
DIPLOMARBEIT Information Extraction – Utilizing Table Patterns

Interaction between clay minerals and
hydrocarbon-utilizing indigenous
microorganisms in high concentrations of
heavy oil: implications for bioremediation
SITI KHODIJAH CHAERUN1 ,* , KAZUE TAZAKI
2, RYUJI ASADA
2AND
KAZUHIRO KOGURE3
1Graduate School of Natural Science and Technology, Kanazawa University, Kakuma, Kanazawa 920-1192,
2
Department of Earth Sciences, Faculty of Science, Kanazawa University, Kakuma, Kanazawa 920-1192, and3Ocean
Research Institute, University of Tokyo, Minamidai, Nakano, Tokyo 164-8639, Japan
(Received 10 February 2004; revised 23 July 2004)
ABSTRACT: This study focused on whether the presence of clay minerals (montmorillonite and
kaolinite) in marine or coastal environments contaminated with high concentrations of heavy-oil
spills were able to support the growth of hydrocarbon degraders to enable bioremediation. The
bacterial growth experiment utilizing ~150 g/l of heavy oil (from the Nakhodka oil spill) was
conducted with 1500 mg/l of montmorillonite or kaolinite. Bacterial strain Pseudomonas aeruginosa
(isolated from Atake seashore, Ishikawa Prefecture, Japan), capable of degrading heavy oil, was
employed in combination with other hydrocarbon degraders inhabiting the heavy oil and seawater
(collected from the Sea of Japan). The interactions among microbial cells, clay minerals and heavy
oil were studied. Both clays were capable of promoting microbial growth and allowed
microorganisms to proliferate (to a greater degree than in a control sample which contained no
clay) in an extremely high concentration of heavy oil. Observation by transmission electron
microscopy of the clay-oil-cell complexes showed that microbial cells tended to be bound primarily
on the edges of the clays. X-ray diffraction analysis showed that the clay-oil and clay-oil-cell
complexes involved the adsorption of microbial cells and/or heavy oil on the external surfaces of the
clays. How do the interactions among clay minerals, microbial cells and heavy oil contribute to
environmental factors influencing the bioremediation process? To our knowledge, there are no
previous reports on the use of clay minerals in the bioremediation of the Nakhodka oil spill in
combination with biofilm formation.
KEYWORDS: clay minerals, montmorillonite, kaolinite, hydrocarbon degraders, heavy oil, bioremediation,
Nakhodka, oil spill, Pseudomonas aeruginosa, biofilm formation.
Microorganisms are the major agents mineralizing
organic pollutants in terrestrial and aquatic environ-
ments, and their adsorption on soil mineral colloids
(e.g. clay minerals) is of great importance (Ortega-
Calvo & Saiz-Jimenez, 1998). Laboratory and field
investigations have shown that the biodegradation
of organic pollutants is enhanced or inhibited in the
presence of minerals, clays and organic soil
constituents (Weber & Coble, 1968; Ogram et al.,
1985; Robinson et al., 1990; Miller & Alexander,
1991; Scow & Alexander, 1992; Guerin & Boyd,
1992; Knaebel et al., 1994), and that clay minerals
have a role in the sorption of organic and inorganic
compounds as we l l a s mic roo rgan i sms
(Schiffenbauer & Stotzky, 1982; Lipson &
* E-mail: [email protected]
DOI: 10.1180/0009855054010159
ClayMinerals (2005) 40, 105±114
# 2005 The Mineralogical Society

Stotzky, 1983; Lipson & Stotzky, 1984; Boyd et al.,
1988; Lee et al., 1990; Khanna & Stotzky, 1992;
Tapp & Stotzky; 1995; Vettori et al., 1999; Lin &
Puls, 2000). Nevertheless, no data exist on the
simultaneous use of clay minerals as adsorbents and
carriers in degrading heavy oil. A mechanism
describing the interactions between clay minerals,
heavy oil and microorganisms is also required.
Microorganisms are generally able to grow at low
substrate concentrations. However, not all micro-
organisms are able to grow on, and consume, high
concentrations of substrate (i.e. heavy oil) as the
carbon and energy source, since high substrate
concentrations may cause inhibition of some
microbial enzymatic reactions (known as substrate
inhibition (Shuler & Kargi, 1992)). Hence, a study
was conducted to address the interactions between
clay minerals and microorganisms in high concen-
trations of heavy oil (~150 g/l).
The primary objective of this study was to
investigate the interactions between clay minerals
and hydrocarbon-utilizing indigenous microorgan-
isms in high concentrations of heavy oil (collected
from the Nakhodka oil spill). These interactions
were studied by X-ray powder diffractrometry
(XRD), scanning electron microscopy (SEM) and
transmission electron microscopy (TEM) in an
attempt to determine where and how microbial
cells are bound to the clay minerals (montmorillon-
ite and kaolinite) that are abundant in nature. These
techniques were also used to evaluate the interac-
tions between microorganisms, clay minerals and
heavy oil. Such understanding might provide
information about the factors influencing the
bioremediation of oil spills.
MATER IALS AND METHODS
Clay minerals
The montmorillonite standard JCSS-3102 (from
Mikawa, Japan) and kaolinite standard JCSS-1101
(from Kanpaku, Japan) used in this study were
obtained from The Clay Science Society of Japan
(Okayama, Japan). Both clays were used in batch
experiments without pretreatment.
Bacterial strain
Apart from microorganisms inhabiting natural
seawater (NSW) and heavy oil, a bacterium was
also added as an inoculum in order to promote the
degradation of the heavy oil. The bacterium was
isolated from Atake seashore, Ishikawa Prefecture,
Japan. Using 16S rDNA sequencing, this bacterium
was affiliated to Pseudomonas aeruginosa (98%
similarity). This bacterium was selected for study
because it produced high extracellular emulsifying
activity after 2 days of incubation in the presence of
heavy oil (2% v/v) and 1 g/l of yeast extract
(Chaerun et al., 2003).
Bacterial culture and batch experimental
conditions
Batch bacterial experiments were conducted with
montmorillonite standard JCSS-3102 and kaolinite
standard JCSS-1101. The growth medium contained
the necessary components needed for bacterial
growth. The following were included for each
litre of medium: 25 g of NH4NO3, 0.5 g of
FeC6H5O7´nH2O (ferric citrate), and 0.5 g of
K2HPO4/l of distilled, deionized water (NDW).
Natural seawater without filtration (collected from
the Sea of Japan) was also added to each flask as
additional medium (Table 1). Briefly, the batch
reaction vessels consisted of acid-washed 1000 ml
Erlenmeyer flasks containing 600 ml of growth
medium: 480 ml of NSW was mixed with 120 ml
of an autoclaved NDW. The montmorillonite or
kaolinite (1500 mg/l) was added to the flask and the
pH of the medium was adjusted to pH 7.8 with 1N
NaOH solution. Immediately after sample prepara-
tion, before introducing the inoculum which
originated from the heavy oil and the NSW
medium into the flasks, all five flasks with their
contents were autoclaved, allowed to cool to room
TABLE 1. Elemental composition of natural seawater
(NSW) collected from the Sea of Japan that was used
for medium analysed by ED-XRF.
Element Wt.%
Na 8.27
Mg 1.49
Si 0.26
S 2.94
Cl 79.94
K 2.99
Ca 3.57
Co 0.05
Br 0.42
Sr 0.06
106 S. K. Chaerun et al.

temperature (24ëC), and then inoculated with NSW
medium, the stock liquid culture of bacterial strain
Pseudomonas aeruginosa (4.5% v/v), and heavy oil
(collected from the Nakhodka oil spill) to a final
concentration of ~150 g/l, serving as the sole
carbon and energy sources (Table 2) (Chaerun &
Tazaki, 2003; Chaerun et al., 2003; Tazaki, 2003).
Overall, five experimental flasks were established:
two main flasks contained medium, bacteria, plus
montmorillonite (coded as R1) or kaolinite (coded as
R2); two abiotic control flasks contained medium
plus montmorillonite (coded as R3) or medium plus
kaolinite (coded as R4); and one biotic control flask
contained medium plus bacteria (coded as R5). To
avoid microbial growth and to ensure sterility during
the experiment, abiotic control flasks were auto-
claved three times and heavy oil solution was
sterilized by filtration through a 0.22 mm pore-
sized filter. Cultures were incubated for 105 days at
room temperature (24ëC) with shaking at 125 rpm,
allowing microorganisms to grow. Samples were
removed periodically at sampling periods of 0, 36,
64 and 105 days. Separate sets of samples were
made up and prepared for analysis by XRD, SEM
and TEM. In addition, the liquid medium of three
experimental systems (i.e. R1, R2 and R5) was
supplemented with 1 g/l of yeast extract after
36 days of the experiments.
X-ray diffractometery
Relative changes in the basal spacings of
montmorillonite or kaolinite were measured by
XRD. Samples were prepared by spreading 2 mg
of the suspended clays over a 2.5 cm2area of a
glass slide. The slides were air-dried, placed in a
desiccator containing silica gel to prevent rehydra-
tion, and observed using Cu-Ka generated at 40 kV
and 30 mA by using the 2y/y method and a scan
speed 2ë/min (Rigaku Rinto 1200 X-ray diffract-
ometer). To determine whether the montmorillonite
remained expandable or not, samples of mont-
morillonite-oil and montmorillonite-oil-cell
complexes were treated with ethylene glycol and
then dried at room temperature before being
analysed by XRD.
Microscopic techniques
For observation by SEM, freeze drying was used
for sample preparation (Suzuki et al., 1995).
Briefly, the suspension samples were fixed with
2.5% (vol/vol) glutaraldehyde, pipette-drawn,
mounted on 0.22 mm membrane filter, washed and
fixed with t-butyl alcohol, subsequently frozen in
liquid nitrogen, and dried up with low-vacuum
SEM. After freeze-drying completely, samples were
transferred to the brass-stub with double-sided
adhesive carbon tape, coated with carbon, and
then observed by using a scanning electron micro-
scope (JEOL JSM-5200 LV). For TEM observation,
samples were fixed with 2.5% v/v glutaraldehyde
for 2 h at 4ëC, mounted on copper specimen grids,
allowed to dehydrate at room temperature, and then
viewed using a JEOL JEM-2000EX transmission
electron microscope.
Analytical techniques
The pH in solution was monitored using a pH
meter (Horiba) at set time intervals. To determine
the total cells in the biofilm and planktonic milieu,
1 ml of the homogenized solution-containing cells
(free-living and attached to clay surfaces) was
placed in vials, sonicated at 4ëC for 3 min and
vortexed for 10 s to disperse the microorganisms
attached to clay surfaces. The resuspended biofilm
or planktonic sample was serially diluted with fresh
minimal salts media, and plated in nutrient agar
(NA) plates supplemented with 1 g/l yeast extract
and 0.85% (w/v) NaCl, then incubated at 25ëC for 3
to 5 days. For statistical purposes, sample analyses
were performed in triplicate. The data are presented
as the arithmetic mean Ô standard deviation of the
mean.
RESULTS AND DISCUSS ION
Microbial growth
During the first 36 days of incubation, there was
no significant increase in the number of microbial
TABLE 2. The chemical composition of heavy oil
collected from the Nakhodka oil spill that was used as
the sole carbon and energy source of microorganisms
analysed by NCS analyser.
Chemical composition of heavy oil Wt.%
Carbon 82.9
Sulphur 1.2
Nitrogen 0.22
Clay minerals and oil-utilizing indigenous microorganisms 107
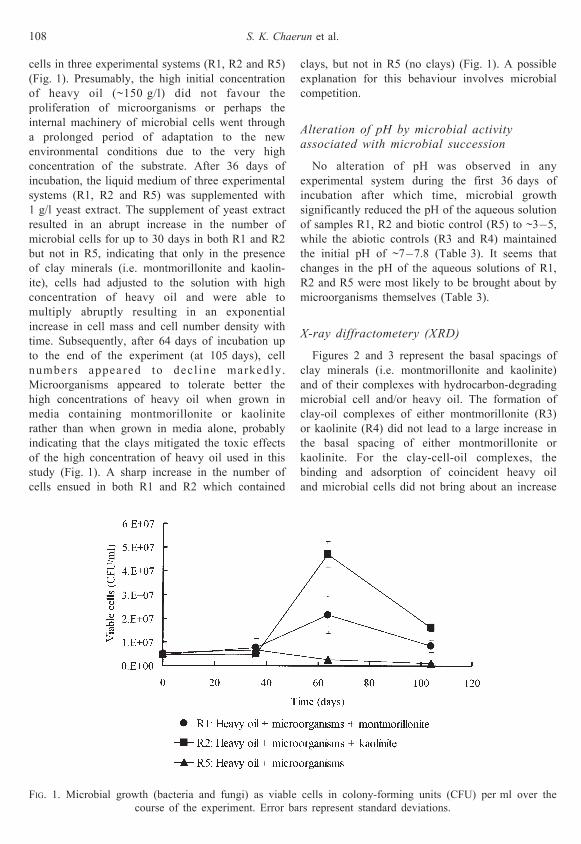
cells in three experimental systems (R1, R2 and R5)
(Fig. 1). Presumably, the high initial concentration
of heavy oil (~150 g/l) did not favour the
proliferation of microorganisms or perhaps the
internal machinery of microbial cells went through
a prolonged period of adaptation to the new
environmental conditions due to the very high
concentration of the substrate. After 36 days of
incubation, the liquid medium of three experimental
systems (R1, R2 and R5) was supplemented with
1 g/l yeast extract. The supplement of yeast extract
resulted in an abrupt increase in the number of
microbial cells for up to 30 days in both R1 and R2
but not in R5, indicating that only in the presence
of clay minerals (i.e. montmorillonite and kaolin-
ite), cells had adjusted to the solution with high
concentration of heavy oil and were able to
multiply abruptly resulting in an exponential
increase in cell mass and cell number density with
time. Subsequently, after 64 days of incubation up
to the end of the experiment (at 105 days), cell
numbers appeared to decl ine markedly.
Microorganisms appeared to tolerate better the
high concentrations of heavy oil when grown in
media containing montmorillonite or kaolinite
rather than when grown in media alone, probably
indicating that the clays mitigated the toxic effects
of the high concentration of heavy oil used in this
study (Fig. 1). A sharp increase in the number of
cells ensued in both R1 and R2 which contained
clays, but not in R5 (no clays) (Fig. 1). A possible
explanation for this behaviour involves microbial
competition.
Alteration of pH by microbial activity
associated with microbial succession
No alteration of pH was observed in any
experimental system during the first 36 days of
incubation after which time, microbial growth
significantly reduced the pH of the aqueous solution
of samples R1, R2 and biotic control (R5) to ~3ÿ5,
while the abiotic controls (R3 and R4) maintained
the initial pH of ~7ÿ7.8 (Table 3). It seems that
changes in the pH of the aqueous solutions of R1,
R2 and R5 were most likely to be brought about by
microorganisms themselves (Table 3).
X-ray diffractometery (XRD)
Figures 2 and 3 represent the basal spacings of
clay minerals (i.e. montmorillonite and kaolinite)
and of their complexes with hydrocarbon-degrading
microbial cell and/or heavy oil. The formation of
clay-oil complexes of either montmorillonite (R3)
or kaolinite (R4) did not lead to a large increase in
the basal spacing of either montmorillonite or
kaolinite. For the clay-cell-oil complexes, the
binding and adsorption of coincident heavy oil
and microbial cells did not bring about an increase
FIG. 1. Microbial growth (bacteria and fungi) as viable cells in colony-forming units (CFU) per ml over the
course of the experiment. Error bars represent standard deviations.
108 S. K. Chaerun et al.
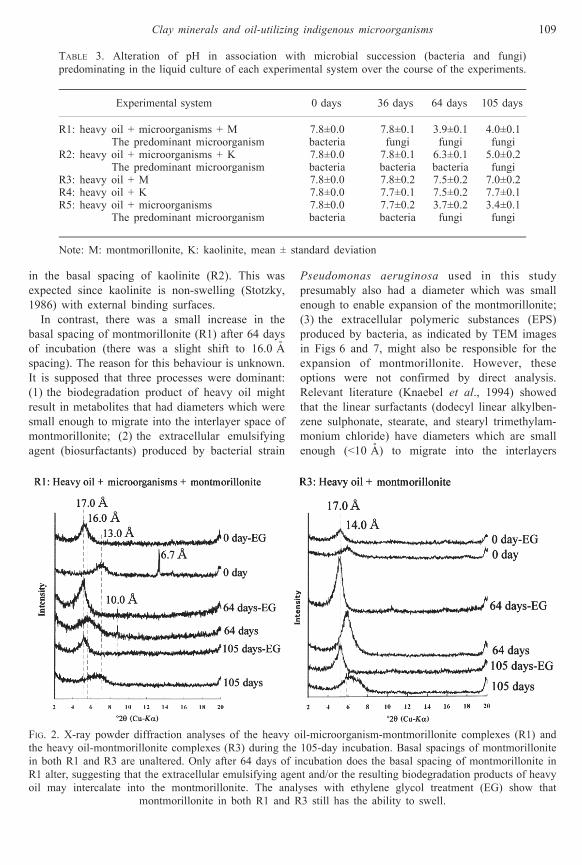
in the basal spacing of kaolinite (R2). This was
expected since kaolinite is non-swelling (Stotzky,
1986) with external binding surfaces.
In contrast, there was a small increase in the
basal spacing of montmorillonite (R1) after 64 days
of incubation (there was a slight shift to 16.0 AÊ
spacing). The reason for this behaviour is unknown.
It is supposed that three processes were dominant:
(1) the biodegradation product of heavy oil might
result in metabolites that had diameters which were
small enough to migrate into the interlayer space of
montmorillonite; (2) the extracellular emulsifying
agent (biosurfactants) produced by bacterial strain
Pseudomonas aeruginosa used in this study
presumably also had a diameter which was small
enough to enable expansion of the montmorillonite;
(3) the extracellular polymeric substances (EPS)
produced by bacteria, as indicated by TEM images
in Figs 6 and 7, might also be responsible for the
expansion of montmorillonite. However, these
options were not confirmed by direct analysis.
Relevant literature (Knaebel et al., 1994) showed
that the linear surfactants (dodecyl linear alkylben-
zene sulphonate, stearate, and stearyl trimethylam-
monium chloride) have diameters which are small
enough (<10 AÊ ) to migrate into the interlayers
TABLE 3. Alteration of pH in association with microbial succession (bacteria and fungi)
predominating in the liquid culture of each experimental system over the course of the experiments.
Experimental system 0 days 36 days 64 days 105 days
R1: heavy oil + microorganisms + M 7.8Ô0.0 7.8Ô0.1 3.9Ô0.1 4.0Ô0.1
The predominant microorganism bacteria fungi fungi fungi
R2: heavy oil + microorganisms + K 7.8Ô0.0 7.8Ô0.1 6.3Ô0.1 5.0Ô0.2
The predominant microorganism bacteria bacteria bacteria fungi
R3: heavy oil + M 7.8Ô0.0 7.8Ô0.2 7.5Ô0.2 7.0Ô0.2
R4: heavy oil + K 7.8Ô0.0 7.7Ô0.1 7.5Ô0.2 7.7Ô0.1
R5: heavy oil + microorganisms 7.8Ô0.0 7.7Ô0.2 3.7Ô0.2 3.4Ô0.1
The predominant microorganism bacteria bacteria fungi fungi
Note: M: montmorillonite, K: kaolinite, mean Ô standard deviation
FIG. 2. X-ray powder diffraction analyses of the heavy oil-microorganism-montmorillonite complexes (R1) and
the heavy oil-montmorillonite complexes (R3) during the 105-day incubation. Basal spacings of montmorillonite
in both R1 and R3 are unaltered. Only after 64 days of incubation does the basal spacing of montmorillonite in
R1 alter, suggesting that the extracellular emulsifying agent and/or the resulting biodegradation products of heavy
oil may intercalate into the montmorillonite. The analyses with ethylene glycol treatment (EG) show that
montmorillonite in both R1 and R3 still has the ability to swell.
Clay minerals and oil-utilizing indigenous microorganisms 109
(10 to 20 AÊ ) of montmorillonite. Harter and Stotzky
(1973) also reported that several forms of the
protein intercalated the H- and Na-smectites which
caused an expansion to 16.4 AÊ . Pepsin also caused
a slight expansion of air-dried Na-smectite to
15.2 AÊ (Harter & Stotzky, 1971).
Furthermore, the XRD spectra of the intermediate
complex (i.e. montmorillonite-oil-cell complex) at
105 days tended to have somewhat more irregular
and broader peaks than those in the initial complex
(0 days), indicating that some metabolic products
mentioned above might remain to intercalate the
montmorillonite. In addition, the analyses with
ethylene glycol treatment showed that the expand-
ability of montmorillonite in both R1 and R3 was
retained throughout the experiments.
Scanning electron microscopy
Figures 4 and 5 present the images of the unique
or attractive dense network of microbial cells and
several clusters of clay particles (R1 and R2) after
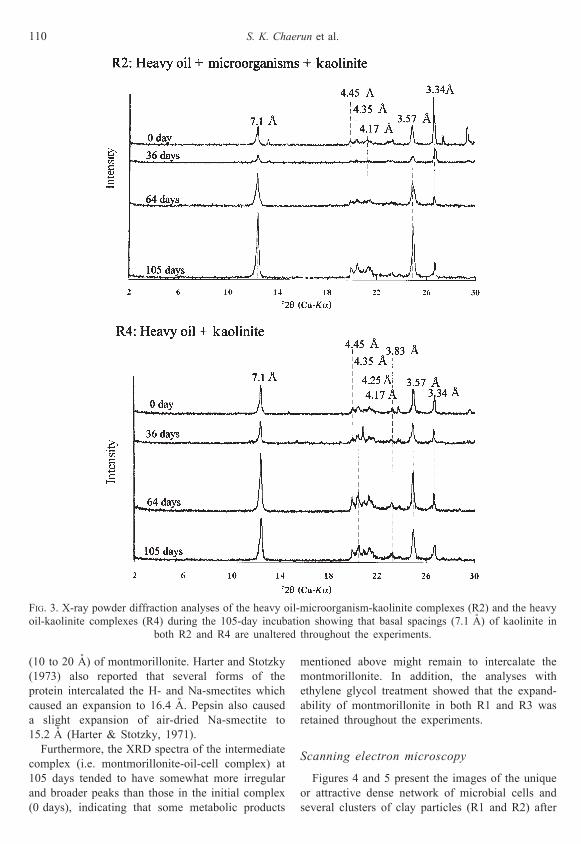
FIG. 3. X-ray powder diffraction analyses of the heavy oil-microorganism-kaolinite complexes (R2) and the heavy
oil-kaolinite complexes (R4) during the 105-day incubation showing that basal spacings (7.1 AÊ ) of kaolinite in
both R2 and R4 are unaltered throughout the experiments.
110 S. K. Chaerun et al.
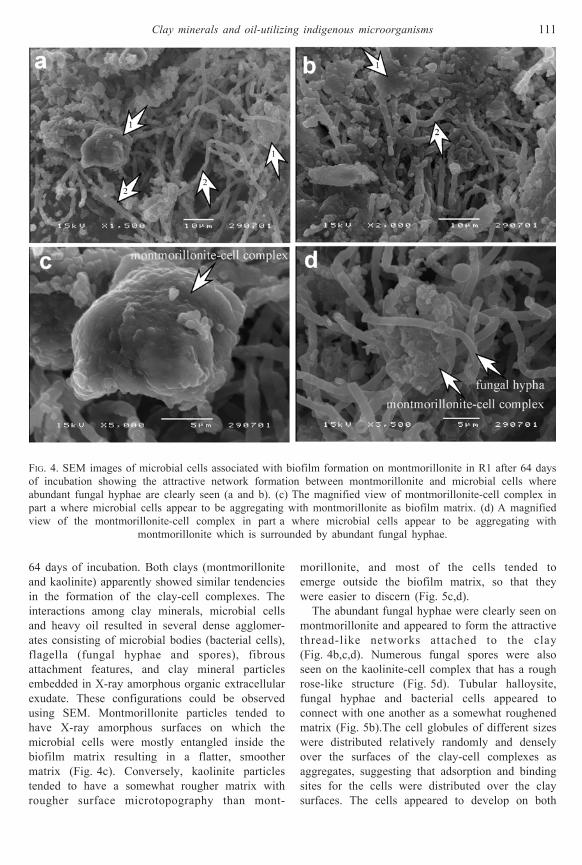
64 days of incubation. Both clays (montmorillonite
and kaolinite) apparently showed similar tendencies
in the formation of the clay-cell complexes. The
interactions among clay minerals, microbial cells
and heavy oil resulted in several dense agglomer-
ates consisting of microbial bodies (bacterial cells),
flagella (fungal hyphae and spores), fibrous
attachment features, and clay mineral particles
embedded in X-ray amorphous organic extracellular
exudate. These configurations could be observed
using SEM. Montmorillonite particles tended to
have X-ray amorphous surfaces on which the
microbial cells were mostly entangled inside the
biofilm matrix resulting in a flatter, smoother
matrix (Fig. 4c). Conversely, kaolinite particles
tended to have a somewhat rougher matrix with
rougher surface microtopography than mont-
morillonite, and most of the cells tended to
emerge outside the biofilm matrix, so that they
were easier to discern (Fig. 5c,d).
The abundant fungal hyphae were clearly seen on
montmorillonite and appeared to form the attractive
thread-like networks attached to the clay
(Fig. 4b,c,d). Numerous fungal spores were also
seen on the kaolinite-cell complex that has a rough
rose-like structure (Fig. 5d). Tubular halloysite,
fungal hyphae and bacterial cells appeared to
connect with one another as a somewhat roughened
matrix (Fig. 5b).The cell globules of different sizes
were distributed relatively randomly and densely
over the surfaces of the clay-cell complexes as
aggregates, suggesting that adsorption and binding
sites for the cells were distributed over the clay
surfaces. The cells appeared to develop on both
FIG. 4. SEM images of microbial cells associated with biofilm formation on montmorillonite in R1 after 64 days
of incubation showing the attractive network formation between montmorillonite and microbial cells where
abundant fungal hyphae are clearly seen (a and b). (c) The magnified view of montmorillonite-cell complex in
part a where microbial cells appear to be aggregating with montmorillonite as biofilm matrix. (d) A magnified
view of the montmorillonite-cell complex in part a where microbial cells appear to be aggregating with
montmorillonite which is surrounded by abundant fungal hyphae.
Clay minerals and oil-utilizing indigenous microorganisms 111

clays as biofilm matrix, indicating that the clays
acted as materials supporting their growth.
Observation by SEM showed that both clays
served as growth-supporting materials supporting
the data on microbial growth given in Fig. 1. The
results obtained here confirmed our previous results
that clay minerals such as bentonite and kaolinite
could have a role as microbial growth-support
media (Chaerun et al., 2003).
Transmission electron microscopy
The TEM images show more clearly than SEM
images the binding and adsorption of the bacterial
cells on the clays (Figs 6, 7). The unique and
attractive network formation of bacterial cells was
clearly seen as biofilm formation, consisting of
bacterial colonies embedded in extracellular poly-
meric substances (EPS) and appearing to attach
firmly to the clays. The cells appeared to colonize
clay surfaces, aggregate and/or grow into multi-
cellular colonies, and embed themselves in EPS
matrix (Figs 6a, 7a,b). The matrix of biofilms is a
complex mixture consisting mainly of exopolysac-
charides, protein and nucleic acids (Sutherland,
2001), and bacteria residing within biofilms
probably play a key role in the degradation of
organic pollutants (Davey & O'Toole, 2000).
Furthermore, the cells appeared to be predomi-
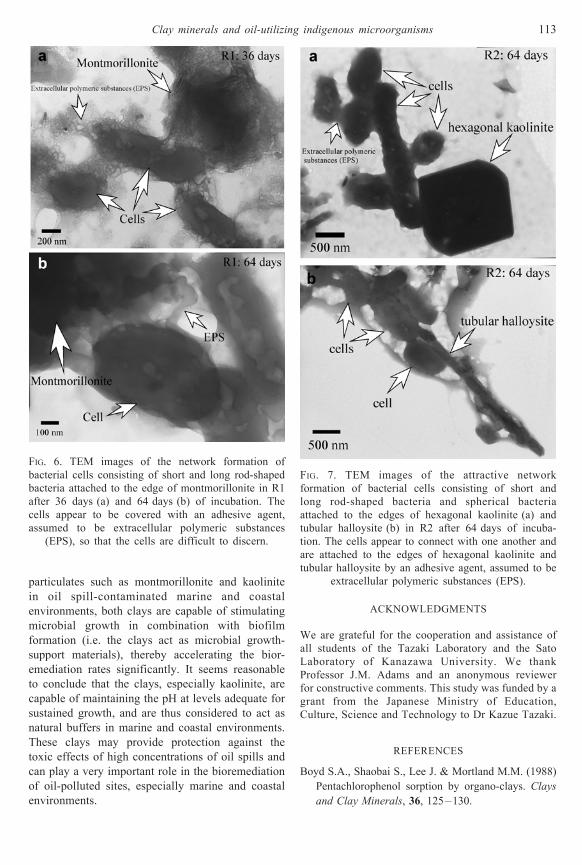
nantly bound on the edges of the clays.
Montmorillonite appeared as a dispersed flake-like
shape (Fig. 6a), whereas kaolinite displayed tube-like
(tubular halloysite) and hexagonal forms (Fig. 7a,b).
CONCLUS ION
The results show that when microorganisms are
associated with naturally occurring inorganic
FIG. 5. SEM images of microbial cells associated with biofilm formation on kaolinite in R2 after 64 days of
incubation showing the network formation between kaolinite and microbial cells. (a and b) The features of
tubular halloysite and kaolinite in aggregation with bacterial cells and the fungal hyphae and spores. (c and
d) The kaolinite-cell complexes showing the development of fungal-bacterial cells on kaolinite as biofilm matrix.
112 S. K. Chaerun et al.
particulates such as montmorillonite and kaolinite
in oil spill-contaminated marine and coastal
environments, both clays are capable of stimulating
microbial growth in combination with biofilm
formation (i.e. the clays act as microbial growth-
support materials), thereby accelerating the bior-
emediation rates significantly. It seems reasonable
to conclude that the clays, especially kaolinite, are
capable of maintaining the pH at levels adequate for
sustained growth, and are thus considered to act as
natural buffers in marine and coastal environments.
These clays may provide protection against the
toxic effects of high concentrations of oil spills and
can play a very important role in the bioremediation
of oil-polluted sites, especially marine and coastal
environments.
ACKNOWLEDGMENTS
We are grateful for the cooperation and assistance of
all students of the Tazaki Laboratory and the Sato
Laboratory of Kanazawa University. We thank
Professor J.M. Adams and an anonymous reviewer
for constructive comments. This study was funded by a
grant from the Japanese Ministry of Education,
Culture, Science and Technology to Dr Kazue Tazaki.
REFERENCES
Boyd S.A., Shaobai S., Lee J. & Mortland M.M. (1988)
Pentachlorophenol sorption by organo-clays. Clays
and Clay Minerals, 36, 125ÿ130.
FIG. 6. TEM images of the network formation of
bacterial cells consisting of short and long rod-shaped
bacteria attached to the edge of montmorillonite in R1
after 36 days (a) and 64 days (b) of incubation. The
cells appear to be covered with an adhesive agent,
assumed to be extracellular polymeric substances
(EPS), so that the cells are difficult to discern.
FIG. 7. TEM images of the attractive network
formation of bacterial cells consisting of short and
long rod-shaped bacteria and spherical bacteria
attached to the edges of hexagonal kaolinite (a) and
tubular halloysite (b) in R2 after 64 days of incuba-
tion. The cells appear to connect with one another and
are attached to the edges of hexagonal kaolinite and
tubular halloysite by an adhesive agent, assumed to be
extracellular polymeric substances (EPS).
Clay minerals and oil-utilizing indigenous microorganisms 113

Chaerun S.K. & Tazaki K. (2003) Hydrocarbon-
degrading bacteria in the heavy oil polluted soil
and seawater after 5 years bioremediation. Pp. 187ÿ
204 in : Water and Soi l Env i ronments :
Microorganisms Play an Important Role (K.
Tazaki, editor). Kanazawa University Press, 21st
Century COE Kanazawa University, Japan.
Chaerun S.K., Tazaki K. & Asada R. (2003) Double
function of bentonite and kaolinite as adsorbents and
`microbial growth-support media' for degradation of
crude oil. Pp. 253ÿ277 in: Heavy Oil Spilled from
Russian Tanker `Nakhodka' in 1997: Towards Eco-
responsibility, Earth Sense (K. Tazaki, editor).
Kanazawa University Press, 21st
Century COE
Kanazawa University, Japan.
Davey M.E. & O'Toole G.A. (2000) Microbial biofilms:
from ecology to molecular genetics. Microbiology
and Molecular Biology Reviews, 64, 847ÿ867.
Guerin W.F. & Boyd S.A. (1992) Differential bioavail-
ability of soil-sorbed naphthalene to two bacterial
species. Applied and Environmental Microbiology,
58, 1142ÿ1152.
Harter R.D. & Stotzky G. (1971) Formation of clay-
protein complexes. Soil Science Society of America
Proceedings, 35, 383ÿ389.
Harter R.D. & Stotzky G. (1973) X-ray diffraction,
electron microscopy, electrophoretic mobility, and
pH of some stable smectite-protein complexes. Soil
Science Society of America Proceedings, 37,
116ÿ123.
Khanna M. & Stotzky G. (1992) Transformation of
Bacillus subtilis by DNA bound on montmorillonite
and effect of DNase on the transforming ability of
bound DNA. Applied and Environmental
Microbiology, 58, 1930ÿ1939.
Knaebel D.B., Federle T.W., McAvoy D.C. & Vestal
J.R. (1994) Effect of mineral and organic soil
constituents on microbial mineralization of organic
compounds in a natural soil. Applied and
Environmental Microbiology, 60, 4500ÿ4508.
Lee J., Mortland M.M., Chiou C.T., Kile D.E. & Boyd
S.A. (1990) Adsorption of benzene, toluene, and
xylene by two tetramethylammonium-smectites hav-
ing different charge densities. Clays and Clay
Minerals, 38, 113ÿ120.
Lin Z. & Puls R.W. (2000) Adsorption, desorption and
oxidation of arsenic affected by clay minerals and
aging process. Environmental Geology, 39, 753ÿ759.
Lipson S.M. & Stotzky G. (1983) Adsorption of reovirus
to clay minerals: effects of cation-exchange capacity,
cation saturation, and surface area. Applied and
Environmental Microbiology, 46, 673ÿ682.
Lipson S.M. & Stotzky G. (1984) Effect of proteins on
adsorption to clay minerals. Applied and
Environmental Microbiology, 48, 525ÿ530.
Miller M.E. & Alexander M. (1991) Kinetics of
bacterial degradation of belzylamine in a montmor-
illonite suspension. Environmental Science and
Technology, 25, 240ÿ245.
Ogram A.V., Jessup R.E., Ou L.T. & Rao P.S.C. (1985)
Effects of sorption on biological biodegradation rates
of (2, 4-dichlorophenoxy) acetic acid in soils. Applied
and Environmental Microbiology, 49, 582ÿ587.
Ortega-Calvo J.J. & Saiz-Jimenez C. (1998) Effect of
humic fractions and clay on biodegradation of
phenanthrene by a Pseudomonas fluorescens strain
isolated from soil. Applied and Environmental
Microbiology, 64, 3123ÿ3126.
Robinson K.G., Farmer W.S. & Novak J.T. (1990)
Availability of sorbed toluene in soils for degradation
of acclimated bacteria.Water Resources, 24, 345ÿ350.
Schiffenbauer M. & Stotzky G. (1982) Adsorption of
Coliphages T1 and T7 to clay minerals. Applied and
Environmental Microbiology, 43, 590ÿ596.
Scow K.M. & Alexander M. (1992) Effect of diffusion
on the kinetics of biodegradation: experimental
results with synthetic aggregates. Soil Science
Society of America Journal, 56, 128ÿ134.
Shuler M.L. & Kargi F. (1992) Bioprocess Engineering
Basic Concepts, pp. 61ÿ78. Prentice Hall
International, New Jersey, USA.
Stotzky G. (1986) Influence of soil minerals colloids on
metabolic processes, growth, adhesion, and ecology
of microbes and viruses. Pp. 305ÿ428 in:
Interactions of Soil Minerals with Natural
Organics and Microbes (P.M. Huang and M.
Schnitzer, editors). Soil Science Society of
America, Madison, Wisconsin, USA.
Sutherland I.W. (2001) The biofilm matrix ÿ an
immobilized but dynamic microbial environment.
Trends in Microbiology, 9, 222ÿ227.
Suzuki T., Shibata M., Tanaka K., Tsuchida K. & Toda
T. (1995) A new drying method: low vacuum SEM
freeze drying and its application to plankton
observation. Bulletin of the Planktonic Society of
Japan, 42, 53ÿ62.
Tapp H. & Stotzky G. (1995) Insecticidal activity of the
toxins from Bacillus thuringiensis subspecies kur-
staki and tenebrionis adsorbed and bound on pure
and soil clays. Applied and Environmental
Microbiology, 61, 1786ÿ1790.
Tazaki K. (editor) (2003) Heavy Oil Spilled from Russian
Tanker `Nakhodka' in 1997: Towards Eco-responsi-
bility, Earth Sense. Kanazawa University Press, 21st
Century COE Kanazawa University, Japan.
Vettori C., Calamai L., Yoder M., Stotzky G. & Gallori
E. (1999) Adsorption and binding of AmpliTaq1
DNA polymerase on the clay minerals, montmor-
illonite and kaolinite. Soil Biology and Biochemistry,
31, 587ÿ593.
Weber J.B. & Coble H.D. (1968) Microbial decomposi-
tion of diquat adsorbed on montmorillonite and
kaolinite clays. Journal of Agricultural and Food
Chemistry, 16, 475ÿ478.
114 S. K. Chaerun et al.
Copyright © 2022 FDOKUMEN




















