inen, P. Kallio, E-L. Nurmiaho-Lassila, H. Ranta, A. Siitonen, J. Elo, S.B. Svenson & C. Svanborg-Ed6n (1982) The role of pili in the adhesion of Escherichia coli to human urinary tract epithelial cells. Scand. J. Infect. Dis. 33 (suppl.): 26-33
Korhonen, T.K., V. V~iis~nen-Rhen, M. Rhen, A. Porc, J. Parkkinen & J. Finne (1984) Escher- ichia coli fimbriae recognizing sialyl galactosides. J. Bacteriol. 159:762-766
Krone, W.J.A., P. de Vries, G. Koningstein, A.J.R. De Jonge, F.K. De Graaf & B. Oudega (1986) Uptake of cloacin DF13 by susceptible cells: removal of immunity protein and frag- mentation of cloacin molecules. J. Bacteriol. 166:260-268
Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680--685
Lindberg, F., B. Lund, L. Johansson & S. Normark (1987) Localization of the receptor-binding protein adhesin at the tip of the bacterial pilus. Nature 328:84-87
Lund, B. (1987) The molecular mechanism of uroepithelial binding by uropathogenic Escherichia coli; variation of adhesion specificity. PhD. thesis. University of Ume/i. Umea, Sweden
Maniatis, T., E.F. Fritsch & J. Sambrook (1982) In: Molecular Cloning, A Laboratory Manual. Cold Spring Harbor Laboratory
Minshew, B., J. Jorgensen, G.W. Counts & S. Falkow (1978) Association of hemolysin produc- tion, hemagglutination of human erythrocytes and virulence of chicken embryos of extraintesti- nal Escherichia coli isolates. Infect. Immun. 28:359-367
Normark, S., M. Bhga, M. G6ransson, F. Lindberg, B. Lund, M. Norgren & B-E. Uhlin (1985) Genetic analysis of bacterial pilus adhesins. In: T.K. Korhonen, E.A. Dawes & P.H. M~ikela (Eds) Enterobacterial Surface Antigens: Methods for Molecular Characterisation (pp. 39-52). Elsevier Science Publishers, Amsterdam, The Netherlands
Orskov, I., F. Orskov, A. Birch-Andersen, M. Kanamori & C. Svanborg-Ed6n (1982) O, K, H and fimbrial antigens in E. coli serotypes associated with pyelonephritis and cystitis. Scand. J. Infect. Dis. (suppl.) 33:18--25
~lrskov, I. & F. Orskov (1983) The serology of E. coli fimbriae. Progress in Allergy 33:80-105 Orskov, F. & I. Orskov (1984) Serotyping of Escherichia coli. In: T. Bergan (Ed) Methods in
Microbiology. Vol. 14 (pp. 43-112). Academic Press, London Orskov, I. & F. Orskov (1985) Escherichia coli in extra intestinal infections. J. Hyg. 95:551-575 Orskov, I., A. Birch-Andersen, J. Duguid, J. Stenderup & F. Orskov (1985) An adhesive protein
capsule of Escherichia coli. Infect. Immun. 47:191-200 Oudega, B., P. Klaasen-Boor, G. Sneeuwloper & F.K. de Graaf (1977) Interaction of the
complex between cloacin and its immunity protein and of cloacin with the outer and cytoplasmic membrane of sensitive cells. Eur. J. Biochem. 78:445-453

Dog uropathogenic Escherichia coli 163
Rhen, M., P. Klemm, E. Wahistr6m, S.B. Svenson, G. K~illenius & T.K. Korhonen (1983) P-fimbriae of Escherichia coli; immuno- and protein-chemical characterization of fimbriae from two pyelonephretic strains. FEMS Microbiol. Letters 18:233-238
Salit, I.E., J. Vavougios & T. Hofmann (1983) Isolation and characterization of Escherichia coli pili from diverse clinical scources. Infect. Immun. 42:755-762
Schmoll, T., J. Hacker & W. Goebel (1987) Nucleotide sequence of the sfaA gene coding for the S-fimbrial protein subunit of Escherichia coli. FEMS Microbiol. Letters 41:229--235
Southern, E.M. (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98:503-517
Svanborg-Ed6n, C., L.A. Hanson, U. Jodal, U. Lindberg & A. Sahl-Akerlund (1976) Variable adherence to normal human urinary epithelial cells of E. coli strains associated with various forms of urinary tract infections. Lancet i: 490--492
Svanborg-Ed6n, C. & H.A. Hansson (1978) Escherichia coli pili as possible mediators of attach- ment to human urinary tract epithelial cells. Infect. Immun. 21:229-237
Svanborg-Ed6n, C., A. Fasth, L. Hagberg, L.A. Hanson, T. Korhonen & H. Leffler (1982) Host interaction with Escherichia coli in the urinary tract. In: L. Weinstein & B.N. Fields (Eds) Seminars in Infectious Disease. Vol. 4 (pp. 113-131). Bacterial vaccines. Thieme-Stratton, New York
Vais/inen, V., T.K. Korhonen, M. Jokinen, C.G. Gahmberg & C. Ehnholm (1982) Blood group M specific haemagglutinin in pyelonephritic Escherichia coll. Lancet i: 1192
Viiisiinen-Rhen, V., J. Elo, E. V~iisiinen, A. Siitonen, I. Orskov, F. Orskov, S.B. Svenson, P.H. M~ikela & T.K. Korhonen (1984) P-fimbriated clones among uropathogenic E. coli strains. Infect. Immun. 43:149-155
Van den Bosch, J.F., U. Verboom-Sohmer, P. Postma, J. de Graaff & D.M. MacLaren (1980) Mannose sensitive and mannose resistant adherance to human uroepithelial cells and urinary virulence of Escherichia coll. Infect. Immun. 29:226-233
Van den Bosch, J.F., P. Postma, J. de Graaff & D.M. MacLaren (1981) Haemolysis by urinary Escherichia coli and virulence in mice. J. Med. Microbiol. 14:321-331
Van den Bosch, J.F., P. Postma, P.A.R. Koopman, J. de Graaff & D.M. MacLaren (1982) Virulence of urinary and faecal Escherichia coli in relation to serotype, haemolysis and hae- magglutination. J. Hyg. 88:567-577
Van Die, I., G. Spierings, I. van Megen, E. Zuidweg, W. Hoekstra & H. Bergmans (1985) Cloning and genetic organization of the gene cluster encoding F71 fimbriae of a uropathogenic E. coli and comparison with the F72 gene cluster. FEMS Microbiol. Letters 28:329-334
Van Die, I., E. Zuidweg, W. Hoekstra & H.E.N. Bergmans (1986) The role of fimbriae of uropathogenic Escherichia coli as carrier of the adhesin involved in mannose-resistant hae- magglutination. Microbial. Pathogenesis 1:51-56
Westerlund, B., A. Pere, T.K. Korhonen, A-K. J~irvinen, A. Siitonen & P.H. Williams (1987) Characterisation of Escherichia coli associated with canine urinary tract infections. Res. Vet. Science 42:404-406




















