
Involvement of the sensor kinase EnvZ in the in vivo activation of the response-regulator PhoB by...
13
Molecular Microbiology (1996) 22(1), 135-147 Involvement of the sensor kinase EnvZ in the in vivo activation of the response-regulator PhoB by acetyl phosphate Soo-Ki Kim, Mary R. Wilmes-Hesenberg+ and Barry L. Wanner* Department of Biologics/ Sciences, Purdue University, West Lafayette, Indiana 47907, USA. Summary Three signalling pathways lead to activation of the phosphate (Pho) regulon by phosphorylation of the response-regulator PhoB in Eschefichia coli. One pathway responds to the extracellular inorganic phosphate (Pi) level and leads to activation by the Pi sensor kinase, PhoR. The other two pathways are Pi independent and are apparent in the absence of PhoR. One Pi-independent pathway responds to the level of an unknown catabolite and leads to activation by the catabolite regulatory sensor kinase, CreC (originally called PhoM); the other Pi-independent pathway responds to acetyl phosphate and leads to activation by a process requiring acetyl phosphate. Here we show that activation of PhoB by acetyl phosphate can require the sensor kinase EnvZ. Accordingly, we propose that the in vivo activation of PhoB by acetyl phosphate (and perhaps other two- component response-regulators as well) probably always requiresa certain kinase that can vary depend- ing upon the growth conditions. Introduction The phosphate (Pho) regulons of Escherichia coliand Sal- monella typhimurium consist of at least 31 and 21 genes, respectively, that are co-regulatedby inorganic phosphate (Pi; Jiang eta/., 1995;Wanner, 1996).These include a total of 38 different genes that are arranged in 11 transcriptional units. One unit, the pho6R operon, encodes the two- component regulatory system comprising the response- regulator PhoB and its partner sensor kinase, PhoR. Each Pi-regulated Pho regulon promoter contains one or more ‘Pho Box’ sequences (Makino eta/., 1988) required Received 6 June, 1996; revised 25 July, 1996; accepted 2 August, 1996. +Present address: Department of Biology, Washington Univer- sity, St. Louis, Missouri 63130, USA. *For correspondence. E-mail BLWQbilbo.bio.purdue.edu; Tel. (317) 494 8034; Fax (317) 494 0876. for regulation. The expression of most of these promoters is known to require phospho-PhoBfor transcriptional acti- vation (Makino eta/., 1992; S.-K. Kim and B. L. Wanner, unpublished). The primary signalling pathway for activation of PhoB responds to the environmental (extracellular) Pi level. This signalling pathway involves activation under conditions of Pi limitation and inhibition when Pi is in excess. Both of these processes require PhoR. Inhibition requires, in addi- tion, an intact, high-affinity Pi-specifictransporter (the Pst system) and a proteincalled PhoU. Control by Pi is brought about via the phosphorylation of PhoB by PhoR when environmental Pi is limiting (less than about 4pM) and dephosphorylation of phospho-PhoB by PhoR (perhaps together with a Pst component(s) or PhoU) when Pi is in excess (greater than about 4 pM; for a review see Wanner, 1 996). Two additional signalling pathways lead to activation of PhoB in the absence of PhoR. One requires the catabolite regulatory sensor kinase CreC (originally called PhoM), and the other requires acetyl phosphate (Wanner and Wilmes-Riesenberg, 1992). Both of these signalling path- ways appear to be coupled to carbon, energy, and Pi metabolism. Accordingly, activation of PhoB by CreC and acetyl phosphate may both be indicative of cross-regula- tion. Cross-regulation may be especially important for the overall global control of cell growth and metabolism and for regulatory connections between different signalling pathways (Wanner, 1992; Wanner eta/., 1995). It has long been known that PhoB is activated by both PhoR and CreC (Wanner and Latterell, 1980). Further- more, using purified components, the cytoplasmic (soluble) domains of PhoR and CreC (denoted ’PhoR and ’CreC, respectively) were shown to be autophosphorylated by ATP and both phospho’PhoR and phospho’CreC were shown to transfer the phosphoryl group to PhoB in vitro (Makino et al., 1988; Amemura et a/., 1990). That PhoB is also activated by acetyl phosphate was revealed by the discovery that suppressor mutations that restored activation in the absence of PhoR and CreC affect acetyl phosphate metabolism (Wanner and Wilmes-Riesenberg, 1 992). During growth on glucose, acetyl phosphate is synthe- sized from acetyl CoA and Pi with the release of free CoA by phosphotransacetylase (Pta). Acetyl phosphate 1996 Blackwell Science Ltd
-
Upload
hms-harvard -
Category
Documents
-
view
1 -
download
0
Transcript of Involvement of the sensor kinase EnvZ in the in vivo activation of the response-regulator PhoB by...

Molecular Microbiology (1996) 22(1), 135-147
Involvement of the sensor kinase EnvZ in the in vivo activation of the response-regulator PhoB by acetyl phosphate
Soo-Ki Kim, Mary R. Wilmes-Hesenberg+ and Barry L. Wanner* Department of Biologics/ Sciences, Purdue University, West Lafayette, Indiana 47907, USA.
Summary
Three signalling pathways lead to activation of the phosphate (Pho) regulon by phosphorylation of the response-regulator PhoB in Eschefichia coli. One pathway responds to the extracellular inorganic phosphate (Pi) level and leads to activation by the Pi sensor kinase, PhoR. The other two pathways are Pi independent and are apparent in the absence of PhoR. One Pi-independent pathway responds to the level of an unknown catabolite and leads to activation by the catabolite regulatory sensor kinase, CreC (originally called PhoM); the other Pi-independent pathway responds to acetyl phosphate and leads to activation by a process requiring acetyl phosphate. Here we show that activation of PhoB by acetyl phosphate can require the sensor kinase EnvZ. Accordingly, we propose that the in vivo activation of PhoB by acetyl phosphate (and perhaps other two- component response-regulators as well) probably always requires a certain kinase that can vary depend- ing upon the growth conditions.
Introduction
The phosphate (Pho) regulons of Escherichia coliand Sal- monella typhimurium consist of at least 31 and 21 genes, respectively, that are co-regulated by inorganic phosphate (Pi; Jiang eta/., 1995; Wanner, 1996). These include a total of 38 different genes that are arranged in 11 transcriptional units. One unit, the pho6R operon, encodes the two- component regulatory system comprising the response- regulator PhoB and its partner sensor kinase, PhoR. Each Pi-regulated Pho regulon promoter contains one or more ‘Pho Box’ sequences (Makino eta/., 1988) required
Received 6 June, 1996; revised 25 July, 1996; accepted 2 August, 1996. +Present address: Department of Biology, Washington Univer- sity, St. Louis, Missouri 63130, USA. *For correspondence. E-mail BLWQbilbo.bio.purdue.edu; Tel. (317) 494 8034; Fax (317) 494 0876.
for regulation. The expression of most of these promoters is known to require phospho-PhoB for transcriptional acti- vation (Makino eta/., 1992; S.-K. Kim and B. L. Wanner, unpublished).
The primary signalling pathway for activation of PhoB responds to the environmental (extracellular) Pi level. This signalling pathway involves activation under conditions of Pi limitation and inhibition when Pi is in excess. Both of these processes require PhoR. Inhibition requires, in addi- tion, an intact, high-affinity Pi-specific transporter (the Pst system) and a protein called PhoU. Control by Pi is brought about via the phosphorylation of PhoB by PhoR when environmental Pi is limiting (less than about 4pM) and dephosphorylation of phospho-PhoB by PhoR (perhaps together with a Pst component(s) or PhoU) when Pi is in excess (greater than about 4 pM; for a review see Wanner, 1 996).
Two additional signalling pathways lead to activation of PhoB in the absence of PhoR. One requires the catabolite regulatory sensor kinase CreC (originally called PhoM), and the other requires acetyl phosphate (Wanner and Wilmes-Riesenberg, 1992). Both of these signalling path- ways appear to be coupled to carbon, energy, and Pi metabolism. Accordingly, activation of PhoB by CreC and acetyl phosphate may both be indicative of cross-regula- tion. Cross-regulation may be especially important for the overall global control of cell growth and metabolism and for regulatory connections between different signalling pathways (Wanner, 1992; Wanner eta/., 1995).
It has long been known that PhoB is activated by both PhoR and CreC (Wanner and Latterell, 1980). Further- more, using purified components, the cytoplasmic (soluble) domains of PhoR and CreC (denoted ’PhoR and ’CreC, respectively) were shown to be autophosphorylated by ATP and both phospho’PhoR and phospho’CreC were shown to transfer the phosphoryl group to PhoB in vitro (Makino et al., 1988; Amemura et a/., 1990). That PhoB is also activated by acetyl phosphate was revealed by the discovery that suppressor mutations that restored activation in the absence of PhoR and CreC affect acetyl phosphate metabolism (Wanner and Wilmes-Riesenberg, 1 992).
During growth on glucose, acetyl phosphate is synthe- sized from acetyl CoA and Pi with the release of free CoA by phosphotransacetylase (Pta). Acetyl phosphate
1996 Blackwell Science Ltd

136 S.-K. Kim, M. R. Wilmes-Riesenberg and B. L. Wanner
and ADP are then converted to acetate and ATP by acetate kinase (AckA). Several mutations that resulted in activation of PhoB in phoRcreC mutants were found to lie in ackA. An ackA mutation is expected to lead to elevated acetyl phosphate levels because of a block in its breakdown during growth on glucose (and on many other carbon sources). Other mutations abolished acti- vation caused by an ackA mutation. Several of these were found to lie in pta. A pfa mutation is expected to lead to lowered acetyl phosphate levels because of a block in its synthesis during growth on carbon sources other than acetate (as the Pta-AckA pathway is freely reversible in vivo). By testing the effects of ackA, pfa, and A(ackApfa) mutations on activation during growth on various carbon sources we showed that acetyl phos- phate is responsible for activation of PhoB in the absence of PhoR and CreC (Wanner and Wilmes-Riesenberg, 1992; see also Wanner, 1994a; 1995; Wanner ef a/., 1 995).
Accordingly, we suggested two potential mechanisms for activation of PhoB by acetyl phosphate: (i) indirect activation of PhoB by acetyl phosphate via a sensor kinase or (ii) direct activation of PhoB by acetyl phosphate binding or covalent modification (presumably phosphory- lation) of PhoB (Wanner, 1992; Wanner and Wilmes- Riesenberg, 1992). We examined the first possibility by searching for mutants lacking the putative ‘acetyl phos- phate sensor,’ but we found no evidence of a sensor kinase for acetyl phosphate (unpublished data cited in Wanner and Wilmes-Riesenberg, 1992).
Our previous studies identified many mutations that lead to regulatory effects on the Pho regulon in the absence of PhoR, in addition to ones in creC, ackA, and pta (Wanner,
1994a; Wanner and Wilmes-Riesenberg, 1992). In parti- cular, we discovered that a mutation of ompR (for the osmolarity response-regulator OmpR) leads to activation of PhoB in the absence of PhoR and CreC. We have now examined the basis for activation of PhoB because of ompR mutations. We found that under certain conditions both EnvZ and acetyl phosphate are required for activation of PhoB. We therefore propose that the in vivoactivation of PhoB by acetyl phosphate involves a sensor kinase(s) and that the specific activating conditions determine the identity of the kinase. These findings have important implications in the control of the Pho regulon and other regulatory sys- tems by acetyl phosphate.
Results
Effects of creC, ackA, and pta mutations on activation of PhoB
Bacterial alkaline phosphatase (Bap; the phoA gene product) is a convenient reporter of the Pho regulon, whose expression is inhibited when Pi is in excess. Conse- quently, the wild-type strain BW13711 synthesizes only small amounts of Bap under all conditions in the presence of excess Pi (Table 1). Small, but significant, differences in the basal levels of Bap are observed between cells grown on different carbon sources, however. Carbon- source effects are especially evident when these activities are compared with those of the otherwise isogenic AphoA mutant BW13745. The residual activities of the AphoA mutant are due to other non-specific phosphatases detect- able in our assay. The small amounts of Bap synthesis in BWl3711 during growth with excess Pi are a conse- quence of Pi inhibition. The same strain synthesizes
Table 1. Effects of phoR, creC, pta, ackA, ompR, and envZ mutations on activation of PhoB.
Bap Specific Activityb On: ’
Strain Relevant genotypea glucose pyruvate TYE TYEG
BW13711 1.5 3.2 1.6 1.1 BW13745 AphoA 0.7 0.5 0.9 NDC
353 46.3 364 BW13713 phoR 124 BW13736 phoR AcreABCD 2.3 95.4 1.0 231 BW16467 phoR A(ackA pfa) BW16470 phoR AcreABCD A(ackA pta) 0.7
BWl6469 phoR AcreABCD ackA 177 1120 70.5 519 BW11886 phoRAompR101 114 371 45.9 514 BW16558 phoR AcreABCD AompRlOl 89.4 251 35.8 544 BWl3973 phoR AcreABCD envZ22 0.5 129 ND ND BW13971 phoR AcreABCD A(ompR envZ)900 0.3 25.5 ND ND
a. All phoR mutants carry the ochre phoR68 allele. See the Experimenfal procedures for complete descriptions. b. Cells were grown on glucose or pyruvate MOPS agar with 2 mM Pi, TYE or TYEG agar for 16 to 20 h prior to assay of Bap activities as described in the Experimental procedures. All media have excess Pi. Specific activity units are nanomoles product made per min per cell optical den- sity (OD) at 420nm. Values are averages of three or more determinations. c. ND, not determined.
26.5 21 1 38.9 5.0 0.6 1.4 0.2
BW16546 phoR AcreABCD pfa 0.8 1.3 2.2 4.5
0 1996 Blackwell Science Ltd, Molecular Microbiology, 22, 135-147

In vivo activation of PhoB by acetyl phosphate 137
higher activation during growth on TYE (compare BW13713 with BW16467). Effects due to acetyl phosphate are much more evident in the absence of CreC, however. No activation is seen in the phoRcreC A(ackApta) mutant BW16470 under all conditions. Acetyl phosphate may also be responsible for low-level Bap synthesis in phoRcreC mutants during growth on glucose. Under these conditions, both the pta and A(ackApta) mutations further reduced Bap synthesis approx. threefold in phoR - creC mutants (compare BW16470 and BW16546 with BWl3736). The significance of these latter effects is unclear as the levels are already quite low.
about 280 units per cell OD420 during growth on glucose under conditions of Pi limitation (data not shown).
Pi inhibition of Pho regulon gene expression is abolished by null phoR mutations. The isogenic phoR mutant BW13713 synthesizes large amounts of Bap in the presence of excess Pi (Table 1). These levels are highly regulated by the carbon and energy source, however. In the absence of PhoR, activation of PhoB is due to CreC, acetyl phosphate, or both, depending upon the growth conditions. Activation by CreC is evident from compari- son of Bap levels in BW13713 with those in the phoR AcreABCD mutant, BW13736. The AcreABCD mutation reduced activation c. 50-fold during growth on glucose or tryptone-yeast extract (TYE), 3.7-fold during growth on pyruvate, and 1.6-fold during growth on TYE containing glucose (TYEG). Reduced activation under these condi- tions is due to loss of CreC because a nonsense creC allele has effects similar to the AcreABCD mutation (data not shown).
Evidence for activation of PhoB by acetyl phosphate is primarily supported by the contrasting effects of pta and ackA mutations when either occurs singly (Wanner and Wilmes-Riesenberg, 1992). A pta mutation prevents acti- vation during growth on glucose or pyruvate and leads to low, but significant, activation during growth on TYEG (compare BW16546 with BW13736, Table 1). During growth on these but not on all carbon sources (acetate is an exception), loss of Pta is expected to result in lowered acetyl phosphate levels due to a block in its synthesis. In contrast, an ackA mutation leads to high activation on these media (compare BW16469 with BWl3736). Under these conditions, loss of AckA is expected to lead to an accumulation of acetyl phosphate due to a block in its breakdown. It is notable that the levels of activation of PhoB due to acetyl phosphate are as high or higher than those due to PhoR under conditions of Pi limitation.
Furthermore, under certain conditions acetyl phosphate and CreC appear to act in concert, resulting in higher activation of PhoB when together than when alone. This is shown by the 4.7-fold higher activation in the CreC' phoR mutant BW13713 than in the CreC' phoR A(ackA pta) mutant BW16467 during growth on glucose (Table 1). As a consequence, BW13713 shows 175-fold higher activation of PhoB than the phoRcreC A(ackApta) mutant BW16470. During growth on glucose, CreC and acetyl phosphate can also act independently. CreC is seen to lead to c. 50- and 40-fold activation in CreC' phoR mutants that are capable and incapable of acetyl phosphate synthesis, respectively (compare BW13713 with BW13736 and BW16467 with BWl6470). In the pre- sence of CreC, acetyl phosphate is seen to lead to c. 4.7-fold higher activation during growth on glucose, 73- fold higher activation during growth on TYEG, 1.7-fold higher activation during growth on pyruvate, and 1.2-fold
0 1996 Blackwell Science Ltd, Molecular Micrd~iology, 22, 135-1 47
Effects of AompR1 01 , envZ, and A(ompR envZ)900 mutations on activation of PhoB
We previously identified numerous mutations that dramati- cally altered Bap synthesis in phoR mutants. In addition to ones in phoA, phoB, creC, ackA, and pta, we identified mutations lying in arcA (for aerobic respiratory control response-regulator), cyaA (for adenylate cyclase), crp (for catabolite activator protein), icd (for isocitrate dehy- drogenase), mdh (for malate dehydrogenase), ompR (for osmotic response-regulator), ops (for exopolysaccharide production), ppk (for polyphosphate kinase), ptsHl (for phosphohistidinoprotein or enzyme I), pur (for purine bio- synthesis), rpiA (for ribose phosphate isomerase), and other (unknown) genes. Although most of these mutations appear to affect activation of PhoB indirectly via CreC or acetyl phosphate, the precise mechanism(s) is unknown (Wanner et a/., 1988; Wanner and Wilmes-Riesenberg, 1992; Wanner, 1994a; M. Wilmes-Riesenberg and B. L. Wanner, unpublished).
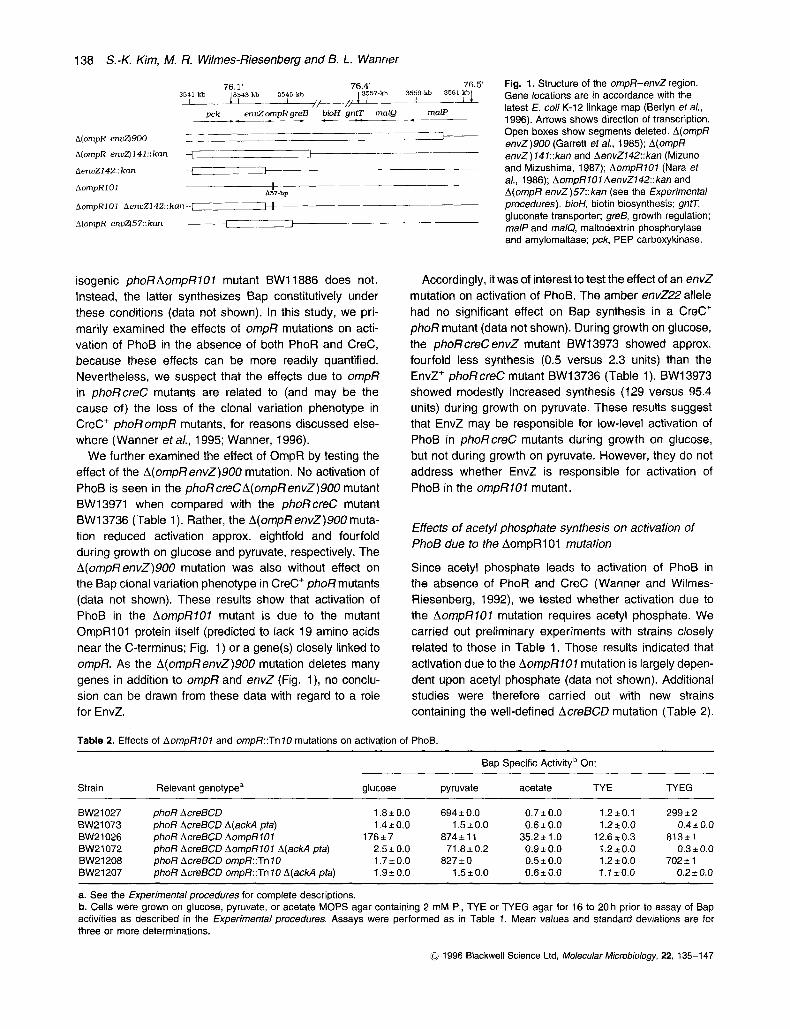
In order to determine how an ompR mutation leads to activation of PhoB, we examined the effect of the well- characterized AompR707 mutation. OmpR and EnvZ are encoded in the ompR-envZ (ompB) operon. The AompRlOl mutation is an in-frame deletion of 57 bp near the carboxyl terminus of OmpR (Fig. 1). It is therefore non-polar, conferring an OmpR-, EnvZ' phenotype. The AompRl07 mutation resulted in high activation of PhoB in the absence of PhoR and CreC. During growth on glucose, the phoRcreCAompRlO7 mutant BW16558 syn- thesized c. 40-fold more Bap than the phoR creC mutant BW13736 (Table 1). BW16558 synthesized c. 2.5-fold more Bap during growth on pyruvate, c. 35-fold more during growth on TYE, and c. 2-fold more during growth on TYEG. Determining the basis for these increased levels brought about by the AOmpR707 mutation is the subject of this study. In addition, we observed that the AompRlOl mutation abolishes the Bap clonal variation phenotype of a CreC' phoR mutant (Wanner, 1987). Whereas the phoR mutant BW13713 shows the Bap clonal variation phenotype during growth on TYE, the
13% S.-K. Kim, M. R. Wilmes-Riesenberg and 8. L. Wanner
76.4 76.5' 3557-kb 3559 kb 3561-kb
76.1' 3541-kb 3543-hb 3545-kb
I I A - 1 1 I I pck enuZompHgreB bioH gntT ma@
_c_t_4 t . .
_ - I AIornpR enua9OO _ _ A(ornpR enuZ)141::kan 4
AenuZ142:kun i -7 AompRlOl
AompR101 AenuZl42:: kan
A(ornpR enuZ)57:kan
n A57-bp
__ I E
isogenic phoRAompRlO1 mutant BW11886 does not. Instead, the latter synthesizes Bap constitutively under these conditions (data not shown). In this study, we pri- marily examined the effects of ompR mutations on acti- vation of PhoB in the absence of both PhoR and CreC, because these effects can be more readily quantified. Nevertheless, we suspect that the effects due to ompR in phoffcreC mutants are related to (and may be the cause of) the loss of the clonal variation phenotype in CreC' phoRompR mutants, for reasons discussed else- where (Wanner et a/., 1 995; Wanner, 1996).
We further examined the effect of OmpR by testing the effect of the A(ompRenvZ)900 mutation. No activation of PhoB is seen in the phoff creCA(ompRenvZ)900 mutant BW13971 when compared with the phoff creC mutant BW13736 (Table 1). Rather, the A(ompRenvZ)SOO muta- tion reduced activation approx. eightfold and fourfold during growth on glucose and pyruvate, respectively. The A(ompRenvZ)900 mutation was also without effect on the Bap clonal variation phenotype in CreC' phoff mutants (data not shown). These results show that activation of PhoB in the AompfflOl mutant is due to the mutant OmpRiOl protein itself (predicted to lack 19 amino acids near the C-terminus; Fig. 1) or a gene(s) closely linked to ompR. As the A(ompRenvZ)900 mutation deletes many genes in addition to ompff and envZ (Fig. l ) , no conclu- sion can be drawn from these data with regard to a role for EnvZ.
Fig. 1. Structure of the ompR-envZ region. Gene locations are in accordance with the latest E. coli K-12 linkage map (Berlyn et a/., 1996). Arrows shows direction of transcription. Open boxes show segments deleted. A(ompR envZ)900 (Garrett eta/., 1985); A(ompR envZ)f41::kan and AenvZf42::kan (Mizuno and Mizushima, 1987); AompRlOf (Nara et a/., 1986); AompR10f AenvZ142::kan and A(ompR envZ)57::kan (see the Experimental procedures). bioH, biotin biosynthesis; gntT, gluconate transporter: greB, growth regulation: malP and ma/Q, maltodextrin phosphorylase and amylomaltase; pck, PEP carboxykinase.
Table 2. Effects of AompR101 and ompR::TnfO mutations on activaion of PhoB
Accordingly, it was of interest to test the effect of an envZ mutation on activation of PhoB. The amber envZ22 allele had no significant effect on Bap synthesis in a CreC' phoR mutant (data not shown). During growth on glucose, the phoRcreC envZ mutant BW13973 showed approx. fourfold less synthesis (0.5 versus 2.3 units) than the EnvZ' phoRcreC mutant BW13736 (Table 1). BW13973 showed modestly increased synthesis (129 versus 95.4 units) during growth on pyruvate. These results suggest that EnvZ may be responsible for low-level activation of PhoB in phoRcreC mutants during growth on glucose, but not during growth on pyruvate. However, they do not address whether EnvZ is responsible for activation of PhoB in the ompRlO7 mutant.
Effects of acetyl phosphate synthesis on activation of PhoB due to the AompRlOl mutation
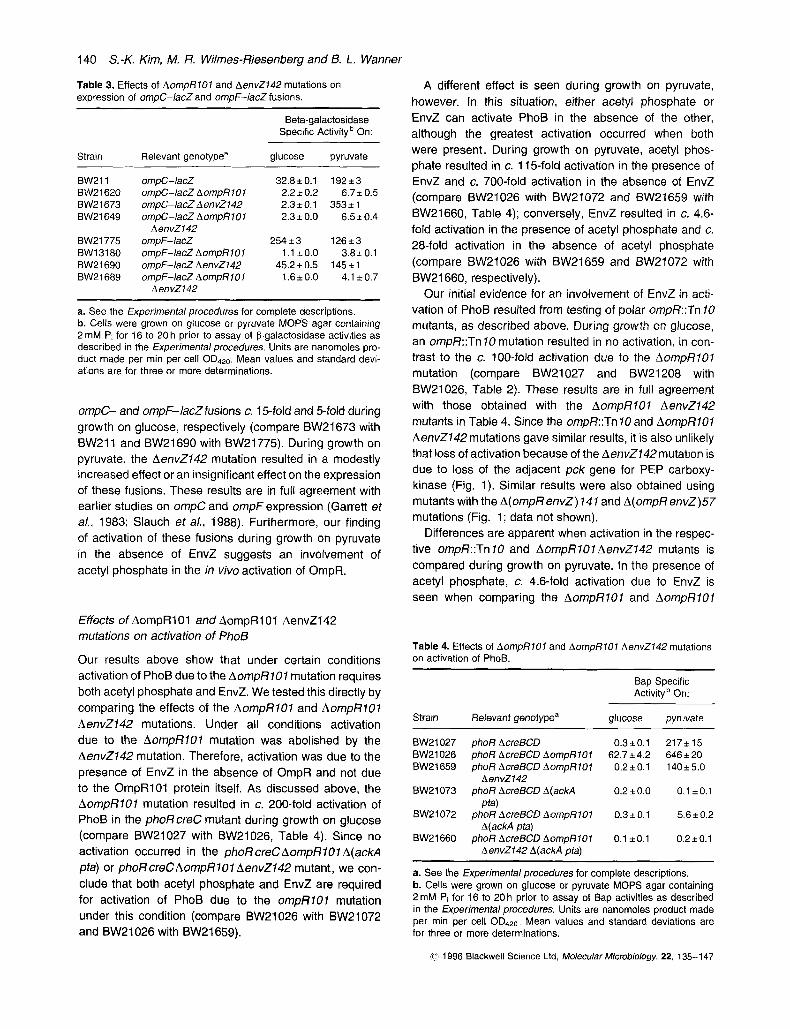
Since acetyl phosphate leads to activation of PhoB in the absence of PhoR and CreC (Wanner and Wilmes- Riesenberg, 1992), we tested whether activation due to the AompRlOl mutation requires acetyl phosphate. We carried out preliminary experiments with strains closely related to those in Table 1. Those results indicated that activation due to the AompRlOl mutation is largely depen- dent upon acetyl phosphate (data not shown). Additional studies were therefore carried out with new strains containing the well-defined AcreBCD mutation (Table 2).
Bap Specific Activityb On:
Strain Relevant genotypea glucose pyruvate acetate TY E TY EG
BW21027 phoR AcreBCD 1.8 f 0.0 694 f 0.0 0.7 5 0.0 1.2*0.1 299 f 2 BW21073 phoR AcreBCD A(ackA pta) 14fO.O 1.5 5 0.0 0.6 f 0.0 1.21 0.0 0.4 i 0.0 BW21026 phoR AcreBCD AompRlOl 176 i7 874f11 35.2f 1 .O 12.6f0.3 813*1 0W21072 phoR AcreBCD AompRIOf A(ackA pfa) 2.5fO.O 71.8 & 0.2 0.9f 0.0 1.2*0.0 0.3iO.O BW21208 phoR AcreBCD ompR::Tn 10 1.7fO.O 827fO 0.5 i 0.0 1.2 f 0.0 702 f 1 BW21207 phoR AcreBCD ompR::Tn 10 A(acW1 pta) 1.9 f 0.0 1.5 f 0.0 0.6iO.O 1.1 *o.o 0.210.0
a. See the Experimental procedures for complete descriptions. b. Cells were grown on glucose, pyruvate, or acetate MOPS agar containing 2 mM P,, TYE or TYEG agar for 16 to 20 h prior to assay of Bap activities as described in the Experimental procedures. Assays were performed as in Table 1. Mean values and standard deviations are for three or more determinations.
0 1996 Blackwell Science Ltd, Molecular Microbiology, 22, 135-1 47

In vivo activation of Ph05 by acetyl phosphate 139
conditions activation of PhoB in phoRcreCAompR707 mutants requires acetyl phosphate. In addition, no acti- vation occurred because of the A(ompRenvZ) mutation (Table 1). Therefore, a gene in the vicinity of the ompR locus is partly responsible for activation of PhoB in AompR707 mutants. In order to determine whether this gene is envZ, we examined a number of other ompR and envZ mutations. Unlike the AompR707 mutation (which is OmpR- EnvZ'), an ompR::Tn 70 mutation is OmpR- and EnvZ- because of polarity. As shown in Table 2, no activation resulted because of the ompR::Tn 70 mutation. Both the phoRcreC mutant BW21027 and the phoR creCompR::Tn 70 mutant BW21208 synthesized similar amounts of Bap during growth on all media except TYEG, on which a 2.3-fold increase was observed. The latter condition is not particularly relevant to this study as this effect was entirely due to acetyl phosphate (compare BW21207 with BW21208).
Therefore activation of PhoB because of the AompR701 mutation may be due to the OmpRl 01 protein or to EnvZ. During growth on glucose, acetate, and TYE, activation due to the AompR707 mutation also requires acetyl phosphate, however. To clarify these regulatory effects, we tested the influence of the ompR337::Tn 70, A(ompR envZ)57, A(ompRenvZ) 747, and AenvZ742 mutations (Fig. 1) on activation of PhoB under these growth condi- tions in both phoRcreCand phoRcreCA(ackApta) mutants. Under all conditions, the ompR337::Tn 70, (ompRenvZ)57, and A(ompRenvZ) 747 mutants synthesized amounts of Bap similar to those of the corresponding ompR::Tn70 mutants in Table 2 (data not shown). The AenvZ742 mutants synthesized amounts similar to those of the envZ22 mutants above (Table 1 and data not shown). These results reinforce our conclusion that the OmpRlO1 protein or EnvZ is responsible for activation of PhoB resulting from the AompR107 mutation.
As expected, the phoR creC mutant BW21027 synthe- sized small amounts of Bap during growth on glucose (1.8 units), acetate (0.7 units), and TYE (1.2 units), while it synthesized large amounts during growth on pyruvate (694 units) and TYEG (299 units): the phoRcreCA(ack4 pta) mutant BW21073 synthesized small amounts under all conditions. The regulatory effects on Bap synthesis in these OmpR' phoRcreC mutants are attributable entirely to acetyl phosphate; the mutant BW21027 is expected to synthesize small amounts of acetyl phosphate during growth on glucose, acetate, or TYE and to synthesize large amounts of acetyl phosphate during growth on pyru- vate or TYEG (Wanner and Wilmes-Riesenberg, 1992).
Importantly, comparison of the phoRcreCAompR707 mutant BW21026 and the OmpR' phoRcreC mutant BW21027 revealed that the AompR707 mutation resulted in c. 100-fold activation during growth on glucose, c. 50- fold activation during growth on acetate, c. 10-fold acti- vation during growth on TYE, and c. 2.7-fold activation during growth on TYEG. No significant effect was appa- rent during growth on pyruvate, probably because high activation had already occurred in the OmpR' phoRcreC mutant.
Several experiments showed that activation of PhoB caused by the AompR707 mutation is largely, but not entirely, due to acetyl phosphate. As mentioned above, activation of PhoB is primarily attributable to the AompR707 mutation under three conditions: during growth on glucose, acetate, and TYE (compare BW21026 with BW21027, Table 2). Under these conditions activation in AompR707 mutants is also almost entirely due to acetyl phosphate. In AompR707 mutants, the A(ackApta) muta- tion reduced activation c. 70-fold (2.5 versus 176 units) during growth on glucose, c. 40-fold (0.9 versus 35.2 units) during growth on acetate, and c. 10-fold (1.2 versus 12.6units) during growth on TYE (compare BW21072 with BW21026). During growth of AompR707 mutants on pyruvate, the A(ackApta) mutation reduced activation only 12-fold (71.8 versus 874 units), however. This is the only case in which substantial (c. 50-fold) activation of PhoB was observed in the absence of PhoR, CreC, and acetyl phosphate (compare BW21072 with BW21073). We show later that under certain conditions activation of PhoB due to the AompR707 mutation requires both acetyl phosphate and EnvZ and that under other conditions activation due to the AompR707 mutation requires only EnvZ. The residual activation of PhoB in the phoRcreC AompR707 A(ackApta) mutant during growth on pyruvate is due to EnvZ (see below).
Effects of other ompR and envZ mutations on activation of PhoB
The above results show that under some, but not all,
0 1996 Blackwell Science Ltd, Molecular Microbio/ogy, 22, 135-147
Effects of AompR101, AenvZ142, and AompR1 01 AenvZl42 mutations on ompC and ompF expression
We constructed double mutants carrying both the AompR707 and AenvZ742 mutations together in order to determine whether EnvZ is required for activation of PhoB due to the ompR707 mutation. We verified these mutations by testing for regulatory effects on expression of ompC- and ompf-lacZ transcriptional fusions (Hall and Silhavy, 1981). Both the single AompR707 mutation and the double AompR707 AenvZ742 mutation reduced expression of the ompC- and ompF-laczfusions c. 15- fold and c. 230-fold during growth on glucose, respec- tively, and c. 30-fold during growth on pyruvate (Table 3; compare BW21620 and BW21649 with BW211; and compare BW13180 and BW21689 with BW21775). The AenvZ742 mutation alone reduced expression of the
140 S.-K. Kim, M. R. Wilmes-Riesenberg and B. L. Wanner
Table 3. Effects of AompRlOl and AenvZ142 mutations on expression of ompC-/acZ and ompf-lacZ fusions.
Strain
BW211 BW21620 BW21673 BW21649
BW21775 BW13180 BW21690 BW21689
Relevant genotypea
ompC-lacZ ompC-IacZ AompR101 ompC-lacZ AenvZ142 ompC-lacZ AompRlOl
ompf-lacZ ompf-lacZ AompR101 ompf-lac2 AenvZl42 ompf-lacZ AompR101
AenvZ142
AenvZ142
Beta-galactosidase Specific Activityb On:
glucose pyruvate
32.8k0.1 192+3 2.2k0.2 6.7~t0.5 2.3r0.1 353k1 2.3 f 0.0 6.5 f 0.4
254*3 126k3
145 i 1 1.1 ~ t 0 . 0 3.8i0.1
45.2 ~t 0.5 1.6 r 0.0 4.1 f 0.7
a. See the Experimental procedures for complete descriptions. b. Cells were grown on glucose or pyruvate MOPS agar containing 2 mM P, for 16 to 20 h prior to assay of 0-galactosidase activities as described in the Experimental procedures. Units are nanomoles pro- duct made per min per cell OD420. Mean values and standard devi- ations are for three or more determinations.
ompC- and ompf-IacZfusions c. 15-fold and 5-fold during growth on glucose, respectively (compare BW21673 with BW211 and BW21690 with BW21775). During growth on pyruvate, the AenvZ742 mutation resulted in a modestly increased effect or an insignificant effect on the expression of these fusions. These results are in full agreement with earlier studies on ompC and ompF expression (Garrett et a/., 1983; Slauch et a/., 1988). Furthermore, our finding of activation of these fusions during growth on pyruvate in the absence of EnvZ suggests an involvement of acetyl phosphate in the in vivo activation of OmpR.
Effects of AompR101 and AompRlOl AenvZl42 mutations on activation of PhoB
Our results above show that under certain conditions activation of PhoB due to the AompRIO1 mutation requires both acetyl phosphate and EnvZ. We tested this directly by comparing the effects of the AompR101 and AompRlOl AenvZ142 mutations. Under all conditions activation due to the AompRIO1 mutation was abolished by the AenvZ142 mutation. Therefore, activation was due to the presence of EnvZ in the absence of OmpR and not due to the OmpR101 protein itself. As discussed above, the AompR101 mutation resulted in c. 200-fold activation of PhoB in the phoRcreC mutant during growth on glucose (compare BW21027 with BW21026, Table 4). Since no activation occurred in the phoRcreCAompR101 A(ackA pta) or phoRcreCAompR107 AenvZ142 mutant, we con- clude that both acetyl phosphate and EnvZ are required for activation of PhoB due to the ompRlOl mutation under this condition (compare BW21026 with BW21072 and BW21026 with BW21659).
A different effect is seen during growth on pyruvate, however. In this situation, either acetyl phosphate or EnvZ can activate PhoB in the absence of the other, although the greatest activation occurred when both were present. During growth on pyruvate, acetyl phos- phate resulted in c. 115-fold activation in the presence of EnvZ and c. 700-fold activation in the absence of EnvZ (compare BW21026 with BW21072 and BW21659 with BW21660, Table 4); conversely, EnvZ resulted in c. 4.6- fold activation in the presence of acetyl phosphate and c. 28-fold activation in the absence of acetyl phosphate (compare BW21026 with BW21659 and BW21072 with BW21660, respectively).
Our initial evidence for an involvement of EnvZ in acti- vation of PhoB resulted from testing of polar ompR::Tn 10 mutants, as described above. During growth on glucose, an ompR::Tn 10 mutation resulted in no activation, in con- trast to the c. 100-fold activation due to the AompR101 mutation (compare BW21027 and BW21208 with BW21026, Table 2). These results are in full agreement with those obtained with the AompR101 AenvZ142 mutants in Table 4. Since the ompR::Tn 10 and AompR101 AenvZ142 mutations gave similar results, it is also unlikely that loss of activation because of the AenvZ742mutation is due to loss of the adjacent pck gene for PEP carboxy- kinase (Fig. 1). Similar results were also obtained using mutants with the A(ompRenvZ) 141 and A(ompRenvZ)57 mutations (Fig. 1; data not shown).
Differences are apparent when activation in the respec- tive ompR::Tn 10 and AompR101 AenvZ142 mutants is compared during growth on pyruvate. In the presence of acetyl phosphate, c. 4.6-fold activation due to EnvZ is seen when comparing the AompR101 and AompR101
Table 4. Effects of AompRlOl and AompR101 AenvZ142 mutations on activation of PhoB.
Bap Specific Activityb On:
Strain Relevant genotypea glucose pyruvate
BW21027 BW21026 BW21659
BW21073
BW21072
BW21660
phoR AcreBCD 0.3k0.1 217k15 phoR AcreBCD AompRIOI 62.7k4.2 646*20 phoR AcreBCD AompR101 0.2r0.1 140i5.0
AenvZ142 phoR AcreBCD A(ackA 0.2Zto.o 0.1 *O.l
Pta) phoR AcreBCD AompRIO1 0.3k0.1 5.6k0.2
phoR AcreBCD AompR101 0.1 i O . 1 0.2k0.1 A(ackA pta)
AenvZ142 A(ackA pta)
a. See the Experimental procedures for complete descriptions. b. Cells were grown on glucose or pyruvate MOPS agar containing 2 mM P, for 16 to 20 h prior to assay of Bap activities as described in the Experimenfal procedures. Units are nanomoles product made per min per cell OD420. Mean values and standard deviations are for three or more determinations.
0 1996 Blackwell Science Ltd, Molecular Microbiology, 22, 135-147

In vivo activation of PhoB by acetyl phosphate 141
regulator by acetyl phosphate is due to the cognate kinase. This would account for the apparent stimulation, because of acetyl phosphate, of activation of PhoB by PhoR and CreC (see below).
Acetyl phosphate appears to be involved in activation of PhoB in both the presence and the absence of PhoR and CreC. It is also clear that activation by PhoR, by CreC, and by acetyl phosphate are largely independent of each other (Wanner and Wilmes-Riesenberg, 1992; Wanner, 1993; 1994a; 1995; Wanner, 1996). PhoR, CreC, or acetyl phos- phate alone result in 400- to 2000-fold activation of PhoB under various growth conditions. Acetyl phosphate appears to be stimulatory in both the presence and the absence of PhoR and CreC, however. Acetyl phosphate leads to approx. threefold higher activation of PhoB by PhoR under conditions of Pi limitation (A. Haldimann and B. L. Wanner, unpublished), approx. fivefold higher activation by CreC during growth on glucose, and approx. 160-fold higher activation in the absence of both PhoR and CreC during growth on pyruvate (Wanner and Wilmes-Riesenberg, 1992; this paper). In accordance with our hypothesis, we propose that, when present, activation of PhoB by PhoR and CreC is stimulated by acetyl phosphate and that when these kinases are absent activation of PhoB by acetyl phosphate is due to a different sensor kinase(s). Furthermore, multiple kinases are probably capable of activating PhoB by acetyl phosphate under different condi- tions. Accordingly, activation of PhoB by acetyl phosphate and an additional (unknown) kinase(s) may account for the substantial activation of PhoB due to acetyl phosphate in the absence of PhoR, CreC, and EnvZ during growth on pyruvate (Table 4).
Our most compelling evidence for the involvement of a kinase in the in vivoactivation of PhoB by acetyl phosphate resulted from the study of AompRlOl mutants. The AompR707 mutation resulted in high activation of PhoB in phoRcreC mutants. Furthermore, these effects are attributable to EnvZ because they were abolished in AompRlOl AenvZ742 mutants. Apparently, EnvZ is cap- able of activating PhoB because of cross-talk (or cross- regulation) in the absence of its partner, OmpR. This is functionally similar to the activation of a response-regula- tor by a non-partner kinase in the absence of the cognate kinase. It is also fundamentally different. For example, high activation of PhoB by CreC occurs in phoRmutants in both the presence and the absence of the CreC partner response-regulator, CreB (Wanner ef a/., 1995). In con- trast, high activation of PhoB by EnvZ was observed only in the absence of both PhoR and OmpR. This is the first case in which the activation of a response-regulator by a non-partner kinase resulted from loss of the non- partner response-regulator. This may be a general phenom- enon having important implications for other regulatory systems as well.
AenvZl42 mutants in Table 4. No effect due to EnvZ is apparent when comparing the AompR101 and ompR:: TnlO mutants in Table 2, however. Instead, in Table 2 the same high activation resulted in AompRlOf and ompR::Tn 10 mutants in the presence of acetyl phosphate. These apparently conflicting results are reconciled if the ompR::Tn 10 mutation is not completely polar. A polarity effect can reduce EnvZ synthesis to such a level that no activation resulted during growth on glucose in the pre- sence of acetyl phosphate or on pyruvate (or TYEG) in the absence of acetyl phosphate while allowing sufficient EnvZ synthesis so that a stimulatory effect was observed during growth on pyruvate (or TYEG) in the presence of acetyl phosphate. Although we cannot rule out the possibi- lity that lowered activation in AompR707 AenvZ142 mutants during growth on pyruvate is due to loss of PEP carboxykinase, it seems more likely that EnvZ would also be involved in activation under this condition because EnvZ is clearly involved during growth on glucose.
Effects of ompC and ompF mutations on activation of
We considered that the AompR107 mutation may indirectly lead to activation of PhoB by abolishing expression of the porin regulon. The OmpC and OmpF porins are required for entry of metabolites into the periplasm (Pratt and Silhavy, 1995); they may also be required for leakage of cellular metabolites from the cell. Therefore, in the absence of OmpC and OmpF, mixed acids produced from glucose may accumulate in the periplasm, leading in turn to acti- vation of EnvZ, which is known to respond to a variety of signals including acidic conditions (Heyde and Portalier, 1987). To rule out this possibility, we examined the effects of ompC and ompFmutations. Under various growth con- ditions neither an ompC nor an ompF mutation alone or in combination affected activation of PhoB in otherwise iso- genic phoRcreC mutants (data not shown). Therefore the effects observed in this study are unlikely to be indirect ones caused by loss of OmpC or OmpF.
Ph05
Discussion
Our results lead us to conclude that under certain conditions, activation of PhoB by acetyl phosphate also requires EnvZ. We therefore propose that the in vivo activation of PhoB by acetyl phosphate, and perhaps by other response-regulators as well, is likely to involve a different kinase(s) under different conditions. It is especi- ally notable that conditions leading to activation of PhoB by acetyl phosphate and EnvZ, in particular growth on glu- cose or acetate, are conditions in which acetyl phosphate levels are expected to be low. In addition, we propose that under certain conditions, activation of a response-
0 1996 Blackwell Science Ltd, Molecular Microbio/ogy, 22, 135-147

142 S.-K. Kim, M. R. Wilmes-Riesenberg and B. L. Wanner
Our earlier finding that acetyl phosphate leads to activation of PhoB indicated that acetyl phosphate leads to the phosphorylation of PhoB (Wanner and Wilmes- Riesenberg, 1992) since phosphorylation is the only known mechanism of PhoB activation. Whether activation by acetyl phosphate was indirect or direct remained unresolved as we were unable to find mutants lacking the putative 'acetyl phosphate sensor.' The existence of multiple kinases capable of activation of PhoB by acetyl phosphate now explains why our earlier attempts to find mutants lacking the 'acetyl phosphate sensor' were unsuc- cessful (Wanner and Wilmes-Riesenberg, 1992); see also
Condition EEstp g:ayi Proposed Mechanism
ATP ADP EnvZ'RaAckA 0.3 5.6 E n v Z u E n v Z P PhoB@
E n v Z q P h o B
A c e Ac EnvZ A E n v Z
E~~vZ+l'ta+AckA+ 62.7 646 Or EnvZ A c e Ac
PhoB *a A c e Ac
Kinase X(?) EnvZ- Ra'AckA' 0.2 140 PhoB L P h o B a
(Wanner, 1992; 1994a; Wanner et a/., 1995). It was subsequently shown that acetyl phosphate can
act as a chemical phosphorylating agent, directly activat- ing the response-regulator CheY (Lukat et a/., 1992) as
1992; Deretic eta/., 1992; Holman et a/., 1994), including
Fig. 2. Mechanisms for activation of PhoB by EnvZ, acetyl phosphate, and Kinase X. All conditions are for activation of PhoB in AompR101 mutants in the absence of PhoR and CreC. The EnvZ', Pta-, Ackp- condition refers to the phoR creC AompRIOI A(ackA Pta) mutant BW21072; the EnvZ+, Pta+, AckA' condition
EnvZ-, Pta', AckA+ condition refers to the phoR creC AompRlOl AenvZ142 mutant BW21659. Bap sclecific activities are from Table
well as many other response-regulators (Feng et a/., refers to the phofl creCAompR'O1 mutant Bw21026; and the
OmpR and PhoB (McCleary and Stock, 1994; Hiratsu et a/., 1995). While these experiments provided new infor- mation on the catalytic mechanism of response-regulator phosphorylation (Stock et a/., 1995), it remained unclear as to how acetyl phosphate acts in vivo. As discussed else- where (Wanner eta/., 1995; Wanner, 1996), the concen- trations of acetyl phosphate required for efficient in vitro phosphorylation of response-regulators (e.g., PhoB has an estimated K, of lOmM or more for acetyl phosphate; see also McCleary, 1996) far exceed those attainable in vivo, which, according to the strain and growth condition, range from c. 20pM to 1 mM (Hunt and Hong, 1980; Hunt, 1980; Bochner and Ames, 1982; Pruss and Wolfe, 1994; McCleary and Stock, 1994). The apparent rates of direct response-regulator phosphorylation by acetyl phosphate are also extremely slow compared to those achieved by the cognate sensor kinases. In contrast, the in vivo activation of PhoB by acetyl phosphate (which is expected to reflect in vivo phosphorylation rates in log-phase cells) is similar in magnitude to activation of PhoB by PhoR (Wanner and Wilmes-Riesenberg, 1992; Wanner, 1995; 1996; Wanner et a/., 1995).
Biochemical mechanisms explaining how EnvZ and acetyl phosphate lead to activation of PhoB under various conditions are shown in Fig. 2. Under the EnvZ', Pta-, AckA- condition, c. 19-fold activation of Pt-108 by EnvZ alone resulted during growth on pyruvate (relative to values during growth on glucose). Under this condition, EnvZ is apparently autophosphorylated by ATP in response to a signal emanating from growth on pyruvate and phospho-EnvZ then subsequently transfers the phosphoryl group to PhoB. Whether this activation of PhoB by EnvZ also occurs in the presence of OmpR or is of physiological relevance remains to be examined. These results also lead to the prediction that pyruvate may cause activation of OmpR by EnvZ. Curiously, both
4. Circled P refers to the phosphoryl group on acetyl phosphate (Ac-P), phospho-EnvZ (EnvZ-P), or phospho-PhoB (PhoB-P). Ac, acetate. The proposed mechanisms are described in the text.
the ompC- and ompf-/acZ fusions are expressed at high levels in OmpR', EnvZ' strains during grow..h on pyruvate (Table 3). How much of this activation is cue to EnvZ, acetyl phosphate, or another factor(s) rema ns to be determined.
Under the EnvZ', Pta', AckA+ condition, both EnvZ and acetyl phosphate are required for high activation of PhoB. During growth on glucose, c. 200-fold activation resulted. This activation is entirely dependent upon both acetyl phosphate and EnvZ because it is abolished when either is absent. During growth on pyruvate, greater than 1000- fold activation resulted from acetyl phosphate and EnvZ, as compared to conditions in which both are absent (Table 4). While this activation appears to be mainly due to the concerted action of acetyl phosphate and EnvZ, substantial activation resulted in the presence of EnvZ or acetyl phosphate alone. Under this condition, activation was reduced c. 115-fold in the absence of acetyl phosphate (EnvZ', Pta-, AckA- condition) and c. 4.6- fold in the absence of EnvZ (EnvZ-, Pta+, AckA' condition).
Two different mechanisms of how acetyl phosphate and EnvZ may act together are illustrated in Fig. 2. In the first, EnvZ is shown to be autophosphorylated by acetyl phosphate and then phospho-EnvZ is shown to transfer the phosphoryl group to PhoB. In the second, EnvZ is shown to catalyse the autophosphorylation of PhoB by acetyl phosphate. Several other possibilities also exist. For example, acetyl phosphate may be an effector mol- ecule that stimulates the autophosphorylation of EnvZ by ATP or that stimulates the phosphotransfer reaction from phospho-EnvZ to PhoB (not shown).
0 1996 Blackwell Science U d , Molecular Mjcrobiobgy, 22, 135-147

In vivo activation of PhoB by acety/ phosphate 143
described elsewhere (Wanner, 1994b). Carbon sources were added at 0.2% for glucose and 1.0% for potassium acetate and pyruvate. TYE contained l o g of tryptone (Difco), 5 g of yeast extract (Difco), and 8 g of NaCl per litre. Glucose (1%) was added after autoclaving for preparation of TYEG. Agar media contained 1.5% Bacto agar (Difco). Xgal (Bachem) was used at 40 pg ml-' for detection of p-galactosidase. Anti- biotics were added at 12.5 pg ,I-' for chloramphenicol, 50pg ml-' for kanamycin, and 12.5pg ml-' for tetracycline.
Under the EnvZ-, Pta', AckA' condition, c. 700-fold activation of PhoB by acetyl phosphate resulted during growth on pyruvate. As mentioned above, we propose that under this condition activation by acetyl phosphate is due to an unknown kinase(s), denoted previously as X (Wanner, 1992; Wanner and Wilmes-Riesenberg, 1992). Accordingly, the mechanism of activation of PhoB by acetyl phosphate due to KinaseXwould be the same as for acetyl phosphate with EnvZ.
Our finding that activation of PhoB by acetyl phosphate involves a sensor kinase(s) provides further support for the hypothesis that acetyl phosphate acts as an important effector molecule in two-component and other regulatory systems, as previously proposed by ourselves (Wanner, 1992; Wanner and Wilmes-Riesenberg, 1992) and others (McCleary and Stock, 1994; Nystrom, 1994; Pruss and Wolfe, 1994; Shin and Park, 1995). Acetyl phosphate may activate response-regulators by means of cognate or non-cognate kinases in several situations. Acetyl phos- phate may be responsible for basal level activation of PhoB when Pi is in excess (Table 1) or for high activation of PhoB by PhoR under conditions of Pi limitation (see above). Acetyl phosphate may also be responsible for the carbon-source effects on activation of OmpR by EnvZ (Table 3). Interestingly, activation of OmpR occurs in response to many signals in addition to changes in osmolarity (Pratt and Silhavy, 1995; Pratt et a/., 1996). Acetyl phosphate may lead to activation of OmpR by EnvZ in response to a subset of these signals. Further- more, osmolarity changes lead to activation of OmpR even in the absence of EnvZ (Forst eta/., 1988). Accord- ingly, acetyl phosphate may be involved in osmoregu- lation, leading to activation of OmpR by EnvZ when present and by a different kinase in the absence of EnvZ.
Acetyl phosphate may also be involved in the activation of the VanR-Vans two-component regulatory system controlling inducible vancomycin resistance in Entero- coccus faecium BM4147 (Arthur eta/., 1995) in response to vancomycin (Arthur et a/., 1992), and other cell-wall inhibitors (Handwerger and Kolokathis, 1990; Allen and Hobbs, 1995). In this case, induced vancomycin resis- tance occurs even in the absence of the sensor kinase Vans (Arthur et a/., 1992). Therefore, acetyl phosphate may have a role in the activation of the response-regulator VanR by Vans in its presence and the activation of VanR by a different kinase in the absence of Vans. If so, inhibi- tion of acetyl phosphate synthesis may provide a new target for antimicrobial chemotherapy.
Experimental procedures
Media
Defined media were based on MOPS and prepared as
(C) 1996 Blackwell Science Ltd, Molecular Microbiology, 22, 135-147
Strains
All strains assayed are fully described in Table 5. AT141 (MC4100 AfompR envZ) 147::kan) and AT142 (MC4100 AenvZ742::kan) were from S. Mizushima; CAG12094 (zcb- 3059::Tn 70) and CAG12177 (zeh-298::Tn 70) were from C. Gross via M. Levinthal; DC846 (ornpR337::Tn 70) was from D. Clark; ECD57 (MC4100 recD7903::Tn70-77 A(ompR envZ)57::kan) was from M. Igo; FD500 (nuoG::Tn70 purf) was from F. Dailey; L1022 (MC4100 ma/Q ompC-/acZ,o-22 (hpl[209])) and L1025 (MC4100 ma/Q o m p C - / a ~ Z ~ ~ - ~ ~ ~ (hpl[209])) were from M. Hall; LP64 (MC4100 ompR::Tn 70) was from L. Pratt; MH513 (MC4100 ompf-/acZ16-13 (hpl[209])) and SG477 (MC4100 envZ22(Am)) were from S. Garrett; MHI 160 (MC4100 ompR707) and SG480 (MC4100 A(envZ ompR bioH gntM ma/QP)900::neo) were from J. Slauch; and TTlO916 (Sa/mone//a typhirnurium LT2 F' 128 lacY476::Tn 70d-CMlAproSA47) was from L. Csonka. Other bacteria and phages are from the laboratory collection. Strains used to test for effects of the ornpR337::Tn70, (ompR envZ)57::kan, A(ompR envZ) 747::kan, AenvZ742:: kan, ompC, and ompf mutations included: BW21280, BW21374 to BW21378, BW21415, BW21416, BW21476 to BW21478, BW22180, and BW22209 (data not shown). Trans- ductions were carried out using P1 kc.
Construction of AompRlOl AenvZ142 mutations
An ompRompC-lacZor ompRompf-IacZstrain is Lac- during growth on glucose or pyruvate, an envZompC-laczstrain is Lac- during growth on glucose and Lac+ on pyruvate, and an envZompf-lacZstrain is Lac+ during growth on glucose or pyruvate (S.-K. Kim and B. L. Wanner, unpublished). There- fore we expected AompR707 AenvZ742and OmpR' AenvZ142 transductants of an ompC or ompf-laczfusion strain to Lac- and Lac' on pyruvate, respectively. Accordingly, BW21620 (Table 5) was infected with a P1 lysate of AT142 and kana- mycin-resistant (KanR) transductants were selected on pyru- vate MOPS agar containing 2 mM Pi and Xgal. Among about 900 transductants, 3 were white and the remainder were blue. Less than 1% of the transductants were expected to be AompR707 AenvZ742::kan recombinants because of the close linkage of these mutations. One white transductant was named BW21649.
Construction of A(creABCD)l14/104::Tn5-112 and AcreBCD153 mutants
The A(creABCD) 7 74/704::Tn5-7 72 mutation is deleted of sequences between the creA 7 74::Tn5 and creD104::TnS

144 S.-K. Kim, M. R. Wilmes-Riesenberg and 6. L. Wanner
Table 5. Bacterial strains used in this work.
Straina Genotype Pedigreeb Source'
BW211
BW10469 BWl1886 BW13180 BW 13971
BW13973 BW16467 BW16558
BW21026
BW21027
BW21072
BW21073
BW21207 BW21208 BW21620
BW21649
BW21659 BW21660
BW21673
BW21689
BW21690
BW21775
Alac-169 ompC-lacZ10.22(hp1[209]) rpsL150 relAl araD139 thi? flbB5301
Alac-169 phoR68 A(pck envZ ompR gre6 bioH gntT malQP)900::neo Alac-169 phoR68 AompR101 Alac-169 phoR68 AompRlOl ompF-la~Z~~.~~(hp1[209]lacY476::Tn 10dCM) Alac-169 phoR68 A(creABCD) 114/104::Tn5 132 A(pck envZ ompR greB
Alac - 169 phoR68 AcreA6CD 1 f4/ 104::Tn5- 132 envZ22
Alac- 169 AompR 10 1 phoR68 AcreA BCD I 14/ 104::Tns f 12
Alac-169 AompR101 phoR68 AcreBCD153
Alac-169 phoR68 AcreBCD 153
Alac-169 AompR101 phoR68 AcreBCD153 A(pta ackA h i d hiSP)~,~,16
deoCl ptsF25
bioH gntT malQP )900::neo
DE3(lac)X74 ph0R68 A(pta acW\ hisQ hiSP)~A3516 zej-223::Tn 10
Alac-169 phoU68 AcreBCD153 A(pta ackA hisQ hiSP)~~3516
Alac-169 phoU68 AcreBCD153 ompR::Tn 10 A(pta ackA his0 h k P ) ~ ~ 3 5 1 6
Alac-169 phoR68 ompR::Tn10 AcreBCD153 Alac-169 ompC-lacZio.22(hp1[209]) AompRlOI rpsL150 relAl araD139 thi?
Alac- 169 ompClacZi
Alac- 169 phoR68 AompR101 AenvZ142::kan Alac-169 phoR68 AcreBCD153 A(pta ackA hisQ h is f )TA35,6 AompRlOl
Alac-169 ompC-lacZio.22(hpl [209]) AenvZ142::kan rpsL 150 relA I araD139
Alac-169 phoR68 AompRlOl AenvZ142::kan o m p F - / a ~ Z ~ ~ . ~ ~
Alac- 169 phoR68 AenvZ142:: kan ompf-lacZ, 6.1
Alac-169 phoR68 o m ~ F l a c Z ~ ~ . ~ ~ (hpl[209]lacY476::Tn 10dCM)
flbB5301 deoCl ptsF25
araD139 thi? flb65301 deoCl ptsF25 (hpl[209]) AompR 101 AenvZ142:: kan rpsL 750 relA 1
AenvZ142:: kan
thi? flbB5301 deoCl ptsF25
(hpl[209]lacY476::Tn 10dCM)
(hpl[209]lacY476::Tn 10dCM)
L1022
ED792 via BW3627 BD792 via BW10469 BD792 via BW11886 BD792 via BW10469
BD792 via BW12131 BD792 via BW13713 BD792 via BW13616
BD792 via
BD792 via
BD792 via
BD792 via
BD792 via BW21073 BD792 via BW21027 L1022
BW13727d
BW13048d
BW21045'
BW21046'
MC4100 via BW21620
BD792 via BW21027 BD792 via BW21073
MC4100 via BW21620
BD792 via BW13180
BD792 via BW13180
BD792 via BW3627
Mal' with P1
KanR with P1 on SG480 Mal' with P1 on MH1160 CmR with P1 on BW12126 TetR with P1 on BW13590
TetR with P1 on BW13590 TetR with P1 on BW16169 KanR, TetS swap with
Thr' with P I on BW20891
Thr' with P1 on BW20891
Pur' with P I on BW18551
Pur' with P1 on BW18551
TetR with P1 on LP64 TetR with P I on LP64 Mal' with P1 on MH1160
See text
KanR with P1 on BW21649 KanR with P I on BW21649
KanR with P I on AT142
KanR with P1 on BW21649
KanR with P1 on AT142
CmR with P1 on BW13180
h::Tn5-112
~~
a. Strains described earlier are BW4714 (Wanner and Boline, 1990); BW3627, BW13590, BW13711, BW13713, BW13736, BW13590, BW16169, BW16469, BW16470, BW16546 (Wanner and Wilmes-Riesenberg, 1992); and BW13745 (Wilmes-Riesenberg and Wanner, 1992). b. All are derivatives of E. coli K-12. Pedigree gives the parent from another laboratory and its immediate ancestor in this laboratory. c. Other strains new to this study are BW12126, BW12131, BWl3616, BW18551, and BW20891. d. BW13048 and BW13727 carry KanR and TetR versions of A(creABCD arcA thr) 704 from BW12070 (Fisher eta/., 1995), respectively. e. BW21045 and BW21046 carry nuoG::Tn 10 and purFfrom FD500.
insertions and was made as described elsewhere (Wanner and Wilmes-Riesenberg, 1992). The AcreSCD753 mutation is deleted of 2.6kbp between the Kpnl sites in creB and creD. It was constructed in vitro and recombined onto the chromosome using the pir-dependent allele replacement vector pWM41 (Metcalf et a/., 1996). The 6.5 kbp EcoRl to Hindlll fragment carrying the creABCDoperon was subcloned from pMW2 (Wilmes-Riesenberg and Wanner, 1992) into pBGS130+ (Spratt ef a/., 1986). The resulting plasmid, pSK3, was digested with Kpnl and religated to create pSKl2. A 3.2 kbp Sall fragment containing the Kpnl deletion was sub- cloned into pWM41 to create pSKl3, which was then trans- formed by electroporation into the non-pir host BW13713, in which the plasmid cannot replicate, resulting in its integration into the chromosome by homologous recombination. Several tetracycline-resistant (TetR) transformants were purified once non-selectively and then subjected to positive selection for loss of tetracycline resistance. A tetracycline-sensitive (Tets) and Bap- recombinant (BW20891) was verified with the polymerase chain reaction (PCR).
PCR verification
PCR was performed using cells grown on TYE agar. An iso- lated colony was added to 20p1 of lysis buffer (1 x PCR buffer: 50 mM KCI, 10 mM Tris-HCI (pH 8.3), 2.5 mM MgC12, 0.1 mgml-' nuelease-free BSA, 0.05mgml-' Proteinase K, 20 mM dithiothreitol, 1.8 mM SDS) and incubated at 37°C for 30 min, after which the tube was heated to 95°C for 5 min to inactivate the Proteinase K. Lysed cells (5 PI) were used for PCR. Amplification was carried out using the upstream creB primer (P36, ttgaattCAACGGGAAACGGGTCTGGTTAG, in which mismatched bases are in lowercase) and the down- stream creD primer (P93, TGCTTTGCCAGTGCGCCTG) at 50pmol each and 2 U of Taq DNA polymerase (Perkin- Elmer Cetus) in a volume of 100 pl containing 1 x PCR buffer, 2.5 mM MgC12, 200 pM each deoxynucleoside tri- phosphate. The mixture was subjected to 25 cycles (denaturing, 1 min at 94°C; annealing, 2min at 55°C; and extension, 1 min at 72°C). DNAs were analysed by elec- trophoresis on agarose gels before and after digestion with
0 1996 Blackwell Science Ltd, Molecular Microbiology, 22, 135-147

In vivo activation of PhoS by acetyl phosphate 145
References
Allen, N.E., and Hobbs, Jr, J.N. (1995) Induction of vancomycin resistance in Enterococcus faecium by non- glycopeptide antibiotics. E M S Microbiol Left 132: 107-1 14.
Amemura, M., Makino, K., Shinagawa, H., and Nakata, A. (1990) Cross talk to the phosphate regulon of Escherichia coli by PhoM protein: PhoM is a histidine protein kinase and catalyzes phosphorylation of PhoB and PhoM open reading frame 2. J Bacteriol 172: 6300-6307.
Arthur, M., Molinas, C., and Courvalin, P. (1992) The VanS- VanR two-component regulatory system controls synthesis of depsipeptide peptidoglycan precursors in Enferococcus faecium BM4147. J Bacteriol 174: 2582-2591.
Arthur, M., Depardieu, F., Holman, T., Wu, Z., Wright, G., Walsh, C.T., and Courvalin, P. (1995) Regulation of glyco- peptide resistance genes of enterococcal transposon Tn 7546 by the VanR-Vans two-component regulatory system. In Two-component Signal Transduction. Hoch, J.A., and Silhavy, T.J. (eds). Washington, DC: American Society for Microbiology, pp. 387-391.
Berlyn, M.K.B., Low, K.B., and Rudd, K.E. (1996) 109. Linkage map of Escherichia coli K-12, edn 9. In Escheri- chia coli and Salmonella typhimurium: Cellular and Mol- ecular Biology. Neidhardt, F.C., Curtiss, R.I., Ingraham, J.L., Lin, E.C.C., Low, Jr, K.B., Magasanik, B., Reznikoff, W., Riley, M., Schaechter, M., and Umbarger, H.E. (eds). Washington, DC: American Society for Microbiology, pp.
Bochner, B.R., and Ames, B.N. (1982) Complete analysis of cellular nucleotides by two-dimensional thin layer chroma- tography. J Biol Chem 257: 9759-9769.
Deretic, V., Leveau, J.H.J., Mohr, C.D., and Hibler, N.S. (1992) In vitro phosphorylation of AIgR, a regulator of mucoidy in Pseudomonas aeruginosa, by a histidine protein kinase and effects of small phospho-donor mol- ecules. Mol Microbiol6: 2761 -2767.
Feng, J., Atkinson, M.R., McCleary, W., Stock, J.B., Wanner, B.L., and Ninfa, A.J. (1992) Role of phosphorylated meta- bolic intermediates in the regulation of glutamine synthe- tase synthesis in Escherichia coli. J Bacteriol 174: 6061 - 6070.
Fisher, S.L., Jiang, W., Wanner, B.L., and Walsh, C.T. (1995) Cross-talk between the histidine protein kinase Vans and the response regulator PhoB: characterization and identi- fication of a Vans domain that inhibits activation of PhoB. J Biol Chem 270: 23143-23149.
Forst, S., Delgado, J., Ramakrishnan, G., and Inouye, M. (1988) Regulation of ompC and ompF expression in Escherichia coli in the absence of envZ. J Bacferiol 170:
Garrett, S., Taylor, R.K., and Silhavy, T.J. (1983) Isolation and characterization of chain-terminating nonsense muta- tions in a porin regulator gene, envZ J Bacterioll56: 62- 69.
Garrett, S., Taylor, R.K., Silhavy, T.J., and Berman, M.L. (1985) Isolation and characterization of AompB strains of Escherichia coli by a general method based on gene fusions. J Bacteriol 162: 840-844.
Hall, M.N., and Silhavy, T.J. (1981) Genetic analysis of the omp6 locus in Escherichia coli K-12. J MolBioll51: 1-15.
Handwerger, S., and Kolokathis, A. (1990) Induction of
1715-1902.
5080-5085.
Kpnl to confirm the AcreBCD753 mutation (data not shown).
Construction of the ompC-lacZ10-22(Lp 1[209]lacY 476::TnlO d-CM), ompF-lacZ16-13(Lp 1[209]lacY476:: TnlOd-CM), and ompC-lacZlo-25(Mu d1-1734) mutations
Strains carrying an ompC or ompf-lacZ fusion with an antibiotic-resistance marker were constructed to simplify transfer of the fusions. One with a chloramphenicol-resistance marker was constructed using hpl[209]lacY476::Tn 70dCM. This phage was made by in vivo recombination. In brief, a mixed lysate was prepared by propagation of hp1[209] on an exconjugant of BW4945 (Alac-769 hsdR574 rpsL) carrying F' 128 lacY476::Tn 70d-CM (from TTlO916). Lysogens carry- ing hpl[209]/acY476::Tn f0d-CM were selected as chlor- amphenicol-resistant (CmR) transductants of BW4714. One such transductant was UV-irradiated and, following plaque purification, the resulting phage was shown to carry chlor- amphenicol resistance (D. C. Young and 8. L. Wanner, unpublished). BW12125 was isolated as a CmR, LacZ', and Lacy- transductant of BW210 (a Mal' transductant of L1022) following infection with hpl[209]lacY476::Tn 70dCM. BW12056 was isolated as a KanR, Lac+, and hS transductant of BW220 (a Mal' transductant of L1025) after infection with a heat-induced lysate of MALI11 734 as described previously (Wanner and Boline, 1990). P1 grown on these strains was used to transfer the respective ompC and ompF mutations by selecting CmR and KanR transductants, as appropriate. Transductants were shown to carry the ompC-lacZlo_25 (Mu d7-7734) or ompf-lacZ,6-ls(hpl[209]lacY476::Tn 70d- CM) mutation using a P i lysate of CAGl2094 or CAG12177. The latter carry Tn 70 c. 70% (175/251) linked to ompC' or c. 38% (26/68) linked to ompF+, respectively. In each case, the expected portion of TetR transductants simultaneously became kanamycin sensitive and LacZ- or chloramphenicol, hs, and LacZ-, respectively.
Growth and enzyme assays
Cells were grown on glucose MOPS agar with 2mM Pi overnight, then simultaneously plated on the same or other media, and incubated for an additional 16-20h before assay. Cells were scraped from the surface and suspended at a cell ODdp0 of c. 0.3-0.5 in Bap assay buffer (1 M TrisHCI, pH8.0, containing 1 mM merthiolate and 1OOpg per ml chloramphenicol) or in p-galactosidase assay buffer (2 buffer containing 1% p-mercaptoethanol and 1OOpg per ml chloramphenicol). Enzyme activities were measured as described elsewhere (Wanner, 1994b). All values in each table are for experiments carried out side by side.
Acknowledgements
We thank individuals cited in the text for generous gifts of strains, and we thank Andreas Haldimann and Jill Hutch- croft for carefully reading the manuscript. This research was supported by NSF Grant MCB9405929.
1996 Blackwell Science Ltd, Molecular Microbiology, 22, 135-147

146 S.-K. Kim, M. R. Wilmes-Riesenberg and B. L. Wanner
vancomycin resistance in Enterococcus faecium by inhibi- tion of transglycosylation. FEMS Microbiol Lett 70: 167- 170.
Heyde, M., and Portalier, R. (1987) Regulation of major outer membrane porin proteins of Escherichia coli K-12 by pH. Mol Gen Genet 208: 511-517.
Hiratsu, K., Makino, K., Nakata, A., and Shinagawa, H. (1995) Autophosphorylation and activation of transcriptional acti- vator PhoB of Escherichia coli by acetyl phosphate in vitro. Gene 161: 7-10.
Holman, T.R., Wu, Z., Wanner, B.L., and Walsh, C.T. (1994) Identification of the DNA-binding site for the phosphory- lated VanR protein required for vancomycin resistance in Enterococcus faecium. Biochemistry 33: 4625-4631 .
Hunt, A.G. (1980) Micromethod for the measurement of acetyl phosphate and acetyl coenzyme A. Meth Enzymol
Hunt, A.G., and Hong, J.-S. (1980) A micromethod for the measurement of acetyl phosphate and acetyl coenzyme A. Anal Biochem 108: 290-294.
Jiang, W., Metcalf, W.W., Lee, K.-S., and Wanner, B.L. (1995) Molecular cloning, mapping, and regulation of Pho regulon genes for phosphonate breakdown by the phosphonatase pathway of Salmonella typhimurium LT2. J Bacteriol 177:
Lukat, G.S., McCleary, W.R., Stock, A.M., and Stock, J.B. (1 992) Phosphorylation of bacterial response regulator proteins by low molecular weight phospho-donors. Roc Natl Acad Sci USA 89: 71 8-722.
McCleary, W.R. (1996) The activation of PhoB by acetyl phosphate. Mol Microbiol 20: 1 155-1 163.
McCleary, W.R., and Stock, J.B. (1994) Acetyl phosphate and the activation of two-component response regulators. J Biol Chem 269: 31 567-31 572.
Makino, K., Shinagawa, H., Amemura, M., Kimura, S., Nakata, A,, and Ishihama, A. (1988) Regulation of the phosphate regulon of Escherichia coli: activation of pstS transcription by PhoB protein in vitro. J Mol Biol 203: 85-95.
Makino, K., Amemura, M., Kim, S.-K., Shinagawa, H., and Nakata, A. (1992) Signal transduction of the phosphate regulon in Escherichia coli mediated by phosphorylation. In Adenine Nucleotides in Cellular Energy Transfer and Signal Transduction. Papa, S., Azzi, A., and Tager, J.M. (eds). Basel: Birkhaeuser Verlag, pp. 191 -200.
Metcalf, W.W., Jiang, W., Daniels, L.L., Kim, S.-K., Haldi- mann, A., and Wanner, B.L. (1996) Conditionally replicative and conjugative plasmids carrying IacZci for cloning, mutagenesis, and allele replacement in bacteria. flasmid
Mizuno, T., and Mizushima, S. (1987) Isolation and charac- terization of deletion mutants of ompR and envZ, regula- tory genes for expression of the outer membrane proteins OmpC and OmpF in Escherichia coli. J Biochem 101 : 387- 396.
Nara, F., Matsuyama, S.-I., Mizuno, T., and Mizushima, S. (1986) Molecular analysis of mutant ompR genes exhibiting different phenotypes as to osmoregulation of the ompFand ompc genes of Escherichia coli. Mol Gen Genet 202: 194- 199.
Nystrom, T. (1 994) The glucose-starvation stimulon of Escheri- chia coli: induced and repressed synthesis of enzymes of
122: 43-50.
641 1-6421.
35: 1-13.
central metabolic pathways and role of acetyl phosphate in gene expression and starvation survival. Mol Microbioll2:
Pratt, L.A., and Silhavy, T.J. (1995) Porin regulation of Escherichia coli. In Two-component Signal Transduction. Hoch, J.A., and Silhavy, T.J. (eds). Washington, DC: American Society for Microbiology, pp. 105-127.
Pratt, L.A., Hsing, W., Gibson, K.E., and Silhavy, T.J. (1996) From acids to osmZ multiple factors influence synthesis of the OmpF and OmpC porins in Escherichia coli. Mol Microbiol 20: 91 1-917.
Pruss, B.M., and Wolfe, A.J. (1994) Regulation of acetyl phosphate synthesis and degradation, and the control of flagellar expression in Escherichia coli. Mol Microbiol 12:
Shin, S., and Park, C. (1995) Modulation of flagellar expres- sion in Escherichia coli by acetyl phosphate and the osmoregulator OmpR. J Bacteriol 177: 4696-4702.
Slauch, J.M., Garrett, S., Jackson, D.E., and Silhavy, T.J. (1988) EnvZ functions through OmpR to control porin gene expression in Escherichia coli K- 12. J Bacteriol 170: 439- 441.
Spratt, B.G., Hedge, P.J., te Heesen, S., Edelman, A,, and Broome-Smith, J.K. (1 986) Kanamycin-resistant vectors that are analogues of plasmids pUC8, pUC9, pEMBL8, and pEMBL9. Gene 41 : 337-342.
Stock, J.B., Surette, M.G., Levit, M., and Park, P. (1995) Two- component signal transduction systems: structure-function relationships and mechanisms of catalysis. In Two-compo- nent Signal Transduction. Hoch, J.A., and Silhavy, T.J. (eds). Washington, DC: American Society for Microbiology,
Wanner, B.L. (1 987) Control of phoR-dependent bacterial alkaline phosphatase clonal variation by the phoM region. J Bacteriol 169: 900-903.
Wanner, B.L. (1992) Is cross regulation by phosphorylation of two-component response regulator proteins important in bacteria? J Bacteriol'l74: 2053-2058.
Wanner, B.L. (1993) Gene regulation by phosphate in enteric bacteria. J Cell Biochem 51 : 47-54.
Wanner, B.L. (1 994a) Multiple controls of the Escherichia coli PHO regulon by the Pi sensor PhoR, the catabolite regulatory Sensor CreC, and acetyl phosphate. In Phos- phate in Microorganisms, Cellular and Molecular Biology. Torriani-Gorini, A., Yagil, E., and Silver, S. (eds). Washing- ton, DC: American Society for Microbiology, pp. 13-21.
Wanner, B.L. (1994b) Gene expression in bacteria using TnphoA and TnphoA' elements to make and switch phoA gene, lacZ (op), and lacZ (pr) fusions. In Methods in Molecular Genetics. Vol 3. Adolph, K.W. (ed.). Orlando: Academic Press, pp. 291 -31 0.
Wanner, B.L. (1 995) Signal transduction and cross regulation in the Escherichia coli phosphate regulon by PhoR, CreC, and acetyl phosphate. In Two-component Signal Transduc- tion. Hoch, J.A., and Silhavy, T.J. (eds). Washington, DC: American Society for Microbiology, pp. 203-221.
Wanner, B.L. (1 996) Phosphorus assimilation and control of the phosphate regulon. In Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology. Neidhardt, F.C., Curtiss, R.I., Ingraham, J.L., Lin, E.C.C., Low, Jr, K.B., Magasanik, B., Reznikoff, W., Riley, M., Schaechter,
833-843.
973-984.
pp. 25-51.
0 1996 Blackwell Science Ltd, Molecular Microbiology, 22, 135-1 47

M., and Umbarger, H.E. (eds). Washington, DC: American Society for Microbiology, pp. 1357-1 381.
Wanner, B.L., and Boline, J.A. (1990) Mapping and molecular cloning of the phn (psiD) locus for phosphonate utilization in Escherichia coli. J Bacteriol 172: 1 186-1 196.
Wanner, B.L., and Latterell, P. (1980) Mutants affected in alkaline phosphatase expression: evidence for multiple positive regulators of the phosphate regulon in Escherichia coli. Genetics 96: 242-266.
Wanner, B.L., and Wilmes-Riesenberg, M.R. (1 992) Involve- ment of phosphotransacetylase, acetate kinase, and acetyl phosphate synthesis in the control of the phosphate regulon in Escherichia coli. J Bacteriol 174: 2124-2130.
Wanner, B.L., Witmes, M.R., and Young, D.C. (1 988) Control
In vivo activation of PhoB by acetyl phosphate 147
of bacterial alkaline phosphatase synthesis and variation in an Escherichia coli K-12 phoR mutant by adenyl cyclase, the cyclic AMP receptor protein, and the phoM operon. J Bacteriol 170: 1092-1 102.
Wanner, B.L., Jiang, W., Kim, S.-K., Yamagata, S., Haldi- mann, A., and Daniels, L.L. (1995) Are the multiple signal transduction pathways of the Pho regulon due to cross talk or cross regulation? In Regulation of Gene Expression in Escherichia coli. Lin, E.C.C., and Lynch, AS. (eds). Austin, Texas: R. G. Landes Co., pp. 297-315.
Wilmes-Riesenberg, M.R., and Wanner, B.L. (1 992) TnphoA and TnphoA’ elements for making and switching fusions for study of transcription, translation, and cell surface local- ization. J Bacteriol 174: 4558-4575.
0 1996 Blackwell Science Ltd, Molecular Microbidogy, 22, 135-147




















