
Base de données de capteurs à localisation mobile Modèle et langage
Upload
khangminh22Category
view
0download
0
1
MUSEUM NATIONAL D’HISTOIRE NATURELLE Ecole Doctorale Sciences de la Nature et de l’Homme – ED 227
Année 2017 N°attribué par la bibliothèque
|_|_|_|_|_|_|_|_|_|_|_|_|
THESE
Pour obtenir le grade de
DOCTEUR DU MUSEUM NATIONAL D’HISTOIRE NATURELLE
Spécialité : Parasitologie/Immunologie
Présentée et soutenue publiquement par
Frédéric FERCOQ
Le 29 Septembre 2017
Interactions filaire/poumon dans le modèle murin de filariose
Litomosoides sigmodontis
Sous la direction du Dr. Coralie MARTIN
JURY :
Nom, prénom Qualité Lieu d’exercice
Dr. MILON Geneviève DR Institut Pasteur, Paris Président du jury
Dr. WOLOWCZUK Isabelle DR2 UMR8204-U1019 – CII, Lille Rapporteur
Dr. DELHAES Laurence PU-PH CHU de Bordeaux Rapporteur
Dr. HUEBNER Marc CR (Allemagne) IMMIP, Bonn Examinateur
Dr. LANDMANN Frédéric CR1 UMR5237 – CRBM, Montpellier Examinateur
Dr. MARTIN Coralie CR1 UMR 7245 – MNHN, Paris Directeur de Thèse

2

3

4
Remerciements Je tiens tout d’abord à remercier les membres du jury d’avoir accepté d’évaluer le
travail effectué au cours de cette thèse.
Un grand merci ensuite :
A ma directrice de thèse, le Dr. Coralie Martin, pour m’avoir accueilli au sein du
laboratoire et encadré durant ces trois années, me permettant d’acquérir de nombreuses
compétences.
Aux Dr. Marc Huebner et Leo Carlin de m’avoir accueilli et guidé à plusieurs reprises,
dans leur laboratoire respectif, permettant ainsi l’initiation et le déploiement durable de
fructueuses collaborations
A Grégory qui, par ses résultats de thèse, non seulement a permis d’initier cette thèse
mais en outre, a transmis de nombreuses connaissances nécessaires à la mise en œuvre rapide
d’approches expérimentales délicates
A Emilie pour les pauses thé/café/bière qui ont ensoleillé mes deux premières années.
Merci aussi pour ses conseils inestimables en biologie moléculaire.
A Géraldine pour son aide précieuse avec la microscopie électronique à balayage et
Françoise pour les conseils en histologie.
A Joy pour s’être bien occupée de mes hordes de souris (et fait le café du matin).
A Christiane pour les moments passées à refaire le monde (et à parler science aussi !).
A Nathaly pour son aide précieuse et ses histoires rocambolesques.
A Estelle pour son aide lors de la dernière ligne droite.
Et à toutes les personnes de l’équipe : Linda & Linda, Brice, Dhikra, Lotfi,
Amandine... qui m'ont permis de passer trois années mémorables.
Enfin, je tiens à remercier Julie qui m'a toujours soutenu.

5
Table des matières
Remerciements ......................................................................................................................... 4
Table des figures ..................................................................................................................... 12
Liste et titre des tableaux ....................................................................................................... 15
Table des Annexes .................................................................................................................. 16
Liste des abréviations ............................................................................................................. 17
Résumé .................................................................................................................................... 20
Abstract ................................................................................................................................... 21
INTRODUCTION .................................................................................................................... 23
1. Filaires ................................................................................................................................. 25
1.1 Diversité des filaires ....................................................................................................... 25
1.2 Biologie .......................................................................................................................... 25
1.3 Programme de développement ...................................................................................... 27
1.4 Wolbachia ....................................................................................................................... 28
2. Filarioses humaines ............................................................................................................ 29
2.1 Epidémiologie et coendémicité ...................................................................................... 31
2.2 Programme de développement ...................................................................................... 33
2.2.1 Les filaires lymphatiques : Wuchereria bancrofti, Brugia/B. malayi et B. timori .. 33
2.2.2 Onchocerca volvulus ............................................................................................... 34
2.2.3 Loa loa ..................................................................................................................... 34
2.2.4 Les mansonelles ...................................................................................................... 35
2.3 Principales manifestations cliniques .............................................................................. 36
2.3.1 Filarioses lymphatiques ........................................................................................... 36
2.3.2 Onchocercose .......................................................................................................... 41
2.3.3 Loase ....................................................................................................................... 44
3. Les filarioses pulmonaires, cardio-pulmonaires et pleurales ......................................... 48
3.1 Dirofilaria immitis .......................................................................................................... 49
3.2 Mansonella perstans ...................................................................................................... 56
3.3 Litomosoides sigmodontis .............................................................................................. 57

6
4. Programmes multivalents visant à « traiter » les filarioses .... Erreur ! Signet non défini.
4.1 Traitements à grande échelle ou traitements de masse (Mass Drug Administration,
MDA) ................................................................................................................................... 61
4.1.2 Programmes de lutte contre l'onchocercose ............................................................ 63
4.1.3 Les traitements individuels ...................................................................................... 63
4.1.4 Considérations actuelles .......................................................................................... 64
4.2 Mode d'action des molécules ciblant les filaires ........................................................... 65
4.2.2 Diéthylcarbamazine ................................................................................................. 67
4.2.3 Albendazole ............................................................................................................. 67
4.2.4 Tétracyclines ........................................................................................................... 69
4.2.5 Traitements en cours d'évaluation ........................................................................... 69
5. Cinétique de la « réponse immunitaire » chez les souris infestées par L. sigmodontis . 71
5.1 « Réponse immunitaire » au site d'inoculation : apport des différents modèles ........ 73
5.2 Migration des larves de L. sigmodontis vers la cavité pleurale .................................. 74
5.3 « Réponses immunitaires » dans la cavité pleurale lors de la phase pré-patente ....... 75
5.4 « Réponse » dans la cavité pleurale au cours de la phase patente .............................. 77
6. Rappels anatomiques/histologiques sur les poumons, le cœur et les systèmes
vasculaires .............................................................................................................................. 81
6.1 Plèvre et mésothélium .................................................................................................... 81
6.2 Le poumon, les bronches, les bronchioles et les cellules alvéolaires de type I et II : .... 83
6.3 Vaisseaux sanguins et lymphatiques pulmonaires ......................................................... 85
6.3.1 Vascularisation sanguine pulmonaire:..................................................................... 85
6.3.2 Les lymphatiques des poumons et de la plèvre : le système pleuropulmonaire ...... 87
6.4 Espace périvasculaire ..................................................................................................... 90
6.5 Les macrophages pulmonaires ....................................................................................... 91
7. Dommages pulmonaires due aux anchylostomes ...................... Erreur ! Signet non défini.
7.1 Cycle de vie des anchylostomes ..................................................................................... 95
7.2 Neutrophiles et amplification de la pathologie pulmonaire. .......................................... 97
7.3 Macrophages : contrôle de sous populations de parasites et réparation des tissus......... 97
OBJECTIFS DE LA THESE ................................................................................................... 99

7
MATERIEL ET METHODES ............................................................................................... 101
1. Ethique .............................................................................................................................. 102
2. Parasites, souris et inoculation des larves /L3 de L. sigmodontis ................................. 102
2.1 Maintien du cycle de L. sigmodontis pour obtention des larves L3 ............................. 102
2.2 Les souris de laboratoire............................................................................................... 103
2.2.1 Souris BALB/c ...................................................................................................... 103
2.2.2 Souris BALB/c IL4R/IL5 KO ............................................................................... 103
2.2.3 Souris C57BL/6 S100A9-/- .................................................................................... 103
2.3 Inoculation aux souris .................................................................................................. 103
2.4 Suivi de la présence ou non de microfilarires chez les souris .................................... 104
3. Autopsie des souris, collecte des filaires, prélèvement et fixation des poumons. ........ 104
3.1 Exsanguination et préparation du sérum ...................................................................... 104
3.2 Lavage de la cavité pleurale ......................................................................................... 104
3.3 Evaluation du développement filarien .......................................................................... 105
3.4 Lavage bronchoalvéolaire ............................................................................................ 105
3.5 Dénombrement des L3 dans les poumons .................................................................... 106
3.6 Préparation et fixation des poumons pour différentes analyses. .................................. 106
4. Les analyses histologiques et immunohistologique.sur coupes fines 105
4.1 Déshydratation, inclusion en paraffine, préparation des lames .................................... 107
4.2 Colorations histologiques ............................................................................................. 108
4.2.1 Coloration à l'Hémalun-Eosine (HE) .................................................................... 108
4.2.2 Coloration au Trichrome de Masson ..................................................................... 109
4.2.3 Coloration à l'Acide Periodique-Schiff (PAS) ...................................................... 111
4.2.4 Colorations immunohistologiques avec révélation enzymatique .......................... 111
4.2.5 Marquage du S100A9 ............................................................................................ 112
4.2.6 Marquage de la cytokératine 7 .............................................................................. 114
4.2.7 Marquage de la podoplanine ................................................................................. 115
5. Immunohistochimie sur coupes épaisses ........................................................................ 117
5.1 Préparation des coupes épaisses de poumons (Precision Cut Lung Slices ou PCLS) .. 117
5.2 Colorations immunohistologiques en fluorescence ...................................................... 117
6. Analyse des images obtenues par microscopie confocale............................................. 119

8
7. Microscopie électronique à balayage .............................................................................. 120
8. Cytométrie en flux ............................................................................................................ 121
8.1 Préparation des cellules ................................................................................................ 121
8.2 Identification des populations leucocytaires. ............................................................... 121
9. Analyse de transcrits de cytokines inflammatoires dans le poumon ........................... 124
9.1 Extraction des ARN totaux et rétrotranscription .......................................................... 124
9.2 Obtention des profils d'expression cytokiniques .......................................................... 126
9.3 Analyse in silico de réseaux biologiques ..................................................................... 127
9.4 Validation des résultats de l'analyse transcriptionelle. ................................................. 128
9.4.1 Rétrotranscription .................................................................................................. 128
9.4.2 Conception des amorces ........................................................................................ 129
9.4.3 PCR quantitative ................................................................................................... 130
10. Purification des microfilaires à partir de sang de mérions hôtes de L. sigmodontis 131
11. Détection et quantification des microfilaires pulmonaires par PCR quantitative ... 132
11.1 Extraction de l'ADN ................................................................................................... 132
11.2 Préparation de la gamme ............................................................................................ 133
11.3 Détection des microfilaires et normalisation .............................................................. 133
12. Cultures cellulaires ......................................................................................................... 135
12.1 Purification des neutrophiles de souris ...................................................................... 135
12.2 Cultures de neutrophiles de souris ............................................................................ 136
12.3 Culture et stimulation des splénocytes de souris ........................................................ 136
13. Dosage des cytokines et autres molécules inflammatoires par une méthode ELISA137
14. Dosage de l'ADN extracellulaire ................................................................................... 138
15. Analyses statistiques ....................................................................................................... 138

9
RESULTATS ......................................................................................................................... 141
CHAPITRE 1 : La migration des larves infestantes/L3 de Litomosoides sigmodontis chez la
souris BALB/c révèle l’existence de dommages pulmonaires transitoires ... Erreur ! Signet non
défini.
Introduction .......................................................................................................................... 144
Résultats ................................................................................................................................ 147
1. Cinétique de la migration des L3; comparaison de différents modes d'inoculation. . 147
2. Le passage de L3 dans le poumon induit une inflammation caractérisée par des
hémorragies locales, des « granulomes » et une accumulation de neutrophiles dans les
espaces périvasculaires.......... .............................................................................................. 150
3. Des processus inflammatoires physiologiques se déploient quelques heures après
l'inoculation des L3. ......................................................................... Erreur ! Signet non défini.
4. Les neutrophiles murins exposés in vitro à des L3 peuvent produire des « NETs ». . 158
Discussion .............................................................................................................................. 161
CHAPITRE 2 : L'hétérodimère S100A8/A9 - calprotectine- favorise-t-il la survie de
Litomosoides sigmodontis ? .................................................................................................. 171
Introduction .......................................................................................................................... 172
Résultats ................................................................................................................................ 178
1. S100A9, un marqueur de l’inflammation aigue chez les souris BALB/c infestées par L.
sigmodontis ? ......................................................................................................................... 178
1.1 Augmentation précoce des transcrits s100a8 et s100a9 dans le poumon .................... 178
1.2 La protéine S100A9 dans les Lavages Broncho Alvéolaires/LBA et fluides des cavités
pleurales…………………………………………………………………………………..179
1.3 Augmentation du nombre de neutrophiles S100A9+ dans les poumons ..................... 181
2. S100A9, un acteur dans le contrôle de l’établissement des L3 de L. sigmodontis ?
Analyse des souris C57BL/6 déficientes en s100a9 ........................ Erreur ! Signet non défini.
2.1 L’absence de s100a9 se traduit par un moindre établissement des larves L3 ...... Erreur !
Signet non défini.
2.2 L’absence de S100A8/A9 se traduit par une moindre recrutement cellulaire dans le
LBA mais facilite celui dans la cavité pleurale .................................................................. 185
2.3 L’absence de S100A8/A9 augmente le nombre de cellules spléniques ....................... 187

10
2.4 Le profil cytokinique des cellules spléniques est caractérisé par une dominance pro-
inflammatoire en l’absence de S100A8/A9 ....................................................................... 188
2.5 Observation de dommages des vaisseaux sanguins pulmonaires et de processus
inflammatoires bronchiques indépendants du S100A8/A9 chez les souris C57BL/6 ........ 189
2.6 En l’absence de S100A8/A9, quelles fonctions pour les neutrophiles ? ..................... 193
Discussion .............................................................................................................................. 196
CHAPITRE 3 : Dommages pulmonaires prolongés chez les rongeurs chez lesquels est
générée la descendance des microfilaires de Litomosoides sigmodontis ...... Erreur ! Signet non
défini.
Introduction : ........................................................................................................................ 203
Résultats ................................................................................................................................ 206
1. Microfilarémie et dommages pulmonaires chez la gerbille .......................................... 206
2. Analyse cinétique de la phase patente chez la souris BALB/c, localisation des parasites
et distinction des souris en deux groupes selon la présence ou non de microfilaires ..... 210
3. Analyse de profils de biomolécules-transcrits et protéines- au niveau des poumons . 212
4. Production de mucus par l'épithélium bronchique de souris hôtes de microfilaires . 216
5. Les macrophages périvasculaires augmentent chez les souris microfilarémiques. .... 219
6. Dommages du mésothélium pleural chez les souris hôtes de microfilaires ................ 225
7. Inflammation pleurale/bronchoalvéolaire exacerbée chez les souris microfilarémiques.
................................................................................................................................................ 230
8. L'initiation des dommages pulmonaire chez les souris microfilarémiques signe-t-elle
un profil de type 2, ............................................................................ Erreur ! Signet non défini.
Discussion .............................................................................................................................. 239
CONCLUSIONS ET PERSPECTIVES ................................................................................. 250
Valorisation des résultats ......................................................................................................
254

11
Annexes ................................................................................................................................. 260
Bibliographie ......................................................................................................................... 265

12
Table des figures
Figure 1. Diversité des filaires. ................................................................................................ 24
Figure 2. Phylogénie des filaires basée sur une analyse multigénique. ................................... 24
Figure 3. Cycle de vie des filaires et diversité d'hôte et de localisation. .................................. 26
Figure 4. Répartition des filarioses humaines. ......................................................................... 30
Figure 5. Evolution du lymphoedème filarien. ........................................................................ 37
Figure 6. Histopathologie du « poumon eosinophile tropical » .............................................. 38
Figure 7. Hypothèse mécanistique sur la pathogénèse des atteintes oculaires. ........................ 42
Figure 8. Synthèse des travaux sur la localisation de Dirofilaria. immitis. ............................. 50
Figure 9. Hypothèses de migration des stades infestants de D. immitis. .................................. 52
Figure 10. Progression de la dirofilariose pulmonaire chez le chien. ...................................... 54
Figure 11. Cycle de vie de Litosomoides sigmodontis. ............................................................ 58
Figure 12. Structure chimique de l'Ivermectine (A), du Diéthylcarbamazine (B) et de
l'Albendazole (C). ..................................................................................................................... 66
Figure 13. Structure chimique de la Doxycycline (A), du Flubendazole (B), et de
l'Emodepside (C). ..................................................................................................................... 68
Figure 14. Coupe transversale de la cavité cavité thoracique. ................................................. 80
Figure 15. Comparaison de la structure des voies aériennes murines et humaines.................. 82
Figure 16. Structure des alvéoles. ............................................................................................ 84
Figure 17. Circulations sanguines pulmonaire et bronchique. ................................................. 86
Figure 18. Réseau lymphatique de la cavité pleurale et drainage du liquide pleural. .............. 88
Figure 19. Résumé des principales pathologies pulmonaires. ................................................. 94
Figure 20. Réponses pulmonaires lors de l'infestation par Nippostrongylus brasiliensis. ....... 96
Figure 21. Stratégie d'analyse du contenu cellulaire de l'espace périvasculaire. ................... 119
Figure 22. Stratégie de discrimination des populations cellulaires. ....................................... 123
Figure 23. Obtention du cycle seuil (CT) après l'acquisition des PCR quantitatives. ............ 126
Figure 24. Purification des microfilaires sur gradient de Percoll. .......................................... 131
Figure 25. Gamme de détection de l’ADN de microfilaires pulmonaires.............................. 134
Figure 26. Voie de migration suggérée des L3 de L. sigmodontis. ........................................ 145
Figure 27. Cinétique de l’infestation post inoculation des L3au niveau de la peau. .............. 146
Figure 28. Localisation tissulaire des L3 lors de la phase de migration. ............................... 146
Figure 29. Cinétique de l’infestation post inoculation intraveineuse de L3 . ......................... 148
Figure 30. Hémorragies dans le poumon de souris où sont détectables les L3 ..................... 150
Figure 31. Granulomes dans les poumons des souris infestées. ............................................. 151
Figure 32. Présence d’infiltrats neutrophiliques dans l’espace périvasculaire des poumons des
souris infestées. ...................................................................................................................... 152
Figure 33. Quantification des neutrophiles dans les espaces broncho-alvéolaires et pleuraux.
................................................................................................................................................ 153
Figure 34. Quantification des éosinophiles et des macrophages dans les espaces
bronchoalvéolaire et pleural. .................................................................................................. 154
Figure 35. Inflammation pulmonaire précoce transitoire ....................................................... 155
Figure 36. Régulation de l’inflammation pulmonaire ............................................................ 157
Figure 37. Neutrophiles de souris C57BL/6 exposés in vitro à des L3 de L. sigmodontis. .. 158
Figure 38. L3 et Nétose de neutrophiles de souris C57BL/6 exposés à des L3 . ................... 159
Figure 39. Synthèse des voies de migrations des L3 après inoculation soit sous-cutanée (SC)
soit intraveineuse (IV). ........................................................................................................... 162
Figure 40. Lors de la migration des L3 de L. sigmodontis au sein des poumons- espaces
pleuraux compris- des processsus inflammatoires physiologiques opèrent-ils ? ................. 166

13
Figure 41. Les protéines de la famille S100, une famille détectée chez les vertébrés. .......... 173
Figure 42. Effets cellulaires potentiels de l’hétérodimère S100A8/S100A9. ........................ 174
Figure 43. Augmentation précoce et transitoire des transcrits s100a8 / s100a9 au niveau des
poumons de souris BALB/c hôtes de L. sigmodontis. ............................................................ 178
Figure 44. Augmentation précoce et transitoire du S100A9 dans les fluides broncho-
alvéolaires et pleuraux de souris BALB/c ............................................................................. 179
Figure 45. Augmentation précoce et transitoire de neutrophiles exprimant S100A9 dans les
poumons de souris BALB/c .................................................................................................. 181
Figure 46. Succès parasitaire en fonction du mode d’inoculation et de la souris C57BL/6 WT
ou S100A9- ............................................................................................................................ 182
Figure 47. Absence de S100A9 dans les granulomes pulmonaires (BALB/c) ...................... 182
Figure 48. Dénombrement - J12/13- des cellules dans les fluides pleuraux et le LBA ......... 184
Figure 49. Correlation entre le nombre de parasites dans la cavité pleurale et le nombre de
cellules dans le « Lavage Broncho-Alvéolaire (LBA) ». ....................................................... 184
Figure 50. Quantification des macrophages et des neutrophiles pleuraux et alvéolaires. ...... 186
Figure 51. Effectif accru des populations de leucocytes/lymphocytes spléniques chez les
souris C57BL/6 invalidées pour le gène s100A9 hôtes de L3 ............................................... 187
Figure 52. Comparaison de la concentration de cytokines dans les surnageants de cultures de
cellules spléniques .................................................................................................................. 188
Figure 53. Coupes histologiques de poumons de souris C57BL/6 s100A9-/- ......................... 190
Figure 54. Nbre de vaisseaux sanguins inflammés et de larves dans la cavité pleurale ........ 191
Figure 55. Nbre de larves dans la cavité pleurale et % de bronches contenant des cellules
caliciformes ............................................................................................................................ 192
Figure 56. Comparaison in vitro de neutrophiles prélevés de souris s100A9+/+ ou s100A9-/- 194
Figure 57. Anomalie du mésothélium pulmonaire de gerbilles hôtes de L. sigmodontis ...... 205
Figure 58. Structure « fibrotique de nodules » au niveau du mésothèlium de gerbilles ........ 207
Figure 59. Microfilaires à proximité de la plèvre viscérale de gerbilles hôtes de L.sigmodontis
................................................................................................................................................ 208
Figure 60. Dénombrement de L. sigmodontis dans la cavité pleuralep, le cœur et le sang de
souris BALB/c ........................................................................................................................ 209
Figure 61. Granulomes et microfilaires libres au jour 70 chez les souris BALB/c ............... 211
Figure 62. Profils de transcrits au niveau de poumons de souris BALB/c hôtes de
L.sigmodontis ......................................................................................................................... 213
Figure 63. Cytokines et chimiokines dans le LBA et la cp de BALB/c Mfposvs Mf neg ......... 214
Figure 64. Analyse de réseaux cytokiniques et fonctions biologiques potentielles. .............. 215
Figure 65. Présence de cellules PAS+/calicifomes mucus+ chez les souris BALB/c Mfpos ... 217
Figure 66. Cellules PAS+ : bronches/ bronchioles de souris BALB/c mfpos vs neg ................. 218
Figure 67. Présence de cellules CD169+ dans les espaces périvasculaires et péribronchiques.
................................................................................................................................................ 219
Figure 68. Les macrophages CD169+ expriment le CD68. .................................................... 220
Figure 69. Infiltrats périvasculaires de macrophages CD169+ détectés uniquement ches les
souris BALB/c microfilarémiques. ........................................................................................ 221
Figure 70. Quantification des infiltrats périvasculaires. ........................................................ 222
Figure 71. Présence de cellules CD169+ dans la plèvre viscérale.. ........................................ 223
Figure 72. Structures lymphoïdes associées aux bronches et à la plèvre. .............................. 224
Figure 73. Epaississement pleural des souris microfilarémiques. .......................................... 225
Figure 74. Hypertrophie des cellules mésothéliales chez les souris infestées........................ 226
Figure 75. Hyperplasie du mésothélium chez les souris microfilarémiques. ......................... 227
Figure 76. Surexpression de la podoplanine par le mésothélium hypertrophique. ................ 228
Figure 77. Fibrose du mésothélium hyperplasique. ............................................................... 229

14
Figure 78. Nbre de cellules dans la cavité pleurale et le LBA de BALB/c Mfposvs Mf neg .... 230
Figure 79. Augmentation des granulocytes et des macrophages pleuraux chez les souris
microfilariennes ...................................................................................................................... 231
Figure 80. Augmentation des éosinophiles et des lymphocytes T CD4+ dans l'espace
bronchoalvéolaire chez les souris infestées. ........................................................................... 232
Figure 81. Rendement filarien et taille des parasites dans les souris IL-4R-/-/IL5-/-............... 233
Figure 82. Fertilité des femelles et production de microfilaires. ........................................... 234
Figure 83. Détection des microfilaires pulmonaires. ............................................................. 235
Figure 84. Localisation des microfilaires chez les souris IL-4R-/-/IL5-/-. ............................... 236
Figure 85. Chez les souris Mfpos les dommages tissulaires sont dépendantes de l’IL4R et de
l’IL5. ....................................................................................................................................... 237
Figure 86. Le recrutement des éosinophiles et des neutrophiles est dépendant de l’IL-4R et de
l’IL-5. ..................................................................................................................................... 238
Figure 87. Synthèse des résultats et hypothèses mécanistiques. ............................................ 246

15
Liste et titre des tableaux
Tableau 1. Caractéristiques des filarioses humaines. ............................................................... 29
Tableau 2. Similarités entre TPE et asthme. ............................................................................ 38
Tableau 3. Filaires cardio-pulmonaires et pleurales. ................................................................ 47
Tableau 4. Comparaison entre M. perstans et L. sigmodontis. ................................................ 58
Tableau 5. Recommandations pour la distribution des traitements à grande échelle, les
traitements individuels et le contrôle des symptômes. ............................................................. 62
Tableau 6. Réponses cellulaires lors des différentes phases du programme de développment
de Litomosoides sigmodontis chez les rongeurs ....................................................................... 72
Tableau 7. Phénotype des macrophages pulmonaires. ............................................................. 92
Tableau 8. Protocole de coloration à l'Hémalun-Eosine. ....................................................... 108
Tableau 9. Protocole de la coloration au Trichrome de Masson. ........................................... 109
Tableau 10. Protocole de la coloration à l'acide périodique-Schiff. ....................................... 110
Tableau 11. Protocole de marquage de la protéine S100A9 .................................................. 113
Tableau 12. Protocole de marquage de la cytokératine 7 ....................................................... 114
Tableau 13. Protocole de marquage de la podoplanine .......................................................... 115
Tableau 14. Origine des anticorps utilisés pour l'immunofluorescence. ................................ 116
Tableau 15. Marquages utilisés en cytométrie en flux. .......................................................... 122
Tableau 16. Séquences des amorces utilisées pour les PCR quantitatives ............................. 129
Tableau 17. Cytokines testées. ............................................................................................... 156

16
Table des Annexes
Annexe 1. Liste des gènes pour l’étude des profils des transcrits pulmonaires à J4 et J70. .. 261 Annexe 2. Décollement des cellules mésothéliales de la plèvre viscérale. ............................ 263

17
Liste des abréviations
α-SMA : Alpha-Smooth Muscle Actin
γ-GT : γ-glutaryltranspeptidase
ADCC : Antibody-dependent cell-mediated cytotoxicity / Cytotoxicité cellulaire dépendante des
anticorps
ADN : Acide désoxyribonucléique
ADNc : ADN complémentaire
ALB : Albendazole
AM : Alveolar macrophage / Macrophage alvéolaire
ANOVA : Analyse Of Variance
APC : Allophycocyanine
APOC : African Programme for Onchocerciasis Control / Programme Africain pour le contrôle de
l'Onchocercose
Arg-1 : Arginine 1
ARN : Acide ribonucléique
ATP : Adénosine triphosphate
A.WOL : Consortium Anti-Wolbachia
BALT : Bronchus-associated lymphoid tissue / Tissus Lymphoïdes Associés aux Bronches
Bm : Brugia malayi
BSA :Bovine Serum Albumin / Albumine de sérum bovin
CCLX : C-C chemokine motif ligand X (CCL11 (Eotaxin-1), CCL5 (RANTES) , CCL3, CCL17 )
CCRX : C-C chemokine receptor type X (CCR2)
CD : Cluster of differentiation
CMHII : Complex majeur d’histocompatibilité de type II
ConA : Concanavaline A
COX-2 : Cyclo-oxygénase 2
CPA : Cellules présentatrices d'antigène
CT : Cycle Threshold / Cycle seuil
CTLA-4 : Cytotoxic T-Lymphocyte Antigen 4
CXCLX : C-X-C chemokine motif ligand X (CXCL1 et CXCL2)
CXCRX : C-X-C chemokine receptor type X (CXCR4)
CX3CR1 : CX3C chemokine receptor 1
DAMP : Damage associated molecular pattern / Motifs moléculaires associés aux dommages
cellulaires
DC : Dendritic cell / Cellule dendritique
DEC : Déthylcarbamazine
DNDi : Drugs for Neglected Diseases initiative
DOX : Doxycycline
dsDNA : Double stranded DNA / ADN double brin
ECF : Extracellular fluid / Fluide extracellulaire
ECM : Extra-cellular matrice / Matrice extracellulaire
ECP : Eosinophil cationic protein / protéine cationique des éosinophiles
ELISA : Enzyme Linked Immunosorbent Assay
EMT : Transition Epithélo-Mésenchymateuse
EPO : Peroxydase des Eosinophiles
FACS : Fluorescence-activated cell sorting

18
FALCs : Fat-associated lymphoid cluster / structures lymphoïdes associés aux graisses
Fc : Fragment crystallizable region
FC : Foldchange
FITC : Fluorescein isothiocyanate
FLBZ : Flubendazole
FSP-1 (S100A4): Fibroblast-specific protein 1
G: Granulome
GITR : Glucocorticoid induced TNFR-related protein
GPELF : Global Programme to Eliminate Lymphatic Filariasis / Programme mondial pour
l’élimination de la filariose lymphatique
HE : Hemalun-Eosine
HMDS : Héxaméthyldisylasane
HRP : Horse Raddish Peroxydase / Peroxydase de raifort
IFN-γ : Interferon-γ
Ig : Immunoglobuline (IgG, IgE IgG1, IgG4 IgM)
ILC : Cellules lymphoïdes innées (ILC2)
IL-X: Interleukine X (I IL-1β ; L-4, IL-5, IL-6, IL-8, IL-9, IL-10, IL-13, IL-17A, IL-25, IL-33 IL-1β)
IL-1R1 : Interleukin 1 receptor, type I / Récepteur à IL-1 de type 1
IL-1Ra : Interleukin 1 receptor antagonist/ Antagoniste du récepteur à l'IL-1
IL-4Rα : Chaine alpha du récepteur à l'IL-4
IM : Interstitial macrophage / Macrophage interstitiel
iNOS : Oxyde nitrique synthase inductible
IPA : Ingenuity Pathway Analysis
IVM : Ivermectine
KO : Knock-Out
LBA : liquide de lavage bronchoalvéolaire
LPS : Lipopolysaccharide
LX : Stade larvaire X
MBP : Major Basic Protein / Protéine Basique Majeure
MCP-1 (CCL2) : Monocyte chemoattractant protein 1/ Protéine chimioattractive monocytaire 1
MDA : Mass Drug Administration / Administration médicale de masse
MDP : Muramyldipeptide
MEB : Microscopie électronique à balayage
Mf s: Microfilaires
Mfpos : Souris microfilarémique
Mfneg : Souris amicrofilarémique
MMP : Matrix metalloproteinase
MPO : Myelopéroxydase
MRP8 (S100A8) : Myeloid-related protein 8
MRP14 (S100A9) : Myeloid-related protein 14
NaCl : Chlorure de sodium
nd : Non décrit
NGS : Next-Generation Sequencing
NOD2 : Nucleotide-binding oligomerization domain-containing protein 2
OCP : Onchocerciasis Control Programme / Programme de lutte contre l'onchocercose
Pam3Cys : 3HCl, (S)-[2,3-Bis(palmitoyloxy)-(2-RS)-propyl]-N-palmitoyl-(R)-Cys-(S)-Ser-(S)-Lys4-
OH
OMS : Organisation mondiale de la santé

19
PAS : Acide périodique-Schiff
PBS : Phosphate-buffered saline / tampon phosphate salin
PCLS : Precision Cut Lung Slices
PD-1 : Programmed Death 1
PDPN : Podoplanine
PE : R-Phycoérythrine
PE-Cy5 : R-Phycoérythrine-Cyanine 5
PFA : Paraformaldéhyde
p.i. : post inoculation
PleCs : Cellules de l’exsudat pleural
PMA : Phorbol Myristate Acetate
PVS : PeriVascular Space / espace périvasculaire
RPMI : Roswell Park Mémorial Institute 1640
Siglec : Sialic acid-binding immunoglobulin-type lectin
SP-A : Surfactant protein A
SP-D : Surfactant protein D
ST2 : Récepteur à l'IL-33
SVF : Sérum de veau fœtal
Teff : Lymphocytes T effecteurs
TGF-β : Tumor Growth Factor β
Th1 : T helper cells type 1
Th17 : T helper cells type 17
Th2 : T helper cells type 2
TLR : récepteur Toll-Like (TLR-2, TLR-6, TLR-1)
TMB : 3,3',5,5'-Tetramethylbenzidine
TNF-α : Tumor necrosis factor α
TPE : Poumon éosinophile tropical
Treg : Lymphocytes T régulateurs
TSLP : Thymic stromal lymphopoietin
TX : Triton
VEGF : Vascular endothelial growth factor
WoLP : Wolbachia Lipopeptide
WSP : Protéine de surface de Wolbachia

20
Résumé
Les filaires sont des nématodes parasites transmis à des vertébrés par des arthropodes
hématophages. Les espèces filariennes qui s'installent dans les cavités cœlomiques, les
vaisseaux lymphatiques ou des tissus conjonctifs ont leur stades infestants (ou L3) qui migrent
via le système lymphatique après leur inoculation dans la peau. En utilisant le modèle murin
avec la filaire Litomosoides sigmodontis dont les adultes résident dans la cavité pleurale, deux
phases d'interaction des filaires avec les poumons des souris BALB/c sont décrites 1) lors de
la migration des L3 de la peau à la cavité pleurale ; 2) pendant la phase patente de l’infection
quand les adultes pondent des microfilaires dans la cavité pleurale.
Au cours de la phase précoce les L3 rejoignent le système sanguin pulmonaire puis
traversent les poumons pour entrer dans la cavité pleurale. Ce passage se traduit par des
processus inflammatoires transitoires : tout d'abord des hémorragies consécutives à la rupture
des capillaires pulmonaires, accompagnées d'une augmentation du nombre de neutrophiles
pulmonaires et de la libération transitoire d'IL-1β et des alarmines IL-33 et S100A9 dans la
cavité pleurale. Le S100A9 semble faciliter la survie des filaires, probablement en facilitant la
migration rapide des L3 lors du démarrage de la phase contre-inflammatoire initiée dans le
poumon. En témoigne le recrutement de macrophages et d'éosinophiles, la production d'IL-4,
de CCL2 et d'IL9, ainsi que l’atténuation/extinction de l’expression de molécules pro-
inflammatoires. La formation de granulomes est également observée dans le tissu pulmonaire.
Lors de la phase patente de l'infestation, la descendance transmissible
(Microfilaires/Mf) est générée uniquement chez 60% des souris BALB/c -souris Mf pos -. Une
telle observation a permis de comparer les cavités pleurales et les poumons de souris Mfpos et
Mfneg. Chez les souris Mfpos des processus inflammatoires prolongés au niveau de la plèvre -
dont témoigne une fibrose de la plèvre viscérale- sont couplés au niveau du poumon et des
bronches respectivement à une accumulation périvasculaire de macrophages et une
inflammation bronchoalvéolaire (en témoigne la productionon de mucus par les cellules
caliciformes et une éosinophilie). Le contrôle de la charge parasitaire (adultes et
microfilaires), mais aussi la mise en place de la pathologie sont dépendantes de l'IL-5 et des
axes de signalisation dépendants de l'IL-4R.

21
Abstract
Filariae are parasitic nematodes transmited to vertebrates by haematophagous
arthropods. The filarial species that settle in the coelomic cavities, the lymphatic vessels or
the connective tissues have their infectious stages (or L3) which migrate via the lymphatic
system after their inoculation into the skin. Using the murine model with the
filaria Litomosoides sigmodontis, whose adults reside in the pleural cavity, two phases
of interaction between filariae and the lung of BALB/c mice are described 1) during the L3
migration from the skin to the pleural cavity; 2) during the patent phase of infection, when
adults realease microfilariae in the pleural cavity.
During the 1st phase L3 join the pulmonary blood system and then cross through the
lungs to enter the pleural cavity. This passage induces a transient acute pathology: first
haemorrhages following the rupture of the pulmonary capillaries, together with an increase in
the number of pulmonary neutrophils and the transient release of IL-1β and the alarmins IL-
33 and S100A9 in the pleural cavity. S100A9 appears to facilitate the survival of the filariae
either by an anti-inflammatory effect or by facilitating the migration of L3. Neutrophils can
release NETs in response to L3. Within days following the infection, a regulatory response
takes place in the lungs, with recruitment of macrophages and eosinophils, production of IL-4,
CCL2 and IL-9, and downregulation of inflammatory molecules. The formation of
granulomas is also observed in pulmonary tissue. The passage of L3 also induces an
inflammation of pulmonary blood vessels, in C57BL/6 mice only.
During the patent phase of the infection, 40% of the mice do not develop blood
microfilaraemia. Comparison of responses of microfilaremic and amicrofilaremic mice shows
an exacerbation of pleural inflammation induced by microfilariae. In addition, microfilaremic
mice develop microfilaria-dependent pulmonary pathology consisting on fibrosis of the
visceral pleura, perivascular accumulation of macrophages and bronchoalveolar inflammation
(mucus production and eosinophilia). The control of the filariae (adults and microfilariae), but
also the establishment of the pathology are dependent on IL-5 and IL-4R.

22

23
INTRODUCTION

24
Figure 1. Diversité des filaires.
Chaque portion du cercle correspond à une sous-famille. Le premier chiffre représente le
nombre d'espèces de la sous-famille et le deuxième chiffre entre parenthèses indique le
nombre de genres. D'après [1].
Figure 2. Phylogénie des filaires basée sur une analyse multigénique.
Soixante spécimens de filaires (représentant 48 espèces) ont été analysés. Filaria latala et
Protospirura muricola ont été utilisées comme groupes externes. Les séquences de coxI, rbp1,
hsp70, myoHC et des ADN ribosomiques 12S, 18S et 28S ont été analysées en utilisant
l'inférence bayésienne. Les résultats supportent l'existence de cinq clades appelés ONC1 à
ONC5. Les sous-familles sont indiquées par couleur : bleu pour les Onchocercinae, vert foncé
pour les Dirofilariinae, pourpre pour les Splendidofilariinae, vert pâle pour les Setariinae,
jaune pour les Waltonellinae, orange pour les Icosiellinae et rouge pour les Oswaldofilariinae.
Issu de [2].

25
1. Filaires
1.1 Diversité des filaires
La famille des Onchocercidae regroupe des nématodes parasites plus communément
appelés filaires. Ces parasites présentent un large spectre d'hôtes : ils peuvent infester aussi
bien des squamates, des crocodiliens, des amphibiens, des oiseaux et des mammifères -y
compris l'homme [3,4]. Environ 600 espèces de filaires sont actuellement décrites [1,4] et Figure
1). L'adaptation des filaires au mode de vie parasite a conduit à une régression de la
morphologie, rendant difficile l'étude phylogénétique. Les filaires sont actuellement réparties
en 8 sous-familles incluant 87 genres [1,4,5]. Une phylogénie récente basée sur une analyse
multigénique [2] suggère l'existence de 5 clades monophylétiques et remet en cause la
délimitation de certaines de ces sous-familles (Figure 2).
1.2 Biologie
Comme tous les nématodes, les filaires adultes sont des vers ronds, longs (jusqu’à
plusieurs dizaines de centimètres pour certaines onchocerques), à corps non segmenté,
composés d'un hypoderme et d'une couche de muscles longitudinaux organisés en quadrants
recouverts par une cuticule [6]. Leur système digestif est composé d'une capsule buccale
antérieure, d'un œsophage divisé en une partie musculaire et une glandulaire, d'un intestin et
d'un rectum postérieur. Leur système nerveux est composé d'un anneau nerveux péri-
œsophagien associé à des ganglions et des cordons nerveux [6,7]. Les nerfs rejoignent des
récepteurs tactiles appelés papilles (céphaliques ou caudales) ou des chémorécepteurs
nommés amphides et phasmides. De plus, les filaires ont un système d'excrétion et
d'osmorégulation. Un dimorphisme sexuel de taille est plus ou moins marqué en fonction de
l'espèce, avec des femelles plus longues que les mâles. Le système reproducteur des femelles
est composé de deux ovaires reliés par un oviducte à un utérus bifurqué terminé en vagin
conduisant à une vulve antérieure. Le système reproducteur des mâles est constitué d'un
testicule relié à une vésicule séminale conduisant à un canal déférent s'ouvrant sur un orifice
génital fusionné avec l'anus appelé cloaque. L'appareil copulatoire des mâles est composé de
deux spicules [7].
26
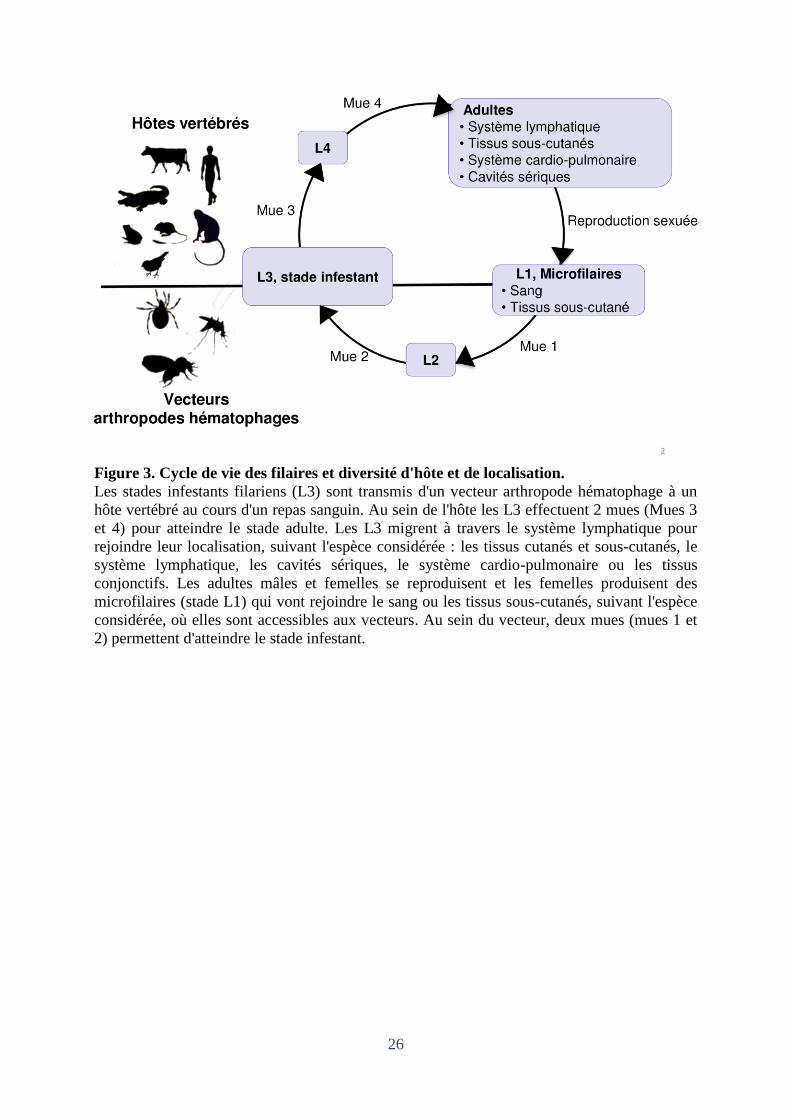
Figure 3. Cycle de vie des filaires et diversité d'hôte et de localisation.
Les stades infestants filariens (L3) sont transmis d'un vecteur arthropode hématophage à un
hôte vertébré au cours d'un repas sanguin. Au sein de l'hôte les L3 effectuent 2 mues (Mues 3
et 4) pour atteindre le stade adulte. Les L3 migrent à travers le système lymphatique pour
rejoindre leur localisation, suivant l'espèce considérée : les tissus cutanés et sous-cutanés, le
système lymphatique, les cavités sériques, le système cardio-pulmonaire ou les tissus
conjonctifs. Les adultes mâles et femelles se reproduisent et les femelles produisent des
microfilaires (stade L1) qui vont rejoindre le sang ou les tissus sous-cutanés, suivant l'espèce
considérée, où elles sont accessibles aux vecteurs. Au sein du vecteur, deux mues (mues 1 et
2) permettent d'atteindre le stade infestant.

27
1.3 Programme de développement
Les filaires ont un cycle de vie dixène caractérisé par 5 stades incluant 4 stades
larvaires (L1, L2, L3, L4) suivis d'un stade adulte (mâles et femelles) (Figure 3). Le premier
stade larvaire (L1) est aussi appelé microfilaire (Mf). Dans les filaires femelles adultes, les
œufs présentent une fine membrane appelée gaine. Chez certaines filaires, les Mfs éclosent in
utero (Mfs nues) alors que d'autres conservent la membrane de l'œuf (Mfs gainées) [4]. Les
Mfs sont expulsées par les femelles et migrent, suivant les espèces filariennes, dans le sang ou
la peau de leur hôte ; elles peuvent alors passer dans un arthropode hématophage au moment
du repas sanguin de celui-ci [3,4]. Les vecteurs des filaires peuvent être des poux, des puces,
des diptères (moustiques, simulies, ceratopogonidés, tabanidés) mais aussi des tiques et autres
acariens [3,4] selon l’espèce filarienne considérée. Les microfilaires se retrouvent dans
l'estomac du vecteur après le repas sanguin et doivent traverser l'épithélium de ce dernier pour
poursuivre leur développement [8].
Le développement larvaire se poursuit dans le vecteur avec deux mues successives
amenant au troisième stade, le stade infestant (L3). Ce développement s'effectue, selon
l'espèce filarienne considérée, dans différents sites préférentiels : les tissus graisseux, les
muscles, l'hémocèle ou encore les tubes de Malpighi [4]. Les larves qui atteignent
l'hémolymphe vont ensuite migrer vers la tête de l'arthropode et les pièces buccales, les
glandes salivaires ou l'hémocèle [4,9]. La croissance de la filaire se poursuit lorsque
l'arthropode infecté transfère les L3 dans un nouvel hôte vertébré au cours d'un repas sanguin.
La localisation des filaires adultes est variable selon l’espèce considérée : certaines
espèces s'installent dans les tissus cutanés et sous-cutanés, mais d’autres résident dans le
système lymphatique, les cavités sériques, le système cardio-pulmonaire ou les tissus
conjonctifs [4]. Malgré cette diversité dans la localisation finale, un comportement identique
semble avoir été sélectionné chez les filaires : les L3 survivent en entrant dans un vaisseau
lymphatique de la peau et ils migrent à travers l'hôte en utilisant ce système vasculaire [3,10,11].
Au sein de l'hôte vertébré, le développement des L3 continue avec deux mues
supplémentaires permettant d'atteindre les stades adultes mâles et femelles [4].

28
1.4 Wolbachia
Wolbachia est une α-proteobacterie intracellulaire Gram-négative de l'ordre des
Rickettsiales. Elle est présente chez les arthropodes mais on la trouve aussi chez les filaires.
Sur les 85 espèces de filaires étudiées à ce jour, 52.9% sont infestées par une souche de
Wolbachia [12]. En l'absence de description formelle d'espèces, les différentes souches de
Wolbachia ont été classées en supergroupes représentant les subdivisions phylogénétiques
majeures [13]. Plus de 10 supergroupes de Wolbachia infectent exclusivement les arthropodes,
3 supergroupes infectent spécifiquement les filaires (groupes C, D et J) et un groupe infecte
les deux (groupe F) [12].
Au sein des filaires, Wolbachia a été identifiée principalement dans l'appareil
reproducteur des femelles et dans les chordes latérales de l'hypoderme. [12,14,15]. La densité en
Wolbachia varie en fonction des tissus colonisés et du stade de développement de la filaire.
Du stade microfilaire au stade infestant, la quantité de Wolbachia est faible et est restreinte à
quelques cellules des chordes latérales [15]. La quantité de Wolbachia augmente dans les
chordes latérales des larves de stade 4 et des jeunes adultes. Dans les femelles fécondées,
Wolbachia migre des chordes latérales vers les ovaires et les embryons [15,16]. La présence de
la bactérie dans les gonades des filaires femelles est liée à sa transmission verticale, de la
femelle à sa descendance.
Seules les espèces de Wolbachia appartenant aux supergroupes C et J semblent
présenter une coévolution stricte avec leur hôte [12]. Les clades filariens hébergeant les
supergroupes D et F présentent de nombreux cas de pertes secondaires et de transferts
horizontaux de la bactérie [12]. Lorsque la bactérie est présente, la symbiose entre Wolbachia
et les Onchocercidae est considérée comme mutualiste, car les deux partenaires bénéficient de
l'association [17]. Wolbachia est nécessaire à la survie et la fertilité des filaires [18]. En effet,
des traitements antibiotiques sur diverses espèces de filaires ont démontré que l'élimination de
Wolbachia est associée à une apoptose des cellules germinales adultes et des cellules
somatiques des embryons [19] ainsi qu'à une perte de la polarité antéropostérieure des
embryons [20]
.
29
2. Filarioses humaines
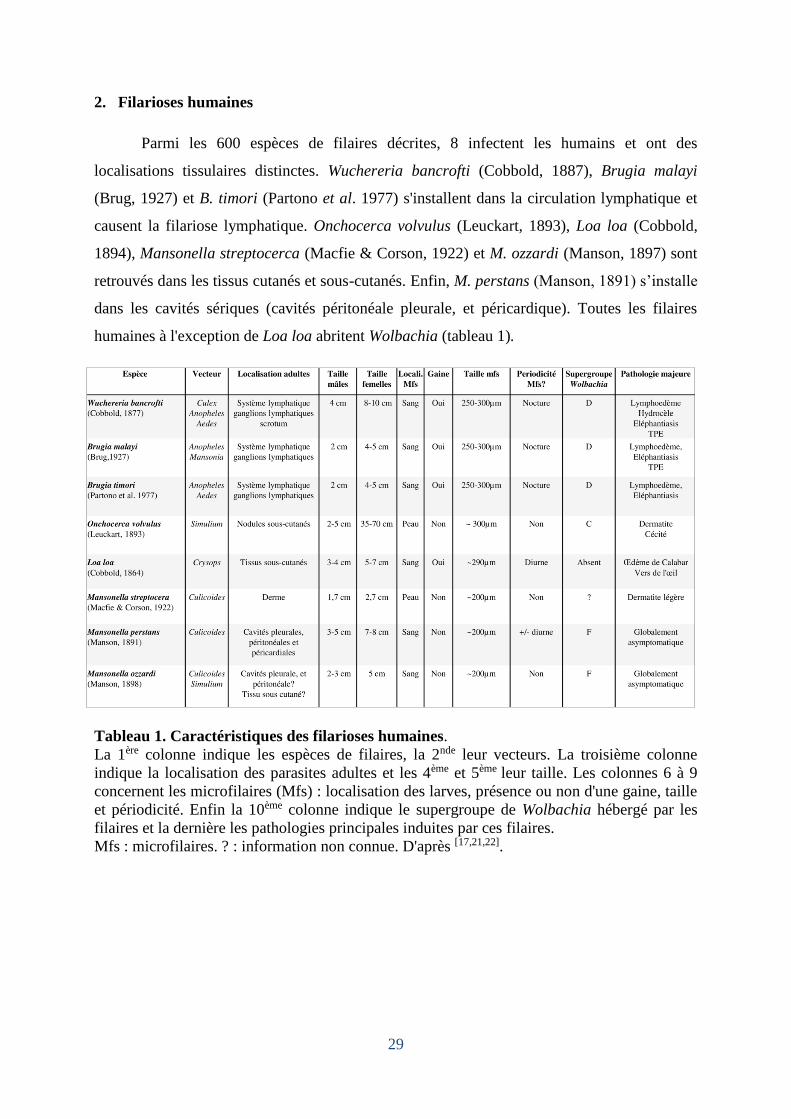
Parmi les 600 espèces de filaires décrites, 8 infectent les humains et ont des
localisations tissulaires distinctes. Wuchereria bancrofti (Cobbold, 1887), Brugia malayi
(Brug, 1927) et B. timori (Partono et al. 1977) s'installent dans la circulation lymphatique et
causent la filariose lymphatique. Onchocerca volvulus (Leuckart, 1893), Loa loa (Cobbold,
1894), Mansonella streptocerca (Macfie & Corson, 1922) et M. ozzardi (Manson, 1897) sont
retrouvés dans les tissus cutanés et sous-cutanés. Enfin, M. perstans (Manson, 1891) s’installe
dans les cavités sériques (cavités péritonéale pleurale, et péricardique). Toutes les filaires
humaines à l'exception de Loa loa abritent Wolbachia (tableau 1).
Tableau 1. Caractéristiques des filarioses humaines.
La 1ère colonne indique les espèces de filaires, la 2nde leur vecteurs. La troisième colonne
indique la localisation des parasites adultes et les 4ème et 5ème leur taille. Les colonnes 6 à 9
concernent les microfilaires (Mfs) : localisation des larves, présence ou non d'une gaine, taille
et périodicité. Enfin la 10ème colonne indique le supergroupe de Wolbachia hébergé par les
filaires et la dernière les pathologies principales induites par ces filaires.
Mfs : microfilaires. ? : information non connue. D'après [17,21,22].
30
A
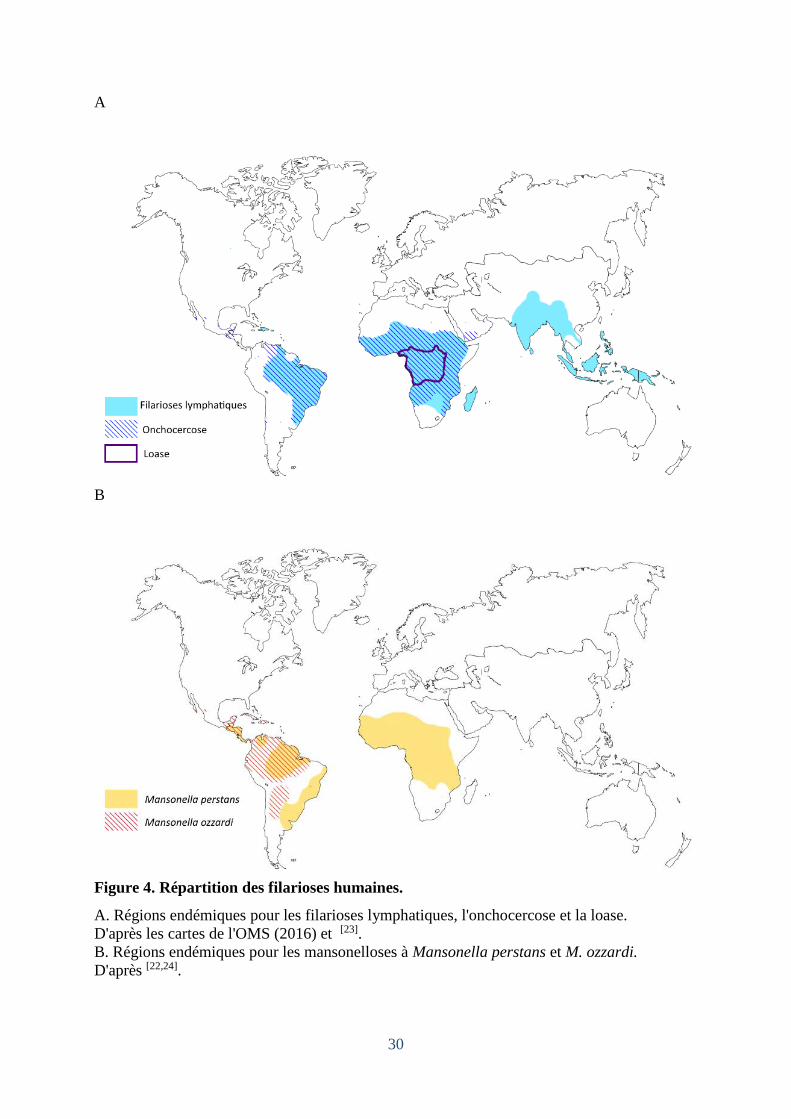
B
Figure 4. Répartition des filarioses humaines.
A. Régions endémiques pour les filarioses lymphatiques, l'onchocercose et la loase.
D'après les cartes de l'OMS (2016) et [23].
B. Régions endémiques pour les mansonelloses à Mansonella perstans et M. ozzardi.
D'après [22,24].

31
2.1 Epidémiologie et coendémicité
Si des filaires animales sont présentes sur tout le globe, les filarioses humaines sont
restreintes aux régions tropicales et subtropicales. Dans certaines régions d'Afrique, la
filariose lymphatique, l'onchocercose, la mansonellose à M. perstans et/ou la loase sont co-
endémique et certains patients peuvent présenter des multi-infections. De même, en Amérique
latine, on peut retrouver des zones co-endémiques pour M. perstans et M. ozzardi, la filariose
lymphatique et/ou l'onchocercose (Figure 4). Ces coïnfections sont importantes à connaître
car elles peuvent entrainer des erreurs d'identification des parasites, soit parce qu’une espèce
est minoritaire et non identifiée, soit par réaction croisée des kits de détection. Le problème
majeur provient des coïnfections avec L. loa qui peut entraîner de graves complications lors
des traitements.
En 2014, la filariose lymphatique était endémique dans 73 pays tropicaux et
subtropicaux d'Asie, d'Afrique, d'Amérique centrale et du Sud et du Pacifique [25,26]. Environ 1
milliard de personnes vivent en zone d'endémie et les 68 millions de cas avérés de filariose
lymphatique comprennent 36 millions de personnes présentant des symptômes tels qu'un
hydrocèle ou un lymphoedème secondaire [26].
Onchocerca volvulus est endémique dans 31 pays d'Afrique et infecte environ 15,5
millions de personnes [27,28]. 99% des personnes infestées vivent en Afrique sub-saharienne,
mais il existe encore quelques foyers en Amérique Latine (à la frontière Brésil-Venezuela) et
au Yémen [29]. Il a été estimé que 187 millions de personnes vivent dans des zones où le
parasite peut être transmis dont 500 000 en Amérique Latine [27,30]. 26 pays en Afrique
présentent une coendémicité pour la filariose lymphatique et l'onchocercose [26].
Les infections par Loa loa sont restreintes aux forêts tropicales d'Afrique de l'Ouest et
du Centre ou elles représentent au moins 10 millions de cas [23,31-33]. Dans ces zones, de
nombreux cas de co-infection avec O. volvulus, W. bancrofti et/ou M. perstans sont décrits
[22,31,34,35].

32
Les mansonelloses sont relativement asymptomatiques et leur prévalence est, de ce
fait, méconnue [22,24]. Cependant la forte incidence des coinfections avec M. perstans ou M.
ozzardi, et la présence d’une Wolbachia du supergroupe F dans ces mansonelles [12,36,37] sont à
l’origine d’un regain d’intérêt pour ces filaires [22,24,38,39].
Les infections par Mansonella ozzardi sont localisées exclusivement en Amérique
Latine, du Mexique au Nord de l'Argentine et dans les Caraïbes. Suivant les régions, la
prévalence varie entre 0 et 50% de la population qui est infestée rendant difficile l'estimation
du nombre total de personnes infestées [24]. Au Brésil, M. ozzardi est retrouvée dans des
régions d'Amazonie où est présente O. volvulus et induit régulièrement des erreurs
d'identifications bien que les microfilaires soient morphologiquement très différentes ([4] et
Tableau 1).
M. perstans est largement distribuée en Afrique et présente aussi des cas en Amérique
latine. Au total, 33 pays d'Afrique subsaharienne semblent être endémiques pour la
transmission avec un total d'environ 580 millions d'habitants à risque (Banque mondiale,
2008). Simonsen [22] estime que 114 millions de personnes pourraient être infestées par M.
perstans en Afrique ce qui en ferait un des parasites les plus prévalent en Afrique. Au
Cameroun, la prévalence de M. perstans est hétérogène : les zones forestières humides
présentent une prévalence et une intensité de l'infection élevées, la prévalence pouvant
atteindre 70% de la population ; en revanche, la prévalence est faible dans les zones de savane
[40]. Au Ghana, plus de 30% de la population pourrait être infestée [41].

33
2.2 Programme de développement et biologie
2.2.1 Les filaires lymphatiques : Wuchereria bancrofti, Brugia malayi et
B. timori
La filaire W. bancrofti est responsable de 90% des cas connus de filariose lymphatique
et B. malayi est à l'origine de la plupart des cas restants (OMS et [18]). Les filaires sont
transmises par des moustiques des genres Culex, Anopheles, Mansonia et Aedes lors d'un
repas sanguin [42].
Après la piqure, les larves s'installent, muent et se reproduisent dans le système
lymphatique. Les adultes de Wuchereria bancrofti se retrouvent généralement dans les
vaisseaux lymphatiques ou les sinus des ganglions lymphatiques des membres inférieurs chez
les patients et aussi dans les vaisseaux lymphatiques du scrotum chez les hommes [4,43].
Brugia malayi s'installe préférentiellement dans le système lymphatique de l'aine et des
aisselles [44]. Les adultes peuvent survivre de 5 à 8 ans dans leur hôte [18].
Les microfilaires sont pondues dans les vaisseaux lymphatiques et migrent dans le
sang. Chez des singes Semnopithèques (Presbytis melalophos et Trachypithecus cristatus)
infestés avec des larves infestantes de W. bancrofti, la phase pré-patente dure 200-280 jours
[45-47]. Les microfilaires de W. bancrofti et B. malayi ont une grande gaine dépassant des deux
extrémités de la microfilaire [4] et mesurent entre 250 et 300µm de long. La partie postérieure
des Mfs de B. malayi présente un noyau qui est absent chez W. bancrofti [4]. Les Mfs
présentent également une périodicité circadienne qui a été associée au mode de vie du vecteur
[48]. Les microfilaires se réfugient dans les capillaires pulmonaires pendant la journée et sont
retrouvées dans le sang périphérique la nuit où elles sont accessibles au vecteur [49]. La
périodicité serait contrôlée par les différences de concentrations en oxygène dans le sang entre
les périodes de veille et de sommeil jouant sur la dilatation des capillaires sanguins [4,50]. Une
étude in vitro a montré que les Mfs de B. malayi peuvent adhérer aux cellules endothéliales
[50]. L'adhérence implique la gaine de la microfilaire et nécessite un environnement riche en
oxygène ainsi qu'un flux élevé, comme ceux trouvés dans l'hôte pendant la journée [50].

34
2.2.2 Onchocerca volvulus
Les vecteurs de O. volvulus sont des mouches hématophages faisant partie du
complexe Simulium damnosum. Ce complexe est composé d'espèces génétiquement
différentes, mais qui ont une morphologie identique [51]. 63 espèces ont été définies sur la base
de leurs variations chromosomiques [30]. Le vecteur nécessite des cours d'eau pour se
reproduire et par conséquent l'infection se produit à proximité des rivières.
Le vecteur effectue une morsure superficielle en dilacérant la peau et les capillaires
sanguins avec ses maxillaires et ses mandibules. Les L3 pénètrent dans la peau par la plaie
due à cette morsure. Les filaires femelles adultes restent dans le tissu sous-cutané et des
nodules cellulaires se forment autour d'elles, les onchocercomes. Les femelles adultes peuvent
vivre plus de 10 ans au sein de ces nodules onchocerquiens [52]. Les femelles s'accouplent et
sont fécondées par les mâles. Les microfilaires sont gainées et mesurent environ 300µm. Elles
sont pondues dans le nodule où elles sont retrouvées en grandes quantités [53]. Des vaisseaux
sanguins et lymphatiques irriguent le nodule [53]. Aucune microfilaire n'est retrouvée dans les
vaisseaux sanguins et quelques microfilaires sont retrouvées dans les vaisseaux lymphatiques
[53], suggérant qu'elles pourraient les utiliser pour rejoindre le derme où elles sont accessibles
aux simulies [53-55]. Dans le derme, les microfilaires ne sont pas observées dans les vaisseaux
lymphatiques même si ces derniers sont anormalement dilatés [53]. Les microfilaires peuvent
se retrouver dans les tissus sous-conjonctivaux de l'œil [56,57].
2.2.3 Loa loa
Les humains sont le seul hôte connu de Loa loa. Les mouches du genre Chrysops
(majoritairement C. silace et C. dimidiate) sont les vecteurs de la loase [4]. Ces mouches
vivent dans les forêts d'Afrique équatoriale et effectuent leur repas sanguin pendant la
journée.
Après infection, les filaires pénètrent dans les tissus sous cutanés et maturent
rapidement. Chez la gerbille, le stade adulte est atteint en 25 jours [58]. Ils migrent à travers les
tissus sous-cutanés pendant toute leur durée de vie (jusqu'à 17 ans) [59]. Les infections
expérimentales chez les primates non humains ont montré que la période pré-patente est
d'environ 140 jours [60]. Les microfilaires sont retrouvées dans la circulation sanguine. Les
microfilaires mesurent environ 290µm par 7.5µm et présentent une gaine et ont 3 noyaux

35
caractéristiques dans la région terminale [4]. De plus, comme W. bancrofti, les Mfs de L. loa
affichent une périodicité, mais celle-ci est inversée. Les microfilaires sont présentes dans le
sang périphérique pendant la journée et résident probablement dans le tissu pulmonaire
pendant la nuit bien que cela n'ait pas été formellement démontré [61,62]. Ces caractéristiques
aident à différencier les microfilaires sanguines de L. loa de celles de W. bancrofti et de
Mansonella perstans dont la distribution géographique chevauche celle de L. loa (Tableau 1).
2.2.4 Les mansonelles
Les trois espèces de mansonelles infectant les humains, Mansonella perstans
(synonymes : Dipetalonema perstans, Tetrapetalonemaperstans, Acanthocheilonema
perstans), M. ozzardi et M. streptocerca (synonymes : Dipetalonema streptocerca,
Tetrapetalonema streptocerca), varient par leur distribution, leur spécificité vectorielle et leur
site. Toutefois bien que les humains soient l'hôte principal, des infections ont été décrites chez
d'autres primates et M. ozzardi a aussi été trouvé chez des oiseaux et des amphibiens [22,63,64].
Le cycle de vie des mansonelles est mal connu. M. streptocerca est spécifiquement
transmis par la mouche Culicoides grahamii [65] mais plusieurs espèces de Culicoides peuvent
transmettre les larves de M. perstans et M. ozzardi [66]. Les adultes de M. ozzardi ont
initialement été décrits dans la cavité abdominale et dans les mésentères [4] mais leur
localisation exacte est toujours incertaine car l'inoculation de larves de M. ozzardi dans des
primates induit un développement sous-cutané du parasite [67]. Concernant M. streptocerca,
les adultes résident dans le derme de la peau. Les adultes de M. perstans sont rarement
observés, mais ils ont été occasionnellement récupérés dans les cavités sériques (péritoine,
plèvre, péricarde) lors de chirurgies ou d'autopsies [22].
Les microfilaires des trois espèces sont dépourvues de gaine. Les Mfs mesurent
environ 190-200µm de long par 4-5µm de diamètre. Les Mfs vivantes ont de fortes capacités
de contraction et d'élongation ce qui fait que les tailles décrites varient beaucoup [22,68]. Les
Mfs de M. streptocerca sont cutanées alors que celles de M. ozzardi et M. perstans sont
sanguines [69].

36
2.3 Principales manifestations cliniques
La plupart des infections filariennes chez les vertébrés non humains sont considérées
comme étant généralement asymptomatiques. Les signes cliniques n'apparaissent pas avant
que les parasites n’aient atteint le stade adulte et ils dépendent des espèces ainsi que de la
localisation des adultes et des microfilaires [17].
Les pathologies associées à W. bancrofti, B. malayi, O. volvulus et L. loa sont décrites
ci-après :
2.3.1 Filarioses lymphatiques
Les symptômes des filarioses lymphatiques sont majoritairement dus à la présence des
parasites adultes dans les vaisseaux lymphatiques. Certains patients présentent une filariose
dite "occulte" avec des adultes mais pas de microfilaires sanguines. Cependant chez ces
patients, des microfilaires sont parfois retrouvées dans les tissus où elles induisent des
symptômes cliniques.
Symptômes associés aux adultes
Après leur installation dans le circuit lymphatique, les parasites adultes induisent une
dilatation des vaisseaux lymphatiques, une baisse de la contraction des muscles des
lymphatiques et une perte de la fonction des valves lymphatiques [43,70,71]. Au fur et à mesure
que la filariose progresse et que les parasites meurent, une insuffisance lymphatique
chronique se met en place, caractérisée par un ralentissement du flux lymphatique. Les
symptômes se produisent le long d'un continuum allant de la dilatation lymphatique
asymptomatique (environ 2/3 des patients) aux hydrocèles et aux lymphoedèmes secondaires
(les lymphoedèmes primaires provenant de facteurs congénitaux) pouvant progresser vers
l'éléphantiasis [43,72].
Les fonctions principales du système lymphatique sont l'élimination des molécules de
haut poids moléculaire des espaces interstitiels et le transport des antigènes et des cellules
immunitaires vers les ganglions lymphatiques. L'insuffisance lymphatique induit donc la
rétention de protéines et l'augmentation du risque d'infections [73]. Le lymphoedème
secondaire se forme suite à l'accumulation des protéines, qui augmente la pression oncotique,
entraînant la formation d'un œdème riche en protéines qui stimule les fibroblastes. Ceux-ci
sont à l'origine d'une fibrose cutanée hyperplasique avec un épaississement du derme et une

37
accumulation de lobules graisseux dans l'hypoderme. Les infections bactériennes et fongiques
sont une complication fréquente des lymphoedèmes filariens (Figure 5). Ceci conduit à
l'éléphantiasis, un lympœdème irréversible même après la mort des parasites adultes 1995 [74].
L'hydrocèle est l'autre forme courante des infections par W. bancrofti chez des patients
masculins. Elle consiste en l'accumulation de fluide entre les deux couches de la tunique
vaginale dans le scrotum. Dans ce cas, l'élargissement des vaisseaux lymphatiques et leur
obstruction ultérieure conduisent à la pathologie indépendamment des infections bactériennes
[73,75]. Ces symptômes sont associés à la présence de parasites adultes dans les vaisseaux
lymphatiques du scrotum [76,77].
Figure 5. Evolution du lymphoedème filarien.
Les parasites adultes entrainent une dilatation lymphatique conduisant à des lymphoedèmes.
De nombreux facteurs participent à la pathogénèse et aggravent la pathologie comme la mort
des parasites, l'inflammation chronique et les infections secondaires. Différents stades
permettent de caractériser le lymphoedème : Stade 2 : gonflement non réversible pendant la
nuit; Stade 3 : plis superficiels de la peau à la cheville. Stade 4 : altération de la texture de la
peau et formation de boutons. Stade 5: plis profonds de la peau en plus des altérations de la
phase 4; Stade 6 : lichénification généralisée en plus des altérations de la phase 5. Étape 7:
incapacité du patient à effectuer un travail quotidien. D'après Dreyer et al. 2000 [70].

38
Symptôme TPE Asthme
Obstruction bronchique/mucus + +
Eosinophilie sanguine + +
Eosinophilie bronchoalvéolaire + +
Infiltrats pulmonaires d'éosinophiles + +
Infiltrats pulmonaires de mastocytes ? +
IgE et IgG circulants + +
Fibrose du tissu pulmonaire + +
Tableau 2. Similarités entre TPE et asthme.
Comparaison des symptômes (1ère colonne) observés lors du TPE (2ème colonne) et de l'asthme
(3ème colonne). + : symptôme observé. ? : information non connue. D'après [78-80] .
Figure 6. Histopathologie du « poumon eosinophile tropical ».
(A) Microfilaire (flèche) dans le parenchyme pulmonaire avec infiltrats de macrophages ; (B)
Infiltrats péribronchiques d'éosinophiles ; B : Bronche ; C. Fibrose pulmonaire : zone claire
avec forte concentration de fibres. Images issues de [78].

39
Symptômes associés aux Mfs :
Le poumon éosinophile tropical (TPE) est le principal symptôme des filarioses
occultes. Il est dû à une hyperréactivité immunitaire qui se développe chez certains individus
infectés par W. bancrofti et B. malayi [77].
Les symptômes sont proches de ceux de l'asthme avec des toux récurrentes et des
difficultés respiratoires (voir tableau 2). Ces patients ne présentent en général pas de
microfilaires circulantes mais des microfilaires ont été observées dans des biopsies
pulmonaires de patients TPE (Figure 6A), suggérant que les Mfs seraient piégées dans la
microcirculation pulmonaire [78,81]. Très peu d'études ont été effectuées pour comprendre les
mécanismes du TPE. La dégradation des microfilaires dans les capillaires pulmonaires
libèrerait des constituants antigéniques qui déclencheraient la réponse immunitaire observée,
caractérisée par une éosinophilie très élevée et des forts niveaux d'IgE et d'IgG sanguins et
alvéolaires spécifiques aux filaires [79,80]. Les patients présentent de plus une forte
concentration d'éosinophiles dans les alvéoles pulmonaires [80]. Ces réponses sont associées à
un défaut de ventilation avec obstruction des voies aériennes [80]. Les études de biopsies
pulmonaires ont mis en évidence des réactions histopathologiques constituées d'infiltrats
d'éosinophiles et/ou de macrophages dans les espaces interstitiels, péribronchiaux et
périvasculaires des poumons associés parfois à une fibrose interstitielle marquée [78] (Figure
6).
Des modèles de souris sensibilisées par des injections répétées d'antigènes ou de
microfilaires mortes de B. malayi développent les symptômes du TPE [82,83]. Chez ces souris,
le recrutement des éosinophiles est corrélé au développement de l'hypersensibilité aérienne et
de grandes quantités de MBP (Major Basic Protein, un constituant principal des granules
éosinophiles) sont retrouvées dans le liquide bronchoalvéolaire [83] et sur les cellules
épithéliales bronchiques [84]. De plus, l'expression de nombreux gènes anti-apoptotiques est
augmentée dans les éosinophiles entraînant une survie accrue de ces cellules [82]. Les souris
déficientes en IL-5 ne présentent pas d'infiltrats d’éosinophiles ni de symptômes asthmatiques
[83] indiquant un rôle prépondérant des éosinophiles et de leurs granules dans l'instauration de
la pathologie observée lors du TPE. Enfin, les macrophages alvéolaires prennent un
phénotype alternativement activé avec une hausse de l'expression de marqueurs spécifiques
tels que l'arginase 1 (Arg-1), Ym-1 et Relm-α [82] et pourraient participer à la pathologie. En

40
effet, les souris déficientes en IL-4, molécule importante dans polarisation des macrophages,
présentent des atteintes pulmonaires réduites [85].
D'autres études suggèrent une origine auto-immune de la maladie. Un antigène de B.
malayi, Bm23-25, a été identifié chez des patients atteints de TPE. Cette molécule est
majoritairement produite par les microfilaires et induit une production d’IgE après stimulation
de cellules sanguines de patients filariens TPE ou non [86]. Le sérum et le liquide
bronchoalvéolaire des patients TPE contiennent des IgE reconnaissant Bm23-25. Cet antigène
filarien est un homologue d'une sous-unité d'une enzyme humaine, la γ-glutaryl transpeptidase
(γ-GT) [87]. Les γ-GT de mammifères sont des enzymes transmembranaires présentes en
grande quantité sur la face apicale des cellules épithéliales, dont les cellules épithéliales
pulmonaires. Du fait de ce mimétisme moléculaire, les IgE antiᵧ-GT filarienne peuvent aussi
reconnaitre la γ-GT humaine présente sur les épithéliums pulmonaires et pourraient induire
une rupture de tolérance immunitaire [88]. Les niveaux d'IgE spécifiques pour le Bm23-25 sont
associés avec la pathologie pulmonaire chez les patients filariens ; les patients TPE présentent
des niveaux d'IgE spécifiques pour Bm23-25 environ 50 fois plus élevés que les autres
patients filariens [89]. Le mécanisme entraînant la pathologie n'a pas été étudié mais il est
probable que la sensibilisation des mastocytes par des IgE spécifiques pour la protéine
humaine entraîne leur activation et la dégranulation de protéines cytotoxiques. Des
mécanismes de cytotoxicité dépendante des anticorps (ADCC) pourraient aussi mettre en jeu
les éosinophiles et leurs granules cytotoxiques.

41
2.3.2 Onchocercose
Les manifestations cliniques de l'infection par O. volvulus sont liées à la réponse
immunitaire induite par la mort de microfilaires dans la peau et les yeux et très peu à la
présence des parasites adultes. Environ 30% des personnes infestées ne présentent pas de
signes cliniques d'infection.
Symptômes associés aux adultes
La genèse des onchocercomes semble induite par les filaires femelles puisqu'on ne
retrouve jamais de nodule contenant un mâle seul [90]. Ils se trouvent dans des tissus
intramusculaires sous-cutanés ou plus profonds entourés d'un noyau fibreux qui contient des
vaisseaux sanguins et des vaisseaux lymphatiques néoformés [53]. Les vaisseaux sanguins
permettraient de faciliter l’arrivée de cellules immunitaires mais aussi de fournir des
nutriments aux vers [91-93]. Les filaires adultes dans les nodules contribuent peu à la
pathogenèse en dehors du fait qu'ils peuvent être douloureux et gênants [94]. Les microfilaires
sont libérées par les filaires femelles dans le nodule d'où elles peuvent migrer sur de longues
distances dans le corps de leur hôte [95,96].
Symptômes associés aux Mfs
La pathologie est essentiellement dermatologique. Il a été estimé que chez une
personne fortement infestée, plus de 100 000 microfilaires peuvent mourir chaque jour dans la
peau [97,98]. La pathologie de l'onchocercose est principalement une conséquence de réponses
inflammatoires chroniques de type Th2 [99,100]. La réponse cutanée aux Mfs présente un large
éventail clinique suivant la résistance/susceptibilité de l'hôte. Les patients présentant une forte
microfiladermie développent généralement une faible inflammation de la peau et de
nombreux marqueurs anti-inflammatoires comme l'interleukine (IL)-10 et le TGF-β (Tumor
Growth Factor β) [101-103]. Les patients infectés chroniquement peuvent présenter une perte
d'élasticité de la peau, probablement due à des protéases issues du parasite ou de l'hôte [104],
[105], ainsi qu'une dépigmentation de la peau (peau de léopard) [100]. De rares cas de réponses
hyper-réactives existent et sont appelées sowda, qui dérive du mot aswad, signifiant "de
couleur noire" en arabe [94].
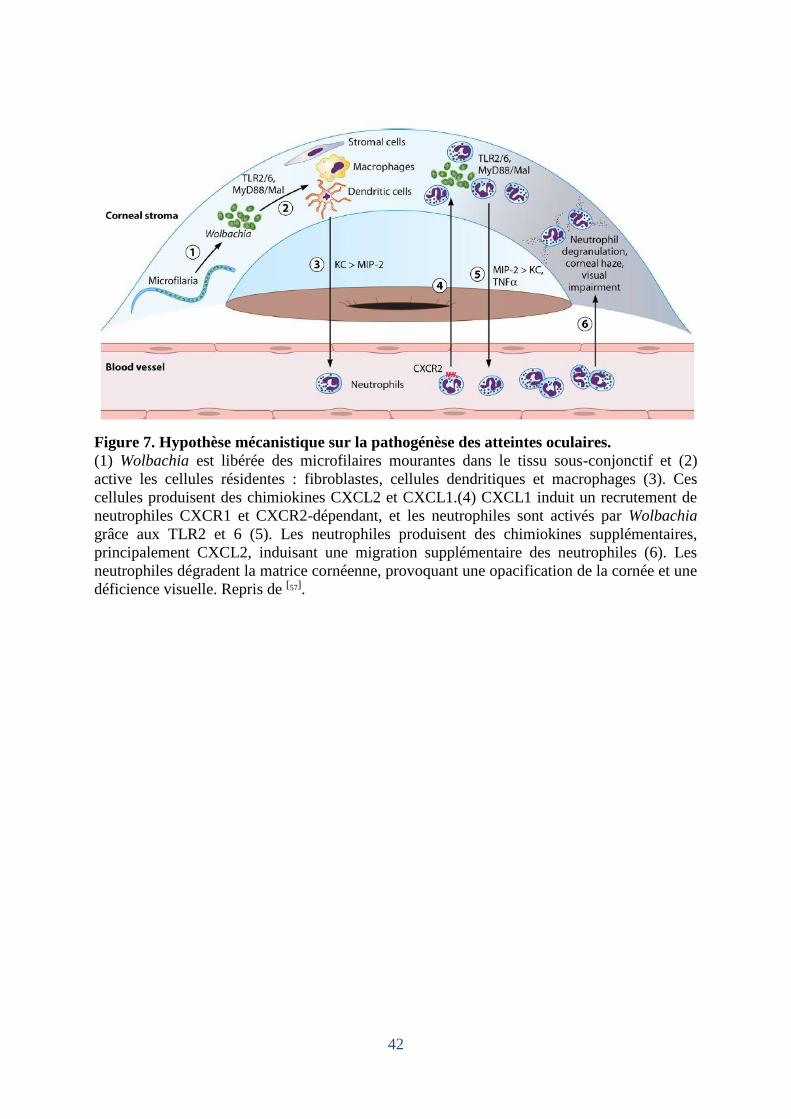
42
Figure 7. Hypothèse mécanistique sur la pathogénèse des atteintes oculaires.
(1) Wolbachia est libérée des microfilaires mourantes dans le tissu sous-conjonctif et (2)
active les cellules résidentes : fibroblastes, cellules dendritiques et macrophages (3). Ces
cellules produisent des chimiokines CXCL2 et CXCL1.(4) CXCL1 induit un recrutement de
neutrophiles CXCR1 et CXCR2-dépendant, et les neutrophiles sont activés par Wolbachia
grâce aux TLR2 et 6 (5). Les neutrophiles produisent des chimiokines supplémentaires,
principalement CXCL2, induisant une migration supplémentaire des neutrophiles (6). Les
neutrophiles dégradent la matrice cornéenne, provoquant une opacification de la cornée et une
déficience visuelle. Repris de [57].

43
Ces réactions sont associées à une grave pathologie cutanée. Chez ces patients le
nombre de microfilaires dermiques est très faible et des infiltrats cellulaires massifs sont
observés dans les onchocercomes qui ne contiennent que peu de parasites [99]. Les quelques
microfilaires vivantes ou mortes se retrouvent principalement entourées d'éosinophiles et de
macrophages, et plus rarement de neutrophiles, formant de multiples micro-granulomes [99].
Les patients sowda présentent des niveaux plus faibles de TGF-β [102], et des réponses de type
Th2 et Th17 accrues [106]. De plus, les éosinophiles et des neutrophiles montrent des capacités
chimiotactiques plus importantes [107]. L'action de ces cellules permettrait une destruction
efficace des microfilaires, mais au prix d'une pathologie cutanée accrue. En effet, les patients
présentent des plaques hyperpigmentées sur la peau, d'intenses démangeaisons et des œdèmes
sévères. Des surinfections bactériennes peuvent également se produire en raison des plaies
dues aux démangeaisons [108].
Chez les patients présentant des charges importantes en microfilaires, celles-ci peuvent
migrer dans le tissu conjonctif des yeux et la cornée. Les yeux sont un organe immuno-
privilégié où en temps normal les cellules immunitaires ne peuvent pas pénétrer.
L'accumulation de microfilaires mortes induit une rupture de cette protection et une
inflammation de la cornée et une infiltration de macrophages, de neutrophiles et
d'éosinophiles. L'étude de modèles murins suggère que Wolbachia joue un rôle important
dans le développement des pathologies dues à O. volvulus. La mort des microfilaires dans la
stroma de la cornée permettrait la libération de la bactérie et sa détection par les cellules
résidentes de la cornée grâce au récepteur TLR2 [109]. Ensuite, la production de cytokines pro-
inflammatoires telles que l'IL-1β, l'interféron (IFN)-γ, CXCL1 et CXCL2 [57,109,110] entrainent
le recrutement de neutrophiles et de macrophages par l'intermédiaire des récepteurs CCR2 et
IL-1R1 [57,109-114]. La dégranulation des neutrophiles avec la libération d'oxyde nitrique, de
myelopéroxydase (MPO) et de réactifs de l'oxygène en association avec les molécules
produites par les éosinophiles pourraient participer à la mise en place de la pathologie. Si
l'inflammation perdure, cela peut entraîner une fibrose de la cornée conduisant à la perte
progressive de la vue [57,99].

44
2.3.3 Loase
De nombreux cas de Loa loa sont asymptomatiques et plus de 40% des personnes
infestées par L. loa présentent une loase occulte sans microfilaires détectables [94,115]. La
pathogénèse des infections à L. loa est encore largement méconnue. Cependant, contrairement
aux autres filarioses, L. loa n'héberge pas Wolbachia. Les symptômes observés ne sont donc
dus qu'à la présence du parasite et non à celle de la bactérie [116].
Symptômes associés aux adultes
Les parasites adultes migrent continuellement dans les tissus sous cutanés où ils sont
palpables et ne causent pas de symptômes particuliers. Il arrive que les parasites migrent dans
les tissus sous-conjonctifs de l'œil où ils sont alors visibles [117-119]. Ce passage oculaire peut
durer de quelques minutes à plus d'une journée [62,115]. Il est associé à une inflammation
locale, des démangeaisons, une sensibilité à la lumière et une douleur sévère [115]. Après le
passage des parasites, les symptômes disparaissent rapidement sans laisser de séquelles.
Des œdèmes appelés "œdèmes de Calabar" sont souvent observés chez les personnes
infestées par L. loa. Ils ne sont pas persistants et sont associés à des démangeaisons localisées
ou généralisées. Ils peuvent se produire à distance des parasites adultes, et sont plus
fréquemment observés sur les membres, en particulier les avant-bras. Ils disparaissent
spontanément, généralement après plusieurs jours, et réapparaissent, à intervalles irréguliers
[62]. Ces œdèmes sont des angioœdèmes (c'est-à-dire d'un type allergique) et peuvent se
produire chez des individus amicrofilarémiques, suggérant qu'ils seraient dus à la présence
des adultes [62]. Ils sont observés chez 50% des patients vivant en zone endémique, mais
jusqu’à 80 et 90% des résidents temporaires [120-122]. La pathogénèse des œdèmes n'est pas
connue mais elle est associée aux éosinophiles. Les résidents temporaires présentent une
éosinophilie sanguine et des IgG spécifiques pour les filaires plus élevées que les personnes
vivant en zone d'endémie [121,123]. De plus, de nombreuses molécules issues des granules
éosinophiles sont retrouvées dans le sang des résidents temporaires [121]. Cette différence
pourrait être due à une tolérance au parasite chez les personnes vivant dans les zones
d'endémie entraînant de plus hautes microfilarémies mais des œdèmes moins intenses et
moins nombreux [62,121]. Les neutrophiles ne semblent pas participer aux réponses contre les
adultes de L. loa, peut-être à cause de l'absence de Wolbachia. En effet, l'étude de biopsies a
montré l'absence de neutrophiles à proximité des parasites [116].

45
Symptômes associés aux Mfs
Une des particularités de la loase est que les patients peuvent présenter des
microfilarémies considérables comparées aux autres filarioses [121]. Les individus présentant
une forte microfilarémie sont généralement asymptomatiques. Cependant, chez ces patients, la
prise d’ivermectine, utilisés contre W. bancrofti ou O. volvulus (voir 4.2), peut induire de
graves encéphalopathies pouvant conduire au coma et à la mort [122,124-126]. Toutes les
personnes présentant une forte microfilarémie ne développent cependant pas ces effets
secondaires après traitement [127] ; il est probable que d'autres facteurs, comme des
coïnfections, puissent participer au développement de l'encéphalopathie [128]. Une fois de plus,
très peu de données sont disponibles quand à l'origine de ce syndrome [128]. L'ivermectine
induit une paralysie des microfilaires en bloquant leurs canaux chlorure post-synaptiques [129].
La pathologie viendrait d'une embolie des vaisseaux sanguins cérébraux par les microfilaires
et de l'inflammation associée à la mort des parasites [127]. Les microfilaires peuvent aussi
traverser la barrière hématoencéphalique, entraînant une inflammation du tissu cérébral, avec
des infiltrats de macrophages et de lymphocytes [126] et la présence de microfilaires et de sang
dans le fluide cérébrospinal [125,130].

46
Sous famille Parasite Hôte
(expérimental/accidentel) Stade décrit Localisation pulmonaire Ref
Oswaldofilariinae Oswaldofilaria bacillaris
(Molin, 1858) Crocodylidae Adultes Poumons entiers [2]
Oswaldofilaria chlamydosauri
(Breinl, 1913)
Agamidae
(Agamidae) Adultes Poumons entiers [131]
Conispiculum flavescens
(Castellani et Willey, 1905) Agamidae Adultes Poumons entiers [132]
Piratuba queenslandensis
Mackerras, 1962 Varanidae Adultes Poumons entiers [131]
Piratuba varanicola Mackerras,
1962 Varanidae Adultes Poumons entiers [131]
Dirofilariinae Dirofilaria immitis
(Leidy, 1856) Canidae Adultes, mf Artères pulmonaires, Bronches [133]
(Humains) Adultes Poumons entiers [134]
Dirofilaria spectans
Freitas et Lent, 1949 Mustelidae Adultes Artères pulmonaires [135]
Foleyella dolichoptera
Wehr et Causey, 1939 Ranidae Adultes Tissu pulmonaire [136]
Onchocercinae Acanthocheilonema spirocauda
(Leidy, 1858) Phocidae Adultes
Poumons entiers,
Artères pulmonaires [137]
Acanthocheilonema viteae
(Krepkogorskaya, 1933)
nd,
(Rongeurs) L3, L4 Poumons entiers [10]
Bisbalia vossi
Bain & Guerrero, 2003 Geomyoidea Adultes Cavité pleurale [138]
Brugia pahangi
(Buckley & Edeson, 1956)
nd,
(Rongeurs) L3, L4, Adultes
Poumons entiers,
Artères pulmonaires [10,139,140]
Brugia malayi
(Brug, 1927) Humains mf Poumons entiers [81]
Brugia malayi
(Brug, 1927) (Rongeurs) L4, Adultes, mf
Poumons entiers,
Artères pulmonaires [10,140]
Brugia buckleyi Dissanaike et
Paramananthan, 1961 Leporidae Adultes Artères pulmonaires [141]
Cardiofilaria chabaudi
Dissanaike et Fernando, 1965 Cuculidae Adultes Poumons entiers [142]
Deraiophoronema evansi
(Lewis, 1882) Camelidae Adultes Artères pulmonaires [143]
Litomosoides petteri
Bain, Petit et Berteaux, 1980 Didelphidae Adultes Cavité pleurale [144]
Litomosoides sigmodontis
Chandler, 1931
Rongeurs
(Rongeurs) Adultes, L3, mf
Cavité pleurale
Tissu pulmonaire [10] [11]
Mansonella perstans
(Manson, 1891) Humains Adultes Cavité pleurale [22]
Molinema sprendi
(Anderson, 1953) Castoridae Adultes Cavité pleurale [2]
Monanema globulosa
(Muller et Nelson, 1975)
nd,
(Rongeurs) L4, Adultes
Poumons entiers
Artères pulmonaires [10]
Monanema martini
Bain, Bartlett et Petit, 1986
nd,
(Rongeurs) L3, L4, Adultes Artères pulmonaires [10]
Wuchereria bancrofti
(Cobbold, 1877) Humains Mf Cavité pleurale [145]
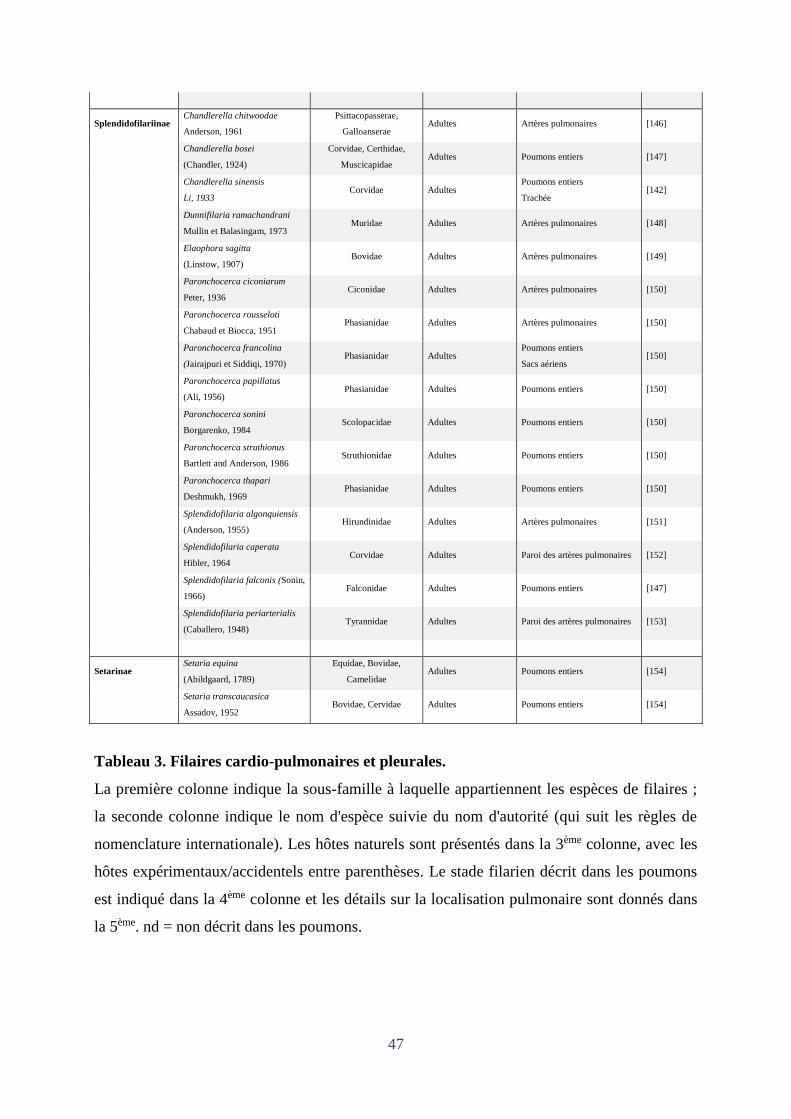
47
Splendidofilariinae Chandlerella chitwoodae
Anderson, 1961
Psittacopasserae,
Galloanserae Adultes Artères pulmonaires [146]
Chandlerella bosei
(Chandler, 1924)
Corvidae, Certhidae,
Muscicapidae Adultes Poumons entiers [147]
Chandlerella sinensis
Li, 1933 Corvidae Adultes
Poumons entiers
Trachée [142]
Dunnifilaria ramachandrani
Mullin et Balasingam, 1973 Muridae Adultes Artères pulmonaires [148]
Elaophora sagitta
(Linstow, 1907) Bovidae Adultes Artères pulmonaires [149]
Paronchocerca ciconiarum
Peter, 1936 Ciconidae Adultes Artères pulmonaires [150]
Paronchocerca rousseloti
Chabaud et Biocca, 1951 Phasianidae Adultes Artères pulmonaires [150]
Paronchocerca francolina
(Jairajpuri et Siddiqi, 1970) Phasianidae Adultes
Poumons entiers
Sacs aériens [150]
Paronchocerca papillatus
(Ali, 1956) Phasianidae Adultes Poumons entiers [150]
Paronchocerca sonini
Borgarenko, 1984 Scolopacidae Adultes Poumons entiers [150]
Paronchocerca struthionus
Bartlett and Anderson, 1986 Struthionidae Adultes Poumons entiers [150]
Paronchocerca thapari
Deshmukh, 1969 Phasianidae Adultes Poumons entiers [150]
Splendidofilaria algonquiensis
(Anderson, 1955) Hirundinidae Adultes Artères pulmonaires [151]
Splendidofilaria caperata
Hibler, 1964 Corvidae Adultes Paroi des artères pulmonaires [152]
Splendidofilaria falconis (Sonin,
1966) Falconidae Adultes Poumons entiers [147]
Splendidofilaria periarterialis
(Caballero, 1948) Tyrannidae Adultes Paroi des artères pulmonaires [153]
Setarinae Setaria equina
(Abildgaard, 1789)
Equidae, Bovidae,
Camelidae Adultes Poumons entiers [154]
Setaria transcaucasica
Assadov, 1952 Bovidae, Cervidae Adultes Poumons entiers [154]
Tableau 3. Filaires cardio-pulmonaires et pleurales.
La première colonne indique la sous-famille à laquelle appartiennent les espèces de filaires ;
la seconde colonne indique le nom d'espèce suivie du nom d'autorité (qui suit les règles de
nomenclature internationale). Les hôtes naturels sont présentés dans la 3ème colonne, avec les
hôtes expérimentaux/accidentels entre parenthèses. Le stade filarien décrit dans les poumons
est indiqué dans la 4ème colonne et les détails sur la localisation pulmonaire sont donnés dans
la 5ème. nd = non décrit dans les poumons.

48
3 Les filarioses pulmonaires, cardio-pulmonaires et pleurales
La présence de filaires dans l'environnement pulmonaire (poumon sensu stricto et/ou
système cardio-pulmonaire et/ou cavité pleurale) a été décrite chez de nombreuses espèces de
filaires dans un large éventail d'hôtes comprenant des oiseaux, des crocodiles, des chiens et
les humains (Tableau 3). La plupart des études ont documenté la présence de vers adultes ou
de microfilaires, mais très rarement des L3 du fait de leur faible nombre et du caractère
transitoire de leur migration.
La/les voies de migration des filaires présentant des localisations pulmonaires ne sont
pas connues mais pourraient s'expliquer anatomiquement : lors de la migration lymphatique
les larves s'engageraient dans le canal thoracique et arriveraient ainsi dans la veine cave
supérieure, le cœur droit puis les artères pulmonaires. De là, suivant l’espèce filarienne
considérée, les L3 pourraient s'installer dans la circulation sanguine pulmonaire (comme
Dirofilaria immitis), pénétrer le tissu pulmonaire (comme certaines Oswaldofilariae de
reptiles) ou rejoindre la cavité pleurale (M. perstans ou des espèces des genres Litomosoides).
Chez d'autres espèces (par exemple B. malayi chez les rongeurs) la présence d'adultes dans les
poumons est parfois décrite, elle pourrait être accidentelle et résulter de parasites emportés par
le flux lymphatique à travers le canal thoracique jusqu'à la circulation sanguine pulmonaire
[10].
La présence de filaires dans l'environnement pulmonaire est souvent considérée
globalement asymptomatique comme chez M. perstans mais peut causer de graves
pathologies dans les dirofilarioses pulmonaires dues aux adultes de D. immitis ou dans le
Poumon Eosinophile Tropical induit par les microfilaires de W. bancrofti et B. malayi.
La biologie et les interactions avec les poumons de D. immitis et de M. perstans sont
développées ci-après, de même que celles de Litomosoides sigmodontis, le modèle murin
utilisé lors de cette étude.

49
3.1 Dirofilaria immitis
Biologie et cycle de vie
Les dirofilarioses sont dues aux filaires du genre Dirofilaria qui comprend une
trentaine d’espèces [155]. D. immitis (Leidy, 1856) et D. repens (Railliet et Henry, 1911) sont
les agents d’importantes filarioses canines en raison de leurs effets pathologiques et de leur
forte prévalence et incidence [134]. D. immitis et D. repens présentent un large spectre d’hôtes
comprenant de nombreuses espèces de mammifères [155] mais les chiens domestiques et
sauvages sont les hôtes privilégiés de ces filaires. Les vecteurs sont des moustiques de la
famille Culicidae [156] appartenant aux genres Culex, Aedes, Anopheles, Mansonia,
Armigeres, Ochlerotatus, et Coquillettidia [157].
Les infections par D. immitis ont lieu dans les régions tropicales et tempérées du
monde entier alors que la présence D. repens est restreinte à l'Europe, l'Afrique et l'Asie. De
nombreux cas zoonotiques ont été décrits chez l’homme pour ces deux espèces. D. immitis est
responsable des cas humains de dirofilarioses pulmonaires, majoritairement localisés en
Amérique [134]. Dans ces zoonoses, les filaires ne finissent pas leur développement et meurent
avant la production de microfilaires [158] [134] [155].
Chez le chien, les adultes de D. immitis vivent dans les artères pulmonaires et le
ventricule droit du cœur. Les adultes sont identifiables 70 jours post infection dans ces
localisations. Toutefois, la voie de migration qu'empruntent les L3 après inoculation dans la
peau pour s’y rendre n’est pas connue. Lors d'autopsies de chiens et de furets infestés
expérimentalement, la localisation des parasites à différents temps après infection a été
étudiée dans la peau, les muscles, les cavités thoraciques et abdominales, le cœur et l'artère
pulmonaire [159] [160] [161] [162]. La figure 8 synthétise les résultats de ces études. Le rendement
parasitaire est relativement stable dans ces organes pendant 70 jours après l'inoculation des L3
avec une moyenne de 30% des larves inoculées qui sont retrouvées (les rendements un peu
plus faibles dans les temps très précoces sont probablement dus au fait que les L3 sont plus
difficiles à trouver que les L4 et les adultes). Durant cette période, les parasites ont été
récupérés dans les tissus sous-cutanés et les muscles, parfois à distance du point d'inoculation,
où ils semblent pouvoir se développer jusqu'au stade jeune adulte.
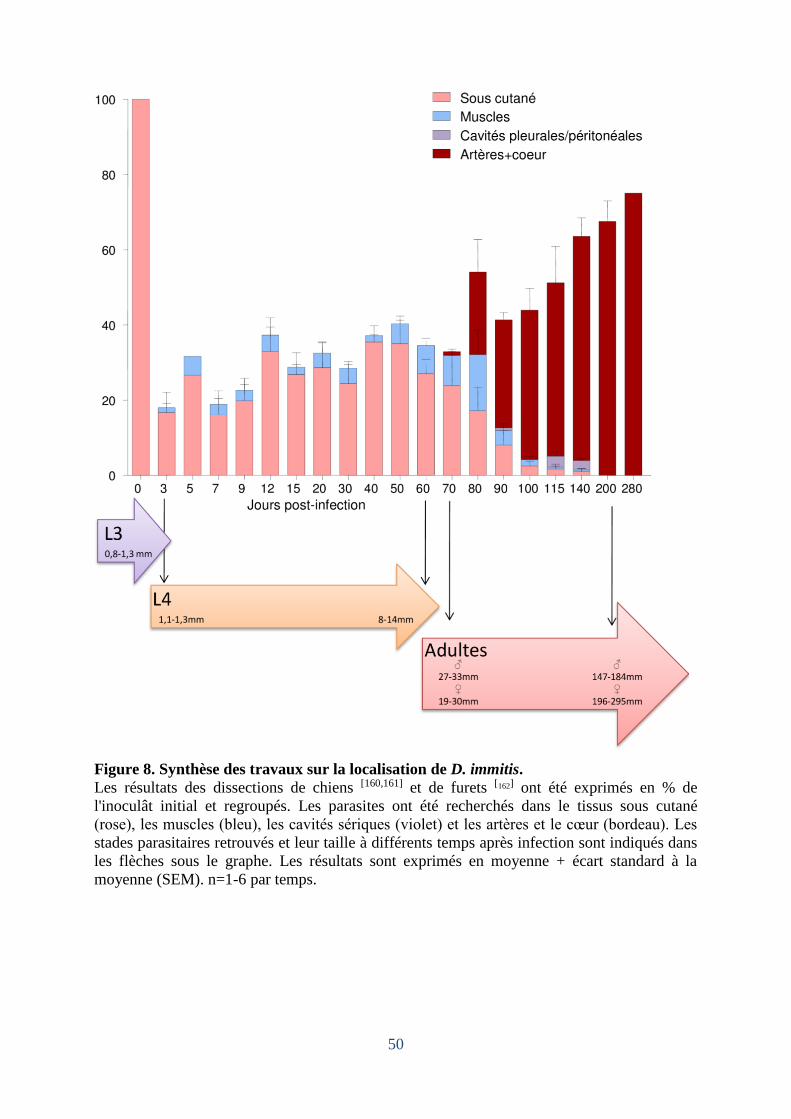
50
Figure 8. Synthèse des travaux sur la localisation de D. immitis.
Les résultats des dissections de chiens [160,161] et de furets [162] ont été exprimés en % de
l'inoculât initial et regroupés. Les parasites ont été recherchés dans le tissus sous cutané
(rose), les muscles (bleu), les cavités sériques (violet) et les artères et le cœur (bordeau). Les
stades parasitaires retrouvés et leur taille à différents temps après infection sont indiqués dans
les flèches sous le graphe. Les résultats sont exprimés en moyenne + écart standard à la
moyenne (SEM). n=1-6 par temps.

51
A partir de 70 jours après infection, le nombre de filaires dans la peau et les muscles
chute alors que celui des parasites présents dans les artères pulmonaires et le cœur augmente
progressivement et peut atteindre plus de 70% de l'inoculât initial ([160] et Figure 8).
L'hypothèse qui a alors été émise (Kume et Itagaki 1955 [159], Kotani et Powers 1982 [161],
Figure 9 - Hypothèse 1) est que les L3 migreraient dans les tissus sous-cutanés et les muscles
où seraient réalisées les 3ème et 4ème mues. Après 60 à 90 jours, les parasites adultes entreraient
dans une veine et rejoindraient les artères pulmonaires et le cœur droit, portés par la
circulation veineuse. Cette hypothèse pourrait être étayée par le fait que des jeunes adultes,
récupérés après leur migration dans la circulation sanguine pulmonaire, sont capables de re-
migrer vers les artères pulmonaires après une transplantation dans les tissus sous-cutanés de
chiens non-infestés [159,163]. Il semble néanmoins étonnant qu'un parasite mesurant plusieurs
centimètres arrive à pénétrer l'épaisse paroi des vaisseaux sanguins, sans engendrer de fortes
hémorragies.
La limitation principale de ces études est que ni le système lymphatique, ni le tissu
pulmonaire n'ont été analysés. En effet, l'idée d'un passage lymphatique avait été rejetée par
Kume (1955) en argumentant que les larves du nématode Metastrongylus elongatus migrent
par les lymphatiques en causant des hémorragies des ganglions lymphatiques. On sait
aujourd'hui que de nombreuses filaires migrent à travers le circuit lymphatique sans induire de
tels dégâts [10,164]. Il est donc possible, comme l'a suggéré Wenk (1967 [11]) que les larves
pénètrent rapidement dans le système lymphatique avant de remonter le canal thoracique pour
rejoindre la circulation sanguine pulmonaire. Dans ce cas là, les 30% de larves observées dans
la peau/les muscles dans les temps précoces de l'infection correspondraient à celles qui ont
échoué à pénétrer les vaisseaux lymphatiques et qui périraient progressivement après leur
dernière mue; les 70% restants rejoindraient les artères pulmonaires où elle pourraient
atteindre la maturité sexuelle.
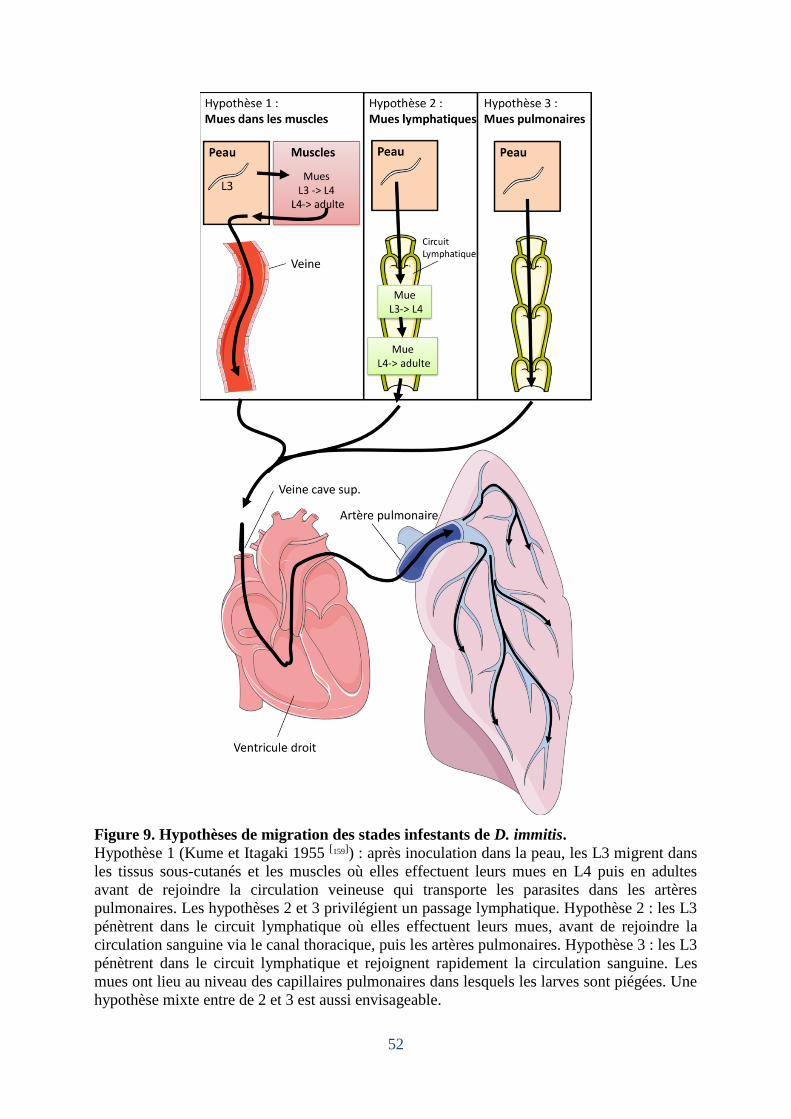
52
Figure 9. Hypothèses de migration des stades infestants de D. immitis.
Hypothèse 1 (Kume et Itagaki 1955 [159]) : après inoculation dans la peau, les L3 migrent dans
les tissus sous-cutanés et les muscles où elles effectuent leurs mues en L4 puis en adultes
avant de rejoindre la circulation veineuse qui transporte les parasites dans les artères
pulmonaires. Les hypothèses 2 et 3 privilégient un passage lymphatique. Hypothèse 2 : les L3
pénètrent dans le circuit lymphatique où elles effectuent leurs mues, avant de rejoindre la
circulation sanguine via le canal thoracique, puis les artères pulmonaires. Hypothèse 3 : les L3
pénètrent dans le circuit lymphatique et rejoignent rapidement la circulation sanguine. Les
mues ont lieu au niveau des capillaires pulmonaires dans lesquels les larves sont piégées. Une
hypothèse mixte entre de 2 et 3 est aussi envisageable.

53
Une fois dans le système lymphatique il existe encore au moins 2 options (Figure 9-hypothèse
2 et 3):
• Les parasites effectuent leurs mues dans le système lymphatique pour rejoindre la
circulation sanguine avant leur dernière mue. Cela est d'autant plus probable que la
migration d'adultes du circuit lymphatique vers la circulation sanguine a été décrite
chez Monanema martini¸ notamment après des traitements [10,165].
• Les larves peuvent rejoindre directement la circulation sanguine pulmonaire par le
canal thoracique et rester bloquées dans les capillaires pulmonaires. Ensuite les
parasites continueraient leur développement dans les capillaires ou les alvéoles avant
de remonter vers les artères pulmonaires. Cette hypothèse semble moins plausible car
dans ce cas, il est probable que des symptômes respiratoires apparaitraient avant la
dernière mue.
Ces deux hypothèses sont soutenues par la mue en L4 très précoce des larves de D.
immitis (vers 2-3 jours p.i.). En effet, la plus grande taille des L4 pourrait permettre
d'augmenter les chances qu'elles soient bloquées dans la circulation pulmonaire ou qu'elles se
maintiennent dans le flux lymphatique. Il est aussi possible que D. immitis puisse utiliser
différentes voies de migration pour atteindre les artères pulmonaires : une partie des larves
pénètreraient rapidement dans la circulation lymphatique pour poursuivre leur développement
et les autres matureraient dans la peau avant de migrer.
Pathologie pulmonaire
Les parasites sont tout d'abord retrouvés dans les artères pulmonaires profondes
(Figure 9), où ils induisent des embolies (obstruction des vaisseaux sanguins) et des lésions
vasculaires. La sévérité des lésions dépend du nombre de parasites présents dans les poumons,
pouvant aller de 1 à 250 [166]. Au fur et à mesure que le parasite croît en taille, il remonte à
contre-courant vers l'artère pulmonaire principale. Ceci est probablement rendu possible par le
fait que la circulation sanguine pulmonaire est une circulation à basse pression [167].
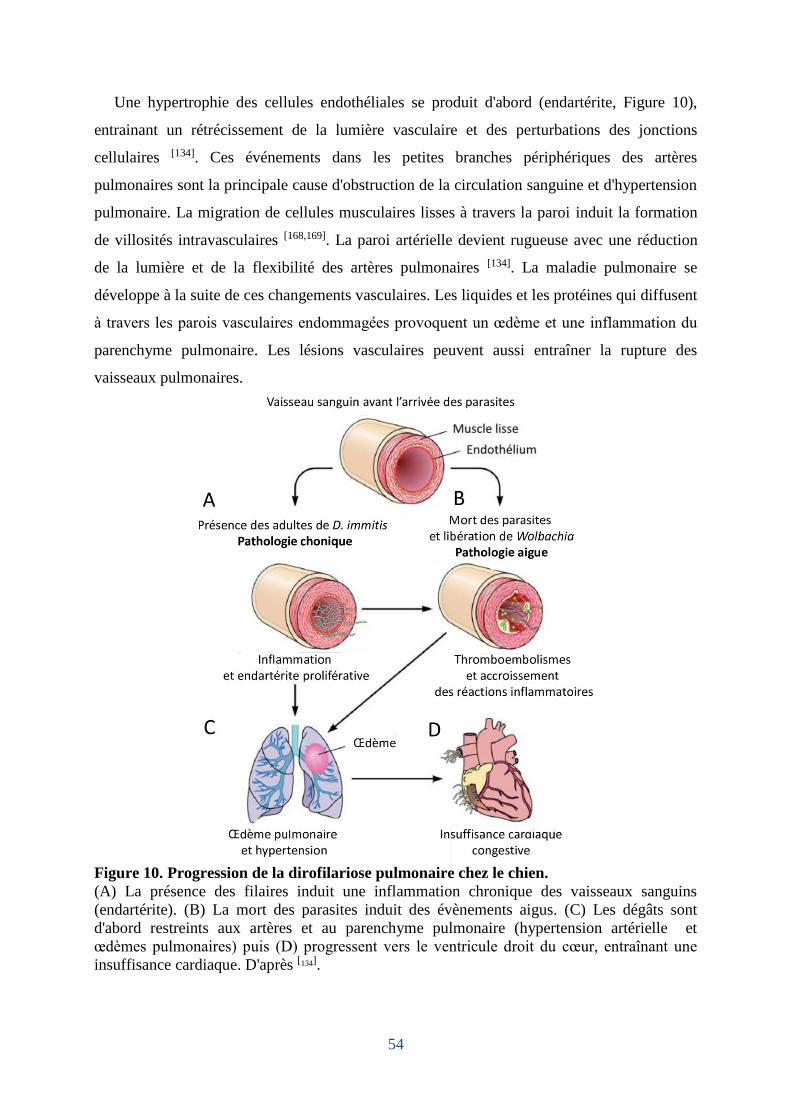
54
Une hypertrophie des cellules endothéliales se produit d'abord (endartérite, Figure 10),
entrainant un rétrécissement de la lumière vasculaire et des perturbations des jonctions
cellulaires [134]. Ces événements dans les petites branches périphériques des artères
pulmonaires sont la principale cause d'obstruction de la circulation sanguine et d'hypertension
pulmonaire. La migration de cellules musculaires lisses à travers la paroi induit la formation
de villosités intravasculaires [168,169]. La paroi artérielle devient rugueuse avec une réduction
de la lumière et de la flexibilité des artères pulmonaires [134]. La maladie pulmonaire se
développe à la suite de ces changements vasculaires. Les liquides et les protéines qui diffusent
à travers les parois vasculaires endommagées provoquent un œdème et une inflammation du
parenchyme pulmonaire. Les lésions vasculaires peuvent aussi entraîner la rupture des
vaisseaux pulmonaires.
Figure 10. Progression de la dirofilariose pulmonaire chez le chien.
(A) La présence des filaires induit une inflammation chronique des vaisseaux sanguins
(endartérite). (B) La mort des parasites induit des évènements aigus. (C) Les dégâts sont
d'abord restreints aux artères et au parenchyme pulmonaire (hypertension artérielle et
œdèmes pulmonaires) puis (D) progressent vers le ventricule droit du cœur, entraînant une
insuffisance cardiaque. D'après [134].

55
Parallèlement à la progression chronique de la maladie, la mort des adultes provoque
des événements aigus caractérisés par une thromboembolie (obstruction de vaisseaux dus à
des caillots sanguins et à des fragments de filaires mortes) et une inflammation sévère qui
peuvent être létale pour l'animal ([170,171] et Figure 10). L'altération de la paroi artérielle et la
réduction du diamètre de la lumière des artères pulmonaires, qui peuvent être occultées par les
villosités, les thrombus et/ou la présence de vers, en conjonction avec des médiateurs
inflammatoires, peuvent entraîner une hypertension pulmonaire. Cela génère une surcharge
sanguine dans le ventricule droit du cœur, induisant une insuffisance cardiaque congestive
[172].
La dirofilariose pulmonaire humaine se caractérise par la formation de nodules
pulmonaires autour de jeunes adultes morts. Lorsque les parasites atteignent une branche
petite ou moyenne de l'artère pulmonaire, ils en bloquent le passage, provoquant une embolie
et une inflammation localisée [173,174]. Des zones fibrotiques et nécrotiques ainsi que des
granulomes peuvent être observées dans les poumons autour des parasites morts [173-176].
Différents traitements existent pour éliminer les parasites des chiens atteints par D.
immitis. L’élimination des parasites n’est pas sans risque en particulier quand l’infestation est
importante du fait des effets secondaires liés à la destruction massive de vers dans la
circulation sanguine. Comme chez l'humain, des lactones macrocycliques, comme
l'ivermectine, sont utilisées pour éliminer les microfilaires et arrêter la transmission. Il existe
néanmoins un traitement capable de tuer les adultes. La mélarsomine hydrochloride
(Immiticide), est une composé organo-arsénié ayant une efficacité supérieure à 96% pour tuer
les parasites adultes [166] mais son mode d'action n'est pas connu. La thromboembolie
pulmonaire est une conséquence inévitable du traitement macrofilaricide et peut être grave
voire mortelle chez les chiens fortement infectés [166,177]. Les risques sont généralement
minimes chez les animaux porteurs d’un petit nombre de vers adultes car une embolie légère
peut passer cliniquement inaperçue [178]. Un choc anaphylactique (Hypersensibilité de type I)
peut aussi faire suite à la libération massive d’antigènes suite à la destruction des parasites
[166].

56
3.2 Mansonella perstans
Biologie et cycle de vie
Le cycle de vie de Mansonella perstans est mal connu. Plusieurs espèces de
Culicoides peuvent transmettre les larves de M. perstans [66]. Les adultes de M. perstans sont
rarement observés, mais ils ont été occasionnellement récupérés dans les cavités sériques
(cavités péritonéale, pleurale, et péricardique) lors de chirurgies ou d'autopsies [22].
Les microfilaires sont dépourvues de gaine et mesurent environ 190-200µm de long
par 4-5µm de diamètre. Les Mfs vivantes ont de fortes capacités de contraction de
d'élongation ce qui fait que les tailles décrites varient beaucoup [22,68]. Les Mfs sont sanguines
[69].
Pathologie pulmonaire
Les adultes de M. perstans s'installeraient majoritairement dans les cavités sériques,
dont la cavité pleurale. Les infections par M. perstans ne sont pas associées à un tableau
clinique précis. Pour cette raison, elles ne sont pas considérées comme un problème de santé
publique. Les rapports cliniques traitent majoritairement des expatriés qui retournent dans leur
pays d'origine ou des patients ayant accès à des établissement médicaux [22]. La plupart des
manifestations signalées suggèrent que les symptômes sont plutôt liées à la présence des
parasites adultes dans les cavités sériques et moins aux microfilaires ou à d'autres stades
larvaires [22]. M. perstans peut provoquer une éosinophilie sanguine [179] pouvant atteindre une
hyper éosinophilie chez les expatriés [180], des gonflements sous-cutanés, des douleurs et des
éruptions cutanées chez de nombreux patients [22]. Certains cas de pathologies associées aux
poumons ont aussi été décrits: une hypertension pulmonaire chez un enfant [180] mais surtout
des effusions pleurales associées à des microfilaires pleurales [181,182]. Si la présence d'adultes
dans la cavité pleurale n'a pas été démontrée dans ces cas, il est possible que les microfilaires
retrouvées aient été produites localement et soient responsables de l'inflammation pleurale.
Plus récemment, une tomographie assistée par ordinateur d'une patiente infestée par M.
perstans a révélé des infiltrats péribronchiques [183].

57
3.3 Litomosoides sigmodontis
Modèle murin de filariose- justifications
L'étude des filaires humaines et vétérinaires est limitée par le fait qu'aucune ne peut
effectuer son cycle de vie complet chez la souris immunocompétente. Il est préférable d’avoir
un modèle murin car peu d'outils immunologiques sont disponibles pour étudier la réponse
immunitaire chez d’autres rongeurs comme la gerbille dans laquelle peut se développer B.
malayi [184]. Chez la souris, les études de B. malayi sont limitées à l'injection ou à
l'implantation de L3, d'adultes ou de microfilaires, qui survivent de quelques semaines (larves
et microfilaires) à des mois (adultes), en fonction de la souche et du sexe de la souris [185-187]
[82,188].
Loa loa peut se développer partiellement chez des souris immunodéficientes avec
traitements par des corticostéroïdes [189] ou suite à des modifications génétiques [190]. Chez les
souris IL-4R/IL5 double déficientes, les parasites survivent au moins 70 jours et atteignent le
stade adulte, indiquant un rôle critique de ces molécules dans la résistance des souris à L. loa.
L'étude n'a pas été prolongée assez longtemps pour observer une éventuelle microfilarémie
[190]. La croissance d'Onchocerca volvulus et les réponses immunitaires de l'hôte ont été
étudiées chez la souris en implantant des chambres de diffusions contenant des L3 sous la
peau de souris [191-193]. Dans les chambres de diffusions, les larves effectuent leur mue en L4
et peuvent survivre plus de 3 semaines [193].
L'avantage de L. sigmodontis est que cette filaire peut effectuer son cycle de vie
complet chez la souris et permet donc une étude approfondie des relations hôte-parasite en
particulier pour la réponse immunitaire, des essais vaccinaux et des tests de drogues anti-
filariennes [55]. De plus L. sigmodontis se développe différemment dans la souris selon la
souche considérée. En effet, les souris BALB/c permettent un développement complet du
parasite jusqu'à la phase patente; les souris CBA/ca sont semi-résistantes car elles ne
développent pas de phase patente avec production de microfilaires mais les filaires atteignent
un stade adulte qui survit autant que dans les BALB/c; les souris C57BL/6 sont résistantes, les
adultes meurent rapidement entre 30 et 50 jours post infection [194-196]. L’analyse comparée de
ces différentes souches peut être utilisée pour caractériser les processus déterminants dans la
résistance et la susceptibilité aux parasites [195].
Enfin L. sigmodontis présente de nombreuses similarités avec Mansonella perstans,
dont la localisation pleurale des adultes (Tableau 4).

58
M. perstans chez l’humain L. sigmodontis chez
souris BALB/c
L. sigmodontis chez la
gerbille
Taille L3 600-900µm 750-800µm 750-800µm
Mue en L4 9 à 10 jours p.i. 9 à 10 jours p.i. 9 à 10 jours p.i.
Taille mâle adulte 35-45mm x 50-60µm 20±5mm x 130µm 25±5mm x 130µm
Taille femelle
adulte
50-80mm x 80-120µm 75±10mm x 220-255µm 90±10mm x 220-255µm
Localisation
adultes
Cavité pleurale, cavité
péritonéale, péricarde.
Cavité pleurale Cavité pleurale, cavité
péritonéale.
Taille Mfs ~ 190 x 4-5 µm ~ 80 x4-5µm ~ 80 x4-5µm
Localisation Mfs Sang Sang Sang
Gaine Mfs NON OUI OUI
Périodicité +/- NON NON
Wolbachia
(supergroupe)
OUI (F) OUI (D) OUI (D)
Tableau 4. Comparaison entre M. perstans et L. sigmodontis.
Des caractéristiques de L3, de L4, d'adultes et de Microfilaires (Mfs) pour M. perstans sont
présentées dans la colonne 1et la seconde celles de L. sigmodontis.
Figure 11. Cycle de vie de L. sigmodontis.
Les L3 sont inoculées dans l’hôte murin lors d'un repas sanguin et migrent jusqu’à la cavité
pleurale en 7-8 jours. À ce niveau elles vont muer vers 9-10 jours post infection (p.i.) en L4
puis vers 30 jours en adultes mâles et femelles qui se reproduisent et libèrent des microfilaires
dans la cavité pleurale qui rejoignent le sang environ 55-60 jours après infection. Les
microfilaires (L1) sont ensuite ingérées par le vecteur acarien lors d'un repas sanguin dans
lequel elles muent jusqu'au stade L3.

59
Biologie et cycle de vie
26 espèces sont actuellement décrites dans le genre Litomosoides [1]. Elles vivent dans
les cavités pleurale et/ou péritonéale des rongeurs, des chauves-souris et de petits marsupiaux
[4,197]. Litomosoides sigmodontis (Chandler, 1931) a pour hôte naturel Sigmodon hispidus (le
rat du coton) et son vecteur est l'acarien Ornithonyssus bacoti [194,198].
Les larves inoculées de L. sigmodontis migrent du site d’inoculation au système
lymphatique et se retrouvent dans la cavité pleurale en 7-8 jours [10,11,199]. Quelques larves
peuvent être retrouvées dans la cavité péritonéale chez le rat du coton et le mérion, mais pas
chez la souris ( [10,200] et Figure 11). La mue en L4 a lieu entre 9 et 10 jours après inoculation
puis une mue autour de 30 jours donne des adultes sexués capables de se reproduire [201-203].
La femelle fécondée pond des Mfs dans la cavité pleurale qui rejoignent le sang où elles sont
identifiables 55-60 jours après inoculation [201,203]). Dans le cycle naturel, les Mfs sont
ingérées par l'acarien pendant un repas sanguin. Les larves peuvent se développer dans de
nombreux organes de l'acarien comme le tissu interstitiel, les glandes salivaires ou les glandes
coaxales [198]. Elles muent en stade L2 en 5 à 7 jours, puis en L3 vers le douzième jour. Les
larves infestantes migrent alors vers l'hémocèle d'où elles s'échapperont et atteindront la peau
de l'hôte du vertébré lors d'un repas sanguin [9].
Pathologie pulmonaire
Lors de leur phase de migration, des L3 de L. sigmodontis ont été observées dans les
poumons dilacérés mécaniquement des souris BALB/c deux jours après inoculation [204] et
encore plus tôt dans leur hôte naturel, le rat du coton [11]. Des granulomes constitués de
lymphocytes T et de macrophages ont aussi été observés dans les poumons de souris infestées
7 jours p.i. [205]. Au cours du développement de la filaire dans la cavité pleurale le volume de
l’exsudat pleural augmente et il contient de nombreux leucocytes avec un pic de cellularité
observé à J30 p.i. [196], ce pic étant plus prononcé chez les souris C57BL/6 que chez les
BALB/c [195,196,202,206]. De plus une hypertrophie des cellules mésothéliales a été observée dans
les souris C57BL/6 30 jours après infection [196].
Plus tard lors de l'infection, les femelles pondent les microfilaires dans la cavité
pleurale; les Mfs rejoignent la circulation sanguine par une voie migratoire inconnue.
Les travaux de Wharton (1946 [207]) et Schneider (1968 [208]) ont décrit des lésions
pulmonaires chez le rat du coton (Sigmodon hispidus) et la gerbille (Meriones unguiculatus)

60
en phase patente d’infestation par L. sigmodontis. Ils ont observé une hyperplasie et une
prolifération des cellules mésothéliales avec la formation de nodules papillaires et une fibrose
de la plèvre. Des microfilaires ont été observées dans la vasculature pulmonaire, mais aussi
dans le parenchyme et des infiltrats cellulaires sont présents autour des bronches [207,208].

61
4 Programmes multivalents visant à « traiter » les filarioses
4.1 Traitements à grande échelle ou traitements de masse (Mass Drug
Administration, MDA)
4.1.1 Programme mondial pour l’élimination de la filariose lymphatique
(GPELF)
L'Organisation mondiale de la santé (OMS) a lancé en 2000 le Programme mondial
pour l’élimination de la filariose lymphatique (GPELF) avec comme objectif l’élimination de
cette maladie d'ici à 2020. La stratégie repose sur un arrêt de la propagation de la maladie
grâce à la chimiothérapie préventive par l’administration à grande échelle de médicaments
(Mass Drug Administration, MDA). Le traitement consiste en la prise unique de 2
médicaments par l'ensemble de la population à risque chaque année. L'albendazole (ALB), en
association avec de l’ivermectine (IVM) ou du déthylcarbamazine (DEC) sont utilisés. Ces
médicaments ne permettent pas de tuer les parasites adultes, mais éliminent efficacement les
microfilaires de la circulation sanguine et préviennent ainsi la propagation des parasites aux
vecteurs. Pour interrompre le cycle de transmission du parasite, l'administration des
traitements doit donc être menée chaque année pendant au moins 5 ans, avec une couverture
de toute la population à risque [209]. Aucun traitement macrofilaricide (éliminant les parasites
adultes) n'est actuellement disponible pour la MDA.
Lors des débuts du programme GPELF, 81 pays étaient considérés comme
endémiques pour la filariose lymphatique. De 2000 à 2015, plus de 6,2 milliards de
traitements ont été fournis à environ 820 millions d’individus dans 64 pays, ce qui a
considérablement réduit la transmission et a fait baisser le nombre de personnes infestées de
120 millions à 68 millions (OMS).
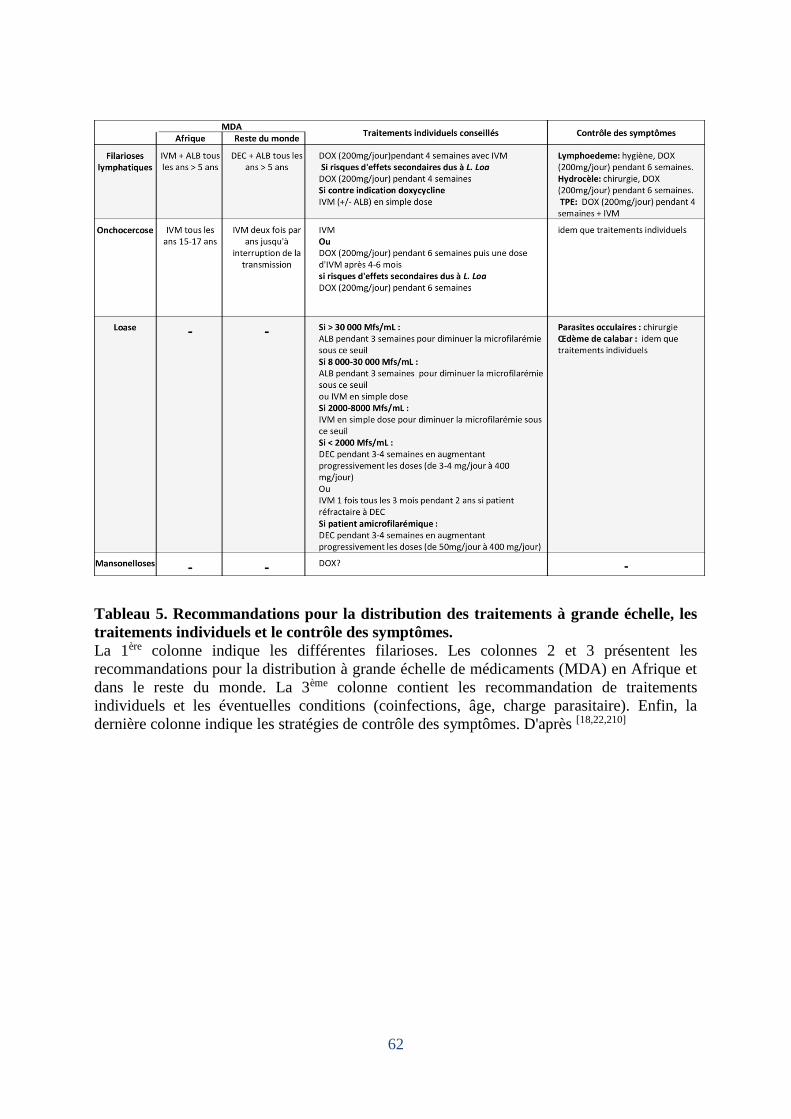
62
Tableau 5. Recommandations pour la distribution des traitements à grande échelle, les
traitements individuels et le contrôle des symptômes.
La 1ère colonne indique les différentes filarioses. Les colonnes 2 et 3 présentent les
recommandations pour la distribution à grande échelle de médicaments (MDA) en Afrique et
dans le reste du monde. La 3ème colonne contient les recommandation de traitements
individuels et les éventuelles conditions (coinfections, âge, charge parasitaire). Enfin, la
dernière colonne indique les stratégies de contrôle des symptômes. D'après [18,22,210]

63
4.1.2 Programmes de lutte contre l'onchocercose
En raison des importantes conséquences de l'onchocercose en Afrique de l'Ouest,
l'OMS a lancé en 1974 le Programme de lutte contre l'onchocercose en Afrique de l'Ouest
(OCP). Pendant des années, le programme a consisté en une lutte anti-vectorielle consistant à
la pulvérisation d’insecticides contre les larves de simulies. En 1987, Merck & Co a annoncé
qu'ils fourniraient gratuitement l’ivermectine pour le traitement de l'onchocercose pendant
aussi longtemps que cela serait nécessaire. A partir de ce moment, la distribution d'IVM à
grande échelle s'est ajoutée à la lutte anti-vectorielle. Lorsque l'OCP a pris fin en 2002, il avait
réussi à éliminer l'onchocercose en tant que problème de santé publique dans 10 des 11 pays
de son aire d’intervention. Basé sur ce succès, le programme Africain pour le contrôle de
l'Onchocercose (APOC) a été lancé entre 1995 et 2015 pour étendre l'OCP. APOC incluait 31
pays d'Afrique sub-saharienne et la stratégie consistait une fois de plus en l'administration à
grande échelle d'ivermectine.
A la clôture du programme en 2015, plus de 114 millions de personnes avaient bénéficié d’un
traitement en Afrique et entre 2005 et 2015 la prévalence de l'infection a chuté de 30% [28].
Trois pays semblent avoir réussi à arrêter la transmission d’Onchocerca volvulus : la
Colombie, l'Equateur et le Mexique [211].
4.1.3 Les traitements individuels
En plus des traitements à grande échelle à visée prophylactique, des traitements
individualisés existent pour les cas avérés d'infection. Ceux-ci utilisent généralement les
mêmes molécules mais avec des posologies pouvant différer (Tableau 5). De plus, des
stratégies visant l'endosymbiote Wolbachia peuvent être mises en place avec des traitements
antibiotiques comme la doxycycline (Dox).
Les stratégies de contrôle et d'élimination des filarioses lymphatiques et de
l'onchocercose sont compliquées dans les régions où une de ces filaires est co-endémique
avec Loa loa. Dans ce cas là, les patients nécessitent un suivi personnalisé. En effet, de graves
effets secondaires ont été observés chez des individus présentant de grandes quantités de
microfilaires de L. loa traités avec l'ivermectine ou la DEC qui peuvent développer des
méningoencephalopathies [33,212].
Seules l'onchocercose et la filariose lymphatique font l'objet de programmes
internationaux avec des distributions de médicaments à grande échelle. Il n'y a actuellement

64
pas de traitement adapté à une application à grande échelle pour traiter les infections à Loa
loa. Boussinesq (2012 [210]) a proposé une stratégie de traitements individuels dépendant de la
charge en microfilaires (Tableau 5). L'objectif de ces traitements est de faire baisser
progressivement la microfilarémie tout en évitant l'apparition d'effets secondaires. Le
problème de ce protocole est qu'il nécessite un suivi régulier voire une hospitalisation, ce qui
est difficilement applicable dans les pays endémiques à L. loa [210].
Dans le cas des mansonelloses, comme les infections sont considérées comme
asymptomatiques, elles sont rarement traitées. Peu d'essais ont été faits pour essayer de les
traiter et aucun programme international n'a été mis en place [213]. Mansonella perstans est
considérée comme une des filaires les plus difficiles à traiter. L'ivermectine, le
déthylcarbamazine et l'albendazole ont peu d'effet sur le parasite [214,215]. M. perstans
hébergeant Wolbachia, quelques essais de traitement par la doxycycline ont été réalisés,
rapportant un effet macrofilaricide de la doxycycline [183,216].
4.1.4 Considérations actuelles
Dans l'ensemble, les programmes de MDA ont conduit à une réduction significative de
la transmission des filaires et de la morbidité causée par les microfilaires. Cependant, les
traitements n'étant pas macrofilaricides (ils ne tuent pas les parasites adultes) et n'entrainant
pas une stérilisation durable des parasites, les traitements doivent être pris tous les ans
pendant des années jusqu'à la mort des parasites (durée de vie de jusqu'à 5 ans pour B. malayi
et jusqu'à 17 ans pour O. volvulus). Des études pilotes sont actuellement en cours pour tester
l'efficacité d'une trithérapie ALB, DEC et IVM en simple dose [217]. En particulier, une étude
pilote sur 12 individus a montré qu'un tel traitement entraîne une absence de microfilarémie
pendant au moins deux ans et ceci suggère une efficacité supérieure aux régimes actuels pour
le traitement des filarioses lymphatiques [218]. On ne sait pas encore si l'efficacité du
traitement est due à une stérilisation des femelles ou à leur élimination. Bien que le nombre de
patients inclus dans cette étude soit faible et malgré le peu de recul sur l 'utilisation de la
trithérapie, ce résultat et les succès des MDA ont conduit l'OMS et une partie de la
communauté scientifique à estimer que la filariose lymphatique pourrait être éliminée comme
problème de santé publique d'ici à 2020 [209]. L'utilisation de cette stratégie pour le traitement
de l'onchocercose semble plus compliquée du fait des effets secondaires oculaires décrits
après les traitements par la DEC [217,219,220].

65
Il est important de noter que l'utilisation massive des trois médicaments actuellement
disponibles (IVM, ALB et DEC) augmente le risque d'émergence et de propagation de la
résistance aux traitements (déjà avéré pour l'ivermectine [221]), ce qui peut constituer une
menace pour les programmes d'éliminations [222]. Comme il n'y a pas de vaccin disponible, il
est important de développer de nouveaux traitements. Parmi les objectifs actuels, l'un a pour
but de développer un traitement oral de courte durée macrofilaricide pouvant traiter à la fois
l'onchocercose et la filariose lymphatique.
4.2 Mode d'action des molécules ciblant les filaires
4.2.1 Ivermectine
L’identification de l’ivermectine comme principale arme thérapeutique pour les
programmes internationaux de lutte contre l’onchocercose justifie pleinement le fait que
Satoshi Omura et William Campbell aient reçu le prix Nobel de médecine 2015.
L'ivermectine est une lactone macrocyclique (Figure 12A) bloquant les canaux post-
synaptiques dépendant du glutamate transportant les ions chlorure (Glutamate-gated Chloride
Channels) [129]. L'intérêt de cette cible est que ces canaux ne sont présents que chez les
nématodes et les arthropodes, limitant ainsi les effets néfastes de l'IVM à ces organismes [223].
Le blocage des canaux inhibe la libération des Mfs par les vers femelles et immobilise les Mfs
circulantes [224]. Les Mfs sont alors drainées vers les ganglions lymphatiques, où les larves
immobilisées sont détruites [225]. L'IVM a donc un effet sur la fertilité des femelles et la
production des Mfs [225] mais cet effet est réversible. Une analyse transcriptomique de
femelles de B. malayi traitées avec de l'ivermectine in vitro a montré que le traitement a aussi
un effet sur de nombreux gènes impliqués dans la méiose et l'embryogénèse ainsi que le
métabolisme de ces parasites [226]. La limitation principale de ce traitement est qu'il est
exclusivement microfilaricide (il élimine les microfilaires et non les adultes) et que la
stérilisation est temporaire [227]. L'objectif du traitement à l'ivermectine est donc d'arrêter la
transmission, avec pour conséquence le fait que les patients doivent être traités durant toute la
vie du parasite. Il est à noter que l'usage à grande échelle depuis plus de 20 ans de
l'ivermectine seule pour traiter l'onchocercose semble avoir entraîné l'émergence de parasites
résistants à ce traitement [228] [222].
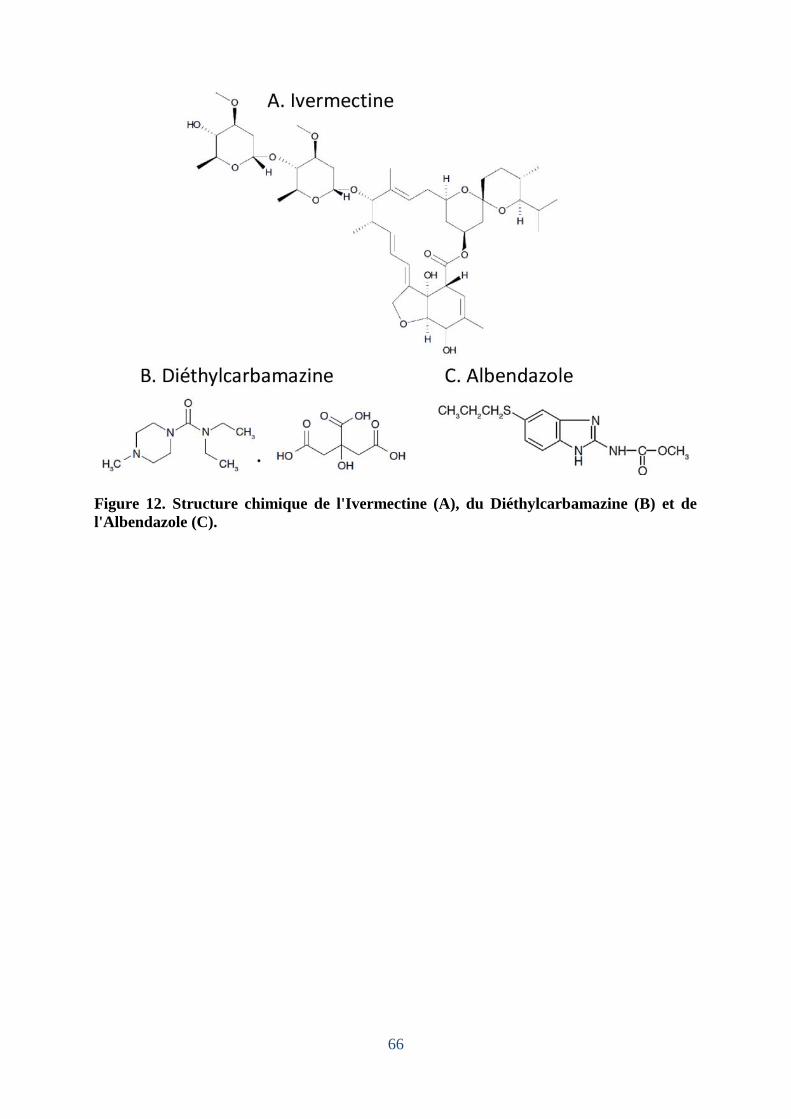
66
Figure 12. Structure chimique de l'Ivermectine (A), du Diéthylcarbamazine (B) et de
l'Albendazole (C).

67
4.2.2 Diéthylcarbamazine
La diéthylcarbamazine (DEC), un dérivé de la pipérazine (Figure 12B), a un effet sur
tous les stades filariens. Cependant, le mécanisme exact d'action de la DEC est mal connu.
L'exposition de microfilaires à de fortes concentrations de DEC in vivo n'est pas suffisant pour
les tuer, suggérant que la DEC n'aurait pas un effet direct majeur sur les filaires. Les
microfilaires disparaissent de la circulation sanguine quelques minutes après le traitement. Le
DEC interfère avec le métabolisme des acides arachidoniques chez les microfilaires et les
cellules endothéliales de l'hôte. Ceci pourrait induire une vasoconstriction et une
augmentation de l'adhésion des microfilaires aux cellules endothéliales [229]. Les microfilaires
immobilisées pourraient ensuite être détruites par les granulocytes [230]. De plus, la DEC peut
orienter la réponse immunitaire avec des effets anti-inflammatoires. Le traitement permet
d'inhiber l'activation du facteur de transcription nucléaire kappa B (NF-kB), un régulateur clé
de gènes pro-inflammatoires tels que TNF-α, IL-1β, ainsi que l'oxyde nitrique synthase
inductible (iNOS) et la cyclooxygénase 2 (COX-2) [231]. La DEC n'est pas recommandée dans
les zones endémiques d'onchocercose où elle peut induire une inflammation locale chez des
sujets présentant des Mfs oculaires [18].
4.2.3 Albendazole
L'albendazole (ALB) est un benzimidazole (Figure 12C) anthelminthique à large
spectre contre les plathelminthes, les cestodes et les nématodes ; il inhibe la polymérisation de
la β-tubuline des vers et la formation des microtubules [232]. De plus, l'ALB bloque
l'absorption de glucose dans les stades larvaires et adultes, entraînant l'épuisement du
glycogène et la réduction des stocks d'adénosine triphosphate (ATP) conduisant à
l'immobilisation et à la mort du parasite [222]. Enfin, chez W. bancrofti l'ALB conduit à une
interruption de la reproduction chez les filaires adultes (d’environ 9 mois) [222].
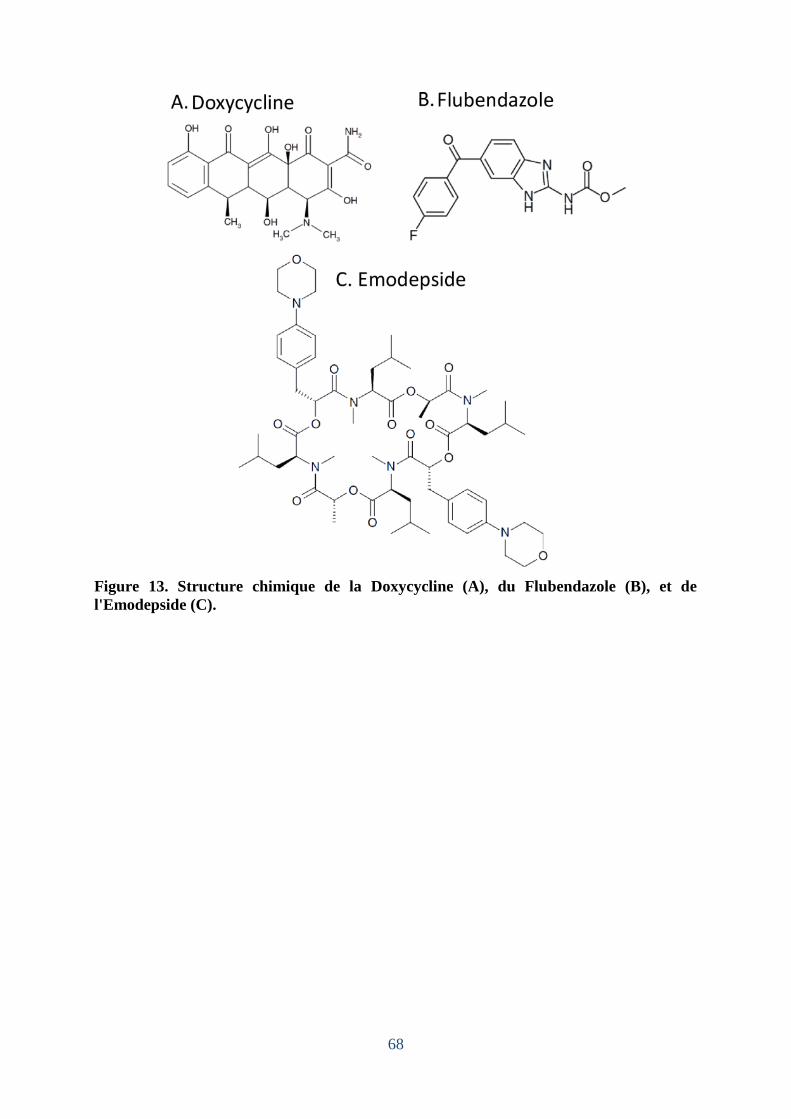
68
Figure 13. Structure chimique de la Doxycycline (A), du Flubendazole (B), et de
l'Emodepside (C).

69
4.2.4 Tétracyclines
Wolbachia est essentielle à la croissance et la survie des filaires humaines (sauf Loa
loa) ce qui en fait une cible thérapeutique. Lors d'essais cliniques [233-236], l'efficacité estimée
de la doxycycline (Dox, Figure 13A) à éliminer Wolbachia chez les femelles adultes d’O.
volvulus était de 91% à 94% en moyenne, quel que soit le schéma thérapeutique utilisé. La
durée de vie des filaires dans lesquelles la population en Wolbachia est fortement diminuée
est réduite de 70% à 80%, passant de 10 ans à 2-3 ans [235]. Un effet macrofilaricide a aussi été
observé chez les patients infectés par Wuchereria bancrofti [237], chez qui un traitement de 4
semaines avec 200mg de Dox permet d'éliminer 80% des adultes. De plus le traitement à la
doxycycline permet de diminuer les symptômes de lymphoedèmes indépendamment de la
présence de parasites adultes vivants en diminuant les infections bactériennes et
l'inflammation [238]. Cependant, son application généralisée dans le cadre des MDA est limitée
par des contre-indications chez les femmes enceintes et les enfants de moins de neuf ans [239]
et par la durée des traitements. La Dox est néanmoins conseillée pour les traitements
individuels (Tableau 5).
4.2.5 Traitements en cours d'évaluation
Flubendazole
Les benzimidazoles (flubendazole, mebendazole, oxfendazole, ciclobendazole,
albendazole, cambendazole, fenbendazole) fonctionnent en interférant dans la formation des
microtubules. Les benzimidazoles peuvent affecter la tubuline des hôtes aussi bien que celle
des parasites et causent une mauvaise séparation chromosomique pendant la mitose [240].
Cependant, les benzimidazoles affectent préférentiellement la tubuline des nématodes plutôt
que celle des mammifères. Ce sont aussi des agents antifongiques ce qui peut être important
lors de l'éléphantiasis qui implique souvent des surinfections fongiques [240].
Le flubendazole (FLBZ, Figure 13B), est actif contre un large spectre de nématodes
gastro-intestinaux chez l'homme et certaines espèces animales. Le FLBZ a aussi un effet
filaricide chez de nombreuses espèces de filaires [240]. Le FLBZ serait le benzimidazole le plus
puissant comme macrofilaricide [241]. Ce composé est déjà approuvé pour une utilisation chez
l’homme pour le traitement de nématodes intestinaux, ce qui peut être un avantage par rapport
aux autres candidats macrofilaricides. L'activité macrofilaricide du FLBZ nécessite cependant
une exposition systémique qui n'est pas atteinte après l'administration orale classique à cause

70
de sa solubilité réduite dans l'eau [242,243]. L'activité in vivo du FLBZ contre plusieurs espèces
filariennes a été rapportée après son administration intramusculaire dans des essais chez
l'animal et chez l'homme [244]. Cependant, des effets secondaires ont été signalés après
l'administration du FLBZ, majoritairement une inflammation du site d'injection et de la fièvre
[244,245]). De plus, le FLBZ a un effet perturbateur sur la mitose [246]. Des recherches sont en
cours pour améliorer la formulation du traitement et la biodisponibilité du traitement tout en
diminuant les effets secondaires [243,246] mais à l'heure actuelle il semble peu probable que le
FLBZ obtienne une autorisation de mise sur le marché pour les filarioses.
Emodepside
L'émodepside est un cyclooctadepsipeptide (Figure 13C) qui agit sur les jonctions
neuromusculaires. L'action principale de l'émodepside passe par l'activation d'un canal
potassique chez les filaires (SLO-1) entrainant la paralysie des parasites [247]. L'émodepside
est entrée en étude de phase I sur volontaires sain en 2016 pour le traitement des patients
atteints d'onchocercose (DNDi : https://www.dndi.org/diseases-projects/portfolio/emodepside/).
Tylosine
Le consortium Anti-Wolbachia (A.WOL), a pour objectif de mettre au point un
traitement ciblant Wolbachia. autre que les tétracyclines. Un dérivé de la tylosine, un
antibiotique vétérinaire ciblant Wolbachia est actuellement en développement préclinique
pour le traitement des filarioses. Il induirait un effet anti-Wolbachia plus robuste que la
doxycycline dans plusieurs modèles in vivo et serait efficace plus rapidement (http://dndi.org).
Une étude de phase I sera réalisée en 2017 pour évaluer la sécurité, la tolérance et la
pharmacocinétique du médicament.

71
5 Cinétique de la réponse immunitaire chez les souris infestées par L. sigmodontis
Réponses immunitaires associées aux filarioses
Lors d'une infection filarienne, une réponse immunitaire se met en place. Grace à de
nombreuses molécules sécrétées et à Wolbachia, les filaires sont capables d'orienter la
réponse immunitaire dès l'entrée dans l'organisme pour favoriser leur migration et leur survie.
Ainsi, les filaires sont capables de survivre de plusieurs mois à plusieurs années dans leur hôte
[248].
Un réseau complexe de cellules et protéines est impliqué dans la résistance et la
susceptibilité aux filaires. La réponse aux filaires est généralement définie comme une
réponse Th2 modifiée. Cette réponse implique la production des cytokines IL-4, IL-5, IL-9,
IL-10 et IL-13, la production des d'anticorps d'isotypes IgG1, IgG4 et IgE (chez l'homme) et
IgG1 et IgE (chez la souris) [248] et des quantités accrues d'éosinophiles et de macrophages
alternativement activés ainsi que des cellules lymphoïdes innées (ILC) [249]. De plus, les
cellules dendritiques ont des capacités de présentation de l'antigène réduites et les
lymphocytes prennent un phénotype régulateur anti-inflammatoire. Les nombreux
mécanismes régulateurs mis en place, s'ils permettent une survie accrue des parasites,
participent à la réparation des tissus et diminuent les risques de pathologies dues à une
réponse inflammatoire trop forte [250].
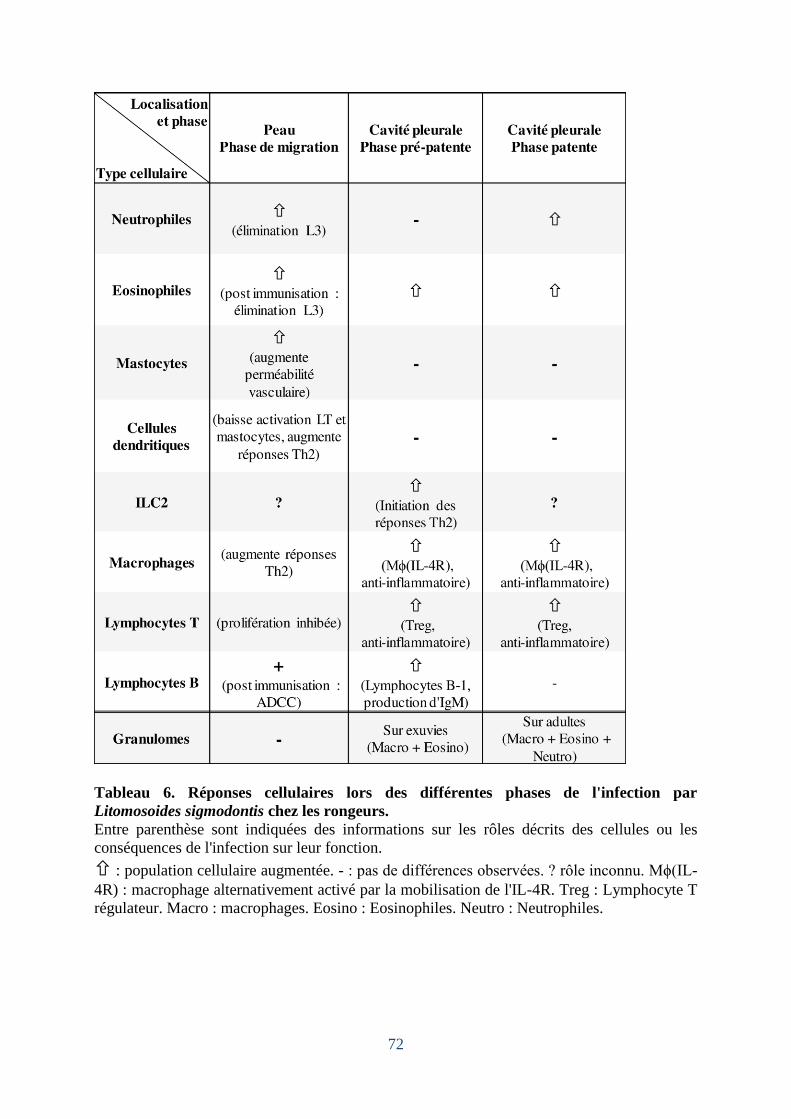
72
Tableau 6. Réponses cellulaires lors des différentes phases de l'infection par
Litomosoides sigmodontis chez les rongeurs.
Entre parenthèse sont indiquées des informations sur les rôles décrits des cellules ou les
conséquences de l'infection sur leur fonction.
: population cellulaire augmentée. - : pas de différences observées. ? rôle inconnu. Mϕ(IL-
4R) : macrophage alternativement activé par la mobilisation de l'IL-4R. Treg : Lymphocyte T
régulateur. Macro : macrophages. Eosino : Eosinophiles. Neutro : Neutrophiles.

73
5.1 Réponse immunitaire au site d'inoculation : apport des différents modèles
Le premier contact entre les filaires et leur hôte a lieu lors de l'entrée des L3 dans la
peau. Cette étape est commune à toutes les filaires et il est probable que la réponse
immunitaire cutanée initiale montre des similarités chez toutes les espèces. Dans les modèles
étudiés chez les rongeurs (B. malayi, L. sigmodontis, Acanthocheilonema viteae, ou
Monanema martini), la réponse inflammatoire détruit une grande quantité de L3 en quelques
heures dans le tissu cutané [10,55]. Cela implique une induction précoce des défenses de l'hôte,
avec le recrutement et l'activation de nombreuses cellules, comme les cellules présentatrices
d'antigène (CPA), les granulocytes et les mastocytes [251,252].
Neutrophiles
Dans la zone inflammatoire du site d'inoculation, de nombreux neutrophiles sont
recrutés [253-258]. Le recrutement des neutrophiles dans la peau est dépendant du récepteur
NOD2 (Nucleotide-binding oligomerization domain-containing protein 2) qui reconnait le
Muramyldipeptide (MDP) de la paroi de certaines bactéries, dont Wolbachia [258]. Les
neutrophiles ont un rôle important dans l'élimination des L3 dans la peau (Tableau 6). En
effet, dans des souris présentant un défaut de maturation des cellules B et de la production
d'anticorps (la souche µMT), la grande majorité des L3 sont détruites par une réponse
dominée par les neutrophiles [254]. La déplétion des neutrophiles avec des anticorps
monoclonaux avant l'inoculation des L3 conduit donc a un rendement parasitaire plus élevé
[257,258]. De plus, l'infection de souris présentant un nombre de neutrophiles constitutivement
plus élevé dans la peau (Cxcr4+/1013) conduit à une baisse importante du succès filarien. Cet
effet protecteur est perdu en évitant la phase de migration sous-cutanée des L3 par une
inoculation intraveineuse des larves [257].
Eosinophiles
L'immunisation de souris avec des L3 irradiées induit une réduction du rendement
filarien dans les premiers jours après l'infection avec des L3 saines (Tableau 6 et [259,260]).
Cette immunité protectrice est médiée dans la peau par les éosinophiles et les anticorps dans
les deux jours suivant l'infection [191,253,261]. Par conséquent, l'efficacité de cette méthode
d'immunisation est diminuée à la fois chez les souris μMT, dans lesquelles des éosinophiles
sont recrutés sur le site d'une infection mais ne libèrent pas leurs granules [254], et chez des
souris déficientes en éosinophiles ou en IgE [262]. Le mécanisme effecteur précis déployé par

74
les éosinophiles lors du ciblage des filaires n'est pas entièrement élucidé, l'élimination de
l'EPO ou du MBP entraînent une augmentation du rendement filarien de L. sigmodontis chez
la souris, ce qui suggère un rôle de ces protéines granulaires dans la défense contre les
parasites. On ne sait pas si cela est principalement dû à la réduction de la capacité effectrice
des éosinophiles à dégranuler ou à un mécanisme indirect impliquant des niveaux altérés d'IL-
10, d'IL-5 et / ou d'IL-4 [263].
Cellules dendritiques
Lorsque des L3 de L. sigmodontis sont injectées dans la peau des souris, la chimiokine
CCL17 est libérée par les cellules dendritiques (dendritic cell, DC) via un mécanisme
dépendant de Wolbachia et du TLR2 [264]. CCL17 limite la libération d'histamine par les
mastocytes, molécule qui augmente la perméabilité vasculaire et facilite l'échappement des
larves vers le système lymphatique. De plus, les L3 de B. malayi agissent sur les cellules de
Langerhans (des DC du tissus cutané) humaines conduisant à une diminution de la
prolifération des lymphocytes T CD4+ [265,266]. La stimulation in vitro de macrophages et de
cellules dendritiques par des extraits solubles de B. malayi induit une réponse majoritairement
Th2 et une baisse de leur production de cytokines inflammatoires comme IL-1β, IL-6, IL-12,
et TNF-α [267-269]. A l'inverse, la stimulation de ces cellules avec des composés de Wolbachia
tels que la protéine de surface (WSP) [270] ou des composés lipoprotéiniques diacétylés comme
WoLP (Wolbachia Lipopeptide) sont activés via un mécanisme dépendant de TLR-2/TLR-6
et dans une moindre mesure TLR-2/TLR-1 2006 [268] et ont un phénotype pro-inflammatoire.
5.2 Migration des larves de L. sigmodontis vers la cavité pleurale
Les L3 présentes dans la cavité pleurale sont celles qui ont quitté le site d'inoculation
en migrant via le système lymphatique en passant par les vaisseaux lymphatiques afférents et
les ganglions lymphatiques drainant [10,11] (Kilarski non publié :
http://www.wired.com/wiredscience/2011/08/nikon-small-world-vote-2011/?pid=1861). Des dosages
réalisés sur des surnageants de culture de cellules isolées de ces ganglions 60 heures après
l'inoculation des L3 ont montré une augmentation de la production d'IL-5 et d'IL-6après
restimulation par des extraits bruts filariens. Il semblerait que l'augmentation de l'IL-5
pourrait servir de signal à la filaire pour réaliser la suite de son développement [195,201]. Dès 6
heures après infection, un recrutement de neutrophiles est observé dans le tissu pulmonaire
[271].

75
5.3 Réponses immunitaires dans la cavité pleurale lors de la phase pré-patente
Les L3 s'installent dans la cavité pleurale et y réalisent leur croissance et leurs mues
jusqu'au stade adulte. Si le rendement filarien final est établi 8 jours post-infection, des L3 ont
déjà été observées dans la cavité pleurale dès 2 jours après infection [204]. Rapidement, une
réaction inflammatoire de type Th2 se met en place. De nombreuses chimiokines telles que
IL-4, IL-5, IL-10, IL-13, CCL2 (MCP-1), CCL11 (Eotaxin-1), CCL5 (RANTES) et CXCL12
sont retrouvées dans la cavité pleurale [196,272,273]. Un fort recrutement de cellules est observé
dès J10 [195,274] avec majoritairement des macrophages et des éosinophiles et de lymphocytes
B [195,273,275] (Tableau 6) et un pic trente jours après inoculation des L3. Les neutrophiles sont
très rares avant la phase patente [196].
Initiation de la réponse Th2 dans la cavité pleurale
Le mésothélium pleural réagit rapidement aux infections et peut participer au
recrutement cellulaire dans la cavité pleurale en produisant des cytokines [276]. Lors de
l'infection par L. sigmodontis, il peut produire CXCL12 qui est un chimioattractant des
neutrophiles, des monocytes, des lymphocytes T et des éosinophiles [196] ainsi que CCL11
(Eotaxine-1) et CCL5 (RANTES) qui sont des molécules extrêmement chimiotactiques pour
les éosinophiles [272].
Dès 5 jours après l'infection par L. sigmodontis, le nombre d'ILC2 (cellules
lymphoïdes innées) augmente dans la cavité pleurale, pour atteindre un maximum juste avant
la phase patente autour de 40 jours post-infection [273]. Les ILC2, productrices d'IL-13, d'IL-5
et d'IL-4 [277] pourraient être les initiateurs de la réponse Th2 dans la cavité pleurale.
7 jours après l'infection, une prolifération de Lymphocytes T régulateurs (Treg)
CD4+Foxp3+ a été décrite [278]. Ils sont capables de produire de l'IL-10 [103,279] et la molécule
immunosuppressive CTLA-4 (Cytotoxic T-Lymphocyte Antigen 4) [280].
De plus l'inflammation de la cavité pleurale induite par L. sigmodontis provoque la
formation sur le médiastin et le péricarde de structures lymphoïdes, les FALCs, composées
majoritairement de lymphocytes B et d'ILC2 (FALC : Fat-associated lymphoid cluster /
structures lymphoïdes associés aux graisses) [281]. Les cellules stromales des FALCs
produisent de l'IL-33 qui active les ILC2. Celles-ci produisent ensuite de l'IL-5 induisant
l'activation des cellules B-1 pleurales et la sécrétion locale d'IgM polyréactifs [281]. Les souris

76
BALB.Xid, qui n'ont pas de lymphocytes B-1, sont plus susceptibles que les BALB/c à une
infection par L. sigmodontis [282].
11 jours après infection, une prolifération et une activation des macrophages pleuraux
par le récepteur à l'IL-4 (IL4Rα, qui fixe aussi l'IL-13) sont observées [281]. L'activation du
récepteurs à l'IL-33 (ST2) participe également à l'activation et à la prolifération de ces
macrophages [281]. L'augmentation du nombre de macrophages dans la cavité pleurale ne
provient pas d'un recrutement de monocytes sanguins mais d'une prolifération des
macrophages résidents [283]. Ces macrophages produisent de grandes quantités de molécules
anti-inflammatoires comme le TGF-β, PGE2, l'antagoniste du récepteur à l'IL-1 (IL-1RA) et
l'arginase [277].
L'action combinée des macrophages et des Treg induit une anergie des lymphocytes T
effecteurs (Teff) qui permet une survie accrue des parasites [268,279,284]. Les Teff expriment des
taux élevés d'IL10, de GITR (glucocorticoid induced TNFR-related protein), CTLA-4 et PD-1
(Programmed Death 1) [279,280,285].
Spécificité des souches murines : comparaison des souches BALB/c et C57BL/6
Le rendement filarien reste stable chez les souris BALB/c quasiment jusqu'au début de
la phase patente vers 50-55 jours post infection [196], indiquant que les mécanismes
d'immunosuppression induits par la présence des filaires sont efficaces. Il est aussi probable
que cette immunosuppression joue un rôle important dans la limitation des dommages
tissulaires qui résulteraient d'une trop forte réaction inflammatoire [277]. Chez les souris
C57BL/6, les parasites se développent jusqu'à effectuer leur dernière mue puis les jeunes
adultes meurent vers 40 jours, bien avant d'atteindre la phase patente [195,203].
La réponse immunitaire est différente dans les deux souches de souris. La cinétique du
recrutement cellulaire dans la cavité pleurale est lente mais de forte amplitude chez les souris
C57BL/6 alors qu’elle est rapide et d'amplitude intermédiaire chez les BALB/c [195]. A 30
jours p.i., aucune différence dans les proportions leucocytaires de la cavité pleurale n'est
observée entre les deux souches de souris mais le recrutement cellulaire pleural est plus
important chez les souris C57BL/6. Ceci est associé à une sécrétion plus élevée de
chimiokines inflammatoires comme CCL2, CCL3 et CCL11 et CXCL12 dans le liquide
pleural des souris C57BL/6 [195,196]. Les souris C57BL/6 présentant la mutation Rag2IL-2Rγ-/-,
déficientes en cellules T, B et NK, ont un phénotype semblable à celui des souris BALB/c,

77
atteignant la phase de production de microfilaires, indiquant un rôle important de ces cellules
dans la résistance à l'infection [286].
Chez les souris BALB/c, des granulomes réactionnels sont observés dans la cavité
pleurale à différent temps au cours de l’infection filarienne : entre 8 et 10 jours post-infection
ils sont essentiellement constitués de macrophages et d'éosinophiles; il n'y a que de rares
neutrophiles. Ces granulomes ne contiennent que des exuvies de L3 [202] et il a été suggéré que
ces exuvies pourraient jouer un rôle de leurre contre la réponse de l’hôte. D'autres granulomes
sont retrouvés entre J27 et J39 contenant de nouveau des exuvies, cette fois-ci post mue en
jeunes adultes, et ils sont composés majoritairement d'éosinophiles (70%) mais pas de
neutrophiles [202]. Chez les souris C57BL/6, des granulomes sont aussi retrouvés entre 27 et 39
jours après infection mais autour de jeunes vers adultes. Ils sont aussi composés
majoritairement d'éosinophiles [202].
5.4 Réponse dans la cavité pleurale au cours de la phase patente
Les souris BALB/c développent la phase patente avec production de microfilaires dans
la cavité pleurale environ 55 jours après infection par L. sigmodontis. Seule une partie des
souris infestées (inoculées avec un nombre connu de filaires ou par contact avec des acariens
infestés) devient patente (environ 50%). Il est intéressant de noter que dans les zones
endémiques des filarioses humaines, une fraction des patients concernés par la filariose
lymphatique, la loase et la mansonellose (à M. perstans) est aussi amicrofilarémique
(respectivement jusqu'à 40%, jusqu'à 70% et proportion inconnue) [18,22,62].
Des études chez des individus présentant des infections asymptomatiques par W.
bancrofti ont révélé que les individus Mfpos présentent des réponses immunitaires plus faibles
que les individus Mfneg [287]. Les infections chez les souris BALB/c avec L. sigmodontis
produisent également de animaux Mfpos et Mfneg et les réponses immunitaires entre ces
groupes sont différentes [276] : les souris Mfpos présentent des concentrations d'IL-5 et d'IL13
dans la cavité pleurale plus élevées [288]. La mise en place de la phase patente est déterminée
notamment par des réponses dépendant du TLR-4. En effet, l'infection de souris déficientes en
TLR-4 conduit à un plus grand pourcentage de souris microfilarémiques, sans différences
dans le nombre de microfilaires circulantes [288]. De même, L. sigmodontis dans les souris
C3H/HeJ naturellement déficientes en TLR-4 produit des microfilaires, comparé aux souris

78
C3H/HeN qui ne développent pas de microfilarémie malgré la présence d'adultes dans la
cavité [289].
Deux pics de libération d'histamine dans le sang sont observés lors de l'infection par L.
sigmodontis suivant l'injection sous cutanée de L3 dans des souris BALB/c : la première a lieu
30 minutes après infection, et coïncide avec l'échappement des filaires dans les lymphatiques;
la seconde a lieu après environ 55 jours [290], correspondant au début de la phase patente. La
libération d'histamine favorise la survie des parasites en diminuant les réponses éosinophiles
[290], mais on peut aussi imaginer que l'effet vasodilatateur de l'histamine aide à la sortie des
microfilaires de la cavité pleurale hors de la cavité pleurale par un mécanisme similaire à celui
observé pour les L3 dans la peau [264].
Les éosinophiles, via l'IL5 ont un effet protecteur pour l'hôte (Tableau 6). Les souris
BALB/c déficientes en IL-5 qui ne présentent quasiment aucun éosinophiles, ont un
rendement parasitaire ainsi qu'une microfilarémie plus élevées que les souris sauvages [255,291].
A l'inverse, le rendement parasitaire à J60 dans des souris éosinophiliques (CBA/Ca
surexprimant l'IL-5) est beaucoup plus faible que chez des souris contrôles [292]. L'action des
éosinophiles peut être directe par la sécrétion des protéines de leurs granules dont la
Peroxydase des Eosinophiles (EPO) et la Protéine Basique Majeure (MBP) ou être indirecte
en influençant la réponse cytokinique [263,292,293].
A partir de la phase patente, des neutrophiles sont observés dans la cavité pleurale [202]
et semblent liés à la formation d'un autre type de granulome. Les granulomes observés à partir
de J60 contiennent des filaires adultes mortes et sont composés de macrophages, de
neutrophiles et d'éosinophiles (Tableau 6 et [202,294]). L'arrivée des neutrophiles dans la cavité
pleurale et leur participation à la formation des granulomes semblent liées à leur capacité à
exprimer l'intégrine CD11b, indispensable au trafic des neutrophiles [202]. L'accumulation des
éosinophiles, mais aussi des neutrophiles dans la cavité pleurale est aussi dépendante de l'IL-5
[294]. Enfin la bactérie Wolbachia, relarguée lors de la mort des parasites pourrait aussi être
liée au recrutement et à l'activation des neutrophiles dans la cavité pleurale [17].

79
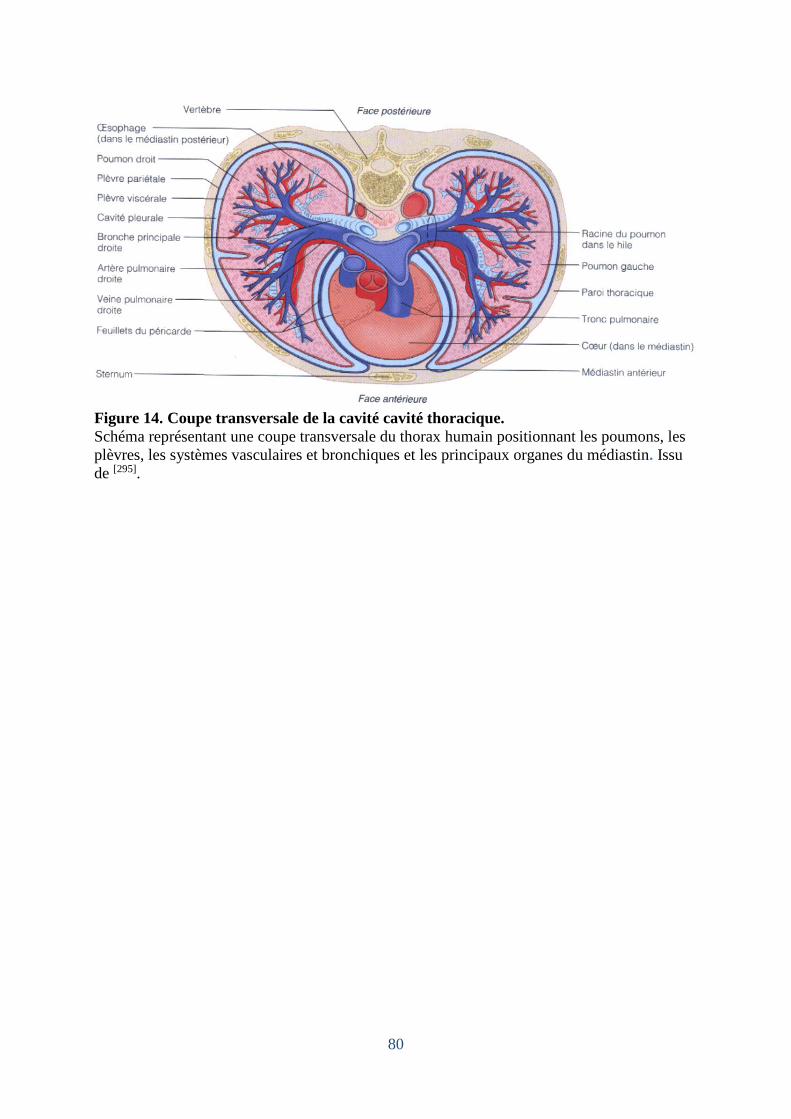
80
Figure 14. Coupe transversale de la cavité cavité thoracique.
Schéma représentant une coupe transversale du thorax humain positionnant les poumons, les
plèvres, les systèmes vasculaires et bronchiques et les principaux organes du médiastin. Issu
de [295].

81
6. Rappels anatomiques/histologiques sur les poumons, le cœur et les systèmes
vasculaires
La cavité thoracique contient le poumon et les plèvres et, dans le médiastin, d’autres
structures dont le cœur (Figure 14).
6.1 Plèvre et mésothélium
La plèvre est une membrane séreuse, fine, brillante, glissante, qui tapisse la paroi
thoracique et le médiastin (plèvre pariétale) puis, à partir du médiastin, elle se réfléchit sur
elle-même pour rejoindre le poumon (plèvre viscérale ou pulmonaire) [296]. La plèvre viscérale
recouvre les poumons et s’insinue dans leurs scissures.
L’espace virtuel qui existe entre ces deux feuillets est appelé « cavité pleurale », il
contient un film liquidien de moins de 20µm d'épaisseur, qui permet un glissement sans
friction entre eux [297]. Il joue un rôle important dans la respiration ; en effet la pression
intrapleurale est négative, ce qui permet l'expansion pulmonaire et maintient le couplage
mécanique entre le poumon et la paroi thoracique pendant tout le cycle respiratoire.
Ces plèvres sont bordées par une monocouche de cellules mésothéliales plates
(épithélium pavimenteux) constituant le mésothélium [298]. Il repose sur une mince membrane
basale supportée par des tissus conjonctifs, des fibroblastes et des vaisseaux sanguins et
lymphatiques [299]. Les cellules mésothéliales sont reliées entre elles par des jonctions serrées,
des jonctions communicantes et des desmosomes [297]. Les cellules mésothéliales fournissent
une barrière de protection continue contre les dégâts mécaniques et synthétisent les
glycosaminoglycans et les phospholipides (semblables à celles formant le surfactant
pulmonaires) nécessaires pour lubrifier la surface pleurale.
Le taux de renouvellement des cellules mésothéliales est très faible (moins de 0,5%)
[300]. Cependant, la prolifération cellulaire augmente après une lésion mésothéliale ou une
exposition à des agents inflammatoires, ce qui suggère un rôle des cellules mésothéliales dans
la cicatrisation des plaies et la mise en place de la fibrose pleurale [297].
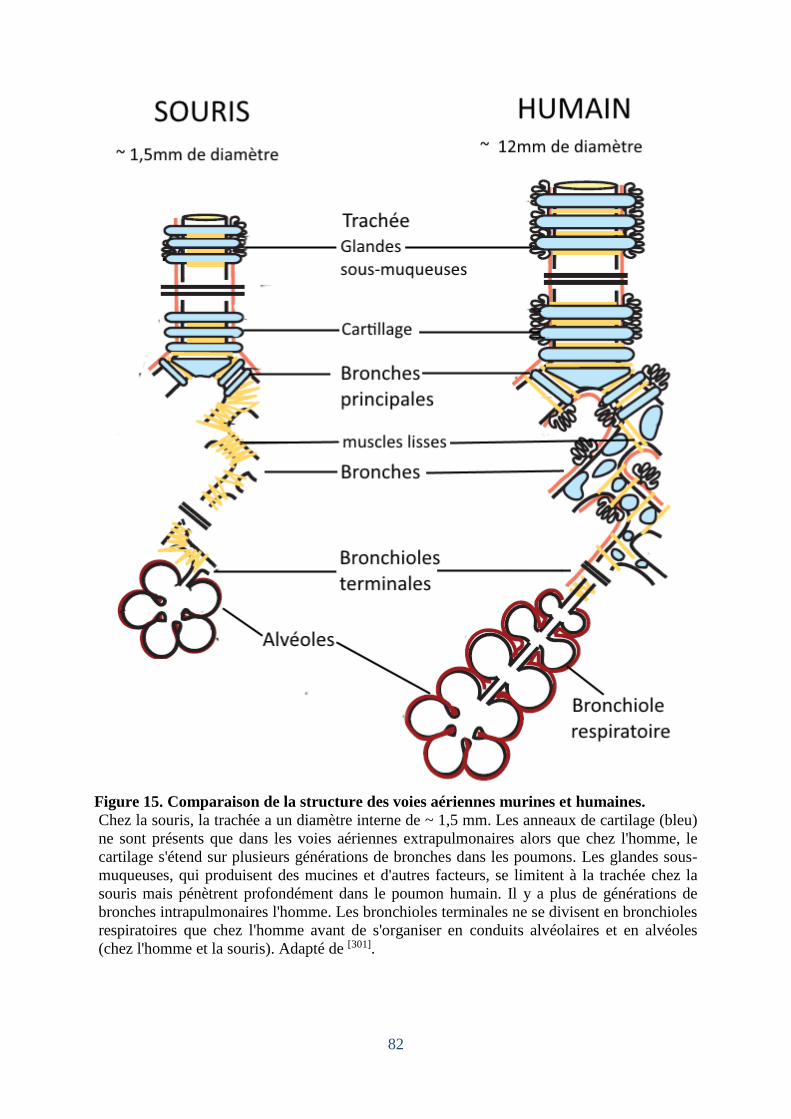
82
Figure 15. Comparaison de la structure des voies aériennes murines et humaines.
Chez la souris, la trachée a un diamètre interne de ~ 1,5 mm. Les anneaux de cartilage (bleu)
ne sont présents que dans les voies aériennes extrapulmonaires alors que chez l'homme, le
cartilage s'étend sur plusieurs générations de bronches dans les poumons. Les glandes sous-
muqueuses, qui produisent des mucines et d'autres facteurs, se limitent à la trachée chez la
souris mais pénètrent profondément dans le poumon humain. Il y a plus de générations de
bronches intrapulmonaires l'homme. Les bronchioles terminales ne se divisent en bronchioles
respiratoires que chez l'homme avant de s'organiser en conduits alvéolaires et en alvéoles
(chez l'homme et la souris). Adapté de [301].

83
6.2 Le poumon, les bronches, les bronchioles et les cellules alvéolaires de type I et
II
La trachée va du larynx vers la cage thoracique où elle se ramifie en deux bronches
principales pénétrant dans chacun des deux poumons (droit et gauche) au niveau du hile. Les
deux bronches principales se divisent et vont subdiviser les poumons droit et gauche en
différents lobes [302]. Suivant les espèces, le nombre de lobes de chaque poumon diffère. Chez
l'homme le poumon droit présente 3 lobes et le gauche 2; chez la souris le poumon droit
présente 4 lobes et le gauche ne présente pas de lobation [303]. Après un certain nombre de
division successives (17-21 générations chez l'humain, 13–17 chez la souris) les bronches
cèdent la place aux bronchioles, qui sont plus petites. Les bronches ont une paroi épaisse
contenant des amas cartilagineux ainsi qu'une couche de cellules musculaires lisses. Les
bronches humaines présentent aussi des glandes séro-muqueuses sous-épithéliales absentes
chez la souris. Les bronchioles ont une paroi plus fine sans cartilage (Figure 15).
Les bronches et les bronchioles présentent un épithélium ciliaire, plus épais dans les
bronches, dont les cils battent de manière coordonnée pour pousser une couche de mucus vers
le pharynx, où elle est avalée ou éliminée par la toux. Ce système joue un rôle important dans
l'élimination de particules inhalées et de microorganismes qui se déposent sur les muqueuses
des voies aériennes [302]. La composition de cet épithélium varie suivant la taille des
bronches/bronchioles et présente des différences suivant les espèces. Globalement, les mêmes
types cellulaires forment l'épithélium des voies respiratoires, mais la proportion relative de
chaque type de cellule varie. Quatre types cellulaires composent l'épithélium des voies
aériennes : 1) les cellules ciliées sont prédominantes ; 2) les cellules caliciformes qui
produisent le mucus ; elles sont abondantes dans les voies aériennes inférieures chez l'humain
mais très rares chez la souris en condition homéostatique ; 3) les cellules Club (ou Clara) sont
des cellules sécrétoires qui produisent des composés antimicrobiens qui contribuent à la
défense de la muqueuse des voies respiratoires ; 4) les cellules basales ont la capacité de
proliférer en réponse à des dommages et de se différencier en d'autres types de cellules
épithéliales [304].

84
Les bronchioles terminales se divisent en bronchioles respiratoires (chez l'humain et
non chez la souris- Figure 15) puis en conduits alvéolaires et enfin en alvéoles. Les poumons
d'un homme de 70kg ont 300 millions d'alvéoles représentant une surface d'environ 143 m2
pour un volume pulmonaire moyen de 4,3L [296,302]. 200 ml de sang se trouvent dans le réseau
capillaire à tout moment. On estime qu'environ 90% de la surface alvéolaire est recouverte par
le réseau des capillaires pulmonaires, ce qui correspond une interface air-sang d'environ 126
m2 [302]. Chez la souris, il y a environ 2 millions d'alvéoles avec une surface alvéolaire de 82,2
cm2 pour un volume pulmonaire moyen de 0,3 mL [305].
Il existe deux types de cellules de revêtement
alvéolaire (Figure 16). Les cellules alvéolaires de type I,
ou pneumocytes de type I, ont un noyau plat central et un
grand cytoplasme qui atteint 50 μm de diamètre pour une
épaisseur de seulement 0,2-0,5 μm permettant les
échanges gazeux. Ces cellules constituent seulement 40%
des cellules alvéolaires mais couvrent 90% de la surface
alvéolaire. Les cellules de type II, ou pneumocytes de type
II, sont des cellules cuboïdales d'un diamètre d'environ 15
μm ; elles constituent 60% des cellules des alvéoles, mais
couvrent seulement 5% de la surface alvéolaire ; elles
deviennent hyperplasiques lors de dégâts alvéolaires [296].
La surface alvéolaire est recouverte d'une couche de
surfactant (composé majoritairement de phospholipides),
qui est synthétisé et libéré par la cellule alvéolaire de type
II.
Figure 16. Structure des alvéoles.
L'épithélium alvéolaire est composé de pneumocytes de type I présentant un noyau plat
central et un grand cytoplasme et de pneumocytes de type II, cellules cuboïdales produisant le
surfactant recouvrant la surface alvéolaire. La barrière alvéole-capillaire est très fine (environ
0,6µm) et permet les échanges gazeux. Les macrophages alvéolaires sont le type majoritaire
de l'espace alvéolaire à l'homéostasie.

85
6.3 Vaisseaux sanguins et lymphatiques pulmonaires
6.3.1 Vascularisation sanguine pulmonaire
La circulation sanguine pulmonaire est constituée de deux systèmes circulatoires
d'origine différente : la circulation pulmonaire et la circulation bronchique, les deux systèmes
étant communicants.
La circulation pulmonaire
Les veines caves supérieure et inférieure collectent le sang veineux au niveau du
ventricule droit qui rejoint l’artère pulmonaire qui se divise en artère pulmonaire gauche et
droite. Les artères pulmonaires apportent aux poumons plus de 90% de leur irrigation sous la
forme de sang de type veineux, peu oxygéné, perfusé sous basse pression. Les artères
pulmonaires suivent étroitement l'arbre bronchique et émettent des ramifications s'achevant
dans les sacs alvéolaires où elles forment de vastes réseaux capillaires enveloppant les
alvéoles, siège principal de l'hématose (oxygénation de sang) (Figure 17- points 1 à 5).
La paroi des capillaires est seulement constituée de quelques cellules endothéliales.
Les lames basales des cellules alvéolaires et de l'endothélium sont confondues, conduisant à
une barrière alvéole-capillaire d'une épaisseur moyenne de 0,6µm [296]. Le diamètre des
capillaires est compris entre 5 et 8 µm [306-308]. Les réseaux capillaires alvéolaires se
condensent ensuite en veines qui vont confluer vers les veines pulmonaires gauche et droite
qui se rejoignent dans l’oreillette gauche.
Les artères pulmonaires sont d'abord des artères élastiques avec une paroi relativement
mince et une adventice épaisse, puis des artères musculaires avec deux limitantes élastiques,
interne et externe. Ensuite, la couche musculaire s'amincit puis disparaît dans les vaisseaux
précapillaires de 30 à 40µm de diamètre. Il y a peu de différences entre artères et veines dans
la circulation pulmonaire, en raison de la minceur de la paroi artérielle, correspondant à la
faible pression sanguine. Les veines, cependant n'ont qu'une limitante élastique interne et elles
ne s'entourent de fibres musculaires qu'au dessus d'un diamètre de 100µm.
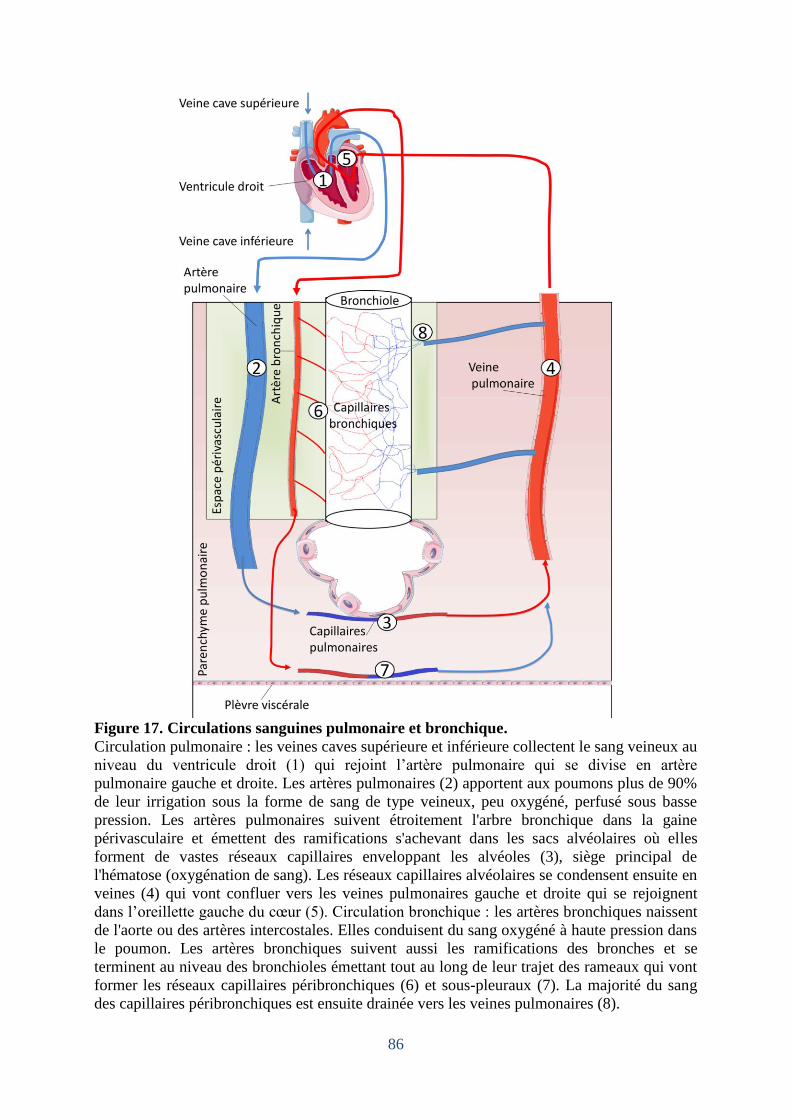
86
Figure 17. Circulations sanguines pulmonaire et bronchique.
Circulation pulmonaire : les veines caves supérieure et inférieure collectent le sang veineux au
niveau du ventricule droit (1) qui rejoint l’artère pulmonaire qui se divise en artère
pulmonaire gauche et droite. Les artères pulmonaires (2) apportent aux poumons plus de 90%
de leur irrigation sous la forme de sang de type veineux, peu oxygéné, perfusé sous basse
pression. Les artères pulmonaires suivent étroitement l'arbre bronchique dans la gaine
périvasculaire et émettent des ramifications s'achevant dans les sacs alvéolaires où elles
forment de vastes réseaux capillaires enveloppant les alvéoles (3), siège principal de
l'hématose (oxygénation de sang). Les réseaux capillaires alvéolaires se condensent ensuite en
veines (4) qui vont confluer vers les veines pulmonaires gauche et droite qui se rejoignent
dans l’oreillette gauche du cœur (5). Circulation bronchique : les artères bronchiques naissent
de l'aorte ou des artères intercostales. Elles conduisent du sang oxygéné à haute pression dans
le poumon. Les artères bronchiques suivent aussi les ramifications des bronches et se
terminent au niveau des bronchioles émettant tout au long de leur trajet des rameaux qui vont
former les réseaux capillaires péribronchiques (6) et sous-pleuraux (7). La majorité du sang
des capillaires péribronchiques est ensuite drainée vers les veines pulmonaires (8).

87
La circulation bronchique
Les artères bronchiques naissent de l'aorte ou des artères intercostales. Elles
conduisent du sang oxygéné à haute pression dans le poumon. C'est la circulation nutritive de
l'arbre bronchique (Figure 17- point 6-8).
Les artères bronchiques suivent aussi les ramifications des bronches et se terminent au
niveau des bronchioles émettant tout au long de leur trajet des rameaux qui vont former les
réseaux capillaires péribronchiques et sous-pleuraux. A leur extrémité, ces capillaires
communiquent avec le réseau capillaire alvéolaire, assurant ainsi une connexion (anastomose)
entre les réseaux artériels bronchique et pulmonaire. La majorité du sang des capillaires
péribronchiques est ensuite drainée vers les veines pulmonaires. Des veines bronchiques
autonomes existent seulement au niveau des plus grosses bronches ; les veines pulmonaires
collectent donc la majeure partie du sang des plexus péribronchiques [306].
6.3.2 Les lymphatiques des poumons et de la plèvre : le système
pleuropulmonaire
Les lymphatiques du thorax drainent les liquides du tissu interstitiel pulmonaire et de
la cavité pleurale. Pourvus de valves empêchant leur reflux, ils les conduisent vers la
circulation sanguine, soit directement dans les confluents veineux sous-claviers, soit via le
canal thoracique. Ils forment des courants ascendants pariétaux et viscéraux anastomosés
entre eux. Les courants pariétaux drainent la lymphe de la paroi thoracique et de la cavité
pleurale. Les courants viscéraux drainent celle des poumons et des organes présents dans le
médiastin. Le canal thoracique monte au niveau du médiastin postérieur et s’abouche en
général dans le confluent veineux sous-clavier gauche. Il reçoit une partie de la lymphe du
thorax et draine celle des membres inférieurs et du tronc. La moitié, voire les deux tiers, de la
lymphe du canal thoracique provient du foie et du tube digestif.
Réseau lymphatique de la plèvre diaphragmatique
Il est particulièrement riche et dense et les vaisseaux drainent la lymphe vers les chaînes
mammaires internes médiastinales et aussi vers les ganglions cœliaques à partir desquels ils
rejoignent l’origine du canal thoracique [309].
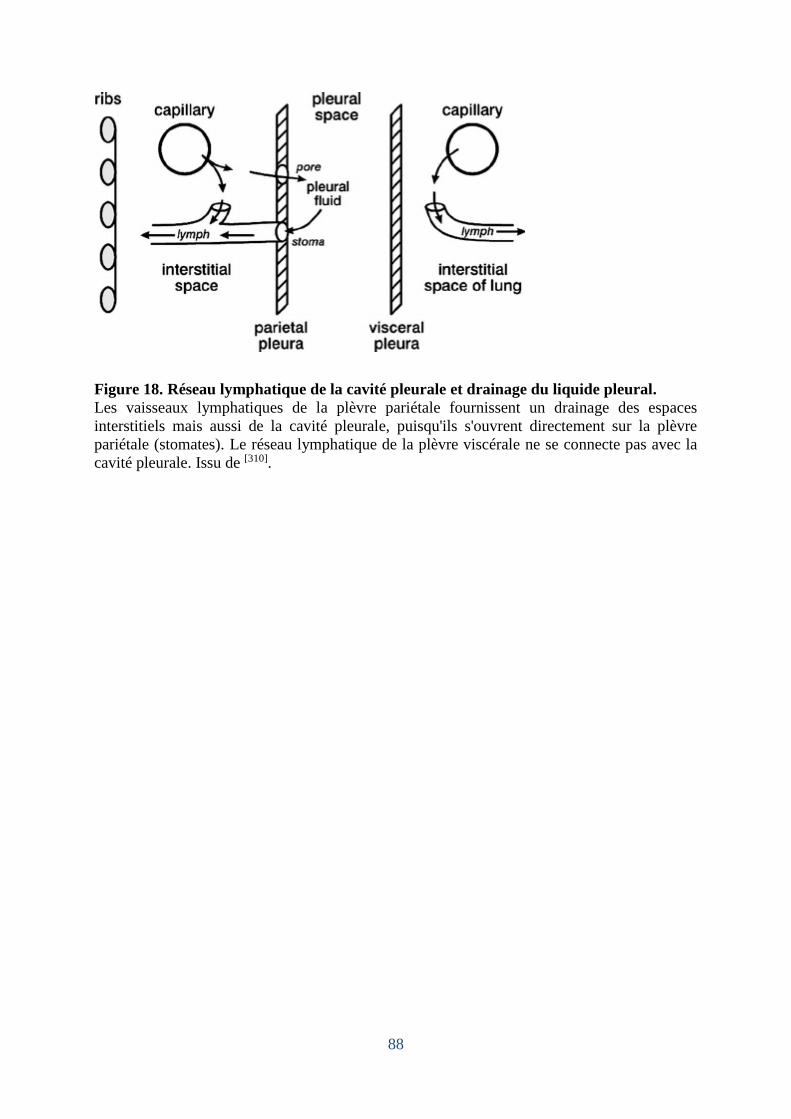
88
Figure 18. Réseau lymphatique de la cavité pleurale et drainage du liquide pleural.
Les vaisseaux lymphatiques de la plèvre pariétale fournissent un drainage des espaces
interstitiels mais aussi de la cavité pleurale, puisqu'ils s'ouvrent directement sur la plèvre
pariétale (stomates). Le réseau lymphatique de la plèvre viscérale ne se connecte pas avec la
cavité pleurale. Issu de [310].

89
Réseau lymphatique de la plèvre pariétale (paroi costale)
Le réseau lymphatique est très important notamment au niveau de la moitié inférieure et
postérieure de la paroi costale. Les vaisseaux lymphatiques sous-pleuraux se connectent
ensuite aux chaînes pariétales mammaires internes et aux courants ascendants latéro-
vertébraux.
Réseau lymphatique de la plèvre médiastinale (et péricardique)
Les capillaires et vaisseaux lymphatiques sont surtout facilement mis en évidence au niveau
du péricarde où les drainages se font vers les chaînes ganglionnaires du médiastin [311].
Réseau lymphatique de la plèvre viscérale pulmonaire
La plèvre viscérale possède un réseau lymphatique étendu. Les vaisseaux lymphatiques qui en
sont issus peuvent rejoindre les chaînes ganglionnaires du médiastin soit en restant sous-
pleuraux soit en se connectant au réseau lymphatique du poumon [312].
Réseau lymphatique et communications avec la cavité pleurale
Les vaisseaux lymphatiques de la plèvre viscérale ne se connectent pas avec la cavité pleurale
(Figure 18). En revanche, la plèvre pariétale et la plèvre diaphragmatique sont dotées de
stomates [313], orifices de 8 à 10 µm qui sont des abouchements directs des capillaires
lymphatiques sous-jacents. La cavité pleurale est ainsi directement drainée par le système
lymphatique (Figure 18).
Les lymphatiques du poumon
La lymphe issue de l’interstitium est véhiculée par des vaisseaux qui prennent
naissance au niveau des alvéoles pulmonaires sous forme de capillaires lymphatiques. Les
lymphatiques partent des segments pulmonaires et remontent le long des axes broncho-
artériels et des veines pulmonaires pour rejoindre le médiastin. Au long de ce parcours les
capillaires évoluent en vaisseaux collecteurs lymphatiques, valvulés et pulsatiles, et qui
véhiculent la lymphe de façon active. Dans le médiastin, ils poursuivent un trajet ascendant
pour aller s’aboucher dans la circulation veineuse au niveau de la veine sous-clavière. Ils
traversent des ganglions dans la plupart des cas. Toutefois, ces ganglions sont irrégulièrement
échelonnés le long de leur trajet et les lymphatiques peuvent avoir des trajets directs
aganglionnaires. Ainsi dans 20 à 35 % des cas, selon le segment pulmonaire drainé, la lymphe
se rend directement au médiastin sans traverser de ganglion intrapulmonaire [314].

90
6.4 Espace périvasculaire
Dans le poumon, les bronches, les artères pulmonaires, et les vaisseaux lymphatiques
passent dans une gaine interstitielle commune [306] ou espace périvasculaire (PVS,
« PeriVascular Space »). C'est un tissu conjonctif composé d'un ensemble de composants de
matrice extracellulaire (ECM) tels que du collagène, de l'élastine, de la fibronectine, des
protéoglycans comme le versican, et des polysaccharides comme de l'acide hyaluronique [315-
317]). Il contient aussi des fibroblastes et des macrophages [318]. Cet espace a un rôle évident de
structure dans le maintien de l'architecture du poumon mais semble aussi avoir un rôle
immunitaire important [317]. Une accumulation de leucocytes est souvent notifiée dans le PVS
dans des modèles animaux d'infections pulmonaires, de fibroses ou de réactions allergiques
[319-323].
Les fibres de collagène présentes dans le PVS pourraient offrir un point d'accroche
permettant aux leucocytes de traverser la paroi épaisse des artères et des veines pulmonaires
[321]. Au cours de l'inflammation pulmonaire aiguë, l’ECM se remodèle pour permettre
l'infiltration de leucocytes. Cette ECM provisoire implique l'accumulation d'acide
hyaluronique et de versican, qui, ensemble, forment une matrice permettant l'entrée des
leucocytes et l'expansion des cellules stromales résidentes [316,317]. Le blocage du principal
récepteur à l'acide hyaluronique, le CD44, induit une baisse de l'accumulation d'éosinophiles
dans un modèle d'asthme [324,325]. Les souris déficientes en versican ne présentent pas
d'infiltrats périvasculaires dans un modèle d'inflammation pulmonaire induite chimiquement
[316]. De plus, le versican aurait aussi un rôle dans le contrôle de l'inflammation : il peut fixer
de nombreuses chimiokines, ce qui permettrait de contrôler leur disponibilité [317,326,327].

91
6.5 Les macrophages pulmonaires
Au moins deux populations de macrophages sont présentes dans les poumons : les
macrophages alvéolaires (Alveolar macrophage, AM) et les macrophages interstitiels
(Interstitial macrophage, IM) [318,328,329] (Tableau 7).
Les AM sont localisés dans les alvéoles - où ils représentent 90 à 95% du contenu
cellulaire à l'homéostasie - ainsi que dans le mucus des voies respiratoires plus grandes [304].
Étant donné qu'il y a plusieurs millions d'alvéoles et seulement environ un à deux millions
d'AM dans un poumon de souris, toutes les alvéoles ne contiennent pas d'AM ; en effet, un
seul AM est détecté pour environ trois alvéoles. Cependant les AM peuvent se déplacer entre
les alvéoles à travers des pores de liaison dits "pores de Kohn" [328]. Leur rôle principal est de
cataboliser le surfactant, d'éliminer les particules, les cellules apoptotiques et les débris
cellulaires des alvéoles afin de maintenir la fonction pulmonaire [304,330]. Les AM reçoivent
constamment des signaux de régulation négative des cellules de l'épithélium alvéolaire ; Ces
signaux comprennent le CD200, une protéine membranaire exprimée par les cellules
épithéliales, le TGF-ß et l'IL-10, qui empêchent l'activation des macrophages alvéolaires
[304,331]. Les AM produisent aussi du TGF-β qui entraîne la génération de cellules Foxp3 +
iTreg à partir de cellules T CD4+ naïves, ces cellules induisant une tolérance aux antigènes
inhalés. Ce processus se produit dans le tissu pulmonaire et non dans les ganglions
lymphatiques drainants et dépend de la présentation de l'antigène par les AM, et non par les
DC [328]. Lors de l'infection, la régulation négative qui contraint normalement l'activation des
AM est perdue (en raison notamment de la mort des cellules épithéliales). Les AM mettent
alors en œuvre des mécanismes de signalisation [304] pour induire le passage d'un mode de
tolérance à un mode inflammatoire via la sécrétion d'IL-1, d'IL-6, de TNF-α et de TGF-β [328].
Les AM ont un rôle majeur dans la défense contre les infections et dans la prévention des
lésions pulmonaires aiguës par la limitation des dommages tissulaires et leur réparation après
l'infection. La capacité phagocytaire des AM augmente considérablement lors des infections,
grâce notamment aux collectines SP-A, SP-D et C1q, présentes dans le surfactant pulmonaire
qui peuvent se fixer sur les pathogènes et les cellules apoptotiques [332].
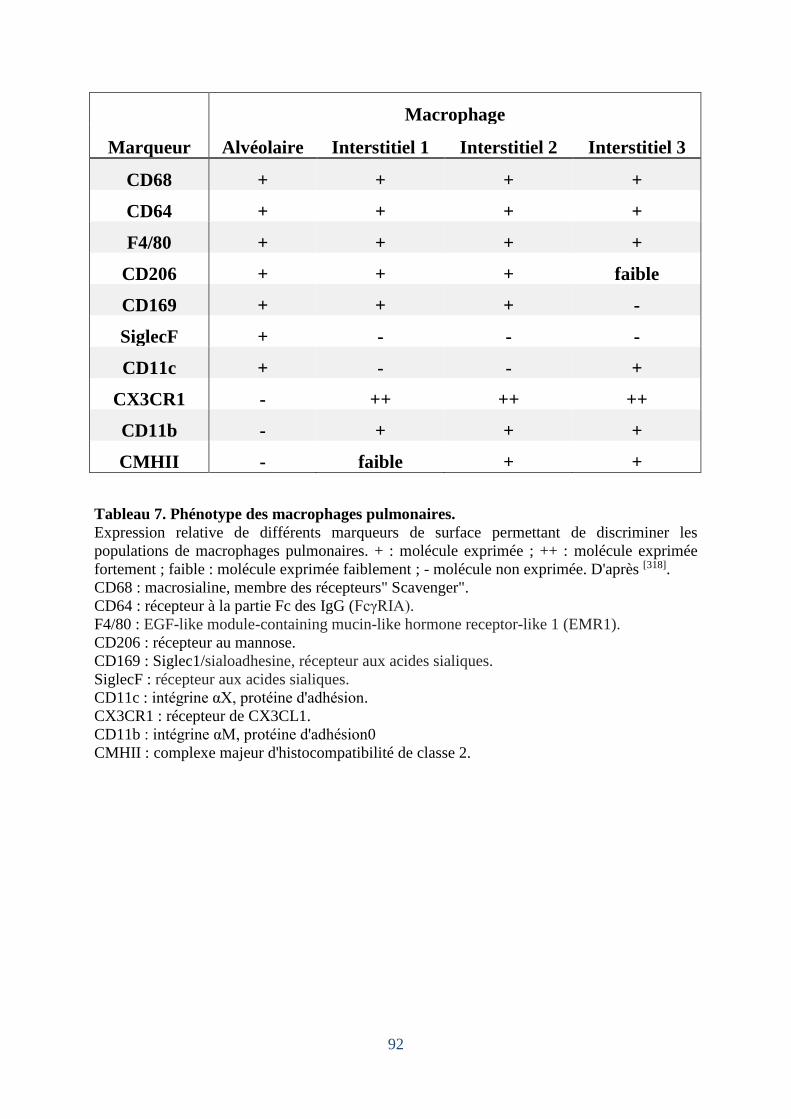
92
Macrophage
Marqueur Alvéolaire Interstitiel 1 Interstitiel 2 Interstitiel 3
CD68 + + + +
CD64 + + + +
F4/80 + + + +
CD206 + + + faible
CD169 + + + -
SiglecF + - - -
CD11c + - - +
CX3CR1 - ++ ++ ++
CD11b - + + +
CMHII - faible + +
Tableau 7. Phénotype des macrophages pulmonaires.
Expression relative de différents marqueurs de surface permettant de discriminer les
populations de macrophages pulmonaires. + : molécule exprimée ; ++ : molécule exprimée
fortement ; faible : molécule exprimée faiblement ; - molécule non exprimée. D'après [318].
CD68 : macrosialine, membre des récepteurs" Scavenger".
CD64 : récepteur à la partie Fc des IgG (FcγRIA).
F4/80 : EGF-like module-containing mucin-like hormone receptor-like 1 (EMR1).
CD206 : récepteur au mannose.
CD169 : Siglec1/sialoadhesine, récepteur aux acides sialiques.
SiglecF : récepteur aux acides sialiques.
CD11c : intégrine αX, protéine d'adhésion.
CX3CR1 : récepteur de CX3CL1.
CD11b : intégrine αM, protéine d'adhésion0
CMHII : complexe majeur d'histocompatibilité de classe 2.

93
Les collectines présentent deux parties : une partie lectine qui se fixe sur les
polysaccharides et une queue collagéneuse, reconnue par le récepteur CD91 des macrophages
[328,332].
Les macrophages interstitiels (IM) ne sont pas bien caractérisés, en partie à cause des
difficultés d'identification et d'isolement [318,333]. Une étude récente suggère que les IM
pulmonaires sont situés dans l'interstitium bronchovasculaire (c’est-à-dire l’espace
périvasculaire ou le PVS) et non dans l'interstitium alvéolaire [318]. Cette étude décrit 3
populations d'IM qui s'auto-renouvèlent et présentent des marqueurs et des profils
transcriptomiques différents (Tableau 7). Deux de ces populations ont un phénotype assez
similaire, avec des capacités de phagocytose et une expression du marqueur CD169. Ces
populations de macrophages CD169+ s'accroissent lors de l'infection par Schistosoma mansoni
[334] et sont présentes à proximité des Tissus Lymphoïdes Associés aux Bronches (BALT) [335].
94
Figure 19. Résumé des principales pathologies pulmonaires.
De nombreux facteurs infectieux et non infectieux peuvent induire des pathologies
pulmonaires. Les pathologies infectieuses peuvent être d'origine virale, bactérienne, fongique
ou parasitaire. Les pathologies non infectieuses sont d'origine environnementale, génétique ou
tumorale.
COPD : Chronic Obstructive Pulmonary Disease / broncho-pneumopathie chronique
obstructive
ARDS : Acute respiratory distress syndrome / syndrome de détresse respiratoire aiguë
RSV : respiratory syncytial virus / virus respiratoire syncytial
TRALI : transfusion related acute lung injury / Œdème pulmonaire lésionnel post-
transfusionnel

95
7. Dommages pulmonaires dus aux anchylostomes
Les poumons, de par leur situation à l'interface entre le milieu extérieur et le sang sont
constamment exposés à des particules dangereuses ou des agents infectieux. Cependant, très
peu d'informations sont disponibles pour les filarioses mais de nombreuses pathologies sont
bien décrites (Figure 19), y compris des pathologies parasitaires à nématodes, dont les
anchylostomes.
7.1 Cycle de vie des anchylostomes
Les anchylostomes (dont Necator Americanus et Anchylostoma duodenale chez
l'humain, Anchylostoma caninus chez le chien et Nippostrongylus brasiliensis chez les
rongeurs) sont des nématodes gastro-intestinaux de mammifères [4]. Les larves infestantes
(L3) sont très mobiles. La contamination se fait par voie transcutanée et les larves migrent
dans l’organisme humain pour devenir adultes dans la région duodéno-jéjunale. La migration
des L3 vers les intestins de leur hôte implique un passage pulmonaire [336]. Les L3 pénètrent
activement dans la peau et il est suggéré que les L3 entrent dans les vaisseaux sanguins et sont
ainsi acheminées via le cœur droit aux poumons [277,337] ; toutefois une entrée dans les
vaisseaux lymphatiques de la peau a été décrite chez le chien [338], ce qui suggère que les L3
pourraient rejoindre la circulation sanguine via le canal thoracique. Les parasites seraient
bloqués dans les capillaires pulmonaires et ils en sortiraient activement pour entrer dans le
parenchyme ; en effet, des hémorragies (pétéchies) sont observées sur les poumons de souris
infestées par N. brasiliensis entre 1 et 3 jours après infection [277,339,340]. Les L3 sont
retrouvées dans des alvéoles partiellement détruites présentant des hémorragies. Là, elles
subissent une mue (L4) et, après 48 h, elles remontent les voies respiratoires et la trachée, où
elles sont toussées et avalées par l'hôte [336]. Elles continuent leur développement dans le tube
digestif où les adultes se reproduisent et pondent leurs œufs [277].
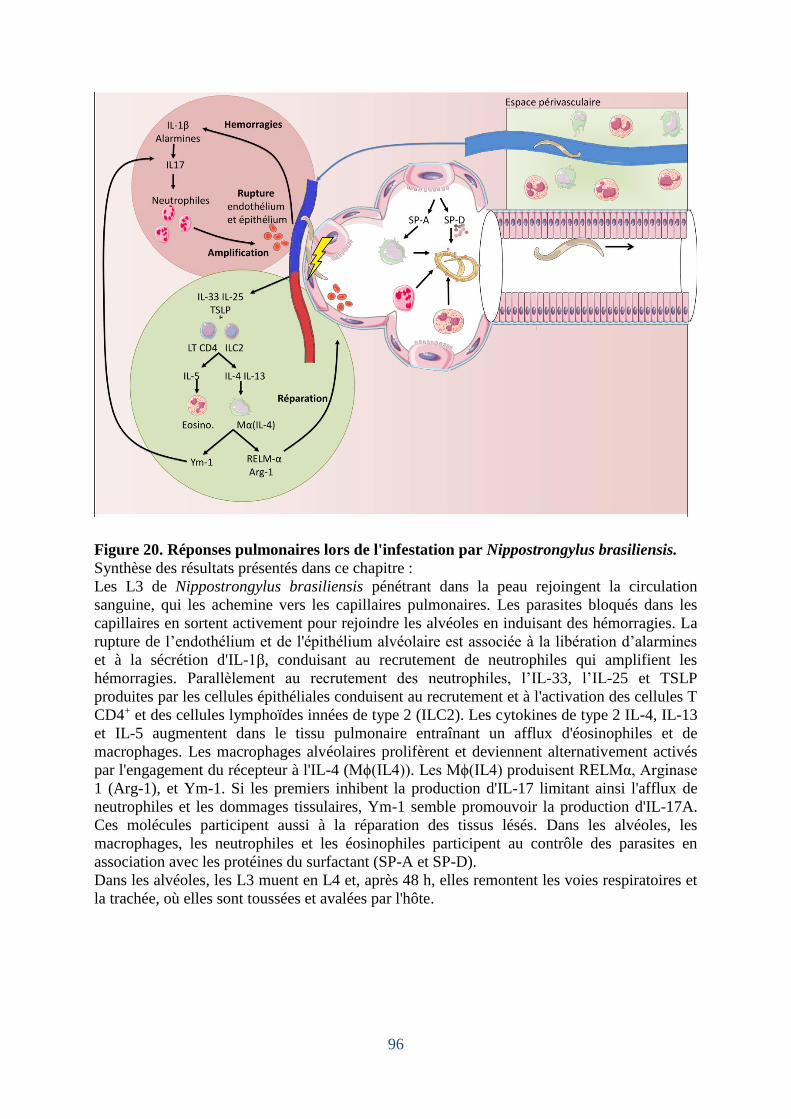
96
Figure 20. Réponses pulmonaires lors de l'infestation par Nippostrongylus brasiliensis.
Synthèse des résultats présentés dans ce chapitre :
Les L3 de Nippostrongylus brasiliensis pénétrant dans la peau rejoingent la circulation
sanguine, qui les achemine vers les capillaires pulmonaires. Les parasites bloqués dans les
capillaires en sortent activement pour rejoindre les alvéoles en induisant des hémorragies. La
rupture de l’endothélium et de l'épithélium alvéolaire est associée à la libération d’alarmines
et à la sécrétion d'IL-1β, conduisant au recrutement de neutrophiles qui amplifient les
hémorragies. Parallèlement au recrutement des neutrophiles, l’IL-33, l’IL-25 et TSLP
produites par les cellules épithéliales conduisent au recrutement et à l'activation des cellules T
CD4+ et des cellules lymphoïdes innées de type 2 (ILC2). Les cytokines de type 2 IL-4, IL-13
et IL-5 augmentent dans le tissu pulmonaire entraînant un afflux d'éosinophiles et de
macrophages. Les macrophages alvéolaires prolifèrent et deviennent alternativement activés
par l'engagement du récepteur à l'IL-4 (Mϕ(IL4)). Les Mϕ(IL4) produisent RELMα, Arginase
1 (Arg-1), et Ym-1. Si les premiers inhibent la production d'IL-17 limitant ainsi l'afflux de
neutrophiles et les dommages tissulaires, Ym-1 semble promouvoir la production d'IL-17A.
Ces molécules participent aussi à la réparation des tissus lésés. Dans les alvéoles, les
macrophages, les neutrophiles et les éosinophiles participent au contrôle des parasites en
association avec les protéines du surfactant (SP-A et SP-D).
Dans les alvéoles, les L3 muent en L4 et, après 48 h, elles remontent les voies respiratoires et
la trachée, où elles sont toussées et avalées par l'hôte.

97
7.2 Neutrophiles et amplification de la pathologie pulmonaire.
Au fur et à mesure que les larves traversent le poumon, la rupture de l’endothélium et
de l'épithélium alvéolaire est associée à la libération d’alarmines (« motifs moléculaires
associés au danger » ou DAMP) et à la sécrétion d'IL-1β par les cellules épithéliales des voies
respiratoires. Les niveaux d'IL-17A augmentent rapidement dans les poumons des souris
infestées par N. brasiliensis. La fixation de l'IL-17 sur son récepteur à la surface des cellules
épithéliales induit la production de chimiokines telles que l'IL-8 ou CXCL1, qui ont un fort
pouvoir chimiotactique sur les neutrophiles [341]. Il s'ensuit donc un recrutement important de
neutrophiles sur le lieu de l'infection [339,342] (Figure 20). La déplétion des neutrophiles avant
infection augmente le rendement parasitaire, mais diminue l'intensité des lésions tissulaires et
les hémorragies [339,343]. Ainsi, les neutrophiles ont un effet anti-parasitaire mais au prix d'une
pathologie pulmonaire accrue [342]. Les molécules impliquées dans l'exacerbation de la
pathologie induite par les neutrophiles ne sont pas connues, cependant la myelopéroxydase
(MPO, qui catalyse la formation d'espèces réactives de l'oxygène) des neutrophiles est
nécessaire à l'élimination des larves [343,344]. Ce stress oxydatif pourrait induire des dommages
collatéraux sur les tissus environnants. Le nombre de neutrophiles culmine 2 jours après
infection et retrouve son niveau normal dès 7 jours après infection [320].
7.3 Macrophages : contrôle de sous populations de parasites et réparation des
tissus
Parallèlement au recrutement des neutrophiles, l’IL-33, l’IL-25 et TSLP produites par
les cellules épithéliales conduisent au recrutement et à l'activation des cellules T CD4+ et des
cellules lymphoïdes innées de type 2 (ILC2) [340,345-347]. Les cytokines de type 2 IL-4, IL-13,
IL-9 et IL-5 augmentent dans le tissu entraînant un afflux d'éosinophiles et de macrophages 4
jours après infection, notamment dans l'espace périvasculaire [320,348]. Les macrophages
alvéolaires prolifèrent et deviennent alternativement activés par l'engagement du récepteur à
l'IL-4 (IL-4Rα / ces macrophages sont notés Mϕ(IL4)) [277,344,346,349]. De plus des molécules
comme les Protéines du Surfactant (SP)-A et SP-D augmentent dans le liquide alvéolaire. Ces
molécules participent à l'activation des macrophages mais aussi à l'élimination des parasites et
à la réparation des tissus [346,349]. SP-D peut se fixer aux carbohydrates présentes à la surface
des L4 et aide les macrophages alvéolaires à tuer les parasites en agissant comme une
opsonine [346]. De même, l'absence de SP-A entraîne des rendements parasitaires plus élevés

98
ainsi que des dommages tissulaires exacerbés [349]. SP-A augmente l'activation et la
prolifération des macrophages médiées par l'IL-4 et favorise la réparation des tissus [349].
Les macrophages sont indispensables à la réparation des tissus après une blessure [350].
La suppression des macrophages dans les souris infestées par N. brasiliensis exacerbe les
hémorragies causées par la migration du parasite et entraîne des lésions pulmonaires plus
sévères [339]. Les Mϕ(IL4) produisent RELMα, Arginase 1 (Arg-1), et Ym-1 [277]. Si les
premiers inhibent la production d'IL-17 limitant ainsi l'afflux de neutrophiles et les dommages
tissulaires, Ym-1 semble promouvoir la production d'IL-17A [339,342]. L'arginase convertit
l'arginine en ornithine, qui peut servir de précurseur pour la synthèse de la proline et
l'hydroxyproline, constituants centraux du collagène nécessaire à cicatrisation. L'arginase est
également anti-inflammatoire car elle est en concurrence pour l'arginine avec iNOS
(produisant l'oxyde nitrique) [350]. La phase de cicatrisation est caractérisée par une
prolifération et une différenciation cellulaires visant à remplacer les cellules détruites, ainsi
que par la création de matrice extracellulaire (ECM) et le dépôt de collagène par les
fibroblastes pour supporter les nouvelles cellules [350].
Cependant, malgré une réparation rapide des tissus, une inflammation pulmonaire persiste
pendant plus de 300 jours après l'infection, soit bien après que les parasites aient été éliminés
par les souris [351].

99
OBJECTIFS DE LA THESE
L'objectif principal de cette étude est de caractériser les interactions qu'entretiennent
les filaires avec l'environnement pulmonaire dans le modèle murin de filariose Litomosoides
sigmodontis lors de deux phases de l'infection :
1. Lors de la phase précoce de l'infection (0h - 12 jours post-infection)
2. Lors de la phase chronique de l'infection (55 jours - 100 jours post-infection)
Les objectifs spécifiques sont :
- Décrire la migration pulmonaire des L3 de L. sigmodontis
- Analyser la réponse inflammatoire/immunitaire associée à cette phase précoce de
l'infection
- Comprendre la fonction des neutrophiles dans cette réponse inflammatoire en étudiant
les NETs
- Préciser le rôle de l'alarmine S100A9 et de l'héterodimère S100A8/S100A9
(calprotectine) lors de la phase précoce et pré-patente de l'infection par L. sigmodontis
- Décrire la pathologie pulmonaire observée en phase patente
- Analyser la réponse inflammatoire/immunitaire associée à la phase patente de
l'infection
- Définir les différences entre souris microfilarémiques et souris amicrofilarémiques

100

101
MATERIEL ET METHODES

102
1. Éthique
Toutes les procédures expérimentales ont été réalisées en conformité avec la Directive
UE 2010/63 / UE et la législation nationale, à savoir le Décret n ° 2013-118 du 1er février
2013, du Ministère de l'Agriculture, de l'Agroalimentaire et de la Forêt". Les protocoles ont
été approuvés par le comité d'éthique du Musée national d'histoire naturelle (68-002) et par la
Direction départementale de la protection des populations (n ° C75- 05-15).
2. Parasites, souris et inoculation des larves /L3 de L. sigmodontis
2.1 Maintien du cycle de Litomosoides sigmodontis
Le cycle de la filaire Litomosoides sigmodontis est maintenu au laboratoire en utilisant
comme vecteur l'acarien hématophage Ornithonyssus bacoti et comme hôte réservoir la
gerbille Meriones unguiculatus [9].
L'élevage d'O. bacoti est maintenu à une température de 26-28°C et une humidité
relative de 60-80% sur des gerbilles saines. Les gerbilles « réservoir » (males ou femelles, 6-8
semaines) sont infestées par une dose de 80 L3 injectées en intrapéritonéal sous 200µL de
RPMI 1640 additionné 25 mM d'Hépès (Eurobio) avec une aiguille 23G (Terumo, Neolus).
Après 3 mois d'infection, un prélèvement 10µl de sang au sinus rétro-orbital est réalisé pour
évaluer la microfilarémie (charge en microfilaires sanguines) ; le sang est étalé sur une lame
de verre, coloré au GIEMSA (solution d'azur éosine-bleu de méthylène, MERCK) à 10% et
les microfilaires sont comptées à l’aide d’un microscope optique. Les gerbilles présentant les
microfilarémies les plus fortes (>1000 mf/10µL) sont utilisées pour infecter les acariens. Pour
ce faire, des acariens sains sont prélevés et mis à jeûner 5 à 7 jours avant contact avec la
gerbille infestée. Le gorgement des acariens s'effectue sur une nuit. Les acariens sont ensuite
récupérés dans des tubes en verre fermés par du coton (50 acariens par tube) et placés dans
une chambre humide (28°C, 80% d'humidité) pendant au moins 12 jours pour obtenir des L3
et avant 19 jours pour limiter la mortalité des acariens infectés. Les acariens infestés de 12-19
jours sont placés dans une boite de Pétri en verre de 5cm de diamètre avec du RPMI 1640
contenant 25 mM d'Hépès et disséqués avec des pinces fines sous une loupe binoculaire pour
que les L3 soient libérées. Les L3 sont récoltées à l’aide d’une pipette Pasteur en verre effilée
pour constituer les doses infectantes.

103
2.2 Les souris
2.2.1 Souris BALB/c
Des souris femelles de la lignée consanguine BALB/c (Envigo) âgées de 6 à 8
semaines ont été utilisées. La lignée BALB/c est la plus sensible à l'infection par L.
sigmodontis et environ 50% des souris développe une phase patente (présence de microfilaires
dans le sang à partir de 55 jours après infection [203] suite à une injection en sous-cutané. Les
souris BALB/c femelles présentent une microfilarémie plus élevée que les souris BALB/c
mâles [194]. Les souris sont maintenues dans les installations du MNHN avec un cycle lumière
/ obscurité de 12 heures.
2.2.2 Souris BALB/c IL-4Rα-/-/IL-5-/-
Des souris BALB/c doublement déficientes en IL-5 et pour la chaîne α du récepteur à
IL-4 (IL-4Rα-/-/IL-5-/-) ont été utilisées [352-354]. Les souris mâles et femelles âgées de 6 à 10
semaines ont été retenues. Les couples reproducteurs initiaux proviennent de l’Université
clinique de Bonn en Allemagne (Dr Marc Huebner) et les souris sont maintenues et
reproduites dans les installations du MNHN. Ces souris sont extrêmement sensibles à
l'infection et présentent des microfilarémies très élevées (Marc Huebner, communication
personnelle).
2.2.3 Souris C57BL/6 S100A9-/-
Des souris C57BL/6 déficientes en S100A9 (S100A9-/-) [355] mâles et femelles, âgées
de 6 à 10 semaines, ont été utilisées à l’Institut de Microbiologie Médicale, d'Immunologie et
de Parasitologie de l'Université clinique de Bonn, où ont eu lieu les expériences en
collaboration avec le Dr Marc Huebner.
2.3 Inoculation aux souris
Les L3 ont été inoculées soit :
- par injection de 40 L3 en sous-cutané sous 0.2 mL de RPMI au niveau de la région lombaire
gauche
- par injection de 40 L3 en intraveineuse caudale sous 50 µL de RMPI.
- par transmission pendant le repas sanguin de l'acarien O. bacoti ("infection naturelle") : la
moitié d’un lot d’acariens est mis en contact avec un groupe de souris pendant 12heures [9]. La
deuxième moitié du même lot d’acarien a été disséqué sous une loupe binoculaire pour

104
évaluer le nombre moyen de L3 par acarien. Comme il a été démontré précédemment
qu'aucune L3 ne reste dans les acariens pendant leur repas sanguin [9], cela nous a permis
d'évaluer le nombre de L3 inoculées par souris.
La cinétique de l'infection a été suivie pendant 100 jours post infection. Les souris ont
été sacrifiées à 2 heures, 6 heures, 4 jours, 6 jours, 8 jours, 12 jours, 14 jours, 30 jours, 70
jours et 100 jours après l'inoculation (p.i.)
2.4 Suivi de la microfilarémie des souris
La microfilarémie est mesurée 60 et 70 jours post-infection. Une goutte de 10 µL de
sang est prélevée soit au sinus rétro-orbital (J60 p.i. et J70 p.i.) ou dans le cœur (J70 p.i. et
J100 p.i.) et déposée sur une lame en verre. La goutte est tournée énergiquement et étalée avec
le coin d'une lame en verre afin d'empêcher la coagulation (défibrination) pour créer une zone
d’environ 15mm de diamètre. La lame est ensuite séchée à l'étuve. Les hématies sont éclatées
en mettant les lames dans l'eau du robinet pendant 2 à 3 minutes et la goutte épaisse est fixée
par un court bain dans du méthanol.
Les lames sont ensuite colorées dans une solution de GIEMSA (solution d'azur éosine-
bleu de méthylène, MERCK) à 10% (solution tampon : hydrogénophosphate disodique à
0,5g/L + phosphate de potassium monopotassique à 0,2g/L), rincées à l'eau et séchées à
l'étuve.
3. Autopsie des souris, collecte des filaires, prélèvement et fixation des poumons
3.1 Exsanguination et préparation du sérum
Les souris sont euthanasiées par exsanguination totale au sinus rétro-orbital. Le sang
est recueilli dans des tubes de 1,5 mL. Après 30 minutes minimum et 1h maximum pour
permettre la coagulation, le sang est centrifugé (16 000g, 5min) puis le sérum est isolé et
conservé à -20°C pour doser ultérieurement différentes cytokines et autres molécules
inflammatoires.
3.2 Lavage de la cavité pleurale
Après avoir pratiqué une légère ouverture du diaphragme, la cavité pleurale de chaque
souris est rincée par flux et reflux de 10 x 1 ml de PBS 1X froid avec une pipette Pasteur en
plastique. Les 2 premiers ml sont déposés dans un tube de 15ml et les 8 ml suivants dans un
deuxième tube de 15ml sur glace. Après décantation de 20 min, 1 ml est prélevé du 1er tube,

105
centrifugé (5min, 500g) et le surnageant est congelé à -20°C pour analyses ultérieures
(cytokines) ; le culot est repris dans le ml restant du premier tube et l'ensemble est déposé
dans le deuxième tube. Les 9 ml obtenus sont décantés plusieurs minutes pour permettre aux
filaires de se déposer au fond du tube mais pas les cellules. Les 8ml supérieurs du deuxième
tube sont isolés dans un nouveau tube de 15ml, centrifugés (5min, 350g), et le culot de
cellules de l’exsudat pleural (PleCs) est repris dans 1mL de PBS- 2% SVF (EUROBIO) pour
phénotypage cellulaire par cytométrie en flux (voir paragraphe 8). Les filaires isolées sont
transférées dans une boite de Pétri en verre (5cm de diamètre) pour comptage et analyse
morphologique.
3.3 Evaluation du développement filarien
Les filaires isolées sont comptées et le rendement filarien (F/L3) est établi. Il
correspond à (100 x nombre de vers récupérés) / nombre de larves inoculées.
Plusieurs autres paramètres sont mesurés :
1. Le nombre de filaires vivantes et entourées d'un manchon de cellules inflammatoires.
Elles sont incluses dans le dénombrement des filaires (cF).
2. Le nombre de filaires mortes entourées de cellules réactionnelles constituant des
granulomes (G). Ces granulomes sont de taille variable ; les plus gros peuvent contenir
des filaires reconnaissables, les petits des débris de filaires. Ils ne sont pas inclus dans le
rendement parasitaire.
3. Le nombre de filaires femelles (nf)
4. Le nombre de filaires mâles (nm)
5. Le sex ratio (f/m +f)
6. La longueur des filaires (L)
7. La présence d'œufs et de microfilaires dans les femelles.
3.4 Lavage bronchoalvéolaire
La trachée est exposée et incisée à la jonction cervico-thoracique. Une canule est
insérée et fixée avec du fil. L'espace bronchoalvéolaire est lavé avec 10 fois 1 ml de PBS 1X
froid. Le premier ml est recueilli dans un premier tube, centrifugé (5min, 350g) et le
surnageant est congelé (-20°C). Le culot est repris dans les 9 ml restants du lavage. Après
centrifugation (5min, 350 g), le culot contenant les cellules du liquide de lavage
bronchoalvéolaire (LBA) est repris dans 1 ml de PBS - 2% SVF.

106
3.5 Dénombrement des L3 dans les poumons
Après avoir réalisé les lavages pleuraux et broncho-alvéolaires, les poumons sont
retirés de la cage thoracique (à H2, H6, J2, J4, J6 et J8 p.i.). Les poumons sont ensuite placés
dans des boîtes de Pétri de 5 cm de diamètre contenant 7 ml de PBS et déchirés en petits
morceaux (environ 1-3mm2) à l’aide de pinces fines. Les boîtes de Pétri sont examinées sous
une loupe binoculaire à différents temps entre 1 h et 24 h pour permettre aux L3 de sortir du
tissu. Les L3 isolées sont ensuite comptées.
3.6 Préparation et fixation des poumons pour différentes analyses
Pour l'observation des poumons en microscopie optique (voir paragraphe 4), les
poumons sont gonflés avec du formaldéhyde (VWR) à 4% dans du PBS, enlevés et plongés
dans du formaldéhyde 4%-PBS pendant 24h. Après 24h, le fixateur est renouvelé pour 24h
supplémentaires puis les poumons sont placés en éthanol 70%.
Pour l'observation des poumons en microscopie confocale (voir paragraphe 5), les
poumons sont remplis par la trachée avec de l'agarose à bas point de fusion à 2% (p / v dans
du PBS 1X, pH 7,4) chauffé à 40°C (Sigma-Aldrich). Après refroidissement de la souris sur
glace quelques minutes pour permettre à l’agarose de durcir, les poumons sont retirés du
thorax et transférés dans du PBS froid pour rinçage, puis fixés 2 h dans du paraformaldéhyde
(PFA, Electron Microscopy Science) froid à 4% dans du PBS et finalement stockés à 4°C
dans du PBS 1X.
Pour l'observation des poumons en microscopie électronique à balayage (voir
paragraphe 8), les poumons sont retirés du thorax et placés dans une boite de Pétri contenant
du PBS. Les poumons sont coupés au scalpel en tranches d'environ 3-4mm. Les sections sont
ensuite fixées pendant 1h dans une solution de glutaraldéhyde à 2.5%.
Pour l'analyse des transcrits pulmonaires à J4 et J70 p.i. (voir paragraphe 9), l'aorte est
coupée et les poumons sont exsanguinés par injection de 10mL de PBS froid dans le
ventricule droit du cœur puis immergés dans du RNAlater et congelés à -80°C.
Pour les extractions d'ADN afin d’évaluer la quantité de microfilaires pulmonaires à
J70 p.i. (voir paragraphe 10), les poumons sont gonflés avec de l'éthanol 70% via une canule
insérée dans la trachée puis ils sont extraits de la cavité et plongés dans un tube de
50mlcontenant de l'éthanol 70% puis stocké à 4°C.

107
4. Analyses histologiques et immunohistologique sur coupes fines
4.1 Déshydratation, inclusion en paraffine, préparation des lames
La paraffine n'est pas miscible à l'eau, les organes doivent donc être déshydratés avant
l'inclusion dans la paraffine. La paraffine n'est pas non plus soluble dans l'alcool utilisé pour
la déshydratation. On procède donc à une double substitution. On remplace l'eau par de
l'alcool (déshydratation) puis on remplace l'alcool par le toluène (substitution).
Les organes fixés au formaldéhyde sont déshydratés par une succession de 3 bains
d'alcool 70%, 3 bains d'alcool 95%, 3 bains d'alcool 100% puis 3 bains de toluène. Chaque
bain a une durée de 30 minutes. Enfin les organes sont mis dans un bain de paraffine liquide
(Leica-Paraplast PLUS) à 57°C pendant 2 jours pour bien imprégner les organes. Les blocs de
paraffine contenant les organes sont coulés dans des moules d'inclusion avec une station
d'inclusion en paraffine (Leica EG1150H) et une cassette permettant le montage sur le
microtome est collée sur le dessus du bloc. Le bloc est ensuit refroidi sur une plaque de
refroidissement (Leica EG1150C) pour figer la paraffine.
Les blocs sont coupés au microtome (Leica RM2245) pour obtenir des coupes d’une
épaisseur de 5µm. Les sections ainsi obtenues sont placées à la surface de l'eau d'un bain-
marie chauffant à 40°C afin de défroisser le tissu inclus dans la paraffine sans faire fondre
cette dernière. Une lame de verre est plongée dans le bain à la verticale puis, en remontant, les
coupes lisses s'attachent à la lame. Les lames sont ensuite égouttées pour chasser l'excès d'eau
puis séchées à l'étuve à 57°C toute une nuit.
Pour les colorations histologiques, des lames de verre de qualité standard ont été
utilisées. Les coupes pour les immunomarquages ont été étalées sur des lames Superfrost
(Superfrost plus, Thermo scientific). Ces lames sont chargées positivement, ce qui augmente
l'adhérence des tissus aux lames.
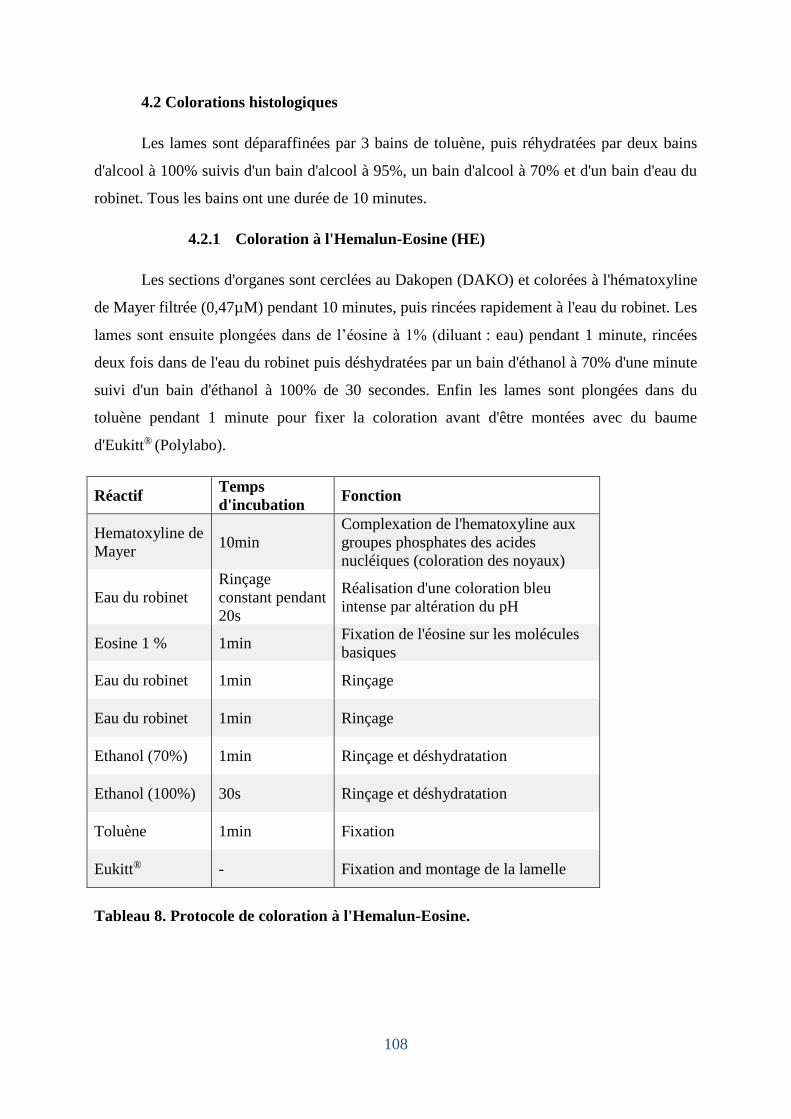
108
4.2 Colorations histologiques
Les lames sont déparaffinées par 3 bains de toluène, puis réhydratées par deux bains
d'alcool à 100% suivis d'un bain d'alcool à 95%, un bain d'alcool à 70% et d'un bain d'eau du
robinet. Tous les bains ont une durée de 10 minutes.
4.2.1 Coloration à l'Hemalun-Eosine (HE)
Les sections d'organes sont cerclées au Dakopen (DAKO) et colorées à l'hématoxyline
de Mayer filtrée (0,47µM) pendant 10 minutes, puis rincées rapidement à l'eau du robinet. Les
lames sont ensuite plongées dans de l’éosine à 1% (diluant : eau) pendant 1 minute, rincées
deux fois dans de l'eau du robinet puis déshydratées par un bain d'éthanol à 70% d'une minute
suivi d'un bain d'éthanol à 100% de 30 secondes. Enfin les lames sont plongées dans du
toluène pendant 1 minute pour fixer la coloration avant d'être montées avec du baume
d'Eukitt® (Polylabo).
Réactif Temps
d'incubation Fonction
Hematoxyline de
Mayer 10min
Complexation de l'hematoxyline aux
groupes phosphates des acides
nucléiques (coloration des noyaux)
Eau du robinet
Rinçage
constant pendant
20s
Réalisation d'une coloration bleu
intense par altération du pH
Eosine 1 % 1min Fixation de l'éosine sur les molécules
basiques
Eau du robinet 1min Rinçage
Eau du robinet 1min Rinçage
Ethanol (70%) 1min Rinçage et déshydratation
Ethanol (100%) 30s Rinçage et déshydratation
Toluène 1min Fixation
Eukitt® - Fixation and montage de la lamelle
Tableau 8. Protocole de coloration à l'Hemalun-Eosine.
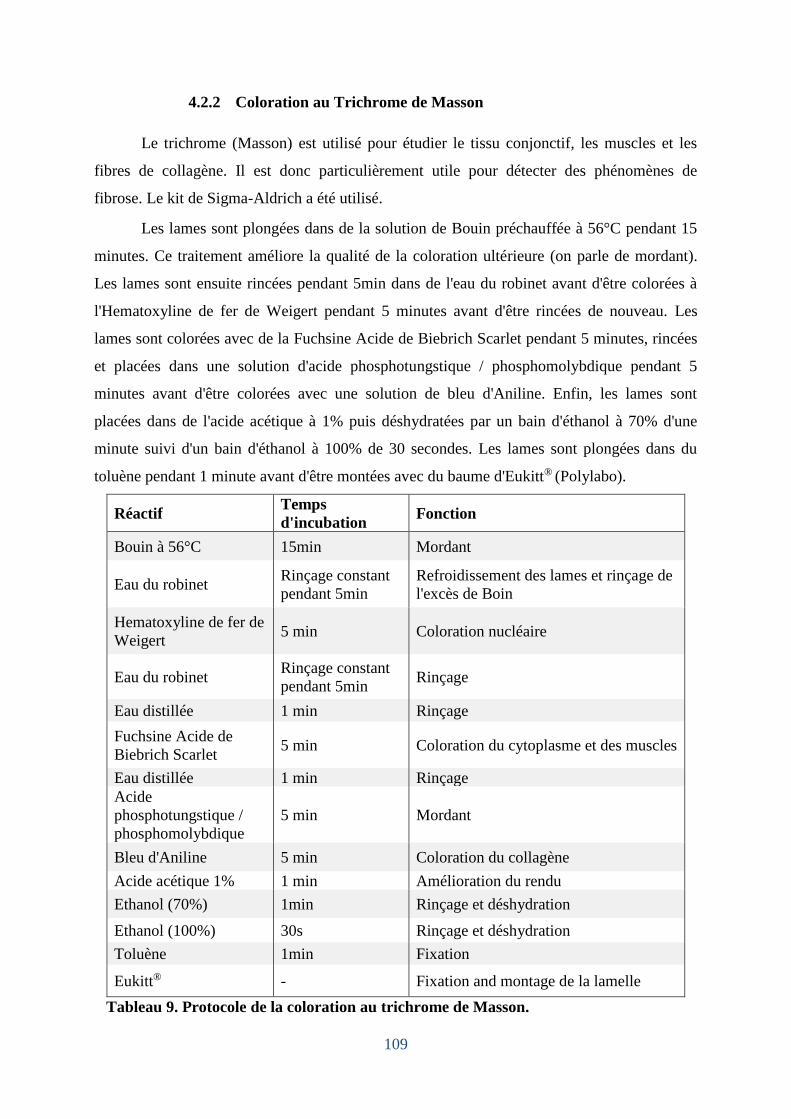
109
4.2.2 Coloration au Trichrome de Masson
Le trichrome (Masson) est utilisé pour étudier le tissu conjonctif, les muscles et les
fibres de collagène. Il est donc particulièrement utile pour détecter des phénomènes de
fibrose. Le kit de Sigma-Aldrich a été utilisé.
Les lames sont plongées dans de la solution de Bouin préchauffée à 56°C pendant 15
minutes. Ce traitement améliore la qualité de la coloration ultérieure (on parle de mordant).
Les lames sont ensuite rincées pendant 5min dans de l'eau du robinet avant d'être colorées à
l'Hematoxyline de fer de Weigert pendant 5 minutes avant d'être rincées de nouveau. Les
lames sont colorées avec de la Fuchsine Acide de Biebrich Scarlet pendant 5 minutes, rincées
et placées dans une solution d'acide phosphotungstique / phosphomolybdique pendant 5
minutes avant d'être colorées avec une solution de bleu d'Aniline. Enfin, les lames sont
placées dans de l'acide acétique à 1% puis déshydratées par un bain d'éthanol à 70% d'une
minute suivi d'un bain d'éthanol à 100% de 30 secondes. Les lames sont plongées dans du
toluène pendant 1 minute avant d'être montées avec du baume d'Eukitt® (Polylabo).
Réactif Temps
d'incubation Fonction
Bouin à 56°C 15min Mordant
Eau du robinet Rinçage constant
pendant 5min
Refroidissement des lames et rinçage de
l'excès de Boin
Hematoxyline de fer de
Weigert 5 min Coloration nucléaire
Eau du robinet Rinçage constant
pendant 5min Rinçage
Eau distillée 1 min Rinçage
Fuchsine Acide de
Biebrich Scarlet 5 min Coloration du cytoplasme et des muscles
Eau distillée 1 min Rinçage
Acide
phosphotungstique /
phosphomolybdique
5 min Mordant
Bleu d'Aniline 5 min Coloration du collagène
Acide acétique 1% 1 min Amélioration du rendu
Ethanol (70%) 1min Rinçage et déshydration
Ethanol (100%) 30s Rinçage et déshydration
Toluène 1min Fixation
Eukitt® - Fixation and montage de la lamelle
Tableau 9. Protocole de la coloration au trichrome de Masson.
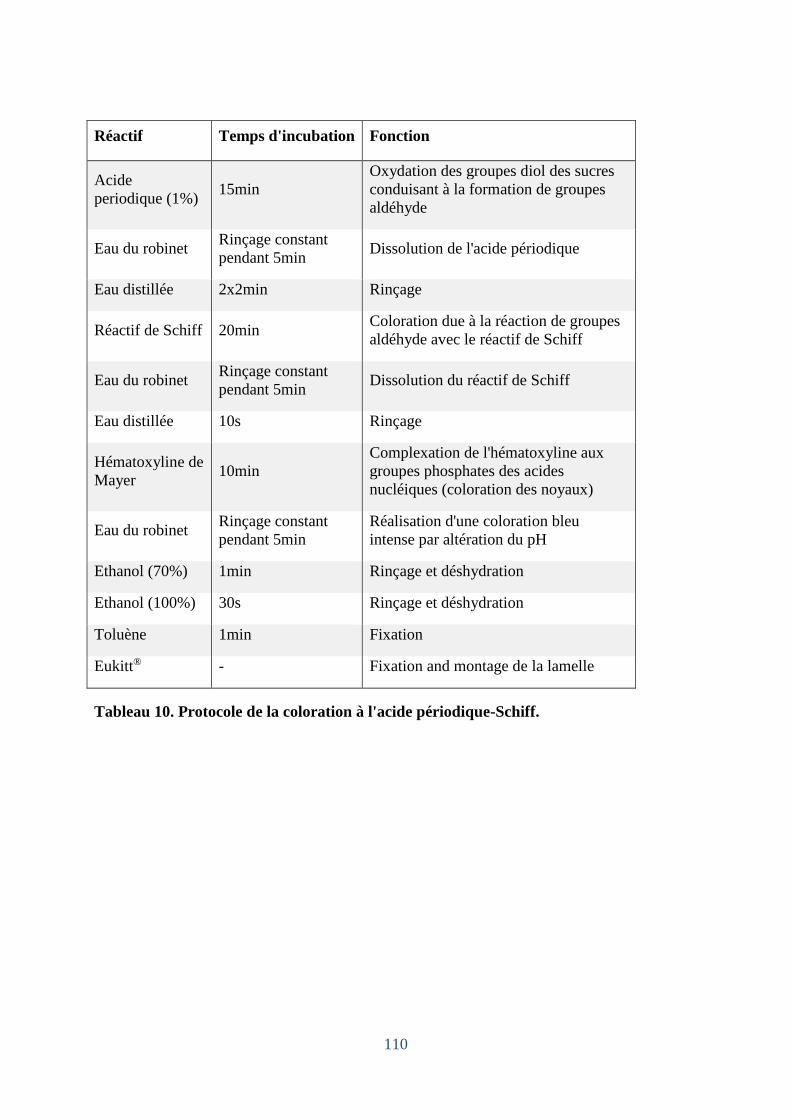
110
Réactif Temps d'incubation Fonction
Acide
periodique (1%) 15min
Oxydation des groupes diol des sucres
conduisant à la formation de groupes
aldéhyde
Eau du robinet Rinçage constant
pendant 5min Dissolution de l'acide périodique
Eau distillée 2x2min Rinçage
Réactif de Schiff 20min Coloration due à la réaction de groupes
aldéhyde avec le réactif de Schiff
Eau du robinet Rinçage constant
pendant 5min Dissolution du réactif de Schiff
Eau distillée 10s Rinçage
Hématoxyline de
Mayer 10min
Complexation de l'hématoxyline aux
groupes phosphates des acides
nucléiques (coloration des noyaux)
Eau du robinet Rinçage constant
pendant 5min
Réalisation d'une coloration bleu
intense par altération du pH
Ethanol (70%) 1min Rinçage et déshydration
Ethanol (100%) 30s Rinçage et déshydration
Toluène 1min Fixation
Eukitt® - Fixation and montage de la lamelle
Tableau 10. Protocole de la coloration à l'acide périodique-Schiff.

111
4.2.1 Coloration à l'Acide Periodique-Schiff (PAS)
Les sections sont colorées selon la méthode de l'acide periodique-Schiff (PAS). Cette
coloration permet de détecter le glycogène et d'autres polysaccharides dans les tissus. Cette
méthode permet de colorer les cellules caliciformes, qui sont des cellules épithéliales
glandulaires remplies de mucine (une glycoprotéine). Ces cellules sont responsables de la
production de mucus (mucine dissoute dans l'eau), caractéristique de l'inflammation
pulmonaire.
Les sections d'organes sont cerclées au Dakopen (DAKO) et traitées à l'acide
periodique à 1% pendant 15 minutes. Les lames sont ensuite rincées à l'eau du robinet pendant
15 minutes, puis par deux bains de 2 minutes d'eau distillée. Les lames sont ensuite colorées
au réactif de Schiff (VWR) pendant 20 minutes, rincées et colorées à l'hématoxyline de Mayer
pendant 10 minutes. Elles sont enfin rincées 5minutes dans de l'eau du robinet et 10 secondes
dans de l'eau distillée puis déshydratées par un bain d'éthanol à 70% d'une minute suivi d'un
bain d'éthanol à 100% de 30 secondes. Enfin les lames sont plongées dans du toluène pendant
1 minute avant d'être montées avec du baume d'Eukitt® (Polylabo).

112
4.2.2 Colorations immunohistologiques avec révélation enzymatique
Les lames Superfrost ont été déparaffinées et réhydratées comme pour les colorations
histologiques. Pour tous les marquages, le démasquage des antigènes est réalisé avec une
solution de Protéinase K (10µg/mL) en tampon Tris-EDTA (TE) (50mM Tris Base, 1mM
EDTA, 0.5% Triton X-100, pH 8) à 37°C pendant 10 minutes en chambre humide. Les lames
sont ensuite laissées à refroidir pendant 10 minutes à température ambiante. Les peroxydases
et les phosphatases alcalines endogènes sont bloquées pendant dix minutes par ajout de
100µL de la solution DualEndogenous Enzyme Block (DAKO) sur les coupes pendant 10
minutes.
4.2.3 Marquage du S100A9
Un blocage des avidines puis des biotines endogènes est réalisé successivement
pendant 15 minutes grâce au kit Avidin/Biotin Blocking kit (Vector Laboratories). Les étapes
suivantes sont réalisées avec le kit Vectastain ELITE ABC (Vector Laboratories). Les sites
non spécifiques sont bloqués avec du sérum de rat à 5% (Vector Laboratories ; diluant : PBS)
pendant 20min. Le surplus de sérum est retiré puis l'anticorps primaire de rat monoclonal anti-
S100A9 (clone MU14-2A5, Hycult biotech) dilué au 1/200 dans le sérum de rat à 5%-PBS)
est ajouté. Les lames sont mises à incuber durant 1h à température ambiante en chambre
humide. L'anticorps secondaire anti-rat couplé à une biotine (Vector Laboratories), puis la
streptavidine (qui va se fixer sur la biotine) couplé à l'enzyme HRP (ABC reagent, Vector
Laboratories), sont ensuite respectivement mis à incuber 30 minutes chacun à température
ambiante. Puis une goutte de EnVision+ Dual Link System-HRP (Dako) est ajoutée pendant 1
minutes à température ambiante. Le marquage donne une coloration rouge vif. L’AEC forme
un produit final qui est soluble dans les composés organiques. Il est donc nécessaire, d’utiliser
un contremarquage non alcoolique et un milieu de montage aqueux. Le contremarquage est
réalisé à l'Hemalun de Mayer pendant 1 minute avant montage à l'Aquatex (Merck).

113
Réactif Temps d'incubation Fonction
Avidin blocking solution 15min Blocage des Avidines endogènes
PBS 1 min Rinçage
Biotin blocking solution 15min Blocage des Biotines endogènes
PBS 2x2min Rinçage
Sérum de rat 5% 20min Blocage es sites non spécifiques
Anticorps primaire
(1/200)
1h en chambre
humide Fixation sur l'antigène d'intérêt
PBS 2x3min Rinçage
Anticorps secondaire
biotinylé
30min en chambre
humide Fixation sur l'anticorps primaire
PBS 2x3min Rinçage
Réactif ABC 30min en chambre
humide
Fixation du complexe avidine/peroxydase
sur la biotine de l'anticorps secondaire
PBS 2x3min Rinçage
AEC (substrat) 1min Oxydation du substrat par la peroxydase
Eau du robinet 2x1min Rinçage
Hématoxyline de Mayer 1min
Complexation de l'hématoxyline aux
groupes phosphates des acides nucléiques
(coloration des noyaux)
Eau du robinet 1min Rinçage
Aquatex - Montage et fixation
Tableau 11. Protocole de marquage de la protéine S100A9
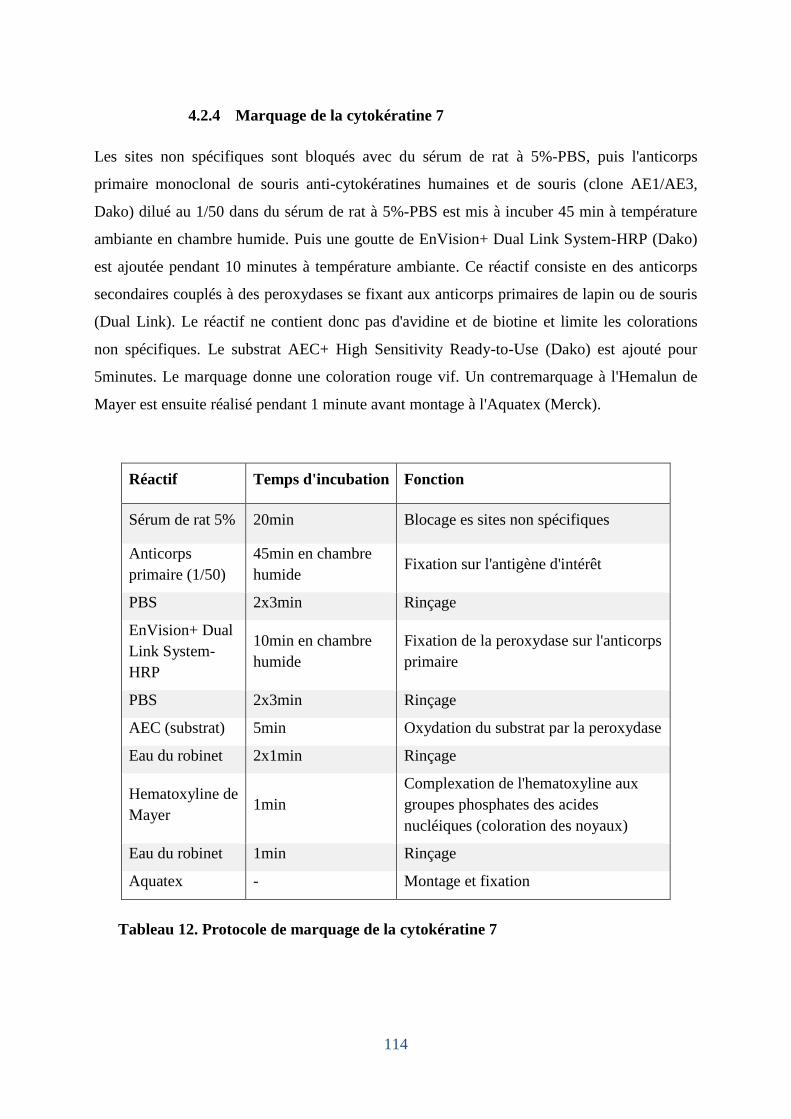
114
4.2.4 Marquage de la cytokératine 7
Les sites non spécifiques sont bloqués avec du sérum de rat à 5%-PBS, puis l'anticorps
primaire monoclonal de souris anti-cytokératines humaines et de souris (clone AE1/AE3,
Dako) dilué au 1/50 dans du sérum de rat à 5%-PBS est mis à incuber 45 min à température
ambiante en chambre humide. Puis une goutte de EnVision+ Dual Link System-HRP (Dako)
est ajoutée pendant 10 minutes à température ambiante. Ce réactif consiste en des anticorps
secondaires couplés à des peroxydases se fixant aux anticorps primaires de lapin ou de souris
(Dual Link). Le réactif ne contient donc pas d'avidine et de biotine et limite les colorations
non spécifiques. Le substrat AEC+ High Sensitivity Ready-to-Use (Dako) est ajouté pour
5minutes. Le marquage donne une coloration rouge vif. Un contremarquage à l'Hemalun de
Mayer est ensuite réalisé pendant 1 minute avant montage à l'Aquatex (Merck).
Réactif Temps d'incubation Fonction
Sérum de rat 5% 20min Blocage es sites non spécifiques
Anticorps
primaire (1/50)
45min en chambre
humide Fixation sur l'antigène d'intérêt
PBS 2x3min Rinçage
EnVision+ Dual
Link System-
HRP
10min en chambre
humide
Fixation de la peroxydase sur l'anticorps
primaire
PBS 2x3min Rinçage
AEC (substrat) 5min Oxydation du substrat par la peroxydase
Eau du robinet 2x1min Rinçage
Hematoxyline de
Mayer 1min
Complexation de l'hematoxyline aux
groupes phosphates des acides
nucléiques (coloration des noyaux)
Eau du robinet 1min Rinçage
Aquatex - Montage et fixation
Tableau 12. Protocole de marquage de la cytokératine 7
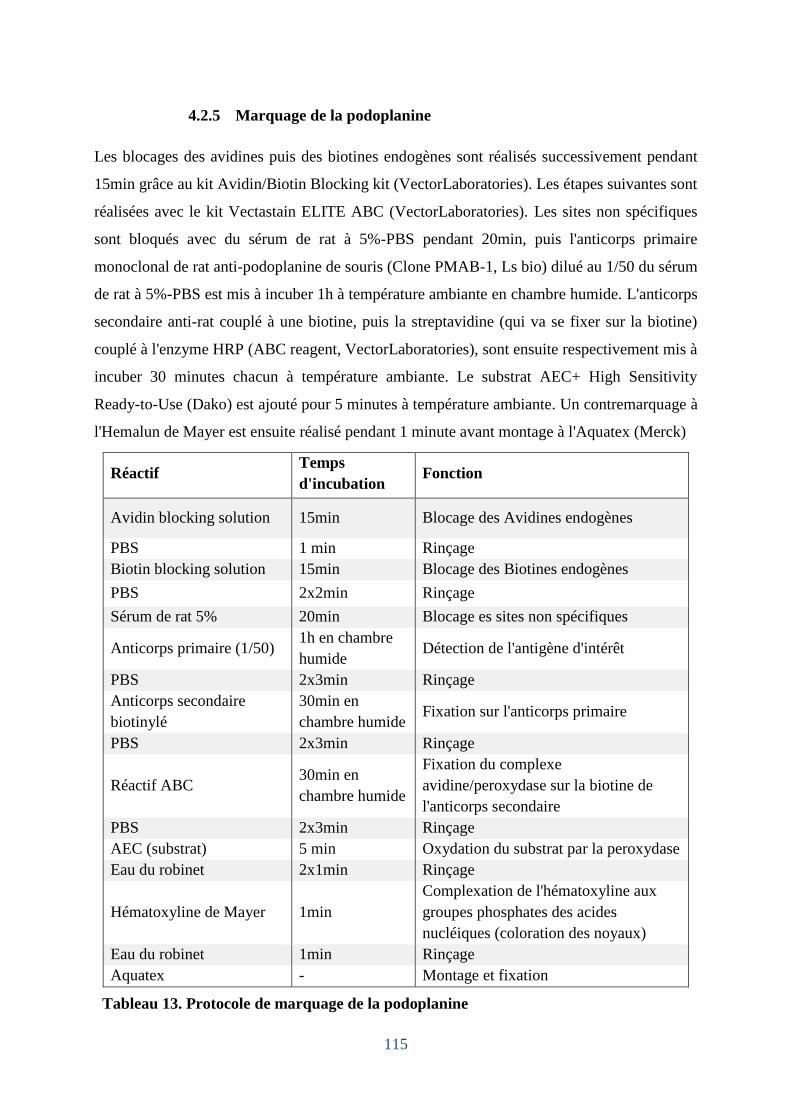
115
4.2.5 Marquage de la podoplanine
Les blocages des avidines puis des biotines endogènes sont réalisés successivement pendant
15min grâce au kit Avidin/Biotin Blocking kit (VectorLaboratories). Les étapes suivantes sont
réalisées avec le kit Vectastain ELITE ABC (VectorLaboratories). Les sites non spécifiques
sont bloqués avec du sérum de rat à 5%-PBS pendant 20min, puis l'anticorps primaire
monoclonal de rat anti-podoplanine de souris (Clone PMAB-1, Ls bio) dilué au 1/50 du sérum
de rat à 5%-PBS est mis à incuber 1h à température ambiante en chambre humide. L'anticorps
secondaire anti-rat couplé à une biotine, puis la streptavidine (qui va se fixer sur la biotine)
couplé à l'enzyme HRP (ABC reagent, VectorLaboratories), sont ensuite respectivement mis à
incuber 30 minutes chacun à température ambiante. Le substrat AEC+ High Sensitivity
Ready-to-Use (Dako) est ajouté pour 5 minutes à température ambiante. Un contremarquage à
l'Hemalun de Mayer est ensuite réalisé pendant 1 minute avant montage à l'Aquatex (Merck)
Réactif Temps
d'incubation Fonction
Avidin blocking solution 15min Blocage des Avidines endogènes
PBS 1 min Rinçage
Biotin blocking solution 15min Blocage des Biotines endogènes
PBS 2x2min Rinçage
Sérum de rat 5% 20min Blocage es sites non spécifiques
Anticorps primaire (1/50) 1h en chambre
humide Détection de l'antigène d'intérêt
PBS 2x3min Rinçage
Anticorps secondaire
biotinylé
30min en
chambre humide Fixation sur l'anticorps primaire
PBS 2x3min Rinçage
Réactif ABC 30min en
chambre humide
Fixation du complexe
avidine/peroxydase sur la biotine de
l'anticorps secondaire
PBS 2x3min Rinçage
AEC (substrat) 5 min Oxydation du substrat par la peroxydase
Eau du robinet 2x1min Rinçage
Hématoxyline de Mayer 1min
Complexation de l'hématoxyline aux
groupes phosphates des acides
nucléiques (coloration des noyaux)
Eau du robinet 1min Rinçage
Aquatex - Montage et fixation
Tableau 13. Protocole de marquage de la podoplanine
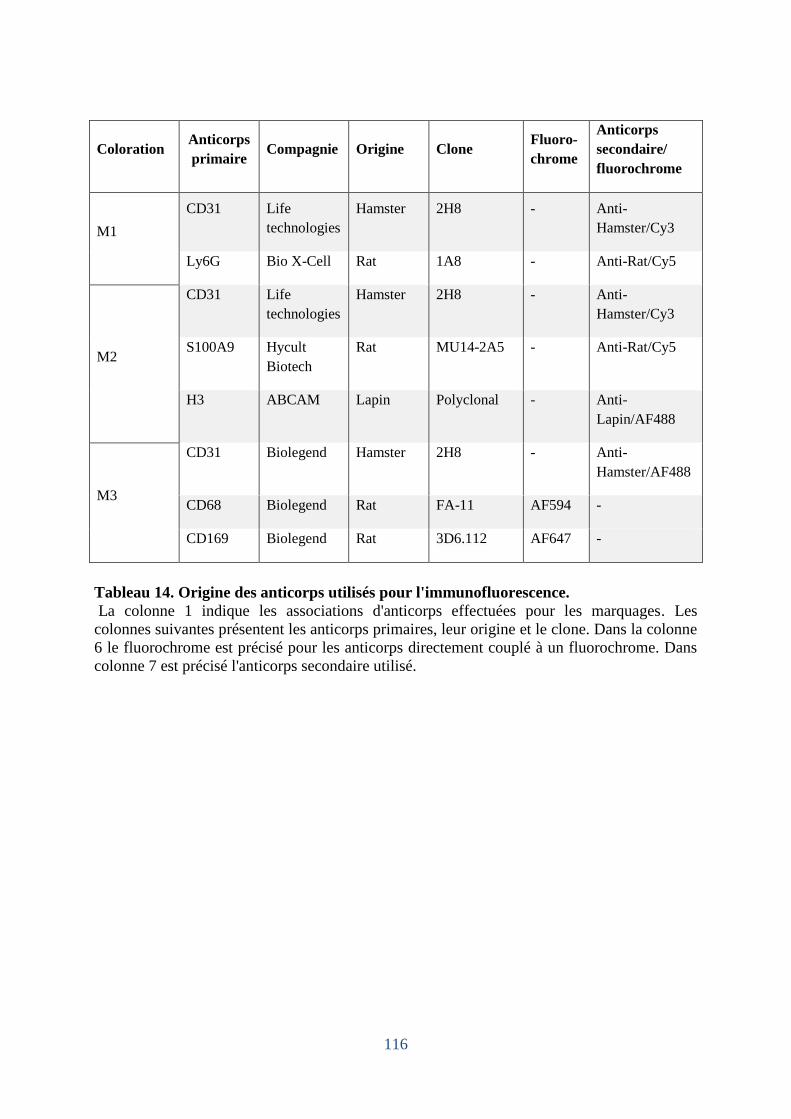
116
Coloration Anticorps
primaire Compagnie Origine Clone
Fluoro-
chrome
Anticorps
secondaire/
fluorochrome
M1
CD31 Life
technologies
Hamster 2H8 - Anti-
Hamster/Cy3
Ly6G Bio X-Cell Rat 1A8 - Anti-Rat/Cy5
M2
CD31 Life
technologies
Hamster 2H8 - Anti-
Hamster/Cy3
S100A9 Hycult
Biotech
Rat MU14-2A5 - Anti-Rat/Cy5
H3 ABCAM Lapin Polyclonal - Anti-
Lapin/AF488
M3
CD31 Biolegend Hamster 2H8 - Anti-
Hamster/AF488
CD68 Biolegend Rat FA-11 AF594 -
CD169 Biolegend Rat 3D6.112 AF647 -
Tableau 14. Origine des anticorps utilisés pour l'immunofluorescence.
La colonne 1 indique les associations d'anticorps effectuées pour les marquages. Les
colonnes suivantes présentent les anticorps primaires, leur origine et le clone. Dans la colonne
6 le fluorochrome est précisé pour les anticorps directement couplé à un fluorochrome. Dans
colonne 7 est précisé l'anticorps secondaire utilisé.

117
5. Immunohistochimie sur coupes épaisses
5.1 Préparation des coupes épaisses de poumons (Precision Cut Lung Slices ou
PCLS)
Les poumons gonflés à l’agarose (voir 3.6) fixés au paraformaldéhyde (PFA) et
stockés à 4°C dans du PBS sont préparés pour réaliser des coupes épaisses. Ces étapes ont été
effectuées dans l'équipe du Dr Léo Carlin (Impérial College de Londres, National Heart &
Lung Institute puis Beatson Institute, Leucocyte Dynamics, Glasgow).
Le poumon gauche est isolé et collé sur un bloc d'aluminium refroidi en utilisant de la
colle Superglue et le bloc est placé dans le vibratome (Microcut H1200 BioRad) avec la partie
supérieure du poumon (le hile) dirigée à l'opposé de la lame. En effet, les bronches principales
situées au niveau du hile sont plus rigides que le tissu environnant et rendent l'initiation de la
coupe plus difficile. Des coupes de 300 μm d'épaisseur sont préparées dans un bain de PBS /
BSA1% / Azide 0.05% refroidi. L'amplitude et la vitesse d'oscillation sont réglées au
maximum et l'avancée de la lame est d'1mm/sec.
5.2 Colorations immunohistologiques en fluorescence
Les coupes de chaque poumon sont perméabilisées et bloquées avec 300 μL de PBS /
NGS10% / BSA1% / TX-100 0,3% / Azide 0,05% (appelé "solution tampon" dans la suite)
dans un puits d’une plaque de 24 puits et placées sur un agitateur à température ambiante
pendant 30 minutes. Les coupes sont ensuite rincées une fois avec du PBS / BSA1% / TX-100
0,1% / Azide 0,05% puis 250 μL de solution d'anticorps primaires (dilution : 1/200 ; voir
Tableau 14 pour les différents marquages) dilué dans la solution tampon sont ajoutés pendant
3 heures sous agitation à température ambiante. Les coupes sont ensuite rincées avec la
solution tampon, puis 3 autres lavages de 2 minutes sont effectués sous agitation.
Les anticorps secondaires (dilution 1/200), les anticorps primaires directement couplé
à un fluorochrome (dilution 1/200) et le DAPI (dilution 1/1500) sont ajoutés sous 250 μL
pendant1 h à température ambiante sous agitation. Les coupes sont rincées 3 fois 2 minutes
avec du PBS / BSA1% / TX-100 0,1% / Azide 0,05% puis deux fois en PBS avant de finaliser
la fixation pendant 2 minutes dans de la PFA à 4% dans du PBS et rincer deux fois en PBS.

118

119
Les coupes sont placées sur les lames et recouvertes avec 150µL de Mowiol (Sigma-
Aldrich) additionné de 2,5% de DABCO puis recouvertes. Les lames ont été observées à
l'aide d'un microscope confocal (TCS-SP5, Leica à Londres ou Airyscann 880 ; Zeiss à
Glasgow).Des zones d'environ 1mm x1mm de côté par 100µm de profondeur contenant des
espaces périvasculaires ont été analyses. Au moins 3 images ont été obtenues pour chaque
poumon.
6. Analyse des images obtenues par microscopie confocale
Les images obtenues en microscopie confocale sont analysées grace aulogiciel
IMARIS (Bitplane). Les différents plans sont empilés pour obtenir une reconstitution en 3
dimensions des différents signaux de fluorescence. Le DAPI et l’anticorps anti-histone H3
(H3) permettent de visualiser le noyau des cellules. L’anticorps anti-CD31 colore les cellules
endothéliales et permet donc d'observer l'architecture générale des poumons (capillaires
pulmonaires, veines, artères et vaisseaux lymphatiques). Ly6G mais aussi S100A9 sont
utilisés pour observer les neutrophiles; les anticorps anti-CD68 et anti-CD169 reconnaissent
des macrophages.
Pour analyser l'espace péri-vasculaire, celui-ci est modélisé par un objet en 3 dimension
permettant d'obtenir son volume (en mm3). Ensuite le nombre de cellules présentes dans
l'espace (DAPI+ et DAPI+CD169+) est déterminé automatiquement grace aux algorithmes du
logiciel. En bref, les éléments dépassant un seuil de fluorescences et ayant une taille entre 10
et 20 µm sont retenus. Les comptages automatiques sont ensuite contrôlés manuellement. Le
nombre de cellules est ensuite normalisé par rapport au volume de l'espace (Figure 21).
<- Figure 21. Stratégie d'analyse du contenu cellulaire de l'espace périvasculaire.
Les images obtenues en microscopie confocale sont analysées grâce au logiciel IMARIS
(Bitplane). (A) Les différents plans sont empilés pour obtenir une reconstitution en 3
dimensions des différents signaux de fluorescence. Le DAPI (Bleu) permet de visualiser le
noyau des cellules. Le marquage du CD31 (vert) colore les cellules endothéliales et permet
donc d'observer l'architecture générale des poumons (capillaires pulmonaires, veines, artères
et vaisseaux lymphatiques). L'espace périvasculaire (PVS) contient un vaisseau sanguin et est
délimité par les bronchioles et les alvéoles. (B) Un objet 3D est crée manuellement pour
modéliser le PVS (fantôme blanc) permettant d'obtenir le volume V du PVS. (C) Le nombre
N de cellules (en rouge) est ensuite comptées par le logiciel (éléments mesurant entre 10 et 20
µm). Les cellules détectées apparaissent comme un point violet. Le nombre de cellules est
ensuite normalisé par rapport au volume du PVS, permettant d'obtenir la concentration de
cellules dans le PVS : N/V cellules/mm3.
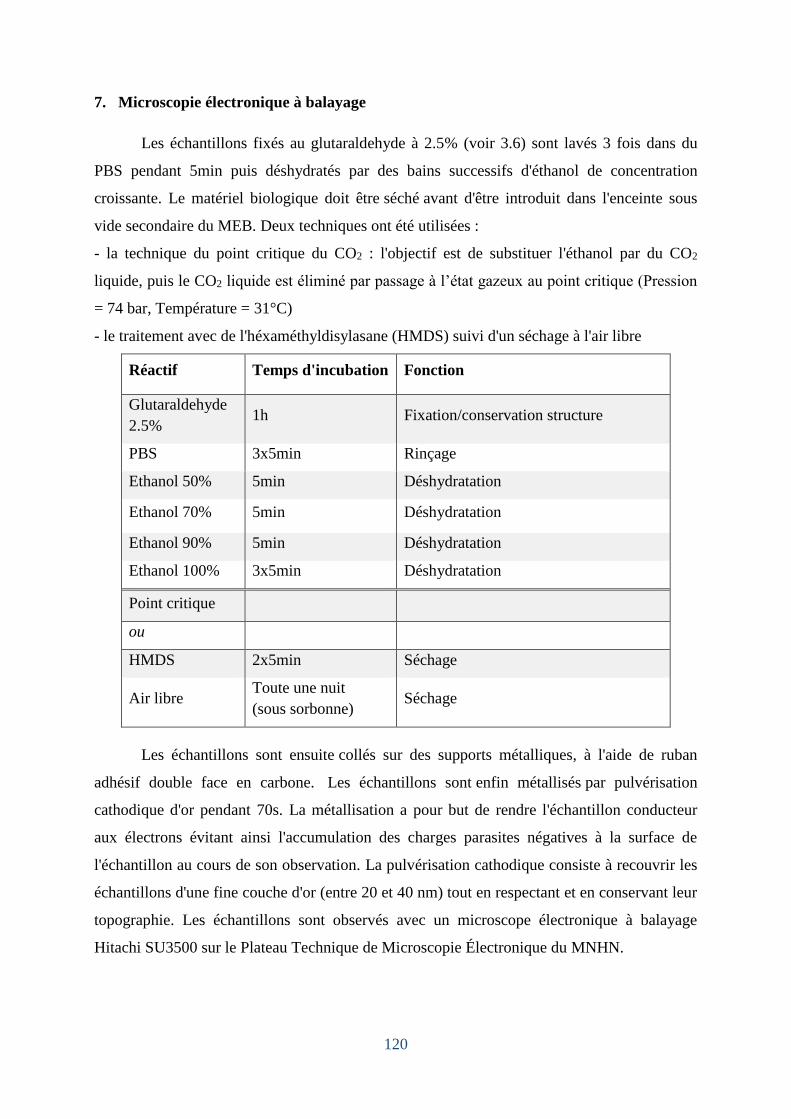
120
7. Microscopie électronique à balayage
Les échantillons fixés au glutaraldehyde à 2.5% (voir 3.6) sont lavés 3 fois dans du
PBS pendant 5min puis déshydratés par des bains successifs d'éthanol de concentration
croissante. Le matériel biologique doit être séché avant d'être introduit dans l'enceinte sous
vide secondaire du MEB. Deux techniques ont été utilisées :
- la technique du point critique du CO2 : l'objectif est de substituer l'éthanol par du CO2
liquide, puis le CO2 liquide est éliminé par passage à l’état gazeux au point critique (Pression
= 74 bar, Température = 31°C)
- le traitement avec de l'héxaméthyldisylasane (HMDS) suivi d'un séchage à l'air libre
Réactif Temps d'incubation Fonction
Glutaraldehyde
2.5% 1h Fixation/conservation structure
PBS 3x5min Rinçage
Ethanol 50% 5min Déshydratation
Ethanol 70% 5min Déshydratation
Ethanol 90% 5min Déshydratation
Ethanol 100% 3x5min Déshydratation
Point critique
ou
HMDS 2x5min Séchage
Air libre Toute une nuit
(sous sorbonne) Séchage
Les échantillons sont ensuite collés sur des supports métalliques, à l'aide de ruban
adhésif double face en carbone. Les échantillons sont enfin métallisés par pulvérisation
cathodique d'or pendant 70s. La métallisation a pour but de rendre l'échantillon conducteur
aux électrons évitant ainsi l'accumulation des charges parasites négatives à la surface de
l'échantillon au cours de son observation. La pulvérisation cathodique consiste à recouvrir les
échantillons d'une fine couche d'or (entre 20 et 40 nm) tout en respectant et en conservant leur
topographie. Les échantillons sont observés avec un microscope électronique à balayage
Hitachi SU3500 sur le Plateau Technique de Microscopie Électronique du MNHN.

121
8. Cytométrie en flux
8.1 Préparation des cellules
Les cellules isolées de la cavité pleurale et du liquide bronchoalvéolaire sont analysées
à l’aide d’un cytomètre en flux. Premièrement, les globules rouges présents dans les culots
cellulaires sont lysés par un choc hypotonique avec 5ml de NaCl 0.2% (puis équilibré avec
l’ajout de 5 ml de NaCl à 1.6%). Les suspensions cellulaires sont centrifugées à 400gpendant
4 min à 4°C, puis les culots cellulaires sont repris dans 1 ml de PBS contenant2% de SVF et
les cellules sont comptées en utilisant un hémocytomètre (KOVA® Glasstic Slide). Au moins
300 000 cellules sont utilisées pour le marquage des cellules pleurales. Pour les cellules du
liquide bronchoalvéolaire, leur quantité étant faible, la totalité des cellules est utilisée pour les
différents marquages. Les cellules sont lavées une fois par ajout de PBS-SVF 2% puis
centrifugées à 400g pendant 4 min à 4°C et le surnageant est éliminé (ces 3 étapes seront
appelées "lavage" par la suite). Les récepteurs aux régions Fc des anticorps sont ensuite
bloqués en incubant à 4°C les cellules pendant 15 minutes avec un anticorps anti-CD16/CD32
(dilution : 1/1000, eBiosciences) pour éviter la fixation non spécifique des anticorps
primaires. Après un lavage, les cellules sont prêtes pour les différents marquages.
8.2 Identification des populations leucocytaires.
Différents marquages avec des anticorps monoclonaux couplés à des fluorochromes
sont effectués pour identifier les populations de leucocytes. Les différentes populations sont
identifiées en utilisant les anticorps de rat anti-souris suivants: anti-F4/80-APC (dilution
1/200, eBioscience, clone BM8), anti-SiglecF-PE (dilution 1/200, BD Biosciences, clone E50-
2440), Ly6G-FITC (dilution 1/200, BD Biosciences, clone 1A8), CD3-PE (dilution 1/200, BD
Biosciences, clone 17A2), CD4-FITC (dilution 1/200, BD Biosciences, clone RM4-5), CD8-
APC/Cy5 (dilution : 1/200, BD Biosciences, clone 53-6.8 ), CD19-APC (dilution : 1/200, BD
Biosciences, clone 1D3) (Tableau 15). Un marquage intracellulaire avec anticorps primaire
polyclonal de lapin anti-Relm-α (1/400) suivi d'un anticorps secondaire couplé à de l'AF488
se fixant sur les anticorps de lapin (1/1000) a été effectué pour identifier les macrophages
alternativement activés (AAM) (Tableau 15). L'acquisition a été réalisée avec un cytomètre en
flux FACSVerse (BD Biosciences) et le logiciel FACSuite (BD Biosciences). Les doublets et
les débris sont exclus (Figure 22). Les analyses ont été effectuées avec le logiciel FACSuite.
La figure 22 présente la stratégie de discrimination des populations cellulaires mise en place
pour le marquage 1.
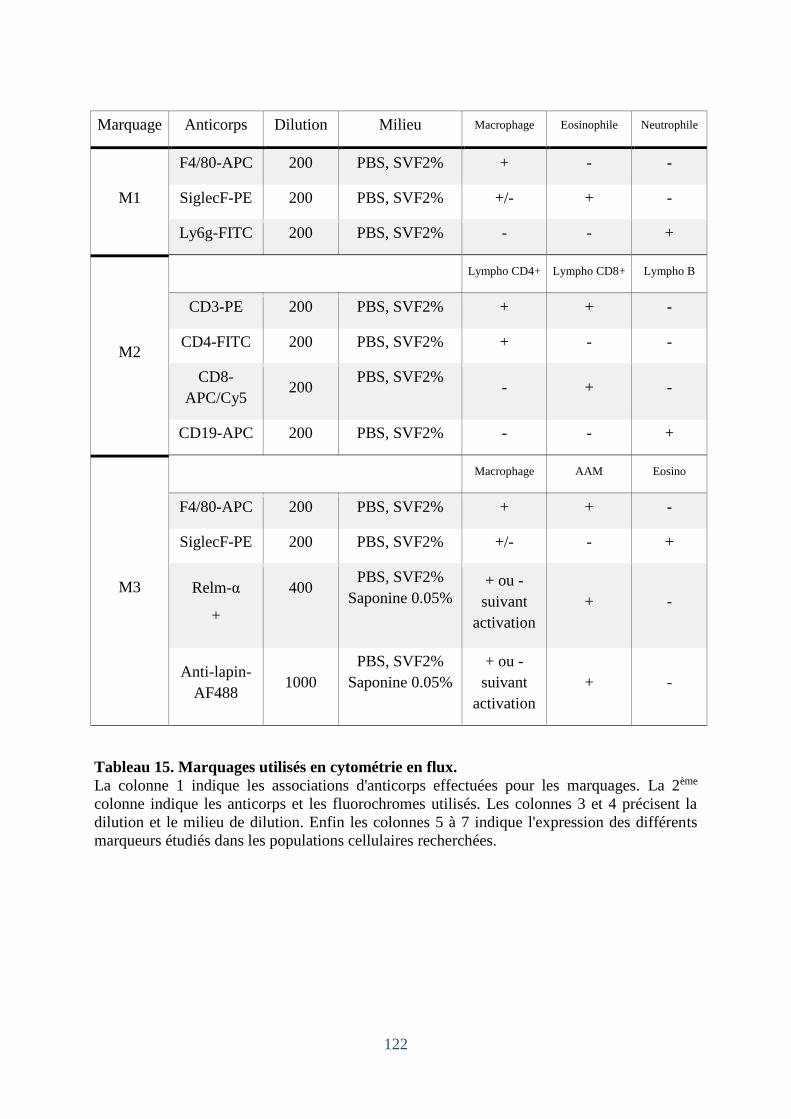
122
Marquage Anticorps Dilution Milieu Macrophage Eosinophile Neutrophile
M1
F4/80-APC 200 PBS, SVF2% + - -
SiglecF-PE 200 PBS, SVF2% +/- + -
Ly6g-FITC 200 PBS, SVF2% - - +
M2
Lympho CD4+ Lympho CD8+ Lympho B
CD3-PE 200 PBS, SVF2% + + -
CD4-FITC 200 PBS, SVF2% + - -
CD8-
APC/Cy5 200
PBS, SVF2% - + -
CD19-APC 200 PBS, SVF2% - - +
M3
Macrophage AAM Eosino
F4/80-APC 200 PBS, SVF2% + + -
SiglecF-PE 200 PBS, SVF2% +/- - +
Relm-α
+
400
PBS, SVF2%
Saponine 0.05% + ou -
suivant
activation
+ -
Anti-lapin-
AF488 1000
PBS, SVF2%
Saponine 0.05%
+ ou -
suivant
activation
+ -
Tableau 15. Marquages utilisés en cytométrie en flux.
La colonne 1 indique les associations d'anticorps effectuées pour les marquages. La 2ème
colonne indique les anticorps et les fluorochromes utilisés. Les colonnes 3 et 4 précisent la
dilution et le milieu de dilution. Enfin les colonnes 5 à 7 indique l'expression des différents
marqueurs étudiés dans les populations cellulaires recherchées.
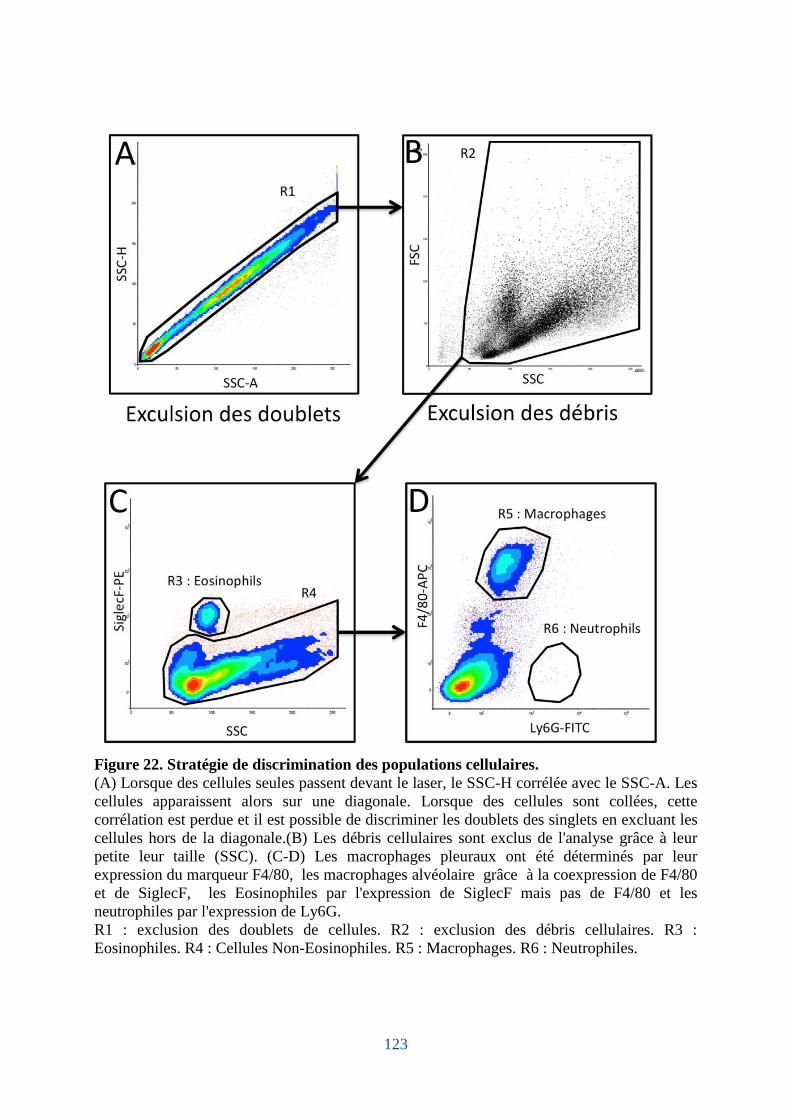
123
Figure 22. Stratégie de discrimination des populations cellulaires.
(A) Lorsque des cellules seules passent devant le laser, le SSC-H corrélée avec le SSC-A. Les
cellules apparaissent alors sur une diagonale. Lorsque des cellules sont collées, cette
corrélation est perdue et il est possible de discriminer les doublets des singlets en excluant les
cellules hors de la diagonale.(B) Les débris cellulaires sont exclus de l'analyse grâce à leur
petite leur taille (SSC). (C-D) Les macrophages pleuraux ont été déterminés par leur
expression du marqueur F4/80, les macrophages alvéolaire grâce à la coexpression de F4/80
et de SiglecF, les Eosinophiles par l'expression de SiglecF mais pas de F4/80 et les
neutrophiles par l'expression de Ly6G.
R1 : exclusion des doublets de cellules. R2 : exclusion des débris cellulaires. R3 :
Eosinophiles. R4 : Cellules Non-Eosinophiles. R5 : Macrophages. R6 : Neutrophiles.

124
9. Analyse des transcrits de cytokines inflammatoires dans le poumon
9.1 Extraction des ARN totaux et rétrotranscription
Les poumons de souris prélevés 4 jours et 70 jours après infection et ceux des souris
contrôle non infestées (8 souris par groupe) ont été préservé en RNA later à -80°C.
L’extraction d’ARN est réalisée sur colonne (kit RNeasy Midi kit; Qiagen). Les poumons sont
placés encore congelés dans un microtube de 2mL contenant 3 billes de tungstène de 5mm de
diamètre et 300µL de tampon de lyse du kit (tampon RLT). Pour éviter toute contamination,
les billes sont préalablement nettoyées avec du RNase Away (VWR) et les tubes sont passés
10 minutes aux UV. Les poumons sont dissociés à l’aide du broyeur à billes Tissue Lyser II
(Qiagen) en effectuant 2 cycles de 1 min à 20 Hz. Le tube est brièvement centrifugé puis le
broyat est transféré dans un tube de 15mL. 4mL de tampon de lyse sont ajouté et le mélange
est vortexé 45-60sec avant de centrifuger 10min à 4000g. Le surnageant est transféré dans un
nouveau tube de 15mL et 4mL d'éthanol 70% sont rajoutés et le tube est secoué pour
resuspendre les précipités. L'échantillon est alors transféré dans une colonne du kit RNeasy
Midi kit (Qiagen) et centrifugé pendant 5 minutes à 4000g. Le liquide du tube collecteur est
jeté et 4mL de tampon RW1 (kit Qiagen) sont ajoutés pour laver la colonne; une nouvelle
centrifugation de 5 minutes (4000g) est réalisée. Puis deux lavages avec 2,5ml de tampon
RPE (Qiagen) sont effectués. Enfin, 250μl d’eau RNase-free sont ajoutés pour éluer l’ARN.
Cette dernière étape est répétée avec un nouveau volume d’eau RNase-free (250μl). Les ARN
sont conservés à -80°C.
La quantité et la qualité des ARN obtenus est vérifiée avec un spectrophotomètre à
spectre complet (Nanodrop2000, Thermo Scientific) et un bioanalyseur Agilent 2100.
Théoriquement le Nanodrop permet d'estimer la quantité d'ADN ou d'ARN (absorbance de
l'échantillon à 260nm) et le rapport des absorbances à 260 nm et 280 nm est utilisé pour
évaluer la pureté de l'ADN et de l'ARN. Un ratio de ~ 1,8 est généralement accepté comme
"pur" pour l’ADN ; Un ratio de ~ 2,0 est généralement accepté comme "pur" pour l'ARN. Si
le rapport est sensiblement inférieur dans les deux cas, il peut indiquer la présence de
protéines, de phénols ou d'autres contaminants qui absorbent fortement à près de 280 nm.
Pour les très petites quantités il est connu que le Nanodrop peut surévaluer la concentration.
Pour la qualité des ARN le bioanalyseur Agilent effectue une électrophorèse en
capillaires ce qui permet de contrôler l’intégrité de des ARN totaux. Un facteur d’intégrité ou

125
RIN (RNA Integrity Number) est attribué à chaque échantillon. Les échantillons présentant un
RIN< 8 (dégradés) ont été exclus.
Pour chaque condition, la retrotranscription est effectuée sur un pool de 6-8
échantillons (suivant leur qualité) en mettant la même quantité d'ARN pour chaque
échantillon. Les ADN complémentaires (ADNc) sont produits avec le kit First-strand cDNA
(Qiagen). Tout d'abord, les contaminations éventuelles avec de l'ADN génomique sont
éliminées avec le mix suivant.
Produit Volume (µL) pour 1 échantillon
ARN 0, 5µg
Tampon GE 2 µL
Eau RNase-free Variable
Volume total 10 µL
Le mélange est mis à incuber pendant 5min à 42°C dans un thermocycleur (Mastercycler®
nexus, Eppendorf) puis placé sur glace au moins 1min pour stopper la réaction. La
retrotranscription est effectuée en ajoutant 10 µL du mix suivant dans les tubes traités à la
DNase, avant d'incuber à 42°C pendant 15min exactement et d'inactiver la réaction en
incubant 5min à 95°C
Produit Volume (µL) pour 1 échantillon
Tampon BC3 5x 4µL
Contrôle P2 2 µL
Mix de Reverse Transcriptase RE3 2µL
Eau RNase-free 3µL
91µL d'eau RNase-free est ensuite ajouté à chaque échantillon qui sont alors prêt pour les
PCR en temps réel.
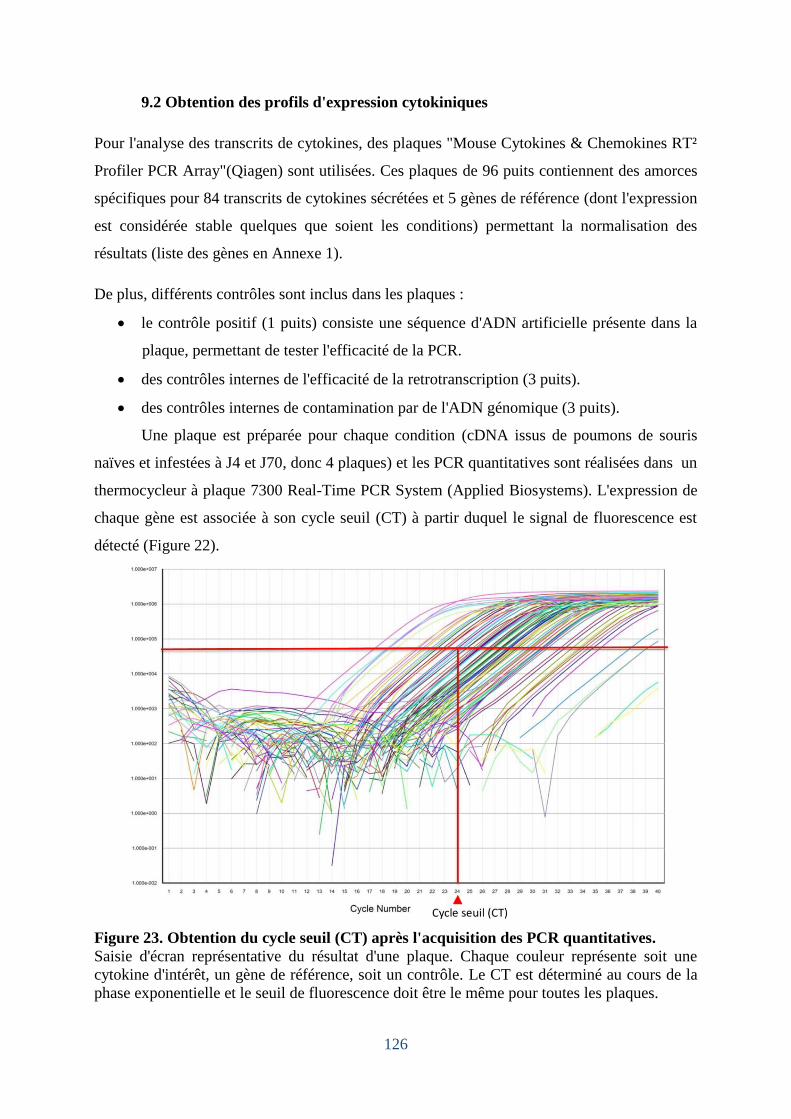
126
9.2 Obtention des profils d'expression cytokiniques
Pour l'analyse des transcrits de cytokines, des plaques "Mouse Cytokines & Chemokines RT²
Profiler PCR Array"(Qiagen) sont utilisées. Ces plaques de 96 puits contiennent des amorces
spécifiques pour 84 transcrits de cytokines sécrétées et 5 gènes de référence (dont l'expression
est considérée stable quelques que soient les conditions) permettant la normalisation des
résultats (liste des gènes en Annexe 1).
De plus, différents contrôles sont inclus dans les plaques :
• le contrôle positif (1 puits) consiste une séquence d'ADN artificielle présente dans la
plaque, permettant de tester l'efficacité de la PCR.
• des contrôles internes de l'efficacité de la retrotranscription (3 puits).
• des contrôles internes de contamination par de l'ADN génomique (3 puits).
Une plaque est préparée pour chaque condition (cDNA issus de poumons de souris
naïves et infestées à J4 et J70, donc 4 plaques) et les PCR quantitatives sont réalisées dans un
thermocycleur à plaque 7300 Real-Time PCR System (Applied Biosystems). L'expression de
chaque gène est associée à son cycle seuil (CT) à partir duquel le signal de fluorescence est
détecté (Figure 22).
Figure 23. Obtention du cycle seuil (CT) après l'acquisition des PCR quantitatives.
Saisie d'écran représentative du résultat d'une plaque. Chaque couleur représente soit une
cytokine d'intérêt, un gène de référence, soit un contrôle. Le CT est déterminé au cours de la
phase exponentielle et le seuil de fluorescence doit être le même pour toutes les plaques.

127
Les données obtenues pour chaque plaque sont traitées en utilisant le logiciel en ligne
"RT2 Profiler PCR Array Data Analysis 3.5" sur le site Web sabiociences.com (Qiagen).
Après vérification des contrôles, l'expression des gènes a été normalisée avec les 5 gènes de
référence (Actb, B2m, Gapdh, Gusb, Hsp90ab1), permettant d'obtenir le ΔCT :
ΔCT (gène X) = CT(gène X)−CT(gènes de ménage)
L'expression relative de chaque gène est ensuite déterminée grâce la méthode des ∆∆Ct :
2-ΔΔCT= 2 –(ΔCT(infesté)−ΔCT(naïf))
Les résultats sont exprimés par un rapport cible/contrôle. Ceci permet d'obtenir
l'augmentation ou la baisse relative (« Foldchange » ou FC) d'expression des différents gènes
entre deux conditions données, ici entre les poumons de souris naïves et ceux de souris
infestées. Un seuil (« threeshold ») d'augmentation/diminution de 90% a été placé (FC ≥ 1.9)
pour la suite des analyses.
9.3 Analyse in silico des réseaux biologiques
Les données transcriptionnelles ont été évaluées à l'aide du logiciel IPA (Ingenuity
Pathway Analysis, Systems Inc., États-Unis). IPA contient une base de données conséquente
(Ingenuity Knowledge Base) permettant de modéliser et d’analyser des systèmes biologiques
et chimiques complexes. Plus particulièrement, l'option « Core Analysis » est utilisée pour
interpréter des jeux de données dans le contexte de processus biologiques, de voies
métaboliques et de réseaux moléculaires. Les algorithmes d'IPA identifient les fonctions
biologiques impliquées dans les changements d'expression génique observés dans les données
analysées en les comparant à la littérature scientifique existante. A partir de cela, les
algorithmes peuvent aussi proposer des mécanismes et des molécules régulatrices pouvant
expliquer les résultats observés.
D'un point de vue pratique, le logiciel calcule un score d'activation (z-score) pour
chaque processus/réseau biologique en fonction des profils d'expressions géniques entrés et de
leur adéquation avec les données enregistrées dans les bases de données. Si le z-score est
supérieur à 2, le logiciel prédit que le processus est probablement activé, s'il est inférieur à - 2,
il serait inhibé.

128
9.4 Validation des résultats de l'analyse transcriptionelle
9.4.1 Retrotranscription
Les résultats des plaques étant obtenus à partir de pools d'échantillons, il est nécessaire
de valider ces résultats pour chaque individu. Pour cela, les ADN complémentaires (ADNc)
sont préparés pour chaque échantillon comme suit :
Les ARN ont été traités à la DNase (Turbo DNase, Ambion) à 37°C pendant 30min pour
éviter toute contamination par de l'ADN génomique en utilisant le mélange réactionnel
suivant :
Produit Volume (µL) pour 1 échantillon
Eau nucléase-free 19
10X turbo DNase Buffer (Ambion) 5
Turbo DNase (Ambion) 1
ARN 25
Ensuite, la réaction est stoppée en ajoutant 5µL de solution d'inactivation (Ambion).
L'échantillon est centrifugé (10000g, 1min30) et le surnageant est placé dans un nouveau tube
de 1ml.
4µL d'ARN traité à la DNase sont ensuite utilisés pour effectuer la retrotranscription des ARN
en ADNc. Le reste des ARN traités est congelé à -80°C.
Produit Volume (µL) pour 1 échantillon
Eau nucléase-free 6
OligodT (0.8µg/µL) (Roche)* 2
dNTP (10mM) 1
ARN traité à la DNase 4
Le mélange réactionnel est mis à incuber 5min à 65°C dans un thermocycleur (Mastercycler®
nexus, Eppendorf) pour permettre l'appariement spécifique des amorces sur les ARN puis
récupéré sur glace au moins une minute. Après une brève centrifugation, l’enzyme
Superscript III (Invitrogen) permettant la rétrotranscription est ajoutée :
Produit Volume (µL) pour 1 échantillon
5X buffer 4
0.1M DTT 1
RNase inhibitor (30U/µL) 1
Superscript III RT 1
Les tubes sont placés dans le thermocycleur à 50°C pendant 45min puis 15min à 70°C
(inactivation de l'enzyme). Enfin 40µL d'eau nucléase-free sont ajoutés avant dosage
(Nanodrop).

129
9.4.2 Conception des amorces
Les résultats obtenus à J4 p.i. ont été validés avec un gène candidat (CXCL1) et ceux à J70
avec 3 gènes (CXCL9, CCL2 et IL-13). Un ou deux gènes de référence ont été utilisés
(Actine-β et GAPDH).
Les amorces ont été conçues avec le logiciel AmplifX avec les conditions suivantes:
les amorces encadrent un intron afin de détecter les contaminations par de l'ADN génomique ;
les amorces mesurent entre 20 et 24 paires de bases (pb), température d'hybridation de
60±5°C, % GC entre 40 et 60%, produit de PCR entre 100 et 200pb ; les amorces ne doivent
pas d'hybrider entre elles ou former des boucles in silico (une double vérification est effectuée
en utilisant l'outil en ligne OligoCalc: http://biotools.nubic.northwestern.edu/OligoCalc.html);
les amorces ne doivent pas s'apparier à des séquences existant chez des filaires. Pour cela, les
séquences ont été comparées aux génomes des bases de données du NCBI
(https://blast.ncbi.nlm.nih.gov/Blast.cgi)
Amorce Séquence
IL 13 sens GGATATTGCATGGCCTCTGTAACC
IL 13 antisens GTGGCGAAACAGTTGCTTTGTG
CXCL9 sens CCATGAAGTCCGCTGTTCTTTTCC
CXCL9 antisens TGGGGCAAACTGTTTGAGGTCT
CCL2 sens ACTGCATCTGCCCTAAGGTCTTCA
CCL2 antisens TAAGGCATCACAGTCCGAGTCACA
ACTINE sens ATTGCTGACAGGATGCAGAAG
ACTINE antisens AGTCCGCCTAGAAGCACTTG
GAPDH sens ACTCTTCCACCTTCGATGCCG
GAPDH antisens ACCCTGTTGCTGTAGCCGTA
CXCL1 sens CACTGCACCCAAACCGAAGTCATA
CXCL1 antisens TCTCCGTTACTTGGGGACACCTTT
Tableau 16. Séquences des amorces utilisées pour les PCR quantitatives

130
9.4.3 PCR quantitative
Les PCR quantitatives sont effectuées dans un thermocycleurLightCycler2.0 (Roche
Diagnostics, France).
Pour CXCL1, CXCL9, CCL2, l’Actine et la GAPDH, le kit MasterPlus SYBR (Roche
Diagnostics, France) a été utilisé et le mélange réactionnel de 10 µL contient :
Produit Volume (µL) pour 1 échantillon
Eau nucléase-free 4 µL
Amorces sens (5mM) 1 µL
Amorces antisens (5mM) 1 µL
Solution A+B 2 µL
ADNc 2 µL
Pour l’IL-13, le kit Master SYBR Green (Roche Diagnostics, France) a été utilisé et le
mélange réactionnel de 10 µL contient :
Produit Volume (µL) pour 1 échantillon
Eau nucléase-free 1.6 µL
MgCl2 (10mM) 0,4 µL
Amorces sens (5mM) 1 µL
Amorces antisens (5mM) 1 µL
Solution 10X 1 µL
ADNc 4 µL
Le programme d'amplification comprend une incubation initiale de 10 min à 95°C suivie de
40 cycles composés de :
- une étape de dénaturation : 10 sec 95°C
- une étape d'appariement des amorces de 8 sec à 60°C
- une étape d'amplification de 10sec à 72°C, au cours desquelles les données de fluorescence
sont recueillies.
Ce programme a été suivi d'une étape de fusion permettant d'obtenir la température de fusion
(Tm) des amplicons.
L'expression de CXCL1 a été déterminée par rapport à l'Actine-β et celle de CXCL9, CCL2 et
IL-13 par rapport à l'Actine- β et au GAPDH en utilisant la méthode 2-ΔΔCT.

131
10. Purification des microfilaires à partir de sang de mérion hôtes de L. sigmodontis
Le sang de mérions microfilarémiques est prélevé au sinus rétro-orbital et déposé dans
un tube contenant de l'héparine pour éviter la coagulation du sang (ratio : 1 µL d'héparine
pour 10µL de sang). Le sang est mélangé avec un volume égal de RPMI 1640. Les
microfilaires sont ensuite purifiées sur gradient de Percoll (Densité : 1,130, pH=9; Sigma
Chemical CO, St Louis, USA). Le Percoll est composé de particules de silice de 15-30 nm de
diamètre, recouvertes de polyvinylpyrrolidone et les gradients de Percoll permettent de
séparer les cellules en fonction de leur densité.
- Une solution de Percoll "100%" est préparée en mélangeant 9 volumes de Percoll pur
avec un volume de sucrose à 2,5M en RPMI 1640.
- Ensuite du Percoll "25%" est préparé en mélangeant 1 volume de Percoll« 100% »
avec 3 volumes de sucrose 0.25M en RPMI 1640.
4mL de Percoll « 25% » sont transférés dans un tube de 15 mL puis 4mL du mélange
Sang/RPMI est déposé délicatement et sans à-coup au dessus. Le tube est centrifugé 35min à
400g à température ambiante et sans frein. L'aspect des tubes avant et après centrifugation est
indiqué en figure 23.
Figure 24. Purification des microfilaires sur gradient de Percoll.
Représentation de l'aspect des tubes avant et après centrifugation avec la position de l'anneau
de Mfs.
La phase contenant le RPMI et le sérum est éliminée à l’aide d’une pipette pasteur en
plastique, puis l'anneau de microfilaires (Mfs) est récupéré délicatement dans un nouveau tube
et dilué avec du RPMI pour réaliser un lavage. Le tube est centrifugé 10 min à 400 g, le
surnageant est éliminé et le culot est resuspendu dans 1mL de RPMI. Le nombre de
microfilaires est déterminé à l’aide d’un hémocytomètre (KOVA® Glasstic Slide).

132
11. Détection et quantification des microfilaires pulmonaires par PCR quantitative
Pour estimer le nombre de microfilaires pulmonaires, une approche de détection par
PCR quantitative a été mise en place. Pour ce faire une gamme standardisée d’ADN extraits
de poumons de souris non infestées mises avec des quantités connues de microfilaires a été
réalisée. Dans un deuxième temps cette gamme a été utilisé pour déterminer le ratio
Mfs/poumons dans les ADN extraits de poumons de souris infestées microfilarémiques ou
non.
11.1 Extraction de l'ADN des poumons
Les poumons gonflés et fixés à l'éthanol à 70° (voir 2.8) sont rincés en PBS. Les
éventuels morceaux de filaires adultes et les granulomes adhérant à la plèvre viscérale
pulmonaire sont retirés. Les poumons sont placés dans un microtube de 2mL contenant 3
billes de tungstène de 5mm de diamètre et 500µL de PBS stérile. Pour éviter toute
contamination, les billes sont préalablement nettoyées avec du RNase Away (VWR - cette
solution élimine les contaminations par des RNases et de l'ADN) et les tubes sont passés
10minutes aux UV. Les poumons sont dissociés à l’aide du broyeur à bille Tissue Lyser II
(Qiagen) en effectuant 2 cycles de 1 min à 20 Hz. Les tubes sont brièvement centrifugés puis
100µL de la solution sont transférés dans un microtube de 1.5mL contenant 180µL de tampon
de lyse (Buffer ATL, Qiagen) et 20µL de protéinase K (Qiagen). Le mélange est vortexé
brièvement et mis à incuber toute une nuit à 56°C avec une agitation régulière. L'ADN est
ensuite purifié en utilisant le kit Qiagen DNA microkit en suivant les spécifications du
fabricant : 200µL de tampon de lyse (Tampon AL) sont ajoutés dans le tube avant de vortexer
par à-coup pendant 15s jusqu'à obtenir une solution homogène. 200µL d'éthanol absolu sont
alors ajoutés et le tube est vortexé à nouveau et mis à incuber 5min à température ambiante.
Le contenu du tube est transféré dans une colonne QiAamp MinElute et centrifugé pendant
1min (6000g). Deux lavages de la membrane de la colonne sont réalisés successivement en
ajoutant 500µL de tampon AW1 puis AW2 avant de centrifuger le tube pendant 1min
(6000g). La membrane est ensuite séchée en centrifugeant pendant 3min (2000g). Enfin, la
colonne est placée sur un nouveau tube collecteur et 75μl d’eau DNAse-free sont ajoutés pour
éluer l’ADN. Cette dernière étape est répétée avec un nouveau volume d’eau DNAse-free
(75μl). Les ADN sont conservés à -20°C.

133
11.2 Préparation de la gamme
Les poumons de dix souris naïves sont récupérés et broyés comme précédemment. Les
broyats sont mélangés puis répartis dans dix microtubes de 2mL pour homogénéiser la
quantité de matériel entre les échantillons. 4 millions de microfilaires sont ensuite broyées de
la même façon sous 500µL de PBS. Une gamme de dilution du broyat de microfilaires est
ensuite réalisée par dilutions successives et ajoutée sous 100µL aux tubes contenant les
broyats de poumons.
Quantité de Mfs/ tube
1000000 100000 10000 1000 500 250 125 0
De ces mélanges poumons-microfilaires 100µL sont transférés dans un microtube de 1.5mL
contenant 180µL de tampon de lyse (Buffer ATL, Qiagen) et 20µL de proteinase K. Le
mélange est vortexé brièvement et mis à incuber toute une nuit à 56°C avec une agitation
régulière. L'ADN est ensuite purifié en utilisant le kit Qiagen DNA microkit en suivant les
spécifications du fabricant (voir 11.1).
11.3 Détection des microfilaires et normalisation
Des PCR quantitatives sont effectuées sur chaque échantillon d'ADN en utilisant le kit
MasterPlus SYBR Green (Roche Diagnostics, France) dans un thermocycleur LightCycler 2.0
(Roche Diagnostics, France).
L'ADN filarien est détecté grâce à des amorces ciblant l'actine de L. sigmodontis et l'ADN
murin avec des amorces ciblant l'actine de Mus musculus.
L.s Actin sens ATCCAAGCTGTCCTGTCTCT
L.s Actin anti-sens TGAGAATTGATTTGAGCTAATG
M. m Actin S ATTGCTGACAGGATGCAGAAG
M. m Actin AS AGTCCGCCTAGAAGCACTTG
Le mélange réactionnel de 10 µL contient :
Produit Volume (µL) pour 1 échantillon
Eau nucléase-free 2 µL
Amorcessens (5mM) 1 µL
Amorcesantisens (5mM) 1 µL
Solution A+B 2 µL
ADNc 4 µL
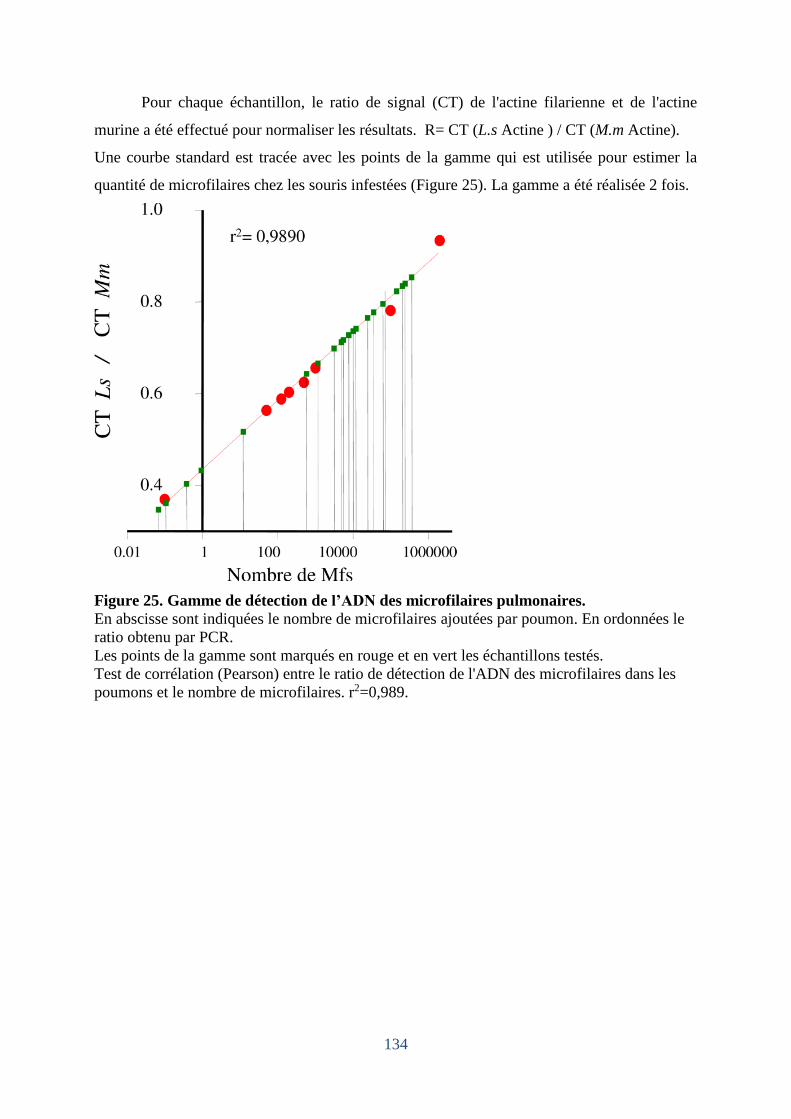
134
Pour chaque échantillon, le ratio de signal (CT) de l'actine filarienne et de l'actine
murine a été effectué pour normaliser les résultats. R= CT (L.s Actine ) / CT (M.m Actine).
Une courbe standard est tracée avec les points de la gamme qui est utilisée pour estimer la
quantité de microfilaires chez les souris infestées (Figure 25). La gamme a été réalisée 2 fois.
Figure 25. Gamme de détection de l’ADN des microfilaires pulmonaires.
En abscisse sont indiquées le nombre de microfilaires ajoutées par poumon. En ordonnées le
ratio obtenu par PCR.
Les points de la gamme sont marqués en rouge et en vert les échantillons testés.
Test de corrélation (Pearson) entre le ratio de détection de l'ADN des microfilaires dans les
poumons et le nombre de microfilaires. r2=0,989.

135
12. Cultures cellulaires
12.1 Purification des neutrophiles
Les fémurs de souris C57BL/6 de type sauvage et de souris C57BL/6 S100A9ko sont isolés et
nettoyés pour enlever les tissus autour des os. Les condyles des os sont coupés avec un scalpel
et la moelle osseuse est poussée hors de l’os avec 5mL de PBS stérile (grâce à une seringue de
5mL et une aiguille 27G) ; elle est récupérée dans un tube de 15mL. Les morceaux de moelle
osseuse sont dissociés par des allers-retours avec la seringue. Le tube est centrifugé à 400g
pendant 4min à 4°C, le surnageant est éliminé et les cellules resuspendues
dans 3mL de PBS.
Les neutrophiles sont purifiés grâce à un gradient de Percoll en 3 couches.
Une solution de Percoll "100%" est préparée en mélangeant 9 volumes de
Percoll pur avec un volume de PBS 10X. Ensuite les différentes
concentrations de Percoll sont préparées comme suit :
Percoll 72% : 7,5mL Percoll 100 + 2,8 mL RPMI
Percoll 64% : 6,4mL Percoll 100 + 3,6 mL PBS
Percoll 52% : 5,2mL Percoll 100 + 4,8 mL RPMI
L'alternance de PBS et de RPMI pour la dilution du Percoll sert uniquement
à mieux visualiser les différentes couches.
Le gradient est ensuite monté à l'aide d'une pipette doucement et sans à-coup dans un
tube de 15mL en ajoutant 3mL de chaque concentration de Percoll pour finir avec le dépôt de
3mL de la suspension de cellules.
Le tube est centrifugé 30min à 1500g à température ambiante et sans frein.
Les neutrophiles matures sont retrouvés à l'interface entre les couches 72% et 64%, où ils sont
récupérés à l'aide d'une pipette pasteur molle. Les cellules sont lavées avec 10mL de PBS,
centrifugées à 400g pendant 4min à 4°C et le surnageant est éliminé. Les globules rouges
présents dans les culots cellulaires sont lysés par un choc hypotonique avec 5ml de NaCl
0.2% (puis équilibré avec l’ajout de 5 ml de NaCl à 1,6%). Après une nouvelle centrifugation,
les cellules sont resuspendues dans 1mL de PBS. Le nombre de cellules est déterminé à l’aide
d’un hémocytomètre (KOVA® GlassticSlide).

136
12.2 Cultures de neutrophiles
Pour étudier les réponses des neutrophiles à différents stimuli, 105 neutrophiles sont cultivés
en triplicats pour chaque condition sous 200µL de RPMI 1640 10% SVF, 1% L-Glutamine,
100U/mL Pénicilline-Streptomycine dans une plaque 96 puits à 37°C pendant 4, 6, 24 et 36h
à 37°C.
Les cellules sont cultivées avec les différents stimuli suivants :
- 10 ou 20 L3
- 100 nM de phorbolmyristateacetate (PMA)
- 100 ng/mL de LPS
- 10 µg/mL de Pam3Cys (un activateur du TLR4)
- 50 mg/mL d'antigène de filaires adultes
- 50 mg/mL d'antigène de filaires adultes traités à la tétracycline (sans Wolbachia)
- ne sont pas stimulées (contrôle)
Après l'incubation, le surnageant est récupéré et congelé à -20°C pour dosage ultérieur de
cytokines et de l'ADN extracellulaire et les cellules sont récupérées pour analyse en
cytométrie en flux. Les cellules sont utilisées pour étudier leur activation en cytométrie en
flux ou en microscopie électronique à balayage. Pour la microscopie électronique à balayage,
les cultures sont effectuées dans des inserts de culture cellulaires Millicell (Merck Millipore)
avec une membrane présentant des pores de 3µm. La culture en insert permet d'éviter de
perdre les échantillons lors des lavages et de la déshydratation.
12.3 Culture et stimulation des splénocytes
Les rates des souris infestées sont prélevées dans des conditions aseptiques. Elles sont
broyées par passage à travers les mailles d'un tamis cellulaires et récupérées dans un tube
15mL. Le tamis est rincé avec 5mL de RPMI 1640 + Pénicilline-Streptomycine (100 U/mL)
pour assurer la meilleure élution possible des cellules. Après centrifugation à 400g pendant
8min à 4°C et élimination du surnageant, les érythrocytes sont lysés à froid dans une solution
avec la solution RBC lysis buffer (Ebioscience). Après une nouvelle centrifugation à 400g
pendant 8min à 4°C, les cellules sont resuspendues dans 10mL de RPMI 1640 10% SVF, 1%
L-Glutamine, 100U/mL Pénicilline-Streptomycine et le nombre de cellules est déterminé avec
un compteur cellulaire (CASY Model TT, CellCounter and Analyzer, Roche).

137
Pour étudier les réponses de splénocytes après restimulation, 2.106 cellules sont
cultivées en triplicats pour chaque condition sous 1mL de RPMI 1640 10% SVF, 1% L-
Glutamine, 100U/mL Pénicilline-Streptomycine dans une plaque 24 puits à 37°C pendant
72h.
Les cellules sont cultivées selon les différentes conditions suivantes :
- 25mg/mL d'antigène de filaires adultes
- 2,5µg/mL de Concanavaline (ConA)
- Sans stimulus (contrôle).
Après l'incubation, le surnageant est récupéré et congelé à -20°C pour dosage ultérieur de
cytokines.
13. Dosage des cytokines et autres molécules inflammatoires par une méthode ELISA
Les molécules d’intérêt ont été mesurées dans le sérum, les liquides de rinçage du
lavage broncho-alvéolaire et de la cavité pleurale ainsi que dans les surnageants de cultures
cellulaires par une approche ELISA "sandwich". La technique ELISA (Enzyme Linked
ImmunoSorbent Assay) est une technique immuno-enzymatique de détection qui permet de
visualiser une réaction antigène-anticorps grâce à une réaction colorée produite par l'action
sur un substrat d'une enzyme préalablement fixée à l'anticorps. Différentes cytokines,
chimiokines et alarmines ont été mesurées en suivant les protocoles standards (IFN-ɣ, TNF-α,
IL-10, IL-4, IL-1β, IL-6 et IL-17 : kit Ready-set-go, Ebiosciences ; CCL2 et CXCL1 : ELISA
development kit, Peprotech ; MPO, S100A9 et IL-33: Duo-Set ELISA®, R&D). Le substrat
de l'enzyme est le TMB (3,3',5,5'-Tetramethylbenzidine), entraînant une réaction chimique
colorée qui est bloquée avant saturation par ajout d'acide sulfurique dilué (H2S4, 2N). La
lecture des échantillons est réalisée sur un spectrophotomètre Multiskan MS (Labsystems) à
450nm. L’acquisition des données a été effectuée sur le logiciel Delta Soft JV.
Les limites de détection sont les suivantes :
- 4 pg / mL pour IL-4, IL-6 et IL-17 ;
- 8 pg / mL pour IL- 1ß, S100A9, CXCL1 et MCP-1 ;
- 15 pg / mL pour INF-y et IL-33 ;
- 30 pg / mL pour IL-10 ;
- 250 pg / mL pour MPO.

138
14. Dosage de l'ADN extracellulaire
L'ADN extracellulaire double brin été quantifié dans le surnageant de culture des
neutrophiles de souris C57BL/6 après 4, 24 et 36 h à l'aide du kit d'analyse dsDNA Quant-
iTPicoGreen (Life Technologies), conformément aux instructions du fabricant. En bref, les
échantillons sont mélangés avec le réactif PicoGreen (dilution 1 : 1) pendant 5 min à l'abri de
la lumière puis la fluorescence des échantillons est mesurée avec un spectrofluoromètre
(FL600 Microplate fluorescence reader, Bio-Tek) avec une excitation à 480 nm et une
émission à 520nm. Une courbe standard d'ADN est utilisée pour déterminer la concentration
d'ADN libre dans les échantillons.
15. Analyses statistiques
Le choix des tests statistiques a été effectué avant l'analyse sur la base de la taille de
l'échantillon, de la normalité (test de Shapiro-Wilk) et de l'homoscédasticité (test de Bartlett).
Les données provenant d'expériences indépendantes ont été regroupées lorsque cela était
possible. Lorsque la normalité et l'homoscédasticité étaient établis, les résultats ont été
analysés par :
• le test t pour déterminer si deux jeux de données sont significativement différents.
• l'ANOVA à un facteur afin de déterminer l'effet d'un facteur (groupe de souris ou
temps).
• l'ANOVA à deux facteurs afin de déterminer les effets de deux facteurs (groupe de
souris et temps). Des comparaisons multiples de Bonferroni ont été effectuées après
les tests.
En cas de non-normalité des jeux de données, les résultats ont été analysés par des tests non-
paramétriques :
• le test U de Mann-Whitney pour déterminer si deux jeux de données sont
significativement différents.
• le test de Kruskal Wallis suivi d'un test de comparaison multiple de Dunn a été utilisé
afin de déterminer l'effet d'un facteur (condition ou temps).
Les corrélations entre deux jeux de données ont été analysées en utilisant le test de Pearson.
La représentation et l'analyse des données ont été réalisées avec le logiciel GraphPadPrism 5.

139

140

141
RESULTATS

142

143
CHAPITRE 1 :
La migration des larves infestantes/L3 de Litomosoides
sigmodontis chez la souris BALB/c révèle l’existence de dommages
pulmonaires transitoires

144
Introduction
Les L3 des filaires sont injectées dans la peau de l'hôte lors du repas sanguin d'un
vecteur arthropode hématophage. La plupart des espèces filariennes migrent à travers le corps
de l'hôte, de la peau jusqu'à leur niche définitive où les adultes se reproduisent et libèrent des
microfilaires. Ce lieu est différent en fonction de l’espèce filarienne considérée mais 4
localisations sont généralement décrites : le système lymphatique, les cavités séreuses, le
système cardio-pulmonaire ou les tissus conjonctifs [195]. Les modèles de filarioses chez les
rongeurs ont été utilisés pour étudier les voies de migration des L3 en particulier pour décrire
le passage lymphatique des L3 ; de plus certains de ces modèles ont permis d’observer la
présence des L3 dans le poumon, ce qui est le cas pour le parasite humain Brugia malayi dans
la gerbille et pour les principaux modèles animaux de filariose à savoir Brugia pahangi,
Acanthocheilonema viteae et Litomosoides sigmodontis [10]. Cette dernière est la seule filaire
capable d'effectuer un développement complet chez les souris ce qui en a fait un modèle
particulièrement étudié [194]. En outre, L. sigmodontis partage diverses caractéristiques
biologiques avec la filaire humaine Mansonella persans telles que la mue en L4 ayant lieu
entre 9 et 10 jours après infection, les adultes vivant dans les cavités séreuses, dont la cavité
pleurale, et les microfilaires circulant dans le sang [55,194,202].
La biologie de M. perstans est mal connue. Notamment, la voie de migration de la
peau vers la cavité pleurale n'est toujours pas clairement identifiée pour M. perstans; c'est
également le cas pour L. sigmodontis. En 1967, Wenk [11] a émis l'hypothèse que les L3 de L.
sigmodontis et de M. perstans traverseraient les poumons pour atteindre la cavité pleurale.
Cette hypothèse semble en effet probable d'un point de vue anatomique : les L3 de L.
sigmodontis inoculées dans la peau pénètrent rapidement dans un vaisseau lymphatique [11,199]
(Kilarski non publié ; http://www.wired.com/wiredscience/2011/08/nikon-small-world-vote-
2011/?pid=1861) ; de là, les larves s'engageraient dans le canal thoracique et arriveraient ainsi
dans la veine cave supérieure, le cœur droit, puis les artères pulmonaires (Figure 26). Dans les
poumons, les L3 seraient bloquées dans les capillaires pulmonaires dont elles s'échapperaient
pour rejoindre la cavité pleurale en traversant le tissu pulmonaire.
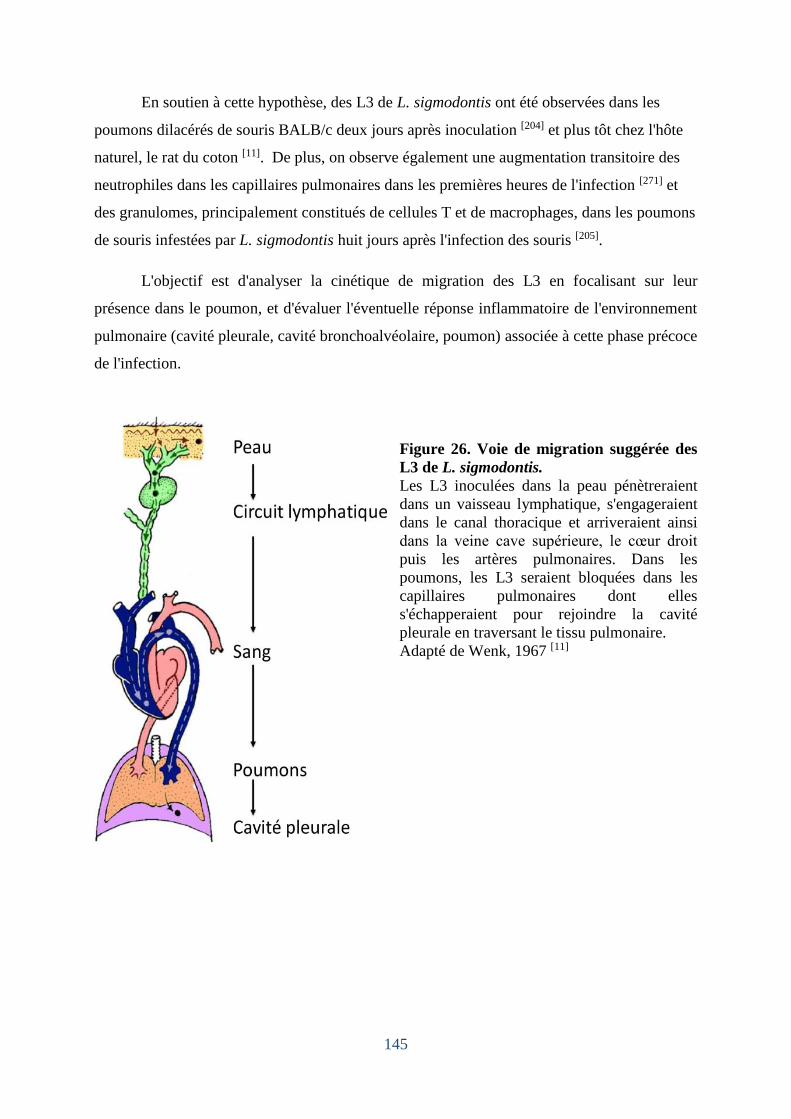
145
En soutien à cette hypothèse, des L3 de L. sigmodontis ont été observées dans les
poumons dilacérés de souris BALB/c deux jours après inoculation [204] et plus tôt chez l'hôte
naturel, le rat du coton [11]. De plus, on observe également une augmentation transitoire des
neutrophiles dans les capillaires pulmonaires dans les premières heures de l'infection [271] et
des granulomes, principalement constitués de cellules T et de macrophages, dans les poumons
de souris infestées par L. sigmodontis huit jours après l'infection des souris [205].
L'objectif est d'analyser la cinétique de migration des L3 en focalisant sur leur
présence dans le poumon, et d'évaluer l'éventuelle réponse inflammatoire de l'environnement
pulmonaire (cavité pleurale, cavité bronchoalvéolaire, poumon) associée à cette phase précoce
de l'infection.
Figure 26. Voie de migration suggérée des
L3 de L. sigmodontis.
Les L3 inoculées dans la peau pénètreraient
dans un vaisseau lymphatique, s'engageraient
dans le canal thoracique et arriveraient ainsi
dans la veine cave supérieure, le cœur droit
puis les artères pulmonaires. Dans les
poumons, les L3 seraient bloquées dans les
capillaires pulmonaires dont elles
s'échapperaient pour rejoindre la cavité
pleurale en traversant le tissu pulmonaire.
Adapté de Wenk, 1967 [11]
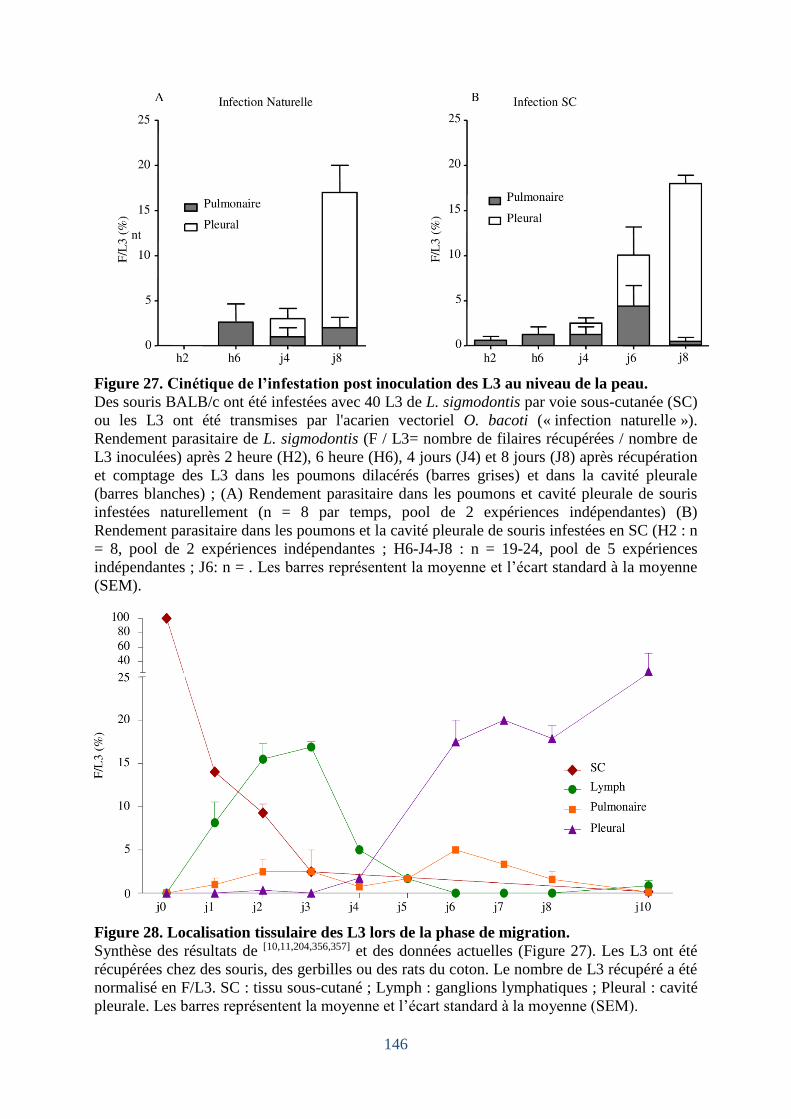
146
Figure 27. Cinétique de l’infestation post inoculation des L3 au niveau de la peau.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée (SC)
ou les L3 ont été transmises par l'acarien vectoriel O. bacoti (« infection naturelle »).
Rendement parasitaire de L. sigmodontis (F / L3= nombre de filaires récupérées / nombre de
L3 inoculées) après 2 heure (H2), 6 heure (H6), 4 jours (J4) et 8 jours (J8) après récupération
et comptage des L3 dans les poumons dilacérés (barres grises) et dans la cavité pleurale
(barres blanches) ; (A) Rendement parasitaire dans les poumons et cavité pleurale de souris
infestées naturellement (n = 8 par temps, pool de 2 expériences indépendantes) (B)
Rendement parasitaire dans les poumons et la cavité pleurale de souris infestées en SC (H2 : n
= 8, pool de 2 expériences indépendantes ; H6-J4-J8 : n = 19-24, pool de 5 expériences
indépendantes ; J6: n = . Les barres représentent la moyenne et l’écart standard à la moyenne
(SEM).
Figure 28. Localisation tissulaire des L3 lors de la phase de migration.
Synthèse des résultats de [10,11,204,356,357] et des données actuelles (Figure 27). Les L3 ont été
récupérées chez des souris, des gerbilles ou des rats du coton. Le nombre de L3 récupéré a été
normalisé en F/L3. SC : tissu sous-cutané ; Lymph : ganglions lymphatiques ; Pleural : cavité
pleurale. Les barres représentent la moyenne et l’écart standard à la moyenne (SEM).

147
Résultats
1 Cinétique de la migration des L3 ; comparaison de différents modes d'inoculation
Pour définir la cinétique d'arrivée des L3 dans la cavité pleurale, des souris BALB/c
ont été infestées par le vecteur acarien O. bacoti ("infection naturelle") ou par voie sous-
cutanée (SC). Les L3 présentes dans la cavité pleurale et dans les poumons ont été
dénombrées à différents temps entre 2 heures (H2) et 8 jours (J8) post-infection (p.i.) (Figures
27A et 27B).
Peu de L3 sont détectées à H2, H6 et J4 p.i. dans la cavité pleurale des souris infestées
en SC ou naturellement (rendement parasitaire <5% de l'inoculât initial). Entre J4 et J8, le
nombre de L3 dans la cavité pleurale augmente progressivement, pour atteindre près de 20%
(Figures 27A et B).
Des L3 sont également observées dans les poumons des souris infestées (infection
naturelle et SC) (Figures 27A et B). Ces résultats concordent avec les observations antérieures
de Wenk et de Bain, qui ont remarqué la présence de L3 de L. sigmodontis dans les poumons
de rongeurs dilacérés mécaniquement [10,11,204,356,357].
Mises ensemble, ces données et les nôtres permettent de proposer le schéma suivant montrant
la progression des L3 de la peau à la cavité pleurale (Figure 28).
Les L3 disparaissent de la peau en quelques jours. Elles sont observées dans les
lymphatiques dès quelques heures après infection avec un pic à J2 et J3 p.i., avant d'atteindre
la cavité pleurale et de s'y accumuler entre J4 et J6 p.i. Une fois dans la cavité pleurale, le
rendement parasitaire reste stable pendant une durée variable selon les espèces hôtes des
rongeurs, de 10 à 30 jours p.i.
La présence de L3 dans les poumons a été observée tout au long de la phase de
migration avec un pic à J6 p.i. (Figure 27B). Le nombre de souris présentant des L3 dans les
poumons augmente également au cours du temps de 25% à H2 p.i. jusqu'à 85% à J6 p.i.
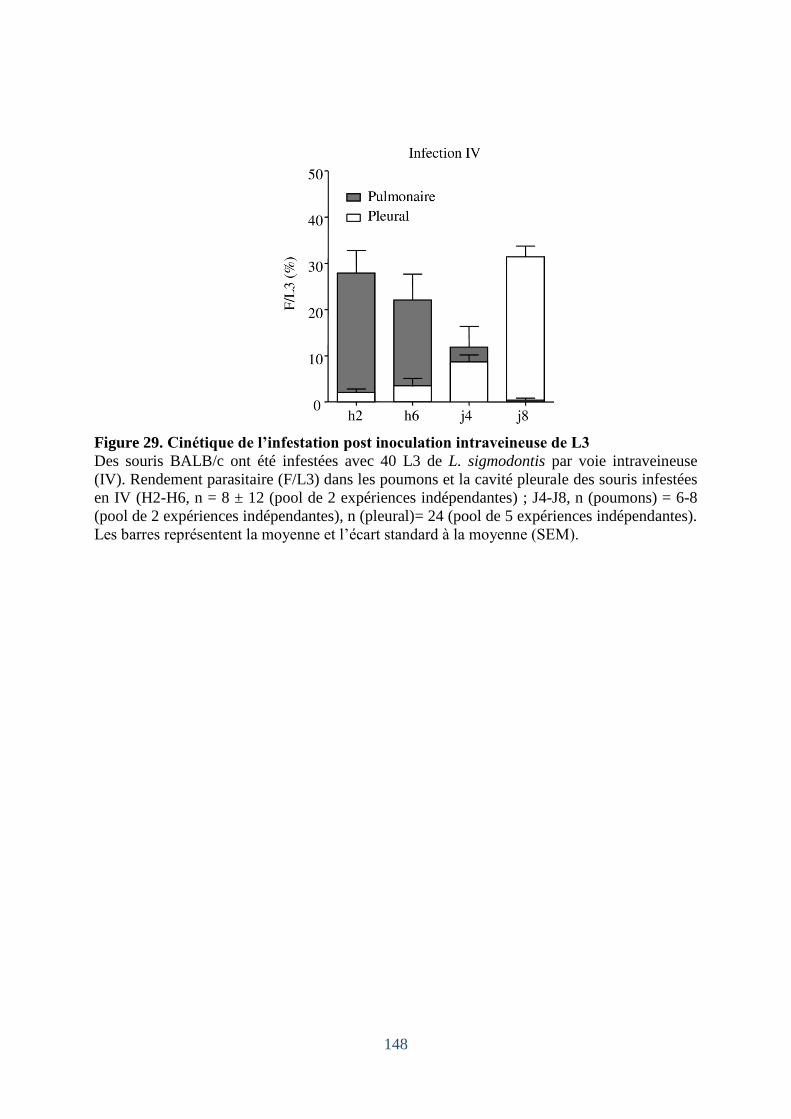
148
Figure 29. Cinétique de l’infestation post inoculation intraveineuse de L3
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie intraveineuse
(IV). Rendement parasitaire (F/L3) dans les poumons et la cavité pleurale des souris infestées
en IV (H2-H6, n = 8 ± 12 (pool de 2 expériences indépendantes) ; J4-J8, n (poumons) = 6-8
(pool de 2 expériences indépendantes), n (pleural)= 24 (pool de 5 expériences indépendantes).
Les barres représentent la moyenne et l’écart standard à la moyenne (SEM).

149
Afin de tester la possibilité d'un passage sanguin, les L3 ont été inoculées par voie
intraveineuse (IV). De plus, cette méthode permet d’éviter la peau, la phase de migration
lymphatique et potentiellement de limiter l'arrivée asynchrone des L3 dans la cavité pleurale.
Le rendement parasitaire est plus élevé dans la cavité pleurale après ce mode
d'inoculation (Figure 29). Il atteint presque le double de celui observé chez les souris infestées
en SC ou naturellement au même moment. Ceci indique d'une part que les L3 survivent dans
la circulation sanguine et d'autre part qu'elles peuvent utiliser ce moyen pour rejoindre la
cavité pleurale. Les larves sont observées dans la cavité pleurale dès H2 p.i., et s'accumulent
régulièrement jusqu'à J8 p.i. (environ 2% à H2, 4% à H6, 10% à J4 et 30% à J8). Au cours de
ce laps de temps, un équilibre entre les poumons et la cavité pleurale est observé : le taux de
récupération dans les poumons est maximal à H2 p.i. et diminue progressivement jusqu'à J8
p.i. (environ 30% à H2, 26% à H6, 13% à J4 et moins de 1% à J8).
Dans les deux modèles (inoculation dans la peau ou en IV), la localisation pulmonaire
suggère que les L3 quittent les capillaires pulmonaires (comme l'a suggéré Wenk [11])
probablement en migrant à travers le parenchyme pulmonaire et le mésothélium viscéral pour
arriver dans la cavité pleurale. L'observation d'une L3 sur une coupe histologique de poumon
d'une souris infestée en IV [271], associée à un recrutement de cellules dont de nombreux
neutrophiles, conforte cette hypothèse.

150
2. Le passage de L3 dans le poumon induit une inflammation caractérisée par des
hémorragies locales, des granulomes et une accumulation de neutrophiles dans
les espaces périvasculaires
Au cours de l’autopsie des souris infestées, des zones hémorragiques localisées
(pétéchies) ont fréquemment été observées à la surface des poumons chez les souris infestées
en IV (Figure 30B par rapport à la Figure 30A). Ce phénomène a été quantifié à H6, J4 et J8
p.i. chez les souris infestées en IV et en SC (Figure 30C).
Figure 30. Hémorragies dans le poumon de souris où sont détectables les L3.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis soit par voie sous-cutanée
(SC), soit par voie intraveineuse (IV). Image représentative de (A) un poumon normal, (B) un
poumon avec de nombreuses hémorragies (cercles rouges bien délimités). (C) Nombre
d'hémorragies pulmonaires superficielles dans les poumons à six heures (H6), quatre jours
(J4) et huit jours (J8) après l'inoculation. n = 6, les barres représentent la moyenne ± SEM;
ANOVA bidirectionnelle suivie de Bonferonni, *** = p <0,001 (différence entre les souris
infestées en IV et SC), ## = p <0,01 (différence entre les points de temps chez les souris
infestées en IV). (D) Test de corrélation (Pearson) entre le nombre de L3 récupéré dans le
poumon et le nombre de hémorragies, r2 = 0,9148.
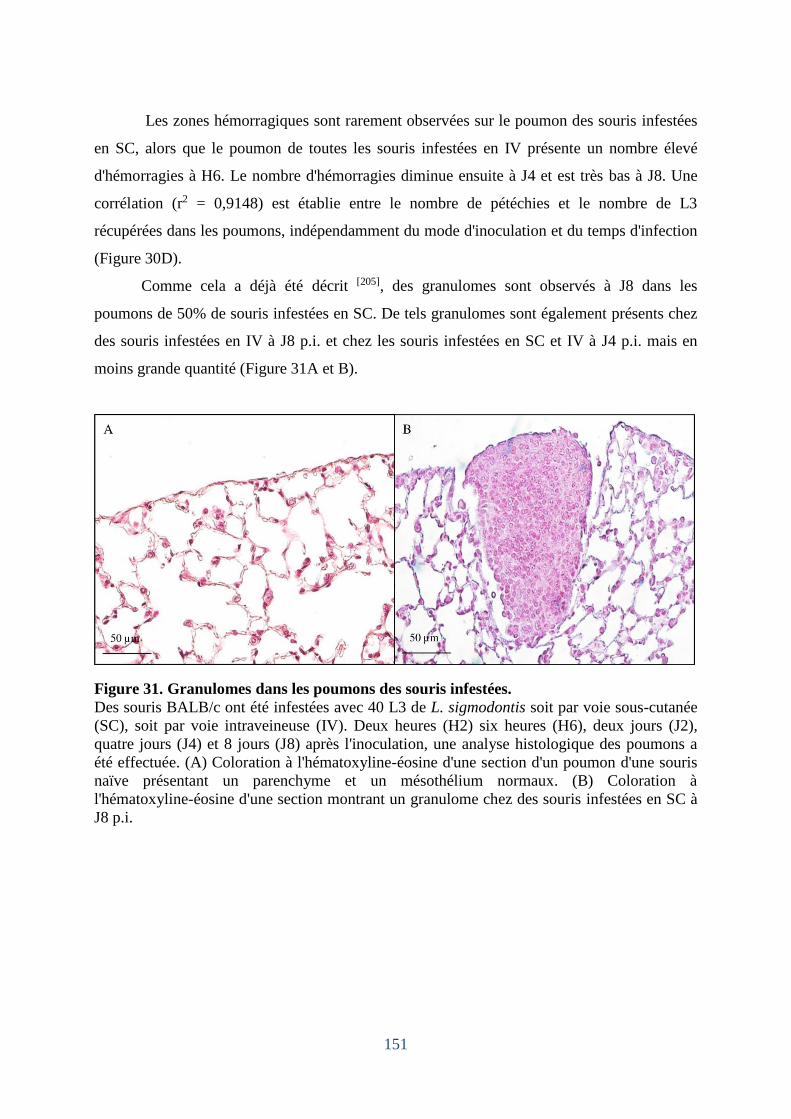
151
Les zones hémorragiques sont rarement observées sur le poumon des souris infestées
en SC, alors que le poumon de toutes les souris infestées en IV présente un nombre élevé
d'hémorragies à H6. Le nombre d'hémorragies diminue ensuite à J4 et est très bas à J8. Une
corrélation (r2 = 0,9148) est établie entre le nombre de pétéchies et le nombre de L3
récupérées dans les poumons, indépendamment du mode d'inoculation et du temps d'infection
(Figure 30D).
Comme cela a déjà été décrit [205], des granulomes sont observés à J8 dans les
poumons de 50% de souris infestées en SC. De tels granulomes sont également présents chez
des souris infestées en IV à J8 p.i. et chez les souris infestées en SC et IV à J4 p.i. mais en
moins grande quantité (Figure 31A et B).
Figure 31. Granulomes dans les poumons des souris infestées.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis soit par voie sous-cutanée
(SC), soit par voie intraveineuse (IV). Deux heures (H2) six heures (H6), deux jours (J2),
quatre jours (J4) et 8 jours (J8) après l'inoculation, une analyse histologique des poumons a
été effectuée. (A) Coloration à l'hématoxyline-éosine d'une section d'un poumon d'une souris
naïve présentant un parenchyme et un mésothélium normaux. (B) Coloration à
l'hématoxyline-éosine d'une section montrant un granulome chez des souris infestées en SC à
J8 p.i.
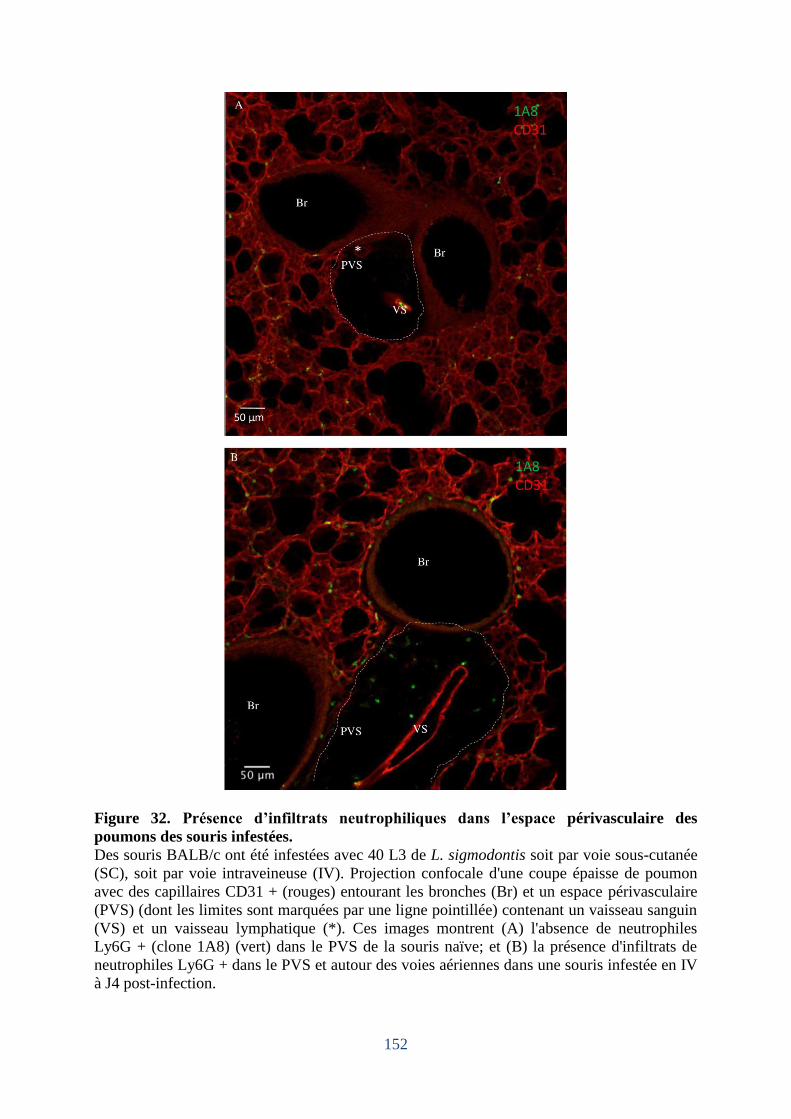
152
Figure 32. Présence d’infiltrats neutrophiliques dans l’espace périvasculaire des
poumons des souris infestées.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis soit par voie sous-cutanée
(SC), soit par voie intraveineuse (IV). Projection confocale d'une coupe épaisse de poumon
avec des capillaires CD31 + (rouges) entourant les bronches (Br) et un espace périvasculaire
(PVS) (dont les limites sont marquées par une ligne pointillée) contenant un vaisseau sanguin
(VS) et un vaisseau lymphatique (*). Ces images montrent (A) l'absence de neutrophiles
Ly6G + (clone 1A8) (vert) dans le PVS de la souris naïve; et (B) la présence d'infiltrats de
neutrophiles Ly6G + dans le PVS et autour des voies aériennes dans une souris infestée en IV
à J4 post-infection.
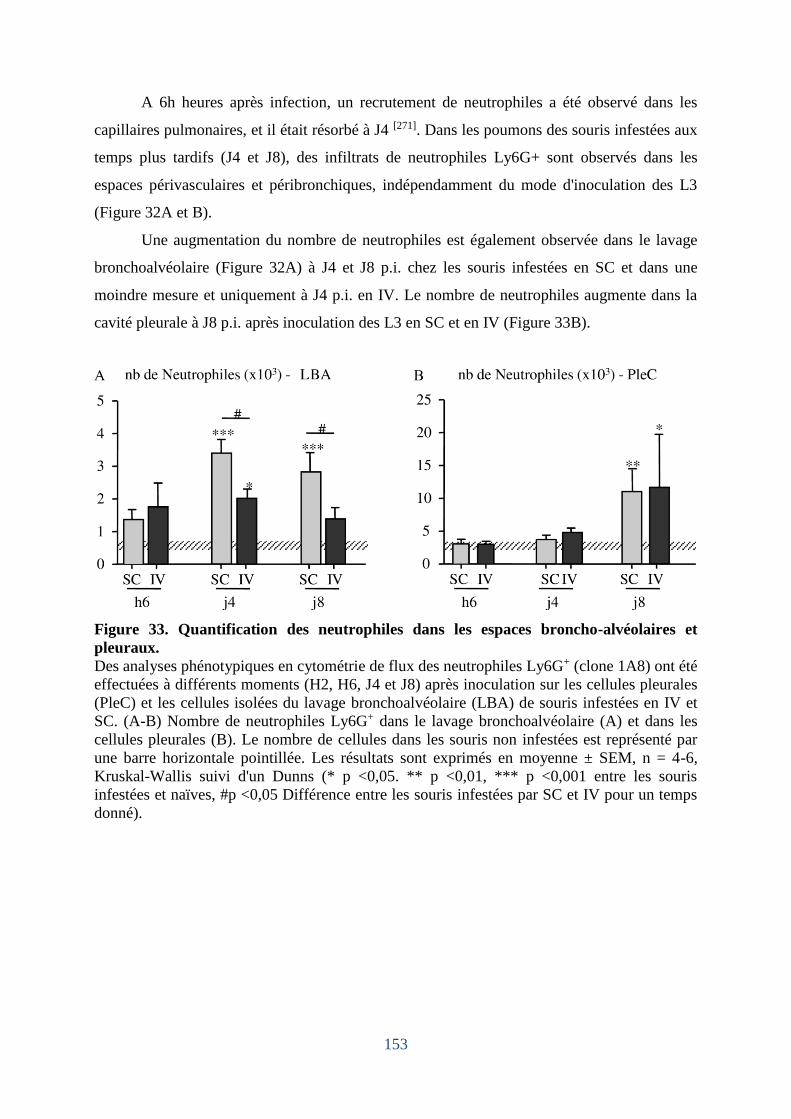
153
A 6h heures après infection, un recrutement de neutrophiles a été observé dans les
capillaires pulmonaires, et il était résorbé à J4 [271]. Dans les poumons des souris infestées aux
temps plus tardifs (J4 et J8), des infiltrats de neutrophiles Ly6G+ sont observés dans les
espaces périvasculaires et péribronchiques, indépendamment du mode d'inoculation des L3
(Figure 32A et B).
Une augmentation du nombre de neutrophiles est également observée dans le lavage
bronchoalvéolaire (Figure 32A) à J4 et J8 p.i. chez les souris infestées en SC et dans une
moindre mesure et uniquement à J4 p.i. en IV. Le nombre de neutrophiles augmente dans la
cavité pleurale à J8 p.i. après inoculation des L3 en SC et en IV (Figure 33B).
Figure 33. Quantification des neutrophiles dans les espaces broncho-alvéolaires et
pleuraux.
Des analyses phénotypiques en cytométrie de flux des neutrophiles Ly6G+ (clone 1A8) ont été
effectuées à différents moments (H2, H6, J4 et J8) après inoculation sur les cellules pleurales
(PleC) et les cellules isolées du lavage bronchoalvéolaire (LBA) de souris infestées en IV et
SC. (A-B) Nombre de neutrophiles Ly6G+ dans le lavage bronchoalvéolaire (A) et dans les
cellules pleurales (B). Le nombre de cellules dans les souris non infestées est représenté par
une barre horizontale pointillée. Les résultats sont exprimés en moyenne ± SEM, n = 4-6,
Kruskal-Wallis suivi d'un Dunns (* p <0,05. ** p <0,01, *** p <0,001 entre les souris
infestées et naïves, #p <0,05 Différence entre les souris infestées par SC et IV pour un temps
donné).
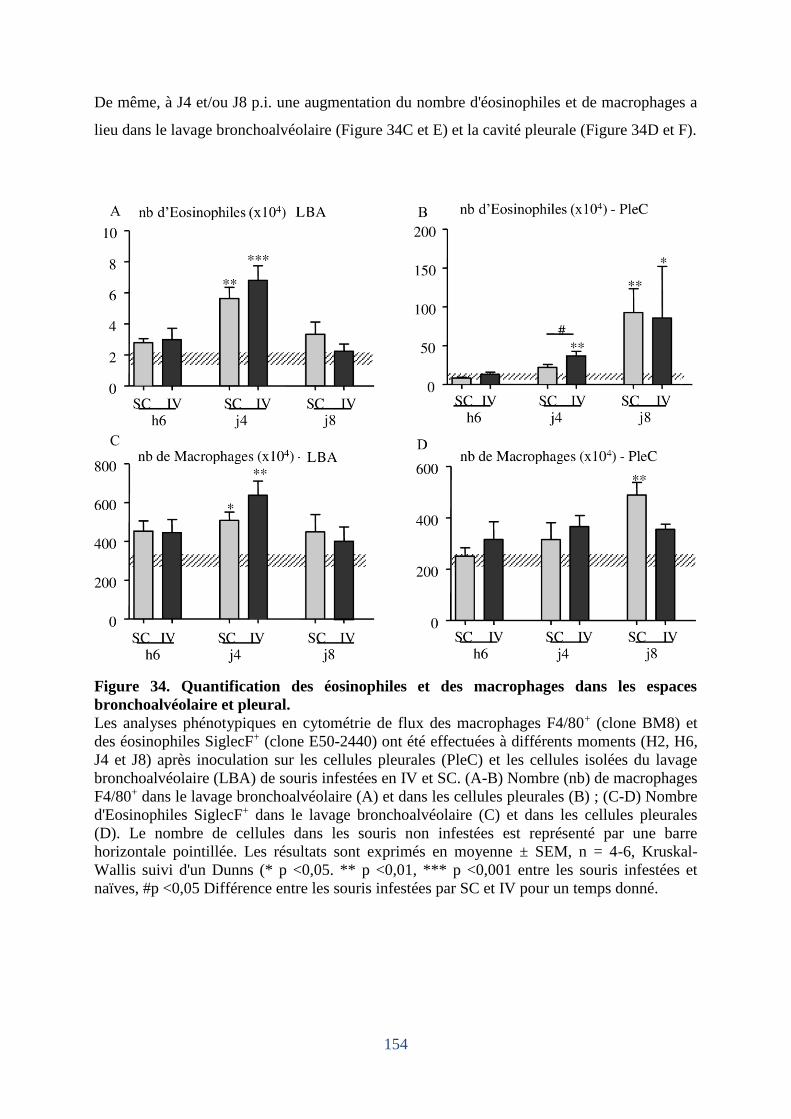
154
De même, à J4 et/ou J8 p.i. une augmentation du nombre d'éosinophiles et de macrophages a
lieu dans le lavage bronchoalvéolaire (Figure 34C et E) et la cavité pleurale (Figure 34D et F).
Figure 34. Quantification des éosinophiles et des macrophages dans les espaces
bronchoalvéolaire et pleural.
Les analyses phénotypiques en cytométrie de flux des macrophages F4/80+ (clone BM8) et
des éosinophiles SiglecF+ (clone E50-2440) ont été effectuées à différents moments (H2, H6,
J4 et J8) après inoculation sur les cellules pleurales (PleC) et les cellules isolées du lavage
bronchoalvéolaire (LBA) de souris infestées en IV et SC. (A-B) Nombre (nb) de macrophages
F4/80+ dans le lavage bronchoalvéolaire (A) et dans les cellules pleurales (B) ; (C-D) Nombre
d'Eosinophiles SiglecF+ dans le lavage bronchoalvéolaire (C) et dans les cellules pleurales
(D). Le nombre de cellules dans les souris non infestées est représenté par une barre
horizontale pointillée. Les résultats sont exprimés en moyenne ± SEM, n = 4-6, Kruskal-
Wallis suivi d'un Dunns (* p <0,05. ** p <0,01, *** p <0,001 entre les souris infestées et
naïves, #p <0,05 Différence entre les souris infestées par SC et IV pour un temps donné.
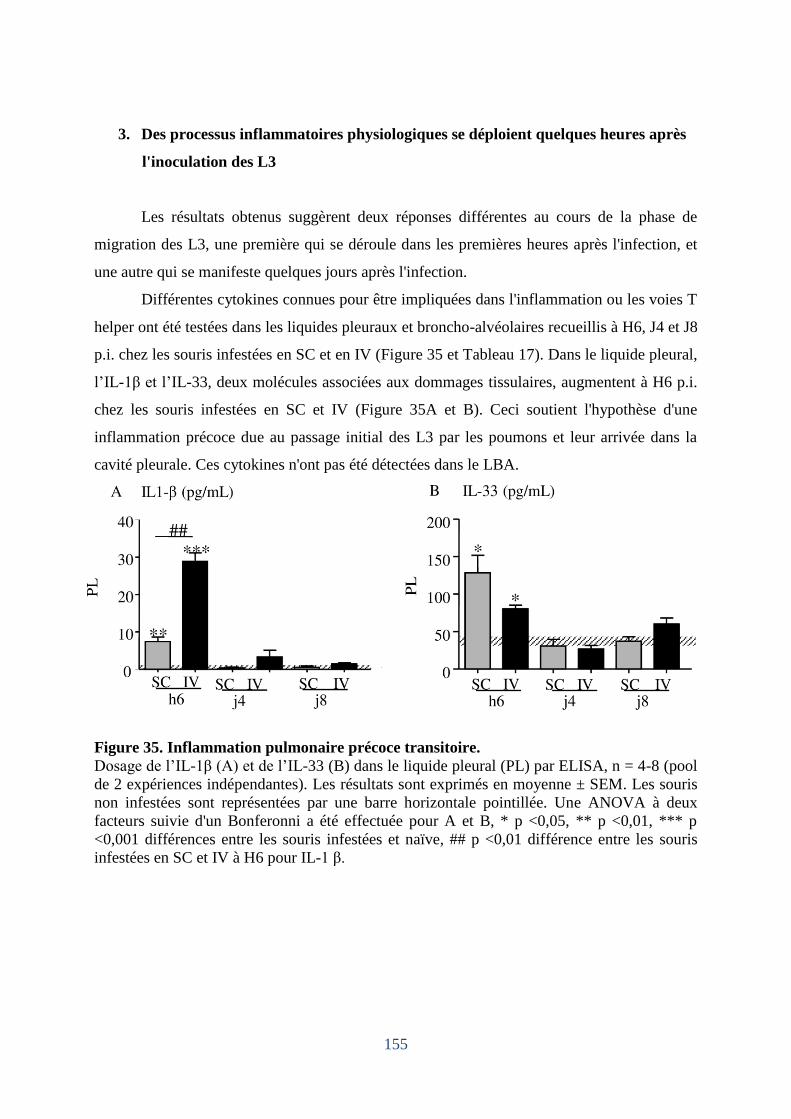
155
3. Des processus inflammatoires physiologiques se déploient quelques heures après
l'inoculation des L3
Les résultats obtenus suggèrent deux réponses différentes au cours de la phase de
migration des L3, une première qui se déroule dans les premières heures après l'infection, et
une autre qui se manifeste quelques jours après l'infection.
Différentes cytokines connues pour être impliquées dans l'inflammation ou les voies T
helper ont été testées dans les liquides pleuraux et broncho-alvéolaires recueillis à H6, J4 et J8
p.i. chez les souris infestées en SC et en IV (Figure 35 et Tableau 17). Dans le liquide pleural,
l’IL-1β et l’IL-33, deux molécules associées aux dommages tissulaires, augmentent à H6 p.i.
chez les souris infestées en SC et IV (Figure 35A et B). Ceci soutient l'hypothèse d'une
inflammation précoce due au passage initial des L3 par les poumons et leur arrivée dans la
cavité pleurale. Ces cytokines n'ont pas été détectées dans le LBA.
Figure 35. Inflammation pulmonaire précoce transitoire.
Dosage de l’IL-1β (A) et de l’IL-33 (B) dans le liquide pleural (PL) par ELISA, n = 4-8 (pool
de 2 expériences indépendantes). Les résultats sont exprimés en moyenne ± SEM. Les souris
non infestées sont représentées par une barre horizontale pointillée. Une ANOVA à deux
facteurs suivie d'un Bonferonni a été effectuée pour A et B, * p <0,05, ** p <0,01, *** p
<0,001 différences entre les souris infestées et naïve, ## p <0,01 différence entre les souris
infestées en SC et IV à H6 pour IL-1 β.
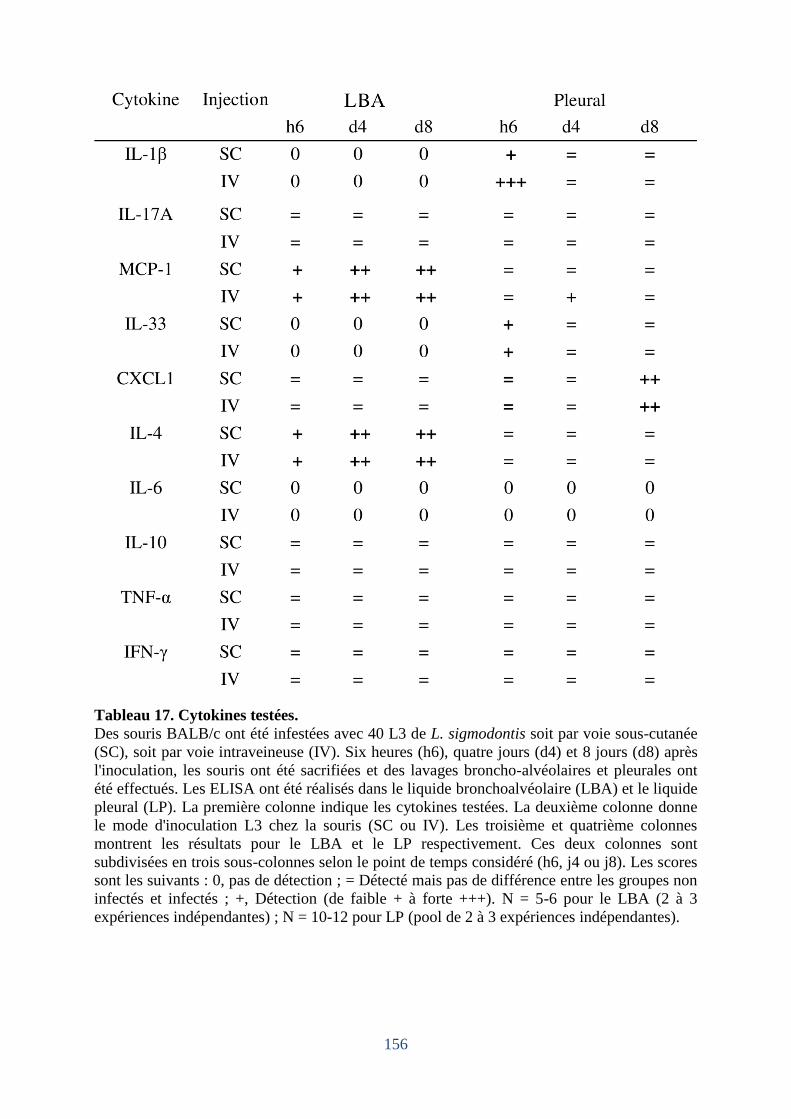
156
Tableau 17. Cytokines testées.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis soit par voie sous-cutanée
(SC), soit par voie intraveineuse (IV). Six heures (h6), quatre jours (d4) et 8 jours (d8) après
l'inoculation, les souris ont été sacrifiées et des lavages broncho-alvéolaires et pleurales ont
été effectués. Les ELISA ont été réalisés dans le liquide bronchoalvéolaire (LBA) et le liquide
pleural (LP). La première colonne indique les cytokines testées. La deuxième colonne donne
le mode d'inoculation L3 chez la souris (SC ou IV). Les troisième et quatrième colonnes
montrent les résultats pour le LBA et le LP respectivement. Ces deux colonnes sont
subdivisées en trois sous-colonnes selon le point de temps considéré (h6, j4 ou j8). Les scores
sont les suivants : 0, pas de détection ; = Détecté mais pas de différence entre les groupes non
infectés et infectés ; +, Détection (de faible + à forte +++). N = 5-6 pour le LBA (2 à 3
expériences indépendantes) ; N = 10-12 pour LP (pool de 2 à 3 expériences indépendantes).

157
Pour caractériser davantage l'environnement inflammatoire dans la deuxième phase,
c'est-à-dire quelques jours après l'infection, les transcrits pulmonaires de plusieurs cytokines
(Annexe 1) ont été analysés à J4 p.i. (Figure 36) chez les souris infestées en SC. Les transcrits
montrent une forte régulation négative de la réponse inflammatoire et des fonctions telles que
la chimiotaxie et le recrutement de neutrophiles et de phagocytes. Par exemple, l'expression
de transcrits de cytokines proinflammatoires comme l'IL-6, le CXCL1 et le CXCL5 sont
diminués. Les transcrits de CXCL1, un chimioattractant puissant pour les neutrophiles, sont
diminués dans le tissu pulmonaire des souris inoculées en SC et IV (Figure 36C). Les niveaux
de la protéine CXCL1 sont néanmoins augmentés dans le liquide pleural à J8 (Tableau 17) et
pourraient être associés au recrutement de neutrophiles observés à ce temps là (Figure 32).
Dans le même temps, l'expression de molécules régulatrices augmente comme les transcrits
d'IL-9 et d'IL-3 et les niveaux d'IL-4 et de MCP-1 dans le LBA (Tableau 17).
Figure 36. Régulation de l’inflammation pulmonaire
(A) Transcrits de cytokines/chimiokines (seuil : facteur d’augmentation/baisse d’expression
d’au moins 2) induite dans le poumon de souris infestées en SC à J4 p.i. et (B) fonctions
associées. Les effets des changements d'expression des gènes dans les poumons ont été prédits
en utilisant Ingenuity Pathways Analysis (IPA) ; la régulation des fonctions est indiquée par
des flèches. (C) Une q-RTPCR a été effectuée pour les transcrits de Cxcl1 pour valider les
résultats du tableau; N = 12 (SC) et 5 (IV). Un test t a été effectué pour Cxcl1, ** p <0,01.
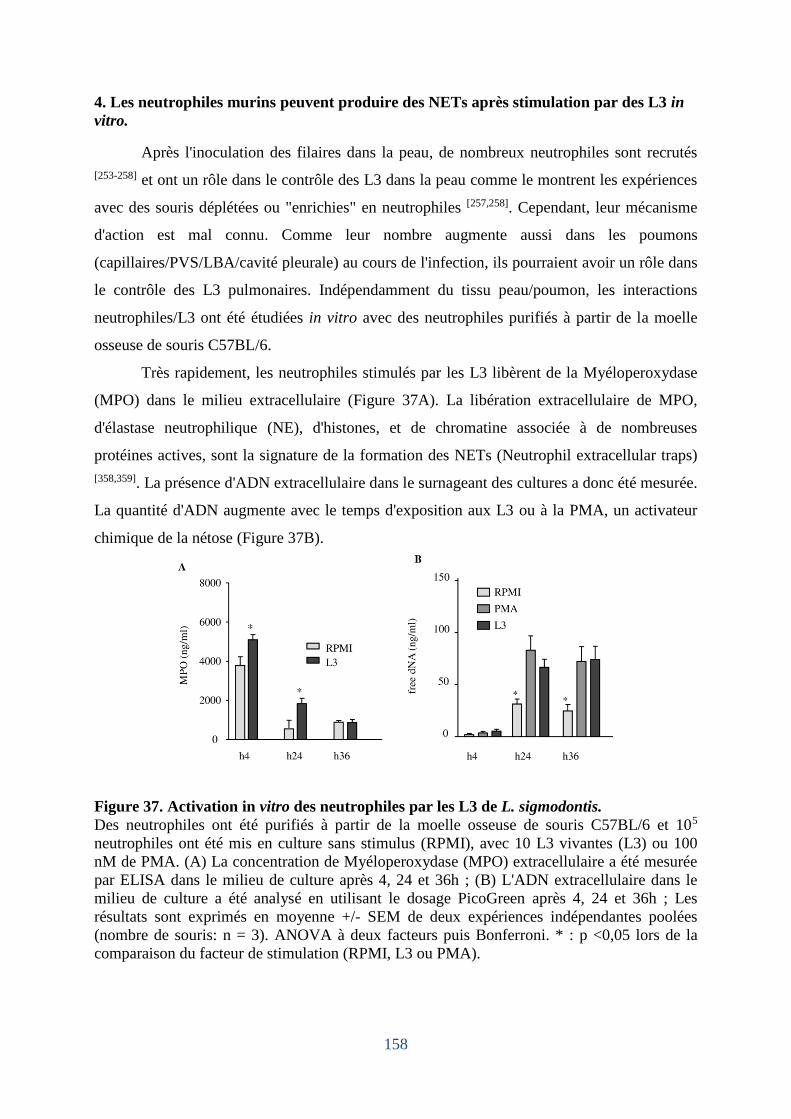
158
4. Les neutrophiles murins peuvent produire des NETs après stimulation par des L3 in
vitro.
Après l'inoculation des filaires dans la peau, de nombreux neutrophiles sont recrutés
[253-258] et ont un rôle dans le contrôle des L3 dans la peau comme le montrent les expériences
avec des souris déplétées ou "enrichies" en neutrophiles [257,258]. Cependant, leur mécanisme
d'action est mal connu. Comme leur nombre augmente aussi dans les poumons
(capillaires/PVS/LBA/cavité pleurale) au cours de l'infection, ils pourraient avoir un rôle dans
le contrôle des L3 pulmonaires. Indépendamment du tissu peau/poumon, les interactions
neutrophiles/L3 ont été étudiées in vitro avec des neutrophiles purifiés à partir de la moelle
osseuse de souris C57BL/6.
Très rapidement, les neutrophiles stimulés par les L3 libèrent de la Myéloperoxydase
(MPO) dans le milieu extracellulaire (Figure 37A). La libération extracellulaire de MPO,
d'élastase neutrophilique (NE), d'histones, et de chromatine associée à de nombreuses
protéines actives, sont la signature de la formation des NETs (Neutrophil extracellular traps)
[358,359]. La présence d'ADN extracellulaire dans le surnageant des cultures a donc été mesurée.
La quantité d'ADN augmente avec le temps d'exposition aux L3 ou à la PMA, un activateur
chimique de la nétose (Figure 37B).
Figure 37. Activation in vitro des neutrophiles par les L3 de L. sigmodontis.
Des neutrophiles ont été purifiés à partir de la moelle osseuse de souris C57BL/6 et 105
neutrophiles ont été mis en culture sans stimulus (RPMI), avec 10 L3 vivantes (L3) ou 100
nM de PMA. (A) La concentration de Myéloperoxydase (MPO) extracellulaire a été mesurée
par ELISA dans le milieu de culture après 4, 24 et 36h ; (B) L'ADN extracellulaire dans le
milieu de culture a été analysé en utilisant le dosage PicoGreen après 4, 24 et 36h ; Les
résultats sont exprimés en moyenne +/- SEM de deux expériences indépendantes poolées
(nombre de souris: n = 3). ANOVA à deux facteurs puis Bonferroni. * : p <0,05 lors de la
comparaison du facteur de stimulation (RPMI, L3 ou PMA).
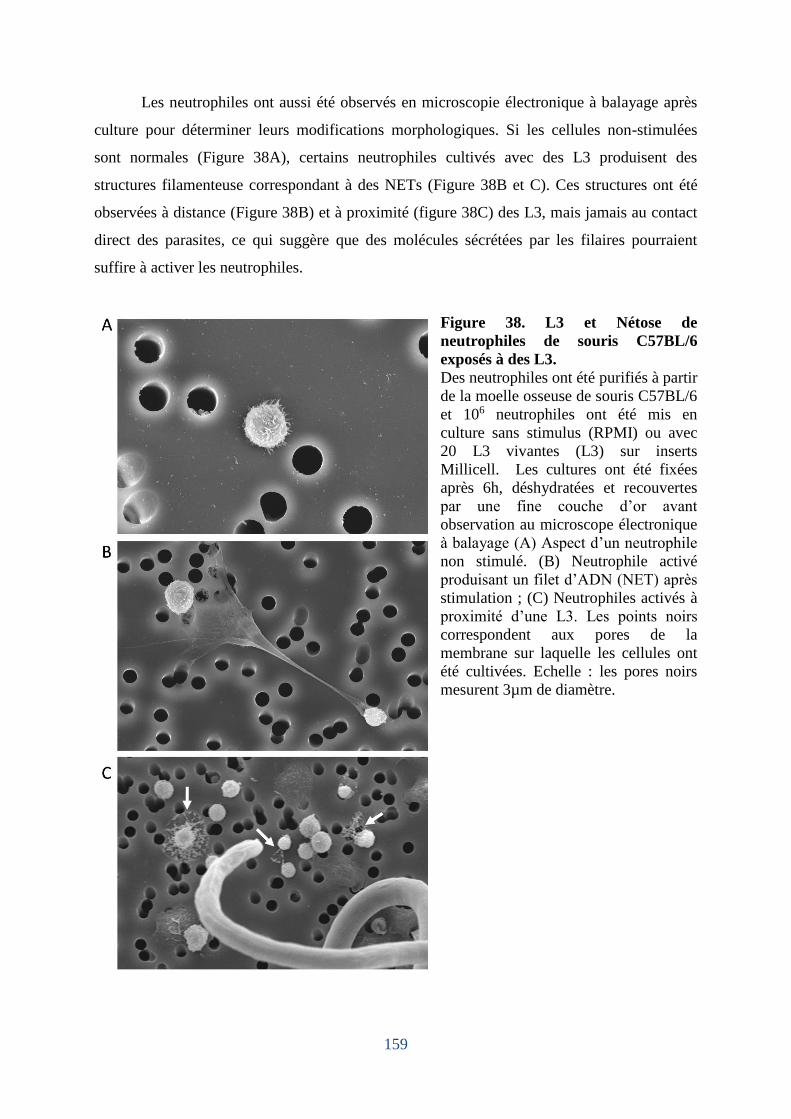
159
Les neutrophiles ont aussi été observés en microscopie électronique à balayage après
culture pour déterminer leurs modifications morphologiques. Si les cellules non-stimulées
sont normales (Figure 38A), certains neutrophiles cultivés avec des L3 produisent des
structures filamenteuse correspondant à des NETs (Figure 38B et C). Ces structures ont été
observées à distance (Figure 38B) et à proximité (figure 38C) des L3, mais jamais au contact
direct des parasites, ce qui suggère que des molécules sécrétées par les filaires pourraient
suffire à activer les neutrophiles.
Figure 38. L3 et Nétose de
neutrophiles de souris C57BL/6
exposés à des L3.
Des neutrophiles ont été purifiés à partir
de la moelle osseuse de souris C57BL/6
et 106 neutrophiles ont été mis en
culture sans stimulus (RPMI) ou avec
20 L3 vivantes (L3) sur inserts
Millicell. Les cultures ont été fixées
après 6h, déshydratées et recouvertes
par une fine couche d’or avant
observation au microscope électronique
à balayage (A) Aspect d’un neutrophile
non stimulé. (B) Neutrophile activé
produisant un filet d’ADN (NET) après
stimulation ; (C) Neutrophiles activés à
proximité d’une L3. Les points noirs
correspondent aux pores de la
membrane sur laquelle les cellules ont
été cultivées. Echelle : les pores noirs
mesurent 3µm de diamètre.

160

161
Discussion
Dans les 8 jours suivant leur inoculation cutanée, les L3 migrent du site d'inoculation
dans la peau, initialement via le système lymphatique, vers la cavité pleurale. Aucune larve ne
se transforme en L4 durant ce processus migratoire car la mue en L4 de L. sigmodontis a lieu
dans la cavité pleurale vers 9-10 jours post-inoculation chez les souris BALB/c [10,199].
Toutefois la taille de la L3 augmente pendant ces 8 jours de migration passant de 750µm de
long à 1400µm [360].
En raison de leur voie de migration complexe, les L3 sont donc confrontées à plusieurs
goulets d'étranglement anatomiques, ce qui entraîne une perte de leur synchronisation dans
l’inoculât initial et leur arrivée dans la cavité pleurale au "compte goutte". Les larves
s'accumulent ensuite pendant 6 à 8 jours après infection atteignant le rendement parasitaire
attendu vers J8.
Si les L3 sont transmises par le vecteur hématophage ou par injection sous-cutanée, la
plupart des larves sont retenues et détruites dans la peau du rongeur. Les L3 échapperaient à la
réponse inflammatoire de la peau en entrant dans le système lymphatique afférent [10,164]. Une
fois dans les lymphatiques, les L3 doivent passer les ganglions lymphatiques drainants pour
atteindre les vaisseaux lymphatiques efférents. La récupération des ganglions lymphatiques à
différents moments chez des rongeurs infectés par L. sigmodontis a révélé la présence
continue de L3 dans le système lymphatique dans les premiers jours de l'infection. Les L3
sont détectées dans les lymphatiques dès quelques heures après infection et pendant environ 5
jours, avec un pic entre 2 et 3 jours p.i. (Figure 28 et [10,11,204,356,357]).
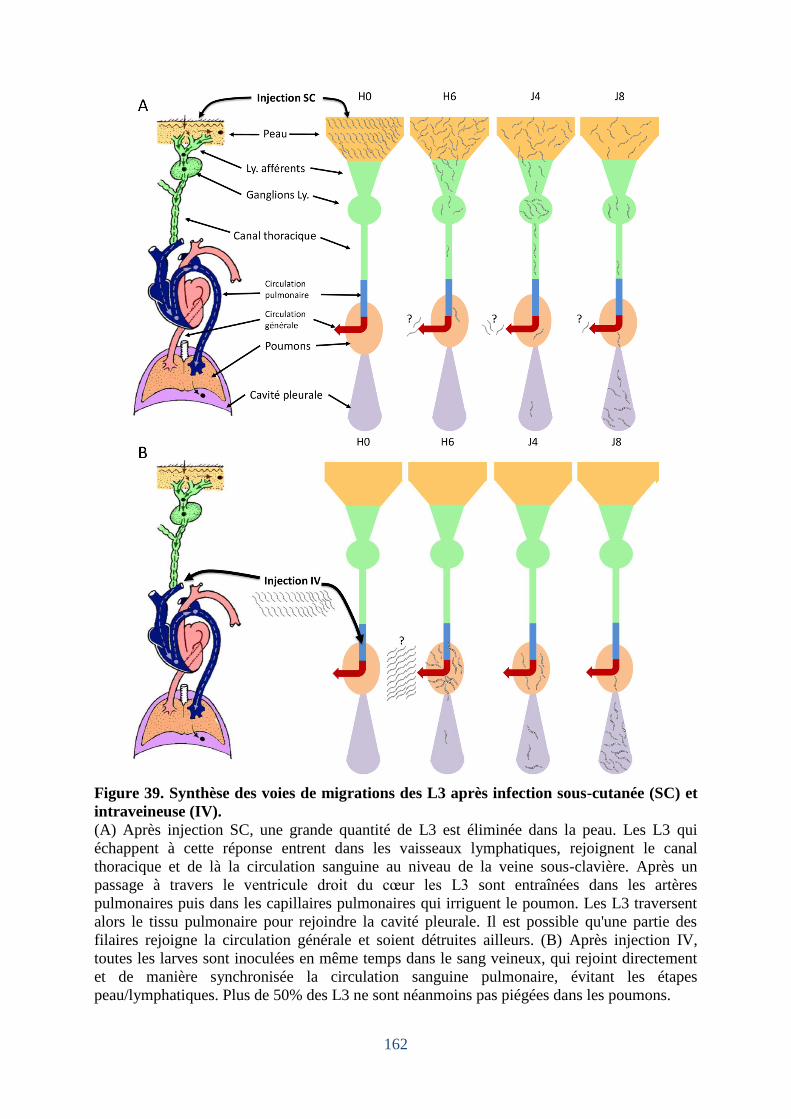
162
Figure 39. Synthèse des voies de migrations des L3 après infection sous-cutanée (SC) et
intraveineuse (IV).
(A) Après injection SC, une grande quantité de L3 est éliminée dans la peau. Les L3 qui
échappent à cette réponse entrent dans les vaisseaux lymphatiques, rejoignent le canal
thoracique et de là la circulation sanguine au niveau de la veine sous-clavière. Après un
passage à travers le ventricule droit du cœur les L3 sont entraînées dans les artères
pulmonaires puis dans les capillaires pulmonaires qui irriguent le poumon. Les L3 traversent
alors le tissu pulmonaire pour rejoindre la cavité pleurale. Il est possible qu'une partie des
filaires rejoigne la circulation générale et soient détruites ailleurs. (B) Après injection IV,
toutes les larves sont inoculées en même temps dans le sang veineux, qui rejoint directement
et de manière synchronisée la circulation sanguine pulmonaire, évitant les étapes
peau/lymphatiques. Plus de 50% des L3 ne sont néanmoins pas piégées dans les poumons.

163
Comprendre la migration des L3 de L. sigmodontis vers la cavité pleurale nécessite de
prendre en compte le système cardio-pulmonaire. L’anatomie de ce système représente une
voie logique d'arrivée dans la cavité pleurale comme le suggérait Wenk [11], et les résultats
d'autopsie semblent confirmer son implication. Des études antérieures [10,204] révélaient la
présence de L3 dans le compartiment cardio-pulmonaire dans les heures et les jours suivant
leur inoculation sous-cutanée. Cependant, la localisation précise (cœur ou poumons) n'avait
pas été déterminée spécifiquement. L'analyse du poumon nous a permis de démontrer la
présence de L3 dans ce tissu (Figure 27).
Comment expliquer la présence de L3 dans les poumons? La structure des valves
bicuspides des vaisseaux lymphatiques rend difficile les mouvements rétrogrades. Les valves
agissent comme des portes unidirectionnelles. Ainsi, une fois dans les vaisseaux
lymphatiques, les filaires n'auraient pas d'autre possibilité que de suivre le flux lymphatique.
Les vaisseaux lymphatiques se jettent dans le canal thoracique et le canal lymphatique droit
qui sont drainés dans les veines sous-clavières. Les L3 passeraient de la lymphe au sang à la
jonction du canal thoracique avec la veine sous-clavière. Après un passage à travers le
ventricule droit du cœur, le sang, et donc les L3, sont entraînés dans les artères pulmonaires
puis dans les capillaires pulmonaires qui irriguent le poumon, qui n'ont que 8-10 μm de
diamètre [305]. Les L3 sont des organismes de grande taille (750-800 μm de longueur pour un
diamètre de 10-12 μm à H0, le double à J8) affichant une musculature puissante permettant
leur motilité [361]. Ils peuvent également libérer des molécules excrétées/sécrétées [362], qui
pourraient faciliter leur passage des capillaires pulmonaires à travers le poumon jusqu'à la
cavité pleurale. Il est aussi possible qu'une partie des L3 arrivant dans les poumons y soit
piégée et soit incapable de se diriger vers la cavité pleurale. Une autre partie des L3 pourrait
ne pas être retenue dans les capillaires mais rejoidrait la circulation générale (Figure 39A).
Concernant cette deuxième hypothèse, aucune L3 n'a été retrouvée dans le sang, mais ces L3
pourraient être détruites dans les organes de nettoyage du sang tels que le foie ou la rate qui
sont fortement vascularisés. Il est aussi possible que ces L3 soient piégées dans les capillaires
des organes distants où elles ne pourraient pas se développer.

164
Pour contourner la peau et les étapes lymphatiques, un nouveau mode d'inoculation a
été utilisé pour infecter les souris (Figure 29). L'inoculation intraveineuse s'est révélée très
efficace comme le montre l'augmentation du rendement filarien. Deux fois plus de L3 ont été
récupérées dans la cavité pleurale des souris infestées en IV comparé à des souris infestées
naturellement ou en SC. Cependant, même chez les souris infestées en IV, plus de la moitié
des L3 est perdue après inoculation, ce qui étaye l'hypothèse selon laquelle une partie
importante des L3 est située ailleurs et ne se développe pas jusqu'au stade adulte. Il est aussi
possible que l'injection intraveineuse soit responsable d'une vasodilatation des capillaires
pulmonaires due à l'apport de liquide dans le sang, ce qui empêcherait certaines filaires d'être
bloquées. Dans le modèle IV, toutes les larves sont inoculées en même temps dans le sang
veineux, qui rejoint directement la circulation sanguine pulmonaire (Figure 39). Ceci explique
pourquoi tant de L3 sont observées dans les poumons dès les premières heures de l'infection.
En effet, en s'affranchissant des goulets d'étranglement que représentent la peau et le système
lymphatique, on favorise une arrivée rapide et synchronisée des L3 dans les poumons. Ce
modèle suggère également que la migration à travers le poumon peut être rapide car les larves
commencent à s'accumuler dans la cavité pleurale dès 2 heures après injection (Figure 29B)
Les différences observées entre les souris infestées en SC et IV pourraient être dues à
ce nombre plus élevé de L3 pulmonaires chez les souris IV. En ce qui concerne la pathologie,
il existe une forte corrélation entre le nombre d'hémorragies et le nombre de L3 récupéré dans
les poumons, le plus grand nombre d'hémorragies étant observé à H6 chez les souris infestées
en IV (Figure 30). L'obstruction des capillaires pulmonaires par les L3, ainsi que le
mouvement des L3, pourraient provoquer les zones hémorragiques observées dans les
poumons. Une telle corrélation n'est vraie ni pour la présence de granulomes ni pour
l'accumulation de neutrophiles dans l'espace périvasculaire des poumons. Ces deux
phénomènes sont observés beaucoup plus tard dans l'infection (J4 et J8), lorsque le nombre de
larves dans les poumons chez les souris infestées en IV ou SC est similaire.
Au cours de la migration des L3, deux phases distinctes peuvent être identifiées,
indépendamment du mode d'inoculation, et sans corrélation avec le nombre de filaires (Figure
40) :
La première, ayant lieu quelques heures après l'infection, se caractérise par une
augmentation des cytokines pro-inflammatoires (Figure 35 et Tableau 17) telles que l'IL-1β et
l'IL-33 (une cytokine cruciale pour la mise en place des réponses Th2 dans les muqueuses
[363,364]) et une augmentation des neutrophiles dans les capillaires pulmonaires [271]. Les

165
neutrophiles diminuent la survie dans les poumons des larves de Nippostrongylus brasiliensis
mais au prix d'une exacerbation des hémorragies et de l'inflammation pulmonaires [339]. Ces
différents effets sont probablement dus au choc oxydatif consécutif à l'activation des
neutrophiles, accompagnée par la libération de myéloperoxydase (MPO) [343].
La deuxième phase, qui se produit entre 4 et 8 jours après l'infection, se caractérise par
l'accumulation des L3 dans la cavité pleurale (Figure 27), la présence de granulomes
pulmonaires et de neutrophiles dans le PVS des poumons (Figures 31 et 32), et une
augmentation de neutrophiles, d'éosinophiles et de macrophages dans les espaces broncho-
alvéolaires et pleuraux (Figures 33 et 34). Ceci est associé à une augmentation des transcrits
pulmonaires de cytokines régulatrices telles que l'IL-9 ou les niveaux d'IL-4 et de MCP-1
(CCL2) dans le lavage bronchoalvéolaire. Les cellules lymphoïdes innées de type 2 (ILC2)
pourraient participer à la production de ces molécules et aux recrutements cellulaires : elles
sont présentes dans les poumons à l'homéostasie [365] et peuvent produire, après stimulation
par l'IL-33, de l'IL-4 et de l'IL-9 mais aussi de l'IL-5 et de l'IL-13 [366]. Elles participeraient
alors à l'activation alternative des macrophages et au recrutement des éosinophiles, qui ont des
effets antihelminthiques et un rôle dans la réparation des tissus [277,320,349,367].
Chez les souris infestées par L. sigmodontis, les infiltrats de neutrophiles observés
dans les capillaires pulmonaires à H6 sont résolus au plus tard à J4 [271]. Ceci concorde avec la
baisse observée à J4 de l'IL-1β et de l'expression des transcrits de cytokines pro-
inflammatoires chimiotactiques pour les neutrophiles, telles que CXCL1 et CXCL5 et dans
une moindre mesure l'IL-6 (Figure 35). Il a notamment été montré que le traitement par des
anticorps anti-CXCL5 atténue l'accumulation de neutrophiles pulmonaires dans un modèle
murin d'inflammation pulmonaire [368].
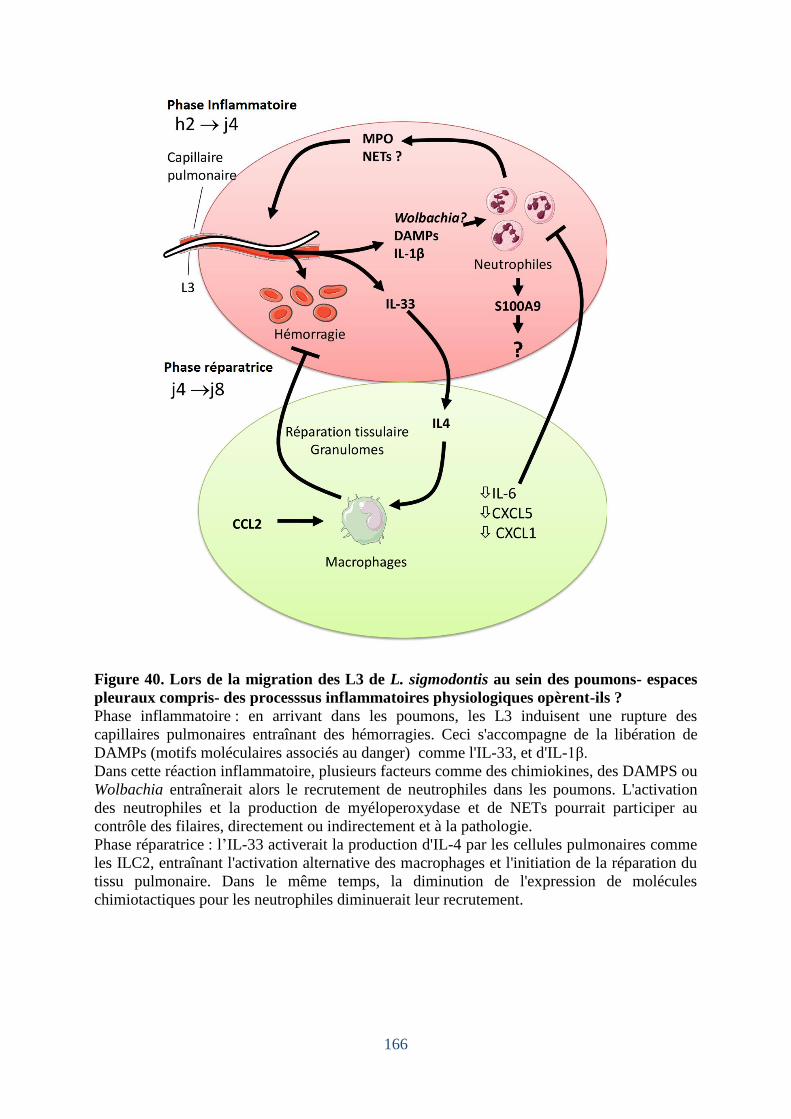
166
Figure 40. Lors de la migration des L3 de L. sigmodontis au sein des poumons- espaces
pleuraux compris- des processsus inflammatoires physiologiques opèrent-ils ?
Phase inflammatoire : en arrivant dans les poumons, les L3 induisent une rupture des
capillaires pulmonaires entraînant des hémorragies. Ceci s'accompagne de la libération de
DAMPs (motifs moléculaires associés au danger) comme l'IL-33, et d'IL-1β.
Dans cette réaction inflammatoire, plusieurs facteurs comme des chimiokines, des DAMPS ou
Wolbachia entraînerait alors le recrutement de neutrophiles dans les poumons. L'activation
des neutrophiles et la production de myéloperoxydase et de NETs pourrait participer au
contrôle des filaires, directement ou indirectement et à la pathologie.
Phase réparatrice : l’IL-33 activerait la production d'IL-4 par les cellules pulmonaires comme
les ILC2, entraînant l'activation alternative des macrophages et l'initiation de la réparation du
tissu pulmonaire. Dans le même temps, la diminution de l'expression de molécules
chimiotactiques pour les neutrophiles diminuerait leur recrutement.

167
Entre J4 et J8, les neutrophiles sont observés dans les espaces périvasculaires
pulmonaires et le liquide bronchoalvéolaire. Ces résultats suggèrent que les neutrophiles
circulent des capillaires vers les alvéoles et les PVS, et le rôle de ces espaces dans le contrôle
des populations de neutrophiles : après la migration des L3 de L. sigmodontis et la mise en
place des réponses régulatrices, les neutrophiles sanguins pourraient rejoindre les alvéoles et
les PVS pour être éliminés par les macrophages résidents. En effet, les macrophages
alvéolaires peuvent phagocyter les neutrophiles apoptotiques après l'inflammation pulmonaire
[369] et les macrophages interstitiels ont des capacités de phagocytose [318].
Après l'arrivée des L3 dans la cavité pleurale, les éosinophiles et les macrophages
pourraient cibler les larves infectantes. Cependant, ces cellules contribuent à la genèse du
granulome réactionnel contre les exuvies et non contre les L3/L4 [202]. De plus, le recrutement
des neutrophiles dans la cavité pleurale à ces temps précoces reste faible et n'est probablement
pas suffisant pour éliminer les L3.
Si les mêmes événements se succèdent chez les souris infestées en SC et en IV, la
cinétique de la réponse est légèrement différente : 1) les taux d'IL-1β mesurés à H6 dans la
cavité pleurale des souris IV sont plus élevés que ceux des souris infestées en SC. Ceci est
probablement la conséquence de l'afflux massif des L3 dans les poumons et de l'arrivée des
premières L3 dans la cavité pleurale ; 2) dans les poumons, la baisse plus importante des
transcrits de CXCL1 à J4 est accompagnée d'une baisse plus précoce du nombre de
neutrophiles dans le LBA, qui revient au niveau des souris naïves à J8. 3) Les infiltrats
d'éosinophiles sont détectés plus tôt chez les souris IV (J4 vs J8), ce qui indique que
l'initiation de la réponse pleurale est plus rapide chez les souris infestées en IV. Ces
différences suggèrent que la mise en place de la phase de régulation de l'inflammation serait
plus précoce chez les souris infestées en IV.
D'autres nématodes parasites présentent une migration pulmonaire similaire, en
particulier Nippostrongylus brasiliensis. Bien que N. brasiliensis soit un parasite gastro-
intestinal de rongeurs, ses L3 pénètrent dans la peau et migrent vers les poumons où se
produit la mue en L4. Des dégâts pulmonaires similaires à ceux que nous observons ont été
décrits chez les souris infestées par N. brasiliensis 23 h après infection, avec une résolution
vers le 7èmejour p.i. [320]. Une forte inflammation pulmonaire est observée dans l'infection par
ce nématode [320]. Elle est caractérisée par un recrutement de neutrophiles et la présence de
nombreuses cytokines inflammatoires comme l’IL-1β etl’IL-17 qui sont impliqués dans

168
l'augmentation du nombre de neutrophiles, mais aussi l’IL-4 et l’IL-5. Ces interleukines
pourraient modifier l’activation des macrophages alvéolaires en les orientant en AAM et ainsi
stopper l’avancée des lésions pulmonaires aiguës, notamment en limitant la production d'IL-
17 grâce aux molécules RELM-α, et Arginase-1 [342]. Il est intéressant de souligner que même
si ces deux espèces de nématodes sont très différentes (une filaire habitant dans les tissus vs
un strongle gastro-intestinal), une partie de la voie de migration des L3 est identique et semble
induire des réponses similaires (hémorragies et infiltrats de neutrophiles suivis d'une
réparation médiée par les macrophages AAM).
Les neutrophiles sont dotés d'un vaste répertoire de PRR (Pattern Recognition
Receptor/ récepteurs de reconnaissance de motifs moléculaires) permettant la reconnaissance
d'agents pathogènes ou de lésions tissulaires [370]. Parmi ces PRR, les neutrophiles expriment
tous les membres de la famille des récepteurs Toll-like (TLR), à l'exception du TLR3 [371] et
des capteurs cytoplasmiques tels que NOD-1, RIG1, MDA5 et IFI16 [370,372,373]. La détection
des agents pathogènes et des lésions tissulaires par ces PRR active les fonctions effectrices
des neutrophiles comme la phagocytose des micro-organismes, la production de réactifs
intermédiaires de l'oxygène, de cytokines, d'enzymes lytiques et de peptides antimicrobiens
[374]. Un autre mécanisme d'action des neutrophiles est la formation de NETs (Neutrophil
Extracellular Traps) ou nétose. La formation NETs ou nétose est un processus graduel
impliquant notamment la génération de réactifs de l'oxygène et la libération extracellulaire de
MPO et de la chromatine [375,376]. Les NETs peuvent capturer des bactéries, des protozoaires et
des virus [377]. Toutefois, les neutrophiles peuvent détecter la taille des microorganismes et
libérer des NETs en présence de pathogènes de grande taille comme les champignons [378]
[379]. Les NET peuvent également immobiliser les larves du nématode Strongyloides
stercoralis facilitant l'élimination des parasites par les cellules du système immunitaire [380].
La destruction des Mfs de B. malayi par des neutrophiles humains in vitro ne fait pas
intervenir la production de NETs [381]. Cependant, les neutrophiles murins en présence de
Litomosoides sigmodontis in vitro libèrent de la MPO et de l'ADN extracellulaire en quelques
heures (Figure 37) et génèrent des réactifs de l'oxygène [257], trois éléments signature de
nétose. L'observation des cultures en microscopie électronique à balayage (Figure 38) ou en
microscopie à fluorescence après marquage de l'ADN extracellulaire [257] montre
effectivement la libération de chromatine par les neutrophiles. Dans les conditions étudiées, la
présence des neutrophiles et la formation des NETs ne semblent pas suffisantes pour piéger
les filaires. En effet, les NETs sont observés à distance des filaires (Figure 38). Il semble

169
probable que les neutrophiles aient été activés par des molécules excrétées par les filaires, par
un contact direct avec les L3 qui sont très mobiles en culture ou via leur endosymbionte
Wolbachia. Un mécanisme de nétose induit par Wolbachia a en effet été démontré dans
l'onchocercose humaine [382].
En conclusion, ces résultats suggèrent que les L3 peuvent induire la nétose chez les
neutrophiles mais la présence de NETs in vivo n'est pas clairement démontrée.

170

171
CHAPITRE 2 :
L’héterodimère S100A8/A9 - calprotectine – favorise-t-il la survie de Litomosoides sigmodontis ?

172
Introduction
Le contenu en protéines du cytosol des neutrophiles est constitué pour 40% de deux
molécules : S100A8 et S100A9, qui sont donc les protéines les plus abondantes dans ces
cellules [383]. Ces molécules sont retrouvées en grandes quantités dans le milieu extracellulaire
lors de l'inflammation et sont souvent associées aux pathologies pulmonaires [384-387]. Elles ont
aussi été retrouvées dans les nodules onchocerquiens contenant des neutrophiles [57]. Dans les
poumons, l'inhalation de S100A9 entraîne un recrutement de neutrophiles en induisant une
augmentation de l'expression de CXCL-10 [388] ; à l'inverse, en cas d'inflammation pulmonaire
due au LPS, S100A9 et l'hétérodimère S100A8/S100A9 diminuent ce recrutement en
réduisant l'expression de gènes codants pour des cytokines comme le TNF-α, l'IL-1β ou l'IL-6
[388]. S100A8 diminue aussi les symptômes asthmatiques en contribuant à l’élimination des
réactifs de l'oxygène nécessaires à l'activation des mastocytes, lesquels en dégranulant libèrent
de l'histamine et de nombreuses cytokines inflammatoires chimiotactiques pour les
éosinophiles [389]. S100A8 et S100A9 sont également présentes dans les monocytes où elles
représentent 5% de leurs protéines ; elles sont absentes des macrophages et des lymphocytes
dans les tissus. L'expression de S100A8 et S100A9 peut être induite sous conditions
inflammatoires dans d'autres types cellulaires comme les kératinocytes ou les cellules
épithéliales [390-393]. De plus, une augmentation du S100A8 et du S100A9 est observée dans
les cellules tumorales de nombreux cancers [387,394-396].
Les protéines S100 tirent leur nom de leur parfaite solubilité dans une solution de
sulfate d’ammonium saturé à 100% [397]. Actuellement 25 protéines S100 ont pu être
identifiées et constituent une famille au sein des calciprotéines à motif EF-hand [398]. Les
motifs EF-hand, composés de deux hélices alpha reliées par une boucle, sont connectés par
une région intermédiaire faisant office de charnière [399] et permettent la fixation du calcium.
Les protéines S100 sont multifonctionnelles et jouent un rôle dans la régulation d’un grand
nombre d’activités cellulaires. Elles interagissent avec de nombreuses protéines effectrices
dans la cellule régulant ainsi des activités enzymatiques, des facteurs de transcription, la
phosphorylation de protéines, la dynamique du cytosquelette, la croissance des cellules et leur
différenciation ainsi que l’homéostasie du calcium. Dans le milieu extracellulaire, elles
exercent des fonctions proches des cytokines et montrent des activités régulatrices sur les
cellules inflammatoires, les neurones, les astrocytes, les microglies, les cellules endothéliales
et épithéliales. Les protéines S100 participent à la migration cellulaire et la chimiotaxie, ainsi
qu’au développement et à la réparation des tissus ([400-402] et Figure 41).

173
Figure 41. Les protéines de la famille S100, une famille détectée chez les vertébrés
Les protéines S100 sont impliquées dans de nombreux processus cellulaires comme la
différenciation et la croissance cellulaire, la migration ou encore l'apoptose. D'après [403].
Jusqu’ici les protéines S100 ont été isolées à partir de vertébrés [404,405] et l’analyse des
génomes disponibles d’invertébrés ne révèle aucune séquence similaire aux gènes des
protéines S100 dans ces organismes. Ainsi, ces protéines représenteraient un groupe
phylogénétique récent. Chez l’homme, la plupart des gènes des protéines S100 sont regroupés
dans la région 1q21 du chromosome 1 [406]. La structure de ces gènes est hautement conservée
[427]. La proximité physique de ces gènes est à l’origine de leur nomenclature officielle [407] ; la
lettre A correspondant au complexe multigénique et le chiffre qui suit à la position relativ de
chaque gène [408].
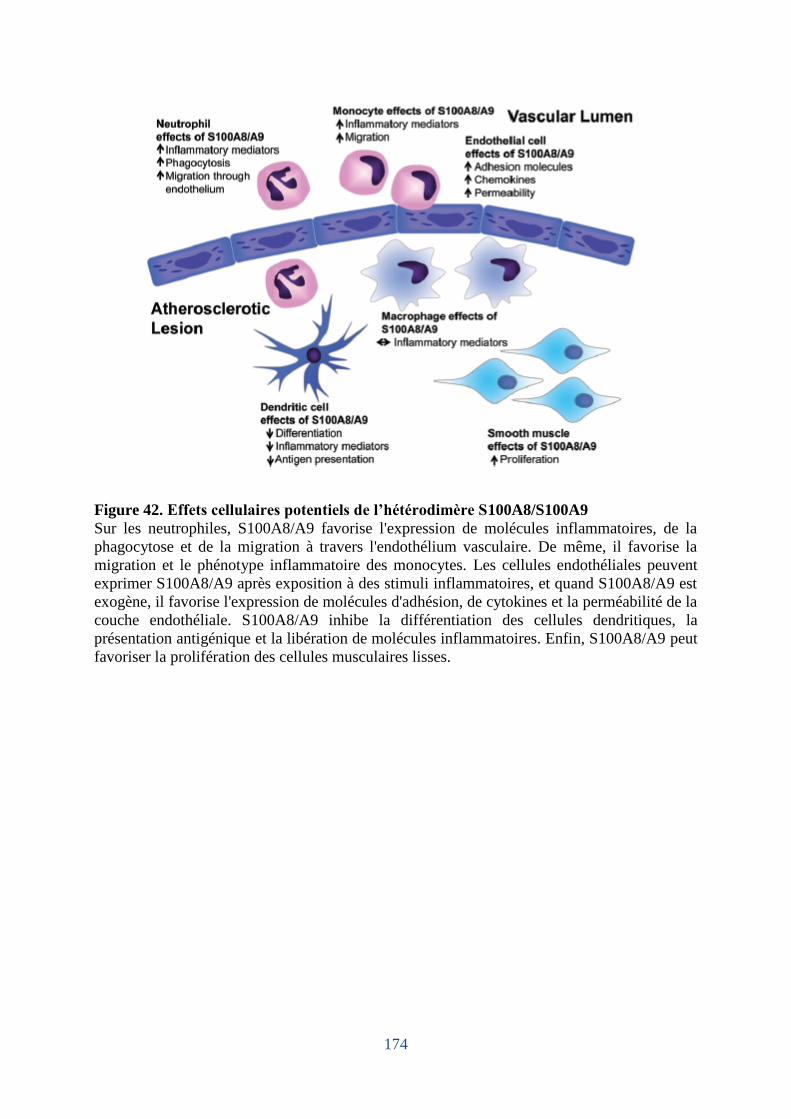
174
Figure 42. Effets cellulaires potentiels de l’hétérodimère S100A8/S100A9
Sur les neutrophiles, S100A8/A9 favorise l'expression de molécules inflammatoires, de la
phagocytose et de la migration à travers l'endothélium vasculaire. De même, il favorise la
migration et le phénotype inflammatoire des monocytes. Les cellules endothéliales peuvent
exprimer S100A8/A9 après exposition à des stimuli inflammatoires, et quand S100A8/A9 est
exogène, il favorise l'expression de molécules d'adhésion, de cytokines et la perméabilité de la
couche endothéliale. S100A8/A9 inhibe la différentiation des cellules dendritiques, la
présentation antigénique et la libération de molécules inflammatoires. Enfin, S100A8/A9 peut
favoriser la prolifération des cellules musculaires lisses.

175
Les protéines S100 sont de petites protéines agencées en dimères [398] et dont le
poids moléculaire d’une sous-unité est compris entre 9 et 14 kilodaltons [409,410]. La plupart
des S100 (S100A4, A6, A7, A8, A10, A11, A12 et S100B) existent sous forme
d’homodimères. Cependant, elles peuvent aussi former des hétérodimères, comme c’est le cas
pour S100A1 et S100B, S100A1 et S100A4, S100A6 et S100B, S100A11 et S100B ainsi que
S100A8 et S100A9 [411-414].
S100A8 et S100A9 forment des hétérodimères associés de manière non covalente,
également appelés calprotectine [415].
Des souris déficientes en S100A8 (S100A8-/-) et en S100A9 (S100A9-/-) ont été produites
[355,416]. Si l'absence de S100A8 est létale pour les embryons [416], la perte de S100A9 conduit à
des souris viables et fertiles [355]. L'absence de S100A9 conduit à une diminution de 30% de
cellules exprimant le S100A8 dans la moelle osseuse et le S100A8 n'est détecté dans aucune
cellule périphérique. Les transcrits d'ARN messager de s100a8 sont détectés dans les cellules
des souris déficientes en S100A9 suggérant une régulation post-transcriptionelle de s100a8
[355]. L'hétérodimère S100A8/S100A9 est donc aussi absent de ces souris. Cependant
l’absence de S100A8 et de S100A9, qui représentent près de 40% des protéines cytosoliques
des neutrophiles normaux, n'affecte pas le développement de neutrophiles viables et matures
[355].
La calprotectine intracellulaire interagit avec le cytosquelette. En présence de calcium,
des tétramères (S100A8/S100A9)2 stabilisent les filaments de tubuline en favorisant la
polymérisation de la tubuline et le regroupement des microtubules [417]. Cette interaction joue
un rôle crucial dans le processus de migration des neutrophiles vers les zones inflammatoires
[418,419]. En plus de ces effets intracellulaires, S100A8 et S100A9 font partie des médiateurs
endogènes exprimés à la suite d'une agression d'origine septique ou non (alarmines) et ont une
activité pro-inflammatoire sur de nombreux types cellulaires, comme les cellules
endothéliales, les monocytes ou les neutrophiles (Figure 42). Les neutrophiles libèrent
S100A8 et S100A9 dans l'espace extracellulaire via des mécanismes passifs et actifs. Leur
libération passive est souvent la conséquence de dommages tissulaires, de la nécrose des
cellules, ou de la formation de NETs (Neutrophil extracellular Traps) [398,420,421]. En revanche,
la sécrétion des complexes S100A8/S100A9 par les phagocytes activés pendant les processus
inflammatoires est un processus spécifique qui est induit lors de leur contact avec
l'endothélium enflammé [398]. Le S100A8/S100A9 extracellulaire stimule l’adhésion et la
transmigration des neutrophiles à travers les cellules endothéliale de plusieurs manières :

176
(1) il active le TLR-4 des neutrophiles qui entraine l'expression des intégrines β2 à leur
surface [422-425]); (2) il contribue à augmenter l’expression d’ICAM-1 (qui fixe l'intégrine β2)
et de VCAM-1 à la surface des cellules endothéliales [418,426] (3) il entraîne la diminution de
l'expression des molécules de jonctions cellulaires, induisant une perméabilité accrue de la
monocouche endothéliale [427,428]. En outre, S100A8/S100A9 régule la survie cellulaire et le
nombre de neutrophiles par un effet anti-apoptotique [429].
Outre leurs propriétés pro-inflammatoires, S100A8/S100A9 présentent des propriétés
régulatrices pour limiter les réponses immunitaires exacerbées et les dommages associés. Les
neutrophiles activés produisent de grandes quantités de réactifs oxygénés pouvant mener à
d’importants dommages cellulaires et tissulaires. S100A8 et S100A9 possèdent des propriétés
antioxydantes et peuvent prendre en charge les ROS (Reactive oxygen species) et les NO
(oxydes nitriques) [430-436]). L'hétérodimère affiche aussi une affinité de liaison élevée pour les
ions Zn2+ et Mn2+ en plus de Ca2+ [437]. La liaison de S100A8/S100A9 à Zn2+ et Mn2+ pourrait
participer à son action antimicrobienne, en chélatant ces ions dans l'espace extracellulaire, les
rendant indisponibles aux bactéries [437,438].
S100A8/S100A9 peut aussi avoir un effet immunosuppresseur sur les cellules
phagocytaires : la pré-stimulation de monocytes avec du S100A8/S100A9 induirait leur
inhibition d'une manière TLR4-dépendante, entraînant l'absence de réponse à une stimulation
ultérieure avec du LPS [439]. De plus, des expériences sur des souris déficientes en
l'hétérodimère S100A8/S100A9 indiquent qu'il pourrait bloquer la différenciation des cellules
dendritiques, l'expression des molécules de co-stimulation (CD80 et CD86) et la présentation
antigénique, entraînant une diminution de la réponse des cellules T [440,441]. Dans la cavité
péritonéale, S100A9 induit une baisse des capacités phagocytaires des macrophages [442] qui
serait due, au moins en partie, à la production d'IL-10 par les cellules B1 [443]. Enfin, la
phagocytose des neutrophiles apoptotiques par les macrophages diminuerait leur activation
d'une manière S100A9 dépendante [444].
Des études de résonance plasmonique de surface indiquent qu’en présence de Ca2+ ou
de Zn2+, S100A9 possède une grande affinité pour le Receptor for Advanced Glycation End-
products (RAGE) alors que S100A8 n’en possède aucune [445]. Ces données suggèrent que
S100A9 pourrait être le ligand principal de RAGE. Plusieurs autres études ont révélé
l’association de S100A8 ou S100A9 avec d’autres récepteurs cellulaires. Les protéoglycans
avec des sulfates héparinés représentent un site de liaison potentiel pour S100A8/S100A9. En
effet, S100A9 possède une forte affinité pour les sulfates d’héparines/héparanes présents à la

177
surface des cellules endothéliales [446]. Le récepteur scavenger CD36 a également été proposé
comme récepteur possible de S100A8/S100A9 [447,448]). Enfin, le TLR-4 a été suggéré comme
récepteur principal de S100A8, alors que le complexe S100A8/S100A9 avait une activité
réduite [449,450]. Une étude a aussi montré que S100A9 se liait avec le CD147 à la surface de
mélanomes métastatiques, ce qui favorisait la production de cytokines inflammatoires et la
libération de MMP [451].
Le rôle de S100A8/S100A9 dans les infections filariennes n'est pas connu. Les
résultats récents de Karadjian [271] suggèrent que S100A8 et S100A9 pourraient être des
marqueurs d’inflammation dans l’infection filarienne avec L. sigmodontis chez la souris
BALB/c. Du S100A8/S100A9 est libéré par les neutrophiles lors de la nétose [379]. Il pourrait
donc participer aux réponses dans la peau et dans les poumons des souris infestées (Chapitre
1). De plus, l'infection par L. sigmodontis de souris déficientes en TLR4 (TLR4-/-), un des
recepteurs de S100A8 et S100A9, a montré un rôle de ce récepteur dans l'initiation de la
microfilarémie [288,289]. Or la stimulation in vitro avec des extraits de filaires de lignées
cellulaires transfectées avec différents TLR, ou de macrophages provenant de souris TLR2 ko
ou TLR4 ko a montré que les réponses à Brugia malayi et Onchocerca volvulus dépendent de
TLR2, mais pas de TLR4 [267]. S100A8 et/ou S100A9, par le biais du TLR4, pourraient donc
moduler les réponses antifilariennes.
Ce chapitre présente les résultats des expériences en cours visant à préciser le rôle de S100A9
lors de la phase pré-patente de l'infection par L. sigmodontis.
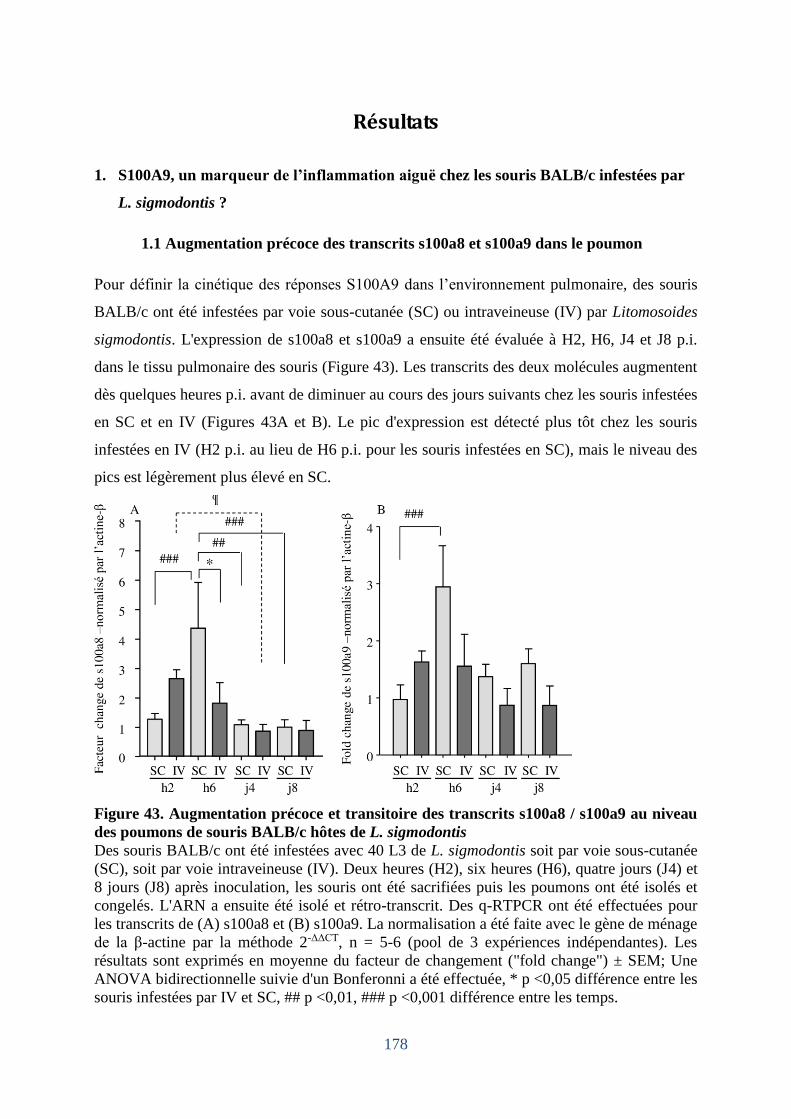
178
Résultats
1. S100A9, un marqueur de l’inflammation aiguë chez les souris BALB/c infestées par
L. sigmodontis ?
1.1 Augmentation précoce des transcrits s100a8 et s100a9 dans le poumon
Pour définir la cinétique des réponses S100A9 dans l’environnement pulmonaire, des souris
BALB/c ont été infestées par voie sous-cutanée (SC) ou intraveineuse (IV) par Litomosoides
sigmodontis. L'expression de s100a8 et s100a9 a ensuite été évaluée à H2, H6, J4 et J8 p.i.
dans le tissu pulmonaire des souris (Figure 43). Les transcrits des deux molécules augmentent
dès quelques heures p.i. avant de diminuer au cours des jours suivants chez les souris infestées
en SC et en IV (Figures 43A et B). Le pic d'expression est détecté plus tôt chez les souris
infestées en IV (H2 p.i. au lieu de H6 p.i. pour les souris infestées en SC), mais le niveau des
pics est légèrement plus élevé en SC.
Figure 43. Augmentation précoce et transitoire des transcrits s100a8 / s100a9 au niveau
des poumons de souris BALB/c hôtes de L. sigmodontis
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis soit par voie sous-cutanée
(SC), soit par voie intraveineuse (IV). Deux heures (H2), six heures (H6), quatre jours (J4) et
8 jours (J8) après inoculation, les souris ont été sacrifiées puis les poumons ont été isolés et
congelés. L'ARN a ensuite été isolé et rétro-transcrit. Des q-RTPCR ont été effectuées pour
les transcrits de (A) s100a8 et (B) s100a9. La normalisation a été faite avec le gène de ménage
de la β-actine par la méthode 2-ΔΔCT, n = 5-6 (pool de 3 expériences indépendantes). Les
résultats sont exprimés en moyenne du facteur de changement ("fold change") ± SEM; Une
ANOVA bidirectionnelle suivie d'un Bonferonni a été effectuée, * p <0,05 différence entre les
souris infestées par IV et SC, ## p <0,01, ### p <0,001 différence entre les temps.
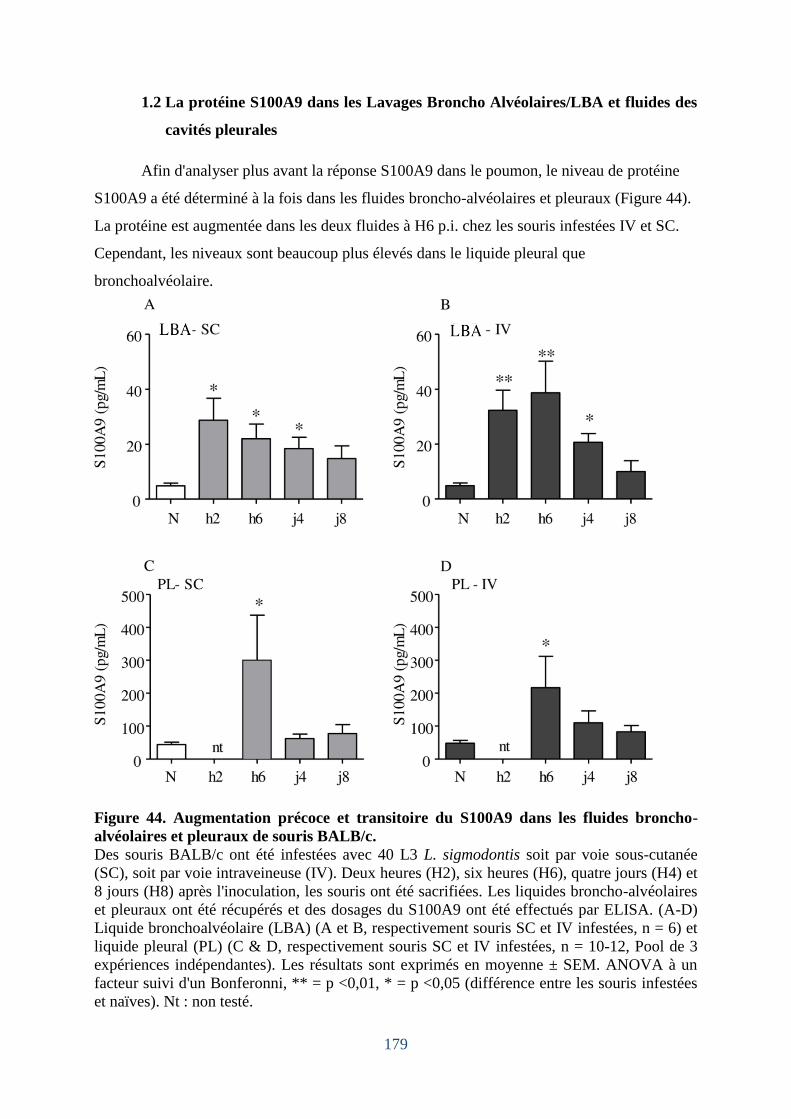
179
1.2 La protéine S100A9 dans les Lavages Broncho Alvéolaires/LBA et fluides des
cavités pleurales
Afin d'analyser plus avant la réponse S100A9 dans le poumon, le niveau de protéine
S100A9 a été déterminé à la fois dans les fluides broncho-alvéolaires et pleuraux (Figure 44).
La protéine est augmentée dans les deux fluides à H6 p.i. chez les souris infestées IV et SC.
Cependant, les niveaux sont beaucoup plus élevés dans le liquide pleural que
bronchoalvéolaire.
Figure 44. Augmentation précoce et transitoire du S100A9 dans les fluides broncho-
alvéolaires et pleuraux de souris BALB/c.
Des souris BALB/c ont été infestées avec 40 L3 L. sigmodontis soit par voie sous-cutanée
(SC), soit par voie intraveineuse (IV). Deux heures (H2), six heures (H6), quatre jours (H4) et
8 jours (H8) après l'inoculation, les souris ont été sacrifiées. Les liquides broncho-alvéolaires
et pleuraux ont été récupérés et des dosages du S100A9 ont été effectués par ELISA. (A-D)
Liquide bronchoalvéolaire (LBA) (A et B, respectivement souris SC et IV infestées, n = 6) et
liquide pleural (PL) (C & D, respectivement souris SC et IV infestées, n = 10-12, Pool de 3
expériences indépendantes). Les résultats sont exprimés en moyenne ± SEM. ANOVA à un
facteur suivi d'un Bonferonni, ** = p <0,01, * = p <0,05 (différence entre les souris infestées
et naïves). Nt : non testé.
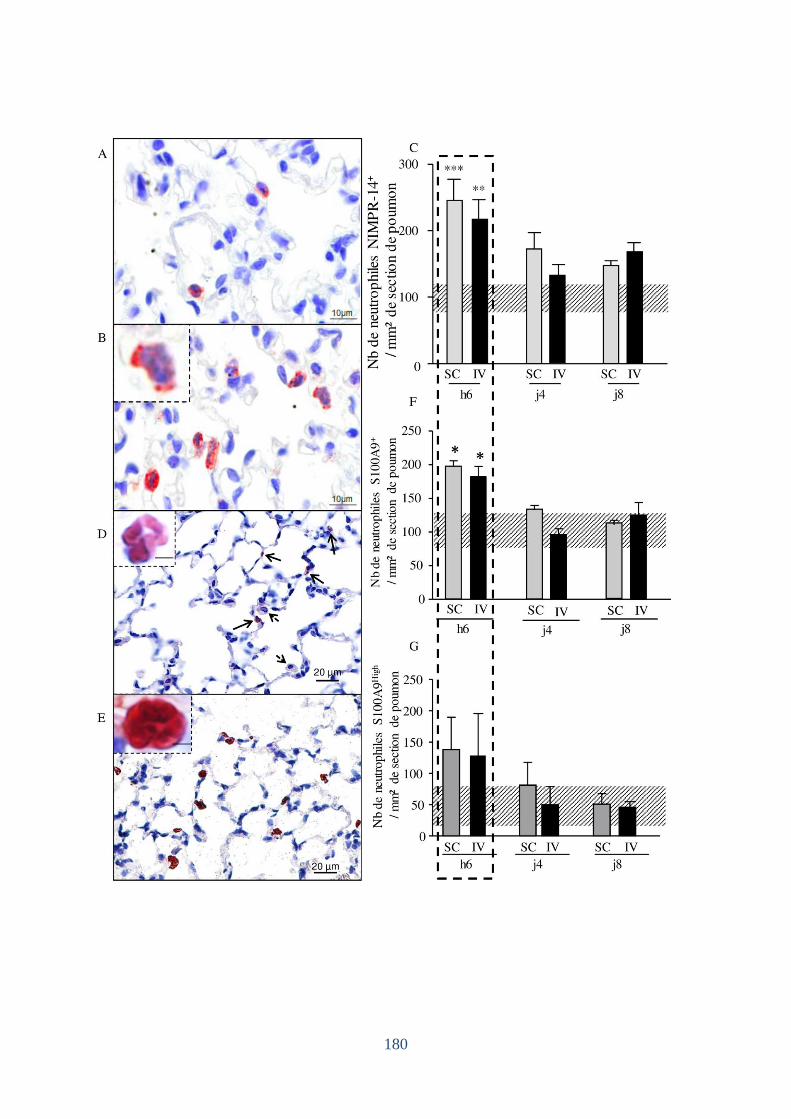
180

181
1.3 Augmentation du nombre de neutrophiles S100A9+ dans les poumons
Dans les premières heures suivant l'inoculation des L3, des neutrophiles sont observés
dans le tissu pulmonaire, le plus souvent dans les capillaires pulmonaires et plus rarement
dans l'espace alvéolaire (Figures 45A et B). Le comptage des neutrophiles révèle une
augmentation transitoire de ces cellules à H6 p.i. chez les souris infestées en SC et IV (Figure
3C). Le marquage spécifique de S100A9 sur des sections pulmonaires indique que toutes les
cellules positives pour S100A9 sont morphologiquement identifiables comme des
neutrophiles (Figure 45D et E) et qu'un grand pourcentage des neutrophiles exprime le
S100A9 (Figure 45F). On distingue deux états en fonction du contenu en S100A9 des
neutrophiles, qui est soit faible (Figure 45D, zoom), soit élevé (Figure 45E, zoom). Le
comptage des neutrophiles S100A9+ (Figure 45F) et de neutrophiles S100A9high (Figure 45G)
révèle un pic de neutrophiles S100A9+ à H6 p.i. chez les souris infestées en SC et IV avec une
majorité de neutrophiles S100A9high (74% et 70% respectivement). Les neutrophiles S100A9+
sont absents des granulomes pulmonaires précédemment décrits (Figure 46 et Chapitre 1)
mais sont présents dans le tissu environnant.
<- Figure 45. Augmentation précoce et transitoire de neutrophiles exprimant S100A9
dans les poumons de souris BALB/c.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis soit par voie sous-cutanée
(SC), soit par voie intraveineuse (IV). Des sections pulmonaires ont été analysées six heures
(h6), quatre jours (j4) et 8 jours (j8) après l'inoculation. (A) Immunomarquage du Ly6G/C
(clone NIMPR-R14) des sections pulmonaires a été réalisée montrant quelques neutrophiles
interstitiels chez des souris naïves (A) et une augmentation des neutrophiles chez la souris
infestée par h6 (B). Les neutrophiles ont été différenciés des monocytes par la forme de leurs
noyaux (B, zoom). (C) Nombre de neutrophiles par mm² sur les sections immunomarquées
pour le Ly6G/C; Les barres représentent la moyenne ± SEM, n = 4-6 (pool de 3 expériences
indépendantes), ANOVA à deux facteurs suivi d'un Bonferonni (** p <0,01, *** p <0,001).
Les souris non infestées sont représentées par une barre horizontale pointillée. (D-G)
Immunomarquage du S100A9 (clone MU14-2A5) sur des sections pulmonaires a été réalisée
montrant (D) une faible coloration de S100A9 dans les neutrophiles (flèches noires). Les
macrophages alvéolaires (flèches pointillées) sont S100A9-. Coin supérieur gauche: zoom sur
un neutrophile S100A9+, (E) coloration importante de S100A9 dans les neutrophiles. Coin
supérieur gauche: zoom sur un neutrophile S100A9high. (F) Nombre total de neutrophiles
S100A9+ et (G) nombre de neutrophiles S100A9high par mm² de poumon. Les souris non
infestées sont représentées par une barre horizontale pointillée. Les résultats sont exprimés en
moyenne ± SEM ; N = 4 souris, 2-3 lames par souris et 2 à 4 sections par lames ont été
analysées.
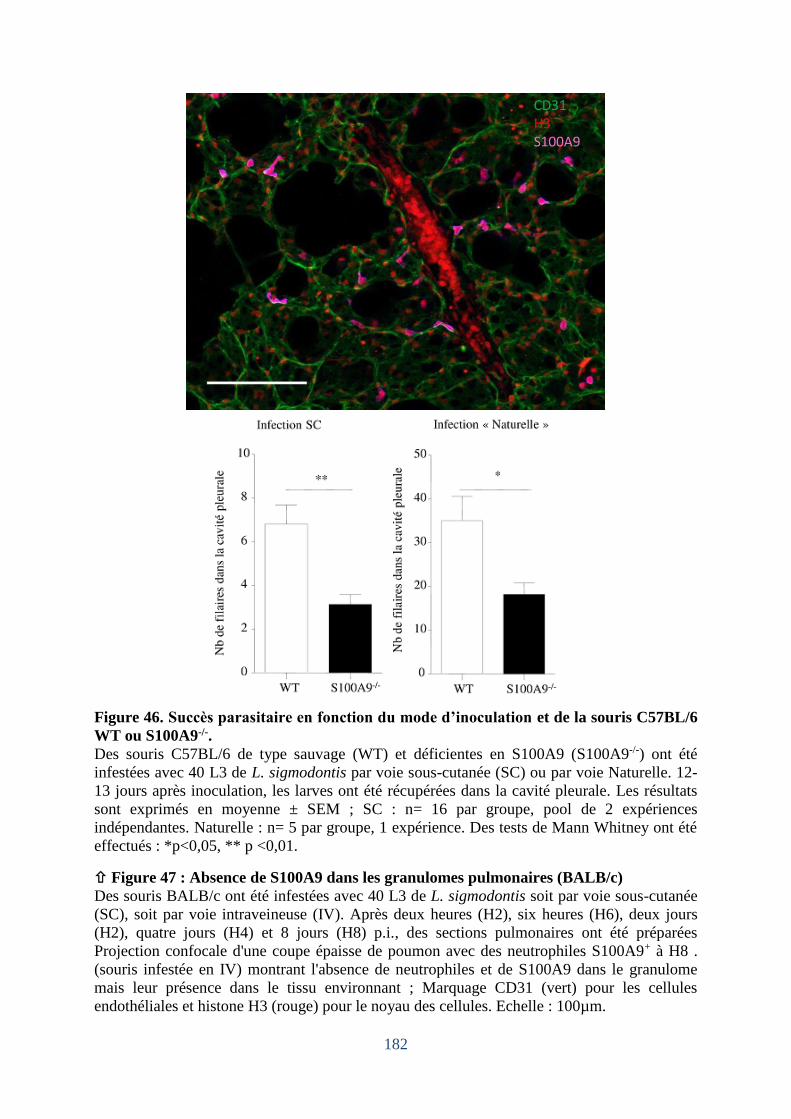
182
Figure 46. Succès parasitaire en fonction du mode d’inoculation et de la souris C57BL/6
WT ou S100A9-/-.
Des souris C57BL/6 de type sauvage (WT) et déficientes en S100A9 (S100A9-/-) ont été
infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée (SC) ou par voie Naturelle. 12-
13 jours après inoculation, les larves ont été récupérées dans la cavité pleurale. Les résultats
sont exprimés en moyenne ± SEM ; SC : n= 16 par groupe, pool de 2 expériences
indépendantes. Naturelle : n= 5 par groupe, 1 expérience. Des tests de Mann Whitney ont été
effectués : *p<0,05, ** p <0,01.
Figure 47 : Absence de S100A9 dans les granulomes pulmonaires (BALB/c)
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis soit par voie sous-cutanée
(SC), soit par voie intraveineuse (IV). Après deux heures (H2), six heures (H6), deux jours
(H2), quatre jours (H4) et 8 jours (H8) p.i., des sections pulmonaires ont été préparées
Projection confocale d'une coupe épaisse de poumon avec des neutrophiles S100A9+ à H8 .
(souris infestée en IV) montrant l'absence de neutrophiles et de S100A9 dans le granulome
mais leur présence dans le tissu environnant ; Marquage CD31 (vert) pour les cellules
endothéliales et histone H3 (rouge) pour le noyau des cellules. Echelle : 100µm.

183
2. S100A9, un acteur dans le contrôle de l’établissement des L3 de L. sigmodontis ?
Analyse des souris C57BL/6 déficientes en s100a9
2.1 L’absence de s100a9 se traduit par un moindre établissement des larves L3
Pour établir le rôle du S100A9 dans l'infection par L. sigmodontis, des souris
déficientes en S100A9 de fond génétique C57BL/6 ont été utilisées. La perte de S100A9
conduit à des souris viables [355]. L'absence de S100A9 conduit à une diminution de 30% de
cellules exprimant le S100A8 dans la moelle osseuse et le S100A8 n'est détecté dans aucune
cellule périphérique. Bien que les transcrits d'ARN messager de s100a8 soient détectés dans
les cellules des souris déficientes en S100A9, la protéine n'est pas produite [355].
L'hétérodimère S100A8/S100A9 est donc aussi absent de ces souris.
Des souris de type sauvages (WT) ou des souris déficientes en S100A9 (S100A9-/-) ont
été infestées soit par transmission des L3 par l'acarien vectoriel O. bacoti (infection
"Naturelle") ou par inoculation de 40 L3 en SC. Le succès parasitaire a été évalué vers 12-13
jours p.i., chez des jeunes L4 (Figure 47) et révèle une forte baisse du nombre de L4 dans la
cavité pleurale des souris déficientes en S100A9 dans les deux modes d’inoculation (infection
SC et Naturelle), suggérant un rôle du S100A9 favorisant la survie des parasites. Lors de
l'infection naturelle, le nombre de parasites inoculé n'est pas connu et dépend du taux
d'infection des acariens. Le nombre de parasites récupéré varie donc d'une expérience à
l'autre. Aucun parasite mort ni aucun granulome n'a été retrouvé à ce temps d’autopsie, ce qui
suggère que l’effet filaricide pourrait être antérieur, potentiellement pendant la phase de
migration des L3.
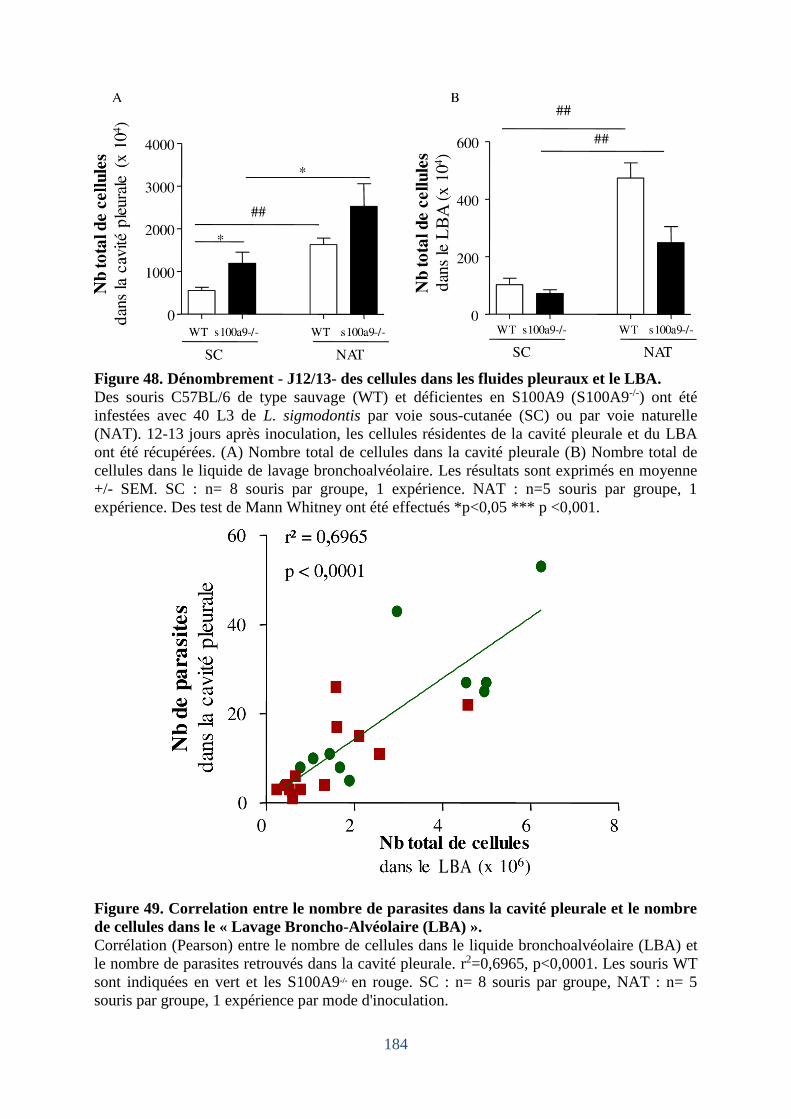
184
Figure 48. Dénombrement - J12/13- des cellules dans les fluides pleuraux et le LBA.
Des souris C57BL/6 de type sauvage (WT) et déficientes en S100A9 (S100A9-/-) ont été
infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée (SC) ou par voie naturelle
(NAT). 12-13 jours après inoculation, les cellules résidentes de la cavité pleurale et du LBA
ont été récupérées. (A) Nombre total de cellules dans la cavité pleurale (B) Nombre total de
cellules dans le liquide de lavage bronchoalvéolaire. Les résultats sont exprimés en moyenne
+/- SEM. SC : n= 8 souris par groupe, 1 expérience. NAT : n=5 souris par groupe, 1
expérience. Des test de Mann Whitney ont été effectués *p<0,05 *** p <0,001.
Figure 49. Correlation entre le nombre de parasites dans la cavité pleurale et le nombre
de cellules dans le « Lavage Broncho-Alvéolaire (LBA) ».
Corrélation (Pearson) entre le nombre de cellules dans le liquide bronchoalvéolaire (LBA) et
le nombre de parasites retrouvés dans la cavité pleurale. r2=0,6965, p<0,0001. Les souris WT
sont indiquées en vert et les S100A9-/- en rouge. SC : n= 8 souris par groupe, NAT : n= 5
souris par groupe, 1 expérience par mode d'inoculation.

185
2.2 L’absence de S100A8/A9 se traduit par un moindre recrutement cellulaire
dans le LBA mais facilite celui dans la cavité pleurale
Les réponses cellulaires dans la cavité pleurale et dans l'espace bronchoalvéolaire ont
été étudiées chez les souris S100A9-/- (Figure 48).
Le nombre total de cellules chez les souris infestées en SC est toujours inférieur à celui des
souris infestées naturellement via les vecteurs, que ce soit dans la cavité pleurale ou dans
l’espace bronchoalvéolaire (Figure 48). Le nombre de cellules pleurales est plus important
chez les souris S100A9-/- (Figure 48A) alors que le nombre de cellules dans l’espace
bronchoalvéolaire est plus faible chez les souris S100A9-/- (Figure 48B), et ceci quel que soit
le mode d’inoculation.
En fait, le nombre de cellules dans le LBA est corrélé au nombre de parasites ayant
rejoint la cavité pleurale (r2=0,6965 et p<0,0001), indépendamment du mode d’inoculation et
de la souche murine (Figure 49). Comme les charges parasitaires sont plus élevées en
infestation naturelle ceci explique la différence observée. De plus les charges parasitaires
étant plus faibles dans les souris S100A9-/- le nombre de cellules est donc légèrement plus
faible dans le LBA des souris S100A9-/- (Figure 48B).
Une telle corrélation entre le nombre de cellules et le nombre de filaires n’est pas observée
dans la cavité pleurale (r2=0,0140 et p=0,5639) où malgré un nombre plus faible de parasites
chez les souris S100A9-/-, le nombre de cellules pleurales augmente.
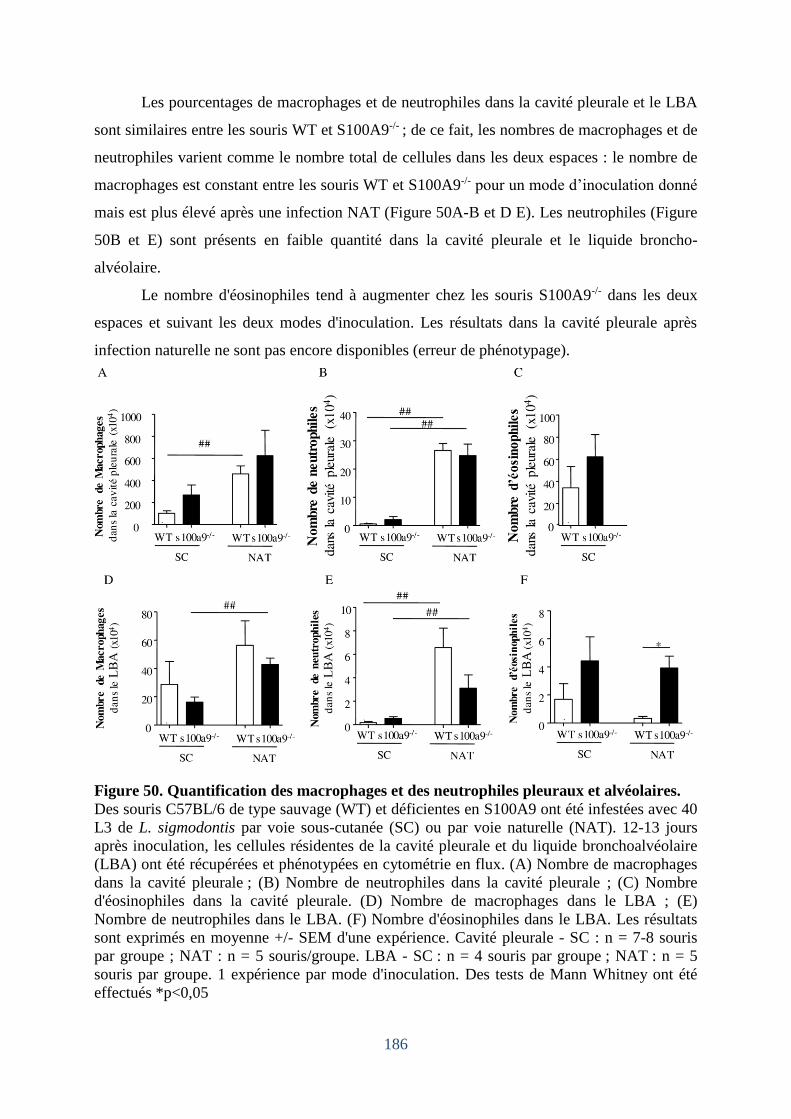
186
Les pourcentages de macrophages et de neutrophiles dans la cavité pleurale et le LBA
sont similaires entre les souris WT et S100A9-/- ; de ce fait, les nombres de macrophages et de
neutrophiles varient comme le nombre total de cellules dans les deux espaces : le nombre de
macrophages est constant entre les souris WT et S100A9-/- pour un mode d’inoculation donné
mais est plus élevé après une infection NAT (Figure 50A-B et D E). Les neutrophiles (Figure
50B et E) sont présents en faible quantité dans la cavité pleurale et le liquide broncho-
alvéolaire.
Le nombre d'éosinophiles tend à augmenter chez les souris S100A9-/- dans les deux
espaces et suivant les deux modes d'inoculation. Les résultats dans la cavité pleurale après
infection naturelle ne sont pas encore disponibles (erreur de phénotypage).
Figure 50. Quantification des macrophages et des neutrophiles pleuraux et alvéolaires.
Des souris C57BL/6 de type sauvage (WT) et déficientes en S100A9 ont été infestées avec 40
L3 de L. sigmodontis par voie sous-cutanée (SC) ou par voie naturelle (NAT). 12-13 jours
après inoculation, les cellules résidentes de la cavité pleurale et du liquide bronchoalvéolaire
(LBA) ont été récupérées et phénotypées en cytométrie en flux. (A) Nombre de macrophages
dans la cavité pleurale ; (B) Nombre de neutrophiles dans la cavité pleurale ; (C) Nombre
d'éosinophiles dans la cavité pleurale. (D) Nombre de macrophages dans le LBA ; (E)
Nombre de neutrophiles dans le LBA. (F) Nombre d'éosinophiles dans le LBA. Les résultats
sont exprimés en moyenne +/- SEM d'une expérience. Cavité pleurale - SC : n = 7-8 souris
par groupe ; NAT : n = 5 souris/groupe. LBA - SC : n = 4 souris par groupe ; NAT : n = 5
souris par groupe. 1 expérience par mode d'inoculation. Des tests de Mann Whitney ont été
effectués *p<0,05
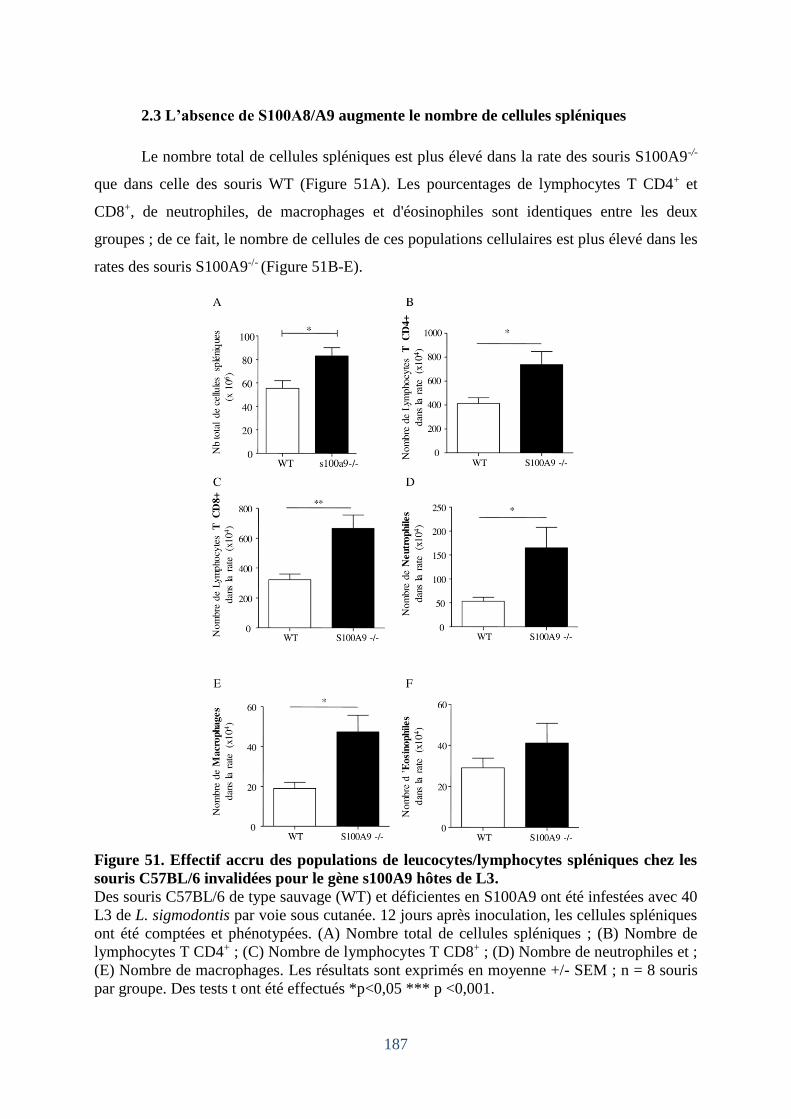
187
2.3 L’absence de S100A8/A9 augmente le nombre de cellules spléniques
Le nombre total de cellules spléniques est plus élevé dans la rate des souris S100A9-/-
que dans celle des souris WT (Figure 51A). Les pourcentages de lymphocytes T CD4+ et
CD8+, de neutrophiles, de macrophages et d'éosinophiles sont identiques entre les deux
groupes ; de ce fait, le nombre de cellules de ces populations cellulaires est plus élevé dans les
rates des souris S100A9-/- (Figure 51B-E).
Figure 51. Effectif accru des populations de leucocytes/lymphocytes spléniques chez les
souris C57BL/6 invalidées pour le gène s100A9 hôtes de L3.
Des souris C57BL/6 de type sauvage (WT) et déficientes en S100A9 ont été infestées avec 40
L3 de L. sigmodontis par voie sous cutanée. 12 jours après inoculation, les cellules spléniques
ont été comptées et phénotypées. (A) Nombre total de cellules spléniques ; (B) Nombre de
lymphocytes T CD4+ ; (C) Nombre de lymphocytes T CD8+ ; (D) Nombre de neutrophiles et ;
(E) Nombre de macrophages. Les résultats sont exprimés en moyenne +/- SEM ; n = 8 souris
par groupe. Des tests t ont été effectués *p<0,05 *** p <0,001.

188
2.4 Le profil cytokinique des cellules spléniques est caractérisé par une
dominance pro-inflammatoire en l’absence de S100A8/A9
Après restimulation pendant 72h par un mitogène (concanavaline A) ou de l’extrait brut de
filaires adultes, la capacité proliférative des splénocytes des souris S100A9-/- est supérieure à
celle des souris WT et en réponse au stimulus filarien ces splénocytes des souris déficientes
en s100a9 produisent davantage des cytokines inflammatoires IL-6, IL-13 ou KC (Figure 52).
Figure 52. Comparaison de la
concentration de cytokines dans les
surnageants de cultures de cellules
spléniques.
Des souris C57BL/6 de type sauvage (WT)
et déficientes en S100A9 ont été infestées
avec 40 L3 de L. sigmodontis par voie sous-
cutanée. 12 jours après inoculation, les
cellules de la rate ont été isolées et 2.106
cellules ont été mises en culture et
stimulées ou non pendant 72h avec de la
Concanavaline A (ConA) à 2.5 µg/mL ou
des antigènes filariens (LsAg) à 25 mg/mL.
Les cultures ont été faites en triplicat pour
chaque souris. Les surnageants de culture
ont été récupérés et des dosages de
cytokines ont été effectués par ELISA. (A)
dosage de l’IL-6 produit après restimulation
de cellules spléniques ; (B) dosage de l’IL-
13; (C) dosage de KC. n = 8 souris par
groupe. Les résultats sont exprimés en
moyenne +/- SEM. n = 8 souris par groupe.
Des tests de Mann Whitney ont été
effectués *p<0,05 ** p <0,01.

189
2.5 Observation de dommages des vaisseaux sanguins pulmonaires et de
processus inflammatoires bronchiques indépendants du S100A8/A9 chez les
souris C57BL/6
L’augmentation précoce de S100A9 dans les poumons (premier paragraphe de ce
chapitre) et l’inflammation bronchoalvéolaire observée chez les souris BALB/c infestées
(Chapitre 1), nous a conduit à réaliser une analyse histologique des poumons mais à un temps
plus tardif (J12-J13) et chez des souris de la lignée C57BL6, WT ou déficientes en S100A9.
Dans 100% des poumons des souris C57BL/6 infestées par voie naturelle et 30% des souris
infestées en SC, des atteintes pulmonaires ont été observées, aussi bien chez les souris WT
que S100A9-/- (Figures 53 et 54). La manifestation principale est une inflammation des
vaisseaux sanguins, qui apparaissent congestionnés et présentent de forts infiltrats
périvasculaires (Figure 53 et 54A-B). Les souris infestées en SC présentent moins de
vaisseaux sanguins enflammés que les souris infestées naturellement (Figures 53 et 54C) et
cette différence ne semble pas due au mode d'inoculation mais au plus grand nombre de
parasites ayant traversé les poumons. En effet, la quantité de vaisseaux sanguins
congestionnés est corrélée au nombre de parasites dans la cavité pleurale (r2=0,45, p>0,0001,
Figure 54D). Le nombre légèrement plus faible de vaisseaux sanguins congestionnés chez les
souris S100A9-/- après l'infection naturelle serait alors dû au rendement parasitaire plus faible
observé chez ces souris.
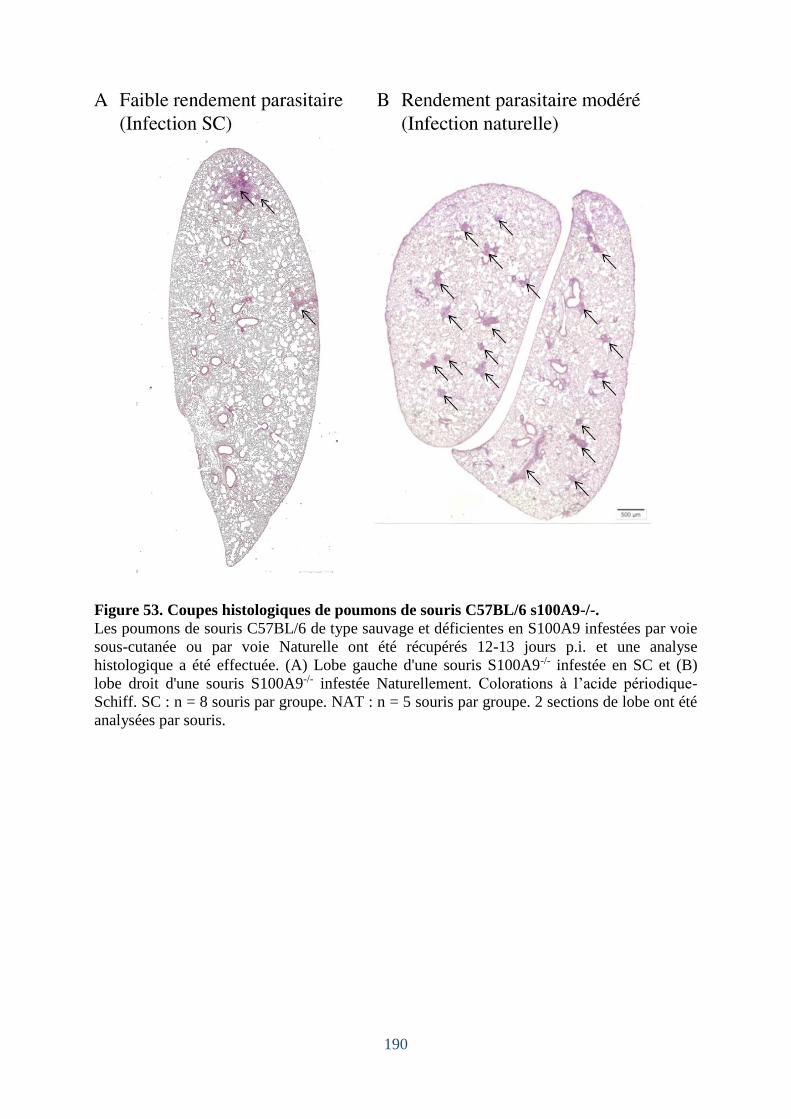
190
Figure 53. Coupes histologiques de poumons de souris C57BL/6 s100A9-/-.
Les poumons de souris C57BL/6 de type sauvage et déficientes en S100A9 infestées par voie
sous-cutanée ou par voie Naturelle ont été récupérés 12-13 jours p.i. et une analyse
histologique a été effectuée. (A) Lobe gauche d'une souris S100A9-/- infestée en SC et (B)
lobe droit d'une souris S100A9-/- infestée Naturellement. Colorations à l’acide périodique-
Schiff. SC : n = 8 souris par groupe. NAT : n = 5 souris par groupe. 2 sections de lobe ont été
analysées par souris.
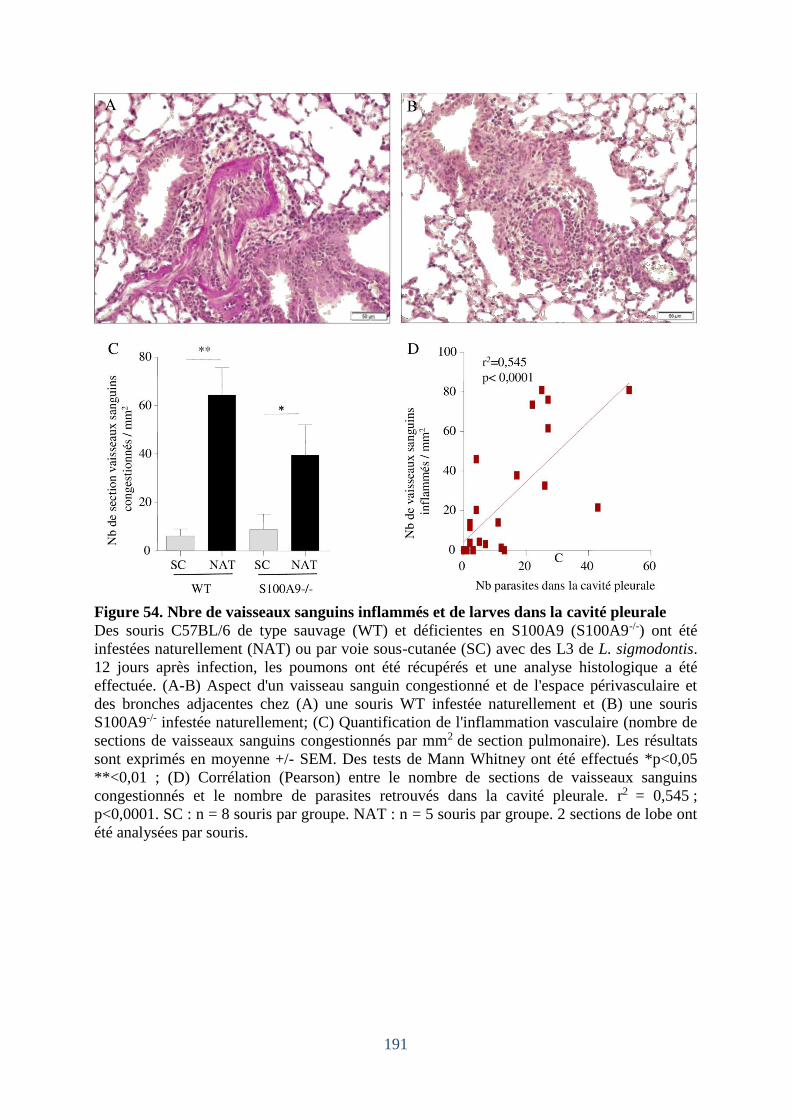
191
Figure 54. Nbre de vaisseaux sanguins inflammés et de larves dans la cavité pleurale
Des souris C57BL/6 de type sauvage (WT) et déficientes en S100A9 (S100A9-/-) ont été
infestées naturellement (NAT) ou par voie sous-cutanée (SC) avec des L3 de L. sigmodontis.
12 jours après infection, les poumons ont été récupérés et une analyse histologique a été
effectuée. (A-B) Aspect d'un vaisseau sanguin congestionné et de l'espace périvasculaire et
des bronches adjacentes chez (A) une souris WT infestée naturellement et (B) une souris
S100A9-/- infestée naturellement; (C) Quantification de l'inflammation vasculaire (nombre de
sections de vaisseaux sanguins congestionnés par mm2 de section pulmonaire). Les résultats
sont exprimés en moyenne +/- SEM. Des tests de Mann Whitney ont été effectués *p<0,05
**<0,01 ; (D) Corrélation (Pearson) entre le nombre de sections de vaisseaux sanguins
congestionnés et le nombre de parasites retrouvés dans la cavité pleurale. r2 = 0,545 ;
p<0,0001. SC : n = 8 souris par groupe. NAT : n = 5 souris par groupe. 2 sections de lobe ont
été analysées par souris.
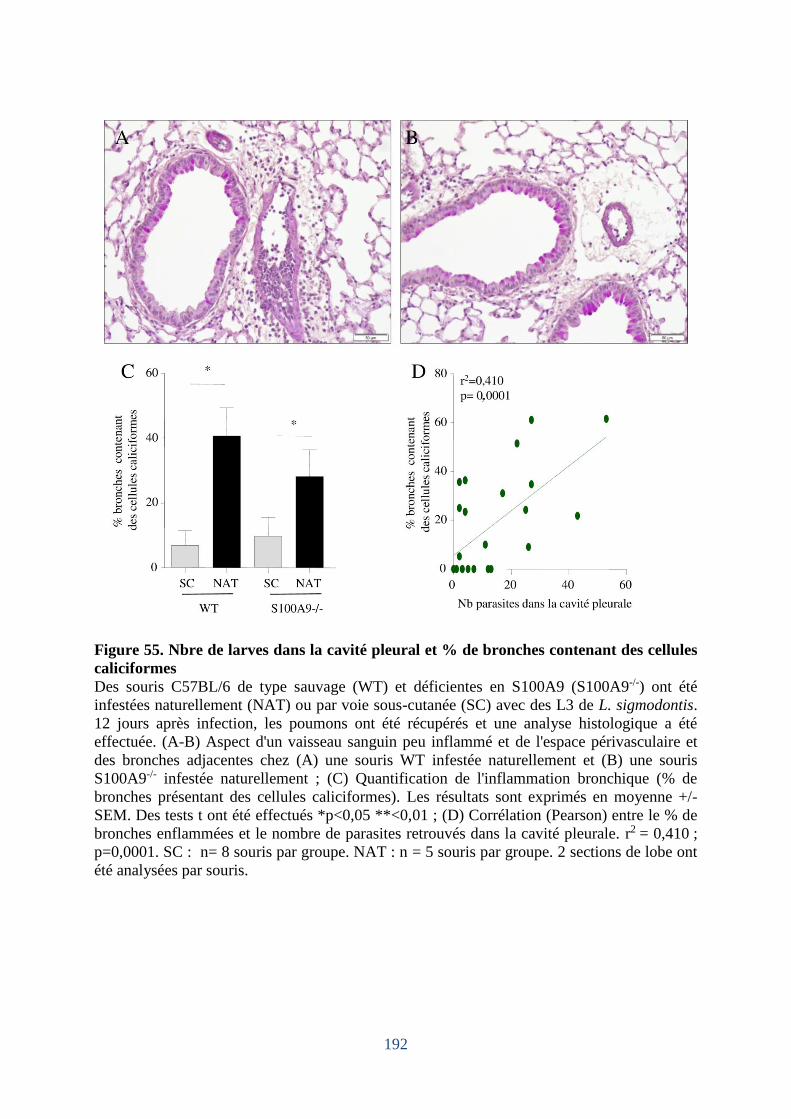
192
Figure 55. Nbre de larves dans la cavité pleural et % de bronches contenant des cellules
caliciformes
Des souris C57BL/6 de type sauvage (WT) et déficientes en S100A9 (S100A9-/-) ont été
infestées naturellement (NAT) ou par voie sous-cutanée (SC) avec des L3 de L. sigmodontis.
12 jours après infection, les poumons ont été récupérés et une analyse histologique a été
effectuée. (A-B) Aspect d'un vaisseau sanguin peu inflammé et de l'espace périvasculaire et
des bronches adjacentes chez (A) une souris WT infestée naturellement et (B) une souris
S100A9-/- infestée naturellement ; (C) Quantification de l'inflammation bronchique (% de
bronches présentant des cellules caliciformes). Les résultats sont exprimés en moyenne +/-
SEM. Des tests t ont été effectués *p<0,05 **<0,01 ; (D) Corrélation (Pearson) entre le % de
bronches enflammées et le nombre de parasites retrouvés dans la cavité pleurale. r2 = 0,410 ;
p=0,0001. SC : n= 8 souris par groupe. NAT : n = 5 souris par groupe. 2 sections de lobe ont
été analysées par souris.

193
La différenciation des cellules épithéliales bronchiques en cellules caliciformes (ou
métaplasie) et leur production de mucus est une des caractéristiques de l'inflammation
pulmonaire. Au niveau des vaisseaux sanguins présentant une inflammation importante,
aucune cellule productrice de mucus n'est observée dans l'épithélium bronchique (Figure 54A
et B). Une métaplasie des cellules caliciformes est néanmoins observée dans les bronches à
proximité d'espaces périvasculaires présentant de faibles infiltrats cellulaires (Figure 55A-B).
Comme pour l'inflammation des vaisseaux sanguins, l'inflammation bronchique est plus
élevée chez les souris infestées naturellement (Figure 55C) et est corrélée au nombre de
parasites dans la cavité pleurale (r2 = 0,410 ; p = 0,0001 ; Figure 55D).
2.6 En l’absence de S100A8/A9, quelles fonctions pour les neutrophiles ?
Les résultats précédents ne sont pas en faveur d’une destruction des larves après leur
arrivée dans la cavité pleurale car aucune L3 ou L4 morte, enkystée ou non, n’a été retrouvée
chez les souris S100A9-/-. Il est donc probable que la baisse du rendement parasitaire
s’établisse en amont de la cavité pleurale, soit au site d’inoculation dans la peau, soit pendant
la phase de migration des L3 y compris dans les poumons.
Dans les deux cas les neutrophiles pourraient être impliqués du fait de la présence
intracellulaire importante de S100A9.
Une expérience préliminaire a été effectuée, dans laquelle des neutrophiles matures
isolés de la moelle osseuse de souris S100A9-/- et WT ont été cultivés avec différents stimuli
pendant 6h, puis des marqueurs d'activations cellulaires ont été mesurés par cytométrie en
flux (Figure 56). Le LPS (ligand de TLR-4), le Pam3Cys (ligand de TLR-2), de l’extrait brut
de filaires adultes (Ls Ag) ou de l’extrait brut de filaires adultes traitées avec de la tétracycline
(Ls Tet), et donc déplétées en Wolbachia, ont été utilisés pour stimuler les cultures.
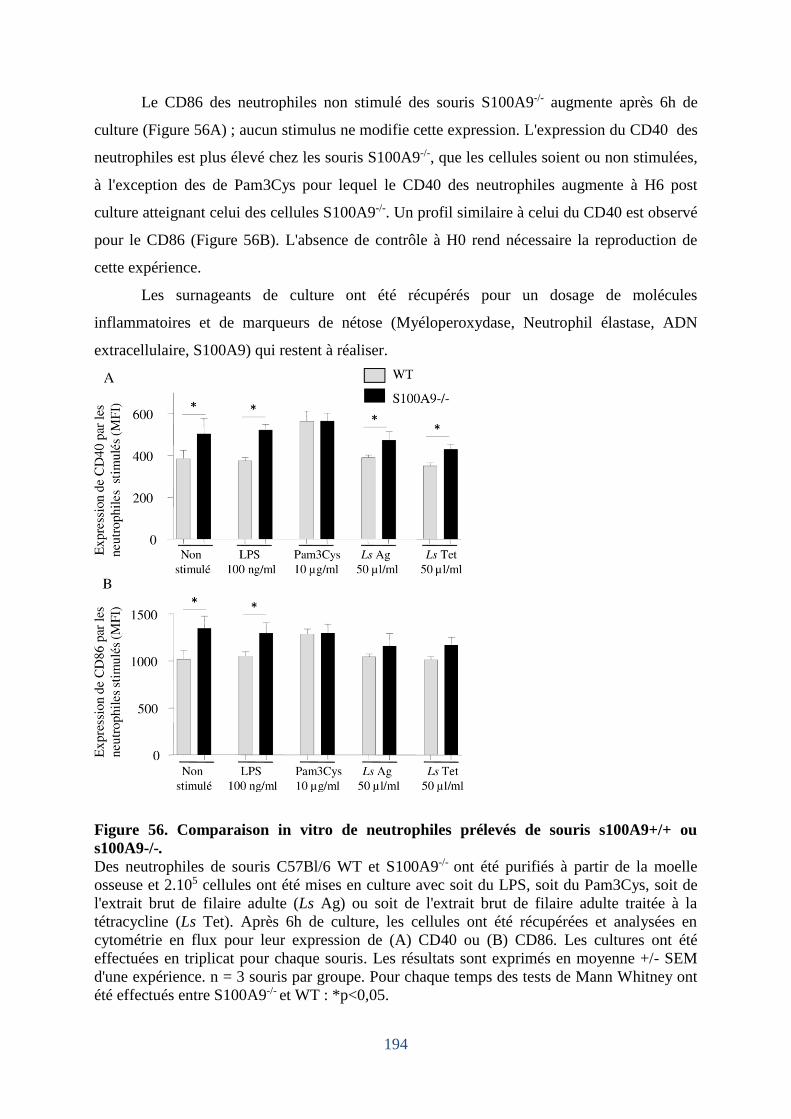
194
Le CD86 des neutrophiles non stimulé des souris S100A9-/- augmente après 6h de
culture (Figure 56A) ; aucun stimulus ne modifie cette expression. L'expression du CD40 des
neutrophiles est plus élevé chez les souris S100A9-/-, que les cellules soient ou non stimulées,
à l'exception des de Pam3Cys pour lequel le CD40 des neutrophiles augmente à H6 post
culture atteignant celui des cellules S100A9-/-. Un profil similaire à celui du CD40 est observé
pour le CD86 (Figure 56B). L'absence de contrôle à H0 rend nécessaire la reproduction de
cette expérience.
Les surnageants de culture ont été récupérés pour un dosage de molécules
inflammatoires et de marqueurs de nétose (Myéloperoxydase, Neutrophil élastase, ADN
extracellulaire, S100A9) qui restent à réaliser.
Figure 56. Comparaison in vitro de neutrophiles prélevés de souris s100A9+/+ ou
s100A9-/-.
Des neutrophiles de souris C57Bl/6 WT et S100A9-/- ont été purifiés à partir de la moelle
osseuse et 2.105 cellules ont été mises en culture avec soit du LPS, soit du Pam3Cys, soit de
l'extrait brut de filaire adulte (Ls Ag) ou soit de l'extrait brut de filaire adulte traitée à la
tétracycline (Ls Tet). Après 6h de culture, les cellules ont été récupérées et analysées en
cytométrie en flux pour leur expression de (A) CD40 ou (B) CD86. Les cultures ont été
effectuées en triplicat pour chaque souris. Les résultats sont exprimés en moyenne +/- SEM
d'une expérience. n = 3 souris par groupe. Pour chaque temps des tests de Mann Whitney ont
été effectués entre S100A9-/- et WT : *p<0,05.

195

196
Discussion
Dès les premières heures suivant l'infection par L. sigmodontis l’expression des
transcrits de s100a8 et s100a9 augmente dans les poumons des souris BALB/c de type
sauvage. Ceci est accompagné d’une augmentation transitoire du nombre de neutrophiles
pulmonaires exprimant S100A9 et des niveaux de la protéine dans le liquide
bronchoalvéolaire et le liquide pleural ; ces modifications sont observées 2-6 heures après
infection et elles sont résolues entre J4 et J8. Le même profil de réponse est observé chez les
souris infestées par voies sous-cutanée et intraveineuse, suggérant un effet probablement
distant et indépendant du nombre de parasites : en effet à H2-H6 p.i., très peu de parasites
sont présents dans les poumons des souris infestées en SC et aucun parasite n'a atteint la
cavité pleurale contrairement aux souris infestées en IV (Chapitre 1). Il est donc plus probable
que la libération de S100A9 dans l'environnement pulmonaire soit avant tout une réponse des
neutrophiles à des antigènes de filaires ou de Wolbachia drainés par le flux lymphatique et le
sang jusqu'aux poumons avant l'arrivée des premières filaires. De plus, S100A8 et S100A9
sont importants dans la réponse aux dommages vasculaires [452] et pourraient participer à la
réparation des capillaires pulmonaires après le passage des L3.
Pour étudier le rôle du S100A9 lors l’infection par L. sigmodontis, des souris C57BL/6
déficientes en cette molécule (S100A9-/-) ont été utilisées. Les souris déficientes en S100A9
sont viables et fertiles [355] contrairement aux souris déficientes en S100A8 [416], le partenaire
d'hétérodimérisation de S100A9. Malgré la présence des transcrits S100A8 dans divers
organes chez les souris S100A9-/-, une forte régulation négative de la protéine S100A8 a été
observée chez ces souris ; en effet l’absence de S100A9 entraîne une réduction de 30% des
cellules S100A8+ dans la moelle osseuse et cette valeur diminue drastiquement pendant la
culture de ces cellules ; de plus la protéine S100A8 n’est pas détectable dans les cellules
S100A9-/- du sang périphérique [355]. Cependant l’absence de S100A8 et de S100A9, qui
représentent près de 40% des protéines cytosoliques des neutrophiles normaux, n'affecte pas
le développement de neutrophiles viables et matures [355].
L’hétérodimère S100A8/A9 qui effectue une grande partie des fonctions biologique de
ces molécules - mais pas seulement - est donc aussi absent dans les souris S100A9-/-.
L'inoculation des filaires dans la peau de souris S100A9-/-, que ce soit après injection de L3
par voie sous-cutanée ou par la piqure du vecteur, se traduit par une baisse du succès
parasitaire dans les deux protocoles : les rendements filariens sont diminués 12 jours après

197
infection par rapport à des souris de type sauvage, suggérant que la présence de S100A9
augmenterait la survie des L3. Le recrutement de cellules dans la cavité pleurale des souris
S100A9-/- infestées est plus importants que chez les souris de type sauvage, mais cette
différence est essentiellement due aux macrophages et dans une moindre mesure aux
éosinophiles ; le recrutement des neutrophiles est semblable entre souris s100A9-/- et souris
WT. Ce résultat pourrait paraitre surprenant compte tenu que le S100A9 est avant tout connu
pour ses propriétés inflammatoires et son rôle favorisant le recrutement des neutrophiles
[418,419,453]). En effet, au niveau cellulaire, les neutrophiles S100A9-/- présentent un système de
microfilaments plus organisé et une réduction de la régulation de CD11b médiée par l'IL-8, ce
qui entraîne un potentiel réduit pour répondre aux molécules chimioattractantes in vitro [355] et
nous aurions pu nous attendre à une diminution du recrutement des neutrophiles dans la cavité
pleurale des souris S100A9-/- infestées. Néanmoins, aucun défaut de migration des
neutrophiles n’a été observé dans deux modèles inflammatoires in vivo (injection d’IL-8 ou de
thioglycolate) [355], ce qui suggère que les molécules S100A9 et S100A8 ne sont pas
indispensables au recrutement des neutrophiles dans ces modèles inflammatoires mais aussi
dans l’infection filarienne.
Un effet immunosuppresseur d’une pré-stimulation par le S100A9 sur les cellules
phagocytaires a été décrit dans un modèle d’inflammation au LPS et est dépendant du TLR4
[439-443]). La stimulation des macrophages pleuraux par le S100A9 en l'absence des parasites
dans les premières heures suivant l'infection pourrait être responsable de la mise en place d’un
phénotype tolérogène des macrophages limitant l’inflammation pleurale. La stimulation des
cellules B1 par S100A9 et leur production d'IL-10 pourrait aussi participer au phénomène,
comme cela a été décrit dans la cavité péritonéale [444].
Chez les souris C57BL/6, le rendement parasitaire est stable pendant au moins 20-30
jours [195,196]. A 12 jours post infection, l’augmentation cellulaire observée dans la cavité
pleurale des souris S100A9-/- est encore très inférieure à celle observée chez des souris
sauvages à J20 ou J30, temps auxquels des cellules adhérant aux filaires sont identifiées ainsi
que des granulomes. Il est donc peu probable que la différence de rendement parasitaire
provienne d’une réaction cellulaire filaricide dans la cavité pleurale. L’action de S100A9 sur
les filaires a probablement lieu plus tôt au cours de l’infection, lors de la phase de migration
des L3 de leur site d’inoculation cutané vers la cavité pleurale, donc dans la peau ou dans les
poumons.

198
L'analyse histologique des poumons des souris C57BL/6 a permis d'observer une
inflammation pulmonaire corrélée au nombre de parasites retrouvés dans la cavité pleurale
mais indépendante du S100A9. Cette réponse inflammatoire pulmonaire n'a pas été observée
jusqu'à présent chez les souris BALB/c, soit parce que l'événement est rare après l'inoculation
de 40 L3 et est passé inaperçu, soit parceque elle est due à la différence de susceptibilité entre
les souris C57BL/6 et BALB/c. Ces atteintes pulmonaires semblent être la conséquence du
passage des L3 dans la circulation sanguine pulmonaire qui entraînerait une réaction de
l'endothélium vasculaire. Une inflammation périvasculaire similaire à celle décrite avec L.
sigmodontis a été observée 4 jours après infection chez des souris infestées par
Nippostrongylus brasiliensis et elle était composée de macrophages et d’éosinophiles [320,339].
L’analyse du contenu cellulaire de l'espace périvasculaire et de la lumière des vaisseaux
sanguins congestionnés permettra de mieux caractériser cette inflammation pulmonaire. Des
mécanismes identiques pourraient être à l'origine de la pathologie des vaisseaux sanguins
pulmonaires ayant lieu chez les chiens infestés par Dirofilaria immitis. En effet,
l’inflammation des artères pulmonaires due à la présence de D. immitis dans la circulation
sanguine pulmonaire induit une obstruction progressive de la lumière vasculaire [134].
La réponse cutanée des souris S100A9-/- infestées avec L. sigmodontis n'a pas encore
été étudiée. L’hétérodimère S100A8/S100A9 a des propriétés antioxydantes [430] qui
pourraient diminuer le choc oxydatif produit par les neutrophiles et donc l’action filaricide des
espèces réactives oxygénées (ROS) sur les L3. L’infection avec des L3 de B. malayi diminue
l’action des cellules dendritiques de la peau [265],[266], or il a été décrit que la stimulation des
cellules dendritiques par du S100A9 peut aussi bloquer leur activation et la présentation
antigénique aux lymphocytes [440,441]. CD40 est une protéine costimulatrice trouvée sur les
cellules présentatrices d'antigène et qui est nécessaire pour leur activation. Les neutrophiles
S100A9-/- présentent des niveaux accrus de CD40, mais aussi de CD86. Une augmentation de
l'expression de ces molécules sur les neutrophiles a aussi été associée à leur apoptose [454] et le
S100A9 induit un effet anti-apoptotique sur les neutrophiles - normaux et allergiques - en
augmentant la sécrétion des cytokines MCP-1, IL-6 et IL-8 par les monocytes [429,455]. Pendant
la nétose S100A9 fait partie des molécules libérées par les neutrophiles [398,420,421] et pourrait
contribuer à contrôler la charge parasitaire dans la peau (cf. Chapitre 1). Enfin,
S100A8/S100A9 induit une perméabilité accrue de la monocouche endothéliale [427,428]. Dans
la peau, l'augmentation de la perméabilité vasculaire pourrait entraîner une augmentation du

199
drainage lymphatique et donc une entrée facilitée des parasites dans le système lymphatique
comme cela a été observé pour l’histamine [264].
S100A9 agit comme un DAMP (damage-associated molecular pattern) via TLR4 ou
RAGE et déclenche des réponses inflammatoires anormales [450,456,457]. La réponse
immunitaire en filariose est complexe, en particulier l'implication et le rôle fonctionnel joué
par TLR4. TLR4 et TLR2 sont capables de lier des produits de la bactérie endosymbiotique
Wolbachia comme WSP, la protéine de surface de Wolbachia, et d'induire une réponse
inflammatoire en sécrétant des cytokines pro-inflammatoires à partir de macrophages et de
cellules dendritiques [270]. Cependant, les filaires visent également TLR4 pour moduler la
réponse immunitaire de l'hôte [289]. L’ES-62, une glycoprotéine conjuguée à la
phosphorylcholine, lie le TLR4 et induit une réponse immunitaire anti-inflammatoire/Th2
favorable à la survie du parasite dans l'hôte [458]. En cas d'infection chronique, la filaire induit
l’apoptose des lymphocytes T pour réguler la réponse de l'hôte et ce processus fonctionne
aussi par TLR4 [77]. Les microfilaires de B. malayi modulent l'expression de TLR4 dans des
cellules dendritiques humaines dérivées de monocytes pour induire un environnement
immunosuppresseur dans l'hôte qui bloque la production de cytokines proinflammatoires et
d'interférons de type 1 [459]. En plus de cet effet inhibiteur des microfilaires sur les cellules
dendritiques, une inhibition partielle de la fonction des cellules de Langerhans humaines a été
observée [459]. Enfin TLR4 est impliqué dans l'activation du système immunitaire conduisant à
l'inhibition de l'embryogenèse filaire ou à la destruction directe des microfilaires. TLR4 a
ainsi un rôle sur la phase patente car presque 100% des souris BALB/c TLR4-/- ont une
microfilarémie sanguine [288].
Les mécanismes inhibant l’embryogénèse de L. sigmodontis via le TLR4 ne sont pas connus.
Les neutrophiles pourraient être impliqués et en particulier leurs protéines S100A8 et
S100A9.

200

201

202
CHAPITRE 3 :
Dommages pulmonaires prolongés chez les rongeurs chez lesquels est générée la descendance de Litomosoides
sigmodontis

203
Introduction :
Peu après leur arrivée dans la cavité pleurale, les L3 muent en L4 puis, environ 30
jours après infection, en adultes. Les adultes mâles et femelles se reproduisent alors et à partir
de 55 jours, les femelles pondent les microfilaires qui vont rejoindre la circulation sanguine.
Le statut microfilarien est variable en fonction du rongeur hôte : la gerbille et le rat du coton
ont d’importantes microfilarémies sanguines (>100 Mf/10µL de sang, atteignant jusqu’à 3000
Mfs/10µL de sang) et plus de 90% des individus sont microfilariens. Chez la souris le statut
microfilarien dépend de la lignée [194] et seules les souris BALB/c peuvent être
microfilariennes ; cependant, seulement 40-60% des souris présentent une microfilarémie
après une inoculation en sous-cutanée de L3 ou via un mode d’infestation naturelle par les
vecteurs ; de plus les microfilarémies des souris BALB/c sont toujours faibles (1-20Mfs/10µl
de sang).
De nombreuses études ont comparé les statuts microfilarien et amicrofilarien de
différentes souches de souris [196,286,460-462] et ont mis en évidence l'importance de la réponse
immunitaire dans l'établissement la phase patente. Par exemple, des souches de souris
C57BL/6 immunodéficientes peuvent devenir microfilariennes [286,462]. Toutefois, peu d'études
ont comparé les réponses pleurales/pulmonaires des souris microfilarémiques (Mf+) et
amicrofilarémiques (Mf-) au sein des souris BALB/c. Il a récemment été montré que les
souris Mfpos présentent des concentrations plus élevées d'IL-5 et d'IL-13 dans la cavité
pleurale que les Mfneg [288].
Des études sur la distribution des microfilaires dans des modèles animaux de filariose
ont conclu que les microfilaires non-circulantes sont situées dans les vaisseaux sanguins des
organes profonds [187,460,463]. Chez les humains, des microfilaires ont été détectées dans la rate,
le foie, la moelle osseuse, les reins, la cavité pleurale, les bronches et les capillaires
pulmonaires [78,81,181,182,464-466]. De même, les microfilaires de L. sigmodontis peuvent
s'accumuler dans ces organes chez les rongeurs, et particulièrement dans les poumons
[196,463,467].

204
Les travaux de Wharton (1946 [207]) et Schneider (1968 [208]) ont décrit des lésions
pulmonaires chez le rat du coton (Sigmodon hispidus) et la gerbille (Meriones unguiculatus)
en phase patente d’infestation par L. sigmodontis. Ils ont alors observé une hyperplasie et une
prolifération des cellules mésothéliales avec la formation de "nodules papillaires" et une
fibrose de la plèvre. Des microfilaires ont été observées dans la vasculature pulmonaire, mais
aussi dans le parenchyme, et des infiltrats cellulaires étaient présents autour des bronches
[207,208]. Les atteintes pulmonaires lors de la phase patente n'ont pas été étudiées chez les souris
BALB/c.
L'objectif de ce chapitre est d’étudier la phase patente de l’infection filarienne dans
l’environnement pleural/pulmonaire chez la souris BALB/c, mais en discriminant les
informations relatives aux souris microfilarémiques de celles liées aux souris
amicrofilarémiques.
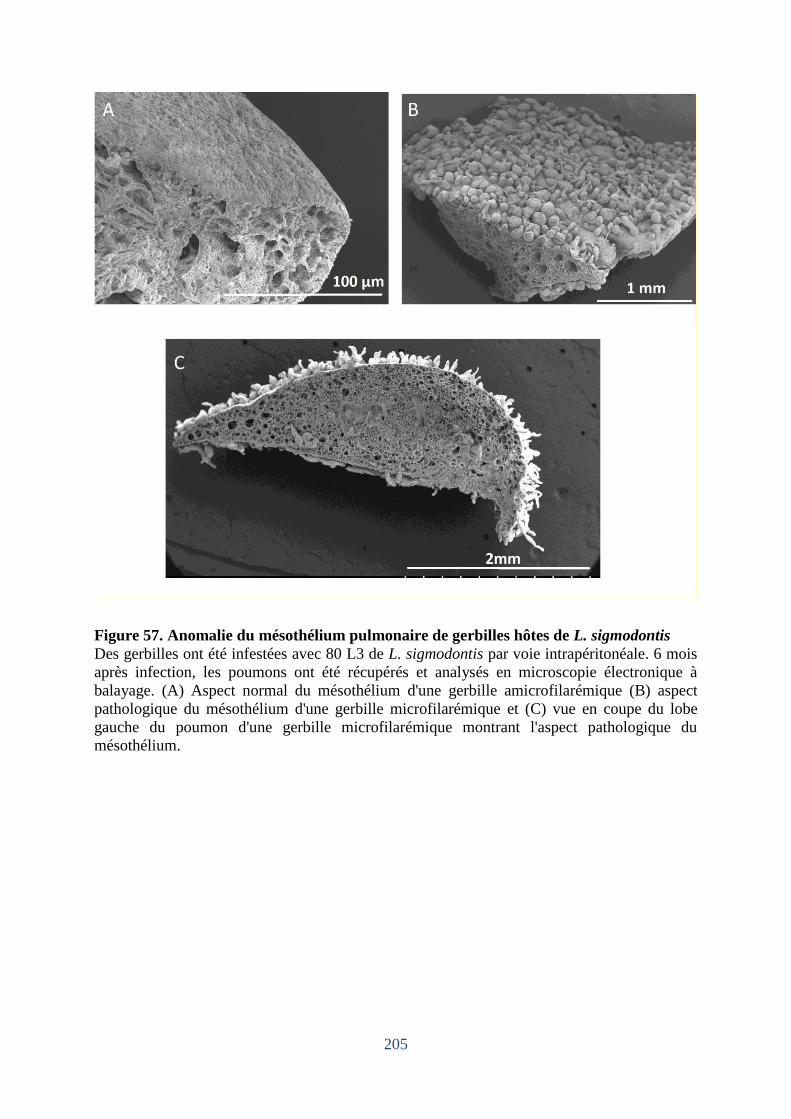
205
Figure 57. Anomalie du mésothélium pulmonaire de gerbilles hôtes de L. sigmodontis
Des gerbilles ont été infestées avec 80 L3 de L. sigmodontis par voie intrapéritonéale. 6 mois
après infection, les poumons ont été récupérés et analysés en microscopie électronique à
balayage. (A) Aspect normal du mésothélium d'une gerbille amicrofilarémique (B) aspect
pathologique du mésothélium d'une gerbille microfilarémique et (C) vue en coupe du lobe
gauche du poumon d'une gerbille microfilarémique montrant l'aspect pathologique du
mésothélium.

206
Résultats
1. Microfilarémie et dommages pulmonaires chez le mérion
Lors d'autopsies de gerbilles M. unguiculatus utilisées pour maintenir le cycle de vie
de Litomosoides sigmodontis au laboratoire, il a souvent été remarqué que les poumons des
animaux présentant une très forte microfilarémie ont un aspect granuleux à l'œil nu. Pour
caractériser ce phénomène, les poumons d'une gerbille infestée depuis 6 mois et
amicrofilarémique et ceux d'une gerbille présentant des microfilaires sanguines (environ 1000
mf/10µL) ont été observés en microscopie électronique à balayage (Figure 57). Comme cela a
été décrit précédemment [207,208], de nombreuses excroissances sont présentes sur la plèvre
viscérale des poumons de la gerbille Mfpos (que les auteurs avaient alors appelé "nodules
papillaires").
L'observation en coupe du tissu permet d'observer la structure interne de ces
excroissances (Figure 58A). Elles sont constituées de fibres (Figure 58B, flèches), ce qui
suggère un phénomène de fibrose. Sous ces excroissances mais parfois à très grande
proximité de la plèvre viscérale des microfilaires sont observées dans les alvéoles
pulmonaires (Figure 59 flèches).
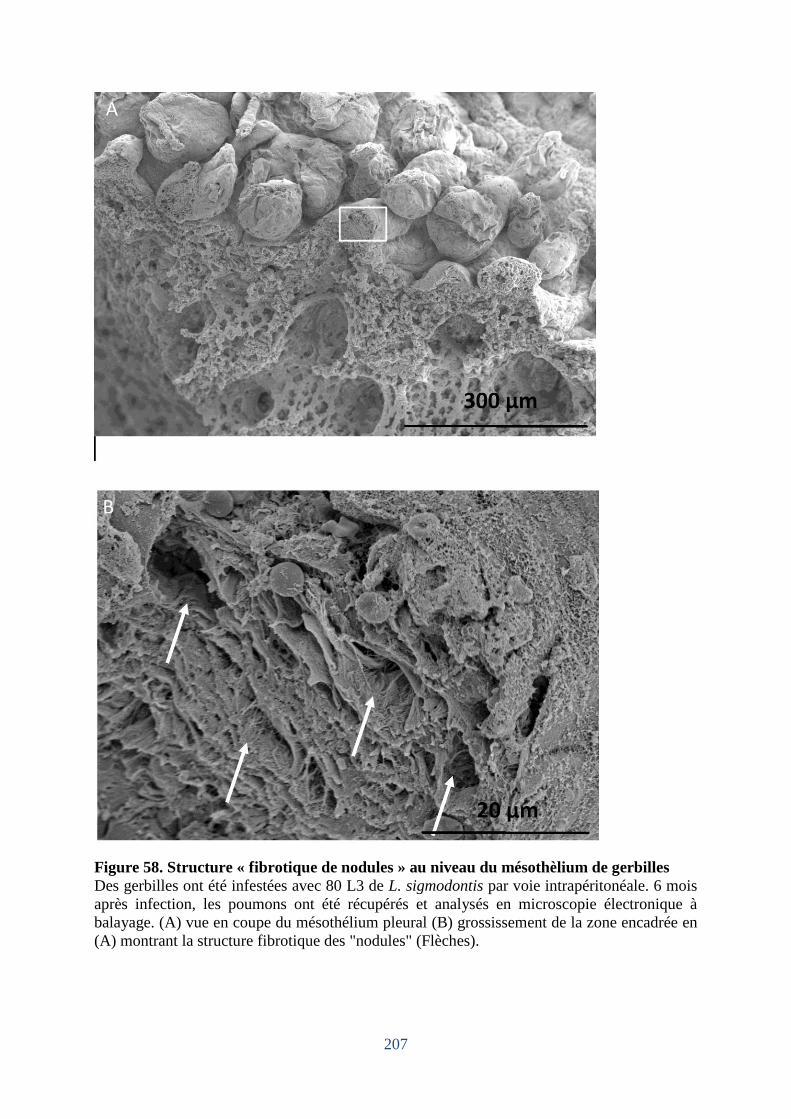
207
Figure 58. Structure « fibrotique de nodules » au niveau du mésothèlium de gerbilles
Des gerbilles ont été infestées avec 80 L3 de L. sigmodontis par voie intrapéritonéale. 6 mois
après infection, les poumons ont été récupérés et analysés en microscopie électronique à
balayage. (A) vue en coupe du mésothélium pleural (B) grossissement de la zone encadrée en
(A) montrant la structure fibrotique des "nodules" (Flèches).

208
Figure 59. Microfilaires à proximité de la plèvre viscérale de gerbilles hôtes de L.
sigmodontis.
Des gerbilles ont été infestées avec 80 L3 de L. sigmodontis par voie intrapéritonéale. 6 mois
après infection, les poumons ont été récupérés et analysés en microscopie électronique à
balayage. Coupe du tissu pulmonaire montrant des microfilaires dans les alvéoles à proximité
de la plèvre viscérale pathologique (Flèches).
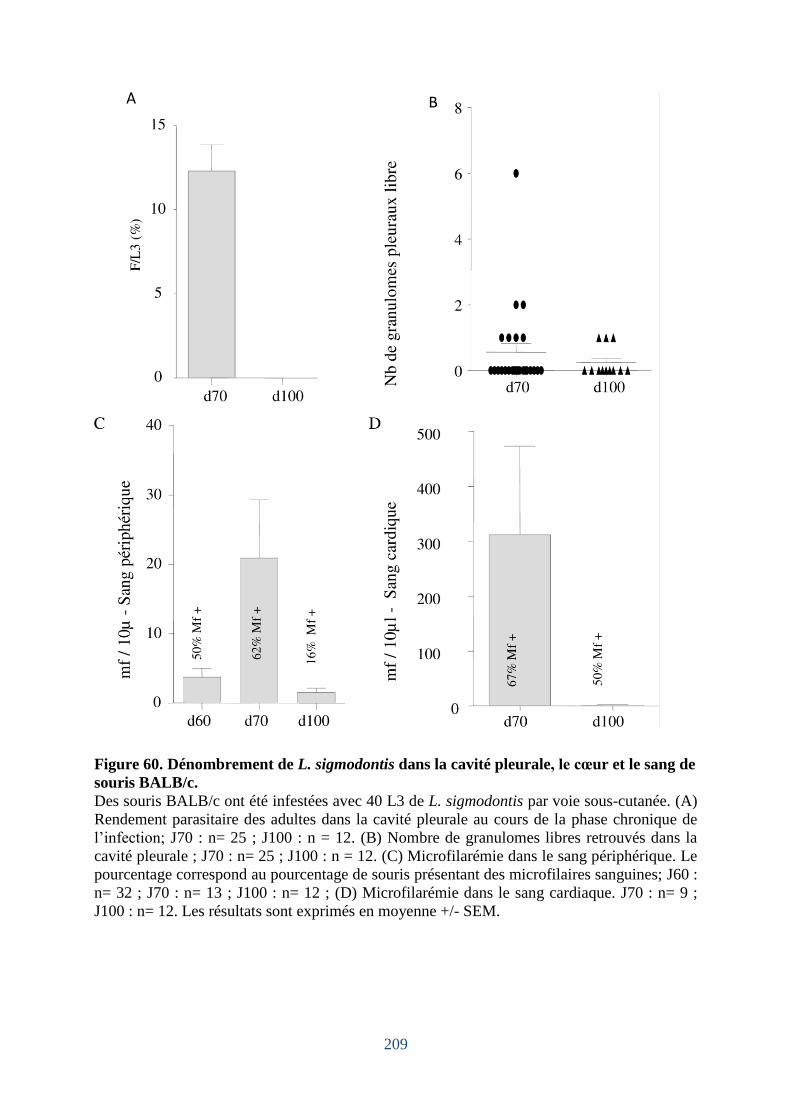
209
Figure 60. Dénombrement de L. sigmodontis dans la cavité pleurale, le cœur et le sang de
souris BALB/c.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. (A)
Rendement parasitaire des adultes dans la cavité pleurale au cours de la phase chronique de
l’infection; J70 : n= 25 ; J100 : n = 12. (B) Nombre de granulomes libres retrouvés dans la
cavité pleurale ; J70 : n= 25 ; J100 : n = 12. (C) Microfilarémie dans le sang périphérique. Le
pourcentage correspond au pourcentage de souris présentant des microfilaires sanguines; J60 :
n= 32 ; J70 : n= 13 ; J100 : n= 12 ; (D) Microfilarémie dans le sang cardiaque. J70 : n= 9 ;
J100 : n= 12. Les résultats sont exprimés en moyenne +/- SEM.

210
2. Analyse cinétique de la phase patente chez la souris BALB/c, localisation des parasites
et distinction des souris en deux groupes selon la présence ou non de microfilaires
Pour étudier les interactions entre L. sigmodontis et l'environnement pulmonaire
durant la phase patente, des souris BALB/c ont été infestées par voie sous-cutanée. A 60
jours, 70 jours et 100 jours post-infection, la microfilarémie a été mesurée. A J70 et J100 p.i.,
les parasites présents dans la cavité pleurale ont été comptés. La microfilarémie cardiaque
(dans le sang ventriculaire) a aussi été déterminée (Figure 60). Contrairement à la gerbille,
chez qui l'infection peut durer plus d'un an [201], les parasites sont éliminés vers 100 jours post-
infection chez la souris BALB/c (Figure 60A). Des granulomes sont régulièrement retrouvés
dans la cavité pleurale des souris infestées à J70 p.i., comme cela a déjà été décrit [202]. Ils
contiennent les parasites adultes morts en voie de dégradation [202]. A J100, très peu de
granulome sont retrouvés (Figure 60), suggérant une élimination relativement rapide (moins
de 30 jours) de ceux-ci. Le nombre de microfilaires augmente dans le sang périphérique entre
J60 et J70 p.i. et diminue à J100 (Figure 60C). A J60 p.i., des microfilaires sont détectées dans
le sang périphérique de la moitié des souris, et dans plus de 60% des souris à J70 p.i. Entre
J70 et J100 p.i., le pourcentage de souris dans lesquelles sont observées des microfilaires dans
le sang périphérique tombe à 16%. Etant donné les faibles microfilarémies observées dans le
sang périphérique, le nombre de microfilaires a aussi été mesuré dans le sang cardiaque lors
des autopsies pour exclure les faux négatifs. En effet, lors de l'exsanguination des animaux, le
sang restant se concentre dans les organes, dont le cœur. La microfilarémie y est donc
beaucoup plus élevée (Figure 60D). Cette technique permet de détecter des individus
microfilariens, non identifiés dans le sang périphérique. Le nombre de souris
microfilarémiques y est donc plus élevé à J70 mais surtout à J100 p.i. Par la suite, le statut
microfilarien est donc déterminé dans le sang cardiaque.
L’analyse macroscopique de la cavité pleurale et du bloc cardio-pulmonaire révèle la
présence de nombreuses filaires dégradées prises dans des granulomes attachés au médiastin.
La présence de parasite a donc été étudiée sur des coupes histologiques du tissu pulmonaire.
Des granulomes partiellement intégrés à la surface des poumons sont observés chez 30% des
souris (Figure 61A). Ces granulomes, comme les granulomes de la cavité, contiennent des
parasites adultes morts (dégradation plus ou moins avancée des tissus filariens, y compris de
la cuticule (Figure 61B)). Quelques microfilaires sont également observées dans les poumons
de 80% des souris Mf+, dans les vaisseaux sanguins (Figure 61C) et dans les espaces
périvasculaires (vaisseaux sanguins : 2,0±0,7 mfs/cm2 de section; PVS : 1,1±0,7 mfs/ cm2 de
section).
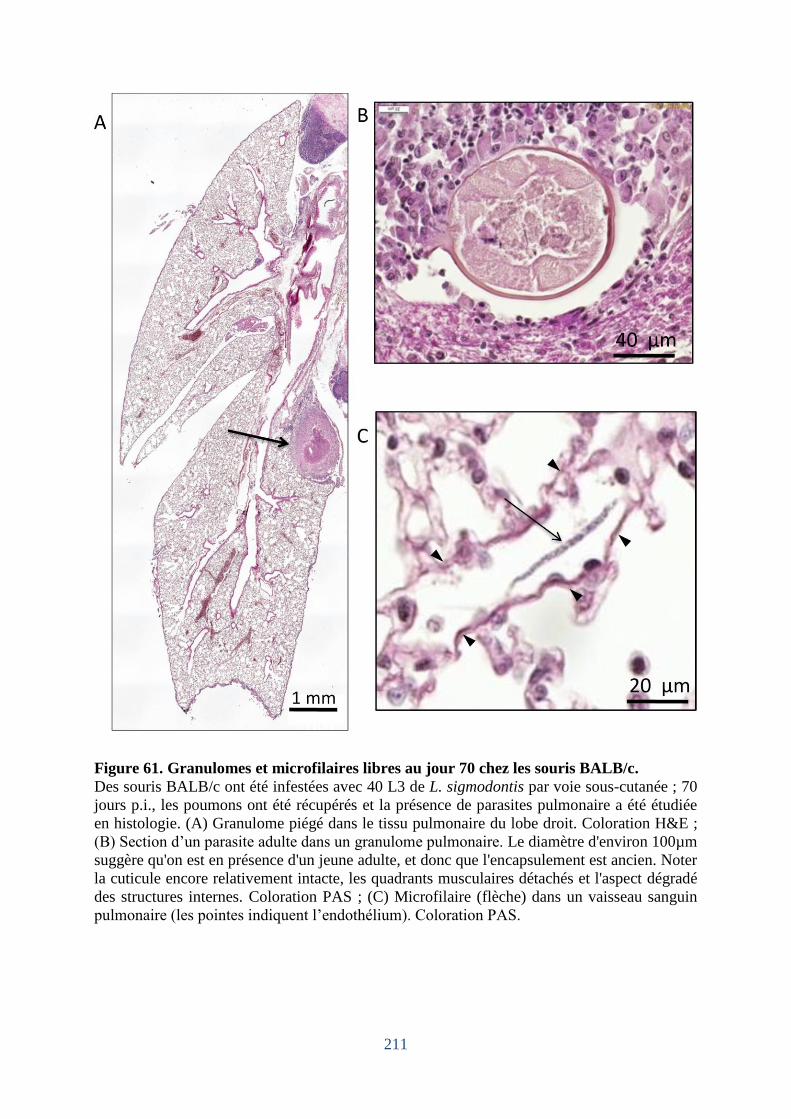
211
Figure 61. Granulomes et microfilaires libres au jour 70 chez les souris BALB/c.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée ; 70
jours p.i., les poumons ont été récupérés et la présence de parasites pulmonaire a été étudiée
en histologie. (A) Granulome piégé dans le tissu pulmonaire du lobe droit. Coloration H&E ;
(B) Section d’un parasite adulte dans un granulome pulmonaire. Le diamètre d'environ 100µm
suggère qu'on est en présence d'un jeune adulte, et donc que l'encapsulement est ancien. Noter
la cuticule encore relativement intacte, les quadrants musculaires détachés et l'aspect dégradé
des structures internes. Coloration PAS ; (C) Microfilaire (flèche) dans un vaisseau sanguin
pulmonaire (les pointes indiquent l’endothélium). Coloration PAS.

212
3. Analyse de profils de biomolécules-transcrits et protéines- au niveau des poumons
Pour caractériser l'environnement inflammatoire pulmonaire dû à la présence des
parasites - adultes et/ou microfilaires - , une analyse des transcrits cytokiniques a été réalisée
sur un pool d'ADNc provenant des poumons de souris infestées à J70 p.i., indépendamment
de leur statut microfilarien (Figure 62A). Sur les 84 gènes compris dans l'analyse (Annexe 1),
30 gènes codant pour des cytokines voient leur expression augmenter lors de l'infection
filarienne (>100% d'augmentation) par rapport à des souris non infestées et 7 gènes sont
régulés à la baisse. L'expression de gènes impliqués dans les réponses de type Th2 sont
augmentés comme l'IL-13, l'IL-4, les eotaxines CCL11 et CCL5, ou l'IL-10 mais aussi de
nombreux gènes inflammatoires associés aux réponses de type Th1 (CXCL9, CXCL10, IFN-
γ, IL-6...). Pour valider ces résultats, des PCR quantitatives pour 3 gènes candidats fortement
régulés (CXCL9, CCL2 et IL13) ont été effectuées sur les ADNc de chaque individu utilisé
dans le pool (Figure 62B).
Ensuite, les protéines candidates ont été mesurées dans le sérum, le liquide pleural et
le liquide de lavage bronchoalvéolaire de ces souris en discriminant les souris Mfpos et Mf-.
CXCL9 et CCL2 sont augmentés dans les 3 fluides des souris Mfpos (Figures 63A et B). En
revanche, l'IL-13 n'a été détectée dans aucun de ces espaces. En plus de ces molécules, l'IL-10
et le TNF-α augmentent dans la cavité pleurale des souris Mfpos mais l'IL-10 diminue dans le
LBA de ces souris (Figures 63C et D). La production de cytokines semble plus faible chez les
souris Mfneg mais au vu du faible nombre de souris analysées, il est nécessaire d'accroitre leur
nombre pour conclure sur la situation des souris Mfneg (des échantillons ont été produits et
restent à analyser).
Pour évaluer les conséquences des changements d'expression de ces gènes, les
résultats de l'analyse des transcrits ont été interprétés avec Ingenuity Pathway Analysis (IPA),
un logiciel d'analyse des réseaux biologiques utilisant une vaste base de données. A partir des
profils d'expression des différents gènes, le logiciel prédit les fonctions biologiques
activées/réprimées et les pathologies associées. Parmi les conséquences prédites de l'infection
(Figure 64A), des pathologies pulmonaires sont suggérées comme l'asthme, la fibrose
pulmonaire et l'hypersensibilité aérienne. De plus, ces gènes sont impliqués dans la
chimiotaxie et l'activation des macrophages, des éosinophiles, des neutrophiles et des
lymphocytes T CD4+.
Particulièrement, l'IL-4 et l'IL-13 semblent avoir un rôle central dans la mise en place
de ces réponses (Figure 64B).
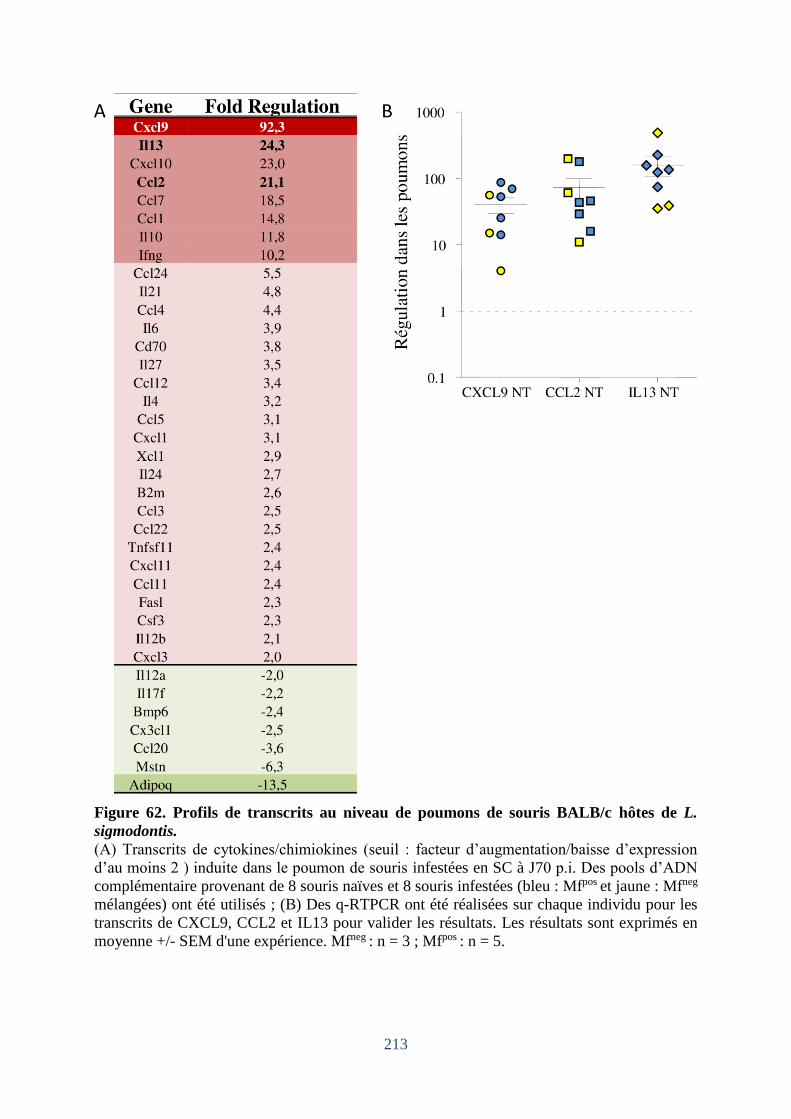
213
Figure 62. Profils de transcrits au niveau de poumons de souris BALB/c hôtes de L.
sigmodontis.
(A) Transcrits de cytokines/chimiokines (seuil : facteur d’augmentation/baisse d’expression
d’au moins 2 ) induite dans le poumon de souris infestées en SC à J70 p.i. Des pools d’ADN
complémentaire provenant de 8 souris naïves et 8 souris infestées (bleu : Mfpos et jaune : Mfneg
mélangées) ont été utilisés ; (B) Des q-RTPCR ont été réalisées sur chaque individu pour les
transcrits de CXCL9, CCL2 et IL13 pour valider les résultats. Les résultats sont exprimés en
moyenne +/- SEM d'une expérience. Mfneg : n = 3 ; Mfpos : n = 5.

214
Figure 63. Cytokines et chimiokines dans le LBA et la CP de BALB/c Mfposvs Mfneg.
Le liquide de lavage bronchoalvéolaire (LBA), le liquide pleural (CP) et le sérum de souris
utilisées pour l'analyse des transcrits ont été récupérés. Des ELISA ont été réalisés pour doser
(A) le CXCL9, (B) le CCL2, (C) l'IL10 et (D) le TNF-α. Les résultats sont exprimés en
moyenne +/- SEM d'une expérience. Les différences ont été testées par le test U de Mann-
Whitney (*p<0,05, **p<0,01, ***p<0,001). N : n=8 souris. Mfneg : n=1-3 souris ; Mfpos : n = 8
souris/groupe.

215
Figure 64. Analyse des réseaux cytokiniques et fonctions biologiques potentielles
(A) Les effets induits par les changements d’expression génique ont été prédits en utilisant le
logiciel Ingenuity Pathway Analysis ; (B) Représentation graphique des effets de différentes
cytokines sur l'activation des macrophages, le recrutement des neutrophiles et la fibrose
pulmonaire. Noter le rôle central de l'IL-4 et de l'IL-13 dans les 3 processus. Flèche : le profil
d’expression du gène favorise la fonction; Râteau : le profil d’expression du gène inhibe la
fonction. Molécule rouge : expression augmentée. Molécule verte : expression diminuée.
Flèches rouge : le résultat amplifie la fonction; Flèches jaunes : le résultat inhibe la fonction;
Flèche grise : la molécule est impliquée dans le mécanisme.

216
4. Production de mucus par l'épithélium bronchique de souris hôtes de microfilaires
La différenciation des cellules épithéliales bronchiques en cellules caliciformes
(appelée métaplasie des cellules caliciformes) et leur production de mucus est une des
caractéristiques de l'inflammation pulmonaire. Afin de déterminer la conséquence de la
présence des filaires dans et autour du poumon, les cellules caliciformes ont été colorées sur
des sections de poumons de souris infestées ou non (J70 p.i.). En effet, l'analyse des transcrits
prédit des réponses asthmatiques dans le poumon avec une stimulation des cellules
épithéliales (Figure 64). La fixation de l'IL-4 mais surtout de l'IL-13 sur leur récepteur (IL-
4R) sont les principales causes de la métaplasie des cellules caliciformes [468,469].
Les bronches des souris non infestées ne contiennent aucune cellule caliciforme
(Figure 65A). A l'inverse, des cellules caliciformes sont observées dans l'épithélium
bronchique des poumons des souris infestées (Figures 65B et 65C). Chez les souris
microfilarémiques (Mf+), les cellules caliciformes sont majoritairement présentes à proximité
des espaces périvasculaires (PVS) (Figure 8C). Des bronches enflammées sont aussi
observées à proximité des granulomes pulmonaires (Figure 66C). La présence des cellules
caliciformes a été quantifiée chez les souris Mfpos et Mfneg (Figure 66D). Les bronches des
souris amicrofilarémiques présentent une faible inflammation bronchique (environ 10% des
sections de bronches contiennent des cellules caliciformes) comparé aux souris Mfpos (environ
35%).
La localisation privilégiée des cellules caliciformes en bordure de PVS pourrait être la
conséquence d'une production localisée d'IL-13 dans ce PVS. Ceci expliquerait pourquoi les
transcrits d'IL-13 étaient augmentés dans les poumons mais pas dans les espaces environnants
(liquide broncho-alvéolaire, sang et cavité pleurale). La présence de Mfs dans le PVS ou dans
les vaisseaux sanguins du PVS pourrait stimuler les cellules résidentes du PVS , comme les
macrophages interstitiels [318], qui produiraient alors de l'IL-13.
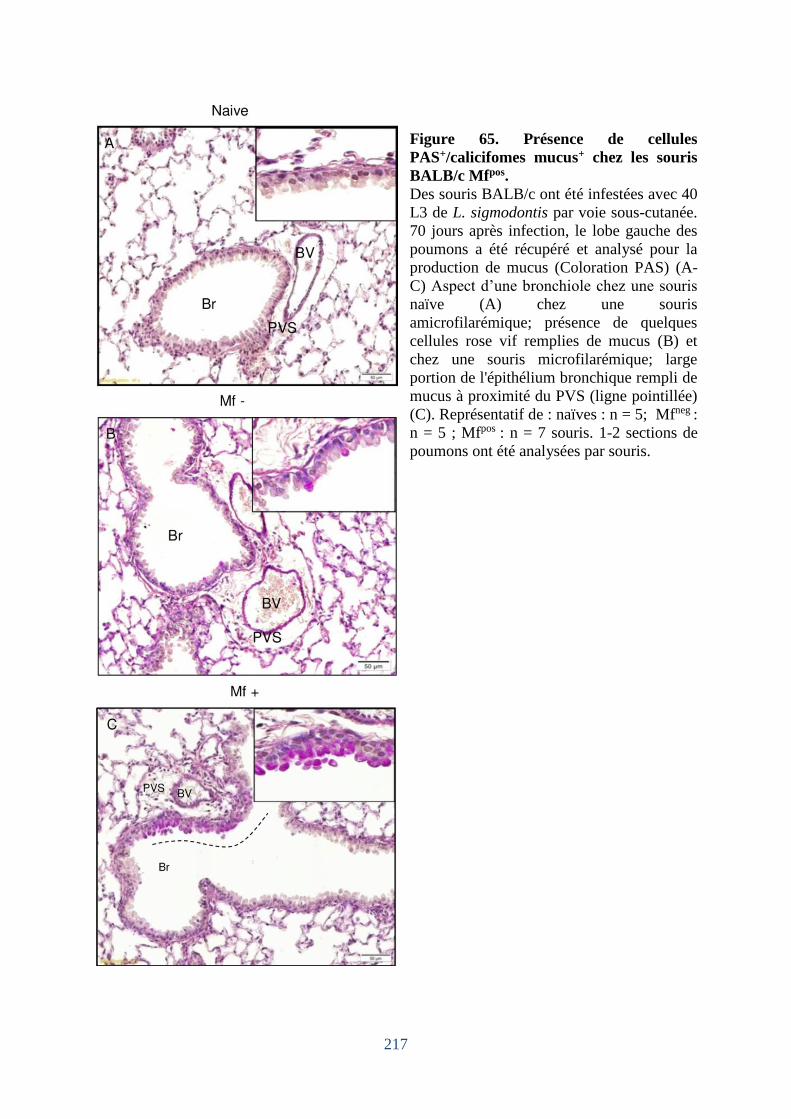
217
Figure 65. Présence de cellules
PAS+/calicifomes mucus+ chez les souris
BALB/c Mfpos.
Des souris BALB/c ont été infestées avec 40
L3 de L. sigmodontis par voie sous-cutanée.
70 jours après infection, le lobe gauche des
poumons a été récupéré et analysé pour la
production de mucus (Coloration PAS) (A-
C) Aspect d’une bronchiole chez une souris
naïve (A) chez une souris
amicrofilarémique; présence de quelques
cellules rose vif remplies de mucus (B) et
chez une souris microfilarémique; large
portion de l'épithélium bronchique rempli de
mucus à proximité du PVS (ligne pointillée)
(C). Représentatif de : naïves : n = 5; Mfneg :
n = 5 ; Mfpos : n = 7 souris. 1-2 sections de
poumons ont été analysées par souris.
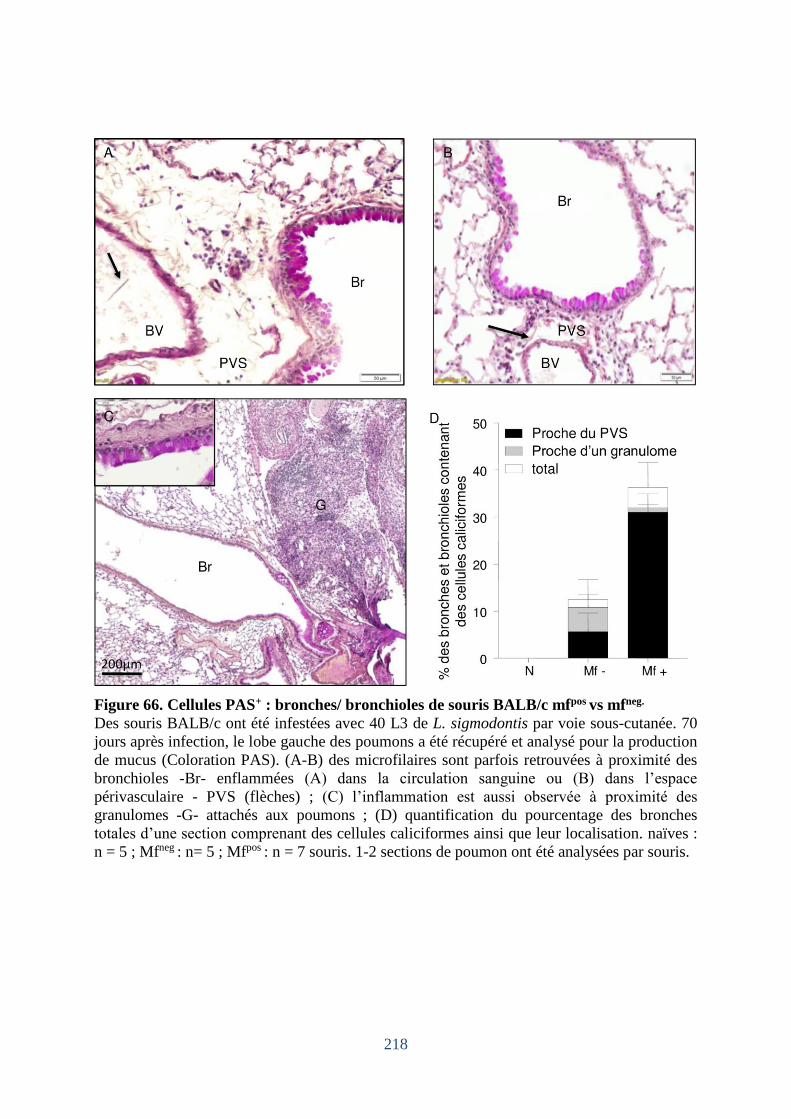
218
Figure 66. Cellules PAS+ : bronches/ bronchioles de souris BALB/c mfpos vs mfneg.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
jours après infection, le lobe gauche des poumons a été récupéré et analysé pour la production
de mucus (Coloration PAS). (A-B) des microfilaires sont parfois retrouvées à proximité des
bronchioles -Br- enflammées (A) dans la circulation sanguine ou (B) dans l’espace
périvasculaire - PVS (flèches) ; (C) l’inflammation est aussi observée à proximité des
granulomes -G- attachés aux poumons ; (D) quantification du pourcentage des bronches
totales d’une section comprenant des cellules caliciformes ainsi que leur localisation. naïves :
n = 5 ; Mfneg : n= 5 ; Mfpos : n = 7 souris. 1-2 sections de poumon ont été analysées par souris.
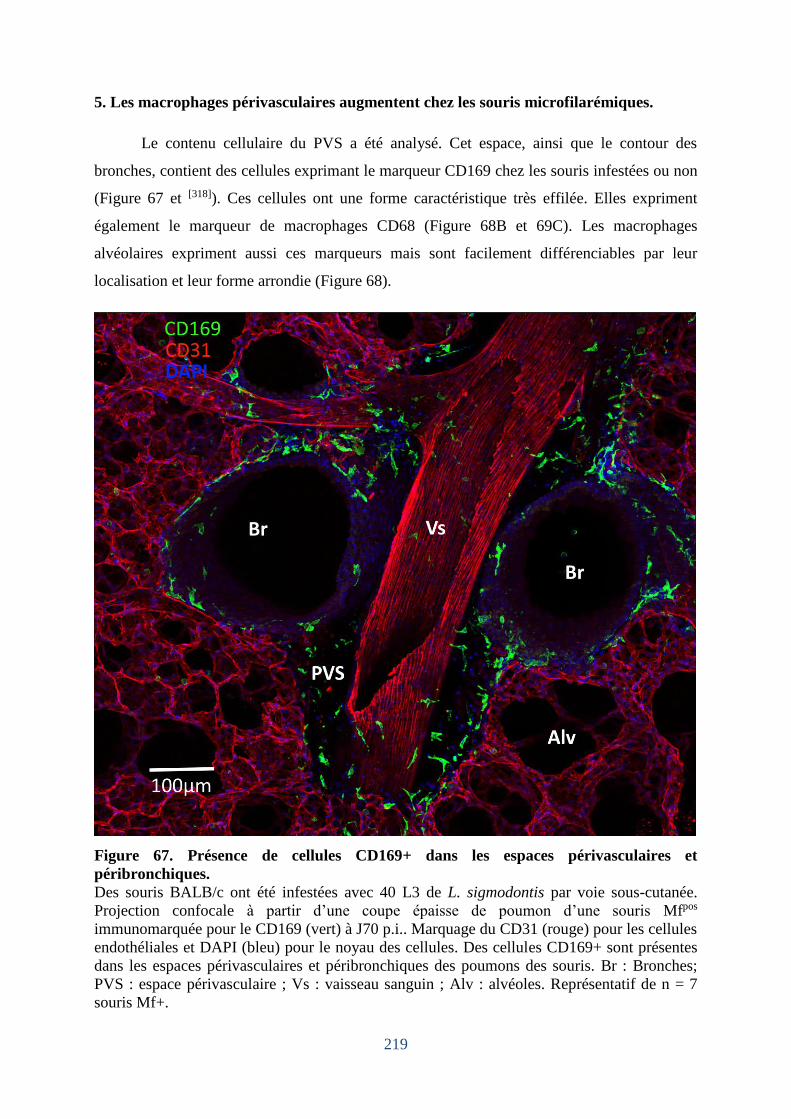
219
5. Les macrophages périvasculaires augmentent chez les souris microfilarémiques.
Le contenu cellulaire du PVS a été analysé. Cet espace, ainsi que le contour des
bronches, contient des cellules exprimant le marqueur CD169 chez les souris infestées ou non
(Figure 67 et [318]). Ces cellules ont une forme caractéristique très effilée. Elles expriment
également le marqueur de macrophages CD68 (Figure 68B et 69C). Les macrophages
alvéolaires expriment aussi ces marqueurs mais sont facilement différenciables par leur
localisation et leur forme arrondie (Figure 68).
Figure 67. Présence de cellules CD169+ dans les espaces périvasculaires et
péribronchiques.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée.
Projection confocale à partir d’une coupe épaisse de poumon d’une souris Mfpos
immunomarquée pour le CD169 (vert) à J70 p.i.. Marquage du CD31 (rouge) pour les cellules
endothéliales et DAPI (bleu) pour le noyau des cellules. Des cellules CD169+ sont présentes
dans les espaces périvasculaires et péribronchiques des poumons des souris. Br : Bronches;
PVS : espace périvasculaire ; Vs : vaisseau sanguin ; Alv : alvéoles. Représentatif de n = 7
souris Mf+.
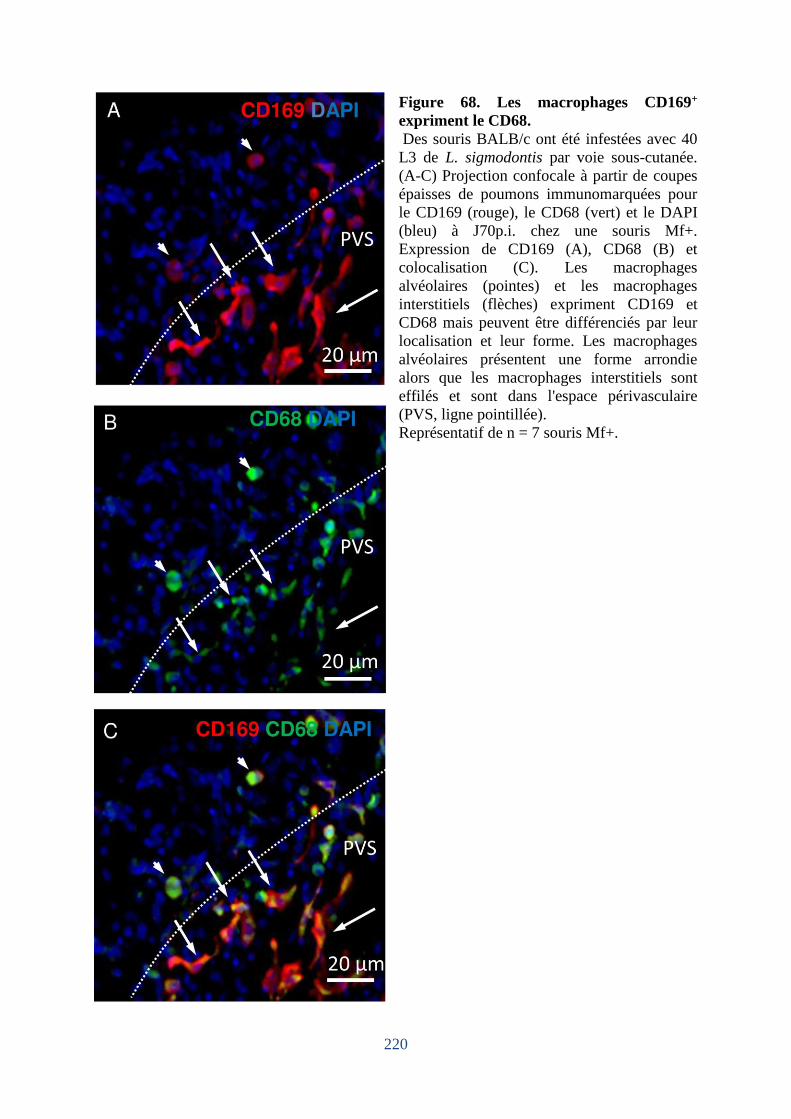
220
Figure 68. Les macrophages CD169+
expriment le CD68.
Des souris BALB/c ont été infestées avec 40
L3 de L. sigmodontis par voie sous-cutanée.
(A-C) Projection confocale à partir de coupes
épaisses de poumons immunomarquées pour
le CD169 (rouge), le CD68 (vert) et le DAPI
(bleu) à J70p.i. chez une souris Mf+.
Expression de CD169 (A), CD68 (B) et
colocalisation (C). Les macrophages
alvéolaires (pointes) et les macrophages
interstitiels (flèches) expriment CD169 et
CD68 mais peuvent être différenciés par leur
localisation et leur forme. Les macrophages
alvéolaires présentent une forme arrondie
alors que les macrophages interstitiels sont
effilés et sont dans l'espace périvasculaire
(PVS, ligne pointillée).
Représentatif de n = 7 souris Mf+.
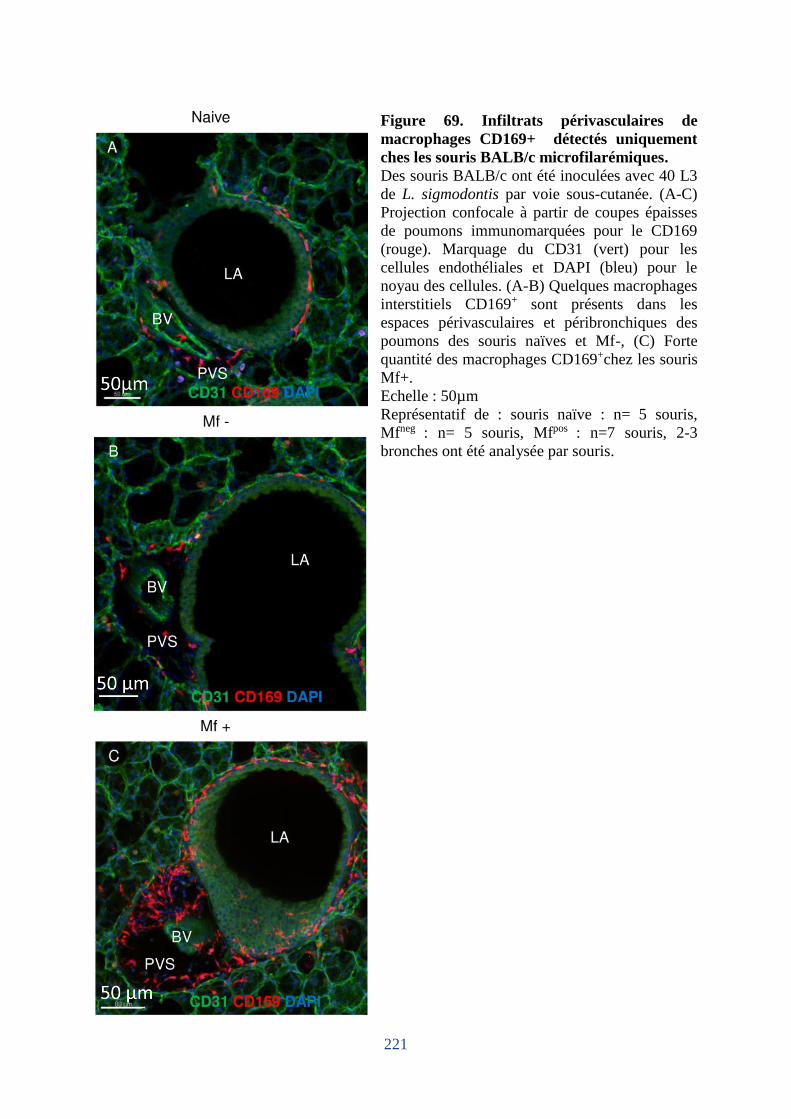
221
Figure 69. Infiltrats périvasculaires de
macrophages CD169+ détectés uniquement
ches les souris BALB/c microfilarémiques.
Des souris BALB/c ont été inoculées avec 40 L3
de L. sigmodontis par voie sous-cutanée. (A-C)
Projection confocale à partir de coupes épaisses
de poumons immunomarquées pour le CD169
(rouge). Marquage du CD31 (vert) pour les
cellules endothéliales et DAPI (bleu) pour le
noyau des cellules. (A-B) Quelques macrophages
interstitiels CD169+ sont présents dans les
espaces périvasculaires et péribronchiques des
poumons des souris naïves et Mf-, (C) Forte
quantité des macrophages CD169+chez les souris
Mf+.
Echelle : 50µm
Représentatif de : souris naïve : n= 5 souris,
Mfneg : n= 5 souris, Mfpos : n=7 souris, 2-3
bronches ont été analysée par souris.
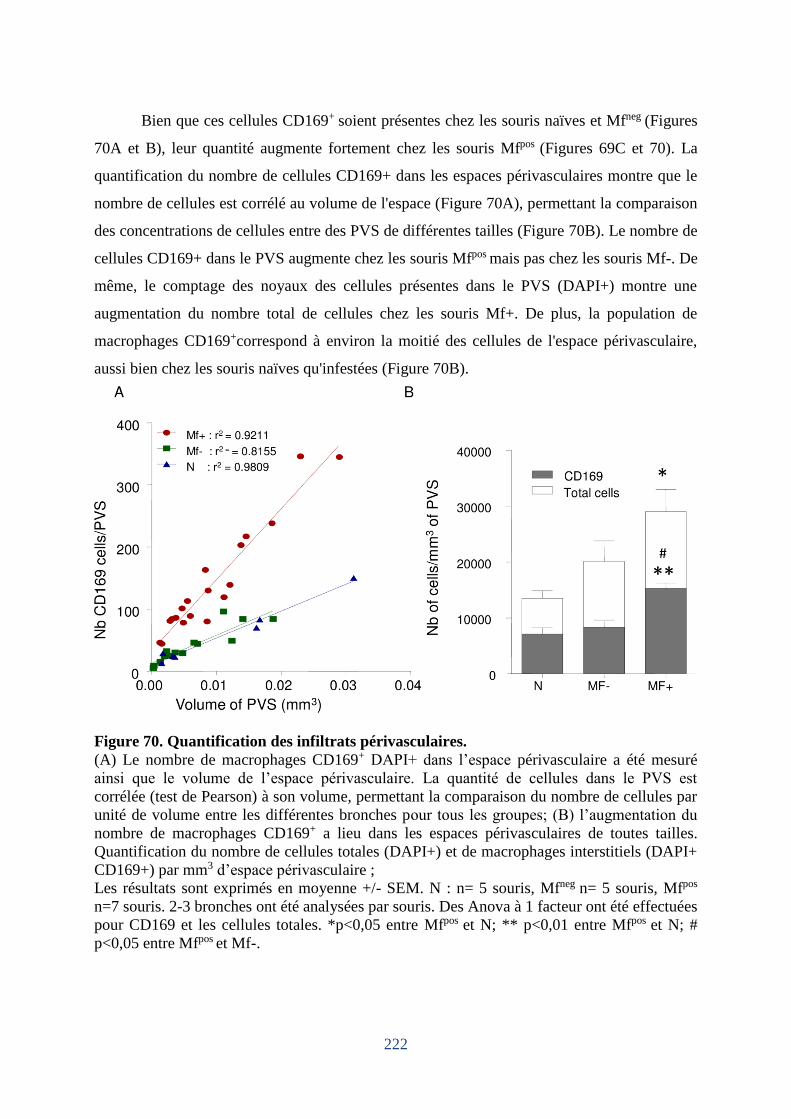
222
Bien que ces cellules CD169+ soient présentes chez les souris naïves et Mfneg (Figures
70A et B), leur quantité augmente fortement chez les souris Mfpos (Figures 69C et 70). La
quantification du nombre de cellules CD169+ dans les espaces périvasculaires montre que le
nombre de cellules est corrélé au volume de l'espace (Figure 70A), permettant la comparaison
des concentrations de cellules entre des PVS de différentes tailles (Figure 70B). Le nombre de
cellules CD169+ dans le PVS augmente chez les souris Mfpos mais pas chez les souris Mf-. De
même, le comptage des noyaux des cellules présentes dans le PVS (DAPI+) montre une
augmentation du nombre total de cellules chez les souris Mf+. De plus, la population de
macrophages CD169+correspond à environ la moitié des cellules de l'espace périvasculaire,
aussi bien chez les souris naïves qu'infestées (Figure 70B).
Figure 70. Quantification des infiltrats périvasculaires.
(A) Le nombre de macrophages CD169+ DAPI+ dans l’espace périvasculaire a été mesuré
ainsi que le volume de l’espace périvasculaire. La quantité de cellules dans le PVS est
corrélée (test de Pearson) à son volume, permettant la comparaison du nombre de cellules par
unité de volume entre les différentes bronches pour tous les groupes; (B) l’augmentation du
nombre de macrophages CD169+ a lieu dans les espaces périvasculaires de toutes tailles.
Quantification du nombre de cellules totales (DAPI+) et de macrophages interstitiels (DAPI+
CD169+) par mm3 d’espace périvasculaire ;
Les résultats sont exprimés en moyenne +/- SEM. N : n= 5 souris, Mfneg n= 5 souris, Mfpos
n=7 souris. 2-3 bronches ont été analysées par souris. Des Anova à 1 facteur ont été effectuées
pour CD169 et les cellules totales. *p<0,05 entre Mfpos et N; ** p<0,01 entre Mfpos et N; #
p<0,05 entre Mfpos et Mf-.
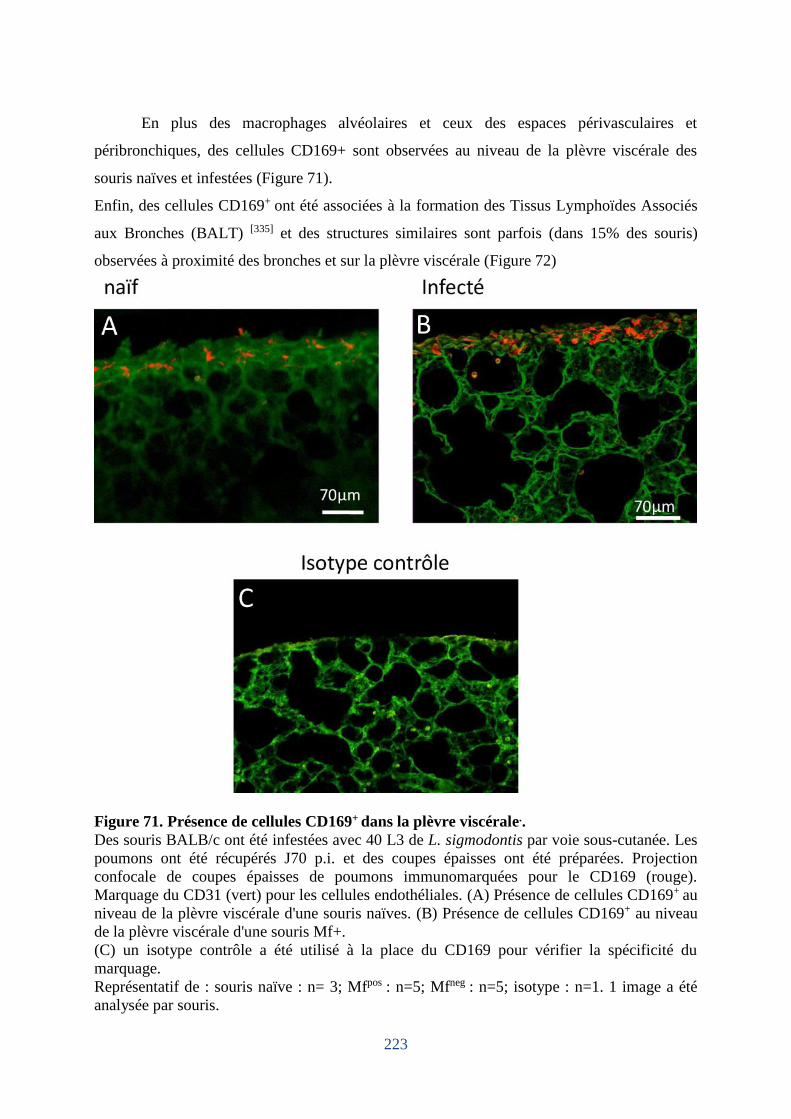
223
En plus des macrophages alvéolaires et ceux des espaces périvasculaires et
péribronchiques, des cellules CD169+ sont observées au niveau de la plèvre viscérale des
souris naïves et infestées (Figure 71).
Enfin, des cellules CD169+ ont été associées à la formation des Tissus Lymphoïdes Associés
aux Bronches (BALT) [335] et des structures similaires sont parfois (dans 15% des souris)
observées à proximité des bronches et sur la plèvre viscérale (Figure 72)
Figure 71. Présence de cellules CD169+ dans la plèvre viscérale..
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. Les
poumons ont été récupérés J70 p.i. et des coupes épaisses ont été préparées. Projection
confocale de coupes épaisses de poumons immunomarquées pour le CD169 (rouge).
Marquage du CD31 (vert) pour les cellules endothéliales. (A) Présence de cellules CD169+ au
niveau de la plèvre viscérale d'une souris naïves. (B) Présence de cellules CD169+ au niveau
de la plèvre viscérale d'une souris Mf+.
(C) un isotype contrôle a été utilisé à la place du CD169 pour vérifier la spécificité du
marquage.
Représentatif de : souris naïve : n= 3; Mfpos : n=5; Mfneg : n=5; isotype : n=1. 1 image a été
analysée par souris.
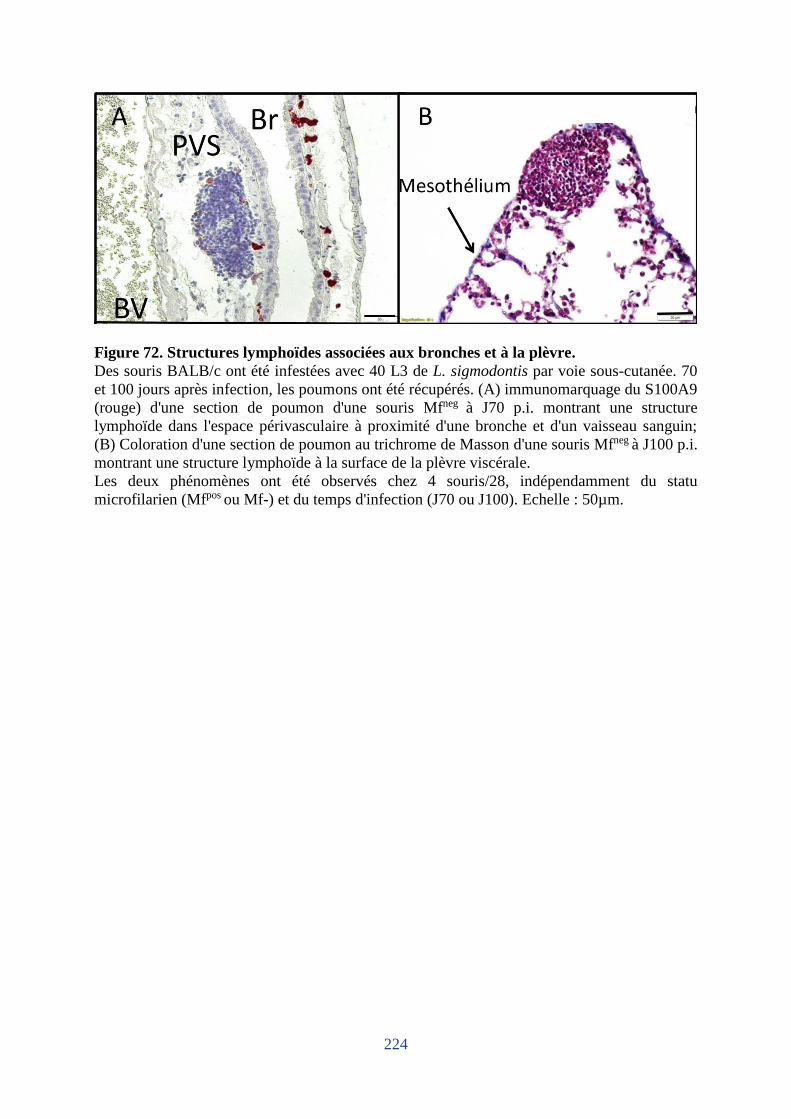
224
Figure 72. Structures lymphoïdes associées aux bronches et à la plèvre.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
et 100 jours après infection, les poumons ont été récupérés. (A) immunomarquage du S100A9
(rouge) d'une section de poumon d'une souris Mfneg à J70 p.i. montrant une structure
lymphoïde dans l'espace périvasculaire à proximité d'une bronche et d'un vaisseau sanguin;
(B) Coloration d'une section de poumon au trichrome de Masson d'une souris Mfneg à J100 p.i.
montrant une structure lymphoïde à la surface de la plèvre viscérale.
Les deux phénomènes ont été observés chez 4 souris/28, indépendamment du statu
microfilarien (Mfpos ou Mf-) et du temps d'infection (J70 ou J100). Echelle : 50µm.

225
6. Dommages du mésothélium pleural chez les souris hôtes de microfilaires.
Au vu de la pathologie pleurale observée chez la gerbille, l'état du mésothélium pleural
a été analysé chez les souris. L'observation de coupes histologiques montre un épaississement
partiel de la plèvre viscérale à J70 et J100 p.i. chez les souris Mf+, phénomène rarement
observé chez les souris Mfneg (Figure 73). L'observation des poumons en microscopie
électronique à balayage montre aussi un changement de morphologie des cellules
mésothéliales. Le mésothélium des souris naïves est lisse (pavimenteux) et il n'est pas
possible de différencier les limites des cellules (Figure 74A). A l'inverse, chez les souris
infestées (Mfpos et Mf-), les cellules mésothéliales sont hypertrophiées (leur volume
augmente) et elles prennent une forme "cubique" (Figure 74B). L'hypertrophie des cellules
mésothéliales est la conséquence de leur activation [470].
Figure 73. Epaississement pleural des souris microfilarémiques.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
et 100 jours après infection, les poumons ont été récupérés et le mésothélium a été analysé.
(A) Aspect global du lobe gauche du poumon d’une souris naïve, (B) d'une souris
amicrofilarémique Mfneg et (C) d'une souris microfilarémique Mfpos montrant épaississement
du mésothélium pleural chez les souris Mf+. Représentatif de : naïve : n = 6 ; Mfneg : n = 7 ;
Mfpos : n = 10 souris.

226
Figure 74. Hypertrophie des cellules mésothéliales chez les souris infestées.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
jours après infection, les poumons ont été récupérés et analysés en microscopie électronique à
balayage. (A) Aspect normal du mésothélium d'une souris naïve (B) aspect hypertrophique du
mésothélium chez une souris infestée. Echelle : 100 µm. n = 1 souris/groupe.
Le mésothélium sain est composé d'une monocouche de cellules pavimenteuses
exprimant la cytokératine 7 (Figure 75A). L'analyse du mésothélium pulmonaire des souris
infestées indique trois états possibles : 1) des cellules mésothéliales "normales"
pavimenteuses ; 2) des cellules mésothéliales hypertrophiques "cuboïdales" (Figure 75B); et
3) du mésothélium présentant une hyperplasie (épaississement de la plèvre du à une
augmentation du nombre de cellules) (Figure 75C). Dans les parties de mésothélium
hyperplasiques, seule la couche supérieure de cellules exprime le marqueur des cellules
mésothéliales (la cytokératine) (Figure 75C). Le périmètre des poumons des souris infestées
peut présenter les trois états mais l'hyperplasie est retrouvée majoritairement chez les souris
Mfpos : environ 40% du périmètre total du poumon des souris Mfpos est hyperplasique, contre
moins de 10% chez les souris Mfneg (Figure 75D). L'hypertrophie des cellules mésothéliales
est observée dès J30 p.i. [196], donc en phase pré-patente, mais à ce temps là, aucune
hyperplasie mésothéliale n'est visible. Les cellules mésothéliales semblent aussi présenter une
baisse d'adhérence à la lame basale. En effet, des zones de mésothélium décollé sont parfois
observées en microscopie électronique à balayage et sur les coupes histologiques (Annexe 2).
Cependant, ces zones pourraient aussi être des artéfacts dus à la manipulation des poumons
lors de la préparation des échantillons.
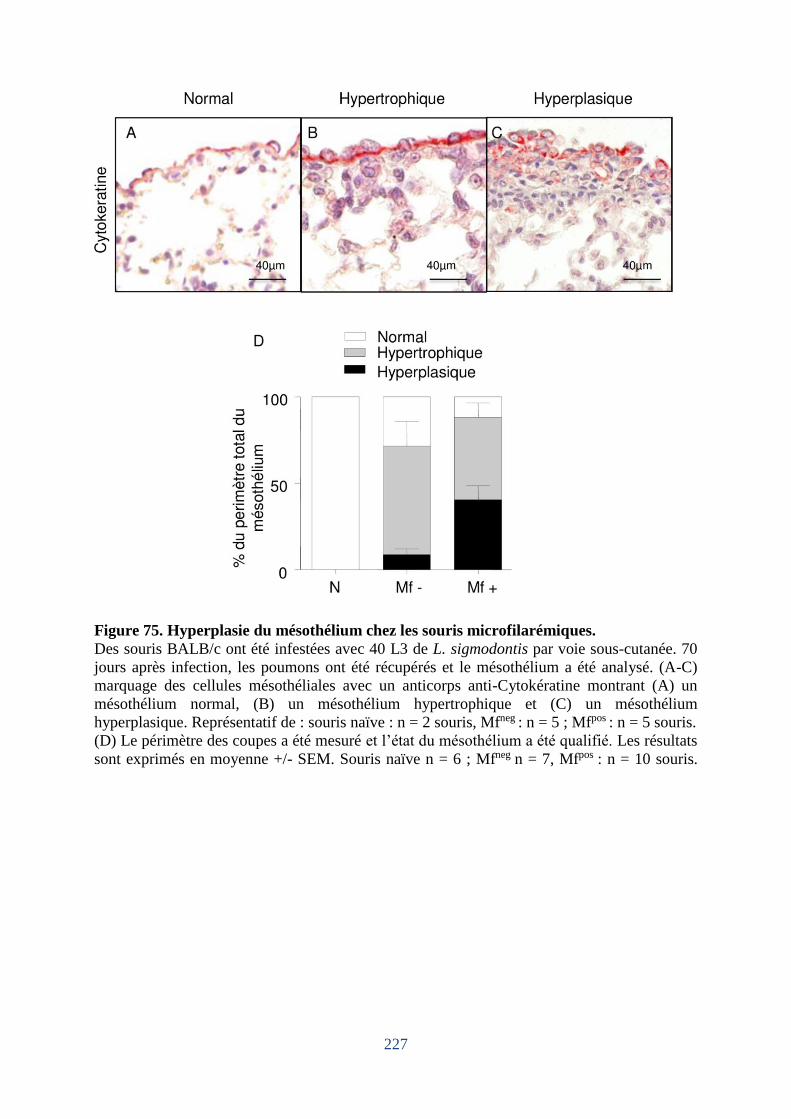
227
Figure 75. Hyperplasie du mésothélium chez les souris microfilarémiques.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
jours après infection, les poumons ont été récupérés et le mésothélium a été analysé. (A-C)
marquage des cellules mésothéliales avec un anticorps anti-Cytokératine montrant (A) un
mésothélium normal, (B) un mésothélium hypertrophique et (C) un mésothélium
hyperplasique. Représentatif de : souris naïve : n = 2 souris, Mfneg : n = 5 ; Mfpos : n = 5 souris.
(D) Le périmètre des coupes a été mesuré et l’état du mésothélium a été qualifié. Les résultats
sont exprimés en moyenne +/- SEM. Souris naïve n = 6 ; Mfneg n = 7, Mfpos : n = 10 souris.
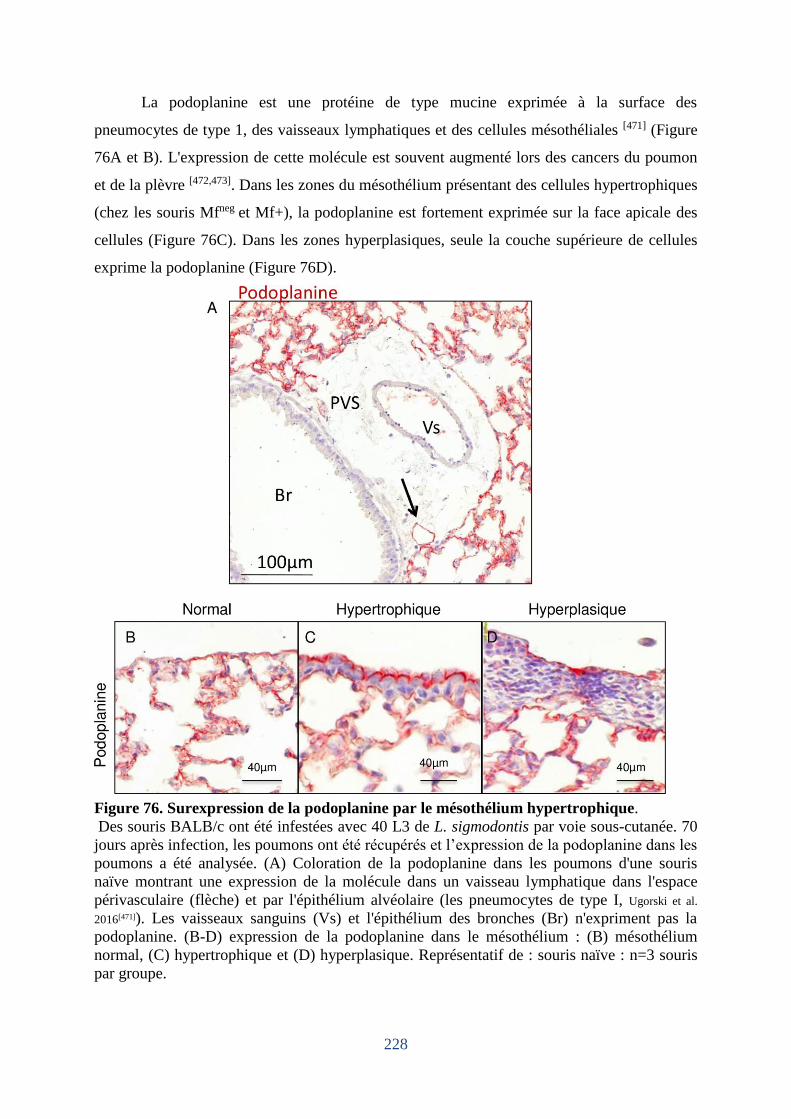
228
La podoplanine est une protéine de type mucine exprimée à la surface des
pneumocytes de type 1, des vaisseaux lymphatiques et des cellules mésothéliales [471] (Figure
76A et B). L'expression de cette molécule est souvent augmenté lors des cancers du poumon
et de la plèvre [472,473]. Dans les zones du mésothélium présentant des cellules hypertrophiques
(chez les souris Mfneg et Mf+), la podoplanine est fortement exprimée sur la face apicale des
cellules (Figure 76C). Dans les zones hyperplasiques, seule la couche supérieure de cellules
exprime la podoplanine (Figure 76D).
Figure 76. Surexpression de la podoplanine par le mésothélium hypertrophique.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
jours après infection, les poumons ont été récupérés et l’expression de la podoplanine dans les
poumons a été analysée. (A) Coloration de la podoplanine dans les poumons d'une souris
naïve montrant une expression de la molécule dans un vaisseau lymphatique dans l'espace
périvasculaire (flèche) et par l'épithélium alvéolaire (les pneumocytes de type I, Ugorski et al.
2016[471]). Les vaisseaux sanguins (Vs) et l'épithélium des bronches (Br) n'expriment pas la
podoplanine. (B-D) expression de la podoplanine dans le mésothélium : (B) mésothélium
normal, (C) hypertrophique et (D) hyperplasique. Représentatif de : souris naïve : n=3 souris
par groupe.
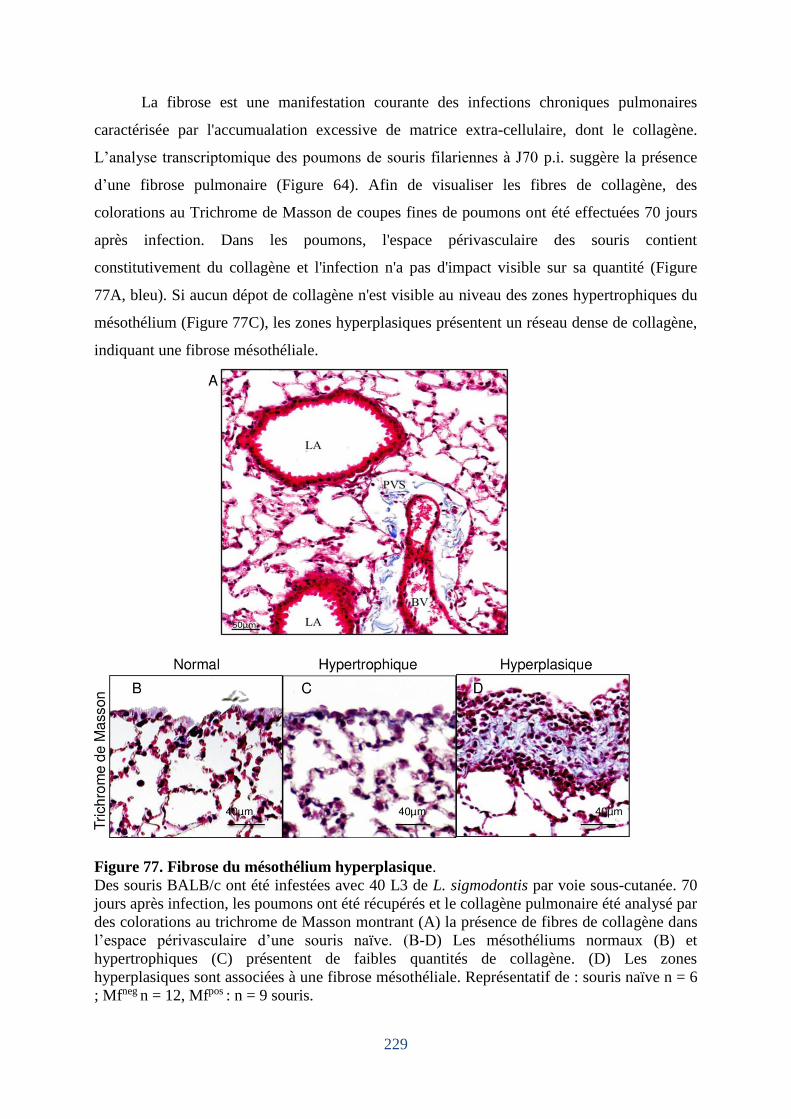
229
La fibrose est une manifestation courante des infections chroniques pulmonaires
caractérisée par l'accumualation excessive de matrice extra-cellulaire, dont le collagène.
L’analyse transcriptomique des poumons de souris filariennes à J70 p.i. suggère la présence
d’une fibrose pulmonaire (Figure 64). Afin de visualiser les fibres de collagène, des
colorations au Trichrome de Masson de coupes fines de poumons ont été effectuées 70 jours
après infection. Dans les poumons, l'espace périvasculaire des souris contient
constitutivement du collagène et l'infection n'a pas d'impact visible sur sa quantité (Figure
77A, bleu). Si aucun dépot de collagène n'est visible au niveau des zones hypertrophiques du
mésothélium (Figure 77C), les zones hyperplasiques présentent un réseau dense de collagène,
indiquant une fibrose mésothéliale.
Figure 77. Fibrose du mésothélium hyperplasique.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
jours après infection, les poumons ont été récupérés et le collagène pulmonaire été analysé par
des colorations au trichrome de Masson montrant (A) la présence de fibres de collagène dans
l’espace périvasculaire d’une souris naïve. (B-D) Les mésothéliums normaux (B) et
hypertrophiques (C) présentent de faibles quantités de collagène. (D) Les zones
hyperplasiques sont associées à une fibrose mésothéliale. Représentatif de : souris naïve n = 6
; Mfneg n = 12, Mfpos : n = 9 souris.
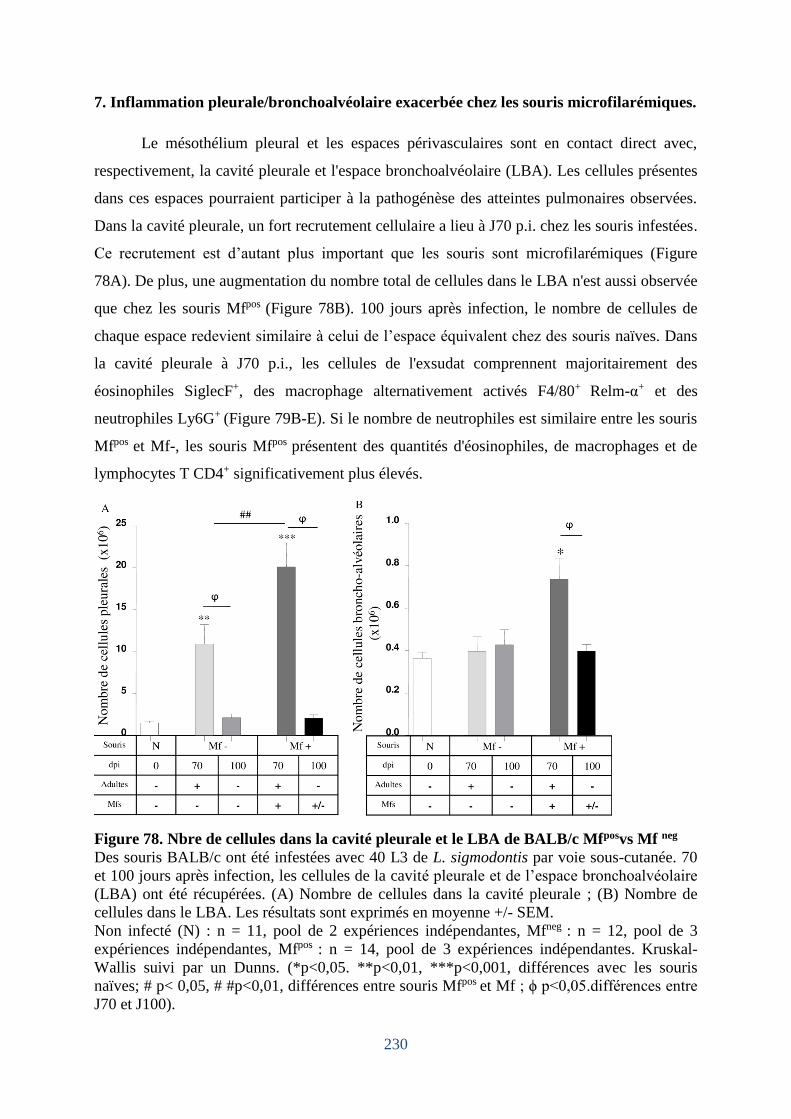
230
7. Inflammation pleurale/bronchoalvéolaire exacerbée chez les souris microfilarémiques.
Le mésothélium pleural et les espaces périvasculaires sont en contact direct avec,
respectivement, la cavité pleurale et l'espace bronchoalvéolaire (LBA). Les cellules présentes
dans ces espaces pourraient participer à la pathogénèse des atteintes pulmonaires observées.
Dans la cavité pleurale, un fort recrutement cellulaire a lieu à J70 p.i. chez les souris infestées.
Ce recrutement est d’autant plus important que les souris sont microfilarémiques (Figure
78A). De plus, une augmentation du nombre total de cellules dans le LBA n'est aussi observée
que chez les souris Mfpos (Figure 78B). 100 jours après infection, le nombre de cellules de
chaque espace redevient similaire à celui de l’espace équivalent chez des souris naïves. Dans
la cavité pleurale à J70 p.i., les cellules de l'exsudat comprennent majoritairement des
éosinophiles SiglecF+, des macrophage alternativement activés F4/80+ Relm-α+ et des
neutrophiles Ly6G+ (Figure 79B-E). Si le nombre de neutrophiles est similaire entre les souris
Mfpos et Mf-, les souris Mfpos présentent des quantités d'éosinophiles, de macrophages et de
lymphocytes T CD4+ significativement plus élevés.
Figure 78. Nbre de cellules dans la cavité pleurale et le LBA de BALB/c Mfposvs Mf neg
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
et 100 jours après infection, les cellules de la cavité pleurale et de l’espace bronchoalvéolaire
(LBA) ont été récupérées. (A) Nombre de cellules dans la cavité pleurale ; (B) Nombre de
cellules dans le LBA. Les résultats sont exprimés en moyenne +/- SEM.
Non infecté (N) : n = 11, pool de 2 expériences indépendantes, Mfneg : n = 12, pool de 3
expériences indépendantes, Mfpos : n = 14, pool de 3 expériences indépendantes. Kruskal-
Wallis suivi par un Dunns. (*p<0,05. **p<0,01, ***p<0,001, différences avec les souris
naïves; # p< 0,05, # #p<0,01, différences entre souris Mfpos et Mf ; ϕ p<0,05.différences entre
J70 et J100).
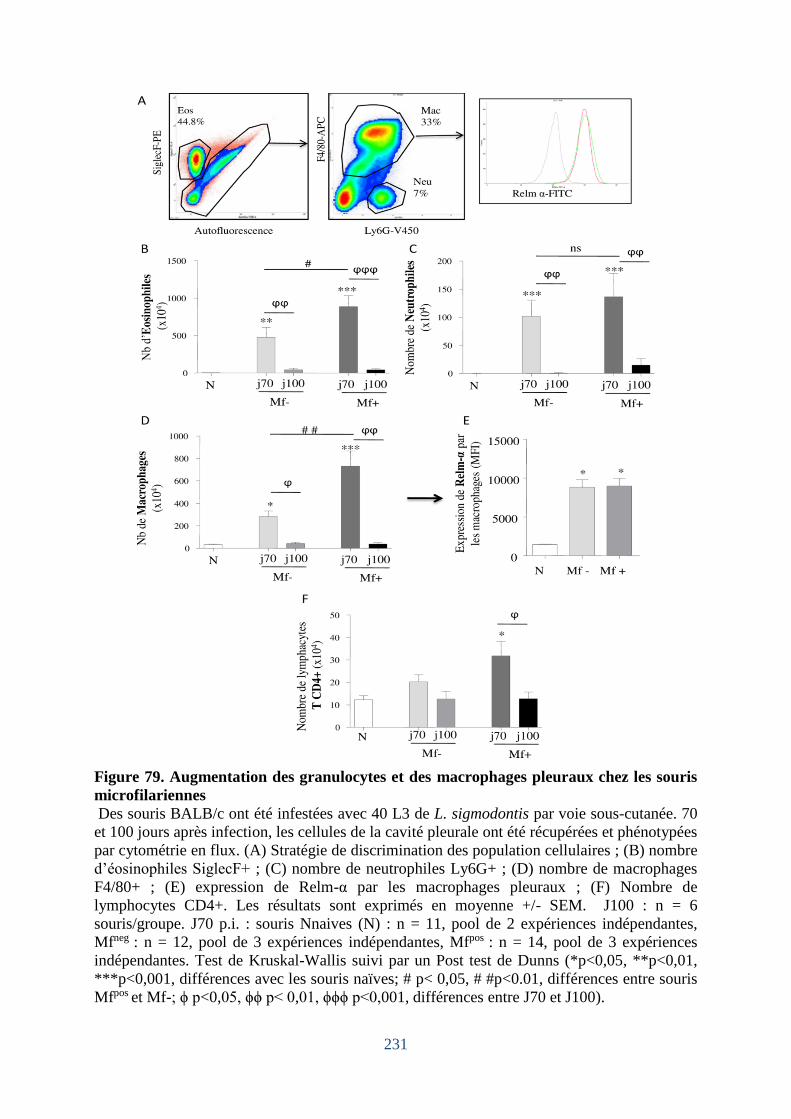
231
Figure 79. Augmentation des granulocytes et des macrophages pleuraux chez les souris
microfilariennes
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
et 100 jours après infection, les cellules de la cavité pleurale ont été récupérées et phénotypées
par cytométrie en flux. (A) Stratégie de discrimination des population cellulaires ; (B) nombre
d’éosinophiles SiglecF+ ; (C) nombre de neutrophiles Ly6G+ ; (D) nombre de macrophages
F4/80+ ; (E) expression de Relm-α par les macrophages pleuraux ; (F) Nombre de
lymphocytes CD4+. Les résultats sont exprimés en moyenne +/- SEM. J100 : n = 6
souris/groupe. J70 p.i. : souris Nnaives (N) : n = 11, pool de 2 expériences indépendantes,
Mfneg : n = 12, pool de 3 expériences indépendantes, Mfpos : n = 14, pool de 3 expériences
indépendantes. Test de Kruskal-Wallis suivi par un Post test de Dunns (*p<0,05, **p<0,01,
***p<0,001, différences avec les souris naïves; # p< 0,05, # #p<0.01, différences entre souris
Mfpos et Mf-; ϕ p<0,05, ϕϕ p< 0,01, ϕϕϕ p<0,001, différences entre J70 et J100).
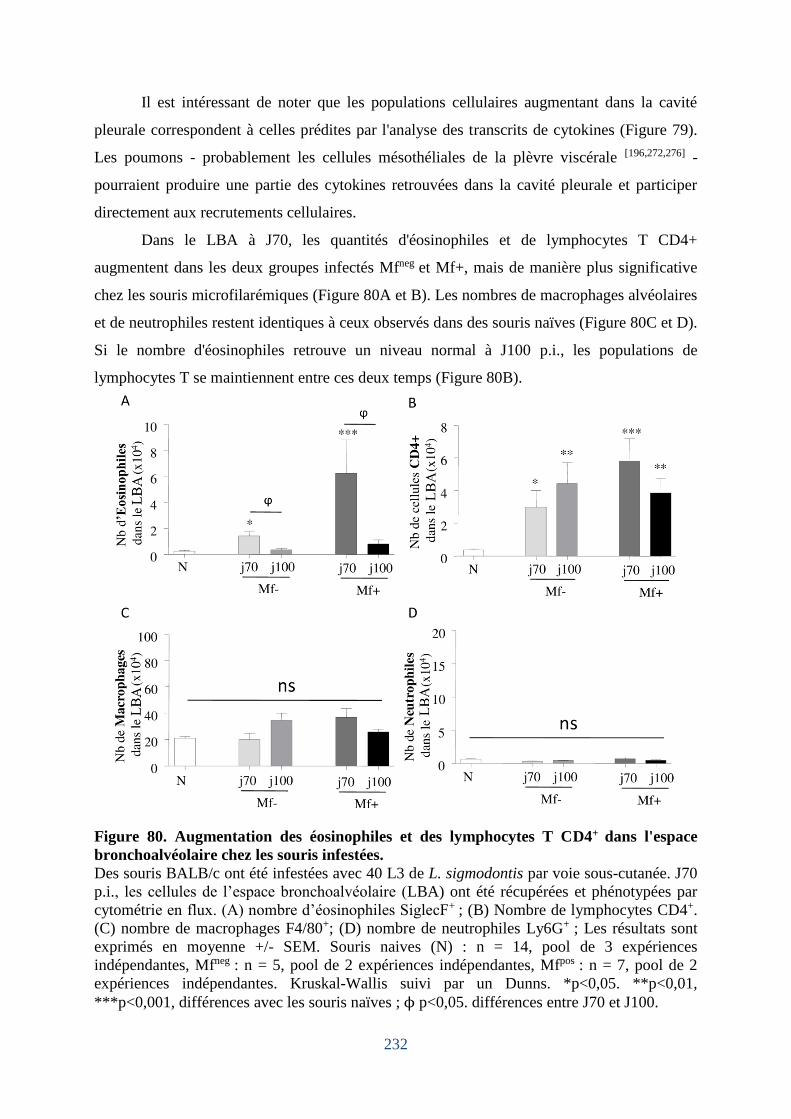
232
Il est intéressant de noter que les populations cellulaires augmentant dans la cavité
pleurale correspondent à celles prédites par l'analyse des transcrits de cytokines (Figure 79).
Les poumons - probablement les cellules mésothéliales de la plèvre viscérale [196,272,276] -
pourraient produire une partie des cytokines retrouvées dans la cavité pleurale et participer
directement aux recrutements cellulaires.
Dans le LBA à J70, les quantités d'éosinophiles et de lymphocytes T CD4+
augmentent dans les deux groupes infectés Mfneg et Mf+, mais de manière plus significative
chez les souris microfilarémiques (Figure 80A et B). Les nombres de macrophages alvéolaires
et de neutrophiles restent identiques à ceux observés dans des souris naïves (Figure 80C et D).
Si le nombre d'éosinophiles retrouve un niveau normal à J100 p.i., les populations de
lymphocytes T se maintiennent entre ces deux temps (Figure 80B).
Figure 80. Augmentation des éosinophiles et des lymphocytes T CD4+ dans l'espace
bronchoalvéolaire chez les souris infestées.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. J70
p.i., les cellules de l’espace bronchoalvéolaire (LBA) ont été récupérées et phénotypées par
cytométrie en flux. (A) nombre d’éosinophiles SiglecF+ ; (B) Nombre de lymphocytes CD4+.
(C) nombre de macrophages F4/80+; (D) nombre de neutrophiles Ly6G+ ; Les résultats sont
exprimés en moyenne +/- SEM. Souris naives (N) : n = 14, pool de 3 expériences
indépendantes, Mfneg : n = 5, pool de 2 expériences indépendantes, Mfpos : n = 7, pool de 2
expériences indépendantes. Kruskal-Wallis suivi par un Dunns. *p<0,05. **p<0,01,
***p<0,001, différences avec les souris naïves ; ϕ p<0,05. différences entre J70 et J100.
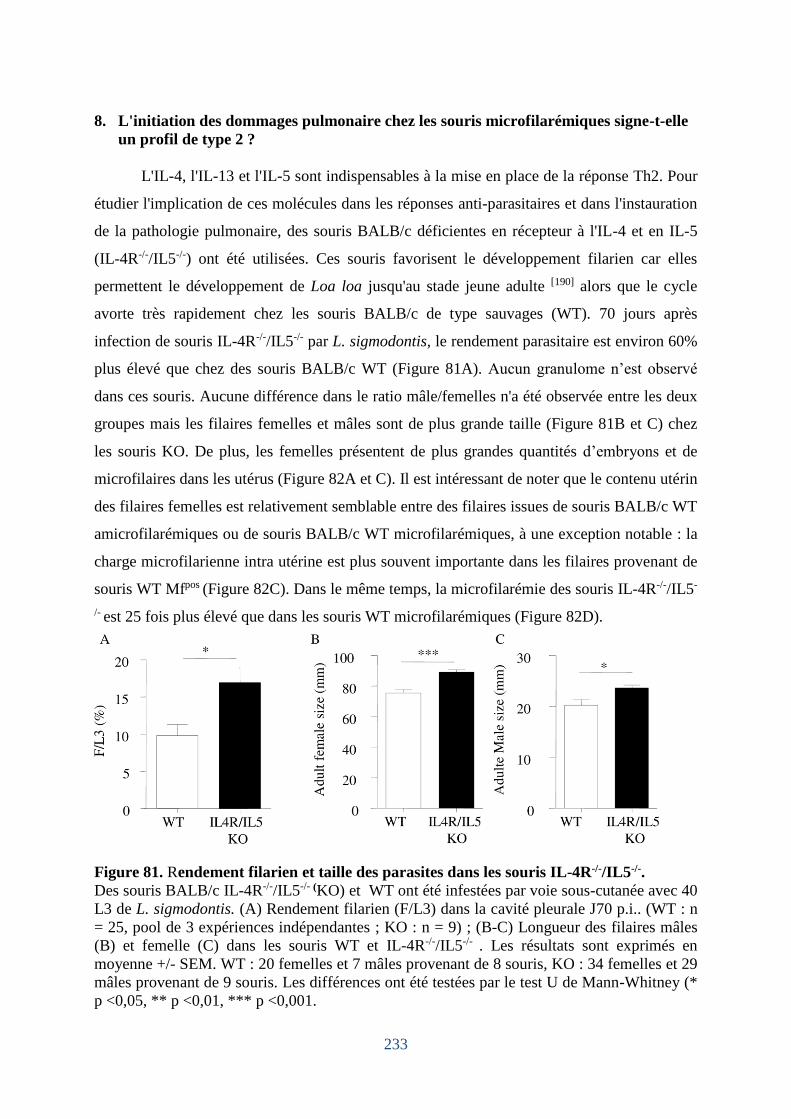
233
8. L'initiation des dommages pulmonaire chez les souris microfilarémiques signe-t-elle
un profil de type 2 ?
L'IL-4, l'IL-13 et l'IL-5 sont indispensables à la mise en place de la réponse Th2. Pour
étudier l'implication de ces molécules dans les réponses anti-parasitaires et dans l'instauration
de la pathologie pulmonaire, des souris BALB/c déficientes en récepteur à l'IL-4 et en IL-5
(IL-4R-/-/IL5-/-) ont été utilisées. Ces souris favorisent le développement filarien car elles
permettent le développement de Loa loa jusqu'au stade jeune adulte [190] alors que le cycle
avorte très rapidement chez les souris BALB/c de type sauvages (WT). 70 jours après
infection de souris IL-4R-/-/IL5-/- par L. sigmodontis, le rendement parasitaire est environ 60%
plus élevé que chez des souris BALB/c WT (Figure 81A). Aucun granulome n’est observé
dans ces souris. Aucune différence dans le ratio mâle/femelles n'a été observée entre les deux
groupes mais les filaires femelles et mâles sont de plus grande taille (Figure 81B et C) chez
les souris KO. De plus, les femelles présentent de plus grandes quantités d’embryons et de
microfilaires dans les utérus (Figure 82A et C). Il est intéressant de noter que le contenu utérin
des filaires femelles est relativement semblable entre des filaires issues de souris BALB/c WT
amicrofilarémiques ou de souris BALB/c WT microfilarémiques, à une exception notable : la
charge microfilarienne intra utérine est plus souvent importante dans les filaires provenant de
souris WT Mfpos (Figure 82C). Dans le même temps, la microfilarémie des souris IL-4R-/-/IL5-
/- est 25 fois plus élevé que dans les souris WT microfilarémiques (Figure 82D).
Figure 81. Rendement filarien et taille des parasites dans les souris IL-4R-/-/IL5-/-.
Des souris BALB/c IL-4R-/-/IL5-/- (KO) et WT ont été infestées par voie sous-cutanée avec 40
L3 de L. sigmodontis. (A) Rendement filarien (F/L3) dans la cavité pleurale J70 p.i.. (WT : n
= 25, pool de 3 expériences indépendantes ; KO : n = 9) ; (B-C) Longueur des filaires mâles
(B) et femelle (C) dans les souris WT et IL-4R-/-/IL5-/- . Les résultats sont exprimés en
moyenne +/- SEM. WT : 20 femelles et 7 mâles provenant de 8 souris, KO : 34 femelles et 29
mâles provenant de 9 souris. Les différences ont été testées par le test U de Mann-Whitney (*
p <0,05, ** p <0,01, *** p <0,001.
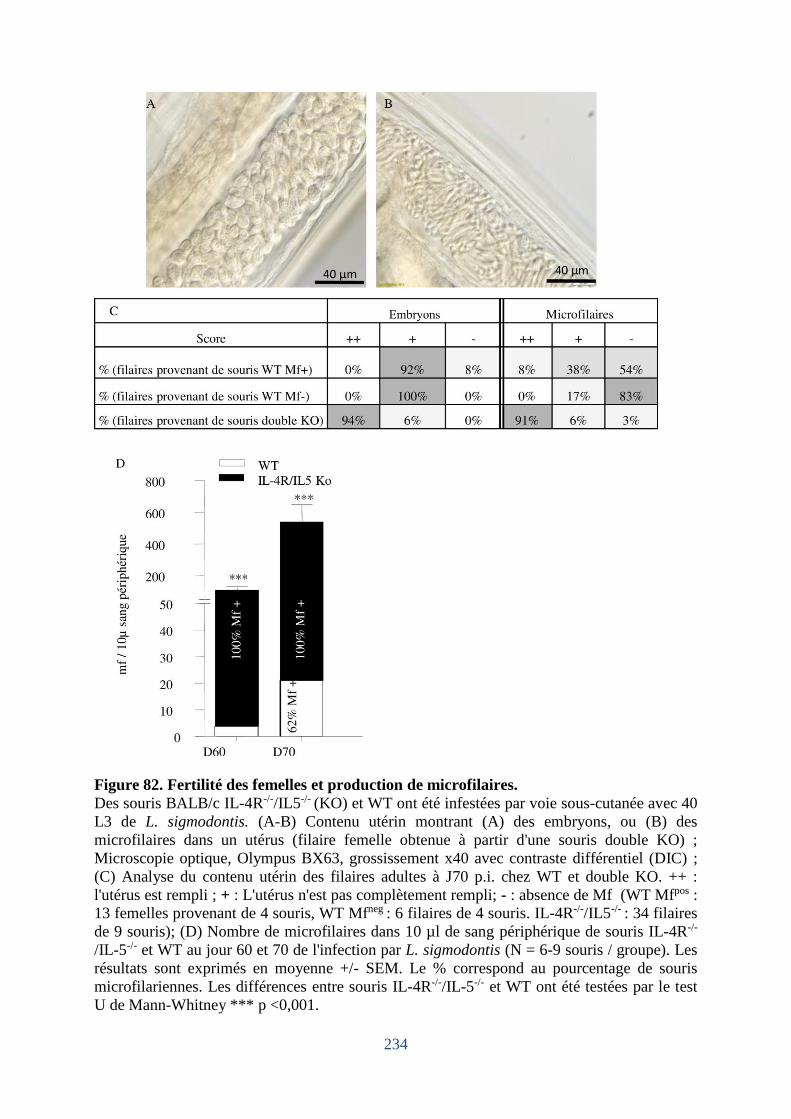
234
Figure 82. Fertilité des femelles et production de microfilaires.
Des souris BALB/c IL-4R-/-/IL5-/- (KO) et WT ont été infestées par voie sous-cutanée avec 40
L3 de L. sigmodontis. (A-B) Contenu utérin montrant (A) des embryons, ou (B) des
microfilaires dans un utérus (filaire femelle obtenue à partir d'une souris double KO) ;
Microscopie optique, Olympus BX63, grossissement x40 avec contraste différentiel (DIC) ;
(C) Analyse du contenu utérin des filaires adultes à J70 p.i. chez WT et double KO. ++ :
l'utérus est rempli ; + : L'utérus n'est pas complètement rempli; - : absence de Mf (WT Mfpos :
13 femelles provenant de 4 souris, WT Mfneg : 6 filaires de 4 souris. IL-4R-/-/IL5-/- : 34 filaires
de 9 souris); (D) Nombre de microfilaires dans 10 µl de sang périphérique de souris IL-4R-/-
/IL-5-/- et WT au jour 60 et 70 de l'infection par L. sigmodontis (N = 6-9 souris / groupe). Les
résultats sont exprimés en moyenne +/- SEM. Le % correspond au pourcentage de souris
microfilariennes. Les différences entre souris IL-4R-/-/IL-5-/- et WT ont été testées par le test
U de Mann-Whitney *** p <0,001.
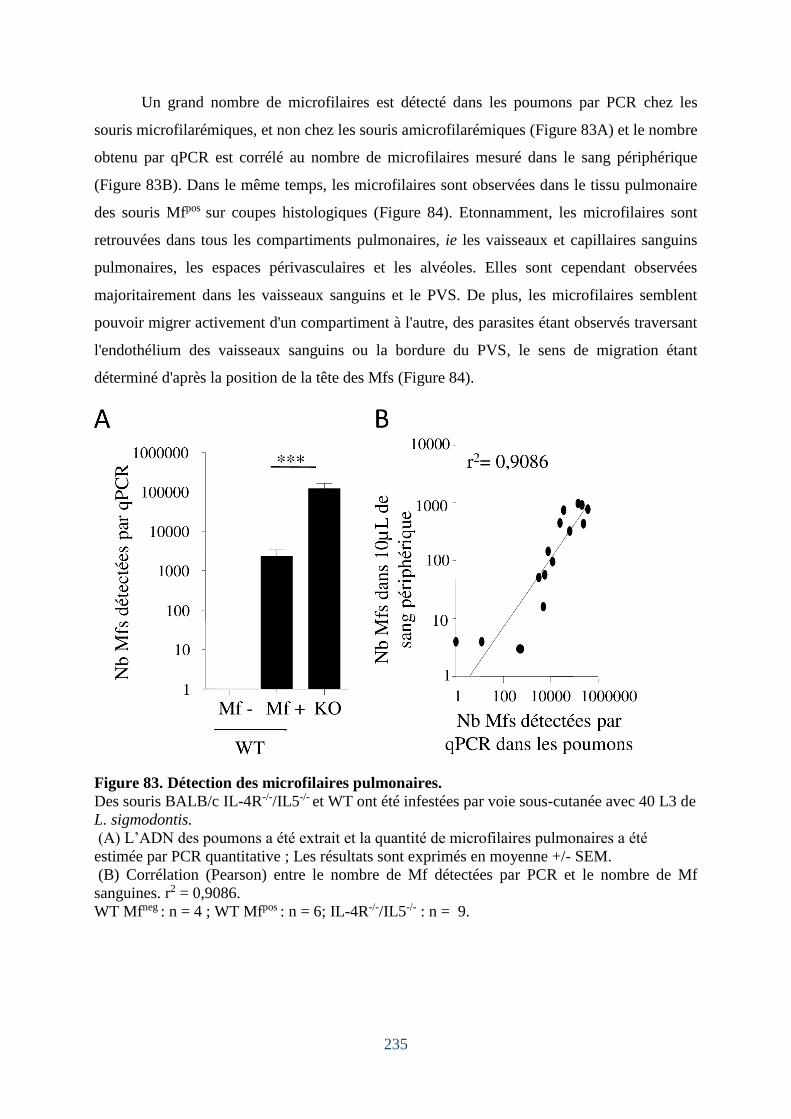
235
Un grand nombre de microfilaires est détecté dans les poumons par PCR chez les
souris microfilarémiques, et non chez les souris amicrofilarémiques (Figure 83A) et le nombre
obtenu par qPCR est corrélé au nombre de microfilaires mesuré dans le sang périphérique
(Figure 83B). Dans le même temps, les microfilaires sont observées dans le tissu pulmonaire
des souris Mfpos sur coupes histologiques (Figure 84). Etonnamment, les microfilaires sont
retrouvées dans tous les compartiments pulmonaires, ie les vaisseaux et capillaires sanguins
pulmonaires, les espaces périvasculaires et les alvéoles. Elles sont cependant observées
majoritairement dans les vaisseaux sanguins et le PVS. De plus, les microfilaires semblent
pouvoir migrer activement d'un compartiment à l'autre, des parasites étant observés traversant
l'endothélium des vaisseaux sanguins ou la bordure du PVS, le sens de migration étant
déterminé d'après la position de la tête des Mfs (Figure 84).
Figure 83. Détection des microfilaires pulmonaires.
Des souris BALB/c IL-4R-/-/IL5-/- et WT ont été infestées par voie sous-cutanée avec 40 L3 de
L. sigmodontis.
(A) L’ADN des poumons a été extrait et la quantité de microfilaires pulmonaires a été
estimée par PCR quantitative ; Les résultats sont exprimés en moyenne +/- SEM.
(B) Corrélation (Pearson) entre le nombre de Mf détectées par PCR et le nombre de Mf
sanguines. r2 = 0,9086.
WT Mfneg : n = 4 ; WT Mfpos : n = 6; IL-4R-/-/IL5-/- : n = 9.
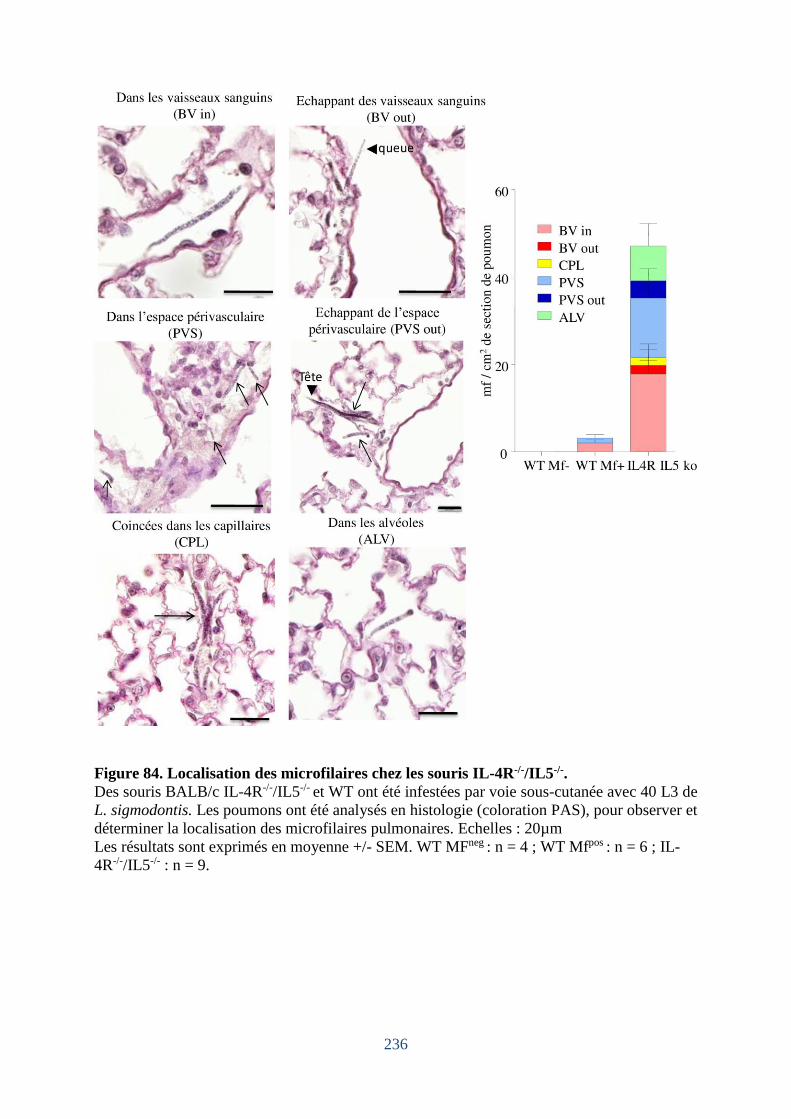
236
Figure 84. Localisation des microfilaires chez les souris IL-4R-/-/IL5-/-.
Des souris BALB/c IL-4R-/-/IL5-/- et WT ont été infestées par voie sous-cutanée avec 40 L3 de
L. sigmodontis. Les poumons ont été analysés en histologie (coloration PAS), pour observer et
déterminer la localisation des microfilaires pulmonaires. Echelles : 20µm
Les résultats sont exprimés en moyenne +/- SEM. WT MFneg : n = 4 ; WT Mfpos : n = 6 ; IL-
4R-/-/IL5-/- : n = 9.
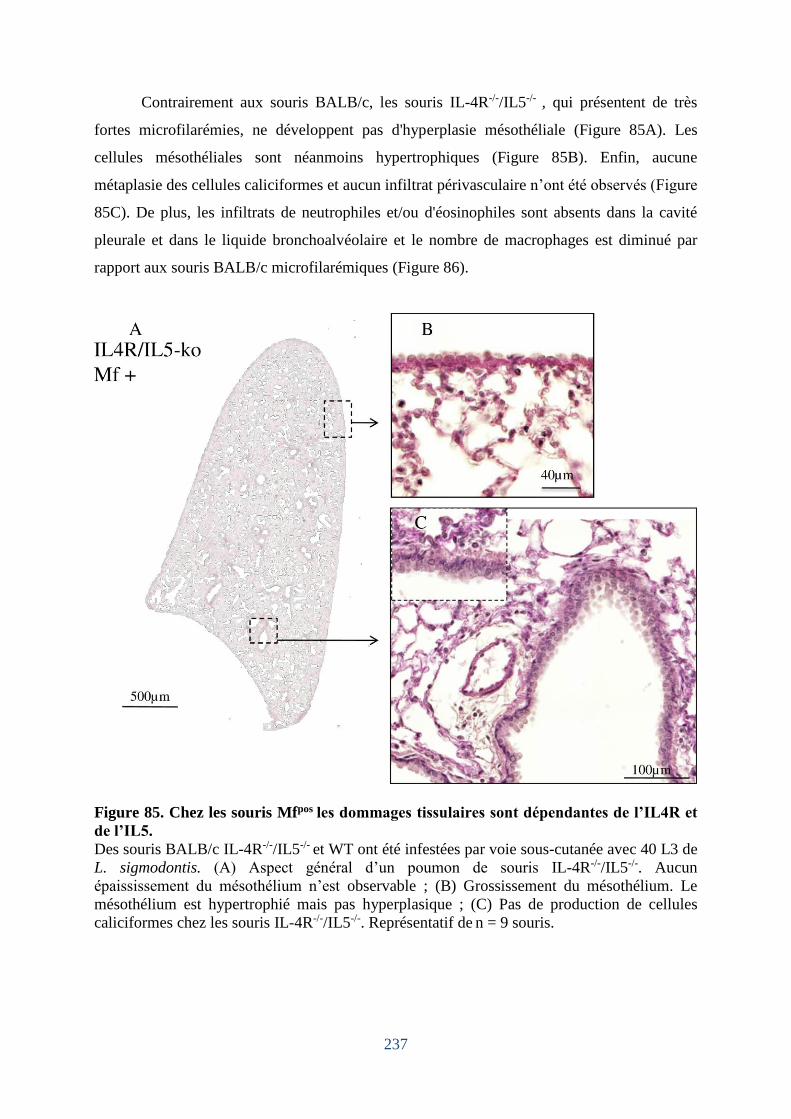
237
Contrairement aux souris BALB/c, les souris IL-4R-/-/IL5-/- , qui présentent de très
fortes microfilarémies, ne développent pas d'hyperplasie mésothéliale (Figure 85A). Les
cellules mésothéliales sont néanmoins hypertrophiques (Figure 85B). Enfin, aucune
métaplasie des cellules caliciformes et aucun infiltrat périvasculaire n’ont été observés (Figure
85C). De plus, les infiltrats de neutrophiles et/ou d'éosinophiles sont absents dans la cavité
pleurale et dans le liquide bronchoalvéolaire et le nombre de macrophages est diminué par
rapport aux souris BALB/c microfilarémiques (Figure 86).
Figure 85. Chez les souris Mfpos les dommages tissulaires sont dépendantes de l’IL4R et
de l’IL5.
Des souris BALB/c IL-4R-/-/IL5-/- et WT ont été infestées par voie sous-cutanée avec 40 L3 de
L. sigmodontis. (A) Aspect général d’un poumon de souris IL-4R-/-/IL5-/-. Aucun
épaississement du mésothélium n’est observable ; (B) Grossissement du mésothélium. Le
mésothélium est hypertrophié mais pas hyperplasique ; (C) Pas de production de cellules
caliciformes chez les souris IL-4R-/-/IL5-/-. Représentatif de n = 9 souris.
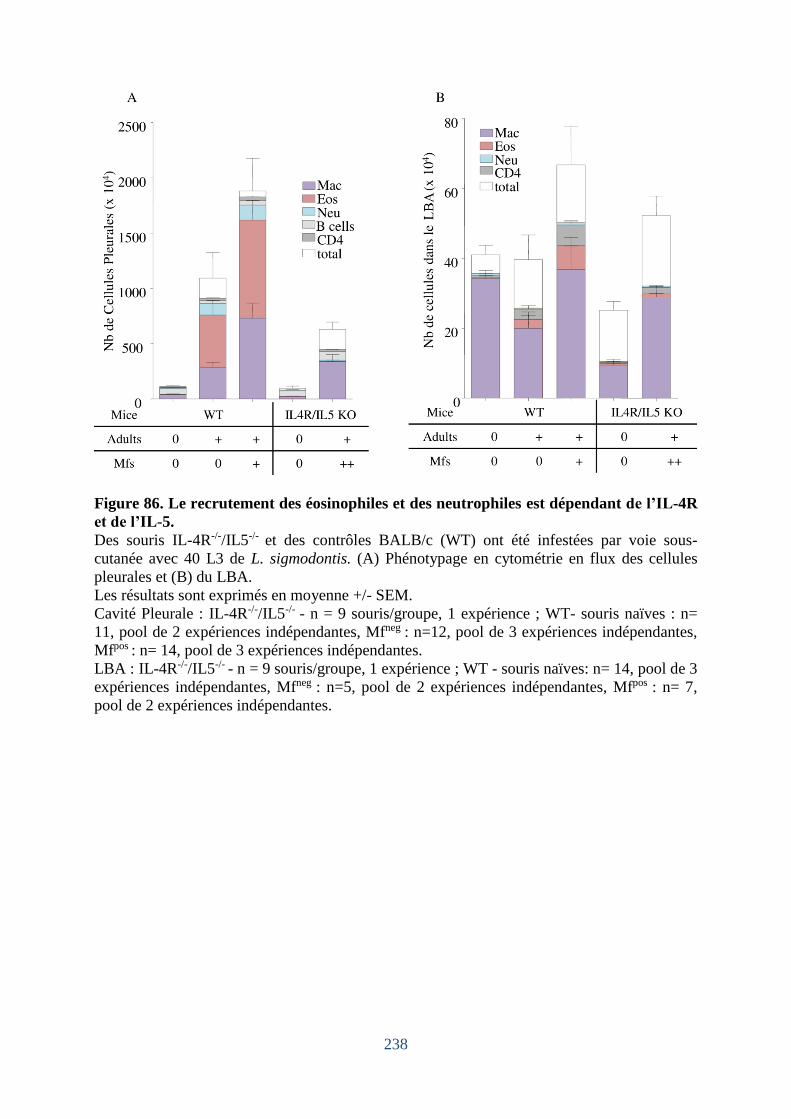
238
Figure 86. Le recrutement des éosinophiles et des neutrophiles est dépendant de l’IL-4R
et de l’IL-5.
Des souris IL-4R-/-/IL5-/- et des contrôles BALB/c (WT) ont été infestées par voie sous-
cutanée avec 40 L3 de L. sigmodontis. (A) Phénotypage en cytométrie en flux des cellules
pleurales et (B) du LBA.
Les résultats sont exprimés en moyenne +/- SEM.
Cavité Pleurale : IL-4R-/-/IL5-/- - n = 9 souris/groupe, 1 expérience ; WT- souris naïves : n=
11, pool de 2 expériences indépendantes, Mfneg : n=12, pool de 3 expériences indépendantes,
Mfpos : n= 14, pool de 3 expériences indépendantes.
LBA : IL-4R-/-/IL5-/- - n = 9 souris/groupe, 1 expérience ; WT - souris naïves: n= 14, pool de 3
expériences indépendantes, Mfneg : n=5, pool de 2 expériences indépendantes, Mfpos : n= 7,
pool de 2 expériences indépendantes.

239
Discussion
Chez les souris BALB/c, le rendement parasitaire est stable pendant environ 60-70
jours après infection [199]. La ponte des microfilaires commence vers J55 p.i. mais environ
40% des souris infestées ne présentent pas de microfilaires, malgré la présence d'œufs et
parfois de microfilaires dans les utérus des filaires femelles (Figure 82C). Les mécanismes
conduisant à l'absence de microfilaires chez les souris amicrofilarémiques (Mf-) ne sont pas
connus mais plusieurs hypothèses pourraient l'expliquer : la première est que certaines souris
pourraient n'être infestées qu'avec des filaires mâles ou femelles, ne permettant donc pas la
reproduction sexuée ; cependant, la quasi totalité des souris BALB/c infestées avec 40L3
présentent des parasites mâles et femelles à J30 p.i., après la dernière mue et donc au moment
de l’accouplement des filaires [274]. Cette hypothèse n'est donc pas suffisante. La deuxième
impliquerait un mauvais développement des embryons en microfilaires ce qui est possible au
vu des embryogrammes réalisés comparant le contenu utérin des filaires femelles issues soit
de souris Mfneg soit de souris Mfpos et montrant davantage de microfilaires utérines dans ces
dernières. La troisième hypothèse serait que les microfilaires soient détruites au fur et à
mesure de leur production mais aucun granulome réactionnel contenant des Mfs n’a été
observé. En fait, la fertilité accrue des filaires dans les souris immunodéficientes (Figure 82C)
conduit à privilégier l'hypothèse d'une baisse de la fertilité dépendant de facteurs
métaboliques et/ou hormonaux en interaction avec la réponse immunitaire. La quatrième
hypothèse impliquerait l’accumulation des microfilaires dans un ou des organes profonds tels
que le poumon, la rate ou le foie mais la quantification des microfilaires dans le poumon ne
montre pas de L1 dans ce tissu chez les souris amicrofilarémiques (Figure 83).
La réponse immunitaire de l'hôte joue en effet un rôle crucial dans l'établissement de la
phase patente comme le montre l'infection de différentes souris immunodéficientes de lignées
BALB/c ou C57BL/6. Dans les souris BALB/c IFNγ-/-, les filaires survivent plus longtemps et
la microfilérémie est plus forte [474]. Les souris C57BL/6, qui normalement éliminent les
parasites avant la phase patente, présentent une microfilarémie sanguine lorsqu'elles ont la
mutation Rag2IL-2Rγ-/- (entraînant une déficience en cellules T, B et NK) [286] ou lorsqu'elles
surexpriment l'IL-10 [462]. De même la mise en place de la phase patente serait déterminée par
des réponses dépendant du TLR-4. En effet, l'infection de souris BALB/c déficientes en TLR-
4 conduit à un plus grand pourcentage de souris microfilarémiques (Mf+), sans différence
dans le nombre de microfilaires circulantes [288]. Ceci pourrait être lié au S100A9 (voir

240
chapitre 2). Les souris BALB/c déficientes à la fois en IL-4R et en IL-5 (IL-4R-/-/IL5-/-)
utilisées dans cette étude présentent des microfilarémies très importantes. L'infection de ces
souris aboutit à 100% de souris Mf+, à une taille et une survie accrue des filaires ainsi qu'à
une fertilité exceptionnelle des femelles (Figures 81 et 82). Volkmann (2001 [475] et 2003 [291])
et Le Goff (2000 [253] et 2002 [476]) ont comparé les réponses de souris de fond génétique
BALB/c (permissif à l'infection) et C57BL/6 (résistant) déficientes soit en IL-5 (IL-5-/-) soit
en IL-4 (IL-4-/-) et/ou à son récepteur (IL-4-/-) au cours de l'infection par L. sigmodontis.
Premièrement les souris BALB/c IL-5-/- permettent le développement d'un plus grand
nombre de parasites adultes dans la cavité pleurale, ont une fécondité et une survie prolongée
des parasites (adultes et Mfs) et ont moins de granulomes pleuraux [291]. Les résultats sont
différents chez les souris C57BL/6 : le rendement parasitaire et la survie des parasites chez
des souris C57BL/6 IL-5-/- restent inchangés et les parasites sont éliminés avant la phase
patente [253]. Ces résultats apparemment contradictoires suggèrent que l'IL-5 n’est un facteur
limitant le développement des filaires que lorsque le parasite infeste un hôte permissif ; chez
un hôte résistant, le parasite serait capable d'éviter les mécanismes dépendants de l'IL-5 [476].
Deuxièmement, les souris BALB/c déficientes dans la signalisation de l'IL-4 (IL-4-/- et
IL-4R-/-) ne développent pas de différences dans la survie des adultes et la formation des
granulomes mais présentent des microfilarémies plus importantes que celles des souris
BALB/c WT [291,476]. Chez les souris C57BL/6 IL-4-/-, les parasites adultes survivent plus de
60 jours et présentent des microfilaires circulantes [476]. Ces résultats montrent que l'IL-5 est
important dans le contrôle de tous les stades parasitaires chez les souris BALB/c et que l'IL-4,
par le biais de son récepteur permet le contrôle des microfilaires chez toutes les souris mais
aussi celui de la survie des parasites chez les souris C57BL/6. Ainsi, les souris BALB/c IL-
4R-/-/IL-5-/- cumulent les deux effets entraînant le phénotype observé.
Les résultats obtenus lors de cette étude montrent une différence dans la réponse
immunitaire entre les souris Mfpos et Mfneg (augmentation des macrophages et des éosinophiles
pleuraux, Figure 79, augmentation de CXCL9 et CCL2 sériques et pleuraux, Figure 63). En
fait, la présence des microfilaires dans la cavité pleurale des souris Mfpos induit un
recrutement pleural plus important que la seule présence des filaires adultes (souris Mf-)
(Figure78A). Le recrutement de ces différentes populations cellulaires a déjà été décrit [202,263]
mais pas leur différentes proportions en fonction de la microfilarémie.

241
La formation de granulomes autour des filaires dans la cavité pleurale est un processus
bien décrit [202]. Cependant, ces granulomes peuvent adhérer à la surface des poumons
auxquels ils sont alors partiellement intégrés (Figure 61A). Des neutrophiles présents dans la
cavité pleurale participent à la formation de ces granulomes [202]. Leur recrutement peut être
dus à de multiples facteurs tels que la sécrétion de Wolbachia ou d'autres produits ES
chemoattractants pour les neutrophiles par les filaires femelles [14,362], ou l’exposition de
Wolbachia lors de la ponte, de la mort des microfilaires et de microfilaires avortées, et/ou de
la mort des adultes [17]. Les neutrophiles, mais aussi les éosinophiles, participent à la
formation des granulomes par un mécanisme dépendant de l'IL-5 [202,294], il est donc logique
que les souris IL-4R-/-/IL-5-/-ne présentent aucun neutrophile ni granulome.
La différence entre les souris Mfpos et Mfneg réside majoritairement dans les quantités
d'éosinophiles et de macrophages, qui sont nécessaires au contrôle des microfilaires [293,477].
Cette différence pourraient être associée à une augmentation de l'IL-13, de l'IL-5 et de l'IL-10
dans la cavité pleurale des souris microfilarémiques [288].
Les souris BALB/c ont éliminé tous les parasites adultes ainsi que la majorité des
granulomes et des microfilaires à J100 (Figure 60). A ce temps, les infiltrats cellulaires de la
cavité pleurale sont aussi résorbés (Figure 78A). Bien que dite « permissive », la résolution de
l’infection chez la souris BALB/c est très différente de celle observée dans le mérion, chez
qui l'infection peut durer plus d'un an. La persistance de l'infection pourrait être à l'origine des
différences de pathologies observées dans les deux modèles, à savoir que la présence des
nodules papillaires décrit chez le mérions n’existe pas chez la souris BALB/c.
La manifestation pulmonaire principale de l'infection à J30 [196] et chez les souris Mfneg
à J70 est une hypertrophie des cellules mésothéliales. Le mésothélium pleural réagit en effet
rapidement aux infections : lorsqu'elles sont activées par des signaux inflammatoires, les
cellules mésothéliales changent de forme, produisent de nombreuses cytokines et diminuent
l'expression des molécules d'adhésion [470]. Le mésothélium activé surexprime la podoplanine,
qui est connue pour induire une augmentation de la mobilité et de la prolifération des cellules
[471,478] et a été associée à la formation de métastases dans des cancers dont le mésothéliome
pleural [478,479]. Ces points pourraient expliquer le décollement du mésothélium parfois
observé (Annexe 2).

242
L'hypertrophie des cellules mésothéliales est associée chez les souris Mfpos à une
hyperplasie et une fibrose du mésothélium caractérisées par une accumulation de cellules et
de collagène dans le tissu sous-pleural (Figure 77). La fibrose se définit comme
l'accumulation excessive de composants de la matrice extracellulaire (ECM ; principalement
l'acide hyaluronique, la fibronectine, des protéoglycans et des collagènes) dans un tissu ou un
organe [480]. Les cellules présentes dans la zone épaissie du mésothélium n'expriment pas les
marqueurs de cellules mésothéliales (podoplanine et cytokératine, Figures 75 et 76) et
pourraient être des fibroblastes ou des myofibroblastes, qui sont les principales cellules
productrices de matrice extracellulaire. Des marquages spécifiques pour ces cellules, comme
l'actine des muscles lisses (α-SMA) pourraient permettre de confirmer leur nature. Les
myofibroblastes peuvent avoir différentes origines. Elles peuvent provenir de fibrocytes issus
de la moelle osseuse se différenciant en myofibroblastes [481]. Ces fibrocytes sont recrutés par
les chimokines CXCL12 et CCL2 [481-483], cette dernière étant fortement augmentée dans les
souris BALB/c infestées et Mf+. Elles peuvent également provenir de fibroblastes résidents
qui prolifèrent et se différencient en myofibroblastes sous l'action du TGF-β [484,485] ; ou, enfin
être issues d'une différenciation des cellules épithéliales alvéolaires ou bronchiques (pour les
voies aériennes) ou des cellules mésothéliales (pour la plèvre) [486]. Ce dernier mécanisme qui
s'appelle Transition Epithélo-Mésenchymateuse (EMT), dépend majoritairement du TGF-β et
de l'IL-13 [486-490] et impliquerait la podoplanine [490]. Les cellules envahissent alors
progressivement la lame basale en se redifférenciant (perte de l'expression de l'E-cadherine,
de la cytokératine, et de la polarité apico-basale et acquisition d'un phénotype de fibroblaste
avec l'expression de α-SMA ou FSP-1) [487,491,492]. Le fait que les souris IL-4R-/-/IL-5-/- ne
présentent pas d'hyperplasie ni de fibrose mésothéliale souligne le rôle de l'IL-13 dans la mise
en place de la pathologie pleurale chez les souris Mfpos (Figure 85). Il est probable que
l'accroissement du nombre de cellules, notamment des éosinophiles, observé chez les souris
Mfpos soit responsable d'une augmentation des cytokines pro-fibrotiques (IL-4, IL-13, TGF-β,
CCL2...) dans la cavité pleurale des souris, entraînant les phénomènes observés. Les dosages
de cytokines pro-fibrotiques dans la cavité pleurale des souris Mfpos et Mfneg permettra de
préciser leur implication et l'infection de souris déficientes en éosinophiles (Δdb/GATA-1)
permettra d'évaluer leur rôle.
Une fois pondues dans la cavité pleurale, les microfilaires rejoignent la circulation
sanguine. Leur voie de migration n'est pas connue. Cependant, deux options semblent
possibles. La plus simple est que les microfilaires s'échappent en utilisant les stomates

243
présents au niveau de la plèvre pariétale. Ces orifices de 8 à 10 µm abouchent directement
dans les capillaires lymphatiques sous-jacents [313]. De là, les microfilaires seraient drainées
par le flux lymphatique jusqu'à la circulation sanguine au niveau des veines sous-clavières.
L'autre possibilité est que les microfilaires traversent directement le mésothélium pleural pour
rejoindre le réseau sanguin pulmonaire. Les microfilaires circulent ensuite dans tout
l'organisme, portées par la circulation sanguine où elles peuvent être prises en charge par leur
vecteur au cours d'un repas sanguin. Une partie des microfilaires est retrouvée dans les
poumons (Figures 61, 66 et 83 et 84). S'il est attendu de les retrouver dans la circulation
sanguine (capillaires, veines et artères), des microfilaires ont aussi été retrouvées dans les
espaces périvasculaires. Dans les souris IL-4R-/-/IL-5-/- présentant des microfilarémies
sanguines extrêmement élevées, les microfilaires sont retrouvées en grande quantité dans les
poumons et sont observées dans tous les compartiments pulmonaires (alvéoles, circulation
sanguine, espaces périvasculaires : Figure 84). Il est probable que la distribution des
microfilaires soit identique chez les souris WT mais leur plus faible nombre pourrait expliquer
leur absence dans ces différents tissus, leur observation étant plus rare. Des microfilaires sont
aussi retrouvées dans l’espace bronchoalvéolaire des poumons de patients infectés par B.
malayi et W. bancrofti [493,494]. Pour rejoindre ces localisations, les microfilaires semblent
capables de traverser les endothéliums et de perturber les épithéliums alvéolaires (Figure 84).
Dans le cas des deux parasites humains, les Mfs sont capables d'adhérer à l'endothélium des
vaisseaux sanguins pulmonaires [50], ce qui pourrait permettre de résister au flux sanguin et
faciliter la traversée des endothéliums. En revanche, les Mfs de L. sigmodontis ne semblent
pas pouvoir faire de même [50]. Il est aussi possible d'imaginer que les Mfs, dans un contexte
inflammatoire favorisant la perméabilité vasculaire, puissent passer entre les cellules
endothéliales pour traverser les vaisseaux sanguins. Ce passage pourrait être facilité par des
molécules sécrétées par les microfilaires [362], ou par la libération de protéases (comme dans la
peau des individus infestés par O. volvulus [104,105]). Dans tous les cas, cette localisation
extravasculaire semble accidentelle car les microfilaires ne sont alors plus disponibles pour
une transmission au vecteur.
Les microfilaires, circulantes et pulmonaires, entraînent des réponses significativement
différentes dans les poumons des souris Mfpos comparés à ceux des souris Mf-. Ces réponses
partagent des caractéristiques observées lors du poumon tropical éosinophile dû aux
microfilaires de B. malayi et W. bancrofti : la présence d'éosinophiles broncho-alvéolaires,
des infiltrats périvasculaires de macrophages, la production de mucus et la fibrose pulmonaire

244
[78-80]. Si les symptômes chez L. sigmodontis paraissent moins sévères que lors du TPE, ces
résultats laissent penser que des mécanismes similaires sont en jeu et permettent de proposer
l'utilisation de l'infection chronique de L. sigmodontis chez des souris Mf+comme modèle de
TPE. Chez les souris Mf+, de nombreuses bronches contiennent des cellules caliciformes
produisant du mucus. Cette augmentation, n'est pas due à la prolifération de cellules
caliciformes préexistantes, mais plutôt à la transdifférenciation (métaplasie) des cellules
ciliées et Clara en cellules caliciformes [468]. La métaplasie des cellules caliciformes est
contrôlée par la fixation de l'IL-4 et l'IL-13 sur leur récepteur (IL-4R) à la surface des cellules
de l'épithélium bronchique [468,469]. Ceci explique pourquoi, chez les souris IL-4R-/-/IL-5-/-,
malgré la quantité importante de microfilaires, aucune inflammation de l'épithélium
bronchique n'est observée (Figure 86). Chez les souris WT Mf+, les cellules caliciformes sont
localisées sur la portion d'épithélium bronchique en contact avec l'espace périvasculaire
(PVS). Il est donc probable que l'IL-13 et l'IL-4 nécessaires à la métaplasie des cellules
caliciformes soient produites par les cellules du PVS. En effet, le contenu cellulaire du PVS
double chez les souris Mfpos et se compose pour moitié de macrophages interstitiels (Figure
70).
Ces macrophages interstitiels du PVS expriment le marqueur CD169 (aussi appelé
Siglec-1 ou Sialoadhesine) qui est aussi exprimé dans les poumons par les macrophages
alvéolaires et par des cellules sous-pleurales de la plèvre viscérale (Figures 67, 68, 69 et [318]).
Ces macrophages interstitiels CD169+ ont été observés dans une autre infection helminthique
avec Schistosoma mansoni et ces cellules augmentent précocement dans les poumons lors
d’une infection par les cercaires de ce parasite [334]. La fonction de ces macrophages CD169+
dans les poumons est cependant méconnue. Ces cellules auraient des capacités de
phagocytose et de prolifération permettant le renouvellement et l'augmentation des
populations sans recrutement de monocytes sanguins [318]. Ils auraient aussi un rôle important
dans le contrôle de l'inflammation : par exemple la suppression des cellules CD169+ chez des
souris CD169 DTR infestées par Plasmodium berghei entraîne une plus forte infection, des
dommages tissulaires et une inflammation importante de nombreux organes comme le foie, la
rate ou les poumons [495]. Lors de la rupture du schizonte, l'hémozoine, un produit de
détoxification du parasite, est normalement libérée dans la circulation et rapidement
phagocytée par les macrophages et les neutrophiles [496]. Dans ces souris CD169 DTR,
d’importants dépôts d'hémozoine sont encore présents dans les tissus [495]. La déficience en

245
CD169 de ces souris ne permet pas d’impliquer spécifiquement les macrophages alvéolaires
et/ou les macrophages interstitiels et/ou les cellules CD169+ de la plèvre.
De part leur localisation à proximité des vaisseaux sanguins, les macrophages
interstitiels présents dans le PVS pourraient avoir un rôle dans la prise en charge des
molécules exogènes circulant dans le sang. CD169 reconnait les acides sialiques [497] et la
gaine des Mfs de L. sigmodontis comporte de tels acides [498], il est donc possible que ces
cellules soient activées directement par les microfilaires ou leurs antigènes. Des macrophages
phénotypiquement proche des macrophages interstitiels pulmonaires CD169+ ont été décrits
dans d'autres organes, et pourraient avoir des fonctions similaires [318]. Dans les reins, les
macrophages périvasculaires CD169+ ont un rôle important dans le contrôle de l'inflammation
en diminuant l'expression de la molécule d'adhésion ICAM-1 des endothéliums et limitant
ainsi les afflux de neutrophiles dans le tissu [499]. Les macrophages interstitiels présents dans
le PVS participent probablement aux infiltrats cellulaires observés lors de toutes les phases de
l'infection (Figures 31, 53 et 70) notamment par leur production de cytokines. Ils pourraient
aussi participer au remodelage de la matrice extracellulaire nécessaire à l'infiltration des
leucocytes ([317] et chapitre 6.4 de l'introduction).
D'autres types cellulaires qui n'ont pas encore été analysés pourraient néanmoins être à
l'origine de la production d'IL-13 dans le PVS, notamment les ILC2 et les éosinophiles. En
effet, ces cellules sont présentes dans le PVS à l'homéostasie, en contact étroit avec la matrice
extracellulaire [365]. Dans l'infection par Nippostrongylus brasiliensis [345,365] ou dans des
modèles d'asthme [500-502], les ILC2 participent à la mise en place des réponses Th2
pulmonaires. Lorsqu'elles sont activées en réponse à la libération des alarmines IL-25, IL-33
et TSLP par les cellules épithéliales/endothéliales (en cas de dégât cellulaire ou de stimulation
par des allergènes), les ILC2 produisent de grandes quantités d'IL-4, d'IL-5 et d'IL-13
[501,503,504]. S'ensuit un recrutement et une survie accrue des éosinophiles entraînant une
accumulation de ces cellules dans les tissus et dans le liquide bronchoalvéolaire [322,323]. Dans
l'asthme, les éosinophiles produisent aussi de l'IL-4, de l'IL-5 et de l'IL-13 qui entretiennent
l'inflammation pulmonaire par des boucles autocrines [505-507].
Pour conclure, ces résultats remettent en question la qualification de Litomosoides
sigmodontis comme asymptomatique et soulignent le rôle des microfilaires et des réponses
Th2 dans l’aggravation de la pathologie pulmonaire (Figure 87).

246
Figure 87. Synthèse des résultats et hypothèses mécanistiques.
Double page suivante :
Dans la cavité pleurale (page de gauche), la présence des filaires adultes entraîne
l’hyperplasie du mésothélium et le recrutement d’éosinophiles, de neutrophiles et la
prolifération des macrophages, sous le contrôle de l’IL-4, de l’IL-5 et de l’IL-13. Des
granulomes composés de ces cellules se forment autour des parasites adultes. En cas de
présence de microfilaires (souris Mf+), les populations de macrophages et d’éosinophiles
s’accroissent, entraînant une augmentation de la sécrétion de cytokines pro-fibrotiques (IL-13,
CCL2, TGF-β). S’ensuit la fibrose de la plèvre viscérale du poumon. En cas d’absence d’IL-5
et de l’IL-4Rα, les recrutements d’éosinophiles et de neutrophiles ainsi que la fibrose sont
inhibés, et les parasites ont une survie et une fertilité accrue.
Dans les poumons (page de droite), l’infection filarienne chronique induit une accumulation
d’éosinophiles dans l’espace bronchoalvéolaire, qui est accrue chez les souris Mf+. Les
microfilaires peuvent rejoindre les espaces périvasculaires, dans lesquels une augmentation du
nombre de macrophages CD169+ est observée. La production d’IL-13 dans cet espace serait à
l’origine de la métaplasie des cellules caliciformes et de la production de mucus dans les voies
aériennes. Les réponses pulmonaires sont inhibées chez les souris IL-4R-/-/IL-5-/- qui
présentent pourtant de grandes quantités de microfilaires intrapulmonaires.
Eosino : éosinophile ; Neutro : neutrophile ; AAM : macrophage alternativement activé par
l'IL-4 ; Mf : microfilaire ; CAM : macrophage activé classiquement ; Fibro : fibroblaste. Colla
: collagène ; AM : macrophage alvéolaire ; IM CD169+ : macrophage interstitiel exprimant le
CD169.

247
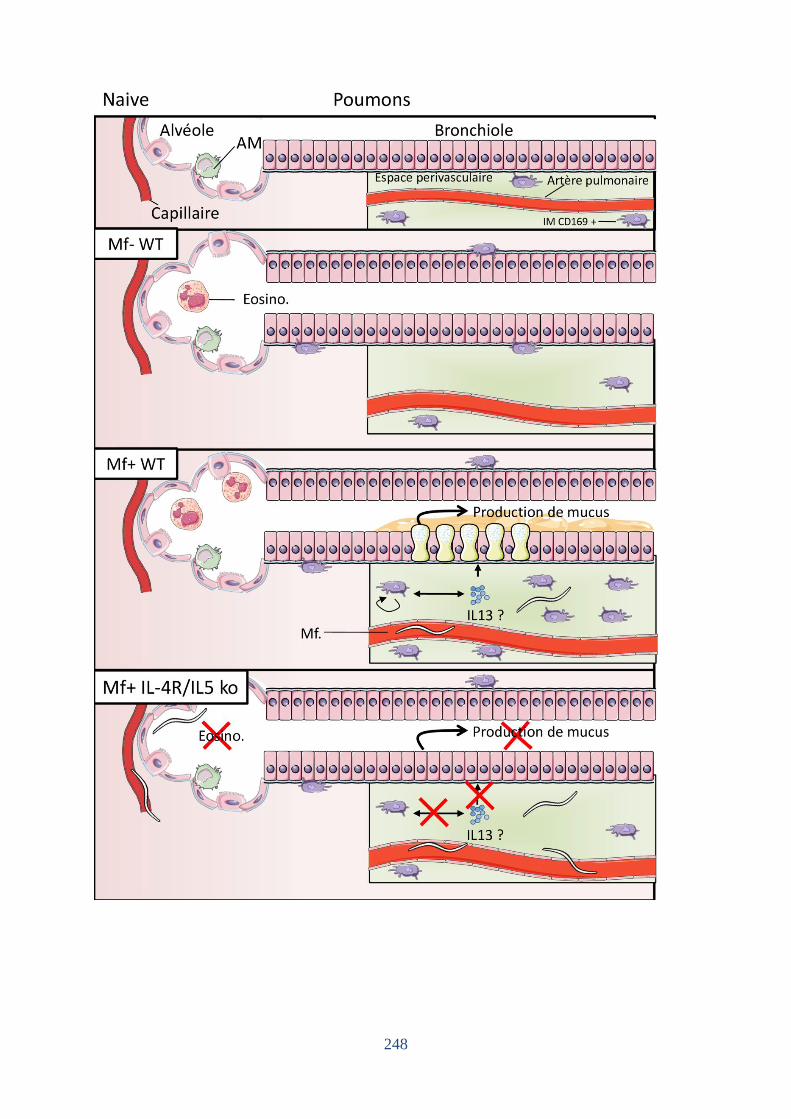
248

249

250
CONCLUSIONS ET PERSPECTIVES

251

252
Conclusions et perspectives
L’ensemble de cette étude a permis de mettre en évidence deux phases d’interaction de
la filaire Litomosoides sigmodontis avec l'environnement pulmonaire de ses hôtes rongeurs.
Premièrement lors de la phase de migration des L3, les résultats présentés démontrent que les
stades infestants filariens migrent dans le sang lorsque ceux-ci sont inoculés en intraveineuse
mais ont aussi une phase sanguine après leur migration lymphatique quand ils sont inoculés
dans la peau. De là, les L3 rejoignent la circulation sanguine pulmonaire et traversent les
poumons pour rejoindre la cavité pleurale. Ce passage induit tout d'abord des hémorragies
consécutives à la rupture des capillaires pulmonaires, accompagnées d'une augmentation du
nombre de neutrophiles pulmonaires et de la sécrétion transitoire d'IL-1β et des alarmines IL-
33 et S100A9 dans la cavité pleurale. Le S100A9 semble faciliter la survie des parasites, soit
par un effet anti-inflammatoire soit en facilitant la migration des L3. Dans les jours suivant
l'infection, une réponse régulatrice se met en place dans les poumons, avec le recrutement de
macrophages et d'éosinophiles, la production d'IL-4, de CCL2 et d'IL9, ainsi que la baisse
d'expression de molécules inflammatoires. Ces réponses visent probablement à réparer les
tissus lésés et conduiraient à la formation des granulomes observés dans le tissu pulmonaire.
Le passage des L3 induit néanmoins une inflammation durable des vaisseaux sanguins
pulmonaires.
Deuxièmement, lors de la phase patente de l'infection, les microfilaires, qui sont
présentes dans le sang de tous les mérions et dans environ 60% des souris BALB/c sont
responsables d’une l'inflammation pleurale forte. De plus, les souris développent une
pathologie pulmonaire dépendante des microfilaires consistant en : (1) la fibrose de la plèvre
viscérale, (2) une accumulation périvasculaire de macrophages et (3) une inflammation
bronchoalvéolaire (production de mucus et éosinophilie). Le contrôle des filaires (adultes et
microfilaires), mais aussi la mise en place de la pathologie sont dépendantes de l'IL-5 et de
l'IL-4R.
Ces résultats présentent donc les poumons comme une zone privilégiée d'interaction
hôte/parasite dans le modèle murin de filariose Litomosoides sigmodontis et ce lors des deux
phases - initiale et chronique - de l'infection. Cette interaction implique tous les
compartiments du poumon: le système sanguin (artères/veines/capillaires), les voies
respiratoires (alvéoles/bronches), les espaces périvasculaires (PVS), les plèvres et la cavité
pleurale. Jusqu'à présent, les réponses dans la cavité pleurale avaient été étudiées à de

253
nombreuses reprises, mais très peu les conséquences pulmonaires de l'infection filarienne,
jusque-là considérée comme asymptomatique.
Parmi les espaces pulmonaires affectés par l'infection, le PVS pourrait avoir un rôle
important dans la mise en place/régulation des réponses pulmonaires. En effet, des infiltrats
cellulaires y sont observés lors de la phase de migration, lors de la phase pré-patente et
lorsque les microfilaires circulent dans le sang. Cependant, la caractérisation de cet espace se
limite ici à quelques types cellulaires (neutrophiles lors de la phase de migration,
macrophages lors de la phase chronique). D'autres types cellulaires sont présents dans le PVS
des souris naïves et infestées mais n'ont pas encore été décrits. Au vu des réponses
pulmonaires (recrutements d'éosinophiles bronchoalvéolaires lors des différentes phases,
productions de cytokines de type Th2 et de mucus), il est probable que le contenu du PVS
reflète ces réponses.
Comment analyser l'espace périvasculaire ?
L'étude fonctionnelle des cellules de l'espace périvasculaire est complexe du fait de
leur localisation. L’analyse des poumons en microscopie confocale a permis d'obtenir des
informations nouvelles sur la localisation des cellules mais cette analyse est limitée à quelques
marqueurs. Une étude par cytométrie en flux sur les cellules des interstitium pulmonaires
(interstitium alvéolaire + périvasculaire + péribronchique) à différents temps après infection
pourrait être réalisée sur une suspension cellulaire obtenue par lyse du tissu préalablement
débarrassé des cellules bronchoalvéolaire (lavage bronchoalvéolaire) et de la circulation
sanguine (injection de PBS additionné d'héparine dans le ventricule droit du cœur). Une
analyse transcriptomique sur des cellules d’intérêt isolées de cette suspension cellulaire avec
un trieur cellulaire (comme réalisé par Gibbings et al 2017 [318]) permettrait ensuite de
préciser leurs fonctions lors des différentes phases de l'infection. Une autre possibilité, serait
d'isoler le PVS par microdissection laser sur des coupes épaisses et de réaliser une analyse
transcriptomique de cette zone. Des microdissections sur coupes épaisses de poumons ont
déjà été effectuées sur des coupes de 200µm d'épaisseur pour isoler le canal alvéolaire [508].
Ceci permettrait d'obtenir des informations sur les réseaux biologiques mis en place dans cette
zone au cours de l'infection.

254
Rôle des ILC2 dans les réponses anti-filariennes ?
Différentes études ont montré que la majorité des cellules produisant des cytokines de
types Th2 dans les poumons de souris naïves ou traitées par l'IL-33 sont les ILC2, et ces
cellules étaient responsables de maintenir et réguler les populations d'éosinophiles [365,509-511].
Contrairement aux lymphocytes T et B, les ILC ne possèdent pas de récepteurs spécifiques
aux antigènes (le TCR et le BCR respectivement). Elles ne requièrent donc pas de phase de
contact avec l'antigène ni de sélection clonale et répondent rapidement aux signaux
cytokiniques. Les ILC ont été classées en 3 groupes dont la diversité fonctionnelle est proche
de celle des lymphocytes T CD4-helper (Th) [512]. Les ILC2 sont stimulées par l'IL-25, l'IL-
33, le TSLP, le TL1A ou le PGD2 et sont capables de produire de l'IL-4, de l'IL-5, de l'IL-9 et
de l'IL-13. De par leur production de cytokines, elles sont donc prépondérantes dans
l'initiation des réponses de type Th2 [512].
La quantité d'ILC2 augmente précocement dans la cavité pleurale et dans les organes
lymphoïdes associés aux graisses (FALC) lors de l'infection par L. sigmodontis [273,281] mais
leur dynamique dans les poumons n'a pas encore été étudiée. Des études dans d'autres
modèles suggèrent qu'une partie de nos résultats pourraient être dus à cette population : tout
d'abord, dans les modèles d'asthme, les ILC2 pulmonaires, par leur production d'IL-13,
contribuent de manière importante à la métaplasie des cellules caliciformes et à la production
de mucus [513]. De plus, des souris traitées pendant 3 semaines avec de l'IL-33 présentent des
infiltrats périvasculaires d'ILC2 produisant de grandes quantités d'IL-5, ce qui entraîne des
accumulations d'éosinophiles et une obstruction des artères pulmonaires semblable à celle
observée dans le chapitre 2 [509]. Enfin, les ILC2 sont importantes dans l'activation alternative
des macrophages pulmonaires et le contrôle des parasites dans les poumons des souris
infestées par Nippostrongylus brasiliensis [345].

255
Les poumons dans le contrôle des parasites ?
Le système capillaire pulmonaire semble jouer un rôle crucial dans la migration des L3
et des microfilaires. Nous avons montré que les L3 migrent jusqu'à la cavité pleurale en
utilisant la circulation sanguine pulmonaire. Cependant, plus de 50% des filaires n'atteignent
pas la cavité pleurale malgré une injection en intraveineuse, alors que celle-ci les achemine
directement vers les capillaires pulmonaires. De plus, une accumulation de microfilaires dans
les poumons est souvent observée lors de l'infection de souris par des microfilaires [196,463,467]
et cette accumulation a été associée à la clairance des microfilaires par les poumons [196,461]
[504]. La disparition de la circulation sanguine de microfilaires inoculées prend des jours ou des
semaines en fonction du fond génétique des souris utilisées [196,461]. Chez les souris BALB/c,
les microfilaires circulent dans le sang pendant plusieurs semaines alors qu'elles disparaissent
en quelques heures chez les souris CBA/Ca ou les C57BL/6 [196]. Les microfilaires
s'accumulent alors dans les poumons et sont éliminées en moins de 4 jours [196]. Les
mécanismes permettant la séquestration et l'élimination des microfilaires dans les poumons
pourraient être analysés en comparant les réponses endothéliales et de l'espace périvasculaire
des différentes souches de souris. Dans le cas de la migration des L3 comme des microfilaires,
le piégeage des filaires dans les capillaires pulmonaires pourrait être modulé par des
traitements vasodilatateurs (papaïne et metformine) ou vasoconstricteurs (phenylephrine).
Enfin la localisation des L3 perdues lors de l'injection IV mais certainement aussi une
partie des L3 inoculées dans la peau (donc celles qui ne sont pas détruites dans la peau et qui
n’arrivent pas à la cavité pleurale) reste inconnue. Cependant le faible nombre inoculé, la
profondeur des tissus concernés, l’absence de filaires GFP, l’impossibilité d’utiliser un
anticorps de surface sur une longue période du fait des mues, rendent difficile une approche
de type tomographie en 3D. Une étude récente [514] a réussi à suivre les stades infestants de
Brugia pahangi pendant quelques jours en suivant la réponse immunitaire élicitée par les
filaires. Pour cela du luminol a été injecté dans les souris. Le luminol produit une lumière
bleue lorsqu'il est activé par la MPO des neutrophiles [515]. Dans cette étude, l'imagerie a
permis de détecter la réponse immunitaire provoquée dans la peau par 5 à 50 L3, 5 jours après
infection. L'utilisation de la lucigenine, qui nécessite une activité NADPH, caractéristique des
macrophages pourrait aussi être envisagée [516].

256

257
Valorisation des résultats
Publications :
Fercoq F, Remion E, Frohberger S, Vallarino N, Hoerauf A, Huebner M, Carlin L, Martin C.
Microfilaremia in Litomosoides sigmodontis infected BALB/c mice is associated with lung
pathology. Manuscrit en préparation.
Karadjian G*, Fercoq F*, Pionnier N, Vallarino-Lhermitte N, Lefoulon E, Nieguitsila A,
Specht S, Carlin L, Martin C. Migratory phase of Litomosoides sigmodontis filarial infective
larvae is associated with pathology and transient increase of S100A9 expressing neutrophils
in the lung. PLOS Neglected Tropical Diseases, 2017. * contribution égale.
Pionnier N, Brotin N, Karadjian G, Hemon P, Gaudin-Nomé F, Vallarino-Lhermitte N,
Nieguitsila A, Fercoq F, Aknin M, Marin-Esteban V, Chollet-Martin S, Schlecht-Louf G,
Bachelerie F, Martin C. Neutropenic mice provide insight into the role of skin-Infiltrating
Neutrophils in the host protective Immunity against Filarial Infective Larvae. PLOS
Neglected Tropical Diseases, 2016.
Congrès:
Presentations orales:
Fercoq F, Karadjian G, Pionnier N, Vallarino-Lhermitte N, Lefoulon E, Nieguitsila A, Specht
S, Carlin L, Martin C. New insights into filarial biology : What Litomosoides sigmodontis
model has taught us about larval migration into lymphatics and lungs? 26th International
Conference of the World Association for the Advancement of Veterinary Parasitology.
Kuala Lumpur 4-8 septembre 2017.
Fercoq F, Karadjian G, Pionnier N, Vallarino-Lhermitte N, Specht S, Carlin L, Martin C.
Filarial infection and pulmonary inflammation in the Litomosoides sigmodontis murine model
of filariasis. EMOP XII – The European Multicolloquium of Parasitology, Turku,
Finlande. 20-24 Juillet 2016.
Karadjian G, Fercoq F, Pionnier N, Vallarino-Lhermitte N, Specht S, Carlin L, Martin C.
Migration des stades infestant de la filaire murine Litomosoides sigmodontiset réponse
inflammatoire associée chez la souris BALB/c. Congrès de la Société Française de
Parasitologie. Grenoble. 23-24 Mars 2016.

258
Posters:
Karadjian G*, Fercoq F*, Pionnier N, Vallarino-Lhermitte N, Specht S, Carlin L, Martin C.
Litomosoides sigmodontis filarial infective larvae migrate through the lungs increasing
S100A9 in neutrophils. Molecular and Cellular Biology of Helminth Parasites IX. Hydra,
Grèce. 31 Aout– 5 September 2015.
Karadjian G*, Fercoq F*, Vallarino-Lhermitte N, Specht S, Carlin L, Kilarski W, Swartz M,
Martin C. Unravelling the filarial larvae migrations : lessons from Litomosoides
sigmodontis mouse model. Molecular and Cellular Biology of Helminth Parasites IX.
Hydra, Grèce. 31 Aout– 5 September 2015.

259

260
ANNEXES

261
Annexe 1. Liste des gènes pour l’étude des profils des transcrits pulmonaires à J4 et J70.
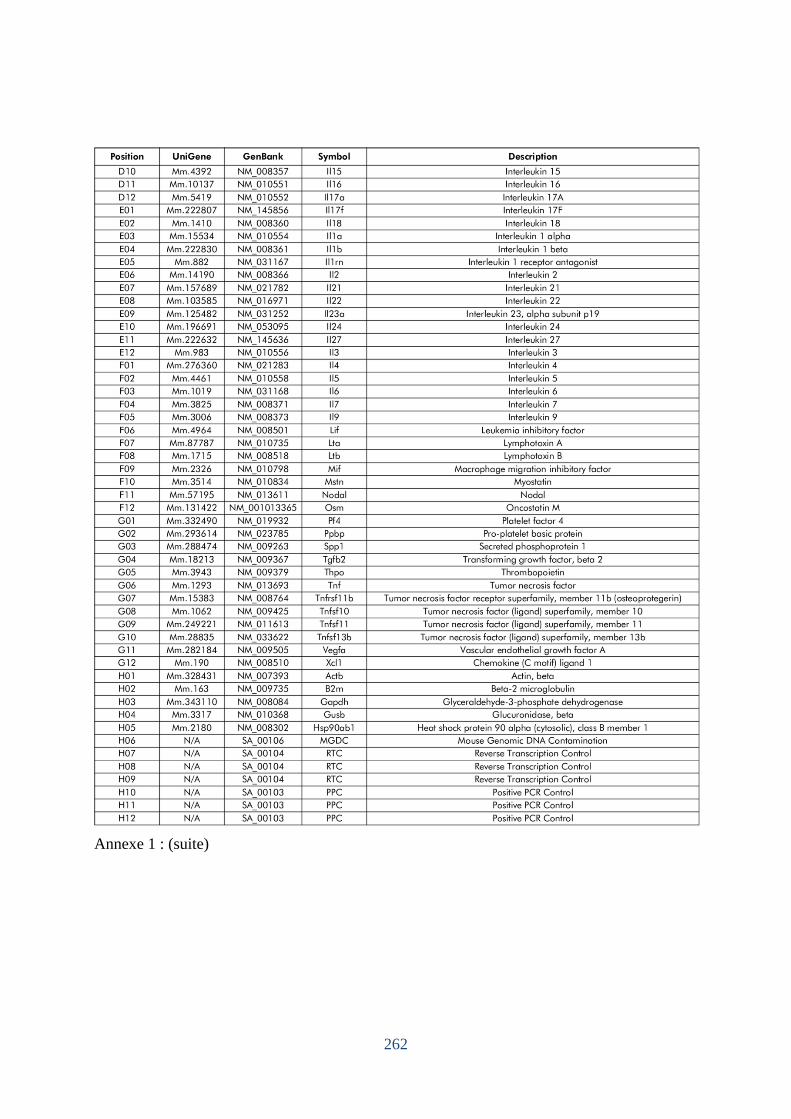
262
Annexe 1 : (suite)
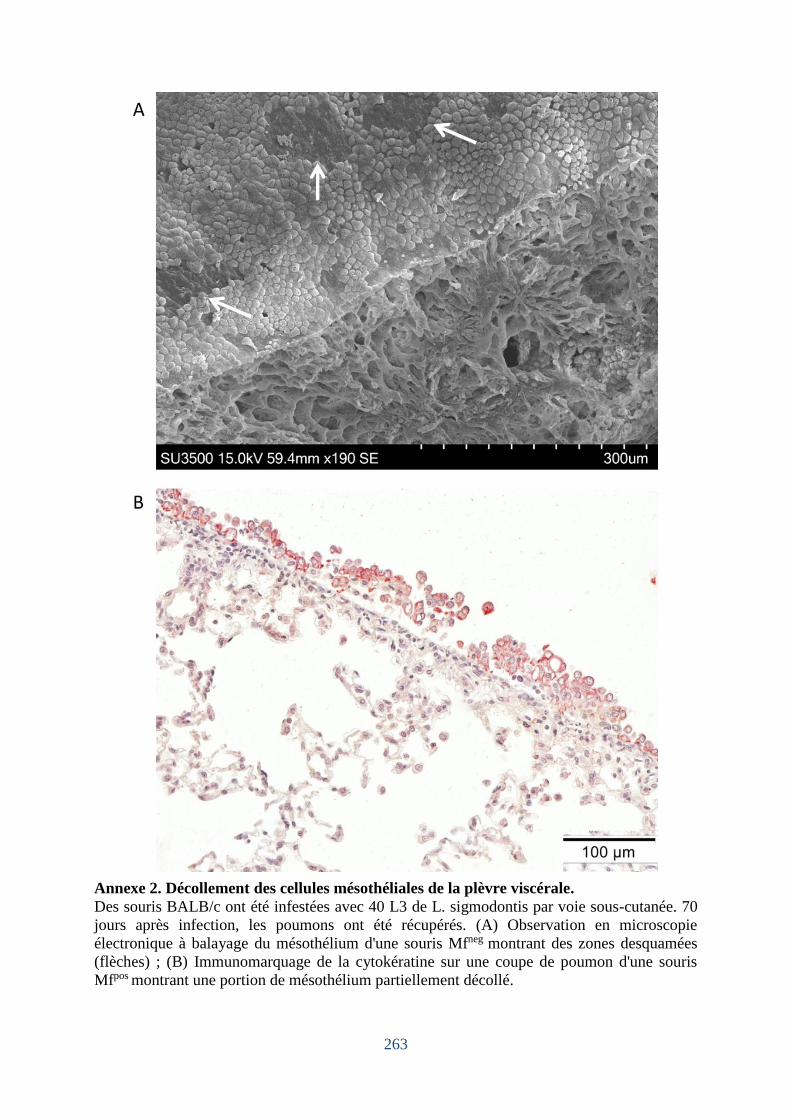
263
Annexe 2. Décollement des cellules mésothéliales de la plèvre viscérale.
Des souris BALB/c ont été infestées avec 40 L3 de L. sigmodontis par voie sous-cutanée. 70
jours après infection, les poumons ont été récupérés. (A) Observation en microscopie
électronique à balayage du mésothélium d'une souris Mfneg montrant des zones desquamées
(flèches) ; (B) Immunomarquage de la cytokératine sur une coupe de poumon d'une souris
Mfpos montrant une portion de mésothélium partiellement décollé.

264

265
BIBLIOGRAPHIE

266
Bibliographie
1. Bain O, Mutafchiev Y, Junker K (2014) Order Spirurida. In: Schmidt-Rhaesa A, editor.
Handbook of Zoology: De Gruyter.
2. Lefoulon E, Bain O, Bourret J, Junker K, Guerrero R, Canizales I, Kuzmin Y, Satoto TB,
Cardenas-Callirgos JM, de Souza Lima S, Raccurt C, Mutafchiev Y, Gavotte L,
Martin C (2015) Shaking the Tree: Multi-locus Sequence Typing Usurps Current
Onchocercid (Filarial Nematode) Phylogeny. PLoS Negl Trop Dis 9: e0004233.
3. Bain O, Babayan S, Gomes J, Rojas G, Guerrero R (2002) First account on the larval
biology of a Litomosoides filaria, from a bat. Parassitologia 44: 89-92.
4. Anderson RC (2000) Nematode parasites of vertebrates : their development and
transmission. Wallingford, Oxon, UK ; New York, NY: CABI Pub. xx, 650 p. p.
5. Uni S, Mat Udin AS, Agatsuma T, Saijuntha W, Junker K, Ramli R, Omar H, Lim YA,
Sivanandam S, Lefoulon E, Martin C, Belabut DM, Kasim S, Abdullah Halim MR,
Zainuri NA, Bhassu S, Fukuda M, Matsubayashi M, Harada M, Low VL, Chen CD,
Suganuma N, Hashim R, Takaoka H, Azirun MS (2017) Morphological and molecular
characteristics of Malayfilaria sofiani Uni, Mat Udin & Takaoka n. g., n. sp.
(Nematoda: Filarioidea) from the common treeshrew Tupaia glis Diard & Duvaucel
(Mammalia: Scandentia) in Peninsular Malaysia. Parasit Vectors 10: 194.
6. Lee DL (2002) The biology of nematodes. London.
7. Chitwood BG, Chitwood MB (1950) An introduction to nematology. Section I. Anatomy.
Washington, D.C.
8. Bain O, Chabaud AG (1975) [The mechanism controlling the crossing of the vector's
stomach wall by microfilariae (Dipetalonema dessetae-Aedes aegypti)]. C R Acad Sci
Hebd Seances Acad Sci D 281: 1199-1202.
9. Nieguitsila A, Frutos R, Moulia C, Lhermitte-Vallarino N, Bain O, Gavotte L, Martin C
(2013) Fitness cost of Litomosoides sigmodontis filarial infection in mite vectors;
implications of infected haematophagous arthropod excretory products in host-vector
interactions. Biomed Res Int 2013: 584105.
10. Bain O, Wanji S, Vuong PN, Marechal P, Le Goff L, Petit G (1994) Larval biology of six
filariae of the sub-family Onchocercinae in a vertebrate host. Parasite 1: 241-254.
11. Wenk P (1967) [The invasion route of the metacyclical larvae of Litomosoides carinii
Chandler 1931 (Filariidae)]. Z Parasitenkd 28: 240-263.
12. Lefoulon E, Bain O, Makepeace BL, d'Haese C, Uni S, Martin C, Gavotte L (2016)
Breakdown of coevolution between symbiotic bacteria Wolbachia and their filarial
hosts. PeerJ 4: e1840.
13. Lo N, Casiraghi M, Salati E, Bazzocchi C, Bandi C (2002) How many wolbachia
supergroups exist? Mol Biol Evol 19: 341-346.
14. Landmann F, Foster JM, Slatko B, Sullivan W (2010) Asymmetric Wolbachia segregation
during early Brugia malayi embryogenesis determines its distribution in adult host
tissues. PLoS Negl Trop Dis 4: e758.
15. Fischer K, Beatty WL, Jiang D, Weil GJ, Fischer PU (2011) Tissue and stage-specific
distribution of Wolbachia in Brugia malayi. PLoS Negl Trop Dis 5: e1174.
16. Landmann F, Bain O, Martin C, Uni S, Taylor MJ, Sullivan W (2012) Both asymmetric
mitotic segregation and cell-to-cell invasion are required for stable germline
transmission of Wolbachia in filarial nematodes. Biol Open 1: 536-547.
17. Bouchery T, Lefoulon E, Karadjian G, Nieguitsila A, Martin C (2013) The symbiotic role
of Wolbachia in Onchocercidae and its impact on filariasis. Clin Microbiol Infect 19:
131-140.

267
18. Taylor MJ, Hoerauf A, Bockarie M (2010) Lymphatic filariasis and onchocerciasis.
Lancet 376: 1175-1185.
19. Landmann F, Voronin D, Sullivan W, Taylor MJ (2011) Anti-filarial activity of antibiotic
therapy is due to extensive apoptosis after Wolbachia depletion from filarial
nematodes. Plos Pathogens 7: e1002351.
20. Landmann F, Foster JM, Michalski ML, Slatko BE, Sullivan W (2014) Co-evolution
between an endosymbiont and its nematode host: Wolbachia asymmetric posterior
localization and AP polarity establishment. PLoS Negl Trop Dis 8: e3096.
21. Huebner MP, Layland LE, Hoerauf A (2014) Lymphatic and Tissue Filariasis. In: Bruschi
F, editor. Helminth Infections and their Impact on Global Public Health. Wien:
Springer-Verlag. pp. 367-409.
22. Simonsen PE, Onapa AW, Asio SM (2011) Mansonella perstans filariasis in Africa. Acta
Trop 120 Suppl 1: S109-120.
23. Zoure HG, Wanji S, Noma M, Amazigo UV, Diggle PJ, Tekle AH, Remme JH (2011)
The geographic distribution of Loa loa in Africa: results of large-scale implementation
of the Rapid Assessment Procedure for Loiasis (RAPLOA). PLoS Negl Trop Dis 5:
e1210.
24. Lima NF, Veggiani Aybar CA, Dantur Juri MJ, Ferreira MU (2016) Mansonella ozzardi: a
neglected New World filarial nematode. Pathog Glob Health 110: 97-107.
25. (2016) Global programme to eliminate lymphatic filariasis: progress report, 2015. Wkly
Epidemiol Rec 91: 441-455.
26. Ramaiah KD, Ottesen EA (2014) Progress and impact of 13 years of the global
programme to eliminate lymphatic filariasis on reducing the burden of filarial disease.
PLoS Negl Trop Dis 8: e3319.
27. (2016) Progress report on the elimination of human onchocerciasis, 2015-2016. Wkly
Epidemiol Rec 91: 505-514.
28. (2016) Global, regional, and national incidence, prevalence, and years lived with disability
for 310 diseases and injuries, 1990-2015: a systematic analysis for the Global Burden
of Disease Study 2015. Lancet 388: 1545-1602.
29. Basanez MG, Pion SD, Churcher TS, Breitling LP, Little MP, Boussinesq M (2006) River
blindness: a success story under threat? PLoS Med 3: e371.
30. Cheke RA (2017) Factors affecting onchocerciasis transmission: lessons for infection
control. Expert Rev Anti Infect Ther 15: 377-386.
31. Metzger WG, Mordmuller B (2014) Loa loa-does it deserve to be neglected? Lancet Infect
Dis 14: 353-357.
32. Diggle PJ, Thomson MC, Christensen OF, Rowlingson B, Obsomer V, Gardon J, Wanji S,
Takougang I, Enyong P, Kamgno J, Remme JH, Boussinesq M, Molyneux DH (2007)
Spatial modelling and the prediction of Loa loa risk: decision making under
uncertainty. Ann Trop Med Parasitol 101: 499-509.
33. Molyneux DH, Hopkins A, Bradley MH, Kelly-Hope LA (2014) Multidimensional
complexities of filariasis control in an era of large-scale mass drug administration
programmes: a can of worms. Parasit Vectors 7: 363.
34. Akue JP, Nkoghe D, Padilla C, Moussavou G, Moukana H, Mbou RA, Ollomo B, Leroy
EM (2011) Epidemiology of concomitant infection due to Loa loa and Mansonella
perstans in Gabon. PLoS Negl Trop Dis 5: e1329.
35. Pion SD, Clarke P, Filipe JA, Kamgno J, Gardon J, Basanez MG, Boussinesq M (2006)
Co-infection with Onchocerca volvulus and Loa loa microfilariae in central
Cameroon: are these two species interacting? Parasitology 132: 843-854.
36. Ferri E, Bain O, Barbuto M, Martin C, Lo N, Uni S, Landmann F, Baccei SG, Guerrero R,
de Souza Lima S, Bandi C, Wanji S, Diagne M, Casiraghi M (2011) New insights into

268
the evolution of Wolbachia infections in filarial nematodes inferred from a large range
of screened species. PLoS One 6: e20843.
37. Keiser PB, Coulibaly Y, Kubofcik J, Diallo AA, Klion AD, Traore SF, Nutman TB (2008)
Molecular identification of Wolbachia from the filarial nematode Mansonella perstans.
Mol Biochem Parasitol 160: 123-128.
38. Gehringer C, Kreidenweiss A, Flamen A, Antony JS, Grobusch MP, Belard S (2014)
Molecular evidence of Wolbachia endosymbiosis in Mansonella perstans in Gabon,
Central Africa. J Infect Dis 210: 1633-1638.
39. Bain O, Mutafchiev Y, Junker K, Guerrero R, Martin C, Lefoulon E, Uni S (2015) Review
of the genus Mansonella Faust, 1929 sensu lato (Nematoda: Onchocercidae), with
descriptions of a new subgenus and a new subspecies. Zootaxa 3918: 151-193.
40. Wanji S, Tayong DB, Layland LE, Datchoua Poutcheu FR, Ndongmo WP, Kengne-Ouafo
JA, Ritter M, Amvongo-Adjia N, Fombad FF, Njeshi CN, Nkwescheu AS, Enyong
PA, Hoerauf A (2016) Update on the distribution of Mansonella perstans in the
southern part of Cameroon: influence of ecological factors and mass drug
administration with ivermectin. Parasit Vectors 9: 311.
41. Debrah LB, Nausch N, Opoku VS, Owusu W, Mubarik Y, Berko DA, Wanji S, Layland
LE, Hoerauf A, Jacobsen M, Debrah AY, Phillips RO (2017) Epidemiology of
Mansonella perstans in the middle belt of Ghana. Parasit Vectors 10: 15.
42. Manguin S, Bangs MJ, Pothikasikorn J, Chareonviriyaphap T (2010) Review on global
co-transmission of human Plasmodium species and Wuchereria bancrofti by
Anopheles mosquitoes. Infect Genet Evol 10: 159-177.
43. Chakraborty S, Manokaran G, Zawieja D, Muthuchamy M (2013) Lymphatic Filariasis:
Perspectives on Lymphatic Remodeling and Contractile Dysfunction in Filarial
Disease Pathogenesis. Microcirculation 20: 349-364.
44. Shenoy RK, Suma TK, Kumaraswami V, Padma S, Rahmah N, Abhilash G, Ramesh C
(2007) Doppler ultrasonography reveals adult-worm nests in the lymph vessels of
children with brugian filariasis. Ann Trop Med Parasitol 101: 173-180.
45. Sucharit S, Harinasuta C, Choochote W (1982) Experimental transmission of subperiodic
Wuchereria bancrofti to the leaf monkey (Presbytis melalophos), and its periodicity.
Am J Trop Med Hyg 31: 599-601.
46. Harinasuta C, Sucharit S, Choochote W (1981) The susceptibility of leaf monkeys to
bancroftian filariasis in Thailand. Southeast Asian J Trop Med Public Health 12: 581-
589.
47. Palmieri JR, Connor DH, Purnomo, Marwoto HA (1983) Bancroftian filariasis.
Wuchereria bancrofti infection in the silvered leaf monkey (Presbytis cristatus). Am J
Pathol 112: 383-386.
48. Khan AM, Dutta P, Das S, Pathak AK, Sarmah P, Hussain ME, Mahanta J (2015)
Microfilarial periodicity of Wuchereria bancrofti in Assam, Northeast India. J Vector
Borne Dis 52: 208-212.
49. Thurston JP (1951) The periodicity of microfilariae. I. The distribution of microfilariae in
the body. Trans R Soc Trop Med Hyg 45: 307-328.
50. Schroeder JH, McCarthy D, Szestak T, Cook DA, Taylor MJ, Craig AG, Lawson C,
Lawrence RA (2017) Brugia malayi microfilariae adhere to human vascular
endothelial cells in a C3-dependent manner. PLoS Negl Trop Dis 11: e0005592.
51. Post RJ, Onyenwe E, Somiari SA, Mafuyai HB, Crainey JL, Ubachukwu PO (2011) A
guide to the Simulium damnosum complex (Diptera: Simuliidae) in Nigeria, with a
cytotaxonomic key for the identification of the sibling species. Ann Trop Med
Parasitol 105: 277-297.

269
52. Plaisier AP, van Oortmarssen GJ, Remme J, Habbema JD (1991) The reproductive
lifespan of Onchocerca volvulus in West African savanna. Acta Trop 48: 271-284.
53. Attout T, Hoerauf A, Denece G, Debrah AY, Marfo-Debrekyei Y, Boussinesq M, Wanji
S, Martinez V, Mand S, Adjei O, Bain O, Specht S, Martin C (2009) Lymphatic
vascularisation and involvement of Lyve-1+ macrophages in the human onchocerca
nodule. PLoS One 4: e8234.
54. Vuong PN, Bain O, Cabaret J, Petit G, Prod'hon J, Ranque P, Chabaud AG (1988) Forest
and savanna onchocerciasis: comparative morphometric histopathology of skin
lesions. Trop Med Parasitol 39: 105-110.
55. Allen JE, Adjei O, Bain O, Hoerauf A, Hoffmann WH, Makepeace BL, Schulz-Key H,
Tanya VN, Trees AJ, Wanji S, Taylor DW (2008) Of mice, cattle, and humans: the
immunology and treatment of river blindness. PLoS Negl Trop Dis 2: e217.
56. Banla M, Tchalim S, Karabou PK, Gantin RG, Agba AI, Kere-Banla A, Helling-Giese G,
Heuschkel C, Schulz-Key H, Soboslay PT (2014) Sustainable control of
onchocerciasis: ocular pathology in onchocerciasis patients treated annually with
ivermectin for 23 years: a cohort study. PLoS One 9: e98411.
57. Tamarozzi F, Halliday A, Gentil K, Hoerauf A, Pearlman E, Taylor MJ (2011)
Onchocerciasis: the role of Wolbachia bacterial endosymbionts in parasite biology,
disease pathogenesis, and treatment. Clin Microbiol Rev 24: 459-468.
58. Bain O, Wanji S, Enyong P, Petit G, Noireau F, Eberhard MI, Wahl G (1998) New
features on the moults and morphogenesis of the human filaria Loa loa by using rodent
hosts consequences. Parasite 5: 37-46.
59. Eveland LK, Yermakov V, Kenney M (1975) Loa loa infection without microfilaraemia.
Trans R Soc Trop Med Hyg 69: 354-355.
60. Orihel TC, Eberhard ML (1985) Loa loa: development and course of patency in
experimentally-infected primates. Trop Med Parasitol 36: 215-224.
61. Hawking F, Moore P, Gammage K, Worms MJ (1967) Periodicity of microfilariae. XII.
The effect of variations in host body temperature on the cycle of Loa loa,
Monnigofilaria setariosa, Dirofilria immitis and other filariae. Trans R Soc Trop Med
Hyg 61: 674-683.
62. Boussinesq M (2006) Loiasis. Ann Trop Med Parasitol 100: 715-731.
63. Peel E, Chardome M (1946) Sur des Filaridés des Chimpanzés Pan paniscus et Pan satyrus
au Congo Belge. Ann Soc Belg Med Trop: 117–156.
64. Habermann RT, Menges RW (1968) Filariasis (Acanthocheilonema perstans) in a gorilla.
Vet Med Small Anim Clin 63: 1040-1043.
65. Duke BO (1954) The uptake of the microfilariae of Acanthocheilonema streptocerca by
Culicoides grahamii, and their subsequent development. Ann Trop Med Parasitol 48:
416-420.
66. Cerqueira ML (1959) Sôbre a transmissao da Mansonella ozzardi. Journal Brasileiro de
Medicina: 900–914.
67. Orihel TC, Lowrie RC, Jr., Eberhard ML, Raccurt C, Kozek WJ, Tidwell MA, Tidwell M
(1981) Susceptibility of laboratory primates to infection with Mansonella ozzardi from
man. Am J Trop Med Hyg 30: 790-794.
68. Manson P (1891) The filaria Sanguinis hominis major and minor, two new species of
hæmatozoa. The Lancet 137: 4-8.
69. Asio SM, Simonsen PE, Onapa AW (2009) Mansonella perstans filariasis in Uganda:
patterns of microfilaraemia and clinical manifestations in two endemic communities.
Trans R Soc Trop Med Hyg 103: 266-273.
70. Dreyer G, Noroes J, Figueredo-Silva J, Piessens WF (2000) Pathogenesis of lymphatic
disease in bancroftian filariasis: a clinical perspective. Parasitol Today 16: 544-548.

270
71. Cahill KM, Kaiser RL (1964) Lymphangiography in Bancroftian Filariasis. Trans R Soc
Trop Med Hyg 58: 356-362.
72. Shenoy RK (2008) Clinical and pathological aspects of filarial lymphedema and its
management. Korean J Parasitol 46: 119-125.
73. Pfarr KM, Debrah AY, Specht S, Hoerauf A (2009) Filariasis and lymphoedema. Parasite
Immunol 31: 664-672.
74. Freedman DO, Bui T, De Almeida Filho PJ, Braga C, Maia e Silva MC, Maciel A,
Furtado AF (1995) Lymphoscintigraphic assessment of the effect of
diethylcarbamazine treatment on lymphatic damage in human bancroftian filariasis.
Am J Trop Med Hyg 52: 258-261.
75. Debrah AY, Mand S, Toliat MR, Marfo-Debrekyei Y, Batsa L, Nurnberg P, Lawson B,
Adjei O, Hoerauf A, Pfarr K (2007) Plasma vascular endothelial growth Factor-A
(VEGF-A) and VEGF-A gene polymorphism are associated with hydrocele
development in lymphatic filariasis. Am J Trop Med Hyg 77: 601-608.
76. Noroes J, Addiss D, Cedenho A, Figueredo-Silva J, Lima G, Dreyer G (2003)
Pathogenesis of filarial hydrocele: risk associated with intrascrotal nodules caused by
death of adult Wuchereria bancrofti. Trans R Soc Trop Med Hyg 97: 561-566.
77. Babu S, Nutman TB (2012) Immunopathogenesis of lymphatic filarial disease. Seminars
in Immunopathology 34: 847-861.
78. Udwadia FE, Joshi VV (1964) A Study of Tropical Eosinophilia. Thorax 19: 548-554.
79. Nutman TB, Vijayan VK, Pinkston P, Kumaraswami V, Steel C, Crystal RG, Ottesen EA
(1989) Tropical pulmonary eosinophilia: analysis of antifilarial antibody localized to
the lung. J Infect Dis 160: 1042-1050.
80. Vijayan VK (2007) Tropical pulmonary eosinophilia: pathogenesis, diagnosis and
management. Curr Opin Pulm Med 13: 428-433.
81. Webb JK, Job CK, Gault EW (1960) Tropical eosinophilia: demonstration of microfilariae
in lung, liver, and lymphnodes. Lancet 1: 835-842.
82. Sharma P, Sharma A, Vishwakarma AL, Agnihotri PK, Sharma S, Srivastava M (2016)
Host lung immunity is severely compromised during tropical pulmonary eosinophilia:
role of lung eosinophils and macrophages. J Leukoc Biol 99: 619-628.
83. Hall LR, Mehlotra RK, Higgins AW, Haxhiu MA, Pearlman E (1998) An essential role
for interleukin-5 and eosinophils in helminth-induced airway hyperresponsiveness.
Infect Immun 66: 4425-4430.
84. Mehlotra RK, Hall LR, Higgins AW, Dreshaj IA, Haxhiu MA, Kazura JW, Pearlman E
(1998) Interleukin-12 suppresses filaria-induced pulmonary eosinophilia, deposition of
major basic protein and airway hyperresponsiveness. Parasite Immunol 20: 455-462.
85. Mehlotra RK, Hall LR, Haxhiu MA, Pearlman E (2001) Reciprocal immunomodulatory
effects of gamma interferon and interleukin-4 on filaria-induced airway
hyperresponsiveness. Infect Immun 69: 1463-1468.
86. Lobos E, Ondo A, Ottesen EA, Nutman TB (1992) Biochemical and immunologic
characterization of a major IgE-inducing filarial antigen of Brugia malayi and
implications for the pathogenesis of tropical pulmonary eosinophilia. J Immunol 149:
3029-3034.
87. Lobos E, Zahn R, Weiss N, Nutman TB (1996) A major allergen of lymphatic filarial
nematodes is a parasite homolog of the gamma-glutamyl transpeptidase. Mol Med 2:
712-724.
88. Gounni AS, Spanel-Borowski K, Palacios M, Heusser C, Moncada S, Lobos E (2001)
Pulmonary inflammation induced by a recombinant Brugia malayi gamma-glutamyl
transpeptidase homolog: involvement of humoral autoimmune responses. Mol Med 7:
344-354.

271
89. Lobos E, Nutman TB, Hothersall JS, Moncada S (2003) Elevated immunoglobulin E
against recombinant Brugia malayi gamma-glutamyl transpeptidase in patients with
bancroftian filariasis: association with tropical pulmonary eosinophilia or putative
immunity. Infect Immun 71: 747-753.
90. Collins RC, Lujan R, Figueroa H, Campbell CC (1982) Early formation of the nodule in
Guatemalan onchocerciasis. Am J Trop Med Hyg 31: 267-269.
91. Parkhouse RM, Bofill M, Gomez-Priego A, Janossy G (1985) Human macrophages and
T-lymphocyte subsets infiltrating nodules of Onchocerca volvulus. Clin Exp Immunol
62: 13-18.
92. Wildenburg G, Korten S, Buttner DW (1998) Mast cell distribution in nodules of
Onchocerca volvulus from untreated patients with generalized onchocerciasis.
Parasitology 116 ( Pt 3): 257-268.
93. Brattig NW, Buttner DW, Hoerauf A (2001) Neutrophil accumulation around Onchocerca
worms and chemotaxis of neutrophils are dependent on Wolbachia endobacteria.
Microbes Infect 3: 439-446.
94. Simonsen PE, Hoerauf A, Fischer A, Weil G (2013) The Filariases. In: Farrar J, editor.
Manson's Tropical Diseases. 23rd Edition ed: Elsevier. pp. pp.737-765.
95. Buttner DW, von Laer G, Mannweiler E, Buttner M (1982) Clinical, parasitological and
serological studies on onchocerciasis in the Yemen Arab Republic. Tropenmed
Parasitol 33: 201-212.
96. Kershaw WE, Duke BO, Budden FH (1954) Distribution of microfilariae of O. volvulus
in the skin; its relation to the skin changes and to eye lesions and blindness. Br Med J
2: 724-729.
97. Duke BO (1993) The population dynamics of Onchocerca volvulus in the human host.
Trop Med Parasitol 44: 61-68.
98. Crainey JL, Medeiros JF, Costa Pessoa FA, Bessa Luz SL (2017
) Onchocerciasis. In: Marcondes CB, editor. Arthropod Borne Diseases: Springer
International Publishing. pp. pp 383-403.
99. Brattig NW (2004) Pathogenesis and host responses in human onchocerciasis: impact of
Onchocerca filariae and Wolbachia endobacteria. Microbes Infect 6: 113-128.
100. Timmann C, Abraha RS, Hamelmann C, Buttner DW, Lepping B, Marfo Y, Brattig N,
Horstmann RD (2003) Cutaneous pathology in onchocerciasis associated with
pronounced systemic T-helper 2-type responses to Onchocerca volvulus. Br J
Dermatol 149: 782-787.
101. Ali MM, Baraka OZ, AbdelRahman SI, Sulaiman SM, Williams JF, Homeida MM,
Mackenzie CD (2003) Immune responses directed against microfilariae correlate with
severity of clinical onchodermatitis and treatment history. J Infect Dis 187: 714-717.
102. Korten S, Hoerauf A, Kaifi JT, Buttner DW (2011) Low levels of transforming growth
factor-beta (TGF-beta) and reduced suppression of Th2-mediated inflammation in
hyperreactive human onchocerciasis. Parasitology 138: 35-45.
103. Satoguina J, Mempel M, Larbi J, Badusche M, Loliger C, Adjei O, Gachelin G, Fleischer
B, Hoerauf A (2002) Antigen-specific T regulatory-1 cells are associated with
immunosuppression in a chronic helminth infection (onchocerciasis). Microbes Infect
4: 1291-1300.
104. Petralanda I, Piessens WF (1994) Pathogenesis of onchocercal dermatitis: possible role
of parasite proteases and autoantibodies to extracellular matrix proteins. Exp Parasitol
79: 177-186.
105. Haffner A, Guilavogui AZ, Tischendorf FW, Brattig NW (1998) Onchocerca volvulus:
microfilariae secrete elastinolytic and males nonelastinolytic matrix-degrading serine
and metalloproteases. Exp Parasitol 90: 26-33.

272
106. Katawa G, Layland LE, Debrah AY, von Horn C, Batsa L, Kwarteng A, Arriens S, D
WT, Specht S, Hoerauf A, Adjobimey T (2015) Hyperreactive onchocerciasis is
characterized by a combination of Th17-Th2 immune responses and reduced
regulatory T cells. PLoS Negl Trop Dis 9: e3414.
107. Rubio de Kromer MT, Medina-De la Garza CE, Brattig NW (1995) Differences in
eosinophil and neutrophil chemotactic responses in sowda and generalized form of
onchocerciasis. Acta Trop 60: 21-33.
108. Hoerauf A, Pfarr K, Mand S, Debrah AY, Specht S (2011) Filariasis in Africa--treatment
challenges and prospects. Clin Microbiol Infect 17: 977-985.
109. Gillette-Ferguson I, Daehnel K, Hise AG, Sun Y, Carlson E, Diaconu E, McGarry HF,
Taylor MJ, Pearlman E (2007) Toll-like receptor 2 regulates CXC chemokine
production and neutrophil recruitment to the cornea in Onchocerca
volvulus/Wolbachia-induced keratitis. Infect Immun 75: 5908-5915.
110. Gentil K, Pearlman E (2009) Gamma interferon and interleukin-1 receptor 1 regulate
neutrophil recruitment to the corneal stroma in a murine model of Onchocerca
volvulus keratitis. Infect Immun 77: 1606-1612.
111. Abiose A (1998) Onchocercal eye disease and the impact of Mectizan treatment. Ann
Trop Med Parasitol 92 Suppl 1: S11-22.
112. Saint Andre A, Blackwell NM, Hall LR, Hoerauf A, Brattig NW, Volkmann L, Taylor
MJ, Ford L, Hise AG, Lass JH, Diaconu E, Pearlman E (2002) The role of
endosymbiotic Wolbachia bacteria in the pathogenesis of river blindness. Science 295:
1892-1895.
113. Pearlman E, Gillette-Ferguson I (2007) Onchocerca volvulus, Wolbachia and river
blindness. Chem Immunol Allergy 92: 254-265.
114. Hall LR, Diaconu E, Patel R, Pearlman E (2001) CXC chemokine receptor 2 but not C-C
chemokine receptor 1 expression is essential for neutrophil recruitment to the cornea
in helminth-mediated keratitis (river blindness). J Immunol 166: 4035-4041.
115. Pion S, Chesnais C (2017) Loiasis. In: Marcondes CB, editor. Arthropod Borne
Diseases: Springer International Publishing.
116. Buttner DW, Wanji S, Bazzocchi C, Bain O, Fischer P (2003) Obligatory symbiotic
Wolbachia endobacteria are absent from Loa loa. Filaria J 2: 10.
117. Penafiel-Freire DM, Herranz-Aguirre M (2017) Loiasis, a Subconjunctival
Manifestation. J Pediatr.
118. Varenne F, Fillaux J, Porterie M, Soler J, Cassagne M, Soler V (2016) [Subconjunctival
loiasis: A case report]. J Fr Ophtalmol 39: e193-e194.
119. Mandal D, Roy D, Bera DK, Manna B (2013) Occurrence of gravid Loa loa in
subconjunctival space of man: a case report from West Bengal, India. J Parasit Dis 37:
52-55.
120. Gobbi F, Postiglione C, Angheben A, Marocco S, Monteiro G, Buonfrate D, Mascarello
M, Gobbo M, Boussinesq M, Bisoffi Z (2014) Imported loiasis in Italy: an analysis of
100 cases. Travel Med Infect Dis 12: 713-717.
121. Herrick JA, Metenou S, Makiya MA, Taylar-Williams CA, Law MA, Klion AD, Nutman
TB (2015) Eosinophil-associated processes underlie differences in clinical
presentation of loiasis between temporary residents and those indigenous to Loa-
endemic areas. Clin Infect Dis 60: 55-63.
122. Ali S, Fisher M, Juckett G (2008) The African eye worm: a case report and review. J
Travel Med 15: 50-52.
123. Nutman TB, Miller KD, Mulligan M, Ottesen EA (1986) Loa loa infection in temporary
residents of endemic regions: recognition of a hyperresponsive syndrome with
characteristic clinical manifestations. J Infect Dis 154: 10-18.

273
124. Nzolo D, Anto F, Hailemariam S, Bakajika D, Muteba D, Makenga JC, Mesia G, Nsibu
C, Mampunza S, Tona G (2017) Central and Peripheral Nervous System Disorders
Following Ivermectin Mass Administration: A Descriptive Study Based on the
Democratic Republic of Congo Pharmacovigilance System. Drugs Real World
Outcomes.
125. Bhalla D, Dumas M, Preux PM (2013) Neurological manifestations of filarial infections.
Handb Clin Neurol 114: 235-242.
126. Kamgno J, Boussinesq M, Labrousse F, Nkegoum B, Thylefors BI, Mackenzie CD
(2008) Encephalopathy after ivermectin treatment in a patient infected with Loa loa
and Plasmodium spp. Am J Trop Med Hyg 78: 546-551.
127. Boussinesq M, Gardon J, Gardon-Wendel N, Chippaux JP (2003) Clinical picture,
epidemiology and outcome of Loa-associated serious adverse events related to mass
ivermectin treatment of onchocerciasis in Cameroon. Filaria J 2 Suppl 1: S4.
128. Mackenzie C, Geary T, Prichard R, Boussinesq M (2007) Where next with Loa loa
encephalopathy? Data are badly needed. Trends Parasitol 23: 237-238.
129. Cully DF, Vassilatis DK, Liu KK, Paress PS, Van der Ploeg LH, Schaeffer JM, Arena JP
(1994) Cloning of an avermectin-sensitive glutamate-gated chloride channel from
Caenorhabditis elegans. Nature 371: 707-711.
130. Ducorps M, Gardon-Wendel N, Ranque S, Ndong W, Boussinesq M, Gardon J,
Schneider D, Chippaux JP (1995) [Secondary effects of the treatment of
hypermicrofilaremic loiasis using ivermectin]. Bull Soc Pathol Exot 88: 105-112.
131. Mackerras M (1962) Filarial parasites (Nematoda : Filarioidea) of Australian animals.
Australian Journal of Zoology 10: 400-457.
132. Bhaskara Menon T, Ramamurti B, Sundarasiva Rao D (1944) Lizard filariasis. an
experimental study. Trans R Soc Trop Med Hyg 37: 373-386.
133. Sevimli FK, Kozan E, Bulbul A, Birdane FM, Kose M, Sevimli A (2007) Dirofilaria
immitis infection in dogs: unusually located and unusual findings. Parasitol Res 101:
1487-1494.
134. Simon F, Siles-Lucas M, Morchon R, Gonzalez-Miguel J, Mellado I, Carreton E,
Montoya-Alonso JA (2012) Human and animal dirofilariasis: the emergence of a
zoonotic mosaic. Clin Microbiol Rev 25: 507-544.
135. Sonin MD (1968) Filariata of animals and man and diseases caused by them. Filarioidea.
Moskow: Nauka Publishers.
136. Sonin MD (1968) Filariata of animals and man and diseases caused by them.
Diplotriaenoidea. Moskow: Nauka Publishers.
137. Measures LN, Gosselin J-F, Bergeron E (1996) Heartworm, Acanthocheilonema
spirocauda (Leidy, 1858), infections in Canadian phocid seals. Canadian Journal of
Fisheries and Aquatic Sciences 54: 842-846.
138. Bain O, Guerrero R (2003) Bisbalia vossi n. g., n. sp. (Nematoda: Onchocercidae), a
filarial worm from a geomyoid rodent, Heteromys anomalus, in Venezuela. Syst
Parasitol 54: 145-151.
139. Ash LR, Riley JM (1970) Development of Brugia pahangi in the jird, Meriones
unguiculatus, with notes on infections in other rodents. J Parasitol 56: 962-968.
140. Vincent AL, Frommes SP, Ash LR (1976) Brugia malayi, Brugia pahangi, and Brugia
patei: pulmonary pathology in jirds, Meriones unguiculatus. Exp Parasitol 40: 330-
354.
141. Dissanaike AS, Paramananthan DC (1961) On Brugia (Brugiella subgen. nov.) buckelyi
n.sp., from the heart and blood vessels of the Ceylon hare. J Helminthol 35: 209-220.

274
142. Anderson RC, Freeman RS (1969) Cardiofilaria inornata (Anderson, 1956) from
Woodcock with a Review of Cardiofilaria and Related Genera (Nematoda:
Filarioidea). Transactions of the American Microscopical Society 88: 68-79.
143. Sazmand A, Tafti MH, Hekmatimoghaddam S, Moobedi I (2013) Dipetalonema evansi
infection in camels of Iran's central area. Pak J Biol Sci 16: 647-650.
144. Bain O, Petit G, Berteaux S (1980) [Description of two new species and infective stages
of filariae from the genus Litomosoides (author's transl)]. Ann Parasitol Hum Comp
55: 225-237.
145. Aggarwal J, Kapila K, Gaur A, Wali JP (1993) Bancroftian filarial pleural effusion.
Postgrad Med J 69: 869-870.
146. Anderson RC (1961) Study of two filarioid nematodes,Chandlerella chitwoodae n.sp.
From Padda oryzivora (L.) and Protofilaria furcata Chandler, 1929. Canadian Journal
of Zoology 39: 317-323.
147. Sonin MD (1966) Filariata of animals and man and diseases caused by them. Aproctidea.
Moskow: Nauka Publisher.
148. Mullin SW, Balasingam S (1973) Dunnifilaria ramachandrani gen. n., sp. n. (Nematoda:
Filarioidea) from the Long-Tailed Giant Rat (Rattus sabanus) in Malaysia. The
Helminthological Society of Washington 40: 47-49.
149. Pletcher JM, Boomker J, de Vos V, Gardiner CH (1989) Lesions in the Heart and Lungs
of Greater Kudu (Tragelaphus strepsiceros) Caused by Cordophilus sagittus
(Nematoda: Filarioidea). Journal of Zoo and Wildlife Medicine 20: 465-470.
150. Bartlett C, Anderson RC (1986) Paronchocerca struthionus n.sp. (Nematoda: Filarioidea)
from ostriches (Struthio camelus), with a redescription of Paronchocerca ciconiarum
Peters, 1936 and a review of the genus. Canadian Journal of Zoology 64: 2480-2491.
151. Bartlett C, Anderson RC (1955) Ornithofilaria algonquiensis n. sp. From Hirundo
erythrogaster with a revision of the genera Paramicipsella Chow, 1939 Emend.
Chabaud and Choquet, 1953 and Ornithofilaria Gönnert, 1937. Canadian Journal of
Zoology 33: 107-112.
152. Hibler CP (1964) New Species of Onchocercidae (Nematoda: Filarioidea) from Pica Pica
Hudsonia (Sabine, 1823). J Parasitol 50: 667-674.
153. Bartlett C, Anderson RC (1985) On the filarioid nematodes (Splendidofilaria spp.) from
the pulmonary arteries of birds. Canadian Journal of Zoology 63: 2373-2377.
154. Sonin MD (1977) Filariata of animals and man and diseases caused by them. Filarioidea.
Moskow: Nauka Publishers.
155. Dantas-Torres F, Otranto D (2013) Dirofilariosis in the Americas: a more virulent
Dirofilaria immitis? Parasit Vectors 6: 288.
156. Cancrini G, Romi R, Gabrielli S, Toma L, M DIP, Scaramozzino P (2003) First finding
of Dirofilaria repens in a natural population of Aedes albopictus. Med Vet Entomol
17: 448-451.
157. Otranto D, Dantas-Torres F, Brianti E, Traversa D, Petric D, Genchi C, Capelli G (2013)
Vector-borne helminths of dogs and humans in Europe. Parasit Vectors 6: 16.
158. Haro A, Tamiya S, Nagashima A (2016) A rare case of human pulmonary dirofilariasis
with a growing pulmonary nodule after migrating infiltration shadows, mimicking
primary lung carcinoma. Int J Surg Case Rep 22: 8-11.
159. Kume S, Itagaki S (1955) On the life-cycle of Dirofilaria immitis in the dog as the final
host. British Veterinary Journal: 16-24.
160. Orihel TC (1961) Morphology of the larval stages of Dirofilaria immitis in the dog. J
Parasitol 47: 251-262.
161. Kotani T, Powers KG (1982) Developmental stages of Dirofilaria immitis in the dog. Am
J Vet Res 43: 2199-2206.

275
162. Supakorndej P, McCall JW, Jun JJ (1994) Early migration and development of
Dirofilaria immitis in the ferret, Mustela putorius furo. J Parasitol 80: 237-244.
163. Hayasaki M (1996) Re-migration of fifth-stage juvenile Dirofilaria immitis into
pulmonary arteries after subcutaneous transplantation in dogs, cats, and rabbits. J
Parasitol 82: 835-837.
164. Wanji S, Cabaret J, Gantier JC, Bonnand N, Bain O (1990) The fate of the filaria
Monanema martini in two rodent hosts: recovery rate, migration, and localization. Ann
Parasitol Hum Comp 65: 80-88.
165. Bain O, Wanji S, Vuong PN, Petit G, Breton B, Gantier JC (1994) Cardio-pulmonary
location of lymphatic filariae. Lymphology 27: 385-388.
166. Bowman DD, Atkins CE (2009) Heartworm biology, treatment, and control. Vet Clin
North Am Small Anim Pract 39: 1127-1158, vii.
167. Suresh K, Shimoda LA (2016) Lung Circulation. Compr Physiol 6: 897-943.
168. Munnell JF, Weldon JS, Lewis RE, Thrall DE, McCall JW (1980) Intimal lesions of the
pulmonary artery in dogs with experimental dirofilariasis. Am J Vet Res 41: 1108-
1112.
169. Schaub RG, Rawlings CA, Keith JC, Jr. (1981) Platelet adhesion and myointimal
proliferation in canine pulmonary arteries. Am J Pathol 104: 13-22.
170. Gonzalez-Miguel J, Morchon R, Carreton E, Montoya-Alonso JA, Simon F (2013)
Surface associated antigens of Dirofilaria immitis adult worms activate the host
fibrinolytic system. Vet Parasitol 196: 235-240.
171. Carreton E, Gonzalez-Miguel J, Montoya-Alonso JA, Morchon R, Simon F, Passeri B,
Cantoni AM, Kramer L (2013) D-dimer deposits in lungs and kidneys suggest its use
as a marker in the clinical workup of dogs with heartworm (Dirofilaria immitis)
disease. Vet Parasitol 191: 182-186.
172. Venco L, Mihaylova L, Boon JA (2014) Right Pulmonary Artery Distensibility Index
(RPAD Index). A field study of an echocardiographic method to detect early
development of pulmonary hypertension and its severity even in the absence of
regurgitant jets for Doppler evaluation in heartworm-infected dogs. Vet Parasitol 206:
60-66.
173. Muro A, Genchi C, Cordero M, Simon F (1999) Human dirofilariasis in the European
Union. Parasitol Today 15: 386-389.
174. Stone M, Dalal I, Stone C, Dalal B (2015) 18-FDG Uptake in Pulmonary Dirofilariasis. J
Radiol Case Rep 9: 28-33.
175. Araya J, Kawabata Y, Tomichi N, Kaneko K, Hayashi K, Iwabuchi K, Terasaki Y,
Kawashima T, Watanabe M (2007) Allergic inflammatory reaction is involved in
necrosis of human pulmonary dirofilariasis. Histopathology 51: 484-490.
176. Karpathiou G, Batistatou A, Steiropoulos P, Stefanou D, Froudarakis ME (2016) An
Unexpected Pulmonary Coin Lesion. Int J Surg Pathol 24: 328-329.
177. Villeneuve A (2014) La dirofilariose canine propositions de traitement et de prévention
appropriées au québec. Faculté de médecine vétérinaire Sainte-Hyacinthe.
178. Hirano Y, Kitagawa H, Sasaki Y (1992) Relationship between pulmonary arterial
pressure and pulmonary thromboembolism associated with dead worms in canine
heartworm disease. J Vet Med Sci 54: 897-904.
179. Bregani ER, Tantardini F, Rovellini A (2007) [Mansonella perstans filariasis].
Parassitologia 49.
180. Fux CA, Chappuis B, Holzer B, Aebi C, Bordmann G, Marti H, Hatz C (2006)
Mansonella perstans causing symptomatic hypereosinophilia in a missionary family.
Travel Med Infect Dis 4: 275-280.

276
181. Kahn JB (1983) Pleural effusion associated with Dipetalonema perstans
(Acanthocheilonema perstans). J Infect Dis 147: 166.
182. Kabego L, Kasengi JB, Mirindi P, Ruhanya V, Lupande D, Bulabula A, Ngoma P (2016)
Pulmonary localization of Mansonella perstans in a 16 months-old male patient in a
tertiary care hospital in Bukavu, Democratic Republic of Congo. Germs 6: 151-154.
183. Asgeirsson H, Harling A, Botero-Kleiven S (2017) Successful treatment of 2 imported
cases of Mansonella perstans infection. PLoS Negl Trop Dis 11: e0005452.
184. Mutafchiev Y, Bain O, Williams Z, McCall JW, Michalski ML (2014) Intraperitoneal
development of the filarial nematode Brugia malayi in the Mongolian jird (Meriones
unguiculatus). Parasitol Res 113: 1827-1835.
185. McSorley HJ, Harcus YM, Murray J, Taylor MD, Maizels RM (2008) Expansion of
Foxp3+ regulatory T cells in mice infected with the filarial parasite Brugia malayi. J
Immunol 181: 6456-6466.
186. Rajan TV, Ganley L, Paciorkowski N, Spencer L, Klei TR, Shultz LD (2002) Brugian
infections in the peritoneal cavities of laboratory mice: kinetics of infection and
cellular responses. Exp Parasitol 100: 235-247.
187. Grove DI, Davis RS, Warren KS (1979) Brugia malayi microfilaraemia in mice: a model
for the study of the host response to microfilariae. Parasitology 79: 303-316.
188. Tanaka M (1986) Clearance of inoculated microfilariae of Brugia malayi by monoclonal
antibodies in BALB/c mice. Jpn J Exp Med 56: 169-175.
189. Wanji S, Tendongfor N, Vuong PN, Enyong P, Bain O (2002) The migration and
localization of Loa loa infective and fourth-stage larvae in normal and
immunosuppressed rodents. Ann Trop Med Parasitol 96: 823-830.
190. Tendongfor N, Wanji S, Ngwa JC, Esum ME, Specht S, Enyong P, Matthaei KI, Hoerauf
A (2012) The human parasite Loa loa in cytokine and cytokine receptor gene knock
out BALB/c mice: survival, development and localization. Parasit Vectors 5: 43.
191. Lange AM, Yutanawiboonchai W, Scott P, Abraham D (1994) IL-4- and IL-5-dependent
protective immunity to Onchocerca volvulus infective larvae in BALB/cBYJ mice. J
Immunol 153: 205-211.
192. Johnson EH, Schynder-Candrian S, Rajan TV, Nelson FK, Lustigman S, Abraham D
(1998) Immune responses to third stage larvae of Onchocerca volvulus in interferon-
gamma and interleukin-4 knockout mice. Parasite Immunol 20: 319-324.
193. Bianco AE, Mustafa MB, Ham PJ (1989) Fate of developing larvae of Onchocerca
lienalis and O. volvulus in micropore chambers implanted into laboratory hosts. J
Helminthol 63: 218-226.
194. Petit G, Diagne M, Marechal P, Owen D, Taylor D, Bain O (1992) Maturation of the
filaria Litomosoides sigmodontis in BALB/c mice; comparative susceptibility of nine
other inbred strains. Ann Parasitol Hum Comp 67: 144-150.
195. Babayan S, Ungeheuer MN, Martin C, Attout T, Belnoue E, Snounou G, Renia L,
Korenaga M, Bain O (2003) Resistance and susceptibility to filarial infection with
Litomosoides sigmodontis are associated with early differences in parasite
development and in localized immune reactions. Infect Immun 71: 6820-6829.
196. Bouchery T, Denece G, Attout T, Ehrhardt K, Lhermitte-Vallarino N, Hachet-Haas M,
Galzi JL, Brotin E, Bachelerie F, Gavotte L, Moulia C, Bain O, Martin C (2012) The
chemokine CXCL12 is essential for the clearance of the filaria Litomosoides
sigmodontis in resistant mice. PLoS One 7: e34971.
197. Brant SV, Gardner SL (2000) Phylogeny of species of the genus Litomosoides
(Nematatoda : Onchocercidae): Evidence of rampant host switching. Journal of
Parasitology 86: 545-554.

277
198. Diagne M, Petit G, Liot P, Cabaret J, Bain O (1990) The filaria Litomosoides galizai in
mites; microfilarial distribution in the host and regulation of the transmission. Ann
Parasitol Hum Comp 65: 193-199.
199. Bain O, Attout T, Babayan S, Kozek W, Martin C (2004) Biology of Filariae in the
Vertebrate Host: Litomosoides sigmodontis Casts Light on Several “Grey Areas”. IX
European Multicolloquium of Parasitology – EMOP. Valencia, Spain: Medimond.
200. Bertram DS (1966) Dynamics of parasitic equilibrium in cotton rat filariasis. Adv
Parasitol 4: 255-319.
201. Babayan SA, Read AF, Lawrence RA, Bain O, Allen JE (2010) Filarial parasites develop
faster and reproduce earlier in response to host immune effectors that determine
filarial life expectancy. PLoS Biol 8: e1000525.
202. Attout T, Martin C, Babayan SA, Kozek WJ, Bazzocchi C, Oudet F, Gallagher IJ, Specht
S, Bain O (2008) Pleural cellular reaction to the filarial infection Litomosoides
sigmodontis is determined by the moulting process, the worm alteration, and the host
strain. Parasitol Int 57: 201-211.
203. Hoffmann W, Petit G, Schulz-Key H, Taylor D, Bain O, Le Goff L (2000) Litomosoides
sigmodontis in mice: reappraisal of an old model for filarial research. Parasitol Today
16: 387-389.
204. Marechal P, Le Goff L, Petit G, Diagne M, Taylor DW, Bain O (1996) The fate of the
filaria Litomosoides sigmodontis in susceptible and naturally resistant mice. Parasite
3: 25-31.
205. Karadjian G, Berrebi D, Dogna N, Vallarino-Lhermitte N, Bain O, Landau I, Martin C
(2014) Co-infection restrains Litomosoides sigmodontis filarial load and plasmodial P.
yoelii but not P. chabaudi parasitaemia in mice. Parasite 21: 16.
206. Graham AL, Taylor MD, Le Goff L, Lamb TJ, Magennis M, Allen JE (2005)
Quantitative appraisal of murine filariasis confirms host strain differences but reveals
that BALB/c females are more susceptible than males to Litomosoides sigmodontis.
Microbes Infect 7: 612-618.
207. Wharton DR (1947) Pathological changes in natural and experimental filariasis in the
cotton rat. J Infect Dis 80: 307-318.
208. Schneider CR, Blair LS, Schardein JL, Boche LK, Thompson PE (1968) Comparison of
early Litomosoides carinii infections in cotton rats and gerbils. J Parasitol 54.
209. Rebollo MP, Bockarie MJ (2017) Can Lymphatic Filariasis Be Eliminated by 2020?
Trends Parasitol 33: 83-92.
210. Boussinesq M (2012) Loiasis: new epidemiologic insights and proposed treatment
strategy. J Travel Med 19: 140-143.
211. Molyneux DH, Savioli L, Engels D (2017) Neglected tropical diseases: progress towards
addressing the chronic pandemic. Lancet 389: 312-325.
212. Kouam MK, Tchatchueng-Mbougua JB, Demanou M, Boussinesq M, Pion SD, Kamgno
J (2013) Impact of repeated ivermectin treatments against onchocerciasis on the
transmission of loiasis: an entomologic evaluation in central Cameroon. Parasit
Vectors 6: 283.
213. Medeiros JF, Crainey JL, Costa Pessoa FA, Bessa Luz SL (2017) Mansonelliasis. In:
Marcondes CB, editor. Arthropod Borne Diseases: Springer International Publishing.
pp. pp 405-426.
214. Hoerauf A (2009) Mansonella perstans--the importance of an endosymbiont. N Engl J
Med 361: 1502-1504.
215. Gardon J, Kamgno J, Gardon-Wendel N, Demanga N, Duke BO, Boussinesq M (2002)
Efficacy of repeated doses of ivermectin against Mansonella perstans. Trans R Soc
Trop Med Hyg 96: 325-326.

278
216. Coulibaly YI, Dembele B, Diallo AA, Lipner EM, Doumbia SS, Coulibaly SY, Konate
S, Diallo DA, Yalcouye D, Kubofcik J, Doumbo OK, Traore AK, Keita AD, Fay MP,
Traore SF, Nutman TB, Klion AD (2009) A randomized trial of doxycycline for
Mansonella perstans infection. N Engl J Med 361: 1448-1458.
217. Fischer PU, King CL, Jacobson JA, Weil GJ (2017) Potential Value of Triple Drug
Therapy with Ivermectin, Diethylcarbamazine, and Albendazole (IDA) to Accelerate
Elimination of Lymphatic Filariasis and Onchocerciasis in Africa. PLoS Negl Trop
Dis 11: e0005163.
218. Thomsen EK, Sanuku N, Baea M, Satofan S, Maki E, Lombore B, Schmidt MS, Siba
PM, Weil GJ, Kazura JW, Fleckenstein LL, King CL (2016) Efficacy, Safety, and
Pharmacokinetics of Coadministered Diethylcarbamazine, Albendazole, and
Ivermectin for Treatment of Bancroftian Filariasis. Clin Infect Dis 62: 334-341.
219. Bryceson AD, Warrell DA, Pope HM (1977) Dangerous reactions to treatment of
onchocerciasis with diethylcarbamazine. Br Med J 1: 742-744.
220. Taylor HR, Murphy RP, Newland HS, White AT, D'Anna SA, Keyvan-Larijani E, Aziz
MA, Cupp EW, Greene BM (1986) Treatment of onchocerciasis. The ocular effects of
ivermectin and diethylcarbamazine. Arch Ophthalmol 104: 863-870.
221. Doyle SR, Bourguinat C, Nana-Djeunga HC, Kengne-Ouafo JA, Pion SDS, Bopda J,
Kamgno J, Wanji S, Che H, Kuesel AC, Walker M, Basanez MG, Boakye DA, Osei-
Atweneboana MY, Boussinesq M, Prichard RK, Grant WN (2017) Genome-wide
analysis of ivermectin response by Onchocerca volvulus reveals that genetic drift and
soft selective sweeps contribute to loss of drug sensitivity. PLoS Negl Trop Dis 11:
e0005816.
222. Cobo F (2016) Determinants of parasite drug resistance in human lymphatic filariasis.
Rev Esp Quimioter 29: 288-295.
223. Geary TG (2012) Are new anthelmintics needed to eliminate human helminthiases? Curr
Opin Infect Dis 25: 709-717.
224. Yates DM, Portillo V, Wolstenholme AJ (2003) The avermectin receptors of
Haemonchus contortus and Caenorhabditis elegans. Int J Parasitol 33: 1183-1193.
225. Vuong PN, Traore S, Wanji S, Diarrassouba S, Balaton A, Bain O (1992) Ivermectin in
human onchocerciasis: a clinical-pathological study of skin lesions before and three
days after treatment. Ann Parasitol Hum Comp 67: 194-196.
226. Ballesteros C, Tritten L, O'Neill M, Burkman E, Zaky WI, Xia J, Moorhead A, Williams
SA, Geary TG (2016) The Effects of Ivermectin on Brugia malayi Females In Vitro: A
Transcriptomic Approach. PLoS Negl Trop Dis 10: e0004929.
227. Ette EI, Thomas WO, Achumba JI (1990) Ivermectin: a long-acting microfilaricidal
agent. DICP 24: 426-433.
228. Osei-Atweneboana MY, Awadzi K, Attah SK, Boakye DA, Gyapong JO, Prichard RK
(2011) Phenotypic evidence of emerging ivermectin resistance in Onchocerca
volvulus. PLoS Negl Trop Dis 5: e998.
229. Kanesa-thasan N, Douglas JG, Kazura JW (1991) Diethylcarbamazine inhibits
endothelial and microfilarial prostanoid metabolism in vitro. Mol Biochem Parasitol
49: 11-19.
230. Maizels RM, Denham DA (1992) Diethylcarbamazine (DEC): immunopharmacological
interactions of an anti-filarial drug. Parasitology 105 Suppl: S49-60.
231. Peixoto CA, Silva BS (2014) Anti-inflammatory effects of diethylcarbamazine: a review.
Eur J Pharmacol 734: 35-41.
232. Horton J (2000) Albendazole: a review of anthelmintic efficacy and safety in humans.
Parasitology 121 Suppl: S113-132.

279
233. Hoerauf A, Specht S, Buttner M, Pfarr K, Mand S, Fimmers R, Marfo-Debrekyei Y,
Konadu P, Debrah AY, Bandi C, Brattig N, Albers A, Larbi J, Batsa L, Taylor MJ,
Adjei O, Buttner DW (2008) Wolbachia endobacteria depletion by doxycycline as
antifilarial therapy has macrofilaricidal activity in onchocerciasis: a randomized
placebo-controlled study. Med Microbiol Immunol 197: 335.
234. Debrah AY, Specht S, Klarmann-Schulz U, Batsa L, Mand S, Marfo-Debrekyei Y,
Fimmers R, Dubben B, Kwarteng A, Osei-Atweneboana M, Boakye D, Ricchiuto A,
Buttner M, Adjei O, Mackenzie CD, Hoerauf A (2015) Doxycycline Leads to Sterility
and Enhanced Killing of Female Onchocerca volvulus Worms in an Area With
Persistent Microfilaridermia After Repeated Ivermectin Treatment: A Randomized,
Placebo-Controlled, Double-Blind Trial. Clin Infect Dis 61: 517-526.
235. Walker M, Specht S, Churcher TS, Hoerauf A, Taylor MJ, Basanez MG (2015)
Therapeutic efficacy and macrofilaricidal activity of doxycycline for the treatment of
river blindness. Clin Infect Dis 60: 1199-1207.
236. Klarmann-Schulz U, Specht S, Debrah AY, Batsa L, Ayisi-Boateng NK, Osei-Mensah J,
Mubarik Y, Konadu P, Ricchiuto A, Fimmers R, Arriens S, Dubben B, Ford L, Taylor
M, Hoerauf A (2017) Comparison of Doxycycline, Minocycline, Doxycycline plus
Albendazole and Albendazole Alone in Their Efficacy against Onchocerciasis in a
Randomized, Open-Label, Pilot Trial. PLoS Negl Trop Dis 11: e0005156.
237. Debrah AY, Mand S, Marfo-Debrekyei Y, Batsa L, Pfarr K, Buttner M, Adjei O, Buttner
D, Hoerauf A (2007) Macrofilaricidal effect of 4 weeks of treatment with doxycycline
on Wuchereria bancrofti. Trop Med Int Health 12: 1433-1441.
238. Mand S, Debrah AY, Klarmann U, Batsa L, Marfo-Debrekyei Y, Kwarteng A, Specht S,
Belda-Domene A, Fimmers R, Taylor M, Adjei O, Hoerauf A (2012) Doxycycline
improves filarial lymphedema independent of active filarial infection: a randomized
controlled trial. Clin Infect Dis 55: 621-630.
239. Kwarteng A, Ahuno ST, Akoto FO (2016) Killing filarial nematode parasites: role of
treatment options and host immune response. Infect Dis Poverty 5: 86.
240. Mackenzie CD, Geary TG (2011) Flubendazole: a candidate macrofilaricide for
lymphatic filariasis and onchocerciasis field programs. Expert Rev Anti Infect Ther 9:
497-501.
241. Zahner H, Schares G (1993) Experimental chemotherapy of filariasis: comparative
evaluation of the efficacy of filaricidal compounds in Mastomys coucha infected with
Litomosoides carinii, Acanthocheilonema viteae, Brugia malayi and B. pahangi. Acta
Trop 52: 221-266.
242. Moreno L, Alvarez L, Mottier L, Virkel G, Bruni SS, Lanusse C (2004) Integrated
pharmacological assessment of flubendazole potential for use in sheep: disposition
kinetics, liver metabolism and parasite diffusion ability. J Vet Pharmacol Ther 27:
299-308.
243. Ceballos L, Mackenzie C, Geary T, Alvarez L, Lanusse C (2014) Exploring the potential
of flubendazole in filariasis control: evaluation of the systemic exposure for different
pharmaceutical preparations. PLoS Negl Trop Dis 8: e2838.
244. Dominguez-Vazquez A, Taylor HR, Greene BM, Ruvalcaba-Macias AM, Rivas-Alcala
AR, Murphy RP, Beltran-Hernandez F (1983) Comparison of flubendazole and
diethylcarbamazine in treatment of onchocerciasis. Lancet 1: 139-143.
245. Ceballos L, Alvarez L, Mackenzie C, Geary T, Lanusse C (2015) Pharmacokinetic
comparison of different flubendazole formulations in pigs: A further contribution to its
development as a macrofilaricide molecule. Int J Parasitol Drugs Drug Resist 5: 178-
184.

280
246. Tweats DJ, Johnson GE, Scandale I, Whitwell J, Evans DB (2016) Genotoxicity of
flubendazole and its metabolites in vitro and the impact of a new formulation on in
vivo aneugenicity. Mutagenesis 31: 309-321.
247. Guest M, Bull K, Walker RJ, Amliwala K, O'Connor V, Harder A, Holden-Dye L,
Hopper NA (2007) The calcium-activated potassium channel, SLO-1, is required for
the action of the novel cyclo-octadepsipeptide anthelmintic, emodepside, in
Caenorhabditis elegans. Int J Parasitol 37: 1577-1588.
248. Hoerauf A, Satoguina J, Saeftel M, Specht S (2005) Immunomodulation by filarial
nematodes. Parasite Immunol 27: 417-429.
249. Bonne-Annee S, Nutman TB (2017) Human innate lymphoid cells (ILCs) in filarial
infections. Parasite Immunol.
250. Babu S, Nutman TB (2014) Immunology of lymphatic filariasis. Parasite Immunol 36:
338-346.
251. Wolff K, Stingl G (1983) The langerhans cell. J Invest Dermatol 80: 17s-21s.
252. Semnani RT, Nutman TB (2004) Toward an understanding of the interaction between
filarial parasites and host antigen-presenting cells. Immunol Rev 201: 127-138.
253. Le Goff L, Loke P, Ali HF, Taylor DW, Allen JE (2000) Interleukin-5 is essential for
vaccine-mediated immunity but not innate resistance to a filarial parasite. Infect
Immun 68: 2513-2517.
254. Martin C, Saeftel M, Vuong PN, Babayan S, Fischer K, Bain O, Hoerauf A (2001) B-cell
deficiency suppresses vaccine-induced protection against murine filariasis but does not
increase the recovery rate for primary infection. Infect Immun 69: 7067-7073.
255. Saeftel M, Arndt M, Specht S, Volkmann L, Hoerauf A (2003) Synergism of gamma
interferon and interleukin-5 in the control of murine filariasis. Infect Immun 71: 6978-
6985.
256. Chirgwin SR, Coleman SU, Porthouse KH, Klei TR (2006) Tissue migration capability
of larval and adult Brugia pahangi. J Parasitol 92: 46-51.
257. Pionnier N, Brotin E, Karadjian G, Hemon P, Gaudin-Nome F, Vallarino-Lhermitte N,
Nieguitsila A, Fercoq F, Aknin ML, Marin-Esteban V, Chollet-Martin S, Schlecht-
Louf G, Bachelerie F, Martin C (2016) Neutropenic Mice Provide Insight into the
Role of Skin-Infiltrating Neutrophils in the Host Protective Immunity against Filarial
Infective Larvae. PLoS Negl Trop Dis 10: e0004605.
258. Ajendra J, Specht S, Ziewer S, Schiefer A, Pfarr K, Parcina M, Kufer TA, Hoerauf A,
Hubner MP (2016) NOD2 dependent neutrophil recruitment is required for early
protective immune responses against infectious Litomosoides sigmodontis L3 larvae.
Sci Rep 6: 39648.
259. Le Goff L, Martin C, Oswald IP, Vuong PN, Petit G, Ungeheuer MN, Bain O (2000)
Parasitology and immunology of mice vaccinated with irradiated Litomosoides
sigmodontis larvae. Parasitology 120 ( Pt 3): 271-280.
260. Martin C, Al-Qaoud KM, Ungeheuer MN, Paehle K, Vuong PN, Bain O, Fleischer B,
Hoerauf A (2000) IL-5 is essential for vaccine-induced protection and for resolution of
primary infection in murine filariasis. Med Microbiol Immunol 189: 67-74.
261. Bleiss W, Oberlander U, Hartmann S, Adam R, Marko A, Schonemeyer A, Lucius R
(2002) Protective immunity induced by irradiated third-stage larvae of the filaria
Acanthocheilonema viteae is directed against challenge third-stage larvae before
molting. J Parasitol 88: 264-270.
262. Abraham D, Leon O, Schnyder-Candrian S, Wang CC, Galioto AM, Kerepesi LA, Lee
JJ, Lustigman S (2004) Immunoglobulin E and eosinophil-dependent protective
immunity to larval Onchocerca volvulus in mice immunized with irradiated larvae.
Infect Immun 72: 810-817.

281
263. Specht S, Saeftel M, Arndt M, Endl E, Dubben B, Lee NA, Lee JJ, Hoerauf A (2006)
Lack of eosinophil peroxidase or major basic protein impairs defense against murine
filarial infection. Infect Immun 74: 5236-5243.
264. Specht S, Frank JK, Alferink J, Dubben B, Layland LE, Denece G, Bain O, Forster I,
Kirschning CJ, Martin C, Hoerauf A (2011) CCL17 controls mast cells for the defense
against filarial larval entry. J Immunol 186: 4845-4852.
265. Semnani RT, Law M, Kubofcik J, Nutman TB (2004) Filaria-induced immune evasion:
suppression by the infective stage of Brugia malayi at the earliest host-parasite
interface. J Immunol 172: 6229-6238.
266. Cotton RN, McDonald-Fleming R, Boyd A, Spates K, Nutman TB, Tolouei Semnani R
(2015) Brugia malayi infective larvae fail to activate Langerhans cells and dermal
dendritic cells in human skin. Parasite Immunol 37: 79-91.
267. Hise AG, Daehnel K, Gillette-Ferguson I, Cho E, McGarry HF, Taylor MJ, Golenbock
DT, Fitzgerald KA, Kazura JW, Pearlman E (2007) Innate immune responses to
endosymbiotic Wolbachia bacteria in Brugia malayi and Onchocerca volvulus are
dependent on TLR2, TLR6, MyD88, and Mal, but not TLR4, TRIF, or TRAM. J
Immunol 178: 1068-1076.
268. Turner JD, Langley RS, Johnston KL, Egerton G, Wanji S, Taylor MJ (2006) Wolbachia
endosymbiotic bacteria of Brugia malayi mediate macrophage tolerance to TLR- and
CD40-specific stimuli in a MyD88/TLR2-dependent manner. J Immunol 177: 1240-
1249.
269. Turner JD, Langley RS, Johnston KL, Gentil K, Ford L, Wu B, Graham M, Sharpley F,
Slatko B, Pearlman E, Taylor MJ (2009) Wolbachia lipoprotein stimulates innate and
adaptive immunity through Toll-like receptors 2 and 6 to induce disease
manifestations of filariasis. J Biol Chem 284: 22364-22378.
270. Brattig NW, Bazzocchi C, Kirschning CJ, Reiling N, Buttner DW, Ceciliani F, Geisinger
F, Hochrein H, Ernst M, Wagner H, Bandi C, Hoerauf A (2004) The major surface
protein of Wolbachia endosymbionts in filarial nematodes elicits immune responses
through TLR2 and TLR4. J Immunol 173: 437-445.
271. Karadjian G (2014) Coinfections naturelles et experimentales : interactions hôte-
parasites chez les oiseaux (Haemosporidies de rapaces) et dans un modèle murin
(Plasmodium sp./Litomosoides sigmodontis). Muséum National d'Histoire Naturelle.
272. Gentil K, Lentz CS, Rai R, Muhsin M, Kamath AD, Mutluer O, Specht S, Hubner MP,
Hoerauf A (2014) Eotaxin-1 is involved in parasite clearance during chronic filarial
infection. Parasite Immunol 36: 60-77.
273. Boyd A, Killoran K, Mitre E, Nutman TB (2015) Pleural cavity type 2 innate lymphoid
cells precede Th2 expansion in murine Litomosoides sigmodontis infection. Exp
Parasitol 159: 118-126.
274. Babayan S, Attout T, Specht S, Hoerauf A, Snounou G, Renia L, Korenaga M, Bain O,
Martin C (2005) Increased early local immune responses and altered worm
development in high-dose infections of mice susceptible to the filaria Litomosoides
sigmodontis. Med Microbiol Immunol 194: 151-162.
275. Loke P, MacDonald AS, Allen JE (2000) Antigen-presenting cells recruited by Brugia
malayi induce Th2 differentiation of naive CD4(+) T cells. Eur J Immunol 30: 1127-
1135.
276. Batra H, Antony VB (2015) Pleural mesothelial cells in pleural and lung diseases. J
Thorac Dis 7: 964-980.
277. Allen JE, Sutherland TE (2014) Host protective roles of type 2 immunity: parasite killing
and tissue repair, flip sides of the same coin. Semin Immunol 26: 329-340.

282
278. Taylor MD, van der Werf N, Harris A, Graham AL, Bain O, Allen JE, Maizels RM
(2009) Early recruitment of natural CD4+ Foxp3+ Treg cells by infective larvae
determines the outcome of filarial infection. Eur J Immunol 39: 192-206.
279. Hartmann W, Schramm C, Breloer M (2015) Litomosoides sigmodontis induces TGF-
beta receptor responsive, IL-10-producing T cells that suppress bystander T-cell
proliferation in mice. Eur J Immunol 45: 2568-2581.
280. Taylor MD, Harris A, Babayan SA, Bain O, Culshaw A, Allen JE, Maizels RM (2007)
CTLA-4 and CD4+ CD25+ regulatory T cells inhibit protective immunity to filarial
parasites in vivo. J Immunol 179: 4626-4634.
281. Jackson-Jones LH, Duncan SM, Magalhaes MS, Campbell SM, Maizels RM, McSorley
HJ, Allen JE, Benezech C (2016) Fat-associated lymphoid clusters control local IgM
secretion during pleural infection and lung inflammation. Nat Commun 7: 12651.
282. Al-Qaoud KM, Fleischer B, Hoerauf A (1998) The Xid defect imparts susceptibility to
experimental murine filariosis--association with a lack of antibody and IL-10
production by B cells in response to phosphorylcholine. Int Immunol 10: 17-25.
283. Ruckerl D, Allen JE (2014) Macrophage proliferation, provenance, and plasticity in
macroparasite infection. Immunol Rev 262: 113-133.
284. van der Werf N, Redpath SA, Azuma M, Yagita H, Taylor MD (2013) Th2 cell-intrinsic
hypo-responsiveness determines susceptibility to helminth infection. Plos Pathogens
9: e1003215.
285. Taylor MD, LeGoff L, Harris A, Malone E, Allen JE, Maizels RM (2005) Removal of
regulatory T cell activity reverses hyporesponsiveness and leads to filarial parasite
clearance in vivo. J Immunol 174: 4924-4933.
286. Layland LE, Ajendra J, Ritter M, Wiszniewsky A, Hoerauf A, Hubner MP (2015)
Development of patent Litomosoides sigmodontis infections in semi-susceptible
C57BL/6 mice in the absence of adaptive immune responses. Parasit Vectors 8: 396.
287. Arndts K, Deininger S, Specht S, Klarmann U, Mand S, Adjobimey T, Debrah AY,
Batsa L, Kwarteng A, Epp C, Taylor M, Adjei O, Layland LE, Hoerauf A (2012)
Elevated adaptive immune responses are associated with latent infections of
Wuchereria bancrofti. PLoS Negl Trop Dis 6: e1611.
288. Rodrigo MB, Schulz S, Krupp V, Ritter M, Wiszniewsky K, Arndts K, Tamadaho RS,
Endl E, Hoerauf A, Layland LE (2016) Patency of Litomosoides sigmodontis infection
depends on Toll-like receptor 4 whereas Toll-like receptor 2 signalling influences
filarial-specific CD4(+) T-cell responses. Immunology 147: 429-442.
289. Pfarr KM, Fischer K, Hoerauf A (2003) Involvement of Toll-like receptor 4 in the
embryogenesis of the rodent filaria Litomosoides sigmodontis. Med Microbiol
Immunol 192: 53-56.
290. Fox EM, Morris CP, Hubner MP, Mitre E (2015) Histamine 1 Receptor Blockade
Enhances Eosinophil-Mediated Clearance of Adult Filarial Worms. PLoS Negl Trop
Dis 9: e0003932.
291. Volkmann L, Bain O, Saeftel M, Specht S, Fischer K, Brombacher F, Matthaei KI,
Hoerauf A (2003) Murine filariasis: interleukin 4 and interleukin 5 lead to
containment of different worm developmental stages. Med Microbiol Immunol 192:
23-31.
292. Martin C, Le Goff L, Ungeheuer MN, Vuong PN, Bain O (2000) Drastic reduction of a
filarial infection in eosinophilic interleukin-5 transgenic mice. Infect Immun 68: 3651-
3656.
293. Cadman ET, Thysse KA, Bearder S, Cheung AY, Johnston AC, Lee JJ, Lawrence RA
(2014) Eosinophils are important for protection, immunoregulation and pathology
during infection with nematode microfilariae. Plos Pathogens 10: e1003988.

283
294. Al-Qaoud KM, Pearlman E, Hartung T, Klukowski J, Fleischer B, Hoerauf A (2000) A
new mechanism for IL-5-dependent helminth control: neutrophil accumulation and
neutrophil-mediated worm encapsulation in murine filariasis are abolished in the
absence of IL-5. Int Immunol 12: 899-908.
295. Marieb E (1999) Anatomie et physiologie humaines. 1194 p.
296. Tomashefeski J, Farver C (2009) Anatomy and Histology of the Lung. Dail and
Hammar's Pulmonary Pathology: Springer New York. pp. 20-48.
297. Negrini D, Moriondo A (2013) Pleural function and lymphatics. Acta Physiol (Oxf) 207:
244-259.
298. Wang NS (1974) The regional difference of pleural mesothelial cells in rabbits. Am Rev
Respir Dis 110: 623-633.
299. Mutsaers SE (2004) The mesothelial cell. Int J Biochem Cell Biol 36: 9-16.
300. Mutsaers SE (2002) Mesothelial cells: their structure, function and role in serosal repair.
Respirology 7: 171-191.
301. Rock JR, Randell SH, Hogan BL (2010) Airway basal stem cells: a perspective on their
roles in epithelial homeostasis and remodeling. Dis Model Mech 3: 545-556.
302. Murray JF (2010) The structure and function of the lung. Int J Tuberc Lung Dis 14: 391-
396.
303. Suarez C, Dintzis S, Frevert C (2012) Respiratory. Comparative Anatomy and Histology
: a Mouse and Human Atlas: Elsevier. pp. 121-134.
304. Iwasaki A, Foxman EF, Molony RD (2017) Early local immune defences in the
respiratory tract. Nat Rev Immunol 17: 7-20.
305. Knust J, Ochs M, Gundersen HJ, Nyengaard JR (2009) Stereological estimates of
alveolar number and size and capillary length and surface area in mice lungs. Anat
Rec (Hoboken) 292: 113-122.
306. Townsley MI (2012) Structure and composition of pulmonary arteries, capillaries, and
veins. Compr Physiol 2: 675-709.
307. Weibel ER, Gomez DM (1962) Architecture of the human lung. Use of quantitative
methods establishes fundamental relations between size and number of lung structures.
Science 137: 577-585.
308. Birks EK, Mathieu-Costello O, Fu Z, Tyler WS, West JB (1994) Comparative aspects of
the strength of pulmonary capillaries in rabbit, dog, and horse. Respir Physiol 97: 235-
246.
309. Okiemy G, Foucault C, Avisse C, Hidden G, Riquet M (2003) Lymphatic drainage of the
diaphragmatic pleura to the peritracheobronchial lymph nodes. Surg Radiol Anat 25:
32-35.
310. Ahmad K, Gandhi MS, Smith WC, Weber KT (2008) Bilateral pleural effusions in
congestive heart failure. Am J Med Sci 335: 484-488.
311. Riquet M, Le Pimpec-Barthes F, Hidden G (2001) Lymphatic drainage of the
pericardium to the mediastinal lymph nodes. Surg Radiol Anat 23: 317-319.
312. Riquet M (2007) Bronchial arteries and lymphatics of the lung. Thorac Surg Clin 17:
619-638, viii.
313. Sevin CM, Light RW (2011) Microscopic anatomy of the pleura. Thorac Surg Clin 21:
173-175, vii.
314. Riquet M (1993) Anatomic basis of lymphatic spread from carcinoma of the lung to the
mediastinum: surgical and prognostic implications. Surg Radiol Anat 15: 271-277.
315. Booth AJ, Hadley R, Cornett AM, Dreffs AA, Matthes SA, Tsui JL, Weiss K, Horowitz
JC, Fiore VF, Barker TH, Moore BB, Martinez FJ, Niklason LE, White ES (2012)
Acellular normal and fibrotic human lung matrices as a culture system for in vitro
investigation. Am J Respir Crit Care Med 186: 866-876.

284
316. Kang I, Harten IA, Chang MY, Braun KR, Sheih A, Nivison MP, Johnson PY, Workman
G, Kaber G, Evanko SP, Chan CK, Merrilees MJ, Ziegler SF, Kinsella MG, Frevert
CW, Wight TN (2017) Versican Deficiency Significantly Reduces Lung Inflammatory
Response Induced by Polyinosine-Polycytidylic Acid Stimulation. J Biol Chem 292:
51-63.
317. Wight TN, Frevert CW, Debley JS, Reeves SR, Parks WC, Ziegler SF (2017) Interplay
of extracellular matrix and leukocytes in lung inflammation. Cell Immunol 312: 1-14.
318. Gibbings SL, Thomas SM, Atif SM, McCubbrey AL, Desch AN, Danhorn T, Leach SM,
Bratton DL, Henson PM, Janssen WJ, Jakubzick CV (2017) Three Unique Interstitial
Macrophages in the Murine Lung at Steady State. Am J Respir Cell Mol Biol.
319. Pabst R (2004) The periarterial space in the lung: its important role in lung edema,
transplantation, and microbial or allergic inflammation. Pathobiology 71: 287-294.
320. Chen F, Wu W, Millman A, Craft JF, Chen E, Patel N, Boucher JL, Urban JF, Jr., Kim
CC, Gause WC (2014) Neutrophils prime a long-lived effector macrophage phenotype
that mediates accelerated helminth expulsion. Nat Immunol 15: 938-946.
321. Wang PM, Kachel DL, Cesta MF, Martin WJ, 2nd (2011) Direct leukocyte migration
across pulmonary arterioles and venules into the perivascular interstitium of murine
lungs during bleomycin injury and repair. Am J Pathol 178: 2560-2572.
322. Komai-Koma M, Brombacher F, Pushparaj PN, Arendse B, McSharry C, Alexander J,
Chaudhuri R, Thomson NC, McKenzie AN, McInnes I, Liew FY, Xu D (2012)
Interleukin-33 amplifies IgE synthesis and triggers mast cell degranulation via
interleukin-4 in naive mice. Allergy 67: 1118-1126.
323. Mizutani N, Nabe T, Yoshino S (2013) Interleukin-33 and alveolar macrophages
contribute to the mechanisms underlying the exacerbation of IgE-mediated airway
inflammation and remodelling in mice. Immunology 139: 205-218.
324. Katoh S, Matsumoto N, Kawakita K, Tominaga A, Kincade PW, Matsukura S (2003) A
role for CD44 in an antigen-induced murine model of pulmonary eosinophilia. J Clin
Invest 111: 1563-1570.
325. Nishimura T, Katoh S, Mori A, Ohtomo T, Saeki M, Hiroi T, Kaminuma O (2016)
Critical role of CD44 in antigen-induced Th2- but not Th17-madiated murine airway
inflammation. Allergol Int 65 Suppl: S59-61.
326. Hirose J, Kawashima H, Yoshie O, Tashiro K, Miyasaka M (2001) Versican interacts
with chemokines and modulates cellular responses. J Biol Chem 276: 5228-5234.
327. Wu YJ, La Pierre DP, Wu J, Yee AJ, Yang BB (2005) The interaction of versican with
its binding partners. Cell Res 15: 483-494.
328. Kopf M, Schneider C, Nobs SP (2015) The development and function of lung-resident
macrophages and dendritic cells. Nat Immunol 16: 36-44.
329. Davies LC, Jenkins SJ, Allen JE, Taylor PR (2013) Tissue-resident macrophages. Nat
Immunol 14: 986-995.
330. Garbi N, Lambrecht BN (2017) Location, function, and ontogeny of pulmonary
macrophages during the steady state. Pflugers Arch 469: 561-572.
331. Bain CC, Mowat AM (2012) CD200 receptor and macrophage function in the intestine.
Immunobiology 217: 643-651.
332. Gardai SJ, Xiao YQ, Dickinson M, Nick JA, Voelker DR, Greene KE, Henson PM
(2003) By binding SIRPalpha or calreticulin/CD91, lung collectins act as dual
function surveillance molecules to suppress or enhance inflammation. Cell 115: 13-23.
333. Scott CL, Henri S, Guilliams M (2014) Mononuclear phagocytes of the intestine, the
skin, and the lung. Immunol Rev 262: 9-24.

285
334. Kambara T, Wilson RA (1990) In situ pulmonary responses of T cell and macrophage
subpopulations to a challenge infection in mice vaccinated with irradiated cercariae of
Schistosoma mansoni. J Parasitol 76: 365-372.
335. Breel M, van der Ende MB, Sminia T, Kraal G (1988) Subpopulations of non-lymphoid
cells in bronchus associated lymphoid tissue and lung of the mouse. Adv Exp Med
Biol 237: 607-613.
336. Ramaswamy K, De Sanctis GT, Green F, Befus D (1991) Pathology of pulmonary
parasitic migration: morphological and bronchoalveolar cellular responses following
Nippostrongylus brasiliensis infection in rats. J Parasitol 77: 302-312.
337. Loukas A, Prociv P (2001) Immune responses in hookworm infections. Clin Microbiol
Rev 14: 689-703, table of contents.
338. Vetter JC, van der Linden ME (1977) Skin penetration of infective hookworm larvae. I.
The path of migration of infective larvae of Ancylostoma braziliense in canine skin. Z
Parasitenkd 53: 255-262.
339. Chen F, Liu Z, Wu W, Rozo C, Bowdridge S, Millman A, Van Rooijen N, Urban JF, Jr.,
Wynn TA, Gause WC (2012) An essential role for TH2-type responses in limiting
acute tissue damage during experimental helminth infection. Nat Med 18: 260-266.
340. Hung LY, Lewkowich IP, Dawson LA, Downey J, Yang Y, Smith DE, Herbert DR
(2013) IL-33 drives biphasic IL-13 production for noncanonical Type 2 immunity
against hookworms. Proc Natl Acad Sci U S A 110: 282-287.
341. Isailovic N, Daigo K, Mantovani A, Selmi C (2015) Interleukin-17 and innate immunity
in infections and chronic inflammation. J Autoimmun 60: 1-11.
342. Sutherland TE, Logan N, Ruckerl D, Humbles AA, Allan SM, Papayannopoulos V,
Stockinger B, Maizels RM, Allen JE (2014) Chitinase-like proteins promote IL-17-
mediated neutrophilia in a tradeoff between nematode killing and host damage. Nat
Immunol 15: 1116-1125.
343. O'Connell AE, Hess JA, Santiago GA, Nolan TJ, Lok JB, Lee JJ, Abraham D (2011)
Major basic protein from eosinophils and myeloperoxidase from neutrophils are
required for protective immunity to Strongyloides stercoralis in mice. Infect Immun
79: 2770-2778.
344. Bonne-Annee S, Kerepesi LA, Hess JA, O'Connell AE, Lok JB, Nolan TJ, Abraham D
(2013) Human and mouse macrophages collaborate with neutrophils to kill larval
Strongyloides stercoralis. Infect Immun 81: 3346-3355.
345. Bouchery T, Kyle R, Camberis M, Shepherd A, Filbey K, Smith A, Harvie M, Painter G,
Johnston K, Ferguson P, Jain R, Roediger B, Delahunt B, Weninger W, Forbes-Blom
E, Le Gros G (2015) ILC2s and T cells cooperate to ensure maintenance of M2
macrophages for lung immunity against hookworms. Nat Commun 6: 6970.
346. Thawer S, Auret J, Schnoeller C, Chetty A, Smith K, Darby M, Roberts L, Mackay RM,
Whitwell HJ, Timms JF, Madsen J, Selkirk ME, Brombacher F, Clark HW, Horsnell
WG (2016) Surfactant Protein-D Is Essential for Immunity to Helminth Infection. Plos
Pathogens 12: e1005461.
347. Turner JE, Morrison PJ, Wilhelm C, Wilson M, Ahlfors H, Renauld JC, Panzer U,
Helmby H, Stockinger B (2013) IL-9-mediated survival of type 2 innate lymphoid
cells promotes damage control in helminth-induced lung inflammation. J Exp Med
210: 2951-2965.
348. Coyle AJ, Kohler G, Tsuyuki S, Brombacher F, Kopf M (1998) Eosinophils are not
required to induce airway hyperresponsiveness after nematode infection. Eur J
Immunol 28: 2640-2647.
349. Minutti CM, Jackson-Jones LH, Garcia-Fojeda B, Knipper JA, Sutherland TE, Logan N,
Rinqvist E, Guillamat-Prats R, Ferenbach DA, Artigas A, Stamme C, Chroneos ZC,

286
Zaiss DM, Casals C, Allen JE (2017) Local amplifiers of IL-4Ralpha-mediated
macrophage activation promote repair in lung and liver. Science 356: 1076-1080.
350. Minutti CM, Knipper JA, Allen JE, Zaiss DM (2017) Tissue-specific contribution of
macrophages to wound healing. Semin Cell Dev Biol 61: 3-11.
351. Marsland BJ, Kurrer M, Reissmann R, Harris NL, Kopf M (2008) Nippostrongylus
brasiliensis infection leads to the development of emphysema associated with the
induction of alternatively activated macrophages. Eur J Immunol 38: 479-488.
352. Webb DC, Mahalingam S, Cai Y, Matthaei KI, Donaldson DD, Foster PS (2003)
Antigen-specific production of interleukin (IL)-13 and IL-5 cooperate to mediate IL-
4Ralpha-independent airway hyperreactivity. Eur J Immunol 33: 3377-3385.
353. Noben-Trauth N, Shultz LD, Brombacher F, Urban JF, Jr., Gu H, Paul WE (1997) An
interleukin 4 (IL-4)-independent pathway for CD4+ T cell IL-4 production is revealed
in IL-4 receptor-deficient mice. Proc Natl Acad Sci U S A 94: 10838-10843.
354. Kopf M, Brombacher F, Hodgkin PD, Ramsay AJ, Milbourne EA, Dai WJ, Ovington
KS, Behm CA, Kohler G, Young IG, Matthaei KI (1996) IL-5-deficient mice have a
developmental defect in CD5+ B-1 cells and lack eosinophilia but have normal
antibody and cytotoxic T cell responses. Immunity 4: 15-24.
355. Manitz MP, Horst B, Seeliger S, Strey A, Skryabin BV, Gunzer M, Frings W, Schonlau
F, Roth J, Sorg C, Nacken W (2003) Loss of S100A9 (MRP14) results in reduced
interleukin-8-induced CD11b surface expression, a polarized microfilament system,
and diminished responsiveness to chemoattractants in vitro. Mol Cell Biol 23: 1034-
1043.
356. Hoffmeister K, Wenk P (1991) Experiments on the regulation of the worm load at the
rodant filariae Litomosoides carinii (Nematoda Filaroidae) in Sigmoda hispidus. Mitt
Österr Ges Tropenmed Parasitol 13: 119-124.
357. Marechal P (1995) Deux filaires du genre Litomosoides chez la souris blanche;
régulation du développement: PhD thesis, Muséum National d'Histoire Naturelle.
358. Papayannopoulos V, Metzler KD, Hakkim A, Zychlinsky A (2010) Neutrophil elastase
and myeloperoxidase regulate the formation of neutrophil extracellular traps. J Cell
Biol 191: 677-691.
359. Masuda S, Nakazawa D, Shida H, Miyoshi A, Kusunoki Y, Tomaru U, Ishizu A (2016)
NETosis markers: Quest for specific, objective, and quantitative markers. Clin Chim
Acta 459: 89-93.
360. Babayan SA, Attout T, Harris A, Taylor MD, Le Goff L, Vuong PN, Renia L, Allen JE,
Bain O (2006) Vaccination against filarial nematodes with irradiated larvae provides
long-term protection against the third larval stage but not against subsequent life cycle
stages. Int J Parasitol 36: 903-914.
361. Bain O, Chabaud AG (1986) [Atlas of infective larvae of filariae]. Trop Med Parasitol
37: 301-340.
362. Armstrong SD, Babayan SA, Lhermitte-Vallarino N, Gray N, Xia D, Martin C, Kumar S,
Taylor DW, Blaxter ML, Wastling JM, Makepeace BL (2014) Comparative analysis
of the secretome from a model filarial nematode (Litomosoides sigmodontis) reveals
maximal diversity in gravid female parasites. Mol Cell Proteomics 13: 2527-2544.
363. Miller AM (2011) Role of IL-33 in inflammation and disease. J Inflamm (Lond) 8: 22.
364. Martin NT, Martin MU (2016) Interleukin 33 is a guardian of barriers and a local
alarmin. Nat Immunol 17: 122-131.
365. Nussbaum JC, Van Dyken SJ, von Moltke J, Cheng LE, Mohapatra A, Molofsky AB,
Thornton EE, Krummel MF, Chawla A, Liang HE, Locksley RM (2013) Type 2 innate
lymphoid cells control eosinophil homeostasis. Nature 502: 245-248.

287
366. van Rijt L, von Richthofen H, van Ree R (2016) Type 2 innate lymphoid cells: at the
cross-roads in allergic asthma. Seminars in Immunopathology 38: 483-496.
367. Madsen DH, Bugge TH (2013) Imaging collagen degradation in vivo highlights a key
role for M2-polarized macrophages in extracellular matrix degradation.
Oncoimmunology 2: e27127.
368. Jeyaseelan S, Manzer R, Young SK, Yamamoto M, Akira S, Mason RJ, Worthen GS
(2005) Induction of CXCL5 during inflammation in the rodent lung involves
activation of alveolar epithelium. Am J Respir Cell Mol Biol 32: 531-539.
369. Schagat TL, Wofford JA, Wright JR (2001) Surfactant protein A enhances alveolar
macrophage phagocytosis of apoptotic neutrophils. J Immunol 166: 2727-2733.
370. Jaillon S, Galdiero MR, Del Prete D, Cassatella MA, Garlanda C, Mantovani A (2013)
Neutrophils in innate and adaptive immunity. Seminars in Immunopathology 35: 377-
394.
371. Prince LR, Whyte MK, Sabroe I, Parker LC (2011) The role of TLRs in neutrophil
activation. Curr Opin Pharmacol 11: 397-403.
372. Clarke TB, Davis KM, Lysenko ES, Zhou AY, Yu Y, Weiser JN (2010) Recognition of
peptidoglycan from the microbiota by Nod1 enhances systemic innate immunity. Nat
Med 16: 228-231.
373. Tamassia N, Bazzoni F, Le Moigne V, Calzetti F, Masala C, Grisendi G, Bussmeyer U,
Scutera S, De Gironcoli M, Costantini C, Musso T, Cassatella MA (2012) IFN-beta
expression is directly activated in human neutrophils transfected with plasmid DNA
and is further increased via TLR-4-mediated signaling. J Immunol 189: 1500-1509.
374. Kolaczkowska E, Kubes P (2013) Neutrophil recruitment and function in health and
inflammation. Nat Rev Immunol 13: 159-175.
375. Metzler KD, Goosmann C, Lubojemska A, Zychlinsky A, Papayannopoulos V (2014) A
myeloperoxidase-containing complex regulates neutrophil elastase release and actin
dynamics during NETosis. Cell Rep 8: 883-896.
376. Nauseef WM, Borregaard N (2014) Neutrophils at work. Nat Immunol 15: 602-611.
377. Abi Abdallah DS, Denkers EY (2012) Neutrophils cast extracellular traps in response to
protozoan parasites. Front Immunol 3: 382.
378. Branzk N, Lubojemska A, Hardison SE, Wang Q, Gutierrez MG, Brown GD,
Papayannopoulos V (2014) Neutrophils sense microbe size and selectively release
neutrophil extracellular traps in response to large pathogens. Nat Immunol 15: 1017-
1025.
379. Urban CF, Ermert D, Schmid M, Abu-Abed U, Goosmann C, Nacken W, Brinkmann V,
Jungblut PR, Zychlinsky A (2009) Neutrophil Extracellular Traps Contain
Calprotectin, a Cytosolic Protein Complex Involved in Host Defense against Candida
albicans. Plos Pathogens 5.
380. Bonne-Annee S, Kerepesi LA, Hess JA, Wesolowski J, Paumet F, Lok JB, Nolan TJ,
Abraham D (2014) Extracellular traps are associated with human and mouse
neutrophil and macrophage mediated killing of larval Strongyloides stercoralis.
Microbes Infect 16: 502-511.
381. McCoy CJ, Reaves BJ, Giguere S, Coates R, Rada B, Wolstenholme AJ (2017) Human
Leukocytes Kill Brugia malayi Microfilariae Independently of DNA-Based
Extracellular Trap Release. PLoS Negl Trop Dis 11: e0005279.
382. Tamarozzi F, Turner JD, Pionnier N, Midgley A, Guimaraes AF, Johnston KL, Edwards
SW, Taylor MJ (2016) Wolbachia endosymbionts induce neutrophil extracellular trap
formation in human onchocerciasis. Sci Rep 6: 35559.

288
383. Edgeworth J, Gorman M, Bennett R, Freemont P, Hogg N (1991) Identification of p8,14
as a highly abundant heterodimeric calcium binding protein complex of myeloid cells.
J Biol Chem 266: 7706-7713.
384. Bargagli E, Olivieri C, Cintorino M, Refini RM, Bianchi N, Prasse A, Rottoli P (2011)
Calgranulin B (S100A9/MRP14): a key molecule in idiopathic pulmonary fibrosis?
Inflammation 34: 85-91.
385. Lorenz E, Muhlebach MS, Tessier PA, Alexis NE, Duncan Hite R, Seeds MC, Peden
DB, Meredith W (2008) Different expression ratio of S100A8/A9 and S100A12 in
acute and chronic lung diseases. Respir Med 102: 567-573.
386. Tirkos S, Newbigging S, Nguyen V, Keet M, Ackerley C, Kent G, Rozmahel RF (2006)
Expression of S100A8 correlates with inflammatory lung disease in congenic mice
deficient of the cystic fibrosis transmembrane conductance regulator. Respir Res 7: 51.
387. Kawai H, Minamiya Y, Takahashi N (2011) Prognostic impact of S100A9
overexpression in non-small cell lung cancer. Tumour Biol 32: 641-646.
388. Hiroshima Y, Hsu K, Tedla N, Wong SW, Chow S, Kawaguchi N, Geczy CL (2017)
S100A8/A9 and S100A9 reduce acute lung injury. Immunol Cell Biol 95: 461-472.
389. Zhao J, Endoh I, Hsu K, Tedla N, Endoh Y, Geczy CL (2011) S100A8 modulates mast
cell function and suppresses eosinophil migration in acute asthma. Antioxid Redox
Signal 14: 1589-1600.
390. Nikitorowicz-Buniak J, Shiwen X, Denton CP, Abraham D, Stratton R (2014)
Abnormally differentiating keratinocytes in the epidermis of systemic sclerosis
patients show enhanced secretion of CCN2 and S100A9. J Invest Dermatol 134: 2693-
2702.
391. Lee MJ, Lee JK, Choi JW, Lee CS, Sim JH, Cho CH, Lee KH, Cho IH, Chung MH, Kim
HR, Ye SK (2012) Interleukin-6 induces S100A9 expression in colonic epithelial cells
through STAT3 activation in experimental ulcerative colitis. PLoS One 7: e38801.
392. Henke MO, Renner A, Rubin BK, Gyves JI, Lorenz E, Koo JS (2006) Up-regulation of
S100A8 and S100A9 protein in bronchial epithelial cells by lipopolysaccharide. Exp
Lung Res 32: 331-347.
393. Frosch M, Metze D, Foell D, Vogl T, Sorg C, Sunderkotter C, Roth J (2005) Early
activation of cutaneous vessels and epithelial cells is characteristic of acute systemic
onset juvenile idiopathic arthritis. Exp Dermatol 14: 259-265.
394. Arai K, Teratani T, Kuruto-Niwa R, Yamada T, Nozawa R (2004) S100A9 expression in
invasive ductal carcinoma of the breast: S100A9 expression in adenocarcinoma is
closely associated with poor tumour differentiation. Eur J Cancer 40: 1179-1187.
395. Cross SS, Hamdy FC, Deloulme JC, Rehman I (2005) Expression of S100 proteins in
normal human tissues and common cancers using tissue microarrays: S100A6,
S100A8, S100A9 and S100A11 are all overexpressed in common cancers.
Histopathology 46: 256-269.
396. Markowitz J, Carson WE, 3rd (2013) Review of S100A9 biology and its role in cancer.
Biochim Biophys Acta 1835: 100-109.
397. Moore BW (1965) A soluble protein characteristic of the nervous system. Biochem
Biophys Res Commun 19: 739-744.
398. Austermann J, Zenker S, Roth J (2017) S100-alarmins: potential therapeutic targets for
arthritis. Expert Opin Ther Targets 21: 739-751.
399. Donato R (2001) S100: a multigenic family of calcium-modulated proteins of the EF-
hand type with intracellular and extracellular functional roles. Int J Biochem Cell Biol
33: 637-668.
400. Donato R, Cannon BR, Sorci G, Riuzzi F, Hsu K, Weber DJ, Geczy CL (2013)
Functions of S100 proteins. Curr Mol Med 13: 24-57.

289
401. Gross SR, Sin CG, Barraclough R, Rudland PS (2014) Joining S100 proteins and
migration: for better or for worse, in sickness and in health. Cell Mol Life Sci 71:
1551-1579.
402. Heizmann CW, Fritz G, Schafer BW (2002) S100 proteins: structure, functions and
pathology. Front Biosci 7: d1356-1368.
403. Chen H, Xu C, Jin Q, Liu Z (2014) S100 protein family in human cancer. Am J Cancer
Res 4: 89-115.
404. Ravasi T, Hsu K, Goyette J, Schroder K, Yang Z, Rahimi F, Miranda LP, Alewood PF,
Hume DA, Geczy C (2004) Probing the S100 protein family through genomic and
functional analysis. Genomics 84: 10-22.
405. Marenholz I, Heizmann CW, Fritz G (2004) S100 proteins in mouse and man: from
evolution to function and pathology (including an update of the nomenclature).
Biochem Biophys Res Commun 322: 1111-1122.
406. Engelkamp D, Schafer BW, Mattei MG, Erne P, Heizmann CW (1993) Six S100 genes
are clustered on human chromosome 1q21: identification of two genes coding for the
two previously unreported calcium-binding proteins S100D and S100E. Proc Natl
Acad Sci U S A 90: 6547-6551.
407. Schafer BW, Wicki R, Engelkamp D, Mattei MG, Heizmann CW (1995) Isolation of a
YAC clone covering a cluster of nine S100 genes on human chromosome 1q21:
rationale for a new nomenclature of the S100 calcium-binding protein family.
Genomics 25: 638-643.
408. Pietas A, Schluns K, Marenholz I, Schafer BW, Heizmann CW, Petersen I (2002)
Molecular cloning and characterization of the human S100A14 gene encoding a novel
member of the S100 family. Genomics 79: 513-522.
409. Zimmer DB, Chessher J, Song W (1996) Nucleotide homologies in genes encoding
members of the S100 protein family. Biochim Biophys Acta 1313: 229-238.
410. Zimmer DB, Cornwall EH, Landar A, Song W (1995) The S100 protein family: history,
function, and expression. Brain Res Bull 37: 417-429.
411. Deloulme JC, Assard N, Mbele GO, Mangin C, Kuwano R, Baudier J (2000) S100A6
and S100A11 are specific targets of the calcium- and zinc-binding S100B protein in
vivo. J Biol Chem 275: 35302-35310.
412. Hunter MJ, Chazin WJ (1998) High level expression and dimer characterization of the
S100 EF-hand proteins, migration inhibitory factor-related proteins 8 and 14. J Biol
Chem 273: 12427-12435.
413. Isobe T, Ishioka N, Okuyama T (1981) Structural relation of two S-100 proteins in
bovine brain; subunit composition of S-100a protein. Eur J Biochem 115: 469-474.
414. Yang Q, O'Hanlon D, Heizmann CW, Marks A (1999) Demonstration of heterodimer
formation between S100B and S100A6 in the yeast two-hybrid system and human
melanoma. Exp Cell Res 246: 501-509.
415. Korndorfer IP, Brueckner F, Skerra A (2007) The crystal structure of the human
(S100A8/S100A9)2 heterotetramer, calprotectin, illustrates how conformational
changes of interacting alpha-helices can determine specific association of two EF-
hand proteins. J Mol Biol 370: 887-898.
416. Passey RJ, Williams E, Lichanska AM, Wells C, Hu S, Geczy CL, Little MH, Hume DA
(1999) A null mutation in the inflammation-associated S100 protein S100A8 causes
early resorption of the mouse embryo. J Immunol 163: 2209-2216.
417. Pruenster M, Vogl T, Roth J, Sperandio M (2016) S100A8/A9: From basic science to
clinical application. Pharmacol Ther 167: 120-131.
418. Vogl T, Ludwig S, Goebeler M, Strey A, Thorey IS, Reichelt R, Foell D, Gerke V,
Manitz MP, Nacken W, Werner S, Sorg C, Roth J (2004) MRP8 and MRP14 control

290
microtubule reorganization during transendothelial migration of phagocytes. Blood
104: 4260-4268.
419. Vogl T, Gharibyan AL, Morozova-Roche LA (2012) Pro-inflammatory S100A8 and
S100A9 proteins: self-assembly into multifunctional native and amyloid complexes.
Int J Mol Sci 13: 2893-2917.
420. Wang Y, Gao H, Kessinger CW, Schmaier A, Jaffer FA, Simon DI (2017) Myeloid-
related protein-14 regulates deep vein thrombosis. JCI Insight 2.
421. Bianchi M, Niemiec MJ, Siler U, Urban CF, Reichenbach J (2011) Restoration of anti-
Aspergillus defense by neutrophil extracellular traps in human chronic granulomatous
disease after gene therapy is calprotectin-dependent. J Allergy Clin Immunol 127:
1243-1252 e1247.
422. Pruenster M, Kurz AR, Chung KJ, Cao-Ehlker X, Bieber S, Nussbaum CF, Bierschenk
S, Eggersmann TK, Rohwedder I, Heinig K, Immler R, Moser M, Koedel U, Gran S,
McEver RP, Vestweber D, Verschoor A, Leanderson T, Chavakis T, Roth J, Vogl T,
Sperandio M (2015) Extracellular MRP8/14 is a regulator of beta2 integrin-dependent
neutrophil slow rolling and adhesion. Nat Commun 6: 6915.
423. Ryckman C, McColl SR, Vandal K, de Medicis R, Lussier A, Poubelle PE, Tessier PA
(2003) Role of S100A8 and S100A9 in neutrophil recruitment in response to
monosodium urate monohydrate crystals in the air-pouch model of acute gouty
arthritis. Arthritis Rheum 48: 2310-2320.
424. Anceriz N, Vandal K, Tessier PA (2007) S100A9 mediates neutrophil adhesion to
fibronectin through activation of beta2 integrins. Biochem Biophys Res Commun 354:
84-89.
425. Newton RA, Hogg N (1998) The human S100 protein MRP-14 is a novel activator of the
beta 2 integrin Mac-1 on neutrophils. J Immunol 160: 1427-1435.
426. Ehlermann P, Eggers K, Bierhaus A, Most P, Weichenhan D, Greten J, Nawroth PP,
Katus HA, Remppis A (2006) Increased proinflammatory endothelial response to
S100A8/A9 after preactivation through advanced glycation end products. Cardiovasc
Diabetol 5: 6.
427. Wang L, Luo H, Chen X, Jiang Y, Huang Q (2014) Functional characterization of
S100A8 and S100A9 in altering monolayer permeability of human umbilical
endothelial cells. PLoS One 9: e90472.
428. Viemann D, Strey A, Janning A, Jurk K, Klimmek K, Vogl T, Hirono K, Ichida F, Foell
D, Kehrel B, Gerke V, Sorg C, Roth J (2005) Myeloid-related proteins 8 and 14
induce a specific inflammatory response in human microvascular endothelial cells.
Blood 105: 2955-2962.
429. Atallah M, Krispin A, Trahtemberg U, Ben-Hamron S, Grau A, Verbovetski I, Mevorach
D (2012) Constitutive neutrophil apoptosis: regulation by cell concentration via S100
A8/9 and the MEK-ERK pathway. PLoS One 7: e29333.
430. Lim SY, Raftery MJ, Geczy CL (2011) Oxidative modifications of DAMPs suppress
inflammation: the case for S100A8 and S100A9. Antioxid Redox Signal 15: 2235-
2248.
431. Lim SY, Raftery MJ, Goyette J, Geczy CL (2010) S-glutathionylation regulates
inflammatory activities of S100A9. J Biol Chem 285: 14377-14388.
432. Lim SY, Raftery MJ, Goyette J, Hsu K, Geczy CL (2009) Oxidative modifications of
S100 proteins: functional regulation by redox. J Leukoc Biol 86: 577-587.
433. Sun Y, Lu Y, Engeland CG, Gordon SC, Sroussi HY (2013) The anti-oxidative, anti-
inflammatory, and protective effect of S100A8 in endotoxemic mice. Mol Immunol
53: 443-449.

291
434. Gomes LH, Raftery MJ, Yan WX, Goyette JD, Thomas PS, Geczy CL (2013) S100A8
and S100A9-oxidant scavengers in inflammation. Free Radic Biol Med 58: 170-186.
435. Sroussi HY, Lu Y, Zhang QL, Villines D, Marucha PT (2010) S100A8 and S100A9
inhibit neutrophil oxidative metabolism in-vitro: involvement of adenosine
metabolites. Free Radic Res 44: 389-396.
436. Sroussi HY, Lu Y, Villines D, Sun Y (2012) The down regulation of neutrophil
oxidative metabolism by S100A8 and S100A9: implication of the protease-activated
receptor-2. Mol Immunol 50: 42-48.
437. Damo SM, Kehl-Fie TE, Sugitani N, Holt ME, Rathi S, Murphy WJ, Zhang Y, Betz C,
Hench L, Fritz G, Skaar EP, Chazin WJ (2013) Molecular basis for manganese
sequestration by calprotectin and roles in the innate immune response to invading
bacterial pathogens. Proc Natl Acad Sci U S A 110: 3841-3846.
438. Corbin BD, Seeley EH, Raab A, Feldmann J, Miller MR, Torres VJ, Anderson KL,
Dattilo BM, Dunman PM, Gerads R, Caprioli RM, Nacken W, Chazin WJ, Skaar EP
(2008) Metal chelation and inhibition of bacterial growth in tissue abscesses. Science
319: 962-965.
439. Austermann J, Friesenhagen J, Fassl SK, Petersen B, Ortkras T, Burgmann J, Barczyk-
Kahlert K, Faist E, Zedler S, Pirr S, Rohde C, Muller-Tidow C, von Kockritz-
Blickwede M, von Kaisenberg CS, Flohe SB, Ulas T, Schultze JL, Roth J, Vogl T,
Viemann D (2014) Alarmins MRP8 and MRP14 induce stress tolerance in phagocytes
under sterile inflammatory conditions. Cell Rep 9: 2112-2123.
440. Petersen B, Wolf M, Austermann J, van Lent P, Foell D, Ahlmann M, Kupas V, Loser K,
Sorg C, Roth J, Vogl T (2013) The alarmin Mrp8/14 as regulator of the adaptive
immune response during allergic contact dermatitis. EMBO J 32: 100-111.
441. Shimizu K, Libby P, Rocha VZ, Folco EJ, Shubiki R, Grabie N, Jang S, Lichtman AH,
Shimizu A, Hogg N, Simon DI, Mitchell RN, Croce K (2011) Loss of myeloid related
protein-8/14 exacerbates cardiac allograft rejection. Circulation 124: 2920-2932.
442. Pagano RL, Sampaio SC, Juliano L, Juliano MA, Giorgi R (2005) The C-terminus of
murine S100A9 inhibits spreading and phagocytic activity of adherent peritoneal cells.
Inflamm Res 54: 204-210.
443. Pagano RL, Moraes NF, De Lorenzo BH, Coccuzzo Sampaio S, Mariano M, Giorgi R
(2014) Inhibition of macrophage functions by the C-terminus of murine S100A9 is
dependent on B-1 cells. Mediators Inflamm 2014: 836491.
444. De Lorenzo BH, Godoy LC, Novaes e Brito RR, Pagano RL, Amorim-Dias MA, Grosso
DM, Lopes JD, Mariano M (2010) Macrophage suppression following phagocytosis of
apoptotic neutrophils is mediated by the S100A9 calcium-binding protein.
Immunobiology 215: 341-347.
445. Bjork P, Bjork A, Vogl T, Stenstrom M, Liberg D, Olsson A, Roth J, Ivars F,
Leanderson T (2009) Identification of human S100A9 as a novel target for treatment
of autoimmune disease via binding to quinoline-3-carboxamides. PLoS Biol 7: e97.
446. Robinson MJ, Tessier P, Poulsom R, Hogg N (2002) The S100 family heterodimer,
MRP-8/14, binds with high affinity to heparin and heparan sulfate
glycosaminoglycans on endothelial cells. J Biol Chem 277: 3658-3665.
447. Hoppmann S, Steinbach J, Pietzsch J (2010) Scavenger receptors are associated with
cellular interactions of S100A12 in vitro and in vivo. Int J Biochem Cell Biol 42: 651-
661.
448. Kerkhoff C, Sorg C, Tandon NN, Nacken W (2001) Interaction of S100A8/S100A9-
arachidonic acid complexes with the scavenger receptor CD36 may facilitate fatty acid
uptake by endothelial cells. Biochemistry 40: 241-248.

292
449. Loser K, Vogl T, Voskort M, Lueken A, Kupas V, Nacken W, Klenner L, Kuhn A, Foell
D, Sorokin L, Luger TA, Roth J, Beissert S (2010) The Toll-like receptor 4 ligands
Mrp8 and Mrp14 are crucial in the development of autoreactive CD8+ T cells. Nat
Med 16: 713-717.
450. Vogl T, Tenbrock K, Ludwig S, Leukert N, Ehrhardt C, van Zoelen MA, Nacken W,
Foell D, van der Poll T, Sorg C, Roth J (2007) Mrp8 and Mrp14 are endogenous
activators of Toll-like receptor 4, promoting lethal, endotoxin-induced shock. Nat Med
13: 1042-1049.
451. Hibino T, Sakaguchi M, Miyamoto S, Yamamoto M, Motoyama A, Hosoi J, Shimokata
T, Ito T, Tsuboi R, Huh NH (2013) S100A9 is a novel ligand of EMMPRIN that
promotes melanoma metastasis. Cancer Res 73: 172-183.
452. Croce K, Gao H, Wang Y, Mooroka T, Sakuma M, Shi C, Sukhova GK, Packard RR,
Hogg N, Libby P, Simon DI (2009) Myeloid-related protein-8/14 is critical for the
biological response to vascular injury. Circulation 120: 427-436.
453. Gopal R, Monin L, Torres D, Slight S, Mehra S, McKenna KC, Fallert Junecko BA,
Reinhart TA, Kolls J, Baez-Saldana R, Cruz-Lagunas A, Rodriguez-Reyna TS, Kumar
NP, Tessier P, Roth J, Selman M, Becerril-Villanueva E, Baquera-Heredia J,
Cumming B, Kasprowicz VO, Steyn AJ, Babu S, Kaushal D, Zuniga J, Vogl T,
Rangel-Moreno J, Khader SA (2013) S100A8/A9 proteins mediate neutrophilic
inflammation and lung pathology during tuberculosis. Am J Respir Crit Care Med
188: 1137-1146.
454. Park HY, Jin JO, Song MG, Park JI, Kwak JY (2007) Expression of dendritic cell
markers on cultured neutrophils and its modulation by anti-apoptotic and pro-
apoptotic compounds. Exp Mol Med 39: 439-449.
455. Lee NR, Park BS, Kim SY, Gu A, Kim DH, Lee JS, Kim IS (2016) Cytokine secreted by
S100A9 via TLR4 in monocytes delays neutrophil apoptosis by inhibition of caspase
9/3 pathway. Cytokine 86: 53-63.
456. Gebhardt C, Nemeth J, Angel P, Hess J (2006) S100A8 and S100A9 in inflammation
and cancer. Biochem Pharmacol 72: 1622-1631.
457. Chen B, Miller AL, Rebelatto M, Brewah Y, Rowe DC, Clarke L, Czapiga M, Rosenthal
K, Imamichi T, Chen Y, Chang CS, Chowdhury PS, Naiman B, Wang Y, Yang D,
Humbles AA, Herbst R, Sims GP (2015) S100A9 induced inflammatory responses are
mediated by distinct damage associated molecular patterns (DAMP) receptors in vitro
and in vivo. PLoS One 10: e0115828.
458. Goodridge HS, Marshall FA, Else KJ, Houston KM, Egan C, Al-Riyami L, Liew FY,
Harnett W, Harnett MM (2005) Immunomodulation via novel use of TLR4 by the
filarial nematode phosphorylcholine-containing secreted product, ES-62. J Immunol
174: 284-293.
459. Venugopal PG, Nutman TB, Semnani RT (2009) Activation and regulation of toll-like
receptors (TLRs) by helminth parasites. Immunol Res 43: 252-263.
460. Hoffmann WH, Pfaff AW, Schulz-Key H, Soboslay PT (2001) Determinants for
resistance and susceptibility to microfilaraemia in Litomosoides sigmodontis filariasis.
Parasitology 122: 641-649.
461. Pfaff AW, Schulz-Key H, Soboslay PT, Geiger SM, Hoffmann WH (2000) Litomosoides
sigmodontis: dynamics of the survival of microfilariae in resistant and susceptible
strains of mice. Exp Parasitol 94: 67-74.
462. Specht S, Taylor MD, Hoeve MA, Allen JE, Lang R, Hoerauf A (2012) Over expression
of IL-10 by macrophages overcomes resistance to murine filariasis. Exp Parasitol 132:
90-96.

293
463. Haas B, Wenk P (1981) Elimination of microfilariae (Litomosoides carinii Filarioidea) in
the patent and in the immunized cotton-rat. Trans R Soc Trop Med Hyg 75: 143-144.
464. Vij M, Kumari N, Krishnani N (2011) Microfilaria in liver aspiration cytology: an
extremely rare finding. Diagn Cytopathol 39: 521-522.
465. Vankalakunti M, Jha PK, Ravishankar B, Vishwanath S, Rampure S, Ballal HS (2011)
Microfilariae-associated nephrotic range proteinuria. Kidney Int 79: 1152.
466. Varghese R, Raghuveer CV, Pai MR, Bansal R (1996) Microfilariae in cytologic smears:
a report of six cases. Acta Cytol 40: 299-301.
467. Wenk P (1986) The function of non circulating microfilariae: Litomosoides carinii
(Nematoda: Filarioidea). Dtsch Tierarztl Wochenschr 93: 414-418.
468. Boucherat O, Boczkowski J, Jeannotte L, Delacourt C (2013) Cellular and molecular
mechanisms of goblet cell metaplasia in the respiratory airways. Exp Lung Res 39:
207-216.
469. Alevy YG, Patel AC, Romero AG, Patel DA, Tucker J, Roswit WT, Miller CA, Heier
RF, Byers DE, Brett TJ, Holtzman MJ (2012) IL-13-induced airway mucus production
is attenuated by MAPK13 inhibition. J Clin Invest 122: 4555-4568.
470. Wang ZB, Li M, Li JC (2010) Recent advances in the research of lymphatic stomata.
Anat Rec (Hoboken) 293: 754-761.
471. Ugorski M, Dziegiel P, Suchanski J (2016) Podoplanin - a small glycoprotein with many
faces. Am J Cancer Res 6: 370-386.
472. Takagi S, Oh-hara T, Sato S, Gong B, Takami M, Fujita N (2014) Expression of
Aggrus/podoplanin in bladder cancer and its role in pulmonary metastasis. Int J
Cancer 134: 2605-2614.
473. Kimura N, Kimura I (2005) Podoplanin as a marker for mesothelioma. Pathol Int 55: 83-
86.
474. Saeftel M, Volkmann L, Korten S, Brattig N, Al-Qaoud K, Fleischer B, Hoerauf A
(2001) Lack of interferon-gamma confers impaired neutrophil granulocyte function
and imparts prolonged survival of adult filarial worms in murine filariasis. Microbes
Infect 3: 203-213.
475. Volkmann L, Saeftel M, Bain O, Fischer K, Fleischer B, Hoerauf A (2001) Interleukin-4
is essential for the control of microfilariae in murine infection with the filaria
Litomosoides sigmodontis. Infect Immun 69: 2950-2956.
476. Le Goff L, Lamb TJ, Graham AL, Harcus Y, Allen JE (2002) IL-4 is required to prevent
filarial nematode development in resistant but not susceptible strains of mice. Int J
Parasitol 32: 1277-1284.
477. Ziewer S, Hubner MP, Dubben B, Hoffmann WH, Bain O, Martin C, Hoerauf A, Specht
S (2012) Immunization with L. sigmodontis microfilariae reduces peripheral
microfilaraemia after challenge infection by inhibition of filarial embryogenesis. PLoS
Negl Trop Dis 6: e1558.
478. Takeuchi S, Fukuda K, Yamada T, Arai S, Takagi S, Ishii G, Ochiai A, Iwakiri S, Itoi K,
Uehara H, Nishihara H, Fujita N, Yano S (2017) Podoplanin promotes progression of
malignant pleural mesothelioma by regulating motility and focus formation. Cancer
Sci 108: 696-703.
479. Padgett DM, Cathro HP, Wick MR, Mills SE (2008) Podoplanin is a better
immunohistochemical marker for sarcomatoid mesothelioma than calretinin. Am J
Surg Pathol 32: 123-127.
480. Wynn TA, Ramalingam TR (2012) Mechanisms of fibrosis: therapeutic translation for
fibrotic disease. Nat Med 18: 1028-1040.

294
481. Inomata M, Kamio K, Azuma A, Matsuda K, Kokuho N, Miura Y, Hayashi H, Nei T,
Fujita K, Saito Y, Gemma A (2014) Pirfenidone inhibits fibrocyte accumulation in the
lungs in bleomycin-induced murine pulmonary fibrosis. Respir Res 15: 16.
482. Phillips RJ, Burdick MD, Hong K, Lutz MA, Murray LA, Xue YY, Belperio JA, Keane
MP, Strieter RM (2004) Circulating fibrocytes traffic to the lungs in response to
CXCL12 and mediate fibrosis. J Clin Invest 114: 438-446.
483. Moore BB, Kolodsick JE, Thannickal VJ, Cooke K, Moore TA, Hogaboam C, Wilke
CA, Toews GB (2005) CCR2-mediated recruitment of fibrocytes to the alveolar space
after fibrotic injury. Am J Pathol 166: 675-684.
484. Tsukui T, Ueha S, Shichino S, Inagaki Y, Matsushima K (2015) Intratracheal cell
transfer demonstrates the profibrotic potential of resident fibroblasts in pulmonary
fibrosis. Am J Pathol 185: 2939-2948.
485. Hoyles RK, Derrett-Smith EC, Khan K, Shiwen X, Howat SL, Wells AU, Abraham DJ,
Denton CP (2011) An essential role for resident fibroblasts in experimental lung
fibrosis is defined by lineage-specific deletion of high-affinity type II transforming
growth factor beta receptor. Am J Respir Crit Care Med 183: 249-261.
486. Takano M, Nekomoto C, Kawami M, Yumoto R (2017) Role of miR-34a in TGF-beta1-
and Drug-Induced Epithelial-Mesenchymal Transition in Alveolar Type II Epithelial
Cells. J Pharm Sci.
487. Zolak JS, Jagirdar R, Surolia R, Karki S, Oliva O, Hock T, Guroji P, Ding Q, Liu RM,
Bolisetty S, Agarwal A, Thannickal VJ, Antony VB (2013) Pleural mesothelial cell
differentiation and invasion in fibrogenic lung injury. Am J Pathol 182: 1239-1247.
488. Stone RC, Pastar I, Ojeh N, Chen V, Liu S, Garzon KI, Tomic-Canic M (2016)
Epithelial-mesenchymal transition in tissue repair and fibrosis. Cell Tissue Res 365:
495-506.
489. Decologne N, Kolb M, Margetts PJ, Menetrier F, Artur Y, Garrido C, Gauldie J, Camus
P, Bonniaud P (2007) TGF-beta1 induces progressive pleural scarring and subpleural
fibrosis. J Immunol 179: 6043-6051.
490. Wu Y, Liu Q, Yan X, Kato Y, Tanaka M, Inokuchi S, Yoshizawa T, Morohashi S,
Kijima H (2016) Podoplanin-mediated TGF-beta-induced epithelial-mesenchymal
transition and its correlation with bHLH transcription factor DEC in TE-11 cells. Int J
Oncol 48: 2310-2320.
491. Kalluri R, Weinberg RA (2009) The basics of epithelial-mesenchymal transition. J Clin
Invest 119: 1420-1428.
492. Mutsaers SE, Birnie K, Lansley S, Herrick SE, Lim CB, Prele CM (2015) Mesothelial
cells in tissue repair and fibrosis. Front Pharmacol 6: 113.
493. Prasad R, Goel MK, Mukerji PK, Agarwal PK (1994) Microfilaria in bronchial aspirate.
Indian J Chest Dis Allied Sci 36: 223-225.
494. Anupindi L, Sahoo R, Rao RV, Verghese G, Rao PV (1993) Microfilariae in bronchial
brushing cytology of symptomatic pulmonary lesions. A report of two cases. Acta
Cytol 37: 397-399.
495. Gupta P, Lai SM, Sheng J, Tetlak P, Balachander A, Claser C, Renia L, Karjalainen K,
Ruedl C (2016) Tissue-Resident CD169(+) Macrophages Form a Crucial Front Line
against Plasmodium Infection. Cell Rep 16: 1749-1761.
496. Barrera V, Skorokhod OA, Baci D, Gremo G, Arese P, Schwarzer E (2011) Host
fibrinogen stably bound to hemozoin rapidly activates monocytes via TLR-4 and
CD11b/CD18-integrin: a new paradigm of hemozoin action. Blood 117: 5674-5682.
497. Klaas M, Crocker PR (2012) Sialoadhesin in recognition of self and non-self. Seminars
in Immunopathology 34: 353-364.

295
498. Rao UR, Chandrashekar R, Subrahmanyam D (1987) Litomosoides carinii:
characterization of surface carbohydrates of microfilariae and infective larvae. Trop
Med Parasitol 38: 15-18.
499. Karasawa K, Asano K, Moriyama S, Ushiki M, Monya M, Iida M, Kuboki E, Yagita H,
Uchida K, Nitta K, Tanaka M (2015) Vascular-resident CD169-positive monocytes
and macrophages control neutrophil accumulation in the kidney with ischemia-
reperfusion injury. J Am Soc Nephrol 26: 896-906.
500. Barlow JL, Peel S, Fox J, Panova V, Hardman CS, Camelo A, Bucks C, Wu X, Kane
CM, Neill DR, Flynn RJ, Sayers I, Hall IP, McKenzie AN (2013) IL-33 is more potent
than IL-25 in provoking IL-13-producing nuocytes (type 2 innate lymphoid cells) and
airway contraction. J Allergy Clin Immunol 132: 933-941.
501. Cheng D, Xue Z, Yi L, Shi H, Zhang K, Huo X, Bonser LR, Zhao J, Xu Y, Erle DJ,
Zhen G (2014) Epithelial interleukin-25 is a key mediator in Th2-high, corticosteroid-
responsive asthma. Am J Respir Crit Care Med 190: 639-648.
502. Divekar R, Kita H (2015) Recent advances in epithelium-derived cytokines (IL-33, IL-
25, and thymic stromal lymphopoietin) and allergic inflammation. Curr Opin Allergy
Clin Immunol 15: 98-103.
503. Mitchell PD, O'Byrne PM (2017) Biologics and the lung: TSLP and other epithelial cell-
derived cytokines in asthma. Pharmacol Ther 169: 104-112.
504. Lloyd CM, Saglani S (2015) Epithelial cytokines and pulmonary allergic inflammation.
Curr Opin Immunol 34: 52-58.
505. Christianson CA, Goplen NP, Zafar I, Irvin C, Good JT, Jr., Rollins DR, Gorentla B, Liu
W, Gorska MM, Chu H, Martin RJ, Alam R (2015) Persistence of asthma requires
multiple feedback circuits involving type 2 innate lymphoid cells and IL-33. J Allergy
Clin Immunol 136: 59-68 e14.
506. Lambrecht BN, Hammad H (2015) The immunology of asthma. Nat Immunol 16: 45-56.
507. Lei Y, Gregory JA, Nilsson GP, Adner M (2013) Insights into mast cell functions in
asthma using mouse models. Pulm Pharmacol Ther 26: 532-539.
508. Bennett RD, Ysasi AB, Belle JM, Wagner WL, Konerding MA, Blainey PC, Pyne S,
Mentzer SJ (2014) Laser microdissection of the alveolar duct enables single-cell
genomic analysis. Front Oncol 4: 260.
509. Ikutani M, Tsuneyama K, Kawaguchi M, Fukuoka J, Kudo F, Nakae S, Arita M, Nagai
Y, Takaki S, Takatsu K (2017) Prolonged activation of IL-5-producing ILC2 causes
pulmonary arterial hypertrophy. JCI Insight 2: e90721.
510. Ikutani M, Yanagibashi T, Ogasawara M, Tsuneyama K, Yamamoto S, Hattori Y, Kouro
T, Itakura A, Nagai Y, Takaki S, Takatsu K (2012) Identification of innate IL-5-
producing cells and their role in lung eosinophil regulation and antitumor immunity. J
Immunol 188: 703-713.
511. Yasuda K, Muto T, Kawagoe T, Matsumoto M, Sasaki Y, Matsushita K, Taki Y,
Futatsugi-Yumikura S, Tsutsui H, Ishii KJ, Yoshimoto T, Akira S, Nakanishi K
(2012) Contribution of IL-33-activated type II innate lymphoid cells to pulmonary
eosinophilia in intestinal nematode-infected mice. Proc Natl Acad Sci U S A 109:
3451-3456.
512. Almeida FF, Belz GT (2016) Innate lymphoid cells: models of plasticity for immune
homeostasis and rapid responsiveness in protection. Mucosal Immunol 9: 1103-1112.
513. Ealey KN, Moro K, Koyasu S (2017) Are ILC2s Jekyll and Hyde in airway
inflammation? Immunol Rev 278: 207-218.
514. Myburgh E, Ritchie R, Goundry A, O'Neill K, Marchesi F, Devaney E (2016) Attempts
to Image the Early Inflammatory Response during Infection with the Lymphatic
Filarial Nematode Brugia pahangi in a Mouse Model. PLoS One 11: e0168602.

296
515. Gross S, Gammon ST, Moss BL, Rauch D, Harding J, Heinecke JW, Ratner L, Piwnica-
Worms D (2009) Bioluminescence imaging of myeloperoxidase activity in vivo. Nat
Med 15: 455-461.
516. Tseng JC, Kung AL (2012) In vivo imaging of inflammatory phagocytes. Chem Biol 19:
1199-1209.
Copyright © 2022 FDOKUMEN




















