
Pre-Trial Detention in Greece: the Achilles heel of the prison system
Upload
dayanandasagarCategory
view
1download
0
Chemico-Biological Interactions 211 (2014) 1–10
Contents lists available at ScienceDirect
Chemico-Biological Interactions
journal homepage: www.elsevier .com/locate /chembioint
Investigation into the cyto-protective and wound healing properties ofcryptic peptides from bovine Achilles tendon collagen
0009-2797/$ - see front matter � 2014 Elsevier Ireland Ltd. All rights reserved.http://dx.doi.org/10.1016/j.cbi.2014.01.002
⇑ Corresponding author. Tel.: +91 416 2202479; fax: +91 416 2243092.E-mail address: [email protected] (C. Shanthi).
Pradipta Banerjee, Alka Mehta, C. Shanthi ⇑School of Bio Science and Technology, VIT University, Vellore 632014, Tamil Nadu, India
a r t i c l e i n f o
Article history:Received 9 October 2013Received in revised form 3 December 2013Accepted 7 January 2014Available online 13 January 2014
Keywords:CollagenCryptic peptidesReactive oxygen speciesAntioxidativeCell adhesionMonolayer wound healing assay
a b s t r a c t
Many proteins have concealed regions in their amino acid sequences that when liberated or exposed byconformational changes can exhibit bioactivity. Two such cryptic bioactive peptides, C2 (with cell adhe-sive properties) and E1 (with cell adhesive and antioxidant properties) have been isolated from bovinetendon collagen. This investigation deals with the efficacy of these peptides in countering externallygenerated stress and imparting cyto-protection in mammalian cell systems. The cell survival activitywas studied with two cell lines, viz., HeLa and Vero, with varying concentrations of five oxidativestress-generating agents. The activities of the peptides in supporting cell adhesion and countering stresswere determined in their coated and dissolved forms. C2 and E1 coated dishes registered 8 times(p < 0.01) higher rate of cell survival against oxidative stress than collagen coated dishes. E1 increasedstress tolerance levels by >100 times in dissolved form and C2, by 8 times in coated form. The peptidessupported faster wound closure than collagen under normal as well as stressed condition. Maximumstress tolerance was observed on C2 coated dishes in the presence of E1 in the medium suggesting thatboth enhanced cell adhesion and antioxidative activities significantly contribute to the cell survivalduring stress. The present study emphasizes that collagen peptides, apart from providing a suitablesurface for cell adhesion, also confer protection to cells against oxidative stress.
� 2014 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
The extracellular matrix (ECM) provides mechanical supportand is also involved in biochemical signalling through membranereceptors with the cells that are in contact with it. These signalscan have a significant influence on cellular cytoskeletal structure,gene transcription and migration. In turn, the cells remodel theECM enzymatically and play an active role in sculpting a microen-vironment suitable for their phenotypes [1]. This dynamic and re-ciprocal communication between the cells and ECM is nowconsidered essential for regular functioning and suitably respond-ing to stress. A key component of such communications involvesthe recognition of a vital ECM component, generally, a short stretchof amino acids, present at a strategic location in the larger protein,by transmembrane receptors such as integrins [2]. The interactionsgenerally require either conformational changes in the ECM pro-tein leading to unmasking of the active region or direct proteolyticaction resulting in the excision of the bioactive region as a peptide.Such concealed segments, which upon enzymatic cleavage or con-formational change can exhibit physiological activities, are termedas cryptic bioactive peptides [3,4]. During ECM remodelling and/or
wound healing, protease-generated specific fragments of constitu-ent proteins can exhibit a wide array of bioactivities including anti-oxidative, chemotactic, wound healing, anti tumorogenic andangiotensin I-converting enzyme inhibition [5–7].
Reactive oxygen species (ROS) are required by living systems tomaintain a number of functions including signal transduction, celladhesion and wound healing [8]. Oxidative stress ensues when thedelicate balance between ROS creation and depletion is hampereddue to tissue injury or faulty detoxification mechanisms. In case ofpersistent wounds or the presence of foreign substances in theproximity of a wound site, the resultant uncontrolled stress dueto ROS or otherwise, leads to delayed healing [9]. Excess ROS actsthrough multiple mechanisms to generate stress including oxida-tion of cellular macromolecules leading to membrane damage, en-zyme dysfunction and hampering of metabolic flux along withimpaired DNA repair, resulting in mutagenesis or cell death [10].Oxidative stress generally has an inverse relation with cell adhe-sion and wound healing; although very low levels of free radicalsare required for integrin attachment to ECM, higher levels resultin cell death. Excess ROS can, along with other effects, alter theexpression of integrin sub-unit genes, consequently leading to im-proper connectivity with the ECM and eventually causing celldeath [11]. Oxidative stress is also involved in reorganization of ac-tin and vimentin arrangements, leading to changes in the focal

2 P. Banerjee et al. / Chemico-Biological Interactions 211 (2014) 1–10
adhesion site resulting in loss of cell-matrix adhesion. This furtherleads to the activation of apoptotic pathways, cell loss, compro-mised tissue integrity and pathologic consequences [12].
In the present study, it is hypothesized that the cell loss due toROS stress can be minimized in the presence of cryptic peptides. Inother words, components of the matrix with prominent cell adhe-sive and antioxidative activities can provide some form of protec-tion to stress-exposed cells. The increased cell survival, in turn, canalso have a substantial effect on cell migration during in vivostress-generating events like tissue remodelling and woundclosure.
The physiological effects of oxidative stress in vitro can be mim-icked by the use of exogenously administered H2O2 and heavy me-tal ions. For the latter scenario, ROS stress can be generatedthrough two distinct mechanisms. With redox-active metal ionslike Fe2+ and Cr6+, a Fenton-like reaction increases ROS while re-dox-inactive toxic metal ions such as Hg2+ and Cd2+ act throughdepletion of major antioxidant components leading to unquenchedfree radicals [13]. In this study, oxidative stress has been generatedby three methods; subjecting cells to H2O2, a combination of Fe2+-H2O2 and heavy metal ions.
The peptides C2 (sequence: GPOGPOGKNGDDGEAGKPGRPG) andE1 (sequence: GETGPAGPAGPIGPVGARGPAGPQGPRGDKGETGEQ)with molecular weights 2.8 and 3.2 kDa respectively have beenisolated from type I collagen in our earlier work [14,15]. The peptideC2 exhibits cell adhesion properties [14] and the RGD-containingpeptide E1 exhibits both cell adhesive and antioxidative properties[15]. The present study was carried out to determine the role of thesepeptides in countering ROS stress and the ability to aid in vitro woundhealing properties.
2. Materials and methods
Bovine Achilles tendons were collected from a slaughterhouseat Vellore, Tamil Nadu, India and type I collagen was purified fromthe tendons through acid extraction/salt precipitation technique[16].
T-flasks (Nunclon surface) were procured from Nunc, Roskilde,Denmark and disposable culture dishes (35 � 10 mm) were ob-tained from Fischer Scientific, Hanover park, IL, USA. PowderedDulbecco’s modified Eagle’s Medium (DMEM), supplemented with2 mM glutamine and 10� antibiotic–antimycotic solution were ob-tained from HiMedia, India. Foetal bovine serum and 10� sterilefiltered trypsin–EDTA solution were obtained from Sigma–Aldrich,St. Louis, MO, USA. Vero and HeLa cell lines were procured fromNational Centre for Cell Science, Pune, India. All bench work asso-ciated with cell lines, including peptide coating has been carriedout inside a class II bio-safety cabinet (Clean Air Systems, Chennai,India) for maintaining sanitized conditions. The metals used forstress generation, K2Cr2O7, FeSO4 and HgCl2, ACS-grade, were ob-tained from Sigma–Aldrich.
2.1. Peptide isolation and coating
The peptides C2 and E1 were isolated as described in earlierstudies [14,15]. Briefly, collagen was cleaved with an extracellularsemi-purified enzyme preparation and peptides fractionatedthrough a combination of ion-exchange and gel permeation chro-matographic techniques. The purified peptides were coated ontothe disposable dishes according to standard procedures [17]. Acoating density of 0.5 nmol/cm2 area, as optimized in our previousstudy [15], has been used throughout this study. Dishes coatedwith extracted collagen type I, termed as CCS (collagen-coated sur-face) was also tested for de-stressor activity.
2.2. Cell maintenance
Vero and HeLa cell lines were maintained according to standardprotocols [18]. Cells were cultured in DMEM supplemented with10% foetal bovine serum and incubated at 37 �C in a 5% CO2 humid-ified incubator. For experiments, cells reaching 80% confluencewere detached from T-flasks with trypsin–EDTA, centrifuged andcell number was enumerated in a Neuber’s chamber through try-pan blue exclusion assay.
2.3. Hydroxyl radical scavenging assay
The hydroxyl radical scavenging efficacy was determinedaccording to the colorimetric assay of Zhang et al. [19]. Knownquantities of peptides C2, E1 and collagen type I were dissolvedin 20 mM KH2PO4-KOH buffer at pH 7.5 and solutions of varyingconcentrations ranging from 0.1 to 100 mM were prepared, the re-agents added and the developed color was measured at 532 nm.BSA and butylated hydroxyl toluene (BHT) was used as negativeand positive control, respectively. Assays were performed intriplicates. Scavenging ability was calculated from the followingequation:
Radical scavenging activity ð%Þ ¼ 1� TestControl
� �� 100
where Test is the absorbance for the test peptide and collagen sam-ples and Control is the absorbance in the absence of peptides andcollagen.
2.4. De-stressor activity of the peptides
Stress was generated by five different stress-inducing agents aslisted in Table 1. Concentrations of the stressors used were chosenfrom previously reported toxicity levels [20–23]. Metal stock solu-tions were prepared in deionized water and sterilized by filtrationthrough 0.2 lm filter. At the time of treatment, stock solutionswere diluted in culture medium to the final concentrations re-quired. For agents 4 and 5, as listed in Table 1, stock H2O2 solutionwas stored in 4 �C and diluted in sterilized Dulbecco’s modifiedCa2+ and Mg2+ free phosphate buffered saline before adding tothe medium.
2.4.1. Assaying de-stressor activity in coated form3.5 � 106 cells were seeded onto the dishes coated with E1, C2
and collagen followed immediately by exposure to the stressoragents in varying concentrations as given in Table 1, keeping thetotal volume constant at 1.5 ml. An equal number of cells seededonto dishes coated with the peptides and collagen, but withoutexposure to any stress, were used as the control. After 6 hincubation, the adhered cells were trypsinized and counted byhaemocytometry.
2.4.2. Assaying de-stressor activity in dissolved form3.5 � 106 cells suspended in DMEM were seeded onto CCS
dishes and subjected to the same level of stress as mentioned inSection 2.4.1. The peptides E1 and C2 were suspended separatelyin 0.5 ml medium and added to the cells to a final concentrationof 0.815 lM [14]. Care was taken to adjust the added amount ofstressors and cells such that the final concentration and volumewould be the same as in Table 1. After 6 h incubation, adherentcells were counted and the stressor concentration ensuing 50% cellsurvival was calculated.

Table 1List of agents used as stressors and their concentration ranges.
Agent Stressor Range of final concentrations (mM)
1 Cr(VI) 0 0.01 0.1 1 10 1002 Fe(II) 0 0.01 0.1 1 10 1003 Hg(II) [lM] 0 1.5625* 3.125* 12.5* 25* 50*
4 H2O2 0 2.5 5 10 25 1005 Fenton’s reagent Fe(II) 0 0.1 5 10 50 100
H2O2 0 0.01 0.1 1 10 50
* Concentrations given in lM.
P. Banerjee et al. / Chemico-Biological Interactions 211 (2014) 1–10 3
2.5. Wound closure assay
The wound closure assay was performed according to the pro-tocols developed by Liang et al. [24]. 3.5 � 105 cells were seededonto peptide coated dishes. An equal number of cells were seededonto collagen coated (positive control) and uncoated dishes (nega-tive controls). When cells have just reached 90–95% confluence,two parallel scratch wounds of approximately 400 lm diameterwere made with a pipette tip in all the dishes. The medium wasreplenished for all the dishes with certain changes as mentionedin Table 2. 10 mM H2O2 was added in set I, 10 mM H2O2 along withE1 in dissolved form (0.815 lM) was added in set II and set III wasincubated as such.
Images of a fixed area of the wound were taken at regular inter-vals over a period of 24 h. Image analysis was carried out by ImageJsoftware (http://rsbweb.nih.gov/ij/) from the National Institute ofHealth, USA. The wound-edge positions of the cells were averagedby digitally drawing lines. The area of the wound was calculated asa rectangle, whose length was taken to be the length of the woundin focus and the average distance between the two advancingedges of the wound as breadth. The decrease in this rectangulararea with time was calculated by measuring the decreasing dis-tance between the advancing edges. Finally, the total was sub-tracted from the original area to determine the area covered andexpressed in % [25].
2.6. Statistical analysis
The hydroxyl radical scavenging and cell adhesion/proliferationassays were performed in triplicates and activities reported asmean ± standard deviation. For the wound healing migration assay,duplicate dishes were used, with each dish having two scratchesand each scratch photographed in two pre-determined areas. Lar-ger datasets were analyzed for statistical significance using oneway and two-way ANOVA. Comparison between multiple groupswas accomplished by post hoc Tukey’s test and student’s t-test. pvalues less than 0.05 were considered significant.
3. Results and discussions
3.1. Hydroxyl radical scavenging assay
Since ROS stress mostly generates �OH radicals, it was necessaryto check for the ability of the peptides to scavenge the same. Theactivities of BHT, the two peptides and collagen were significantlydifferent (p < 0.004) at a confidence level of 95% based on ANOVA.
Table 2Experimental set up for in vitro wound healing studies performed under three conditionspresence of both stressor and a de-stressor (Set III).
Set I C2 coated E1 coatedSet II C2 coated + H2O2 E1 coated + H2O2
Set III C2 coated + E1 dissolved + H2O2 E1 coated + E1 dissolved + H2O2
As depicted in Fig. 1, at 100 nmol, E1 exhibited a scavenging activ-ity of 58.7%, whereas C2 achieved only 15%. BHT, used as the posi-tive control, exhibited highest activity at 100 nmol. To summarize,the results obtained confirmed E1’s effectiveness as a potent �OHscavenger and are also in agreement with our earlier studiesreporting E1 to be a strong antioxidative agent with DPPH radicalscavenging, Fe2+ chelation and extensive reductive ability [14].
3.2. Stress generation by the agents
Stress is a condition in which the homeostasis maintained inliving systems fail, giving rise to diverse pathological conse-quences. Exogenously administered H2O2 induces cell deaththrough apoptosis and necrosis in a concentration-dependentmanner [26]. H2O2 can react with cellular Fe2+ to form reactive rad-icals through Fenton reaction and can stimulate the upregulationand activity of redox-oriented enzymes leading to unregulatedproduction of ROS initiating a vicious cycle [27]. The reactive rad-icals hamper several important cellular signalling pathways andstimulate the release of intracellular iron, further feeding the vis-cous cycle and culminating in pathological conditions.
Cr6+ is a powerful oxidant capable of interaction with cellularcomponents and its reduced forms can react with H2O2 to produceseveral ROS species. Cr3+ in particular has a low tendency to crossthe cell membrane and can form stable complexes with DNA lead-ing to chromosomal aberrations [28,29].
Hg ions are known to generate stress through a wide variety ofmechanisms including protein denaturation, impaired electrontransport leading to increased H2O2 and superoxide production,alteration in Ca levels, and increasing lipid peroxidation. Due tothe higher toxicity of Hg ions in comparison to the other metals[30], concentration ranges in the lM levels were used in the pres-ent study.
3.3. De-stressor activities of peptides
Cell counts in the CCS dishes were found to decrease withincreasing stressor concentration (Correlation = �0.8). However,as displayed in Figs. 2–5, presence of the peptides, both in coatedand dissolved form had a positive impact on cell adhesion and sur-vival (Correlation = �0.4 to �0.6). Table 3 lists the stressor concen-trations in which 50% of the seeded cells survived in the presenceof C2 and E1. On an average for both HeLa and Vero cell lines,C2-coated dishes exhibited cell survival at 5 times higher stressorconcentration for metal-based stress (Figs. 2 and 3) and 10 timeshigher stressor concentration for redox stress (Figs. 4 and 5) when
: in the absence of a stressor (Set I), in the presence of a stressor (Set II) and in the
Collagen coated UncoatedCollagen coated + H2O2 Uncoated + H2O2
Collagen coated + E1 dissolved + H2O2 Uncoated + E1 dissolved + H2O2
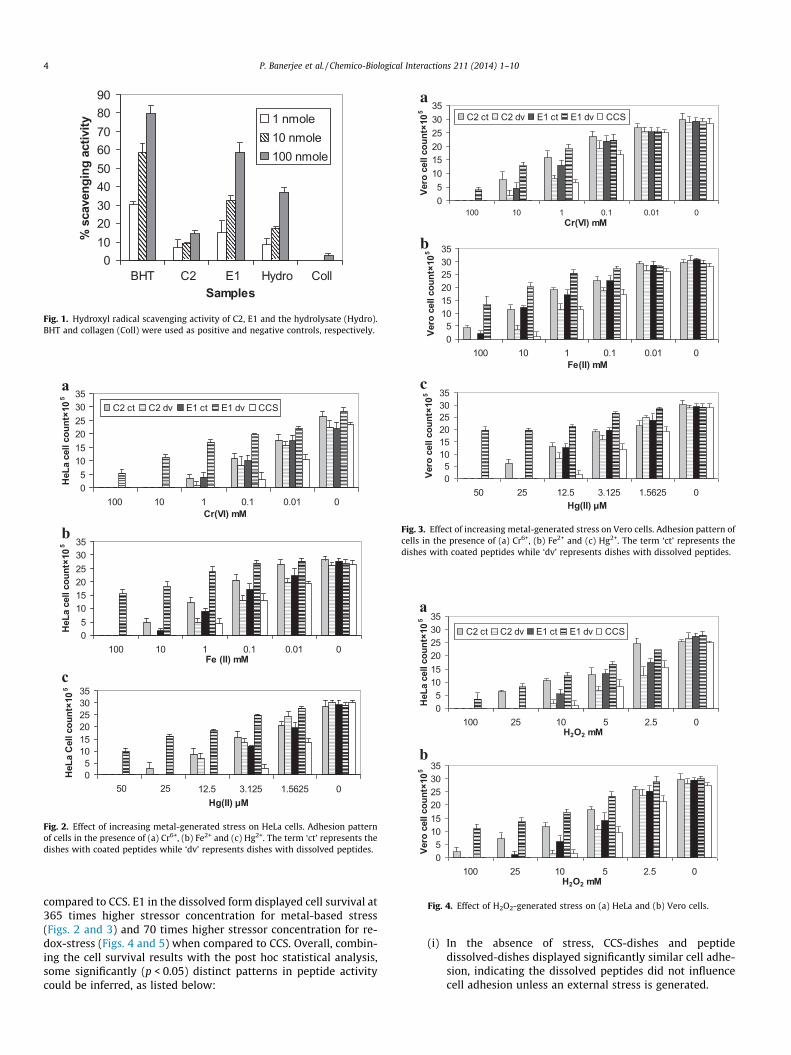
0102030405060708090
BHT C2 E1 Hydro CollSamples
% s
cave
ngin
g ac
tivity 1 nmole
10 nmole100 nmole
Fig. 1. Hydroxyl radical scavenging activity of C2, E1 and the hydrolysate (Hydro).BHT and collagen (Coll) were used as positive and negative controls, respectively.
05
101520253035
100 10 1 0.1 0.01 0Cr(VI) mM
HeL
a ce
ll co
unt×
105
C2 ct C2 dv E1 ct E1 dv CCS
b
a
05
101520253035
100 10 1 0.1 0.01 0Fe (II) mM
HeL
a ce
ll co
unt×
105
c
05
101520253035
50 25 12.5 3.125 1.5625 0Hg(II) µM
HeL
a C
ell c
ount
×105
Fig. 2. Effect of increasing metal-generated stress on HeLa cells. Adhesion patternof cells in the presence of (a) Cr6+, (b) Fe2+ and (c) Hg2+. The term ‘ct’ represents thedishes with coated peptides while ‘dv’ represents dishes with dissolved peptides.
05
101520253035
100 10 1 0.1 0.01 0 Cr(VI) mM
Vero
cel
l cou
nt×1
05 C2 ct C2 dv E1 ct E1 dv CCS
05
101520253035
100 10 1 0.1 0.01 0Fe(II) mM
Vero
cel
l cou
nt×1
05
05
101520253035
50 25 12.5 3.125 1.5625 0Hg(II) µM
Vero
cel
l cou
nt×1
05
a
b
c
Fig. 3. Effect of increasing metal-generated stress on Vero cells. Adhesion pattern ofcells in the presence of (a) Cr6+, (b) Fe2+ and (c) Hg2+. The term ‘ct’ represents thedishes with coated peptides while ‘dv’ represents dishes with dissolved peptides.
05
101520253035
100 25 10 5 2.5 0H2O2 mM
HeL
a ce
ll co
unt×
105
C2 ct C2 dv E1 ct E1 dv CCS
b
a
05
101520253035
100 25 10 5 2.5 0H2O2 mM
Vero
cel
l cou
nt×1
05
Fig. 4. Effect of H2O2-generated stress on (a) HeLa and (b) Vero cells.
4 P. Banerjee et al. / Chemico-Biological Interactions 211 (2014) 1–10
compared to CCS. E1 in the dissolved form displayed cell survival at365 times higher stressor concentration for metal-based stress(Figs. 2 and 3) and 70 times higher stressor concentration for re-dox-stress (Figs. 4 and 5) when compared to CCS. Overall, combin-ing the cell survival results with the post hoc statistical analysis,some significantly (p < 0.05) distinct patterns in peptide activitycould be inferred, as listed below:
(i) In the absence of stress, CCS-dishes and peptidedissolved-dishes displayed significantly similar cell adhe-sion, indicating the dissolved peptides did not influencecell adhesion unless an external stress is generated.
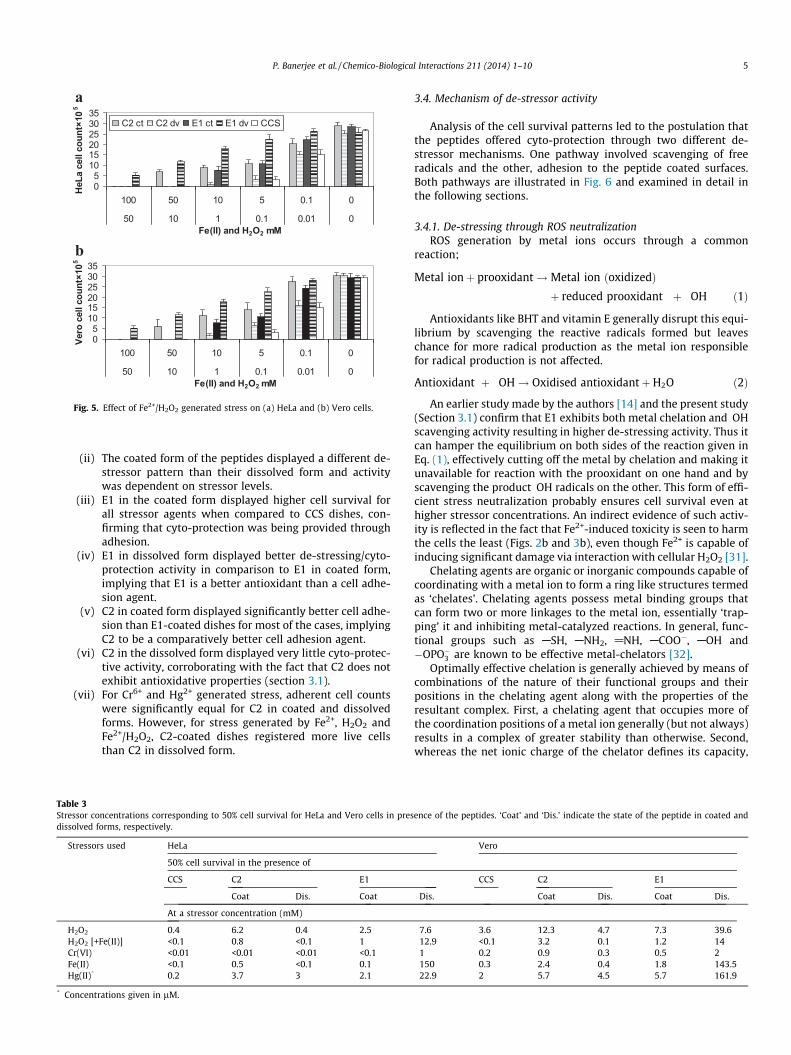
05
101520253035
100 50 10 5 0.1 0
50 10 1 0.1 0.01 0Fe(II) and H2O2 mM
HeL
a ce
ll co
unt×
105
C2 ct C2 dv E1 ct E1 dv CCS
b
a
05
101520253035
100 50 10 5 0.1 0
50 10 1 0.1 0.01 0Fe(II) and H2O2 mM
Vero
cel
l cou
nt×1
05
Fig. 5. Effect of Fe2+/H2O2 generated stress on (a) HeLa and (b) Vero cells.
P. Banerjee et al. / Chemico-Biological Interactions 211 (2014) 1–10 5
(ii) The coated form of the peptides displayed a different de-stressor pattern than their dissolved form and activitywas dependent on stressor levels.
(iii) E1 in the coated form displayed higher cell survival forall stressor agents when compared to CCS dishes, con-firming that cyto-protection was being provided throughadhesion.
(iv) E1 in dissolved form displayed better de-stressing/cyto-protection activity in comparison to E1 in coated form,implying that E1 is a better antioxidant than a cell adhe-sion agent.
(v) C2 in coated form displayed significantly better cell adhe-sion than E1-coated dishes for most of the cases, implyingC2 to be a comparatively better cell adhesion agent.
(vi) C2 in the dissolved form displayed very little cyto-protec-tive activity, corroborating with the fact that C2 does notexhibit antioxidative properties (section 3.1).
(vii) For Cr6+ and Hg2+ generated stress, adherent cell countswere significantly equal for C2 in coated and dissolvedforms. However, for stress generated by Fe2+, H2O2 andFe2+/H2O2, C2-coated dishes registered more live cellsthan C2 in dissolved form.
Table 3Stressor concentrations corresponding to 50% cell survival for HeLa and Vero cells in presdissolved forms, respectively.
Stressors used HeLa
50% cell survival in the presence of
CCS C2 E1
Coat Dis. Coat
At a stressor concentration (mM)
H2O2 0.4 6.2 0.4 2.5H2O2 [+Fe(II)] <0.1 0.8 <0.1 1Cr(VI) <0.01 <0.01 <0.01 <0.1Fe(II) <0.1 0.5 <0.1 0.1Hg(II)* 0.2 3.7 3 2.1
* Concentrations given in lM.
3.4. Mechanism of de-stressor activity
Analysis of the cell survival patterns led to the postulation thatthe peptides offered cyto-protection through two different de-stressor mechanisms. One pathway involved scavenging of freeradicals and the other, adhesion to the peptide coated surfaces.Both pathways are illustrated in Fig. 6 and examined in detail inthe following sections.
3.4.1. De-stressing through ROS neutralizationROS generation by metal ions occurs through a common
reaction;
Metal ionþ prooxidant!Metal ion ðoxidizedÞþ reduced prooxidant þ �OH ð1Þ
Antioxidants like BHT and vitamin E generally disrupt this equi-librium by scavenging the reactive radicals formed but leaveschance for more radical production as the metal ion responsiblefor radical production is not affected.
Antioxidant þ �OH! Oxidised antioxidantþH2O ð2Þ
An earlier study made by the authors [14] and the present study(Section 3.1) confirm that E1 exhibits both metal chelation and �OHscavenging activity resulting in higher de-stressing activity. Thus itcan hamper the equilibrium on both sides of the reaction given inEq. (1), effectively cutting off the metal by chelation and making itunavailable for reaction with the prooxidant on one hand and byscavenging the product �OH radicals on the other. This form of effi-cient stress neutralization probably ensures cell survival even athigher stressor concentrations. An indirect evidence of such activ-ity is reflected in the fact that Fe2+-induced toxicity is seen to harmthe cells the least (Figs. 2b and 3b), even though Fe2+ is capable ofinducing significant damage via interaction with cellular H2O2 [31].
Chelating agents are organic or inorganic compounds capable ofcoordinating with a metal ion to form a ring like structures termedas ‘chelates’. Chelating agents possess metal binding groups thatcan form two or more linkages to the metal ion, essentially ‘trap-ping’ it and inhibiting metal-catalyzed reactions. In general, func-tional groups such as ASH, ANH2, @NH, ACOO�, AOH and�OPO�3 are known to be effective metal-chelators [32].
Optimally effective chelation is generally achieved by means ofcombinations of the nature of their functional groups and theirpositions in the chelating agent along with the properties of theresultant complex. First, a chelating agent that occupies more ofthe coordination positions of a metal ion generally (but not always)results in a complex of greater stability than otherwise. Second,whereas the net ionic charge of the chelator defines its capacity,
ence of the peptides. ‘Coat’ and ‘Dis.’ indicate the state of the peptide in coated and
Vero
CCS C2 E1
Dis. Coat Dis. Coat Dis.
7.6 3.6 12.3 4.7 7.3 39.612.9 <0.1 3.2 0.1 1.2 141 0.2 0.9 0.3 0.5 2150 0.3 2.4 0.4 1.8 143.522.9 2 5.7 4.5 5.7 161.9

Fig. 6. Schematic representation of the mechanisms through which C2 and E1 display cyto-protective effects. The solid arrows represent the pathways leading to cell death inpresence of stress. The dotted arrows represent the action of the peptides. E1 in the dissolved form (Ed) scavenges free radicals along with chelating Fe2+ and Cr6+ and possiblyHg2+. In the coated form, E1 and C2 maintain cell adhesion and possibly activate anti-apoptotic pathways.
6 P. Banerjee et al. / Chemico-Biological Interactions 211 (2014) 1–10
its distribution and ability to reach the metal ion decide the possi-bility of formation of the complex. The final criterion requires thatthe chelation render the metal ion non-toxic and/or making themetal ion unavailable for any redox reactions [33].
In recent times, bioactive peptides have been identified to beeffective chelators and radical scavengers. The activity arises be-cause of the presence of multiple ligand coordination sites due tothe myriad side chain functional groups of the amino acids consti-tuting the peptide [34]. There are two major factors influencingcoordination of metal ions by peptides; reactive state of thepeptide amide group and the side chain residues with coordinationproperties. The alcoholic-OH from S and T, ACOO� from D and Eand ANH2 from K along with the free N- and C-terminal groupsof peptides have been implicated in coordinate bond formationto metal ions [35]. It follows that the metal complexing abilitiesof E1 were due to the presence of side chain residues of E, D, T, Rand K. Fe2+ chelation, in particular, has been reported to occurthrough the carboxylate groups of E [36] present in the peptideE1 with a relative percentage occurrence of 8.3%. There is a chancethat carboxylate functional groups of E1 may also be involved indirect Cr6+ chelation as suggested by some recent studies [37,38].However, complex formation with Hg2+ requires the presence ofC and H residues [39] and the absence of such residues in E1 wouldpossibly restrain it from direct Hg2+ chelation.
Antioxidative therapeutics complemented with chelatingagents has already been reported to be an effective treatmentagainst acute metal toxicity. Combinatorial therapies includingtreatment with antioxidants like N-acetyl cysteine (NAC), lipoicacid, melatonin and gossypin have shown considerable promisein decreasing clinical pathologies related to ROS stress [33].NAC is known to have free radical scavenging properties andthe thiol groups provide complexation sites for metal ions.Co-administration of several antioxidative and metal chelatingagents along with vitamins has led to reversal of the stressaltered parameters. Using antioxidative peptide provides a betteralternative as it combines the antioxidative and the metal chela-tion activities in one agent, making administration simpler.Moreover, the presence of a variety of functional groups in thepeptide impart high solubility in both aqueous and lipid medium.Peptides isolated from fish skin collagen are known to offercyto-protection to a wide array of cell lines including keratino-cytes, fibroblasts and macrophages [40,41].
3.4.2. De-stressing through cell adhesionIntegrin-ECM interaction has been found to be necessary for cell
survival in the event of stress. The dynamic cytoskeleton is aninterconnected continuous network inside cells capable of trans-mitting stress from ECM and other cells. A balance between tensilefilaments and resistive focal points like focal adhesive kinases(FAK) and cellular junctions determines the overall cell shape. Anoutside interference, like the presence of excessive stress can leadto loss of cell-matrix adhesion and a shift in the cellular balance offorces, resulting in changes in cell shape and detachment [10,42].Cell-matrix attachment is a critical factor in cell survival and theloss of which leads to metabolic stress characterized by reducednutrient uptake, decreased entry of glucose into citric acid cycle,decreased ATP production and further increase in ROS, ultimatelyleading to cell division suspension and the activation of pro-apop-totic pathways [43]. ROS induced cell death progresses throughseveral pathways including direct caspase activation, necrosisthrough poly-(ADP-Ribose) polymerase over-activation, activationof sphingomyelinase-generating ceramide and alteration in mito-chondrial redox balance, resulting in release of cytochrome c andthe formation of apoptosome [44,45].
Peptide coating ensured significantly higher cell survival com-pared to CCS, as is evident from Figs. 2–4, emphasizing the roleof cell adhesion in countering stress. Suitable coating materials,particularly those mimicking the ECM environment, improve theadhesion rate due to a firm cell-coated material interaction, conse-quently leading to faster cell spreading. Peptides C2 and E1, by vir-tue of their small size, supplies a relatively greater number ofintegrin recognition domains per cm2, resulting in superior celladhesion compared to collagen [46,47]. Moreover, due to its largesize, collagen would be adsorbed on the dish surface in a randomorientation, resulting in even lesser number of recognizable inte-grin recognition sites per cm2 in comparison to the peptides [48].A firm ECM-integrin attachment increases integrin clustering, has-tening up the focal adhesion sites (FAS) formation. Once FAS isformed, a number of cellular signals including anti-apoptotic, cellspreading and proliferation rate could be effected, all eventuallyleading to higher survival rates [49].
In the presence of low amounts of stress, the activity of E1 andC2 in coated form is almost similar (p > 0.05) but the differencewidens (p < 0.002) in the presence of higher concentration ofstressors, particularly in the case of Fe2+ (Figs. 2b and 3b) and/or

P. Banerjee et al. / Chemico-Biological Interactions 211 (2014) 1–10 7
H2O2 (Figs. 4 and 5) systems. This difference in the efficacy of E1and C2 towards countering specifically redox stress might meanthese peptides initiate different signalling patterns. RGD-contain-ing peptides like E1 interact with aVb3, aVb5, aVb6 and aVb8subtypes of integrin receptor while C2 is thought to interact withthe collagen-binding integrin subtypes: a1b1, a2b1, a10b1 anda11b1 [50]. Integrins preserve cell viability in response to stressat several levels by regulating both the expression and activity ofseveral anti-apoptotic proteins. The ligation of either a5b1 oravb3-integrins or the RGD-binding integrins have been reportedto inhibit apoptosis through a wide variety of stimulations; twomajor pathways include the PI3-K/PKB and the Raf/MEK/ERKsignalling cascade [51]. The downstream signalling leads to ofdeactivation of pro-apoptotic proteins and activation of the Bcl-2family of anti-apoptotic proteins, resulting in inhibition of poreformation in the mitochondria. Ligand-integrin-FAK formationleads to activation of Hsp 2, another anti-apoptotic protein whichinactivates caspases and prevents cytochrome c release. Last butnot the least, it results in the up-regulation of IAP (inhibitors ofapoptotic pathway) class of proteins, which perform a plethoraof functions, including inactivating executioner caspases andregulating cell division [27,51].
3.5. Wound closure monolayer assay
The ‘scratch assay’ is a direct and inexpensive process to studywound healing in vitro [24]. The assay is based on the fact that cre-ation of an artificial ‘scratch’ or a gap on a fully confluent cellmonolayer will cause the cells at the edges of the gap to move to-wards each other for closing the gap and re-establishing contact.The assay was conducted with Vero cell lines because of its fasterdoubling time in comparison to HeLa. The photographs of woundclosure under various conditions are depicted in Fig. 7. The cell
0 3 6 9 a
b
c
d
Fig. 7. Photographs of a wound area showing Vero cell migration for 24 h in the following(Test, Set I), (c) C2 coated in presence of H2O2 (Set II) and (d) C2 coated/E1 dissolved alonhour at which the photograph was taken. The bar in the bottom right photomicrograph
migration was measured by calculation of % area covered by the in-ward moving cells in certain time duration.
In the absence of stress, (Set I; Fig. 8a) the migration of the cellswas significantly faster on peptide coated dishes compared to col-lagen coated ones (p < 0.02) although in general, gap closure oncoated dishes was faster than uncoated ones (p < 0.005). Cellsgrowing on E1 covered 77% of the area within 3 h of the woundcreation and 100% within 6 h. C2 displayed 59% coverage in 3 hand 99.6% in 9 h. Cells growing on collagen closed the wound com-pletely within 12 h whereas cells in uncoated dishes took 24 h todo so.
In the presence of oxidative stress (Set II; Fig. 8b), the uncoateddishes were unable to promote any cell adhesion. Collagen coateddishes exhibited live migrating cells but the gap was only partiallyclosed even after 24 h. On the other hand, peptide coated dishesdisplayed live migrating cells and even though an initial delaywas observed, the gap was closed within 24 h.
Wound closure is a multi-component synergistic effect. To mi-mic the various synergistic effects of the peptides occurring inthe in vitro model of wound healing, E1 in the dissolved formwas added as an additional de-stressor (Set III; Fig. 8c). As is evi-dent from the figure, this had a significant effect on cell survivaland movement; the gap closure rates for C2- and collagen-coateddishes were found to increase (p < 0.04) when compared to Set II.
Interestingly, gap closure in C2-coated/E1-dissolved dishes wasfound to be significantly faster (p < 0.05) than collagen-coated/E1-dissolved dishes. This could be attributed to the synergistic effectsof E1 in dissolved form complementing the activity of C2 in coatedform. E1 in the dissolved form acted as an efficient ROS de-stressor,removing free radicals whereas C2 in the coated form actedthrough integrin receptors to maintain adhesion and as a conse-quence, resulted in faster wound closure. However, gap closurerate in E1-coated/E1-dissolved dishes was significantly lower(p < 0.02) than collagen/E1 and C2/E1 combinations, probably
12 18 24
situations: (a) Uncoated, without stress (Control, Set I), (b) C2 coated, without stressg with H2O2 (Set III). The numbers 0, 3, 6, 9, 12, 18 and 24 at the top represent therepresents a length of 0.1 mm.
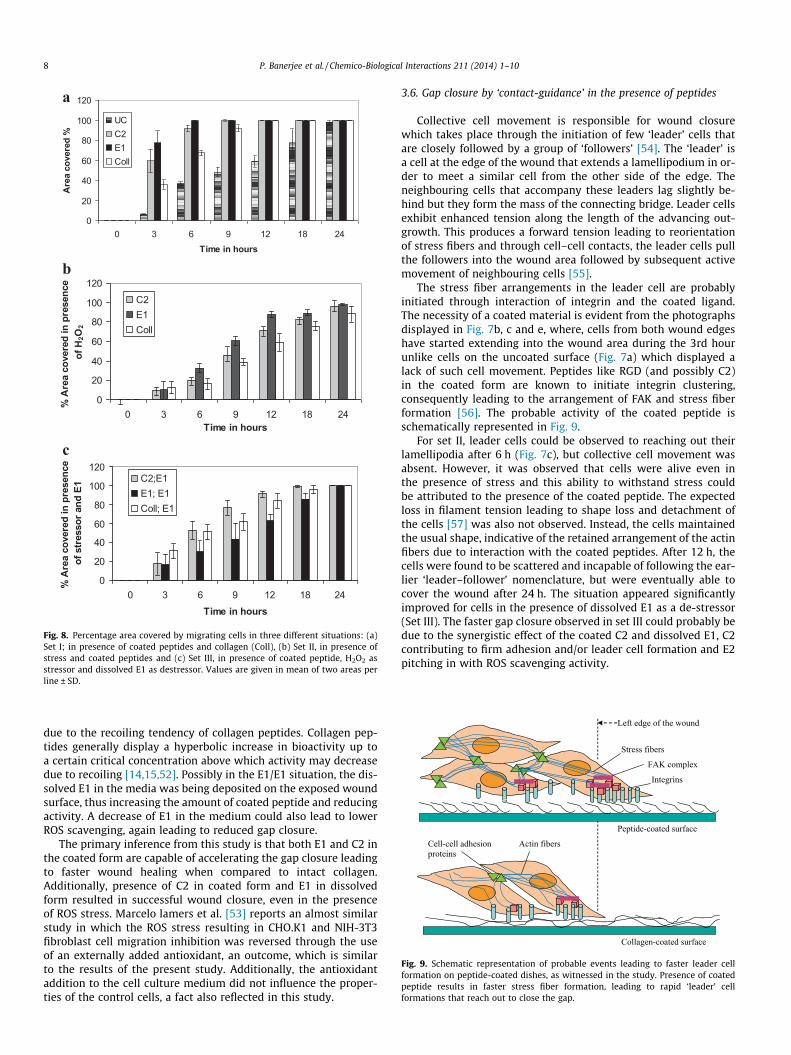
0
20
40
60
80
100
120
0 3 6 9 12 18 24Time in hours
Are
a co
vere
d %
UCC2E1Coll
b
a
0
20
40
60
80
100
120
0 3 6 9 12 18 24Time in hours
% A
rea
cove
red
in p
rese
nce
of H
2O2
C2E1Coll
c
0
20
40
60
80
100
120
0 3 6 9 12 18 24Time in hours
% A
rea
cove
red
in p
rese
nce
of s
tres
sor a
nd E
1 C2;E1E1; E1Coll; E1
Fig. 8. Percentage area covered by migrating cells in three different situations: (a)Set I; in presence of coated peptides and collagen (Coll), (b) Set II, in presence ofstress and coated peptides and (c) Set III, in presence of coated peptide, H2O2 asstressor and dissolved E1 as destressor. Values are given in mean of two areas perline ± SD.
Left edge of the wound
FAK complex
Integrins
Cell-cell adhesion proteins
Stress fibers
Actin fibers
Peptide-coated surface
Collagen-coated surface
Fig. 9. Schematic representation of probable events leading to faster leader cellformation on peptide-coated dishes, as witnessed in the study. Presence of coatedpeptide results in faster stress fiber formation, leading to rapid ‘leader’ cellformations that reach out to close the gap.
8 P. Banerjee et al. / Chemico-Biological Interactions 211 (2014) 1–10
due to the recoiling tendency of collagen peptides. Collagen pep-tides generally display a hyperbolic increase in bioactivity up toa certain critical concentration above which activity may decreasedue to recoiling [14,15,52]. Possibly in the E1/E1 situation, the dis-solved E1 in the media was being deposited on the exposed woundsurface, thus increasing the amount of coated peptide and reducingactivity. A decrease of E1 in the medium could also lead to lowerROS scavenging, again leading to reduced gap closure.
The primary inference from this study is that both E1 and C2 inthe coated form are capable of accelerating the gap closure leadingto faster wound healing when compared to intact collagen.Additionally, presence of C2 in coated form and E1 in dissolvedform resulted in successful wound closure, even in the presenceof ROS stress. Marcelo lamers et al. [53] reports an almost similarstudy in which the ROS stress resulting in CHO.K1 and NIH-3T3fibroblast cell migration inhibition was reversed through the useof an externally added antioxidant, an outcome, which is similarto the results of the present study. Additionally, the antioxidantaddition to the cell culture medium did not influence the proper-ties of the control cells, a fact also reflected in this study.
3.6. Gap closure by ‘contact-guidance’ in the presence of peptides
Collective cell movement is responsible for wound closurewhich takes place through the initiation of few ‘leader’ cells thatare closely followed by a group of ‘followers’ [54]. The ‘leader’ isa cell at the edge of the wound that extends a lamellipodium in or-der to meet a similar cell from the other side of the edge. Theneighbouring cells that accompany these leaders lag slightly be-hind but they form the mass of the connecting bridge. Leader cellsexhibit enhanced tension along the length of the advancing out-growth. This produces a forward tension leading to reorientationof stress fibers and through cell–cell contacts, the leader cells pullthe followers into the wound area followed by subsequent activemovement of neighbouring cells [55].
The stress fiber arrangements in the leader cell are probablyinitiated through interaction of integrin and the coated ligand.The necessity of a coated material is evident from the photographsdisplayed in Fig. 7b, c and e, where, cells from both wound edgeshave started extending into the wound area during the 3rd hourunlike cells on the uncoated surface (Fig. 7a) which displayed alack of such cell movement. Peptides like RGD (and possibly C2)in the coated form are known to initiate integrin clustering,consequently leading to the arrangement of FAK and stress fiberformation [56]. The probable activity of the coated peptide isschematically represented in Fig. 9.
For set II, leader cells could be observed to reaching out theirlamellipodia after 6 h (Fig. 7c), but collective cell movement wasabsent. However, it was observed that cells were alive even inthe presence of stress and this ability to withstand stress couldbe attributed to the presence of the coated peptide. The expectedloss in filament tension leading to shape loss and detachment ofthe cells [57] was also not observed. Instead, the cells maintainedthe usual shape, indicative of the retained arrangement of the actinfibers due to interaction with the coated peptides. After 12 h, thecells were found to be scattered and incapable of following the ear-lier ‘leader–follower’ nomenclature, but were eventually able tocover the wound after 24 h. The situation appeared significantlyimproved for cells in the presence of dissolved E1 as a de-stressor(Set III). The faster gap closure observed in set III could probably bedue to the synergistic effect of the coated C2 and dissolved E1, C2contributing to firm adhesion and/or leader cell formation and E2pitching in with ROS scavenging activity.

P. Banerjee et al. / Chemico-Biological Interactions 211 (2014) 1–10 9
4. Conclusion
The hypothesis that cryptic peptide fragments could impartcyto-protection to cells under stress was proved through thisstudy. E1 in the dissolved form was capable of enhancing cell sur-vival even at high stressor concentrations by virtue of its antioxida-tive properties. C2 in the coated form and in combination with E1in the dissolved form also displayed cyto-protective abilities andrapid wound closure, particularly when subjected to ROS stress.Wound healing is a complex hierarchy of chemical stimulationsand their physiological effects on cells. During collagen remodel-ling during stress or skin wound healing, the incoming fibroblastsdegrade the basal collagen leading to release of several cryptic pep-tides which acts as physiological modulators. This study empha-sizes the probable role of collagen type I cryptic peptides inacting as de-stressor agents during events which may generatestress and assisting in cell adhesion and wound healing.
Conflict of interest
The authors declare no conflict of interest.
Acknowledgements
The authors wish to thank the Department of Science and Tech-nology, Government of India for funding this study. The authors arealso grateful to Vellore Institute of Technology University, wherethe work was carried out.
References
[1] N. Gjorevski, C.M. Nelson, Bidirectional extracellular matrix signaling duringtissue morphogenesis, Cytokine Growth Factor Rev. 20 (2009) 459–465.
[2] D.J. Autelitano, A. Rajic, A.I. Smith, M.C. Berndt, L.L. Ilag, M. Vadas, Thecryptome: a subset of the proteome, comprising cryptic peptides with distinctbioactivities, Drug Discovery Today 11 (2006) 306–314.
[3] S. Schenk, V. Quaranta, Tales from the crypt[ic] sites of the extracellular matrix,Trends Cell Biol. 13 (2003) 366–375.
[4] F. Shahidi, Y. Zhong, Bioactive peptides, J. AOAC Int. 91 (2008) 914–931.[5] H.-G. Byun, J.K. Lee, H.G. Park, J.-K. Jeon, S.-K. Kim, Antioxidant peptides
isolated from the marine rotifer, Brachionus rotundiformis, Process Biochem. 44(2009) 842–846.
[6] G.D. Kamphaus, P.C. Colorado, D.J. Panka, H. Hopfer, R. Ramchandran, A. Torre,Y. Maeshima, J.W. Mier, V.P. Sukhatme, R. Kalluri, Canstatin, a novel matrix-derived inhibitor of angiogenesis and tumor growth, J. Biol. Chem. 275 (2000)1209–1215.
[7] P. Banerjee, C. Shanthi, Isolation of novel bioactive regions from bovineAchilles tendon collagen having angiotensin I-converting enzyme-inhibitoryproperties, Process Biochem. 47 (2012) 2335–2346.
[8] J. Park, J. Lee, C. Choi, Mitochondrial network determines intracellular ROSdynamics and sensitivity to oxidative stress through switching inter-mitochondrial messengers, PLoS ONE 6 (2011) e23211.
[9] M.F. Mian, C. Kang, S. Lee, J.H. Choi, S.S. Bae, S.-H. Kim, Y.-H. Kim, S.H. Ryu, P.-G.Suh, J.-S. Kim, E. Kim, Cleavage of focal adhesion kinase is an early marker andmodulator of oxidative stress-induced apoptosis, Chem. Biol. Interact. 171(2008) 57–66.
[10] H. Lum, K.A. Roebuck, Oxidant stress and endothelial cell dysfunction, Am. J.Physiol. Cell Physiol. 280 (2001) C719–741.
[11] F. Lamari, F. Braut-Boucher, N. Pongnimitprasert, M. Bernard, M.-J. Foglietti, C.Derae, M. Aubery, Cell adhesion and integrin expression are modulated byoxidative stress in EA.hy 926 cells, Free Radic. Res. 41 (2007) 812–822.
[12] L. Zhou, Y. Li, B.Y. Yue, Oxidative stress affects cytoskeletal structure and cell-matrix interactions in cells from an ocular tissue: the trabecular meshwork, J.Cell. Physiol. 180 (1999) 182–189.
[13] N. Ercal, H. Gurer-Orhan, N. Aykin-Burns, Toxic metals and oxidative stresspart I: mechanisms involved in metal-induced oxidative damage, Curr. Top.Med. Chem. 1 (2001) 529–539.
[14] P. Banerjee, G. Suseela, C. Shanthi, Isolation and identification of crypticbioactive regions in bovine Achilles tendon collagen, Protein J. 31 (2012) 374–386.
[15] P. Banerjee, A. Mehta, C. Shanthi, Screening for novel cell adhesive regions inbovine Achilles tendon collagen peptides, Biochem. Cell Biol. (2013) (In print).
[16] M. Radhika, P.K. Sehgal, Studies on the desamidation of bovine collagen, J.Biomed. Mater. Res. 35 (1997) 497–503.
[17] J. García, M.D. Vega, D. Boettiger, Modulation of cell proliferation anddifferentiation through substrate-dependent changes in fibronectinconformation, Mol. Biol. Cell 10 (1999) 785–798.
[18] N. C. Ammerman, M. Beier-Sexton, A. F. Azad, Growth and maintenance of Verocell lines, Curr. Protoc. Microbiol. Appendix 4 (2008) Appendix 4E.
[19] J. Zhang, H. Zhang, L. Wang, X. Guo, X. Wang, H. Yao, Isolation andidentification of antioxidative peptides from rice endosperm proteinenzymatic hydrolysate by consecutive chromatography and MALDI-TOF/TOFMS/MS, Food Chem. 119 (2010) 226–234.
[20] G. Levis, F. Majone, Cytotoxic and clastogenic effects of soluble and insolublecompounds containing hexavalent and trivalent chromium, Br. J. Cancer 44(1981) 219–235.
[21] T. Jungas, I. Motta, F. Duffieux, P. Fanen, V. Stoven, D.M. Ojcius, Glutathionelevels and BAX activation during apoptosis due to oxidative stress in cellsexpressing wild-type and mutant cystic fibrosis transmembrane conductanceregulator, J. Biol. Chem. 277 (2002) 27912–27918.
[22] B. Hultberg, A. Andersson, A. Isaksson, Interaction of metals and thiols in celldamage and glutathione distribution: potentiation of mercury toxicity bydithiothreitol, Toxicology 156 (2001) 93–100.
[23] V. Houot, P. Etienne, A.S. Petitot, S. Barbier, J.P. Blein, L. Suty, Hydrogenperoxide induces programmed cell death features in cultured tobacco BY-2cells, in a dose-dependent manner, J. Exp. Bot. 52 (2001) 1721–1730.
[24] C.-C. Liang, A.Y. Park, J.-L. Guan, In vitro scratch assay: a convenient andinexpensive method for analysis of cell migration in vitro, Nat. Protoc. 2 (2007)329–333.
[25] A. Valster, N.L. Tran, M. Nakada, M.E. Berens, A.Y. Chan, M. Symons, Cellmigration invasion assays, Methods 37 (2005) 208–215.
[26] K. Choi, J. Kim, G.W. Kim, C. Choi, Oxidative stress-induced necrotic cell deathvia mitochondria-dependent burst of reactive oxygen species, Curr. Neurovasc.Res. 6 (2009) 213–222.
[27] D.R. Gough, T.G. Cotter, Hydrogen peroxide: a Jekyll and Hyde signallingmolecule, Cell Death Dis. 2 (2011) e213.
[28] G.R. Borthiry, W.E. Antholine, B. Kalyanaraman, J.M. Myers, C.R. Myers,Reduction of hexavalent chromium by human cytochrome b5: generation ofhydroxyl radical and superoxide, Free Radic. Biol. Med. 42 (2007) 738–755.
[29] K.P. Nickens, S.R. Patierno, S. Ceryak, Chromium genotoxicity: a double-edgedsword, Chem. Biol. Interact. 188 (2010) 276–288.
[30] Y.W. Lee, M.S. Ha, Y.K. Kim, Role of reactive oxygen species and glutathione ininorganic mercury-induced injury in human glioma cells, Neurochem. Res. 26(2001) 1187–11931.
[31] C.C. Winterbourn, Toxicity of iron and hydrogen peroxide: the Fenton reaction,Toxicol. Lett. 82–83 (1995) 969–974.
[32] R.F. Vitor, S. Alves, J.D.G. Correia, A. Paulo, I. Santos, Rhenium(I)- andtechnetium(I) tricarbonyl complexes anchored by bifunctional pyrazole-diamine and pyrazole-dithioether chelators, J. Organomet. Chem. 689 (2004)4764–4774.
[33] S.J.S. Flora, V. Pachauri, Chelation in metal intoxication, Int. J. Environ. Res.Public Health 7 (2010) 2745–2788.
[34] F. Zidane, A. Matéos, C. Cakir-Kiefer, L. Miclo, S. Rahuel-Clermont, J.-M.Girardet, C. Corbier, Binding of divalent metal ions to 1–25 b-caseinophosphopeptide: an isothermal titration calorimetry study, FoodChem. 132 (2012) 391–398.
[35] E. Farkas, I. Sóvágó, Metal complexes of amino acids and peptides, in: J.S.Davies (Ed.), Amino acids, peptides and proteins, Royal Society of Chemistry,Cambridge, 2001, pp. 353–434. Chapter 5.
[36] Y. Lv, Q. Liu, X. Bao, W. Tang, B. Yang, S. Guo, Identification and characteristicsof iron-chelating peptides from soybean protein hydrolysates using IMAC-Fe3+, J. Agric. Food Chem. 57 (2009) 4593–4597.
[37] R. Bala, A.K. Thukral, Phytoremediation of Cr(VI) by Spirodela polyrrhiza (L.)Schleiden employing reducing and chelating agents, Int. J. Phytoremediat. 13(2011) 465–491.
[38] E. Molina-Jijón, G. Zarco-Márquez, O.N. Medina-Campos, Z.L. Zataraín-Barrón,R. Hernández-Pando, E. Pinzón, R.M. Zavaleta, E. Tapia, J. Pedraza-Chaverri,Deferoxamine pretreatment prevents Cr(VI)-induced nephrotoxicity andoxidant stress: role of Cr(VI) chelation, Toxicology 291 (2012) 93–101.
[39] M. Ngu-Schwemlein, J.K. Merle, P. Healy, S. Schwemlein, S. Rhodes,Thermodynamics of the complexation of Hg(II) by cysteinyl peptide ligandsusing isothermal titration calorimetry, Thermochim. Acta 496 (2009) 129–135.
[40] D.-H. Ngo, Z.-J. Qian, B. Ryu, J.W. Park, S.-K. Kim, In vitro antioxidant activity ofa peptide isolated from Nile tilapia (Oreochromis niloticus) scale gelatin in freeradical-mediated oxidative systems, J. Funct. Foods 2 (2010) 107–117.
[41] S. Kato, H. Matsui, Y. Saitoh, N. Miwa, Fish collagen-containing drink issubcutaneously absorbed and attenuates the UVA-induced tissue-integritydestruction and DNA damages in 3D-human skin tissue model, J. Funct. Foods3 (2011) 50–55.
[42] J.V. Zuk, D. Bonventre, K.S. Brown, Matlin, Polarity, integrin, and extracellularmatrix dynamics in the postischemic rat kidney, Am. J. Physiol. 275 (1998)C711–731.
[43] R. Grassian, J.L. Coloff, J.S. Brugge, Extracellular matrix regulation ofmetabolism and implications for tumorigenesis, Cold Spring Harb. Symp.Quant. Biol. 76 (2011) 313–324.
[44] W. Fiers, R. Beyaert, W. Declercq, P. Vandenabeele, More than one way to die:apoptosis, necrosis and reactive oxygen damage, Oncogene 18 (1999) 7719–7730.
[45] H.C. Ha, S.H. Snyder, Poly (ADP-ribose) polymerase is a mediator of necroticcell death by ATP depletion, Proc. Natl. Acad. Sci. U.S.A. 96 (1999) 13978–13982.

10 P. Banerjee et al. / Chemico-Biological Interactions 211 (2014) 1–10
[46] Z. Ma, W. He, T. Yong, S. Ramakrishna, Grafting of gelatin on electrospunpoly(caprolactone) nanofibers to improve endothelial cell spreading andproliferation and to control cell Orientation, Tissue Eng. 11 (2005) 1149–1158.
[47] E.A. Cavalcanti-Adam, T. Volberg, A. Micoulet, H. Kessler, B. Geiger, J.P. Spatz,Cell spreading and focal adhesion dynamics are regulated by spacing ofintegrin ligands, Biophys. J. 92 (2007) 2964–2974.
[48] J.B. Lhoest, E. Detrait, P. van den Bosch de Aguilar, P. Bertrand, Fibronectinadsorption, conformation, and orientation on polystyrene substrates studiedby radiolabeling, XPS, and ToF SIMS, J. Biomed. Mater. Res. 41 (1998) 95–103.
[49] J.E. Meredith Jr., M.A. Schwartz, Integrins, adhesion and apoptosis, Trends CellBiol. 7 (1997) 146–150.
[50] G. Knight, L.F. Morton, A.R. Peachey, D.S. Tuckwell, R.W. Farndale, M.J. Barnes,The collagen-binding A-domains of integrins a1b1 and a2b1recognize thesame specific amino acid sequence, GFOGER, in native (triple-helical)collagens, J. Biol. Chem. 275 (2000) 35–40.
[51] G. Stupack, D.A. Cheresh, Get a ligand, get a life: integrins, signaling and cellsurvival, J. Cell. Sci. 115 (2002) 3729–3738.
[52] N. Saeidi, E.A. Sander, J.W. Ruberti, Dynamic shear-influenced collagen self-assembly, Biomaterials 30 (2009) 6581–6592.
[53] M.L. Lamers, M.E.S. Almeida, M. Vicente-Manzanares, A.F. Horwitz, M.F.Santos, High glucose-mediated oxidative stress impairs cell migration, PLoSONE 6 (2011) e22865.
[54] H. Haga, C. Irahara, R. Kobayashi, T. Nakagaki, K. Kawabata, Collectivemovement of epithelial cells on a collagen gel substrate, Biophys. J. 88(2005) 2250–2256.
[55] T. Omelchenko, J.M. Vasiliev, I.M. Gelfand, H.H. Feder, E.M. Bonder, Rho-dependent formation of epithelial ‘leader’ cells during wound healing, Proc.Natl. Acad. Sci. U.S.A. 100 (2003) 10788–10793.
[56] K. Kawakami, H. Tatsumi, M. Sokabe, Dynamics of integrin clustering at focalcontacts of endothelial cells studied by multimode imaging microscopy, J. CellSci. 114 (2001) 3125–3135.
[57] S. Tojkander, G. Gateva, P. Lappalainen, Actin stress fibers-assembly, dynamicsand biological roles, J. Cell. Sci. 125 (2012) 1855–1864.
Copyright © 2022 FDOKUMEN




















