
UK children's breakfast cereals – an oral health perspective

Journal of Cereal Science 48 (2008) 258e276www.elsevier.com/locate/jcs
Review
Is the in vitro antioxidant potential of whole-graincereals and cereal products well reflected in vivo?
Anthony Fardet*, Edmond Rock, Christian Remesy
Unite de Nutrition Humaine, INRA-UMR 1019, Centre de Recherche de Clermont-Ferrand/Theix, 63122 Saint-Genes-Champanelle, France
Received 14 November 2007; received in revised form 22 January 2008; accepted 23 January 2008
Abstract
There is strong epidemiological evidence that whole-grain cereals protect the body against age-related diseases such as diabetes, cardiovas-cular diseases and some cancers. This may be due to the fibre and micronutrients in the outer layer and germ fractions of the grain acting togetherto combat oxidative stress, inflammation, hyperglycaemia and carcinogenesis. Oxidative stress is associated with these metabolic diseases.Whole-grain cereals are a good source of vitamin E, folates, phenolic acids, zinc, iron, selenium, copper, manganese, carotenoids, phyticacid, lignins, lignans, and alkylresorcinols, all of which have significant antioxidant potential in vitro. Phenolic acids such as ferulic acid arecharacteristic of cereals. They may scavenge free-radical oxygen species both in vitro and in vivo. Phenolics may also act in vivo by triggeringgene induction/repression via cell signalling through transcription factors. Whole-grain cereals are also a good source of betaine, choline andsulphur amino acids that can improve antioxidant status. Betaine, which accounts for about 1% (w/w) of the bran fraction in wheat, acts asa methyl donor that may decrease hyperhomocysteinaemia, a cardiovascular risk factor, but it has been neglected. Cereals and cereal productsare antioxidative in vitro, as are most fruits and vegetables. The in vitro antioxidant capacity of cereals and their constituent fractions is signif-icantly correlated with their polyphenol content, except for maize. However, the in vitro antioxidant capacity of cereals is only an approximatereflection of their in vivo antioxidant effect due to differences in antioxidant solubility/bioavailability within the digestive tract and the metab-olism/conjugation of compounds such as polyphenols. During digestion, the antioxidant capacity of cereals is increased and is likely to providea favourable antioxidative environment for the epithelium tract, notably in the large intestine. Most of the studies performed on animals havebeen concerned with the antioxidant property of coloured rice, especially black rice and its anthocyanin fraction, showing a positive effect onsome antioxidant biomarkers. Those very few studies that have been done on humans have shown that wheat bran and rye product supplementshave no effect on antioxidant status, while a black rice pigment fraction and an avenanthramide-enriched mixture extracted from hulled oats havea positive effect. In vivo studies are therefore needed to further explore the real antioxidant potential of cereals.� 2008 Elsevier Ltd. All rights reserved.
Keywords: Whole-grain cereals; Micronutrients; In vitro Antioxidant potential; In vivo studies
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2592. Whole-grain cereals as a source of antioxidants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 259
2.1. Vitamin E . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2602.2. Folates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2602.3. Minerals and trace elements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 260
Abbreviations: AAPH, 2,20-azobis 2-amidinopropane dihydrochloride; ABTS, 2,20-azinobis 3-ethyl-benzothiazoline-6-sulfonic acid; CAT, catalase; DPPH,
2,2-diphenyl-1-picrylhydrazyl; FRAP, Ferric Reducing Antioxidant Power; GSH, reduced glutathione; GSH-Px, glutathione-peroxidase; GSH-Red, glutathione-
reductase; GSSG, oxidized glutathione; LDL, low-density lipoprotein; MDA, malondialdehydes; ORAC, Oxygen Radical Absorbance Capacity; SOD, superoxide
dismutase; TBARS, thiobarbituric acid reacting substances; TE, trolox equivalent.
* Corresponding author. Tel.: þ33 (0)4 73 62 44 00; fax: þ33 (0)4 73 62 46 38.
E-mail address: [email protected] (A. Fardet).
0733-5210/$ - see front matter � 2008 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jcs.2008.01.002

259A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
2.4. Polyphenols . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2602.5. Alkylresorcinols . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2622.6. g-Oryzanol . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2622.7. Betaine, choline and sulphur amino acids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2622.8. Phytic acid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2622.9. Lignins and lignans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2622.10. Carotenoids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 263
3. In vitro antioxidant potential of whole-grain cereals and their different fractions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2633.1. In vitro antioxidant potential of cereals and cereal products . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2633.2. Wheat . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2643.3. Corn . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2643.4. Rice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2653.5. Oats . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2653.6. Barley . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2653.7. Rye . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2663.8. Sorghum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2663.9. Finger millet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 266
4. In vivo antioxidant potential of whole-grain cereals and their fractions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2664.1. Ex vivo oxidation of LDL . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2664.2. Bioavailability and influence of digestion on the antioxidant capacity of cereal products . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2674.3. Animal studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2674.4. Human intervention studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2704.5. Epidemiological studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2714.6. Whole-grain cereals versus fruits and vegetables . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 271
5. Conclusions and perspectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 271References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 272
1. Introduction
Epidemiological studies have clearly demonstrated thata diet containing whole-grain cereals can protect against met-abolic disorders such as cardiovascular diseases (Anderson,2003; Truswell, 2002), cancer (Chatenoud et al., 1998, 1999)and diabetes (Venn and Mann, 2004). The effect is mainly at-tributed to the fibre and micronutrients in the outer layer of thegrain and in the germ fraction (Slavin et al., 1999; Thompson,1994). However, the bran fraction plays the more importantrole due to the germ released during cereal processing. Theprotective effects of cereal fibres depend on their solubility.Soluble fibre (soluble arabinoxylans and b-glucans) can lowerblood cholesterol (Braaten et al., 1994; Kahlon et al., 1993)and reduce the post-prandial glycaemic response (Casiraghiet al., 2006). Insoluble fibres (cellulose and insoluble arabi-noxylans) increase the speed of transit time and the faeces vol-ume (ballast effect), decreasing the contact betweencarcinogens and the colon epithelial cells, and their fermenta-tion may also produce significant quantities of butyrate, whichitself protects epithelial cells against carcinogenesis (Olmoet al., 2007). Whole-grain cereals also contain micronutrientssuch as vitamin E, folates, phenolic acids, zinc, iron, selenium,copper, manganese, carotenoids, betaine, choline, sulphuramino acids, phytic acid, lignins, lignans, and alkylresorcinols,which all may have potential antioxidant effects. They may actas such in vivo to improve the redox state of various tissues(Slavin et al., 1999). The in vivo bioavailability of each ofthese micronutrients varies, and some, such as the polyphenols
may be metabolised. Thus, some micronutrients may lose theirantioxidant potential or their concentration in the plasma andtissues may be too low for them to act as significant free rad-ical scavengers. Other mechanisms are probably also involved.The in vivo reality is therefore much more complex than it ap-pears in a first view. Obesity and metabolic diseases have beenall associated with increased oxidative stress (Aruoma, 1998;Furukawa et al., 2004; Hwang and Kim, 2007). Antioxidantphytomicronutrients from whole-grain cereals could thereforeprotect the body from this oxidative stress. Exactly how theyexert their protective effect remain to be elucidated.
This review discusses the potential antioxidant effect ofwhole-grain cereals, their various fractions, and cereal prod-ucts, relating their in vitro activities to their antioxidant effectsin vivo. We first discuss the antioxidant properties of cerealmicronutrients and their significance, followed by the in vitroantioxidant capacity of cereals and their fractions. Lastly, wereview the main in vivo studies on the antioxidant effect ofcereals, a brief comparison being made with studies carriedout with fruits and vegetables.
2. Whole-grain cereals as a source of antioxidants
Whole-grain cereals contain a much wider range of com-pounds with potential antioxidant effect than do refined ce-reals. These include vitamin E (mainly in the germ), folates,minerals (iron, zinc) and trace elements (selenium, copperand manganese), carotenoids, phytic acid, lignin and othercompounds such as betaine, choline, sulphur amino acids,

260 A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
alkylresorcinols and lignans found mainly in the bran fraction.Some, such as vitamin E, are considered to be direct free rad-ical scavengers, while others act as cofactors of antioxidantenzymes (selenium, manganese and zinc), or indirect antioxi-dants (folates, choline and betaine).
2.1. Vitamin E
Vitamin E (tocopherols and tocotrienols) is a well-known an-tioxidant: it acts as a free radical scavenger, more specificallywithin cell membranes by preventing the oxidation of polyun-saturated lipids by free radicals such as the hydroxyl radicalOH$. The antioxidant potential of vitamin E is no longer dis-puted. Most in vivo studies have shown that vitamin E improvesvarious parameters of oxidative stress in both animals (Goles-tani et al., 2006) and humans (Martin et al., 1996). Rats wereprotected against the effects of N-nitrosodiethylamine, a hepaticcarcinogen that causes oxidative stress, by pre-treatment withvitamin E (Bansal et al., 2005). Another study, in which obeseZucker rats were fed a diet supplemented with vitamin E, foundtheir plasma 8-epi-prostaglandin F2a was reduced (Laight et al.,1999). Other recent experiments in which rats were fed wheatgerm/wheat germ oil (Leenhardt et al., in press) or breakfast ce-reals (Mitchell et al., 1996) showed that vitamin E bioavailabil-ity from cereals was quite satisfactorily. Similar finding havebeen obtained in humans (Leonard et al., 2004). The cerealsrichest in vitamin E are whole-grain maize and rye whilewhole-grain wheat, barley, oat, millet, sorghum and buckwheatcontain less vitamin E (Table 1). Generally, vitamin E is mainlylocated within the germ of whole-grain cereals, from whichvitamin E-rich oils are extracted. Thus, wheat germ containsaround 25 mg vitamin E/100 g of product (Souci et al., 2000).Refined cereals contain insignificant quantities of vitamin Edue to the loss of vitamin E during refining.
2.2. Folates
Folates (vitamin B9) reduce the concentration of plasma ho-mocysteine (Ward et al., 1997). Hyperhomocysteinaemia is anindependent cardiovascular risk factor, because of its pro-oxi-dative effect. Folates appear to be indirect, but significant, anti-oxidants. Cereals are a moderate source of folates as theycontain from 16 to 143 mg folates/100 g edible portion, depend-ing on the type of cereal (Souci et al., 2000) (Table 1). However,rye (143 mg/100 g grain) and maize (110e170 mg/100 g grain)are better source for folates compared to wheat (87 mg/100 ggrain). Wheat germ (520 mg/100 g) and bran (195 mg/100 g)are good source of folates (Souci et al., 2000).
2.3. Minerals and trace elements
Minerals such as zinc (Zn) and iron (Fe), and trace ele-ments including selenium (Se), copper (Cu) and manganese(Mn), are cofactors of several antioxidant enzymes: superox-ide dismutase is Zn, Cu and Mn dependent, glutathione perox-idase and thioredoxine reductase are Se dependent, andcatalase is Fe dependent. The mineral and trace element
contents vary greatly from one type of cereal to another (Souciet al., 2000) (Table 1) and with the location where they aregrown, especially for Se (Curtin et al., 2006; Zhao et al.,2007). One essential question about these compounds is theirbioavailability, as it is reduced by the phytic acid present inhigh concentrations in whole-grain cereals. However, phyticacid can be broken down by phytases in a pre-fermentationstep, such as that occurring in sourdough breads at pH 5e5.5 (Leenhardt et al., 2005), which significantly increases min-eral bioavailability, especially that of Fe and Zn (Lopez et al.,2003). Whole-grain cereals provide much more minerals thanrefined cereals, and this is quite enough to compensate for thedeleterious effect of phytic acid. While more studies on hu-mans are needed to investigate the bioavailability of mineralsand trace elements from whole-grain cereal products, whole-grain cereals are quite likely to help maintain the body’s min-eral and trace elements status and contribute to the antioxidantpotential in vivo because they are found mainly in the branfraction, especially the aleurone layer. Refined cereals, there-fore, contain significantly less mineral and trace elements. Re-fining leads to the loss of 70% of Fe, and around 75e80% ofZn and Mn (Feillet, 2000).
2.4. Polyphenols
Whole-grain cereals are a major source of polyphenols, es-pecially phenolic acids such as ferulic, vanillic, caffeic, syrin-gic, sinapic and p-coumaric acids (Sosulski et al., 1982). All ofthem have potentially antioxidant properties due to the presenceof an aromatic phenolic ring that can stabilize and delocalizethe unpaired electron within its aromatic ring (Rice-Evanset al., 1997). However, their mechanisms of action in vivoare not fully elucidated. They are believed to act mainly asfree-radical scavengers, and/or chelators of transition metals(minerals or trace elements). But it is unlikely that theirfree-radical scavenging capacity is sufficient to explain theirantioxidant action in vivo, due to their relatively low bioavail-ability within the digestive tract (0.3e26%) (Scalbert and Wil-liamson, 2000). Their mechanisms of action are probablylinked to the activation/repression of particular genes. Recentinvestigations have shown that they can also act on gene ex-pression via transcription factors (e.g. Nrf2), activating theAntioxidant Response Element (Lee et al., 2003; Na andSurh, 2006), which can lead to the transcription of antioxidantcompounds such as glutathione (GSH), or enzymes of gluta-thione metabolism (Myhrstad et al., 2002). In addition, ab-sorbed polyphenols are mostly metabolized and conjugated(Scalbert and Williamson, 2000). For example, limitedamounts of ferulic and sinapic acids are absorbed and thesecome only from the free and soluble portions present in the ce-real (Kern et al., 2003). Ferulic acid is esterified to arabinoseresidues in the cell wall arabinoxylans. However, the boundfractions of ferulic acid and other phenolics can be releasedlater in the colon by fermentation (Adom and Liu, 2002).The ferulic acid and/or its conjugated forms (e.g. glucuro-nides) that crosses the intestinal epithelium exert its antioxi-dant properties from the aqueous phase of the blood plasma,

Table 1
Antioxidant micronutrients in whole-grain cerealsa
Cereal type (variety) Tocopherol and
tocotrienol
(mg/100 g grain)
Folates
(mg/100 g
grain)
Zinc
(mg/100 g
grain)
Iron
(mg/100 g
grain)
Copper
(mg/100 g
grain)
Selenium
(mg/100 g
grain)
Manganese
(mg/100 g
grain)
Polyphenols
(mg gallic acid eq./
100 g grain)
Wheat 1.4 87 2.6 3.2 369 0.5e74.6 3.1 70e1459
Maize (yellow, white & blue) 6.6 110e170 1.7 1.5 240 12.0 0.4 39e711
Rice (brown & black) 1.9 16 1.6 3.2 288 10.0 2.1 54e313
Oats 1.8 33 3.2 5.8 422 7.1 3.1 9e34
Barley 2.2 65 2.8e7.4 2.8e12.8 419e570 7.0 1.5 50e196
Rye 4.1 143 2.9e3.1 2.8e4.4 290e392 1.4 2.9 125e255
Sorghum (white, yellow, red & brown) 1.1 eb 0.3e1.8 1.1e5.7 20e735 13.0 4.3 100e2300
Millet (shucked grain) 4.0 e 2.9e6.6 6.9e20 340e610 2.0 1.1 29e47
Ferulic acid
(mg/100 g
grain)
Betaine
(mg/100 g)
Choline
(mg/100 g)
Phytic acid
(mg/100 g
grain)
Lignin
(g/100 g
grain)
Lignan
(mg/100 g
grain)
Carotenoids
(mg/100 g
grain)
Alkylresorcinols
(mg/100 g
grain)
Wheat 10e198 6.9 (flour & germ) 1.6 (flour & germ) 906 0.6e1.3 490 20e265 28e142
Maize (yellow, white & blue) 177 e e 940 e e 969e1300 eRice (brown & black) 30 0.5 (brown) e 890 e e 14e77 0
Oats 7e30 2.7 (plain) 1.3 (plain) 900 e e 31 e
Barley 36e62 e e 1070 e e 15e105 4e44
Rye 79e102 e e 970 e 95 e 31e278
Sorghum (white, yellow, red & brown) 9 e e e e e 20e22 0
Millet (shucked grain) 29 e e e e e 74e80 9e87
a All values given in the table are adapted from Adom and Liu, 2002; Andreasen et al., 2000; Awika et al., 2003; Choi et al., 2007; Del Pozo-Insfran et al., 2006; Dewanto et al., 2002; Dykes and Rooney, 2006;
Emmons et al., 1999; Hernanz et al., 2001; Li et al., 2004; Liu and Yao, 2007; Liyana-Pathirana and Shahidi, 2006a,b; Maillard et al., 1996; Michalska et al., 2007; Moore et al., 2005; Nilsson et al., 1997; Panfili
et al., 2004; Parikka et al., 2006; Ragaee et al., 2006; Ross et al., 2003b; Schofield and Chen, 1995; Sosulski et al., 1982; Souci et al., 2000; U.S. Department of Agriculture, A.R.S., 1999; Zeisel et al., 2003b; Zhao
et al., 2007; Zhou et al., 2005. When data were expressed on a dry matter basis in literature, the values were modified and expressed on a fresh matter basis, i.e. per 100 g grain (assuming that cereal grains contain
around 13% water).b data not available.
26
1A
.F
ardetet
al./
Journalof
Cereal
Science48
(2008)258e
276

262 A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
where it is a more effective antioxidant against LDL oxidationthan the water-soluble antioxidant ascorbic acid (Castelluccioet al., 1996). Apart from the in vitro antioxidant potential ofphenolic acids, nothing is known, to our knowledge, abouttheir antioxidant potential in vivo when coming from cereals.
2.5. Alkylresorcinols
Alkylresorcinols are phenolic lipids (resorcinol derivativeswith a long hydrocarbon chain at position 5 of the phenolicring) present in significant amounts in whole-grain cereals, es-pecially wheat and rye (>500 m/g). They possess antioxidantproperties in vitro due to their hydrogen donor and radicalscavenging abilities (Parikka et al., 2006), but they are less ef-ficient than vitamin E (Kamal-Eldin et al., 2001). Alkylresor-cinols are considered as membrane-located antioxidants. Theyare readily absorbed (up to 80%) (Ross et al., 2003a) and canbe biologically active. They are therefore potential antioxi-dants in vivo. Their antioxidant potential depends on the chainlength, i.e. their amphiphilic nature, and their incorporationinto cell membranes. Thus, micromolar concentrations of ce-real grain alkylresorcinols can protect erythrocyte membranesagainst hydrogen peroxide-induced lipid oxidation (Kozubekand Nienartowicz, 1995). However, these molecules are read-ily broken down by extrusion processing and fermentationand/or baking (Winata and Lorenz, 1997).
2.6. g-Oryzanol
g-Oryzanol is a mixture of at least 10 phytosteryl ferulates(e.g. methylsterols esterified to ferulic acid). It is specificallyextracted from rice bran and is a powerful inhibitor of iron-driven hydroxyl radical formation. Its antioxidant activityhas been clearly demonstrated both in vitro (Juliano et al.,2005) and in vivo (Suh et al., 2005).
2.7. Betaine, choline and sulphur amino acids
Whole-grain cereals are a rich source of betaine and cho-line. Toasted wheat germ contains 152 mg choline/100 g ofproduct and 1240 mg betaine/100 g product, while the branfraction contains around 1% betaine (Zeisel et al., 2003a).Both compounds have long been neglected as a source of an-tioxidants from cereals. Betaine is a methyl donor to homocys-teine, allowing its conversion to methionine. As stated above,a high concentration of homocysteine is a cardiovascular riskfactor and can lead to oxidative stress. Betaine, which is read-ily available in the digestive tract, may help, together with fo-lates, to decrease oxidative stress in vivo by reducing theconcentration of homocysteine, as shown in rats fed with a be-taine-supplemented diet (Barak et al., 2003). Choline, as a pre-cursor of betaine, is also involved in the body’s antioxidantdefences. The contributions of betaine and choline from ce-reals to the antioxidant status of humans are not known. Stud-ies are therefore needed to investigate the potential antioxidanteffect of these compounds in humans consuming whole-graincereals. Whole-grain and refined cereals are also good sources
of the sulphur amino acids methionine (an essential aminoacid) and cysteine (a non-essential amino acid that can onlybe synthesized from methionine). Dietary cysteine contributesto the synthesis of glutathione, a major endogenous antioxi-dant, particularly in the liver (Mariotti et al., 2004); and methi-onine is readily used by the liver to produce cysteine via thecystathionine pathway, which also favours glutathione synthe-sis (Morand et al., 1997). Whole-grain and white wheat flourshave quite similar sulphur amino acid contents (2.3e2.5 g cys-teine/100 g protein, and 1.2e1.3 g methionine/100 g protein)(Shewry, 2007). Other cereals (barley, oat, rye, rice and maize)have similar cysteine and methionine contents (1.6e2.3 g cys-teine/100 g protein and 1.4e2.5 g methionine/100 g protein)(Shewry, 2007).
2.8. Phytic acid
Phytic acid (inositol hexaphosphate) is a major phosphorusstorage compound in most seeds and cereal grains, contribut-ing about 1e7% of their dry weight. It may account for morethan 70% of the total kernel phosphorus (Zhou and Erdman,1995). Phytic acid is mainly located in the bran fraction ofwhole-grain cereals, especially within the aleurone layer. Ithas long been considered as an antinutrient because it stronglychelates minerals and trace elements (see above). Agro-foodmanufacturers therefore considered that it must be releasedfrom cereals. However, its antioxidant potential in vitro isnow well recognized (Graf and Eaton, 1990). It suppressesFe-catalysed oxidative reactions, because of its capacity tochelate free Fe (Fenton reaction) and may be a potent antiox-idant in vivo, by e.g. suppressing lipid peroxidation (Grafet al., 1987). The hydroxyl radical OH$ generated by the inter-action of Fe with O2
�, causes DNA damage (Halliwell and Ar-uoma, 1991). Phytic acid may reduce the incidence of coloniccancer by a similar mechanism, and protect against otherinflammatory bowel diseases (Graf and Eaton, 1990). Phyticacid also inhibits xanthine oxidase-induced superoxide-depen-dent DNA damage (Muraoka and Miura, 2004). Xanthineoxidase, which generates superoxide anions (O2
�) during theoxidation of xanthine, is abundant within the intestine (Battelliet al., 1972). To summarize, phytic acid can act by formingchelates and suppressing Fe-catalysed damaging redox reac-tions, or by suppressing the oxidative damage caused to thegut epithelium, particularly in the colon where bacteria alsoyield oxygenated radicals. However, phytic acid was shownto have no antioxidant effects in rats (Rimbach and Pallauf,1998), despite these potential effects.
2.9. Lignins and lignans
Lignin biopolymers constitute 30% of plant biomass and be-long to the most abundant organic polymers on earth. Ligninsare a major component of whole-grain cereals, and mayaccount for 3e7% of the bran fraction. They were long con-sidered to be nutritionally inert within the digestive tract.Nevertheless, their polyphenolic structure confers potential an-tioxidant capacities. Little work has been done on the

263A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
bioavailability of lignins in the gut. Begum et al. (2004)showed that rats can metabolize lignins into mammalians li-gnans. Although it is quite reasonable to suppose that ligninshave antioxidant effects in vivo, particularly in the colon,most studies on their antioxidant potential have been per-formed in vitro (Reinosa et al., 1998), or on their influenceon DNA damage (oxidative lesions) in isolated mammaliancells (Labaj et al., 2004; Slamenova et al., 1999, 2000).Thus, diabetic rats (diabetic nephropathy) given subcutaneousinjections of nordihydroguairetic acid (an antioxidant and lip-oxygenase inhibitor from lignin) for 4 weeks showed less renaldysfunction and oxidative stress (antioxidant enzymes, gluta-thione, SOD and catalase, lipid peroxidation) than controls(Anjaneyulu and Chopra, 2004).
Lignans are dietary phyto-oestrogens that are present ina wide variety of plant foods including flax seeds and wholegrains (corn, oats, wheat and rye). The group includes secoiso-lariciresinol, matairesinol, lariciresinol, pinoresinol and syrin-garesinol. They all have a polyphenolic structure and mayhave antioxidant effects. Lignans and their metabolites, themammalian lignans enterodiol and enterolactone, have antiox-idant activity in different lipid and aqueous in vitro model sys-tems and decrease lipid oxidation (Kitts et al., 1999). Lignanshave less marked effects than lignins upon oxidative geneticdamage, as shown in human colon cells incubated with enter-olactone (Pool-Zobel et al., 2000). Moreover, healthy post-menopausal women who ate a low-fat muffin enriched witha lignan complex for 6 weeks showed no change in their serumlipoprotein oxidation resistance or plasma antioxidant capacity(Hallund et al., 2006); while young rats fed flax and lignansecoisolariciresinol diglycoside demonstrated that the lignanmetabolites enterodiol and enterolactone had an antioxidantsparing effect (Yuan et al., 1999).
2.10. Carotenoids
Cereals are a significant source of carotenoids. Maize is thebest source with about 11 mg/kg dry weight; soft wheat has only1.5 mg/kg (Panfili et al., 2004) (Table 1). Wheat germ containsabout 5.5 mg carotenoids/kg dry weight (Panfili et al., 2004).The carotenoids include a þ b-carotenes, b-cryptoxanthin, lu-tein and zeaxanthin: lutein and zeaxanthin are most abundantin maize (6e18 mg/g for zeaxanthin and 4e8 mg/g for lutein)(de Oliveira and Rodriguez-Amaya, 2007), while there appearsto be more lutein in wheat, but the amount varies significantlywith the variety: from 1.2 to 5.8 mg/g dry matter in three varie-ties (Leenhardt et al., 2006), from 0.3 to 1.4 mg/g grain in a studyon 11 soft and hard wheat varieties (Adom et al., 2003), andfrom 0.8 to 1.1 mg/g grain in eight soft wheat varieties (Mooreet al., 2005). Adom et al. (2003) also reported the zeaxanthincontents of 11 wheat varieties from around 8 to 27 mg/g grain,and b-cryptoxanthin contents from around 1.0 to 13.5 mg/ggrain. Carotenoids are more evenly distributed within the grain,with significant quantities within endosperm, in contrast toother micronutrients such as minerals, trace elements and poly-phenols (Konopka et al., 2004). We are aware of no studies that
have investigated the correlation between the in vivo antioxidantcapacity of cereals and their carotenoid content.
In summary, the antioxidant compounds of whole-grain ce-reals are mainly located in the bran and germ fractions, andmost of them are most abundant within the aleurone layer. Re-fined flour products therefore have a much lower antioxidantcapacity than wholemeal cereal products. While whole-graincereals are a significant source of antioxidants, their antioxi-dant potential varies, as discussed below, according to cerealtype, the variety, growing location, the polyphenol contentand the processing.
3. In vitro antioxidant potential of whole-grain cereals andtheir different fractions
Cereal grains have long been thought to be less importantsources of antioxidants than fruit and vegetables (Liu, 2007)although they contain many antioxidants, as discussed above,and are major dietary components worldwide. The most fre-quently used methods to measure the in vitro antioxidant po-tential of cereals and their fractions are the Oxygen RadicalAbsorbance Capacity (ORAC), the 2,20-azinobis 3-ethyl-ben-zothiazoline-6-sulfonic acid (ABTS$þ), the 2,2-diphenyl-1-picrylhydrazyl (DPPH$) and the Ferric Reducing AntioxidantPower (FRAP) assays.
3.1. In vitro antioxidant potential of cereals and cerealproducts
Cereal products have significant antioxidant potentials invitro (Miller et al., 2000; Pellegrini et al., 2006; Wu et al.,2004a). Miller et al. (2000) used the DPPH assay to showthat the average antioxidant activity of cereals and cereal prod-ucts is higher (between 1200 and 3500 mmol Trolox Equiva-lents (TE)/100 g of fresh product) than that of common fruit(mean: 1200 mmol TE/100 g) and vegetables (mean:400 mmol TE/100 g), but lower than that of common berries(around 3880 mmol TE/100 g). The antioxidant capacity ofthe wheat bran fraction was 8500 mmol TE/100 g and that ofthe germ fraction was 5000 mmol TE/100 g (Miller et al.,2000), while that of rice bran was estimated to reach24,300 mmol TE/100 g (Wu et al., 2004b). Wu et al. (2004a)reported similar results using the ORAC assay: the total anti-oxidant capacities (including both lipophilic and hydrophilicfractions of food) of bread, ready-to-eat breakfast cerealsand uncooked breakfast cereals were between 1303 and2479 mmol TE/100 g fresh product, while that of fruit(n ¼ 31 products, berries not included) was 2200 mmol TE/100 g and that of vegetables (same as those used by Milleret al., 2000) was 1200 mmol TE/100 g. Therefore, the antiox-idant activity of cereal products is far from negligible. How-ever, on a dry matter basis, the antioxidant capacity of manyfruits and vegetable is generally higher than that of cerealproducts (Saura-Calixto and Goni, 2006).
From a nutritional point of view, it is particularly relevantto compare daily antioxidant consumption. When this is

264 A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
done, the differences between cereals, fruits and vegetablesbecome less marked. For example, based on the USDA Con-tinuing Survey of Food Intake by Individuals (1994e1996),daily antioxidant consumption per capita was 480 mmol TEfor ready-to-eat breakfast cereals, 600 mmol TE for vegetablesand 760 mmol TE for fruits (Miller et al., 2000). Similarly,application of the FRAP assay to foods of the Spanish dietshowed that daily antioxidant consumption per capita was367 mmol TE for cereals, 418 mmol TE for vegetables and850 mmol TE for fruits (Saura-Calixto and Goni, 2006).
The in vitro antioxidant capacities of cereals (wheat, oats,barley and rye) are significantly correlated with their phenolicacid contents, unlike those of fruit and vegetables. This is es-pecially true for ferulic acid, the major phenolic compound ingrains (the ratio between the free, soluble conjugate and boundforms is 0.1:1:100), except for maize (Adom and Liu, 2002).The bran and germ fractions are the main contributors to theantioxidant potential, but both are generally released duringmilling. Coloured varieties of cereals, such as rice, havemore antioxidant capacity than non-coloured varieties (Huet al., 2003). Zielinski and Kozlowska (2000) reported thatthe antioxidant potentials of methanolic extracts of cerealswere: barley > oat > wheat and rye. The correlation coeffi-cient between total phenolic compounds and antioxidant activ-ity is 0.96 for the whole grain, 0.99 for the external layers,0.80 for the pericarp and testa layers, and 0.99 for the endo-sperm with the embryo (Zielinski and Kozlowska, 2000).Methanolic extracts of grains consumed in Korea (red sor-ghum and black rice) were also shown to have higher antiox-idant activities than white rice, brown rice, foxtail millet,proso millet and barley. Again, the polyphenolic content pos-itively correlated (r > 0.99) with the ABTS radical cationscavenging activity of the extracts (Choi et al., 2007). Anotherstudy reported that maize had the highest in vitro antioxidantpotential (181 mmol vit. C eq/g grain), followed by wheat(77 mmol vit. C eq/g grain), oats (75 mmol vit. C eq/g grain),and rice (56 mmol vit. C eq/g grain) (Adom and Liu, 2002).The differences in the rankings of the cereals according totheir antioxidant potential probably reflect the differentmethods used, but may also depend on the cereal varieties.
However, these studies were concerned only with aqueous-organic extracts of foods whereas the antioxidant capacity inthe residues (bound antioxidant fraction) can be significant(Pellegrini et al., 2006), particularly when considering poly-phenols (Saura-Calixto et al., 2007). Thus, based on three dif-ferent antioxidant assays, Pellegrini et al. (2006) showed thatbound fractions of 18 cereal products had antioxidant capac-ities higher in all cases than those of the free extracts (solubleextracts in organic solvent). In addition, Liyana-Pathirana andShahidi (2006b) demonstrated that the bound phenolic frac-tions of two commercial samples of soft and hard wheat hadsignificantly higher antioxidant capacities than the free and es-terified phenolic fractions. The antioxidant capacity of whole-grain cereals is therefore usually underestimated in vitro,mainly because fibre-associated compounds are generally notincluded. Indeed, the major antioxidant polyphenols are boundpolyphenols, which are 90% of the total in wheat, 87% in
maize, 71% in rice and 58% in barley (Adom and Liu, 2002).Consequently, bound phytomicronutrients are likely to survivedigestion in the stomach and intestine and reach the colon,where they may produce a favourable antioxidant environment(see Section 4.2).
Finally, it appears that water extracts of grain fractions (suchas wheat germ/middlings/bran, fermented wheat bran, rye bran/middlings, buckwheat flour) are of greatest relevance to in vivoactivity: they contain water soluble antioxidants (10e30% oftotal phenolic compounds), that are most readily availablefrom the food matrix within the digestive tract, and thereforeare more likely to have a physiological effect in vivo (Gallardoet al., 2006), while bound phenolics are probably released laterin the colon during fermentation (see Section 4.2).
These results suggest that the antioxidant potential of ce-reals depends on their type, their degree of fractionation andthe extraction medium used. The following section discussesthe reported antioxidant potential of several types of cereals.
3.2. Wheat
The antioxidant activity of wheat and its different fractionsusually depends on the variety examined (Yu et al., 2002a,b),and the area where it is grown (Yu and Zhou, 2004; Yu et al.,2003; Zhou and Yu, 2004). Environmental factors (such astemperature stress, solar radiation and irrigation), and theinteractions between environmental factors and genotype canmodulate the antioxidant activity of wheat (Moore et al.,2006; Yu and Zhou, 2004; Yu et al., 2003; Zhou and Yu,2004).
Ferulic, vanillic and p-coumaric acids are the most abun-dant free phenolic acids in wheat. A study of seven wheat va-rieties showed that ferulic acid is generally the predominantphenolic acid (99e231 mg/g), accounting for about 46e67%of total phenolic acids (Zhou et al., 2005). Ferulic acid isthe main contributor to the antioxidant capacity, suggestingthat ferulic acid could be used as a marker of wheat antioxi-dants (Zhou et al., 2004). The grain fractions (whole-grain,bran and flour) have different antioxidant capacities, withthe wheat bran aleurone layer having an antioxidant capacitycomparable to that of tea and dried raisins (Liyana-Pathiranaand Shahidi, 2007). The aleurone layer is therefore the fractionwith the highest antioxidant activity, followed by the branfraction and whole grain. This is why industrial processeshave been developed to isolate the aleurone layer, and thento enrich cereal products with it (Hemery et al., 2007). How-ever, it may also be interesting to preserve the whole bran frac-tion together with the aleurone layer. The pericarp and testalayers may indeed provide not only insoluble fibre but alsolignin for which in vitro antioxidant potential is now wellrecognized and whose nutritional value remains largelyunknown.
3.3. Corn
Corn is one of the most important cereal grains in theworld. It is consumed in a variety of forms, from sweet

265A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
corn, to nixtamal (cooked kernels), tortillas, chips and polenta.Like other cereals, it is a great source of antioxidants, espe-cially ferulic acid, located in the bran. Ferulic acid may ac-count for 2e4% by dry weight of corn hulls (Hosny andRosazza, 1997). Corn bran may thus contain up to 26e33 gferulic acid/kg product, which is higher than wheat bran(5.3e5.4 g/kg), rye bran (2.8 g/kg), whole wheat (1.3 g/kg),rice endosperm cell wall (9.1 g/kg), and brown rice (0.4 g/kg) (Zhao et al., 2005). Corn technological processes andcorn varieties are also factors that have a significant impacton the antioxidant capacity. Processed sweet corn has a higherantioxidant activity than its raw counterpart, despite the loss ofsome vitamin C (Dewanto et al., 2002), which raises questionsabout the view that processing plant products always leads toa significant loss of their antioxidant potential. High-amylosecorn also has greater antioxidant potential than typical andwaxy genotypes, although the reason remains unclear (Liet al., 2007). The Mexican blue corn genotype has a slightlyhigher antioxidant capacity than the American blue and whitecorn genotypes, which is attributed to its specific anthocyaninsand/or polyphenolic composition (Del Pozo-Insfran et al.,2006). The problem lies in the fact that the phenolic antioxi-dants in corn bran are not readily bioavailable. This was dem-onstrated in a recent study on rats: only 0.4e2.3% of theferulic and p-coumaric acids ingested (via corn bran) wererecovered in the urine, while 64e81% of the phenolic acidswere recovered in the faeces (Zhao et al., 2005).
3.4. Rice
Rice contains potentially antioxidant compounds, notablyin the outer layers of the grain (bran). Significant quantitiesof vitamin E and g-oryzanol can be extracted from rice. Sincethe g-oryzanol content of rice bran is 10-times that of vitaminE, g-oryzanol may contribute more to the reduction of choles-terol oxidation than vitamin E, which is usually considered tobe the major antioxidant in rice bran (Xu et al., 2001). Severalvarieties of rice, including coloured varieties, are likely tohave a greater antioxidant capacity than white rice. For exam-ple, cyanidin-3-glucoside and peonidin 3-glucoside (anthocya-nin pigments) are the major antioxidant compounds in blackrice (Hu et al., 2003), but they are not found in white rice. Pro-cyanidins are the major compounds involved in the antioxidantactivity of red rice (Oki et al., 2002). Pigments suppressed theoxidative changes in human LDL, reduced the formation ofnitric oxide by suppressing inducible nitric oxide synthase ac-tivity in murine macrophage cells, and significantly preventedthe breaks in supercoiled DNA strands induced by ROS (Huet al., 2003). In the end, b-carotene-rich rice varieties (e.g.the yellow Golden Rice) have also been recently developedby genetic engineering, in order to help combat vitamin Adeficiency, notably in Asian countries (Krawinkel, 2007).However, it remains to be determined how the antioxidantproperties of b-carotene in such rice varieties may participatein an improved health. As for wheat, the antiradical efficiencyof rice bran extracts is positively correlated (r ¼ 0.99) withtheir total phenolic content (Goffman and Bergman, 2004).
3.5. Oats
The worldwide consumption of oats is much lower than thatof wheat. However, it is usually consumed as a whole-graincereal. The antioxidant capacity of oats has been discussed byPeterson (2001). Oats, like wheat, contains large amounts ofantioxidant phytomicronutrients, which are also concentratedwithin the outer layer of the grain; these are mainly vitamin E(tocols), phytic acid, phenolic compounds, and avenanthra-mides (alkaloids containing phenolic groups). Avenanthra-mides are typically found in oats and have antioxidant activityin vitro (Peterson et al., 2002) and in vivo (Chen et al., 2007).Avenanthramide is a more powerful antioxidant than some ofthe typical cereal components such as ferulic acid, p-hydroxy-benzoic acid, vanillic acid, and phytic acid (Martinez-Tomeet al., 2004). a-Tocotrienol is predominant in oats and about90% of tocols are a-tocotrienol and a-tocopherol. The tocoph-erols are located in the germ and tocotrienols in the endosperm.The ferulic acid content may reach 30 mg/100 g (Table 1). Oatscontains 8.7 mg/kg free phenolic acids, 20.6 mg/kg solublephenolic acids and 57 mg/kg insoluble phenolic acids. Again,the total phenolic acid content (mainly avenanthramides, vanil-lic and p-hydroxybenzoic acids) was significantly correlatedwith antioxidant activity in vitro (Emmons et al., 1999). A com-parative study also showed that groats had a significantly higherantioxidant activity than did hulls. This is related to the presenceof more avenanthramides and caffeic acid in the groats while thehulls have higher concentrations of many other phenolics (Em-mons and Peterson, 1999). Oat bran and several bran products(crunchy oat bran and oat breakfast cereals) generally havea lower antioxidant activity than wheat bran and derived prod-ucts (bran breakfast cereal, tablet of bran, wheat bran powder)(Martinez-Tome et al., 2004).
3.6. Barley
Barley is widely consumed, but mainly (80e90%) by ani-mals. As for oats and wheat, there is growing interest in barleyproducts because of their high content of antioxidants such asbenzoic and cinnamic acid derivatives, proanthocyanidins,quinines, flavonols, chalcones, flavones, flavanones, andamino phenolic compounds (Hernanz et al., 2001). Barley istherefore an excellent source of natural antioxidants eitherfor food preservation (to inhibit lipid oxidation), or for diseaseprevention (Liu and Yao, 2007). Ferulic acid is the mostabundant phenolic acid in barley, with a concentration of359e624 mg/g dry weight, as measured in 11 barley varieties(Hernanz et al., 2001) (Table 1). The antioxidant capacity ofbarley and barley extracts has been particularly studied inthe field of beer production, since 80% of the phenolics inbeer are derived from barley malt, and are responsible forthe overall stability of beer (Maillard et al., 1996). Unlikewheat and oat, there has been little research on the antioxidantpotential of barley and its fractions. However, the total pheno-lic content of barley is, like that of other cereals, significantlycorrelated with the antioxidant capacity, as measured by theDPPH and ABTS assays (Zhao et al., 2006).

266 A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
3.7. Rye
Rye is typically consumed in Scandinavian countries, espe-cially as whole-grain rye bread. Ferulic acid is the predomi-nant hydroxycinnamic acid, and the concentrations of itsfree and bound forms range from 900 to 1170 mg/g dry matter(Andreasen et al., 2000) (Table 1). The other most abundantphenolic acids in rye are sinapic acid and the dimer 8-O-4-di ferulic acid. These compounds all inhibit LDL oxidationin vitro (Andreasen et al., 2001). The antioxidant activity ofrye extracts (whole-grain, bran and flour) is significantly cor-related with the total content of monomeric and dimerichydroxycinnamic acids, with rye bran having the greatestantioxidant capacity (Andreasen et al., 2001).
3.8. Sorghum
Sorghum is the fifth the most consumed cereal in the world,although human consumption remains limited. It is mainly con-sumed in hot, dry tropical countries (semi-arid regions). Sor-ghum is rich in polyphenols and tannins. There have been fewstudies of its antioxidant capacity (Dykes and Rooney, 2006).The antioxidant potential of sorghum-based products (whole-grain, bran and cooked/extruded products) is strongly corre-lated with their polyphenol content (Awika et al., 2003), withsorghum bran having the highest antioxidant potential (about1000e3000 mmol TE/g dried product). However, another studyof different extracts of white sorghum flour (with water, 60%methanol, 60% ethanol and 60% t-butanol) found that the phe-nolic content was not correlated with the antioxidant capacity(Kamath et al., 2004). Sorghums grains that had a purple/redsecondary colour, a black or dark-red thick pericarp, and a pig-mented testa had greater antioxidant activity. Most of the phe-nolic acids are esterified to cell wall compounds (85% of totalphenolic acids). The anthocyanin content of black sorghum iscorrelated with antioxidant capacity, particularly with that ofthe bran. The anthocyanin content of sorghum bran is three-to four-fold higher than that of the whole grain. Sorghum grainsand bran were also shown to have a greater antioxidant capacity(52e400 mmol TE/g product, ABTS assay) than other cereals(<0.1e34 mmol TE/g product) (Awika et al., 2005). A dietrich in sorghum may therefore be useful for preventing diseasesin which free radicals play a key role.
3.9. Finger millet
Finger millet is mainly consumed in East Africa, Indiaand Sri Lanka after being malted. It is rich in phenolic acidsand therefore has potential antioxidant activity (Dykes andRooney, 2006). The antioxidant capacity of the fraction con-taining free phenolic acids increased (two-fold) after 96 h ofmalting of finger millet, whereas the antioxidant capacity ofthe fraction containing bound phenolic acids decreased(Subba Rao and Muralikrishna, 2002). Finger millet has apotent radical-scavenging activity that is higher than thoseof wheat, rice, and other species of millet (Dykes andRooney, 2006).
The in vitro antioxidant potential of cereals is thereforequite significant. The questions are whether it can be extrapo-lated to the in vivo situation and whether it can explain the pro-tective role of whole-grain cereals. Before discussing animaland human studies using cereals and cereal products, wewill examine ex vivo studies and the influence of digestionon the antioxidant potential of cereals.
4. In vivo antioxidant potential of whole-grain cereals andtheir fractions
4.1. Ex vivo oxidation of LDL
Several studies have measured the susceptibility of LDL tooxidation ex vivo. The LDLs were collected from human(Chen et al., 2004; Handelman et al., 1999; Harder et al.,2004; Hu et al., 2003; Yu et al., 2005), or animal (Chenet al., 2004) plasma. Harder et al. (2004) observed no effectof rye bran consumption by postmenopausal women on subse-quent ex vivo plasma LDL oxidation (CuSO4-induced) in com-parison with their usual diet or with consumption of inertwheat cellulose fibre. The authors suggested that water-solubleantioxidants, including ferulic acid, are separated from LDLfractions during the ultracentrifugation of blood samples, andthat the plasma concentration of ferulic acid was too low tohave any significant effect. Whether the concentration offerulic acid in plasma can become high enough to have invivo effects against LDL oxidation in the long term thus re-mains to be demonstrated. Chen et al. (2004) reported thatno effect was observed on ex vivo resistance of LDL toCu2þ-induced oxidation following oat bran consumption byhamsters, but that the direct addition of oat phenolics inhibitedin vitro Cu2þ-induced human plasma LDL oxidation and thatthis inhibition was dose dependent. Similarly, Handelmanet al. (1999) have shown that oat extracts (milled oat groatpearlings) were able to inhibit ex vivo human LDL oxidationin a dose-dependent manner. Reduced or inhibited oxidationof LDL was also obtained with wheat bran (Yu et al., 2005),rye (Andreasen et al., 2001), black rice pigmented fraction(Hu et al., 2003) and oat bran (Gray et al., 2002) extracts onhuman LDL oxidation. Thus, bran and whole-grain can pro-vide antioxidants that may affect health, via the action of phe-nolic acids on LDL oxidation. The protection against oxidationprovided to LDL by cereal extracts can be extended to lipidspresent in tissues. Parrado et al. (2003) showed that an extractof rice bran fraction (water soluble oryzanol enzymatic extract)protected the proteins and lipids in rat brain homogenates fromoxidation.
Since the objective of in vitro assays is to predict the bio-logical efficiency of antioxidants, a method is appropriateonly if the results correlate well with the in vivo antioxidantcapacity. This may not be the case, as was shown with applepolyphenols in humans (Lotito and Frei, 2004b). The authorsmeasured the in vitro antioxidant potential of one apple extracton human plasma, and then the ex vivo induced oxidation ofplasma of subjects that had consumed five apples. While appleextract protected plasma compounds (i.e. urate, a-tocopherol

267A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
and lipids) from oxidation in vitro, there was no significant dif-ference in the antioxidative protection of plasma ex vivo afterapple consumption by healthy individuals, suggesting that thebioavailability and metabolization of polyphenols from appleextracts modified their initial antioxidant properties (Lotitoand Frei, 2004b). In another study, the same authors demon-strated that the increase in plasma antioxidant capacity follow-ing apple consumption by healthy subjects was due to themetabolic effect of apple fructose on plasma uric acid concen-tration, and was not due to the antioxidant effect of apple poly-phenols (Lotito and Frei, 2004a).
As for apple polyphenols, from cereals do not seem to exerttheir antioxidant capacity in the same way being added di-rectly to plasma or after being consumed, as was shown inhamster with oat extracts (see above the study of Chenet al., 2004). Indeed, most of ex vivo studies used cerealextracts added directly to the reaction medium. However, ce-real antioxidants are metabolized before they can act on theoxidation of LDL in the plasma. Furthermore, their bioavail-ability is often limited, especially that of phenolic acids, lead-ing to very low concentrations that may not be high enough tohave a significant antioxidant effect (Kern et al., 2003).
4.2. Bioavailability and influence of digestion on theantioxidant capacity of cereal products
Most of the antioxidant phytomicronutrients in grain arebound and can survive gastrointestinal digestion to reach thecolon intact, where they provide an antioxidant environment(Perez-Jimenez and Saura-Calixto, 2005). This might partlyexplain why whole-grain consumption helps prevent coloncancer, as demonstrated by epidemiological studies (Adomand Liu, 2002). Otherwise, digestion increases the antioxidantcapacity of cereal products (Liyana-Pathirana and Shahidi,2005; Nagah and Seal, 2005; Perez-Jimenez and Saura-Cal-ixto, 2005). The acidic conditions in the stomach and enzy-matic hydrolysis in the duodenum are likely to increase thesolubility and activity of cereal polyphenols (Perez-Jimenezand Saura-Calixto, 2005). Digestion therefore appears to bean important factor in potentiating the antioxidant capacityof high-bran, whole-grain and refined wheat cereal products(Baublis et al., 2000). Thus, digestion in vitro with enzymaticextracts mimicking conditions in the gastrointestinal tractshowed that the amount of antioxidants released by the cerealmatrix into the human intestine may be higher than that ex-pected from measurements made on the usual aqueous-organicextracts (Perez-Jimenez and Saura-Calixto, 2005). Nagah andSeal (2005) also demonstrated the significant influence of invitro gastrointestinal digestion on the increased antioxidant re-lease from whole-grain cereal foods. Digestion is believed topartially hydrolyse the hydrolysable phenolics. Similarly thedigestion of starch and protein may also increase the releaseof polyphenols. These results are particularly important sincethey suggest that antioxidants exert their effect within the di-gestive tract, providing an environment that protects the intes-tinal epithelium from pro-oxidative compounds. Otherimportant fractions can also be released during fermentation.
Further studies are therefore needed to evaluate the effectsof released antioxidants on the prevention of disorders suchas stomach, colon and rectal cancers. Antioxidants may be an-ticarcinogenic because they stimulate detoxification systems,especially phase II of the conjugation reaction. For example,Wattenberg (1996) classified caffeic and ferulic acids asanti-cancer agents that prevent the formation of carcinogensand block their interaction with critical cell macromolecules.We may therefore ask which part of the gastrointestinal tract,the small intestine or colon, is the major site of antioxidantaction (Halliwell et al., 2000). As the antioxidant polyphenolsof whole-grains are mostly in a bound form, they are morelikely to exert their antioxidant effects in the colon, unlikefruits and vegetables, whose antioxidant polyphenols aremainly free or soluble conjugates that are more available inthe small intestine.
4.3. Animal studies
Few studies are available on the relationship between whole-grain and refined cereal consumption, and in vivo oxidativestress biomarkers (Table 2). Studies were reported on hypercho-lesterolaemic rabbits fed white, red and black varieties of rice,cereals that are rich in anthocyanin, which are potential antiox-idants (Ling et al., 2001, 2002). The liver reactive oxygen spe-cies and aortic MDA were significantly decreased, while thetotal antioxidant capacity of the liver and erythrocyte superoxidedismutase were significantly increased in the rabbits fed col-oured rice, compared to those fed white rice (Ling et al.,2001). This effect was attributed to the outer layers of the col-oured rice. In another study, Ling et al. (2002) tested the antiox-idant capacity of the outer layer fraction of black rice. Theyobserved significant effects on several biomarkers for oxidativestress: decreased serum and aorta MDA and decreased aortic8-OHdG. Xia et al. (2003) tested the effects of an outer layerfraction from black rice on biomarkers for atherosclerosis risk.They showed a significant decrease in oxidized LDL in theserum of ApoE-deficient mice, an established model for athero-sclerosis. Similarly, Kim et al. (2006) showed that a mixture ofbrown and black rice was more effective in reducing liverTBARS and increasing liver glutathione and glutathione-perox-idase than was white rice alone. Cyanidin-3-O-glucoside chlo-ride and peonidin-3-O-glucoside chloride may be responsiblefor the antioxidant effects of black rice extract. Chiang et al.(2006) showed increases in hepatic SOD and catalase activities,and a decrease in plasma TBARS in mice fed black rice extract.The effect of the outer layer fraction of black rice was confirmedin a recent study that showed that plasma TBARS and oxidizedglutathione were significantly reduced in oxidatively stressed(high-fructose diet) rats fed an anthocyanin-rich extract (fromblack rice) (Guo et al., 2007). Otherwise we have shown thatrats fed whole-grain wheat flour or refined wheat flour for2 weeks had similar plasma redox states (vitamins E and C,MDA and FRAP) and urine antioxidant status (MDA andisoprostanes) (Fardet et al., 2007). However, we used a metabo-nomic approach (H NMR-based) to show significant differencesin the redox states of the liver of these rats, with increased in
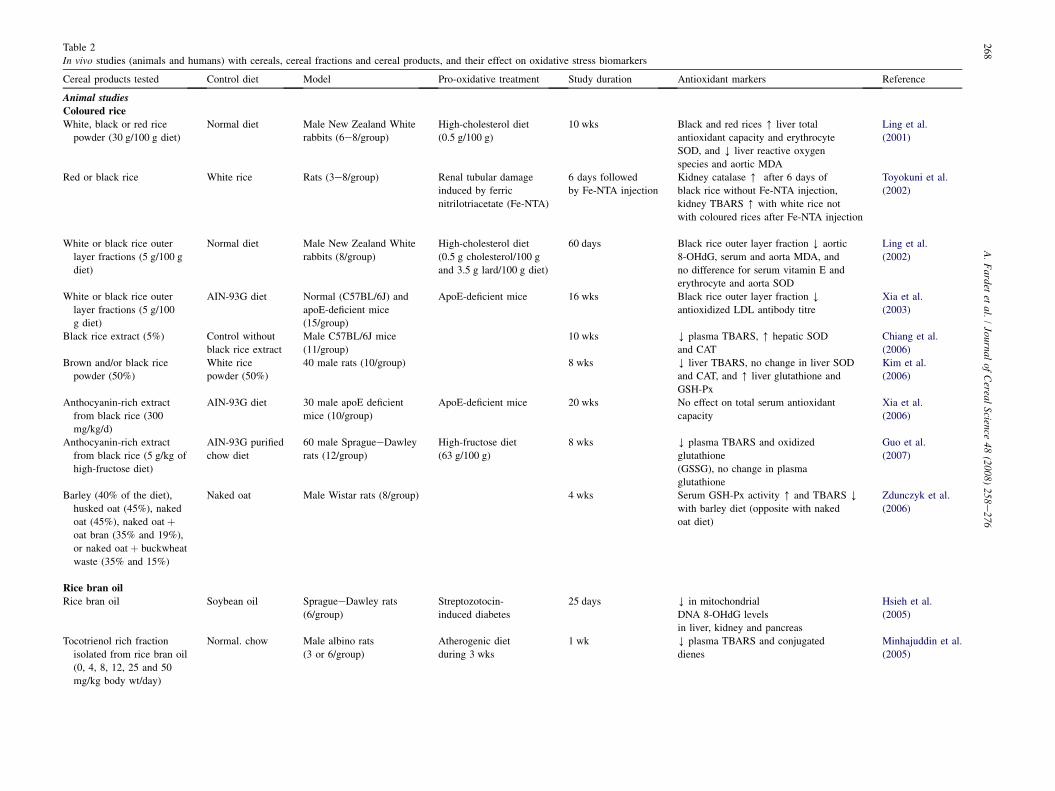
Table 2
In vivo studies (animals and humans) with cereals, cereal fractions and cereal products, and their effect on oxidative stress biomarkers
Cereal products tested Control diet Model Pro-oxidative treatment Study duration Antioxidant markers Reference
Animal studies
Coloured rice
White, black or red rice
powder (30 g/100 g diet)
Normal diet Male New Zealand White
rabbits (6e8/group)
High-cholesterol diet
(0.5 g/100 g)
10 wks Black and red rices [ liver total
antioxidant capacity and erythrocyte
SOD, and Y liver reactive oxygen
species and aortic MDA
Ling et al.
(2001)
Red or black rice White rice Rats (3e8/group) Renal tubular damage
induced by ferric
nitrilotriacetate (Fe-NTA)
6 days followed
by Fe-NTA injection
Kidney catalase [ after 6 days of
black rice without Fe-NTA injection,
kidney TBARS [ with white rice not
with coloured rices after Fe-NTA injection
Toyokuni et al.
(2002)
White or black rice outer
layer fractions (5 g/100 g
diet)
Normal diet Male New Zealand White
rabbits (8/group)
High-cholesterol diet
(0.5 g cholesterol/100 g
and 3.5 g lard/100 g diet)
60 days Black rice outer layer fraction Y aortic
8-OHdG, serum and aorta MDA, and
no difference for serum vitamin E and
erythrocyte and aorta SOD
Ling et al.
(2002)
White or black rice outer
layer fractions (5 g/100
g diet)
AIN-93G diet Normal (C57BL/6J) and
apoE-deficient mice
(15/group)
ApoE-deficient mice 16 wks Black rice outer layer fraction Yantioxidized LDL antibody titre
Xia et al.
(2003)
Black rice extract (5%) Control without
black rice extract
Male C57BL/6J mice
(11/group)
10 wks Y plasma TBARS, [ hepatic SOD
and CAT
Chiang et al.
(2006)
Brown and/or black rice
powder (50%)
White rice
powder (50%)
40 male rats (10/group) 8 wks Y liver TBARS, no change in liver SOD
and CAT, and [ liver glutathione and
GSH-Px
Kim et al.
(2006)
Anthocyanin-rich extract
from black rice (300
mg/kg/d)
AIN-93G diet 30 male apoE deficient
mice (10/group)
ApoE-deficient mice 20 wks No effect on total serum antioxidant
capacity
Xia et al.
(2006)
Anthocyanin-rich extract
from black rice (5 g/kg of
high-fructose diet)
AIN-93G purified
chow diet
60 male SpragueeDawley
rats (12/group)
High-fructose diet
(63 g/100 g)
8 wks Y plasma TBARS and oxidized
glutathione
(GSSG), no change in plasma
glutathione
Guo et al.
(2007)
Barley (40% of the diet),
husked oat (45%), naked
oat (45%), naked oat þoat bran (35% and 19%),
or naked oat þ buckwheat
waste (35% and 15%)
Naked oat Male Wistar rats (8/group) 4 wks Serum GSH-Px activity [ and TBARS Ywith barley diet (opposite with naked
oat diet)
Zdunczyk et al.
(2006)
Rice bran oil
Rice bran oil Soybean oil SpragueeDawley rats
(6/group)
Streptozotocin-
induced diabetes
25 days Y in mitochondrial
DNA 8-OHdG levels
in liver, kidney and pancreas
Hsieh et al.
(2005)
Tocotrienol rich fraction
isolated from rice bran oil
(0, 4, 8, 12, 25 and 50
mg/kg body wt/day)
Normal. chow Male albino rats
(3 or 6/group)
Atherogenic diet
during 3 wks
1 wk Y plasma TBARS and conjugated
dienes
Minhajuddin et al.
(2005)
26
8A
.F
ardetet
al./
Journalof
Cereal
Science48
(2008)258e
276
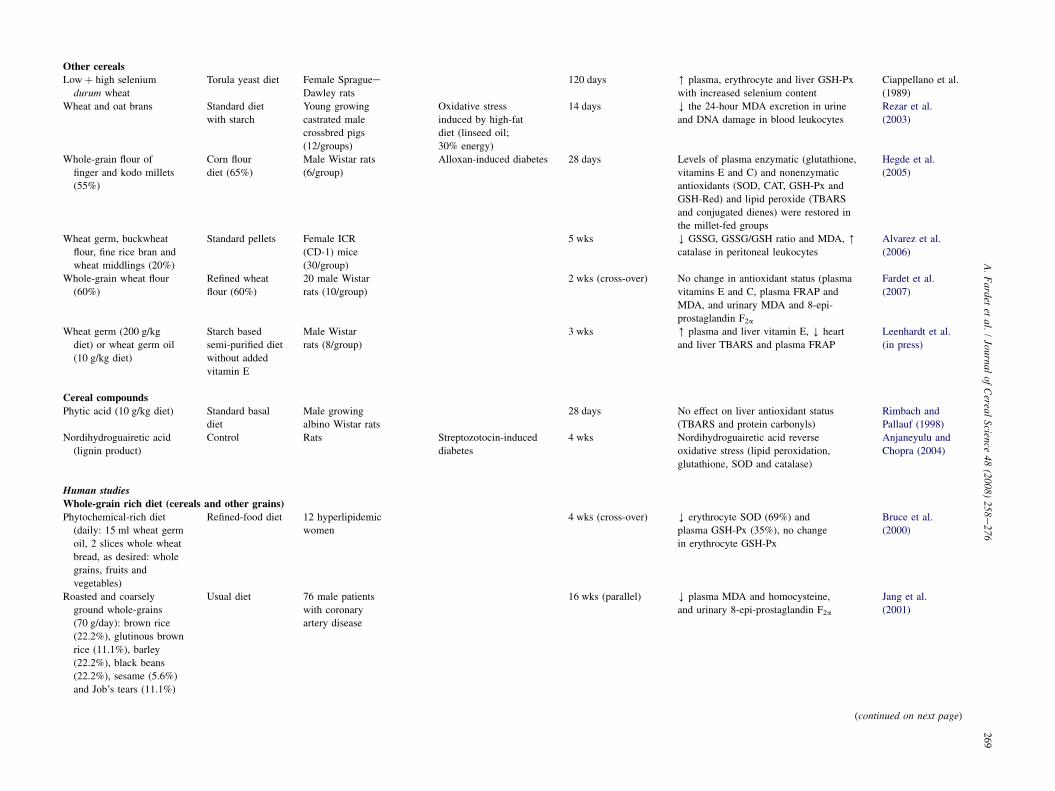
Other cereals
Low þ high selenium
durum wheat
Torula yeast diet Female Spraguee
Dawley rats
120 days [ plasma, erythrocyte and liver GSH-Px
with increased selenium content
Ciappellano et al.
(1989)
Wheat and oat brans Standard diet
with starch
Young growing
castrated male
crossbred pigs
(12/groups)
Oxidative stress
induced by high-fat
diet (linseed oil;
30% energy)
14 days Y the 24-hour MDA excretion in urine
and DNA damage in blood leukocytes
Rezar et al.
(2003)
Whole-grain flour of
finger and kodo millets
(55%)
Corn flour
diet (65%)
Male Wistar rats
(6/group)
Alloxan-induced diabetes 28 days Levels of plasma enzymatic (glutathione,
vitamins E and C) and nonenzymatic
antioxidants (SOD, CAT, GSH-Px and
GSH-Red) and lipid peroxide (TBARS
and conjugated dienes) were restored in
the millet-fed groups
Hegde et al.
(2005)
Wheat germ, buckwheat
flour, fine rice bran and
wheat middlings (20%)
Standard pellets Female ICR
(CD-1) mice
(30/group)
5 wks Y GSSG, GSSG/GSH ratio and MDA, [catalase in peritoneal leukocytes
Alvarez et al.
(2006)
Whole-grain wheat flour
(60%)
Refined wheat
flour (60%)
20 male Wistar
rats (10/group)
2 wks (cross-over) No change in antioxidant status (plasma
vitamins E and C, plasma FRAP and
MDA, and urinary MDA and 8-epi-
prostaglandin F2a
Fardet et al.
(2007)
Wheat germ (200 g/kg
diet) or wheat germ oil
(10 g/kg diet)
Starch based
semi-purified diet
without added
vitamin E
Male Wistar
rats (8/group)
3 wks [ plasma and liver vitamin E, Y heart
and liver TBARS and plasma FRAP
Leenhardt et al.
(in press)
Cereal compounds
Phytic acid (10 g/kg diet) Standard basal
diet
Male growing
albino Wistar rats
28 days No effect on liver antioxidant status
(TBARS and protein carbonyls)
Rimbach and
Pallauf (1998)
Nordihydroguairetic acid
(lignin product)
Control Rats Streptozotocin-induced
diabetes
4 wks Nordihydroguairetic acid reverse
oxidative stress (lipid peroxidation,
glutathione, SOD and catalase)
Anjaneyulu and
Chopra (2004)
Human studies
Whole-grain rich diet (cereals and other grains)
Phytochemical-rich diet
(daily: 15 ml wheat germ
oil, 2 slices whole wheat
bread, as desired: whole
grains, fruits and
vegetables)
Refined-food diet 12 hyperlipidemic
women
4 wks (cross-over) Y erythrocyte SOD (69%) and
plasma GSH-Px (35%), no change
in erythrocyte GSH-Px
Bruce et al.
(2000)
Roasted and coarsely
ground whole-grains
(70 g/day): brown rice
(22.2%), glutinous brown
rice (11.1%), barley
(22.2%), black beans
(22.2%), sesame (5.6%)
and Job’s tears (11.1%)
Usual diet 76 male patients
with coronary
artery disease
16 wks (parallel) Y plasma MDA and homocysteine,
and urinary 8-epi-prostaglandin F2a
Jang et al.
(2001)
(continued on next page)
26
9A
.F
ardetet
al./
Journalof
Cereal
Science48
(2008)258e
276
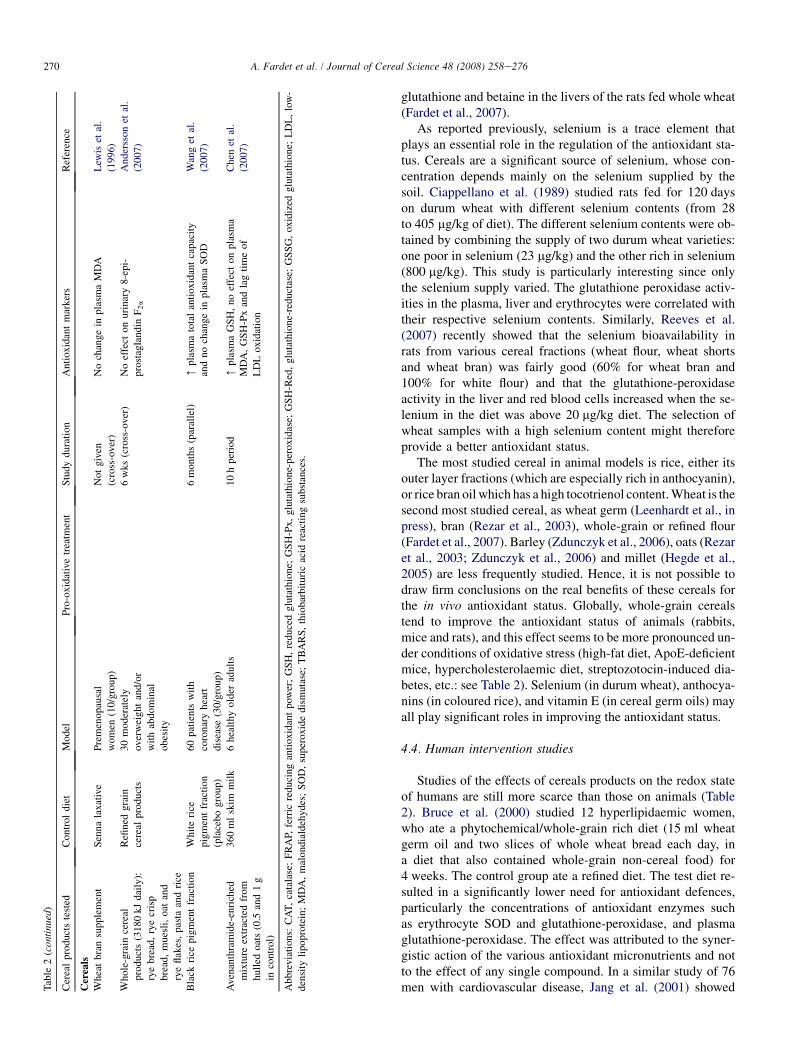
Tab
le2
(con
tinu
ed)
Cer
eal
pro
duc
tste
sted
Co
ntr
old
iet
Mo
del
Pro
-ox
idat
ive
trea
tmen
tS
tud
yd
ura
tio
nA
nti
oxi
dan
tm
ark
ers
Ref
eren
ce
Cer
eals
Wh
eat
bra
nsu
ppl
emen
tS
enn
ala
xat
ive
Pre
men
opa
usa
l
wo
men
(10
/gro
up)
No
tg
iven
(cro
ss-o
ver)
No
chan
ge
inp
lasm
aM
DA
Lew
iset
al.
(19
96)
Wh
ole
-gra
ince
real
pro
duc
ts(3
18
0k
Jd
aily
):
rye
bre
ad,
rye
cris
p
bre
ad,
mu
esli
,o
atan
d
rye
flak
es,
pas
taan
dri
ce
Refi
ned
gra
in
cere
alp
rod
uct
s
30
mo
der
atel
y
over
wei
gh
tan
d/o
r
wit
hab
do
min
al
ob
esit
y
6w
ks
(cro
ss-o
ver)
No
effe
cto
nu
rin
ary
8-e
pi-
pro
stag
lan
din
F2
a
An
der
sson
etal
.
(20
07)
Bla
ckri
cep
igm
ent
frac
tio
nW
hit
eri
ce
pig
men
tfr
acti
on
(pla
ceb
og
rou
p)
60
pat
ien
tsw
ith
coro
nar
yh
eart
dis
ease
(30
/gro
up
)
6m
onth
s(p
aral
lel)
[pla
sma
tota
lan
tioxid
ant
capac
ity
and
no
chan
ge
inp
lasm
aS
OD
Wan
get
al.
(20
07)
Ave
nan
thra
mid
e-en
rich
ed
mix
ture
extr
acte
dfr
om
hu
lled
oat
s(0
.5an
d1
g
inco
ntr
ol)
36
0m
lsk
imm
ilk
6h
ealt
hy
old
erad
ult
s1
0h
per
iod
[p
lasm
aG
SH
,n
oef
fect
on
pla
sma
MD
A,
GS
H-P
xan
dla
gti
me
of
LD
Lo
xid
atio
n
Ch
enet
al.
(20
07)
Abbre
via
tion
s:C
AT
,ca
tala
se;
FR
AP,
ferr
icre
duci
ngan
tioxid
ant
pow
er;
GS
H,
reduce
dglu
tath
ione;
GS
H-P
x,
glu
tath
ione-
per
oxid
ase;
GS
H-R
ed,
glu
tath
ione-
reduct
ase;
GS
SG
,oxid
ized
glu
tath
ione;
LD
L,
low
-
den
sity
lip
op
rote
in;
MD
A,
mal
on
dial
deh
yde
s;S
OD
,su
per
oxi
de
dis
mut
ase;
TB
AR
S,
thio
bar
bitu
ric
acid
reac
tin
gsu
bst
ance
s.
270 A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
glutathione and betaine in the livers of the rats fed whole wheat(Fardet et al., 2007).
As reported previously, selenium is a trace element thatplays an essential role in the regulation of the antioxidant sta-tus. Cereals are a significant source of selenium, whose con-centration depends mainly on the selenium supplied by thesoil. Ciappellano et al. (1989) studied rats fed for 120 dayson durum wheat with different selenium contents (from 28to 405 mg/kg of diet). The different selenium contents were ob-tained by combining the supply of two durum wheat varieties:one poor in selenium (23 mg/kg) and the other rich in selenium(800 mg/kg). This study is particularly interesting since onlythe selenium supply varied. The glutathione peroxidase activ-ities in the plasma, liver and erythrocytes were correlated withtheir respective selenium contents. Similarly, Reeves et al.(2007) recently showed that the selenium bioavailability inrats from various cereal fractions (wheat flour, wheat shortsand wheat bran) was fairly good (60% for wheat bran and100% for white flour) and that the glutathione-peroxidaseactivity in the liver and red blood cells increased when the se-lenium in the diet was above 20 mg/kg diet. The selection ofwheat samples with a high selenium content might thereforeprovide a better antioxidant status.
The most studied cereal in animal models is rice, either itsouter layer fractions (which are especially rich in anthocyanin),or rice bran oil which has a high tocotrienol content. Wheat is thesecond most studied cereal, as wheat germ (Leenhardt et al., inpress), bran (Rezar et al., 2003), whole-grain or refined flour(Fardet et al., 2007). Barley (Zdunczyk et al., 2006), oats (Rezaret al., 2003; Zdunczyk et al., 2006) and millet (Hegde et al.,2005) are less frequently studied. Hence, it is not possible todraw firm conclusions on the real benefits of these cereals forthe in vivo antioxidant status. Globally, whole-grain cerealstend to improve the antioxidant status of animals (rabbits,mice and rats), and this effect seems to be more pronounced un-der conditions of oxidative stress (high-fat diet, ApoE-deficientmice, hypercholesterolaemic diet, streptozotocin-induced dia-betes, etc.: see Table 2). Selenium (in durum wheat), anthocya-nins (in coloured rice), and vitamin E (in cereal germ oils) mayall play significant roles in improving the antioxidant status.
4.4. Human intervention studies
Studies of the effects of cereals products on the redox stateof humans are still more scarce than those on animals (Table2). Bruce et al. (2000) studied 12 hyperlipidaemic women,who ate a phytochemical/whole-grain rich diet (15 ml wheatgerm oil and two slices of whole wheat bread each day, ina diet that also contained whole-grain non-cereal food) for4 weeks. The control group ate a refined diet. The test diet re-sulted in a significantly lower need for antioxidant defences,particularly the concentrations of antioxidant enzymes suchas erythrocyte SOD and glutathione-peroxidase, and plasmaglutathione-peroxidase. The effect was attributed to the syner-gistic action of the various antioxidant micronutrients and notto the effect of any single compound. In a similar study of 76men with cardiovascular disease, Jang et al. (2001) showed

271A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
that the consumption of a whole-grain-based diet (about 55%cereals) favoured antioxidant protection, with decreases inplasma MDA and urinary 8-epi-PGF2a (lipid peroxidationproducts). The reduction in plasma homocysteine could alsohave contributed to the improvement in the antioxidant status.
Other studies were concerned with cereal products only.Supplementation with wheat bran (20 g/day coarse wheatbran) (Lewis et al., 1996) or whole-grain cereal products (An-dersson et al., 2007) had no effect on lipid peroxidation(plasma MDA or urinary 8-epi-PGF2a) in humans (Table 2).Feeding 60 patients with coronary heart disease with blackrice pigment fraction, as used in animal studies (see above),for 6 months increased their total antioxidant plasma capacity(Wang et al., 2007) (Table 2). Other positive effects on antiox-idant status (i.e. increased plasma glutathione) were observedwhen six healthy adults were fed an avenanthramide-enrichedmixture extracted from hulled oats (Chen et al., 2007).
4.5. Epidemiological studies
Epidemiological studies comparing whole and refined ce-reals showed that the consumption of whole-grain cereals isprotective against several diet- and age-related diseases (met-abolic syndrome, cardiovascular diseases, type 2 diabetes andsome cancers) (Anderson, 2003; Chatenoud et al., 1998, 1999;Esmaillzadeh et al., 2005; Truswell, 2002; Venn and Mann,2004), all of these diseases being associated with increased ox-idative stress. It is therefore generally agreed that the synergis-tic action of phytochemicals is responsible for the protectiveaction of whole-grain cereals (Liu, 2007). Due to the antioxi-dant potential of most of cereal phytochemicals, it is verylikely that whole-grain cereals would be protective throughan improvement of the antioxidant status of various tissues.However, besides the free-radical scavenging capacity ofsome phytochemicals, other unknown mechanisms are proba-bly involved. As suggested above, many more human inter-vention studies are needed to elucidate these mechanisms,and to fill the gap between results of epidemiological studiesand those of in vitro experiments relative to the antioxidantcapacity of phytochemicals and cereals fractions.
4.6. Whole-grain cereals versus fruits and vegetables
In a way similar to whole-grain cereals, fruits and vegeta-bles are generally considered protective against chronic pa-thologies, particularly cardiovascular diseases and epithelialcancers, by improving antioxidant status (Eastwood, 1999;Prior, 2003). However, Hung et al. (2004) showed that an in-crease in fruit and vegetable consumption was only associatedwith a modest, and not statistically significant, reduction in thedevelopment of cardiovascular disease, with no effect on can-cer. Similarly, according to Skibsted et al. (2006), there is noproof that the beneficial effect of fruits and vegetables are as-sociated with an antioxidant effect. When considering humanintervention studies measuring the influence of fruit and veg-etable consumption on antioxidant status (plasma and urine),results differ from one study to another, some studies showing
a significant effect of fruit and vegetable consumption onantioxidant status (Cao et al., 1998; Thompson et al., 2005)while other showing no effect (Briviba et al., 2004; Patersonet al., 2006). As for cereals, further studies are needed to betterunderstand the protective effect of fruits and vegetables. Wetherefore cannot say that, in vivo, whole-grain cereals aremore or less protective and/or more or less antioxidativethan fruits and vegetables. However, based on daily consump-tion, it can be estimated that cereal products would providearound 60e70% of the total potential antioxidant capacitythat would be provided by fruits and vegetables (see Section3.1) (Miller et al., 2000; Saura-Calixto and Goni, 2006).Finally, there is a consensus that increasing the consumptionof small amounts of several antioxidant phytochemicals (asthose found in fruits, vegetables and cereals) is preferable tothe consumption of high amounts of one or two selectedantioxidants.
5. Conclusions and perspectives
In vitro antioxidant potential has been measured on whole-grain cereals, refined cereals, brans, and various cereal prod-ucts. However, the results from these different studies aredifficult to compare since the methods used to evaluateantioxidant capacities are often different and have not beenstandardized. Moreover, the antioxidant capacity of cerealsand cereal products have probably been underestimated, sincethe extraction solvents (un-physiological methods) used inmost published in vitro studies do not completely release allantioxidant compounds: an important fraction of antioxidantmicronutrients (especially insoluble phenolic acids) is stronglybound to the fibre fraction of grain outer layers (Serpen et al.,2007). The in vitro antioxidant potential of cereals is generallycorrelated with their polyphenol content (mainly ferulic acid).Correlations with other types of micronutrients remain to beinvestigated. The phenolic compounds in cereals can be free(6%), soluble (17e30%) or bound (66e80%). However, thephenolics are not the most bioavailable group of antioxidantsin vivo. There is a question as to whether the quantity of poly-phenols tested in vitro accurately reflects the amount found inthe plasma or other tissues, or the quantity really consumed.There is good evidence that this is not so.
It is therefore difficult to predict the in vivo efficiency of anantioxidant or of a cereal product based solely on in vitro mea-surements. Numerous factors may influence the antioxidantaction of the defined compound in vivo, such as digestibility,bioavailability and metabolism (e.g. conjugation for polyphe-nols). Thus, the relationship between the antioxidant capacitymeasured in vitro and the real health benefits of cereals and ce-real products is not really known. A strong in vitro antioxidantpotential does not necessarily imply a strong in vivo antioxi-dant effect, especially if the micronutrient is only slightly bio-available. Conversely, a single micronutrient, such as betainewhich is readily bioavailable, may contribute strongly to theredox status in vivo. Moreover, in vitro data ignore other bio-logical actions that can have antioxidant micronutrients invivo: influencing antioxidant enzymatic activities and/or

272 A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
oxidative-related metabolic pathways, activation/repression ofgenes coding for antioxidant compounds/enzymes, or activat-ing signalling pathway. The in vivo contributions and modes ofaction of antioxidant micronutrients included within a complexcereal matrix remain to be elucidated.
The antioxidant capacities of cereals and cereal productsare quite high, especially those of wheat, corn and sorghum(Miller et al., 2000). On a fresh matter basis, it is even equalor higher than those of some fruits and vegetables (Milleret al., 2000; Wu et al., 2004a). Nevertheless, the antioxidantpotential of cereal products has long been neglected. Severalstudies have been conducted on animals and humans, butthey remain scarce in view of the numerous in vitro studies.This difference between the numbers of in vitro and in vivostudies is paradoxical since most cereal antioxidants are onlypartly bioavailable. Studies on the antioxidant potential of ce-real polyphenols have often been carried out on human plasmato investigate their capacity to delay LDL oxidation ex vivo,and not directly in vivo. Thus, no controlled studies in humanshave compared the antioxidant effects of whole-grain and re-fined cereals. However, the few studies that have been donehave provided interesting and encouraging results that stimu-late further investigations. For example, whole-grain cereals-based products seem to provide an antioxidant protectionthroughout the intestinal tract, but this should be confirmedby in vivo studies. While the antioxidant capacity of cerealsis believed to contribute to a better antioxidant status invivo, the mechanisms involved remain poorly understood,and it is difficult to associate the effect of a given micro-con-stituent with a particular metabolic effect. Most of the studiesconclude that they have a global antioxidant effect as a resultof the action in synergy of all the antioxidant compounds inwhole-grain cereals (Slavin, 2003).
Cereals and cereal products are good examples of complexfoods rich in antioxidants of various origins. And the supply ofvarious antioxidants at low concentrations is preferable to sup-plying a very high concentration of a single antioxidant(Thompson, 1994). From a nutritional perspective, it is there-fore preferable not to remove bioactive compounds, ratherthan to isolate and incorporate them into the food.
We must also emphasize that whole-grain cereals are a partic-ularly rich source of betaine (and choline), which are important inhuman nutrition as they decrease the concentration of the cardio-vascular risk factor homocysteine in the plasma. It is paradoxicalthat no studies have yet tried to investigate the antioxidant protec-tive role of cereal betaine. The mechanisms by which cerealbetaine and choline may improve the redox state within organismwould deserve to be further explored. Whole-grain cerealsare also a good source of sulphur amino acids and trace elements,both of which contribute to the body’s antioxidant defences, byfavouring glutathione synthesis or serving as antioxidantenzymes cofactors. The importance of these cereal compoundsfor the antioxidant defences has been largely neglected.
Lastly, to our knowledge, no studies deal with the influenceof whole-grain cereal consumption on the induction/repressionof genes coding for antioxidant compounds. Further studiesare needed to explore this new area of research using the
most recent genomic and transcriptomic techniques. The im-pact of cereal consumption can also be investigated throughanother recent global approach, i.e. metabolomics. This newapproach will allow us to investigate further how complex an-tioxidant-rich foods such as cereals and cereal products canmodify general metabolism, and which metabolic pathways(beyond the usual oxidized lipids) are affected by antioxidants(Fardet et al., 2007; Rezzi et al., 2007). This will provide newinformation on the health benefits of whole-grain cereals.
References
Adom, K.K., Liu, R.H., 2002. Antioxidant activity of grains. Journal of Agri-
cultural and Food Chemistry 50, 6182e6187.
Adom, K.K., Sorrells, M.E., Liu, R.H., 2003. Phytochemical profiles and
antioxidant activity of wheat varieties. Journal of Agricultural and Food
Chemistry 51, 7825e7834.
Alvarez, P., Alvarado, C., Mathieu, F., Jimenez, L., De la Fuente, M., 2006.
Diet supplementation for 5 weeks with polyphenol-rich cereals improves
several functions and the redox state of mouse leucocytes. European
Journal of Nutrition 45, 428e438.
Anderson, J.W., 2003. Whole grains protect against atherosclerotic cardiovas-
cular disease. The Proceedings of the Nutrition Society 62, 135e142.
Andersson, A., Tengblad, S., Karlstrom, B., Kamal-Eldin, A., Landberg, R.,
Basu, S., Aman, P., Vessby, B., 2007. Whole-grain foods do not affect insulin
sensitivity or markers of lipid peroxidation and inflammation in healthy,
moderately overweight subjects. Journal of Nutrition 137, 1401e1407.
Andreasen, M.F., Christensen, L.P., Meyer, A.S., Hansen, A., 2000. Content of
phenolic acids and ferulic acid dehydrodimers in 17 rye (Secale cereale L.)
varieties. Journal of Agricultural and Food Chemistry 48, 2837e2842.
Andreasen, M.F., Landbo, A.K., Christensen, L.P., Hansen, A., Meyer, A.S.,
2001. Antioxidant effects of phenolic rye (Secale cereale L.) extracts,
monomeric hydroxycinnamates, and ferulic acid dehydrodimers on human
low-density lipoproteins. Journal of Agricultural and Food Chemistry 49,
4090e4096.
Anjaneyulu, M., Chopra, K., 2004. Nordihydroguairetic acid, a lignin,
prevents oxidative stress and the development of diabetic nephropathy in
rats. Pharmacology 72, 42e50.
Aruoma, O.I., 1998. Free radicals, oxidative stress, and antioxidants in human
health and disease. Journal of the American Oil Chemists Society 75,
199e212.
Awika, J.M., Rooney, L.W., Wu, X., Prior, R.L., Cisneros-Zevallos, L., 2003.
Screening methods to measure antioxidant activity of sorghum (Sorghum
bicolor) and sorghum products. Journal of Agricultural and Food
Chemistry 51, 6657e6662.
Awika, J.M., Rooney, L.W., Waniska, R.D., 2005. Anthocyanins from black
sorghum and their antioxidant properties. Food Chemistry 90, 293e301.
Bansal, A.K., Bansal, M., Soni, G., Bhatnagar, D., 2005. Protective role of
Vitamin E pre-treatment on N-nitrosodiethylamine induced oxidative
stress in rat liver. Chemico-Biological Interactions 156, 101e111.
Barak, A.J., Beckenhauer, H.C., Mailliard, M.E., Kharbanda, K.K.,
Tuma, D.J., 2003. Betaine lowers elevated S-adenosylhomocysteine levels
in hepatocytes from ethanol-fed rats. Journal of Nutrition 133, 2845e2848.
Battelli, M.G., Corte, E.D., Stirpe, F., 1972. Xanthine oxidase type D (dehy-
drogenase) in the intestine and other organs of the rat. Biochemical Journal
126, 747e749.
Baublis, A.J., Lu, C., Clydesdale, F.M., Decker, E.A., 2000. Potential of
wheat-based breakfast cereals as a source of dietary antioxidants. Journal
of the American College of Nutrition 19, 308Se311S.
Begum, A.N., Nicolle, C., Mila, I., Lapierre, C., Nagano, K., Fukushima, K.,
Heinonen, S.M., Adlercreutz, H., Remesy, C., Scalbert, A., 2004. Dietary
lignins are precursors of mammalian lignans in rats. Journal of Nutrition
134, 120e127.
Braaten, J.T., Wood, P.J., Scott, F.W., Wolynetz, M.S., Lowe, M.K., Bradley-
White, P., Collins, M.W., 1994. Oat beta-glucan reduces blood cholesterol

273A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
concentration in hypercholesterolemic subjects. European Journal of
Clinical Nutrition 48, 465e474.
Briviba, K., Schnabele, K., Rechkemmer, G., Bub, A., 2004. Supplementation
of a diet low in carotenoids with tomato or carrot juice does not affect lipid
peroxidation in plasma and feces of healthy men. Journal of Nutrition 134,
1081e1083.
Bruce, B., Spiller, G.A., Klevay, L.M., Gallagher, S.K., 2000. A diet high in
whole and unrefined foods favorably alters lipids, antioxidant defenses,
and colon function. Journal of the American College of Nutrition 19,
61e67.
Cao, G., Booth, S.L., Sadowski, J.A., Prior, R.L., 1998. Increases in human
plasma antioxidant capacity after consumption of controlled diets high
in fruit and vegetables. American Journal of Clinical Nutrion 68,
1081e1087.
Casiraghi, M.C., Garsetti, M., Testolin, G., Brighenti, F., 2006. Post-prandial
responses to cereal products enriched with barley beta-glucan. Journal of
the American College of Nutrition 25, 313e320.
Castelluccio, C., Bolwell, G.P., Gerrish, C., RiceEvans, C., 1996. Differen-
tial distribution of ferulic acid to the major plasma constituents in
relation to its potential as an antioxidant. Biochemical Journal 316,
691e694.
Chatenoud, L., La Vecchia, C., Franceschi, S., Tavani, A., Jacobs Jr., D.R.,
Parpinel, M.T., Soler, M., Negri, E., 1999. Refined-cereal intake and risk
of selected cancers in italy. American Journal of Clinical Nutrion 70,
1107e1110.
Chatenoud, L., Tavani, A., La Vecchia, C., Jacobs Jr., D.R., Negri, E., Levi, F.,
Franceschi, S., 1998. Whole grain food intake and cancer risk.
International Journal of Cancer 77, 24e28.
Chen, C.Y., Milbury, P.E., Kwak, H.K., Collins, F.W., Samuel, P.,
Blumberg, J.B., 2004. Avenanthramides and phenolic acids from oats are
bioavailable and act synergistically with vitamin C to enhance hamster
and human LDL resistance to oxidation. Journal of Nutrition 134,
1459e1466.
Chen, C.Y.O., Milbury, P.E., Collins, F.W., Blumberg, J.B., 2007. Avenanthra-
mides are bioavailable and have antioxidant activity in humans after acute
consumption of an enriched mixture from oats. Journal of Nutrition 137,
1375e1382.
Chiang, A.N., Wu, H.L., Yeh, H.I., Chu, C.S., Lin, H.C., Lee, W.C., 2006.
Antioxidant effects of black rice extract through the induction of
superoxide dismutase and catalase activities. Lipids 41, 797e803.
Choi, Y., Jeong, H.S., Lee, J., 2007. Antioxidant activity of methanolic
extracts from some grains consumed in Korea. Food Chemistry 103,
130e138.
Ciappellano, S., Testolin, G., Porrini, M., 1989. Effects of durum wheat dietary
selenium on glutathione peroxidase activity and Se content in long-
term-fed rats. Annals of Nutrition & Metabolism 33, 22e30.
Curtin, D., Hanson, R., Lindley, T.N., Butler, R.C., 2006. Selenium concentra-
tion in wheat (Triticum aestivum) grain as influenced by method, rate, and
timing of sodium selenate application. New Zealand Journal of Crop and
Horticultural Science 34, 329e339.
de Oliveira, G.P.R., Rodriguez-Amaya, D.B., 2007. Processed and prepared
corn products as sources of lutein and zeaxanthin: Compositional variation
in the food chain. Journal of Food Science 72. S79-þ.
Del Pozo-Insfran, D., Brenes, C.H., Saldivar, S.O.S., Talcott, S.T., 2006.
Polyphenolic and antioxidant content of white and blue corn (Zea mays
L.) products. Food Research International 39, 696e703.
Dewanto, V., Wu, X., Liu, R.H., 2002. Processed sweet corn has higher
antioxidant activity. Journal of Agriculture and Food Chemistry 50,
4959e4964.
Dykes, L., Rooney, L.W., 2006. Sorghum and millet phenols and antioxidants.
Journal of Cereal Science 44, 236e251.
Eastwood, M.A., 1999. Interaction of dietary antioxidants in vivo: how fruit
and vegetables prevent disease? QJMdan International Journal of
Medicine 92, 527e530.
Emmons, C.L., Peterson, D.M., 1999. Antioxidant activity and phenolic
contents of oat groats and hulls. Cereal Chemistry 76, 902e906.
Emmons, C.L., Peterson, D.M., Paul, G.L., 1999. Antioxidant capacity of oat
(Avena sativa L.) extracts. 2. In vitro antioxidant activity and contents of
phenolic and tocol antioxidants. Journal of Agricultural and Food
Chemistry 47, 4894e4898.
Esmaillzadeh, A., Mirmiran, P., Azizi, F., 2005. Whole-grain consumption and
the metabolic syndrome: a favorable association in Tehranian adults. Euro-
pean Journal of Clinical Nutrition 59, 353e362.
Fardet, A., Canlet, C., Gottardi, G., Lyan, B., Remesy, C., Mazur, A., Paris, A.,
Scalbert, A., 2007. Whole grain and refined wheat flours show distinct
metabolic profiles in rats as assessed by a 1H NMR-based metabonomic
approach. Journal of Nutrition 4, 923e929.
Feillet, P., 2000. Le grain de ble. Composition et utilisation. Editions INRA.
308 pages.
Furukawa, S., Fujita, T., Shimabukuro, M., Iwaki, M., Yamada, Y., Nakajima, Y.,
Nakayama, O., Makishima, M., Matsuda, M., Shimomura, I., 2004.
Increased oxidative stress in obesity and its impact on metabolic syndrome.
Journal of Clinical Investigation 114, 1752e1761.
Gallardo, C., Jimenez, L., Garcia-Conesa, M.T., 2006. Hydroxycinnamic acid
composition and in vitro antioxidant activity of selected grain fractions.
Food Chemistry 99, 455e463.
Goffman, F.D., Bergman, C.J., 2004. Rice kernel phenolic content and its
relationship with antiradical efficiency. Journal of the Science of Food
and Agriculture 84, 1235e1240.
Golestani, A., Rastegar, R., Shariftabrizi, A., Khaghani, S., Payabvash, S.M.,
Salmasi, A.H., Dehpour, A.R., Pasalar, P., 2006. Paradoxical dose- and
time-dependent regulation of superoxide dismutase and antioxidant
capacity by vitamin E in rat. Clinica Chimica Acta 365, 153e159.
Graf, E., Eaton, J.W., 1990. Antioxidant functions of phytic acid. Free Radical
Biology & Medicine 8, 61e69.
Graf, E., Empson, K.L., Eaton, J.W., 1987. Phytic acid. A natural antioxidant.
Journal of Biological Chemistry 262, 11647e11650.
Gray, D.A., Clarke, M.J., Baux, C., Bunting, J.P., Salter, A.M., 2002.
Antioxidant activity of oat extracts added to human LDL particles and
in free radical trapping assays. Journal of Cereal Science 36, 209e218.
Guo, H.H., Ling, W.H., Wang, Q., Liu, C., Hu, Y., Xia, M., Feng, X.,
Xia, X.D., 2007. Effect of anthocyanin-rich extract from black rice (Oryzasativa L. indica) on hyperlipidemia and insulin resistance in fructose-fed
rats. Plant Foods for Human Nutrition 62, 1e6.
Halliwell, B., Aruoma, O.I., 1991. DNA damage by oxygen-derived species.
Its mechanism and measurement in mammalian systems. FEBS Letters
281, 9e19.
Halliwell, B., Zhao, K., Whiteman, M., 2000. The gastrointestinal tract:
a major site of antioxidant action? Free Radical Research 33, 819e830.
Hallund, J., Ravn-Haren, G., Bugel, S., Tholstrup, T., Tetens, I., 2006. A lignan
complex isolated from flaxseed does not affect plasma lipid concentrations
or antioxidant capacity in healthy postmenopausal women. Journal of
Nutrition 136, 112e116.
Handelman, G.J., Cao, G., Walter, M.F., Nightingale, Z.D., Paul, G.L.,
Prior, R.L., Blumberg, J.B., 1999. Antioxidant capacity of oat (Avena
sativa L.) extracts. 1. Inhibition of low-density lipoprotein oxidation and
oxygen radical absorbance capacity. Journal of Agricultural and Food
Chemistry 47, 4888e4893.
Harder, H., Tetens, I., Let, M.B., Meyer, A.S., 2004. Rye bran bread intake
elevates urinary excretion of ferulic acid in humans, but does not affect
the susceptibility of LDL to oxidation ex vivo. European Journal of
Nutrition 43, 230e236.
Hegde, P.S., Rajasekaran, N.S., Chandra, T.S., 2005. Effects of the antioxidant
properties of millet species on oxidative stress and glycemic status in
alloxan-induced rats. Nutrition Research 25, 1109e1120.
Hemery, Y., Rouau, X., Lullien-Pellerin, V., Barron, C., Abecassis, J., 2007.
Dry processes to develop wheat fractions and products with enhanced
nutritional quality. Journal of Cereal Science 46, 327e347.
Hernanz, D., Nunez, V., Sancho, A.I., Faulds, C.B., Williamson, G.,
Bartolome, B., Gomez-Cordoves, C., 2001. Hydroxycinnamic acids and
ferulic acid dehydrodimers in barley and processed barley. Journal of
Agricultural and Food Chemistry 49, 4884e4888.
Hosny, M., Rosazza, J.P.N., 1997. Structures of ferulic acid glycoside esters in
corn hulls. Journal of Natural Products 60, 219e222.
Hsieh, R.H., Lien, L.M., Lin, S.H., Chen, C.W., Cheng, H.J., Cheng, H.H.,
2005. Alleviation of oxidative damage in multiple tissues in rats with

274 A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
streptozotocin-induced diabetes by rice bran oil supplementation. In:
Hsu, C.Y., Le, H.M., Wei, Y.H. (Eds.), Role of the Mitochondria in
Human Aging and Disease: From Genes to Cell Signaling. Blackwell,
New York, pp. 365e371.
Hu, C., Zawistowski, J., Ling, W., Kitts, D.D., 2003. Black rice (Oryza sativaL. indica) pigmented fraction suppresses both reactive oxygen species and
nitric oxide in chemical and biological model systems. Journal of
Agricultural and Food Chemistry 51, 5271e5277.
Hung, H.C., Joshipura, K.J., Jiang, R., Hu, F.B., Hunter, D., Smith-
Warner, S.A., Colditz, G.A., Rosner, B., Spiegelman, D., Willett, W.C.,
2004. Fruit and vegetable intake and risk of major chronic disease. Journal
of the National Cancer Institute 96, 1577e1584.
Hwang, E.S., Kim, G.H., 2007. Biomarkers for oxidative stress status of DNA,
lipids, and proteins in vitro and in vivo cancer research. Toxicology 229,
1e10.
Jang, Y., Lee, J.H., Kim, O.Y., Park, H.Y., Lee, S.Y., 2001. Consumption of
whole grain and legume powder reduces insulin demand, lipid peroxida-
tion, and plasma homocysteine concentrations in patients with coronary ar-
tery disease: randomized controlled clinical trial. Arteriosclerosis,
Thrombosis, and Vascular Biology 21, 2065e2071.
Juliano, C., Cossu, M., Alamanni, M.C., Piu, L., 2005. Antioxidant activity
of gamma-oryzanol: Mechanism of action and its effect on oxidative
stability of pharmaceutical oils. International Journal of Pharmaceutics
299, 146e154.
Kahlon, T.S., Chow, F.I., Knuckles, B.E., Chiu, M.M., 1993. Cholesterol-low-
ering effects in hamsters of beta-glucan-enriched barley fraction, dehulled
whole barley, rice bran, and oat bran and their combinations. Cereal
Chemistry 70, 435e440.
Kamal-Eldin, A., Pouru, A., Eliasson, C., Aman, P., 2001. Alkylresorcinols as
antioxidants: hydrogen donation and peroxyl radical-scavenging effects.
Journal of the Science of Food and Agriculture 81, 353e356.
Kamath, V.G., Chandrashekar, A., Rajini, P.S., 2004. Antiradical properties of
sorghum (Sorghum bicolor L. Moench) flour extracts. Journal of Cereal
Science 40, 283e288.
Kern, S.M., Bennett, R.N., Mellon, F.A., Kroon, P.A., Garcia-Conesa, M.T.,
2003. Absorption of hydroxycinnamates in humans after high-bran cereal
consumption. Journal of Agricultural and Food Chemistry 51, 6050e6055.
Kim, J.Y., Do, M.H., Lee, S.S., 2006. The effects of a mixture of brown and
black rice on lipid profiles and antioxidant status in rats. Annals of
Nutrition and Metabolism 50, 347e353.
Kitts, D.D., Yuan, Y.V., Wijewickreme, A.N., Thompson, L.U., 1999.
Antioxidant activity of the flaxseed lignan secoisolariciresinol diglycoside
and its mammalian lignan metabolites enterodiol and enterolactone.
Molecular and Cellular Biochemistry 202, 91e100.
Konopka, I., Kozirok, W., Rotkiewicz, D., 2004. Lipids and carotenoids of
wheat grain and flour and attempt of correlating them with digital image
analysis of kernel surface and cross-sections. Food Research International
37, 429e438.
Kozubek, A., Nienartowicz, B., 1995. Cereal grain resorcinolic lipids inhibit
H2O2-induced peroxidation of biological membranes. Acta Biochimica
Polonica 42, 309e315.
Krawinkel, M.B., 2007. What we know and don’t know about Golden Rice.
Nature Biotechnology 25. 623e623.
Labaj, J., Slamenova, D., Lazarova, M., Kosikova, M., 2004. Lignin-stimu-
lated reduction of oxidative DNA lesions in testicular cells and
lymphocytes of Sprague-Dawley rats in vitro and ex vivo. Nutrition
and Cancerdan International Journal 50, 198e205.
Laight, D.W., Desai, K.M., Gopaul, N.K., Anggard, E.E., Carrier, M.J., 1999.
F-2-isoprostane evidence of oxidant stress in the insulin resistant, obese
Zucker rat: effects of vitamin E. European Journal of Pharmacology
377, 89e92.
Lee, J.M., Calkins, M.J., Chan, K.M., Kan, Y.W., Johnson, J.A., 2003. Identi-
fication of the NF-E2-related factor-2-dependent genes conferring
protection against oxidative stress in primary cortical astrocytes using
oligonucleotide microarray analysis. Journal of Biological Chemistry
278, 12029e12038.
Leenhardt, F., Levrat-Verny, M.A., Chanliaud, E., Remesy, C., 2005. Moderate
decrease of pH by sourdough fermentation is sufficient to reduce phytate
content of whole wheat flour through endogenous phytase activity. Journal
of Agricultural and Food Chemistry 53, 98e102.
Leenhardt, F., Lyan, B., Rock, E., Boussard, A., Potus, J., Chanliaud, E.,
Remesy, C., 2006. Genetic variability of carotenoid concentration,
and lipoxygenase and peroxidase activities among cultivated wheat
species and bread wheat varieties. European Journal of Agronomy
25, 170e176.
Leenhardt, F., Fardet, A., Lyan, B., Gueux, E., Rock, E., Mazur, A., Chan-
liaud, E., Demigne, C., Remesy, C., in press. Wheat germ supplementa-
tion of a low vitamin E diet in rats affords a more effective antioxidant
protection in tissues than in plasma. Journal of the American College of
Nutrition.
Leonard, S.W., Good, C.K., Gugger, E.T., Traber, M.G., 2004. Vitamin E
bioavailability from fortified breakfast cereal is greater than that from
encapsulated supplements. American Journal of Clinical Nutrition 79,
86e92.
Lewis, S., Bolton, C., Heaton, K., 1996. Lack of influence of intestinal transit
on oxidative status in premenopausal women. European Journal of Clinical
Nutrition 50, 565e568.
Li, W., Bollecker, S.S., Schofield, J.D., 2004. Glutathione and related thiol
compounds. I. Glutathione and related thiol compounds in flour. Journal
of Cereal Science 39, 205e212.
Li, W.D., Wei, C.L., White, P.J., Beta, T., 2007. High-amylose corn exhibits
better antioxidant activity than typical and waxy genotypes. Journal of
Agricultural and Food Chemistry 55, 291e298.
Ling, W.H., Cheng, Q.X., Ma, J., Wang, T., 2001. Red and black rice decrease
atherosclerotic plaque formation and increase antioxidant status in rabbits.
Journal of Nutrition 131, 1421e1426.
Ling, W.H., Wang, L.L., Ma, J., 2002. Supplementation of the black rice outer
layer fraction to rabbits decreases atherosclerotic plaque formation and
increases antioxidant status. Journal of Nutrition 132, 20e26.
Liu, R.H., 2007. Whole grain phytochemicals and health. Journal of Cereal
Science 46, 207e219.
Liu, Q., Yao, H.Y., 2007. Antioxidant activities of barley seeds extracts. Food
Chemistry 102, 732e737.
Liyana-Pathirana, C.M., Shahidi, F., 2005. Antioxidant activity of
commercial soft and hard wheat (Triticum aestivium L.) as affected
by gastric pH conditions. Journal of Agricultural and Food Chemistry
53, 2433e2440.
Liyana-Pathirana, C.M., Shahidi, F., 2006a. Antioxidant properties of
commercial soft and hard winter wheats (Triticum aestivum L.) and their
milling fractions. Journal of the Science of Food and Agriculture 86,
477e485.
Liyana-Pathirana, C.M., Shahidi, F., 2006b. Importance of insoluble-bound
phenolics to antioxidant properties of wheat. Journal of Agricultural and
Food Chemistry 54, 1256e1264.
Liyana-Pathirana, C.M., Shahidi, F., 2007. The antioxidant potential of
milling fractions from breadwheat and durum. Journal of Cereal Science
45, 238e247.
Lopez, H.W., Duclos, V., Coudray, C., Krespine, V., Feillet-Coudray, C.,
Messager, A., Demigne, C., Remesy, C., 2003. Making bread with
sourdough improves mineral bioavailability from reconstituted whole
wheat flour in rats. Nutrition 19, 524e530.
Lotito, S.B., Frei, B., 2004a. The increase in human plasma antioxidant capac-
ity after apple consumption is due to the metabolic effect of fructose on
urate, not apple-derived antioxidant flavonoids. Free Radical Biology &
Medicine 37, 251e258.
Lotito, S.B., Frei, B., 2004b. Relevance of apple polyphenols as antioxidants in
human plasma: contrasting in vitro and in vivo effects. Free Radical
Biology & Medicine 36, 201e211.
Maillard, M.N., Soum, M.H., Boivin, P., Berset, C., 1996. Antioxidant
activity of barley and malt: Relationship with phenolic content. Food Sci-
ence and TechnologydLebensmittel-Wissenschaft & Technologie 29,
238e244.
Mariotti, F., Simbelie, K.L., Makarios-Lahham, L., Huneau, J.F., Laplaize, B.,
Tome, D., Even, P.C., 2004. Acute ingestion of dietary proteins improves
post-exercise liver glutathione in rats in a dose-dependent relationship with
their cysteine content. Journal of Nutrition 134, 128e131.

275A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
Martin, A., Wu, D.Y., Baur, W., Meydani, S.N., Blumberg, J.B., Meydani, M.,
1996. Effect of vitamin E on human aortic endothelial cell responses to
oxidative injury. Free Radical Biology and Medicine 21, 505e511.
Martinez-Tome, M., Murcia, M.A., Frega, N., Ruggieri, S., Jimenez, A.M.,
Roses, F., Parras, P., 2004. Evaluation of antioxidant capacity of cereal
brans. Journal of Agricultural and Food Chemistry 52, 4690e4699.
Michalska, A., Ceglinska, A., Amarowicz, R., Piskula, M.K., Szawara-
Nowak, D., Zielinski, H., 2007. Antioxidant contents and antioxidative
properties of traditional rye breads. Journal of Agricultural and Food
Chemistry 55, 734e740.
Miller, H.E., Rigelhof, F., Marquart, L., Prakash, A., Kanter, M., 2000.
Antioxidant content of whole grain breakfast cereals, fruits and vegetables.
Journal of the American College of Nutrition 19, 312Se319S.
Minhajuddin, M., Beg, Z.H., Iqbal, J., 2005. Hypolipidemic and antioxidant
properties of tocotrienol rich fraction isolated from rice bran oil in
experimentally induced hyperlipidemic rats. Food and Chemical
Toxicology 43, 747e753.
Mitchell, G.V., Grundel, E., Jenkins, M.Y., 1996. Bioavailability for rats of
vitamin E from fortified breakfast cereals. Journal of Food Science 61,
1257e1260.
Moore, J., Hao, Z., Zhou, K., Luther, M., Costa, J., Yu, L.L., 2005. Carotenoid,
tocopherol, phenolic acid, and antioxidant properties of Maryland-grown
soft wheat. Journal of Agricultural and Food Chemistry 53, 6649e6657.
Moore, J., Liu, J.G., Zhou, K.Q., Yu, L.L., 2006. Effects of genotype and
environment on the antioxidant properties of hard winter wheat bran.
Journal of Agricultural and Food Chemistry 54, 5313e5322.
Morand, C., Rios, L., Moundras, C., Besson, C., Remesy, C., Demigne, C.,
1997. Influence of methionine availability on glutathione synthesis and
delivery by the liver. The Journal of Nutritional Biochemistry 8,
246e255.
Muraoka, S., Miura, T., 2004. Inhibition of xanthine oxidase by phytic acid
and its antioxidative action. Life Sciences 74, 1691e1700.
Myhrstad, M.C.W., Carlsen, H., Nordstrom, O., Blomhoff, R., Moskaug, J.O.,
2002. Flavonoids increase the intracellular glutathione level by transactiva-
tion of the gamma-glutamylcysteine synthetase catalytical subunit
promoter. Free Radical Biology and Medicine 32, 386e393.
Na, H.K., Surh, Y.J., 2006. Intracellular signaling network as a prime
chemopreventive target of (�)-epigallocatechin gallate. Molecular
Nutrition and Food Research 50, 152e159.
Nagah, A.M., Seal, C.J., 2005. In vitro procedure to predict apparent
antioxidant release from wholegrain foods measured using three different
analytical methods. Journal of the Science of Food and Agriculture 85,
1177e1185.
Nilsson, M., Aman, P., Harkonen, H., Hallmans, G., Knudsen, K.E.B.,
Mazur, W., Adlercreutz, H., 1997. Content of nutrients and lignans in roller
milled fractions of rye. Journal of the Science of Food and Agriculture 73,
143e148.
Oki, T., Masuda, M., Kobayashi, M., Nishiba, Y., Furuta, S., Suda, I.,
Sato, T., 2002. Polymeric procyanidins as radical-scavenging compo-
nents in red-hulled rice. Journal of Agricultural and Food Chemistry
50, 7524e7529.
Olmo, N., Turnay, J., Perez-Ramos, P., Lecona, E., Barrasa, J.I., de
Silanes, I.L., Lizarbe, M.A., 2007. In vitro models for the study of the ef-
fect of butyrate on human colon adenocarcinoma cells. Toxicology in Vitro
21, 262e270.
Panfili, G., Fratianni, A., Irano, M., 2004. Improved normal-phase high-perfor-
mance liquid chromatography procedure for the determination of
carotenoids in cereals. Public Health Nutrition 52, 6373e6377.
Parikka, K., Rowland, I.R., Welch, R.W., Wahala, K., 2006. In vitro
antioxidant activity and antigenotoxicity of 5-n-alkylresorcinols. Journal
of Agricultural and Food Chemistry 54, 1646e1650.
Parrado, J., Miramontes, E., Jover, M., Marquez, J.C., Angeles Mejias, M.,
Collantes De Teran, L., Absi, E., Bautista, J., 2003. Prevention of brain
protein and lipid oxidation elicited by a water-soluble oryzanol
enzymatic extract derived from rice bran. European Journal of Nutrition
42, 307e314.
Paterson, E., Gordon, M.H., Niwat, C., George, T.W., Parr, L.,
Waroonphan, S., Lovegrove, J.A., 2006. Supplementation with fruit and
vegetable soups and beverages increases plasma carotenoid concentrations
but does not alter markers of oxidative stress or cardiovascular risk factors.
Journal of Nutrition 136, 2849e2855.
Pellegrini, N., Serafini, M., Salvatore, S., Del Rio, D., Bianchi, M.,
Brighenti, F., 2006. Total antioxidant capacity of spices, dried fruits,
nuts, pulses, cereals and sweets consumed in Italy assessed by three differ-
ent in vitro assays. Molecular Nutrition & Food Research 50, 1030e1038.
Perez-Jimenez, J., Saura-Calixto, F., 2005. Literature data may underestimate
the actual antioxidant capacity of cereals. Journal of Agricultural and Food
Chemistry 53, 5036e5040.
Peterson, D.M., 2001. Oat antioxidants. Journal of Cereal Science 33,
115e129.
Peterson, D.M., Hahn, M.J., Emmons, C.L., 2002. Oat avenanthramides
exhibit antioxidant activities in vitro. Food Chemistry 79, 473e478.
Pool-Zobel, B.L., Adlercreutz, H., Glei, M., Liegibel, U.M., Sittlingon, J.,
Rowland, I., Wahala, K., Rechkemmer, G., 2000. Isoflavonoids and lignans
have different potentials to modulate oxidative genetic damage in human
colon cells. Carcinogenesis 21, 1247e1252.
Prior, R.L., 2003. Fruits and vegetables in the prevention of cellular oxidative
damage. American Journal of Clinical Nutrition 78, 570Se578S.
Ragaee, S., Abdel-Aal, E.M., Noaman, M., 2006. Antioxidant activity and
nutrient composition of selected cereals for food use. Food Chemistry
98, 32e38.
Reeves, P.G., Gregoire, B.R., Garvin, D.F., Hareland, G.A., Lindlauf, J.E.,
Johnson, L.K., Finley, J.W., 2007. Determination of selenium bioavailabil-
ity from wheat mill fractions in rats by using the slope-ratio assay and
a modified Torula yeast-based diet. Journal of Agricultural and Food
Chemistry 55, 516e522.
Reinosa, O., Can, B.T., Ricard, J.L., 1998. Yield and antioxidant activity of
lignin obtained at different pH values. Boletin De La Sociedad Chilena
De Quimica 43, 367e373.
Rezar, V., Pajk, T., Marinsek Logar, R., Jese Janezic, V., Salobir, K.,
Oresnik, A., Salobir, J., 2003. Wheat bran and oat bran effectively reduce
oxidative stress induced by high-fat diets in pigs. Annals of Nutrition &
Metabolism 47, 78e84.
Rezzi, S., Ramadan, Z., Fay, L.B., Kochhar, S., 2007. Nutritional metabo-
nomics: Applications and perspectives. Journal of Proteome Research 6,
513e525.
Rice-Evans, C.A., Miller, N.J., Paganga, G., 1997. Antioxidant properties of
phenolic compounds. Trends in Plant Science 2, 152e159.
Rimbach, G., Pallauf, J., 1998. Phytic acid inhibits free radical formation in
vitro but does not affect liver oxidant or antioxidant status in growing
rats. Journal of Nutrition 128, 1950e1955.
Ross, A.B., Shepherd, M.J., Knudsen, K.E.B., Glitso, L.V., Bowey, E.,
Phillips, J., Rowland, I., Guo, Z.X., Massy, D.J.R., Aman, P., Kamal-
Eldin, A., 2003a. Absorption of dietary alkylresorcinols in ileal-cannulated
pigs and rats. British Journal of Nutrition 90, 787e794.
Ross, A.B., Shepherd, M.J., Schupphaus, M., Sinclair, V., Alfaro, B., Kamal-
Eldin, A., Aman, P., 2003b. Alkylresorcinols in cereals and cereal
products. Journal of Agricultural and Food Chemistry 51, 4111e4118.
Saura-Calixto, F., Goni, I., 2006. Antioxidant capacity of the Spanish Mediter-
ranean diet. Food Chemistry 94, 442e447.
Saura-Calixto, F., Serrano, J., Goni, I., 2007. Intake and bioaccessibility of
total polyphenols in a whole diet. Food Chemistry 101, 492e501.
Scalbert, A., Williamson, G., 2000. Dietary intake and bioavailability of
polyphenols. Journal of Nutrition 130, 2073Se2085S.
Schofield, J.D., Chen, X., 1995. Analysis of free reduced and free oxidized
glutathione in wheat-flour. Journal of Cereal Science 21, 127e136.
Serpen, A., Capuano, E., Fogliano, V., Gokmen, V., 2007. A new procedure to
measure the antioxidant activity of insoluble food components. Journal of
Agricultural and Food Chemistry 55, 7676e7681.
Shewry, P.R., 2007. Improving the protein content and composition of cereal
grain. Journal of Cereal Science 46, 239e250.
Skibsted, L.H., Dragsted, L.O., Dyerberg, J., Hansen, H.S., Kiens, B.,
Ovesen, L.F., Tjonneland, A.M., 2006. Antioxidants and health. Ugeskr
Laeger 168, 2787e2789 (in Danish).
Slamenova, D., Horvathova, E., Kosikova, B., Ruzekova, L., Labaj, J., 1999.
Detection of lignin biopolymer- and vitamin E-stimulated reduction of

276 A. Fardet et al. / Journal of Cereal Science 48 (2008) 258e276
DNA strand breaks in H2O2- and MNNG-treated mammalian cells by the
comet assay. Nutrition and Cancer 33, 88e94.
Slamenova, D., Kosikova, B., Labaj, J., Ruzekova, L., 2000. Oxidative/antiox-
idative effects of different lignin preparations on DNA in hamster V79
cells. Neoplasma 47, 349e353.
Slavin, J., 2003. Why whole grains are protective: biological mechanisms.
Proceedings of the Nutrition Society 62, 129e134.
Slavin, J.L., Martini, M.C., Jacobs Jr., D.R., Marquart, L., 1999. Plausible
mechanisms for the protectiveness of whole grains. American Journal of
Clinical Nutrition 70, 459Se463S.
Sosulski, F., Krygier, K., Hogge, L., 1982. Free, esterified, and insoluble-
bound phenolic acids. 3. Composition of phenolic acids in cereal and
potato flours. Journal of Agricultural and Food Chemistry 30, 337e340.
Souci, S.W., Fachmann, W., Kraut, H., 2000. Food Composition and
Nutritional Tables. Medpharm Scientific Publishers/CRC Press, Stuttgart,
1182 pp.
Subba Rao, M.V., Muralikrishna, G., 2002. Evaluation of the antioxidant
properties of free and bound phenolic acids from native and malted finger
millet (ragi, Eleusine coracana Indaf-15). Journal of Agricultural and Food
Chemistry 50, 889e892.
Suh, M.H., Yoo, S.H., Chang, P.S., Lee, H.G., 2005. Antioxidative activity of
microencapsulated gamma-oryzanol on high cholesterol-fed rats. Journal
of Agricultural and Food Chemistry 53, 9747e9750.
Thompson, L.U., 1994. Antioxidants and hormone-mediated health benefits
of whole grains. Critical Reviews in Food Science and Nutrition 34,
473e497.
Thompson, H.J., Heimendinger, J., Sedlacek, S., Haegele, A., Diker, A.,
O’Neill, C., Meinecke, B., Wolfe, P., Zhu, Z.J., Jiang, W.Q., 2005. 8-Iso-
prostane F-2 alpha excretion is reduced in women by increased vegetable
and fruit intake. American Journal of Clinical Nutrition 82, 768e776.
Toyokuni, S., Itani, T., Morimitsu, Y., Okada, K., Ozeki, M., Kondo, S.,
Uchida, K., Osawa, T., Hiai, H., Tashiro, T., 2002. Protective effect of
colored rice over white rice on Fenton reaction-based renal lipid peroxida-
tion in rats. Free Radical Research 36, 583e592.
Truswell, A.S., 2002. Cereal grains and coronary heart disease. European Jour-
nal of Clinical Nutrition 56, 1e14.
U.S. Department of Agriculture, A.R.S, 1999. USDA Nutrient Database for
Standard Reference. Release 13. http://www.nal.usda.gov/fnic/foodcomp.
Venn, B.J., Mann, J.I., 2004. Cereal grains, legumes and diabetes. European
Journal of Clinical Nutrition 58, 1443e1461.
Wang, Q., Han, P.H., Zhang, M.W., Xia, M., Zhu, H.L., Ma, J., Hou, M.J.,
Tang, Z.H., Ling, W.H., 2007. Supplementation of black rice pigment
fraction improves antioxidant and anti-inflammatory status in patients
with coronary heart disease. Asia Pacific Journal of Clinical Nutrition
16, 295e301.
Ward, M., McNulty, H., McPartlin, J., Strain, J.J., Weir, D.G., Scott, J.M.,
1997. Plasma homocysteine, a risk factor for cardiovascular disease, is
lowered by physiological doses of folic acid. QJMdMonthly Journal of
the Association of Physicians 90, 519e524.
Wattenberg, L.W., 1996. Chemoprevention of cancer. Preventive Medicine 25,
44e45.
Winata, A., Lorenz, K., 1997. Effects of fermentation and baking of whole
wheat and whole rye sourdough breads on cereal alkylresorcinols. Cereal
Chemistry 74, 284e287.
Wu, X., Beecher, G.R., Holden, J.M., Haytowitz, D.B., Gebhardt, S.E.,
Prior, R.L., 2004a. Lipophilic and hydrophilic antioxidant capacities of
common foods in the United States. Journal of Agricultural and Food
Chemistry 52, 4026e4037.
Wu, X.L., Gu, L.W., Holden, J., Haytowitz, D.B., Gebhardt, S.E., Beecher, G.,
Prior, R.L., 2004b. Development of a database for total antioxidant
capacity in foods: a preliminary study. Journal of Food Composition and
Analysis 17, 407e422.
Xia, M., Ling, W.H., Ma, J., Kitts, D.D., Zawistowski, J., 2003. Supplementa-
tion of diets with the black rice pigment fraction attenuates atherosclerotic
plaque formation in apolipoprotein e deficient mice. Journal of Nutrition
133, 744e751.
Xia, X.D., Ling, W.H., Ma, J., Xia, M., Hou, M.J., Wang, Q., Zhu, H.L.,
Tang, Z.H., 2006. An anthocyanin-rich extract from black rice enhances
atherosclerotic plaque stabilization in apolipoprotein e-deficient mice.
Journal of Nutrition 136, 2220e2225.
Xu, Z., Hua, N., Godber, J.S., 2001. Antioxidant activity of tocopherols,
tocotrienols, and gamma-oryzanol components from rice bran against
cholesterol oxidation accelerated by 2,20-azobis(2-methylpropionami-
dine) dihydrochloride. Journal of Agricultural and Food Chemistry 49,
2077e2081.
Yu, L., Zhou, K., 2004. Antioxidant properties of bran extracts from ‘Platte’
wheat grown at different locations. Food Chemistry 90, 311e316.
Yu, L.L., Haley, S., Perret, J., Harris, M., 2002a. Antioxidant properties of hard
winter wheat extracts. Food Chemistry 78, 457e461.
Yu, L.L., Haley, S., Perret, J., Harris, M., Wilson, J., Qian, M., 2002b. Free
radical scavenging properties of wheat extracts. Journal of Agricultural
and Food Chemistry 50, 1619e1624.
Yu, L.L., Perret, J., Harris, M., Wilson, J., Haley, S., 2003. Antioxidant
properties of bran extracts from ‘‘Akron’’ wheat grown at different
locations. Journal of Agricultural and Food Chemistry 51, 1566e1570.
Yu, L.L., Zhou, K.Q., Parry, J.W., 2005. Inhibitory effects of wheat bran
extracts on human LDL oxidation and free radicals. Food Science and
Technology-Lebensmittel-Wissenschaft & Technologie 38, 463e470.
Yuan, Y.V., Rickard, S.E., Thompson, L.U., 1999. Short-term feeding of
flaxseed or its lignan has minor influence on in vivo hepatic antioxidant
status in young rats. Nutrition Research 19, 1233e1243.
Zdunczyk, Z., Flis, M., Zielinski, H., Wroblewska, M., Antoszkiewicz, Z.,
Juskiewicz, J., 2006. In vitro antioxidant activities of barley, husked oat,
naked oat, triticale, and buckwheat wastes and their influence on the
growth and biomarkers of antioxidant status in rats. Journal of Agricultural
and Food Chemistry 54, 4168e4175.
Zeisel, S.H., Mar, M.H., Howe, J.C., Holden, J.M., 2003a. Concentrations of
choline-containing compounds and betaine in common foods. Journal of
Nutrition 133, 1302e1307.
Zeisel, S.H., Mar, M.H., Howe, J.C., Holden, J.M., 2003b. Content of choline-
containing compounds and betaine in common foods. FASEB Journal 17.
A766eA766.
Zhao, Z., Egashira, Y., Sanada, H., 2005. Phenolic antioxidants richly
contained in corn bran are slightly bioavailable in rats. Journal of Agricul-
tural and Food Chemistry 53, 5030e5035.
Zhao, H.F., Dong, J.J., Lu, J., Chen, J., Li, Y., Shan, L.J., Lin, Y., Fan, W.,
Gu, G.X., 2006. Effects of extraction solvent mixtures on antioxidant
activity evaluation and their extraction capacity and selectivity for free
phenolic compounds in barley (Hordeum vulgare L.). Journal of Agricul-
tural and Food Chemistry 54, 7277e7286.
Zhao, F., McGrath, S., Gray, C., Lopez-Bellido, J., 2007. Selenium
concentrations in UK wheat and biofortification strategies. Comparative
Biochemistry and Physiology adMolecular & Integrative Physiology
146. S246eS246.
Zhou, J.R., Erdman, J.W., 1995. Phytic acid in health and disease. Critical
Reviews in Food Science and Nutrition 35, 495e508.
Zhou, K., Yu, L., 2004. Antioxidant properties of bran extracts from Trego
wheat grown at different locations. Journal of Agricultural and Food
Chemistry 52, 1112e1117.
Zhou, K., Laux, J.J., Yu, L., 2004. Comparison of Swiss red wheat grain and
fractions for their antioxidant properties. Journal of Agricultural and Food
Chemistry 52, 1118e1123.
Zhou, K.Q., Yin, J.J., Yu, L.L., 2005. Phenolic acid, tocopherol and carotenoid
compositions, and antioxidant functions of hard red winter wheat bran.
Journal of Agricultural and Food Chemistry 53, 3916e3922.
Zielinski, H., Kozlowska, H., 2000. Antioxidant activity and total phenolics in
selected cereal grains and their different morphological fractions. Journal
of Agricultural and Food Chemistry 48, 2008e2016.
Copyright © 2022 FDOKUMEN




















