
DONES DE CINE, AL CINE I AMB EL CINE XII CERTAMEN DE NARRATIVA BREU
Upload
independentCategory
view
1download
0
Volume 11 Number2l 1983 Nucleic Acids Research
Isolation and characterisation of cDNA dones for the Aa- and y-chains of human fibnnogen
A.M.A.Imam, M.A.W.Eaton1, R.Williamson and S.Humphries
Department of Biochemistry, St.Mary's Hospital Medical School, University of London, LondonW2 IPG, and 'Department of Chemistry, Celltech Ltd., 250 Bath Road, Slough SLI 4DY, UK
Received 26 July 1983; Revised and Accepted 13 October 1983
ABSTRACTcDNA clones coding for the Ao(- and Y-chains of human
fibrinogen have been isolated from an adult liver cDNA library.Clones were identified by hybridisation with mixtures ofsynthetic oliqonucleotides 17 bases long, predicted using aminoacid sequence data for each chain. The cDNA insert sizes are1,950bp for Ao(-fibrinogen and 950b] for r-fibrinogen. Theclones do not show any cross-hybridisation. Each cDNAhybridises to a unique sequence in the human genome. In adulthuman liver, Northern blots give an estimated messenger RNAsize of 2.6kb for Ao( -fibrinogen and 1.8kb for Y-fibrinogen.
INTRwUrICNFibrinogen (factor I) is an abundant protein present at 0.3 mg/ml in human
plasna and synthesised in the liver. It is an acute phase protein whichparticipates in the final steps of the coagulation pathway in higher vertebrates.A nuwber of genetically determined variants for human fibrinogen have beenreported (1). Elevated levels of fibrinogen arnd other coagulation factors, suchas factor VII and factor VIII, have been correlated with a predisposition toischanic heart disease (2,3).
Native fibrinogen (MW 340,000) is ncmuposed of dimers ofAo(- (MW 64,000), BoB- (MW 56,000) and )- (MW 47,000) chains linked bydisulphide bridges. The Bp- and >-fibrinogen chains are glycosylated atresidues 364 ard 52 respectively. The N-termini of the A&- ard B)-fibrinogenchains respectively terminate in fibrirpeptides A ard B, which are cleavedsequentially by thrarbin to give fibrin nonamers. These polymerise to form anetwrk of fibres. A transanidation reaction catalysed by factor XIIIacovalently links the fibres, following which platelet aggregaticn ocmpletes clotformation.
The aoplete amino acid sequ of all three chains have been determined(4-7). The chains sh sequen logy, suggestirn that a acaun ancestralsequence underwet duplication and divergence. The chains are rich in Trp andMet, a fact used to our advantage in determning regicns of low ambiguity for thesynthesis of gene-specific oligonucleotide mixtures to be used in isolating theclones.
The isolation of clones codiun for rat and hbnan fibrinogen has recently
©) I R L Press Limited, Oxford, England.
Nucleic Acids ResearchVolume 1 1 Number 21 1983
7427

Nucleic Acids Research
a Chainamino acids-CYS -PRO-GLU -ALA-M ET-ASP-472-477
cDNA 3-ACA-GGN-CTT-CGN-TAC -CT-5G C
y Chainamino acids -TRP - TRP-MET-ASN-LYS - CYS-334-339
cDNA 3ACC-ACC-TAC-TTG-TT -AC-5A C
Fig.l Sequence of synthetic oligonucleotide mixtures used forscreening the adult human liver cr)rA library. The Ao(-fibrinogen probe is a mixture of 64 different oligonucleotides.The )-fibrinogen probe is a mixture of 4 oligonucleotides.
been reported (8-12). Here we report the isolation and characterisation ofcDNAs corresponding to the hunan Ac(- and y-fibrinogen chains. We use theseclones in a preliminary analysis of the structure and expression of hunanfibrinogen genes.
MATERIALS AND METHODS
Screenin9 of an adult human liver cI)NA lirr
The 17-base lonq oligonucleotide probes were synthesised by the solid-phasephosph,triester nmthod as described by Snith et al (13). The cDNA library usedwas generously provided by Dr. Derek Woods and has been previously described(14). 1,000 clones were plated on a 8amn nitrocellulose filter, which wasreplica-plated onto nitrocellulose filters of the same size, amplified usingchloramphenicol and screened with the oligonucleotide mixtures using amodification of the method of Hanahan and Meselson (15). The oligonucleotideswere 5'- labelled with Y-[32P]ATP (5,000 Ci/nM), using T4 polynucleotidekinase (PL Biochenicals) (16). Oligo(dT), 12-18 nucleotides in length(Boehringer), was similarly labelled to a specific activity of lxlO1cpn/ug.Hybridisation to the filter-bound DM was performed in a solution containing 0.9MNaCl/0.09M sodium citrate/standard Denhardt' s solution/0.05% sodiunpyroposphate/50 ug/ml sonicated herring sperm DNA/10 ug/ml poly(A) andlabelled oligonucleotide probe (5x106crmsiq) for four hours at 37 . Afterhybridisation the filters were washed in 0.9M NaCl/0.09M sodium citrate/0.05%sodium pyrophosphate at 370 for 2x30 min and then sequentially at 460 for 15 minand 50for 2 min. Washed filters were exposed to preflashed Fuji X-ray film at -700, using an Okamoto tungstate intensifying screen.Plasmid preparation and DNA analysis
Bacterial clones were grown in L,-broth supplemented with tetracycline (25ug/ml). Plasmid DNA was prepared by a modification of the cleared lvsate method(17) and purified on CsCl gradients. Restriction enzymes were purchased from
7428

Nucleic Acids Research
AB.
Fig.2 Screening of 1,000 recombinants from the cDNA librarywith A.l-oligonucleotide (A) and Y-oligonucleotide (B).Filter hybridisation and post-hybridisation washes were as inMaterials and Methods. Exposure to X-ray film was for 16 hrs.
Bethesda Research Labs; DNA digests were performed according to themanufacturer's recommendation, and the cleaved fragments were analysed onagarose or acrylamide gels. Restricted human DNA samples (10 ugs/slot) were runon 1% agarose gels and blotted onto nitrocellulose filters as ro.ribed bySouthern (18). Plasmid probes were labelled by nick translation (19), uiina 0-[32P]dCTP, 400 Ci/ntM (Amersham), and hybridised with the addition of dextransulphate to 10% in the prehybridisation and hybridisation solutions (20). DNAsequencing was carried out by the enzymatic method of Sanger (21). The cDNAclones were digested with PstI and subcloned into the PstI site of the M13mp9polylinker. The resultinq reccwbinant single strandedM13phaqeDNAwasusedasa template for sequencing reactions, primed with the M13 universal primer.RI preparation and Northern blotting
A post mnrtem specimen of male adult human liver was taken 30 min afterdeath, frozen immediately in liquid nitrogen and stored at -70 RNA was
prepared as described by Auffray and Pougeon (22), denatured by boiling in 2.2Mformaldehyde/50% formamide, run on a 1% agarose/2.2M formaldehyde denaturinggel in MOPS (2QmM Na-MOPS/54 sodium acetate/lmm EDTA) buffer and blotted ontonitrocellulose paper.
RESLTSIdentification of A - and -fibrinogen cDNA clones
1,000 recambinants of an adult liver cDNA library were screenei v'ith 17-k,aselong mixtures of oligonucleotide specific for A o(- and Y-fibrinogens. Fig 1shows the amino acid sequences used to construct the oligonucleotides and thecorresponding mixtures of DNA fragments. The A d-fibrinogen probe is a
7429

Nucleic Acids Research
Clone pAFl insert DNA sequence
91Met Glu Ile Leu Arg Gly Asp
CTGCAGGGGGG GGCGGGGGGCG ATG GAA ATT TTG AGA GGC GAT10 20 30 40
PstI
101 111Phe Ser Ser Ala Asn Asn Arg Asp Asn Thr Tyr Asn Arg Val Ser GluTTT TCC TCA GCC AAT AAC CGT GAT AAT ACC TAC AAC CGA GTG TCA GAG
50 60 70 80 90
121Asp Leu Arg Ser Arg Ile Glu Val Leu Lys Arg Lys Val Ile GluGAT CTG AGA AGC AGA ATT GAA GTC CTG AAG CGC AAA GTC ATA GAA
100 110 120 130
131Lys Val Gln His Ile Gln Leu Leu GlnAAA GTA CAG CAT ATC CAG CTT CTG CAG
(A)140 150 160
PstI
Clone pGF1 insert DNA sequence
CTGCAGGGGGG GGGGGGGGC10 20
PstI
183 193Glu Ile Asp Gly Ser Gly Asn Gly Trp Thr Val Phe Gln Lys Arg LeuGAA ATC GAT GGG TCT GGA AAT GGA TGG ACT GTG TTT CAG AAG AGA CTT
30 40 50 60
203 213Asp Gly Ser Val Asp Phe Lys Lys Asn Trp Ile Gln Tyr Lys Glu GlyGAT GGC AGT GTA GAT TTC AAG AAA AAC TGG ATT CAA TAT AAA GAA GGA70 80 90 100 110
223Phe Gly His Leu Ser Pro Thr Gly Thr Thr Glu Phe Trp Leu Gly AsnTTT GGC CAT CTC TCT CCT ACT GGC ACA ACA GAA TTT TGG CTG GGA AAT
120 130 140 150 160
233 243Glu Lys Ile His Leu Ile Ser Thr Gln Ser Ala Ile Pro TyrGAG AAG ATT CAT TTG ATA AGC ACA CAG TCT GCC ATC CCA TAT
170 180 190 200
Fig.3 Partial nucleuotide sequence of clones pAFl and pGFlinserts and the amino acid sequences derived from them. Thenumbering of amino acids is as previously reported (4-7). Thenucleotide in parentheses is that reported by Kant et al (9).
mixture of 64 different oligonucleotides, and the r'-fibrinogen probe amixture of four oligonucleotides. For Ad- fibrinogen a single positive colony(Fig 2A) was detected, while three positives (Fig 2B) were found using the Y-fibrinogen probe. The positive clones were picked into ordered arrays andrescreened with each of the oligonucleotide mixtures. Plasmid DEA fran each clone
7430
Nucleic Acids Research
A B
kb1 2 3 4 orl-gin kb312 3 Origin!Sb~ ~ riin kb23123.--9-6-'4 9'6- -96- ~~~ 9*-6-66 *664.4- 4.4-:
2-3 23 F2 0 2 0-
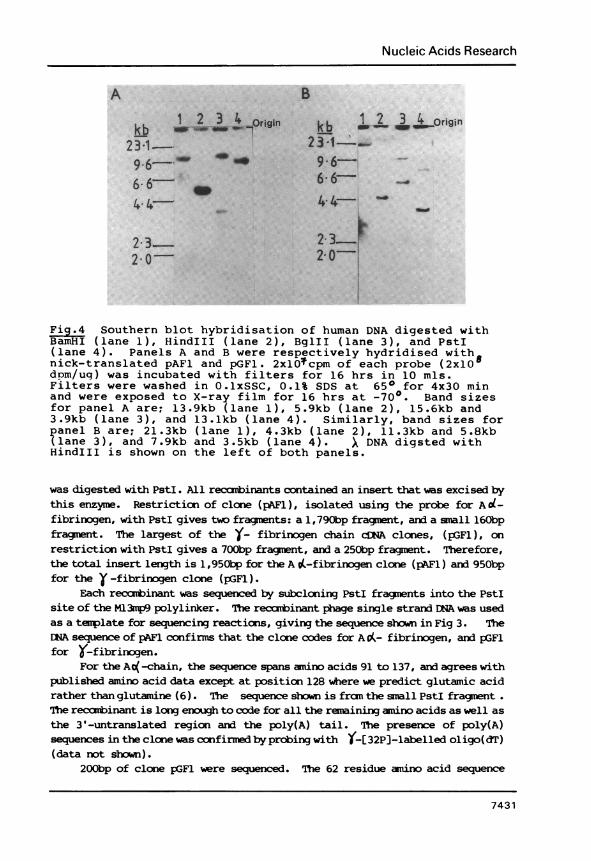
Fig.4 Southern blot hybridisation of human DNA digested withBamHI (lane 1), HindIII (lane 2), BglII (lane 3), and PstI(lane 4). Panels A and B were respectively hydridised withnick-translated pAFl and pGFl. 2xlOcpm of each probe (2x1O8dom/uq) was incubated with filters for 16 hrs in 10 mls.Filters were washed in O.lxSSC, 0.1% SDS at 650 for 4x30 minand were exposed to X-ray film for 16 hrs at -70°. Band sizesfor panel A are; 13.9kb (lane 1), 5.9kb (lane 2), 15.6kb and3.9kb (lane 3), and 13.lkb (lane 4). Similarly, band sizes forpanel B are; 21.3kb (lane 1), 4.3kb (lane 2), 11.3kb and 5.8kb(lane 3), and 7.9kb and 3.5kb (lane 4). X DNA digsted withHindIII is shown on the left of both panels.
was digested with PstI. All recanbinants contained an insert that was excised bythis enzyme. Restriction of clone (pAFl), isolated using the probe for Ad-fibrinogen, with PstI gives two fragments: a 1,790bp fragment, and a small 160bpfragment. The largest of the )- fibrinogen chain cfNA clones, (pGFl), onrestriction with PstI gives a 700bp fragment, and a 250bp fragment. Therefore,the total insert length is 1,950bp for the A i-fibrinogen clone (pAF1) and 950bpfor the ' -fibrinogen clone (pGFl).
Each recambinant was sequenced by subcloning PstI fragments into the PstIsite of the Ml3mp9 polylinker. TIe reccbinant phage single strand DNA was usedas a template for sequencing reactions, giving the sequence shown in Fig 3. TheENA sequence of pAFl confirms that the clone codes for A 0*- fibrinogen, and pGFIfor Y-fibrinogen.
For the Ao -chain, the sequence spans amino acids 91 to 137, and agrees withpublished amino acid data except at positicn 128 where we predict glutamic acidrather than glutamine (6). The sequence shown is fran the small PstI fragment .The recambinant is long enough to code for all the remaining amino acids as well asthe 3 '-untranslated region and the poly(A) tail. The presence of poly(A)sequences in the clone was confirmed by prcbing with J-[32P]-labelled oligo(dT)(data not shown).
200bp of clone pGFl were sequenced. The 62 residue amino acid sequence
7431
Nucleic Acids Research
Origin- a b
kb4-85-
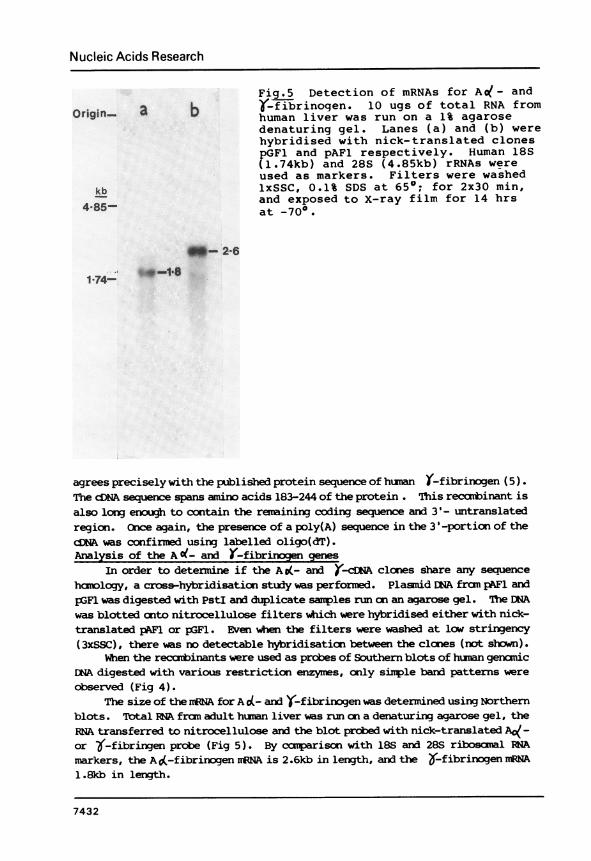
Fig.5 Detection of mRNAs for A l- and(-fibrinogen. 10 ugs of total RNA fromhuman liver was run on a 1% agarosedenaturing gel. Lanes (a) and (b) werehybridised with nick-translated clonespGF1 and pAFl respectively. Human 18S(1.74kb) and 28S (4.85kb) rRNAs wereused as markers. Filters were washedlxSSC, 0.1% SDS at 65'; for 2x30 min,and exposed to X-ray film for 14 hrsat -70°.
in- 2*6
1 74- " -18
agrees precisely with the published protein sequence of hunan (-fibrinogen (5).The cDNA sequence spans amino acids 183-244 of the protein . This recanbinant isalso long enugh to contain the remaining coding sequence and 3'- untranslatedregion. Cnce again, the presence of a poly(A) sequence in the 3'-portion of thedNA was confirmed using labelled oligo(dT).Analysis of the A O(- and (-fibrinogen genes
In order to determine if the A v(- and )-cDMA clones share any sequenceharmlogy, a cross-hybridisation study was performed. Plasmid DNA framn AF1 and
pGFl was digested with PstI and duplicate samples run cn an agarose gel. The DNAwas blotted onto nitrocellulose filters which were hybridised either with nick-translated pAEl or pGFl. Even when the filters were washed at low stringency(3xSSC), there was no detectable hybridisaticn between the clcnes (not shown).
When the recambinants were used as probes of Southern blots of human gencnicDNA digested with various restriction enzymes, only simple band patterns wereobserved (Fig 4).
The size of the rNA for A o(- and Y-fibrinogen was determined using Northernblots. Total RA fran adult human liver was run on a denaturing agarose gel, theRNA transferred to nitrocellulose and the blot prcbed with nick-translated A(-or ~-fibringen probe (Fig 5). By canparison with 18S and 28S ribosamal RNAmarkers, the A 0(-fibrinogen nrNA is 2.6kb in length, and the &-fibrinogen iRNA1.8kb in length.
7432

Nucleic Acids Research
DISCUSSIONThe liver synthesises a number of proteins involved in coagulation,
including the abundant protein fibrinogen. Consequently, only 1,000 clones ofan adult human liver cDNA library were screened with fibrinogen-specificoligonucleotides.Clones pAFl and pGFl, corresponding to the A o- and Y-fibrinogen chains respectively, were isolated by screening with 17-base longoligonucleotides camplementary to regions of each protein chain showing lowambiguity in codon assignment.
The partial DNA sequence of the pAFl insert agrees with the correspondingregion of the cDNA sequence for Ap - fibrinogen reported by Kant et al (9),except at one place. At the codon for glutamine 137, a change from A to G createsa PstI site. This internal PstI site has been confirmed by our restrictiondigest data . The Ad- fibrinogen cDNA sequence of Rixon et al (10) also shows thepresence of this site. At present we are determining the frequency of occurence ofthis PstI site polymorphism in the human population. In agreement with thepublished Ai-fibrinogen cDNA sequences (9,10), we also assign residue 128 asglutamic acid.
The 200bp sequence of pGF1 confirms the identity of the clone, and thepredicted amino acid sequence agrees precisely with the published sequence ofhuman (-fibrinogen. Recently, the entire sequence of the rat (-fibrinogencDNA has been reported (23). By comparison with the rat sequence, there are 8/62amino acid changes (12.9%) and 30/206 base changes (14.5%) in the region we havesequenced. Of the 30 base changes, 19 are silent substitutions; the remaining 11cause 8 amino acid changes. Therefore, this region of >-fibrinogen is not ashighly conserved in evolution as the carboxyterminal region (9).
Both clones give simple band patterns when used as probes of Southern blotsof total human genamic DNA, demonstrating that they represent unique sequencesand donot constitute a multigene family.
The sizes of the A o4- and Y-fibrinogen mRNAs we detect in hunan liver aresimilar to those reported in the rat (8) (A g- 2.3kb , - 1.95kb). Theseobservations in human liver confirm that each fibrinogen chain is coded by an
independent nRA and not by a caruon RNA transcript that either undergoessubsequent differential splicing, or is translated to give a polyprotein. Inthe Northern blot, the intensity of the hybridisation signal with the A £,-
fibrinogen probe is greater than for '(- fibrinogen. However, the Ad -
fibrinogen insert is roughly twice as large as the {(-fibrinogen insert, andtherefore we estimate that the two mRNAs are present in almost equal abundance.
CI Y_LEDGEWe thank Dr. S. Wallis for supplying adult human liver RNA, Paul North for
help with the computer search of protein sequences, and Padi Kosol for help inpreparing the manuscript. This work was su bortedby grants fran the BritishHeart Foundation and the Medical Research Council.
7433

Nucleic Acids Research
REFERENCES1. Bloom,A.L. (1981) in Haemostasis and Thrombosis, Bloom,A.L.
and Thomas,D.P. eds., pp.321-370, Churchill Livingstone,London.
2. Mead,T.W. and North,W.R.S. (1977) Brit.Med.Bull. 33,283-288.
3. Mead,T.W., North,W.R.S., Chakrabarty,R., Stirling,Y.,Haines,A.P. and Thompsom,S.G. (1980) Lancet 17, 1050-1053.
4. Blomback,B., Hessel,B. and Hogg,D. (1976) Thromb.Res. 8,639-658.
5. Lottspeich,R. and Henschen,A. (1977) Hoppe-Seyler's Z.Physiol.Chem. 358, 935-938.
6. Doolittle,R.F., Watt,K.K., Cottrell,B.A., Strong,D.D. andRiley,M. (1979) Nature 280, 464-468.
7. Watt,K.W.K., Tagaki,T. and Doolittle,R.F. (1979)Biochemistry 18, 68-76.
8. Crabtree,G.R. and Kant,J. (1981) J.Biol.Chem. 256,9718-9723.
9. Kant,J.A., Lord,S.T. and Crabtree,G.R. (1983) Proc.Natl.Acad.Sci.USA 80, 3953-3957.
10. Rixon,M.W., Chan,W-Y., Davie,E.W. and Chung,D.W. (1983)Biochemistry 22, 3237-3244.
11. Chunq,D.W., Que,B.G., Rixon,M.W., Mace,M. and Davie,E.W.(1983) Biochemistry 22, 3244-3250.
12. Chung,D.W., Chan,W-Y. and Davie,E.W. (1983) Biochemistry22, 3250-3256.
13. Smith,J., Cook,E., Fotheringham,I., Pheby,S.,Derbyshire,R., Eaton,M.A.W., Doel,M., Lilley,D.M.J.,Pardon,J.F., Patel,T., Lewis,H. and Bell,L.D. (1982) Nucl.Acids Res. 10, 4467-4482.
14. Woods,D.E., Markham,A.F., Ricker,A.T., Goldberger,G. andColten,H.R. (1982) Proc.Natl.Acad.Sci.USA 79, 5661-5665.
15. Hanahan,D. and Meselson,M. (1980) Gene 10, 63-67.16. Maxam,A.M. and Gilbert,W. (1980) Methods Enzymol. 65,
499-560.17. Birnboim,H.C. and Doly,J. (1979) Nucl.Acids Res. 7,
1513-152318. Southern,E. (1979) Methods Enzymol. 68, 152-176.19. Rigby,P.W.J., Deickmann,M., Rhodes,C. and Berg,P. (1977)
J.Mol.Biol. 113, 237-251.20. Jeffreys,A.J. and Flavell,R.A. (1977) Cell 12, 429-439.21. Sanqer,F., Nicklen,S. and Coulson,A.R. (1977) Proc.Natl.
Acad.Sci.USA 74, 5463-5467.22. Auffray,C. and Rougeon,F. (1980) Eur.J.Biochem. 107,
303-314.23. Crabtree,G.R. and Kant,J.A. (1982) Cell 31, 159-166.
7434
Copyright © 2022 FDOKUMEN




















