
Inter-population thermal variability and physiological response in the intertidal fish Scartichthys...
10
ECOPHYSIOLOGY Jose M. Pulgar Francisco Bozinovic F. Patricio Ojeda Local distribution and thermal ecology of two intertidal fishes Received: 26 July 2003 / Accepted: 7 October 2004 / Published online: 24 December 2004 ȑ Springer-Verlag 2004 Abstract Geographic variability in the physiological attributes of widely distributed species can be a result of phenotypic plasticity or can reflect evolutionary re- sponses to a particular habitat. In the field, we assessed thermal variability in low and high intertidal pools and the distribution of resident fish species Scartichthys vir- idis and transitory Girella laevifrons along this vertical intertidal gradient at three localities along the Chilean coast: Antofagasta (the northernmost and warmest habitat), Carrizal Bajo (central coast) and Las Cruces (the southernmost and coldest habitat). In the labora- tory, we evaluated the thermal sensitivity of fish cap- tured from each locality. The response to temperature was estimated as the frequency of opercular movements and as thermal selectivity in a gradient; the former being a indirect indicator of energy costs in a particular envi- ronment and the latter revealing differential occupation of habitat. Seawater temperature in intertidal pools was greatest at Antofagasta, and within each site was greatest in high intertidal pools. The two intertidal fish species showed opposite patterns of local distribution, with S. viridis primarily inhabiting the lower sectors of the intertidal zone, and G. laevifrons occupying the higher sectors of the intertidal zone. This pattern was consistent for all three localities. Locality was found to be a very important factor determining the frequency of opercular movement and thermal selectivity of both S. viridis and G. laevifrons. Our results suggest that S. vir- idis and G. laevifrons respond according to: (1) the thermal history of the habitat from which they came, and (2) the immediate physical conditions of their hab- itat. These results suggest local adaptation rather than plasticity in thermoregulatory and energetic mecha- nisms. Keywords Physiological diversity Intertidal fish Thermal sensitivity Rock pool Introduction Physiological constraints are important determinants of the distributional limits of species and populations (Bozinovic and Rosenmann 1989; Bozinovic et al. 1995; Gaston and Spicer 1998; Chown and Gaston 2000); however, processes associated with environmental tol- erance explaining, at the local scale, differential habitat use or, at the geographical scale, species distribution patterns remain poorly understood. Indeed, latitude, altitude and depth all constitute gradients, which can generate physiological diversity in the responses of individuals and populations (Spicer and Gaston 1999). Among populations, variation in physiological traits can be environmentally induced, through non-genetic, reversible mechanisms of phenotypic flexibility or accli- matization (Piersma and Drent 2003), or can reflect genetic adaptation to local conditions, in which case differences among organisms from different habitats cannot be removed by common garden experiments (Stearns 1982; Spicer and Gaston 1999). Among vertebrate ectotherms, variation in physical factors, particularly temperature, has been documented as one of the most important determinants of distribu- tion (Norris 1963; Spicer and Gaston 1999), affecting all biological processes and life history traits, such as body size (Tracy and Christian 1986; Cossins and Bowler 1987; Weise 1991; Lenski and Bennett 1993; Parsons 1993; Stephen and Porter 1993; Pulgar et al. 1999; Angilletta et al. 2002). Among marine animals with high dispersal capacity, latitudinal genetic differentiation of J. M. Pulgar (&) F. Bozinovic F. P. Ojeda Center for Advanced Studies in Ecology and Biodiversity and Departamento de Ecologı´a, Pontificia Universidad Cato´lica de Chile, Santiago, 6513677, Chile E-mail: [email protected] Tel.: +56-2-6862797 Fax: +56-2-6862621 Present address: J. M. Pulgar Departamento de Ciencias de la Salud, Universidad Andre´s Bello, Santiago, Chile Oecologia (2005) 142: 511–520 DOI 10.1007/s00442-004-1755-4
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Inter-population thermal variability and physiological response in the intertidal fish Scartichthys...

ECOPHYSIOLOGY
Jose M. Pulgar Æ Francisco Bozinovic
F. Patricio Ojeda
Local distribution and thermal ecologyof two intertidal fishes
Received: 26 July 2003 / Accepted: 7 October 2004 / Published online: 24 December 2004� Springer-Verlag 2004
Abstract Geographic variability in the physiologicalattributes of widely distributed species can be a result ofphenotypic plasticity or can reflect evolutionary re-sponses to a particular habitat. In the field, we assessedthermal variability in low and high intertidal pools andthe distribution of resident fish species Scartichthys vir-idis and transitory Girella laevifrons along this verticalintertidal gradient at three localities along the Chileancoast: Antofagasta (the northernmost and warmesthabitat), Carrizal Bajo (central coast) and Las Cruces(the southernmost and coldest habitat). In the labora-tory, we evaluated the thermal sensitivity of fish cap-tured from each locality. The response to temperaturewas estimated as the frequency of opercular movementsand as thermal selectivity in a gradient; the former beinga indirect indicator of energy costs in a particular envi-ronment and the latter revealing differential occupationof habitat. Seawater temperature in intertidal pools wasgreatest at Antofagasta, and within each site wasgreatest in high intertidal pools. The two intertidal fishspecies showed opposite patterns of local distribution,with S. viridis primarily inhabiting the lower sectors ofthe intertidal zone, and G. laevifrons occupying thehigher sectors of the intertidal zone. This pattern wasconsistent for all three localities. Locality was found tobe a very important factor determining the frequency ofopercular movement and thermal selectivity of both S.viridis and G. laevifrons. Our results suggest that S. vir-idis and G. laevifrons respond according to: (1) thethermal history of the habitat from which they came,
and (2) the immediate physical conditions of their hab-itat. These results suggest local adaptation rather thanplasticity in thermoregulatory and energetic mecha-nisms.
Keywords Physiological diversity Æ Intertidal fish ÆThermal sensitivity Æ Rock pool
Introduction
Physiological constraints are important determinants ofthe distributional limits of species and populations(Bozinovic and Rosenmann 1989; Bozinovic et al. 1995;Gaston and Spicer 1998; Chown and Gaston 2000);however, processes associated with environmental tol-erance explaining, at the local scale, differential habitatuse or, at the geographical scale, species distributionpatterns remain poorly understood. Indeed, latitude,altitude and depth all constitute gradients, which cangenerate physiological diversity in the responses ofindividuals and populations (Spicer and Gaston 1999).Among populations, variation in physiological traits canbe environmentally induced, through non-genetic,reversible mechanisms of phenotypic flexibility or accli-matization (Piersma and Drent 2003), or can reflectgenetic adaptation to local conditions, in which casedifferences among organisms from different habitatscannot be removed by common garden experiments(Stearns 1982; Spicer and Gaston 1999).
Among vertebrate ectotherms, variation in physicalfactors, particularly temperature, has been documentedas one of the most important determinants of distribu-tion (Norris 1963; Spicer and Gaston 1999), affecting allbiological processes and life history traits, such as bodysize (Tracy and Christian 1986; Cossins and Bowler1987; Weise 1991; Lenski and Bennett 1993; Parsons1993; Stephen and Porter 1993; Pulgar et al. 1999;Angilletta et al. 2002). Among marine animals with highdispersal capacity, latitudinal genetic differentiation of
J. M. Pulgar (&) Æ F. Bozinovic Æ F. P. OjedaCenter for Advanced Studies in Ecology and Biodiversityand Departamento de Ecologıa,Pontificia Universidad Catolica de Chile,Santiago, 6513677, ChileE-mail: [email protected].: +56-2-6862797Fax: +56-2-6862621
Present address: J. M. PulgarDepartamento de Ciencias de la Salud,Universidad Andres Bello, Santiago, Chile
Oecologia (2005) 142: 511–520DOI 10.1007/s00442-004-1755-4

populations has occurred as an adaptation to localconditions, in spite of the absence of obvious barriers togene flow (Lonsdale and Levinton 1985). Latitudinalphysical variation may generate selective pressures thatresult in thermal tolerance, depending on the geographicorigin of the populations (Spicer and Gaston 1999;Morritt and Ingolfsson 2000).
Intertidal rock pools are characterized by their dis-crete nature in time and space, as well as by their dailyand seasonal variability in temperature, oxygen con-centration and salinity (Newell 1970; Stephenson andStephenson 1972; Truchot and Duhanel-Jouve 1980;Metaxas and Scheibling 1993). These variations, par-ticularly in temperature, depend on the position of therock pool along the intertidal vertical gradient, whichdetermines the length of time that a rock pool remainsisolated from the subtidal system (Gibson 1972, 1982;Horn and Gibson 1988; Horn et al. 1999). Intertidal fishassemblages are composed of resident and transitoryspecies (Horn et al. 1999). Gibson (1982) and Horn andGibson (1988) predicted that intertidal fish zonationpatterns should be similar to those observed amongsessile individuals, being principally associated withphysical conditions (Zander et al. 1999). In this sense,Thompson and Lehner (1976) determined that residentfish species usually occupy sectors of the intertidal zoneclosest to the subtidal zone. These fishes should occupythe lowest sectors of the intertidal zone, and, thus, thehabitats that are the least variable in terms of physicalconditions. Whereas transient fish usually occupy sec-tors most distal from the subtidal zone, which are themost variable in terms of physical conditions
In spite of the fact that intertidal fish present anexcellent study model for research on thermal physiol-ogy, most work on fish has centred on trophic ecology,the identification of guilds, differential habitat utilizationand assemblage persistence and stability (Gosline1966; Thompson and Lehner 1976; Yoshiyama 1980,1981; Gibson 1982; Grossman 1982; Bennett et al. 1983;Bennett and Griffitths 1984, Collette 1986; Grossman1986; Ralston and Horn 1986; Bennett 1987; Varas andOjeda 1990; Prochazca and Griffitths 1987; Munoz andOjeda 1997, 1998; Aldana et al. 2002). Information onthe thermal sensitivity of intertidal fish is scarce. Gra-ham (1970) predicted that a fish’s geographic originwould determine its thermal tolerance and distributionalong intertidal vertical distribution gradients. Davis(1977) indicated that in the fishes of the genus Gibbonsia,those with the greatest capacity for adaptation to hightemperatures had the widest geographic distributions.Bridges (1993), Metaxas and Scheibling (1993) andDavis (2000) established that seasonal variation in theabundance of intertidal fish was mainly associated withseawater temperature and specific microhabitats. Fur-thermore, Pulgar et al. (1999) demonstrated that star-vation did not modify the thermal selectivity of Girellalaevifrons, and Hernandez et al. (2002) reported thatsmall-sized individuals of Graus nigra had a greatertolerance to thermal stress than larger sized individuals,
a response that may explain their differential use of theintertidal vertical gradient. Thus, the distribution andabundance of intertidal fish appear to be stronglyinfluenced by physical habitat conditions, especiallytemperature of the tidal pool (Davis 1977; Zander et al.1999).
Scartichthys viridis (Blenniidae) and G. laevifrons(Kyphosidae) (Nelson 1994), are the most abundant fishspecies inhabiting the rocky intertidal zone along theChilean coast (Stepien 1990; Varas and Ojeda 1990).S. viridis is a permanent resident of the intertidal zone,with the exception of its planktonic larval stage (Varasand Ojeda 1990). G. laevifrons is a transitory speciessince it inhabits the intertidal zone only during thejuvenile stage, later migrating to the subtidal zone(Varas and Ojeda 1990; Munoz and Ojeda 1997). Giventhe wide geographic distribution of both fish species,extending from 18 to 40�S along the Chilean coast(Ojeda et al. 2000), we predicted that populationsinhabiting separate geographic localities would be sub-jected to strong variation in tidal pool temperature, andthis might have generated more thermally sensitivepopulations at higher latitudes than at lower latitudes,and in those environments where the thermal extremesare more marked (low latitudes) fishes inhabiting lowintertidal areas (i.e. thermally more stable) would notoccur in the high intertidal zone (i.e. thermally morevariable). We also predict that in the southern localitiesboth fish groups could occupy the entire intertidal zone.Therefore, the objectives of this work were: (1) to recordthermal variations in tidal pools of the low and highintertidal zones at three localities along the Chileancoast (Antofagasta and Carrizal Bajo in northern Chile,and Las Cruces in central Chile, each separated by ca. 5�latitude), (2) to characterize the intertidal vertical dis-tribution of S. viridis and G. laevifrons at each locality,and (3) to evaluate the thermal sensitivity of S. viridisand G. laevifrons under laboratory conditions. Thermalsensitivity was estimated using two measurements: (1)the frequency of opercular movements as a function ofincreasing temperature, and (2) thermal selectivitywithin a thermal gradient. The frequency of opercularmovements has been directly related to energy expen-diture (see Roger and Weatherley 1993; Van Rooij andVideler 1996; Granterner and Taborsky 1998), whilethermal selectivity reveals differential occupation of thehabitat (Pulgar et al. 1999; Hernandez et al. 2002).
Materials and methods
Thermal variability of the intertidal system
To characterize thermal heterogeneity along the verticalintertidal gradient we placed a thermal logger (TIDBIT;Unisource) in one low and one high intertidal pool ateach of the study locations: Antofagasta (23�24¢S,70�35¢W), Carrizal Bajo (28�04¢S, 70�35¢W), and LasCruces (33�26¢S, 71�41¢W). Seawater temperature in the
512

pools was recorded at half-hour intervals between June2001 (austral winter) and January 2002 (austral sum-mer). The selected low intertidal rock pools were locatedno more than 1–2 m above the low tide mark. Thus, theseawater of these pools was renewed during every tidalcycle. The high intertidal pools were located at least50 m above the breaking waves during low tide and,thus, the seawater in these pools was not renewed duringmost tidal cycles. At each locality, we registered maxi-mum, minimum and average daily water temperatures inboth low and high intertidal pools.
At each geographic locality, specimens were capturedfrom tidepools between June 2001 and January 2002,from high and low intertidal rock pools at three locali-ties along the Chilean coast: Antofagasta (23�20¢S,70�38¢W), Carrizal Bajo (28�04¢S, 71�08¢W) and LasCruces (33�26¢S, 71�41¢W). Specimens were transportedto the University Marine Season at Las Cruces (CoastalMarine Research Station) where they were maintainedunder laboratory conditions, with constantly circulatedfiltered sea water, constant aeration and daily feedingad libitum.
Frequency of opercular movementsin resident S. viridis and transitory G. laevifrons
In the laboratory,we evaluated the frequency of opercularmovement in response to increasing seawater temperature(from 13 to 30�C at a rate of 0.1�C/min over a 2-h period)for fish collected from the three different localities. Inthese tests we used six similarly sized fish from eachlocality [i.e. total length (Lt)=5 cm for G. laevifrons andLt=7 cm forS. viridis]. Fish were placed one per trial, in a12·18·9-cm aquarium, which was submerged in a larger22.5-l (30·30·25 cm) tank. The aquarium contained fil-tered seawater and was equipped with constant aerationand illumination. This design allowed frontal filmingduring 2-h periods, using a Sony Handycam 8 XR videocamera. All experimental assays began at 1200 hours andwere preceded by a 1-h acclimation period in the boxes, atconstant water temperature (13�C). Subsequently, weobserved the recordings and counted the number ofopercular movements per individual per 10 min duringthe 2-h period. Controls were conducted at constant sea-water temperature (13�C) using three similarly sizedindividuals per locality for each species, due to the lownumber of individuals that finally survived.
Thermal acclimation
Fish from each locality were separated into two groups.One group was maintained for 1 month at a constanttemperature of 14±1�C, while the other group was keptat 25±1�C. These acclimation temperatures are thethermal extremes in natural conditions. Both groupswere kept under identical laboratory conditions withconstant aeration and replacement of seawater andunlimited access to food. Ten individuals of each species
from each geographical locality were subjected to one ofthe two thermal treatments. Five individuals were usedfor each control during the thermal selectivity assays.Since S. viridis is a bottom-dwelling species, its thermalsensitivity was evaluated in an horizontal gradient, whileG. laevifrons being a water-column species, was evalu-ated in a vertical thermal gradient.
S. viridis
All of the S. viridis specimens used in the thermalselectivity experiments were similarly sized (6–7 cm).The thermal selectivity of S. viridis was evaluated in a80·15·15-cm aquarium which was filled with seawaterto a depth of 2.5 cm. This depth does not allow verticalthermal stratification within the aquarium, and alsopermits free movement of the fish. The aquarium wassplit into eight equally sized 10-cm compartments, andboth ends were placed on two 15·15·15-cm aquariafilled with seawater. A thermoregulated bath calibratedto 5�C was installed at one end of the tanks, while aheater set to 40�C was placed in the other extreme of thetank. After a period of 45 min, a thermal gradient wasestablished, ranging from 10.1±0.25�C to 28.6±0.49�Calong the 80·15·15-cm aquarium.
Each assay on an individual fish specimen lasted 1 hand was conducted using the following protocol. First,each fish was released in the mid-section of the aquar-ium; after 30 min (i.e. sufficient time for the fish toexplore the gradient), we registered the position of thefish within the thermal gradient. Preliminary assaysindicated that fish explored the entire experimental sys-tem within 20 min (Pulgar et al. 1999). Additionalobservations of position were made at 5-min intervalsduring the following 30 min, thus resulting in a total ofseven observations per fish. An identical protocol wasfollowed with the control fish, which were subjected to asituation with a constant temperature of 14�C. Prior tothe commencement of thermal selectivity assays wecontinuously measured temperature in the gradientusing a digital thermocouple confirming the stability ofthe gradient over this time period.
G. laevifrons
For this species temperature selectivity experiments wereperformed in an aquarium (30·30·60 cm) with a verticalthermal gradient. The aquarium was split into six 10-cmcells, separated by perforated acrylic sheets, which per-mitted fish movement. The bottom of the aquarium wasimmersed in a water bath at 10�C. A heater was placedon the top of the aquarium to maintain the water tem-perature at 31�C. The temperature of each cell wasmonitored for 2 days by means of a digital thermocou-ple to record the stability of the temperature gradient.The average temperature registered at each cell in thethermal gradient was (±1SD): bottom cell 1=10.3�C
513

(0.3), cell 2=12.9�C (1.3), cell 3=16.4�C (1.76), cell4=18.6�C (2.56), cell 5=25.3�C (1.11), upper cell6=30.9�C (1.22). Prior to the commencement of thermalselectivity assays we continuously measured temperaturein the gradient using a digital thermocouple confirmingthe stability of the gradient over this time period.
In each temperature assay one fish was introduced atthe top of the thermal gradient. After 30 min, werecorded the total time spent by the fish in each cell ofthe thermal gradient for a total of 20 min. The sameprotocol was applied to the control fish, but the systemwas maintained at a constant temperature of 14�C.
Statistical analyses
A significance level of P £ 0.05 was consideredthroughout the study. Differences in the frequency ofoccurrence of S. viridis and G. laevifrons were comparedbetween high and low tidepools and between localitiesusing a v2-test applied to a 3·2 contingency table (Zar1996).
At each locality, we evaluated the relationship be-tween temperature and opercular frequency for eachexperimental fish specimen exposed to temperatureincrements and each control fish using linear regression.The slopes obtained from these regressions were thencompared between localities and treatments using a two-way ANOVA (Zar 1996). In order to correct the con-founding effect between time and the temperature ofeach experimental group (control and experimental), theopercular frequency of control fishes was subtractedfrom the opercular frequency of experimental fishes.Later, the slopes of the relationship between tempera-ture and opercular frequency was evaluated betweenlocalities using a one-way ANOVA.
In order to analyse the amount of time spent by G.laevifrons specimens in each of the cells of the thermalgradient, we summed the time spent in the two cells withthe highest temperature (i.e. cells 5 and 6) separatelyfrom the time spent in the two cells with intermediatetemperature (3 and 4), and the two cells with the lowesttemperature (1 and 2) (Table 1). To evaluate thermalselectivity of fish from each locality and acclimationtreatment group, we calculated a standardized temper-ature (ST) as follows:
ST¼ððT�C1�time1ÞþðT �C2�time2ÞþðT �C3�time3ÞÞ
Totaltime
where, T�C 1=the highest temperature of the experi-mental gradient; time 1=duration the fish spent at thehighest temperature; T�C 2=average temperature of theexperimental gradient; time 2=duration the fish spent atthe average temperature, T�C 3=coldest temperature ofthe experimental gradient; time 3= duration the fishspent at the coldest temperature; Total time=time ofeach assay (1,200 s). ST values were compared using atwo-way ANOVA considering the factors geographical
locality (three levels, Antofagasta, Carrizal Bajo and LasCruces) and acclimation treatment (two levels, 14 and25�C).
To evaluate thermal selectivity between experimentalfish (i.e. those exposed to the thermal gradient) andcontrol fish (i.e. those maintained at constant tempera-ture), we utilized the Shannon-Weaver diversity index(H) (Ricklefs 1990), calculated as a measure of fish timedistribution:
H ¼ �Rpi loge pi
where pi is the proportion of time spent in each sector ofthe aquarium. Statistical differences in H values wereanalysed using a one-way ANOVA, considering theexperimental condition as a fixed factor with two levels(treatments and controls).
A similar experimental protocol and statistical anal-yses were followed for S. viridis. In this case, however,we registered the number of times that each fish wasobserved in each of the four sectors of the aquarium.
Results
Intertidal thermal sensitivity
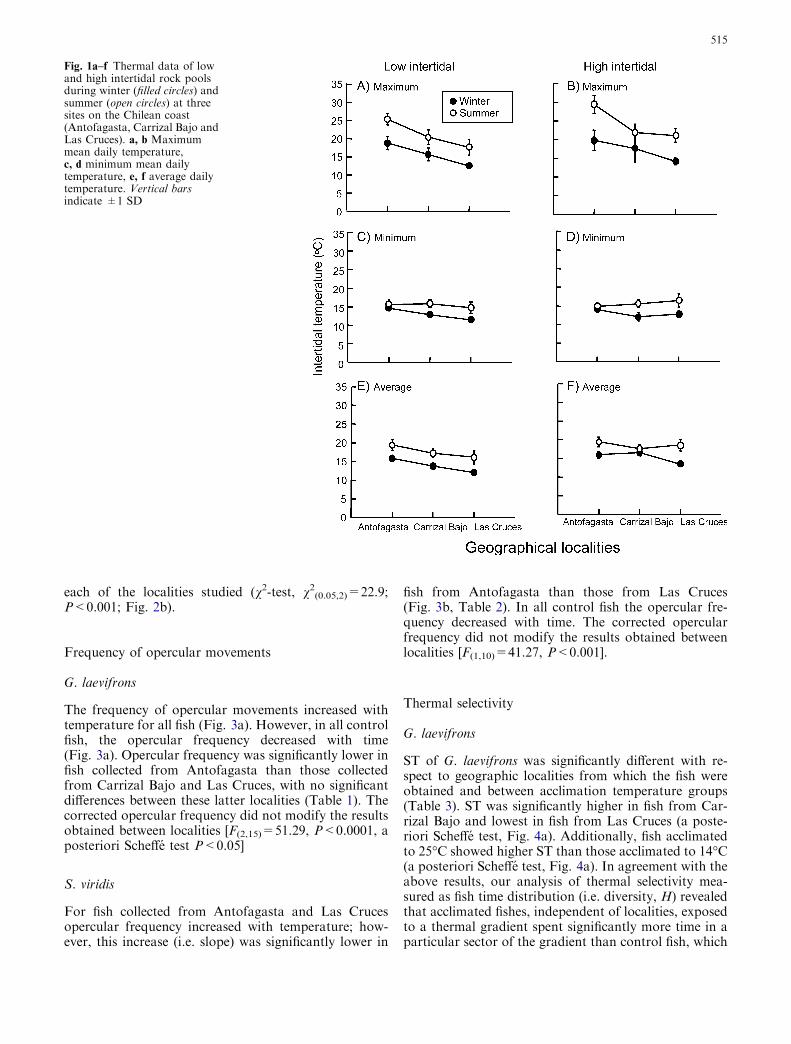
The mean maximal daily temperature recorded in the lowand high intertidal pools at each locality was higher insummer than in winter (Fig. 1a, b). Maximal tempera-tures were significantly higher in Antofagasta than inCarrizal Bajo and Las Cruces (Fig. 1a, b). On the otherhand, the minimum daily temperature diminishes fromsummer to winter in Las Cruces and Carrizal Bajo(Fig. 1c, d). The average daily temperature was higher insummer than inwinter in both intertidal bands (Fig. 1e, f).
Vertical distribution of S. viridis and G. laevifronsalong the intertidal gradient at different localities
The distributions of G. laevifrons and S. viridis in therocky intertidal zone proved to be strikingly different.G. laevifrons was significantly more abundant in highintertidal pools at all geographical localities (v2-test,v2(0.05,2)=25.69; P<0.001; Fig. 2a). In contrast, S. viri-dis was most abundant in the low intertidal rock pools at
Table 1 Results of the two-way ANOVA evaluating opercularfrequencya in Girella laevifrons. Factors are: locality (three levels)and treatment (two levels)
Effect df effect MS F P
Locality 1 403.811 174.93 0.0001Treatment 2 2.37 1.02 0.37Locality·treatment 1 32.44 14.05 0.00012Error 21 2.31
aOpercular frequency of G. laevifrons depends on interactionbetween locality and treatment (i.e. control fishes versus fishesexposed to different temperatures)
514
each of the localities studied (v2-test, v2(0.05,2)=22.9;P<0.001; Fig. 2b).
Frequency of opercular movements
G. laevifrons
The frequency of opercular movements increased withtemperature for all fish (Fig. 3a). However, in all controlfish, the opercular frequency decreased with time(Fig. 3a). Opercular frequency was significantly lower infish collected from Antofagasta than those collectedfrom Carrizal Bajo and Las Cruces, with no significantdifferences between these latter localities (Table 1). Thecorrected opercular frequency did not modify the resultsobtained between localities [F(2,15)=51.29, P<0.0001, aposteriori Scheffe test P<0.05]
S. viridis
For fish collected from Antofagasta and Las Crucesopercular frequency increased with temperature; how-ever, this increase (i.e. slope) was significantly lower in
fish from Antofagasta than those from Las Cruces(Fig. 3b, Table 2). In all control fish the opercular fre-quency decreased with time. The corrected opercularfrequency did not modify the results obtained betweenlocalities [F(1,10)=41.27, P<0.001].
Thermal selectivity
G. laevifrons
ST of G. laevifrons was significantly different with re-spect to geographic localities from which the fish wereobtained and between acclimation temperature groups(Table 3). ST was significantly higher in fish from Car-rizal Bajo and lowest in fish from Las Cruces (a poste-riori Scheffe test, Fig. 4a). Additionally, fish acclimatedto 25�C showed higher ST than those acclimated to 14�C(a posteriori Scheffe test, Fig. 4a). In agreement with theabove results, our analysis of thermal selectivity mea-sured as fish time distribution (i.e. diversity, H) revealedthat acclimated fishes, independent of localities, exposedto a thermal gradient spent significantly more time in aparticular sector of the gradient than control fish, which
Fig. 1a–f Thermal data of lowand high intertidal rock poolsduring winter (filled circles) andsummer (open circles) at threesites on the Chilean coast(Antofagasta, Carrizal Bajo andLas Cruces). a, b Maximummean daily temperature,c, d minimum mean dailytemperature, e, f average dailytemperature. Vertical barsindicate ±1 SD
515

tended to be equally distributed among all the sectors.The time diversity (±1 SE) for control and for experi-mental fishes in the thermal gradient was: controlfish=1.05 (0.02) and experimental fish in the thermalgradient=0.65 (0.04) [ANOVA, F(1, 88)=44.93;P<0.0001].
S. viridis
Similarly to G. laevifrons, the ST of S. viridis wasdependent on the geographic locality from which the fishwere obtained as well as acclimation temperature(Table 4). Standard temperature was significantly higherin fish from Antofagasta and lower in fish from LasCruces (a posteriori Scheffe test, Fig. 4b). Furthermore,fish from Las Cruces acclimated to 25�C showed sig-nificantly higher ST values than those acclimated to14�C, while there was no significant difference betweenacclimation treatments in fish from Antofagasta (aposteriori de Scheffe test, Fig. 4b).
Our analysis of thermal selectivity measured as fishtime distribution (i.e. diversity, H) revealed that accli-mated fishes independent of localities exposed to athermal gradient spent significantly more time in aparticular sector of the gradient than control fish, whichtended to be equally distributed among all the sectors.The time H (±1SE) for control and for experimentalfishes in the thermal gradient was: control fish=5.82(0.16), and experimental fish in the thermal gradi-ent=1.88 (0.24) (ANOVA F(1, 58)=110.95; P<0.00001).
Fig. 2 Intertidal vertical distribution of a Girella laevifrons, whichmainly occupies the high intertidal sectors and b Scartichthysviridis, which mainly occupies the low intertidal sectors. Numbersabove bars indicate the total number of fish collected from all tidalpools at each locality
Fig. 3 Regression slope of frequency of opercular movement ofa G. laevifrons and b S. viridis in seawater with increasingtemperature (open circles) (13–30�C at 0.1�C/min over a 2-hperiod) or stable temperature of 13�C (closed circles). Vertical barsindicate ±1 SE
Table 2 Results of the two-way ANOVA evaluating fish opercularfrequencya in Scartichthys viridis. Factors are: locality (two levels)and treatment (two levels)
Effect df effect MS F P
Locality 1 166.88 224.38 0.0001Treatment 1 14.98 20.15 0.0005Locality·Treatment 1 13.82 18.58 0.0007Error 14 0.74
aOpercular frequency of S. viridis depends on interaction betweenlocality and treatment (i.e. control fishes versus fishes exposed todifferent temperatures)
516

Discussion
Thermal variability of the intertidal system
The main objective of our study was to determine thethermal sensitivity of the intertidal fish, as well as tomeasure the thermal characteristics of the low and highintertidal pools habitats along 10� of latitude. Asmentioned, maximal daily temperature of the thenorthernmost tidepool, at Antofagasta, was higher thanat the other study localities along the Chilean coast
(Fig. 1a, b). The mean minimum daily temperature ofthe rock pools was lower in winter for all localities withthe exception of Antofagasta where it was about 16�C(Fig. 1c, d). The average daily temperature of the poolswas relatively constant among the three geographiclocalities and the two vertical positions (Fig. 3e, f).Moreover tidal pools located in the high intertidal re-flected a strong seasonal component in water tempera-ture (Fig. 1). The water temperature of rock pools atthe three studied localities fluctuated between 13 and30�C. These results are the first to report on the phys-ical variability of the intertidal system of the Chileancoast. Our results are consistent with those reported forintertidal pools at other latitudes, where wide fluctua-tions in the physical conditions of rock pools, includingtemperature, oxygen and salinity, have been reported(Newell 1970; Gibson 1972; Stephenson and Stephenson1972; Metaxas and Scheibling 1993; Horn and Gibson1988). In these studies, the magnitude of variationamong tide pools was dependent on the vertical posi-tion of the pool within the intertidal zone (the furtheraway from the breaking tide, the greater the physicalfluctuations). The same pattern was also observed inour study, where the most distant tide pools attainedthe highest temperature at all localities, particularly insummer months where temperatures in the higherintertidal pools of Antofagasta reached values ofaround 30�C (Fig. 1a). This pattern of thermal condi-tions suggests that fish inhabiting high intertidal rockpools are subjected to the greatest thermal variability,which likely affects their energetic return.
Intertidal vertical distribution of S. viridisand G. laevifrons
S. viridis and G. laevifrons presented opposite patterns ofvertical distribution within the intertidal zone at all threelocalities. S. viridis was most abundant in the lowersectors of the intertidal zone, while G. laevifrons wasmost abundant in the higher sectors of the zone. Thesedifferences in distribution imply that these species aresubject to different regimes of thermal variability(Fig. 2a, b). Previously, Zander et al. (1999) indicatedthat intertidal fish species occupy coastal levels that canvary in their absolute vertical distribution, but not intheir relative position. This observation has been asso-ciated with active searching for specific thermal micro-habitats (Gibson 1982). In this sense, even whenvariations exist in the width of the intertidal band be-tween the sampled localities, G. laevifrons and S. viridiswill continue to present opposite patterns of verticalintertidal distribution (Fig. 2a, b).
Many authors have suggested that high intertidalpools act as refuge areas, especially for juvenile fish (e.g.G. laevifrons), where individuals can avoid negativeinteractions, such as competition with species that aremore abundant in the low intertidal zone (e.g. S. viridis),or subtidal predators that invade the intertidal zone
Fig. 4 Thermal selectivity of a G. laevifrons and b S. viridis fromAntofagasta (filled circles), Carrizal Bajo (open circles) and LasCruces (filled inverted triangles), for fishes acclimated to 14�C and25�C. Vertical bars indicate ±1 SE. Data for S. viridis for CarrizalBajo are not available. The experimental design used to evaluatethermal selectivity include a vertical gradient for G. laevifrons and ahorizontal gradient for S. viridis
Table 3 Results of the two-way ANOVA evaluating thermalselectivitya in G. laevifrons. Factors are: locality (three levels) andacclimation temperature (two levels)
Effect df effect MS F P
Locality 2 327.33 35.71 0.0001Acclimation 1 151.10 16.48 0.0016Locality·acclimation 2 4.11 0.44 0.64Error 54 9.16
aThe standardized temperature (ST) selected by fishes from Carri-zal Bajo and Antofagasta was higher than those selected by fishesfrom Las Cruces. On the other hand, the fish acclimated at 25�Cselected a greater ST than those acclimated at 14�C
517

during high tide (Gibson 1982; Horn and Gibson 1988;Pulgar et al. 1999; Zander et al. 1999; Hernandez et al.2002). However, despite possibly being able to escapestrong biotic interactions in the low intertidal zone (i.e.competition and predation), juvenile fish in the highintertidal zone, such as G. laevifrons, must face largevariations in their physical habitat (Zander et al. 1999;Hernandez et al. 2002). These variations could generatephysiological responses (see Pulgar et al. 1999; Her-nandez et al. 2002) that may be dependent on the localityor microhabitat from where the fish are obtained.
Thermal tolerance of S. viridis and G. laevifrons
The frequency of opercular movements has beenindicated as a direct measure of metabolic costs asso-ciated with a particular thermal environment (Rogersand Weatherley 1993; Van Rooij and Videler 1996).Among intertidal fish, Hernandez et al. (2002) indi-cated that Graus nigra juveniles were able to tolerategreater increments in the temperature of the seawaterthan larger adult fish and to continue responding tothermal stress. In our study, fish from each of thestudied localities responded to increasing temperatureby increasing the frequency of their opercular move-ments. Nevertheless, the response of fish from thenorthernmost locality (Antofagasta) was less intensethan that of fish inhabiting the intertidal pools atCarrizal Bajo, reflecting the observed thermal ends inAntofagasta. Furthermore, fish collected from thesouthernmost locality (Las Cruces), displayed thegreatest magnitude of response to temperatures(Fig. 3a, b). Therefore, our results suggest that theenergy cost associated with a particular thermal con-dition, here estimated indirectly as the frequency ofopercular movements, could be least among fish fromAntofagasta and greatest among fish from Las Cruces(Tables 2, 3; Fig. 3a, b).
Differences in the frequency of opercular movementsof S. viridis and G. laevifrons from the three study sitesclearly coincide with the observed variations in the tidalpool temperatures at each locality (Fig. 1). In this sense,the observed variations in opercular frequency of S.viridis and G. laevifrons from the three populations,indicate geographic variability in the thermal stress re-sponse (Fig. 3a, b), which could be associated with dif-ferent thermal optima for processes such as digestion,assimilation, and reproduction (Angilletta et al. 2002).In this way, natural selection, acting on separate popu-lations subjected to different thermal stresses, wouldensure the normal functioning of biological processesthat have direct effects on fitness (Huey and Bennett1987). Field responses of fish should be determined bythe local physical conditions where fish are obtained. Infurther support of our conclusions, control fish (i.e. fishnot exposed to a thermal gradient) responded bydecreasing opercular movements throughout the dura-tion of experimental runs (Figs. 3, 4).
Thermal selectivity of S. viridis and G. laevifrons
Thermal selection is strongly developed among ecto-therms as a mechanism of reducing possible impacts ofspatial and temporal variations in environmental tem-perature. This property of ectotherms can be affected bymany factors including nutritional state, reproductivestatus, or food quality (Bozinovic and Rosenmann 1988;Angilletta et al. 2002). Pulgar et al. (1999) indicated thatjuvenile G. laevifrons subjected to a thermal gradient,revealed a strong thermophilic tendency. In our study,variations in thermal selectivity were observed in both G.laevifrons, as well as S. viridis (Fig. 4a, b). On the onehand, fish from the northernmost locality were observedto present the greatest frequency of opercular move-ments and to remain longest in the highest temperaturesof the thermal gradient, compared to fish from thelocalities located further south (i.e. Carrizal Bajo andLas Cruces) (Table 4; Fig. 4a, b). On the other hand,thermal selectivity also depended on the temperature towhich fish were acclimated, since specimens from thesame locality that were acclimated to different temper-atures presented differences in their thermal selectivity(Table 4; Fig. 4a, b). These observations of thermalsensitivity, together with measurements of opercularfrequency, indicate that thermoregulatory behaviour inectotherms could reveal the existence of between-popu-lation differences in the optima of physiological pro-cesses that can have a direct bearing on fitness (Conoverand Present 1990; Conover 1998). Moyle and Cech(1982) demonstrated that some fish can select particulartemperatures in order to conserve energy or enhance theefficiency of their enzymatic machinery. Thus, a changein the thermal sensitivities of species that are widelydistributed and subjected to great variability of theirphysical environment, could enable them to respond toenvironmental variability and to persist in a particularhabitat (Huey and Bennett 1987, Angilletta et al. 2002).Our results show that thermal selectivity is dependent onthe temperature to which fish were acclimated in thelaboratory as well as on the particular geographiclocality from which specimens came. The differencesobserved between control fish and individuals subjectedto the experimental gradient support our conclusions(Fig. 4a, b). These results suggest that physiological
Table 4 Results of the two-way ANOVA evaluating thermalselectivitya in S. viridis. Factors are: locality (two levels) andacclimation temperature (two levels)
Effect df effect MS F P
Locality 1 950.62 93.12 0.0001Acclimation 1 83.68 8.19 0.016Locality·Acclimation 1 28.17 2.76 0.11Error 36 10.2
aThe ST selected by fishes from Antofagasta was higher than thoseselected by fishes from Las Cruces. On the other hand, the fishacclimated at 25�C selected a greater ST than those acclimated at14�C
518

differences exist among populations and that these dif-ferences can potentially compensate for the observedvariability in the physical thermal environment.
In conclusion, we revealed geographic differences inthe thermal sensitivity of S. viridis and G. laevifrons,which could not be explained by acclimation or pheno-typic flexibility. These results are ecologically relevantand are able to explain macroscale distribution patternsin terms of physiological responses, and contrast withmost studies, which have evaluated what animals do butnot what they are able to do (Gaston and Spicer 1998).Finally as pointed out by Spicer and Gaston (1999),between-species variations in physiological traits canarise from methodological differences, allometric prob-lems, species-specific or phylogenetic differences and/orthe specific environmental conditions experienced byspecies and populations. Nevertheless, given that ourstudy was intraspecific and since we used the samemethods throughout, we are confident that the thermalsensitivity of the field fish subjected to acclimation isdependent on geographic locality, a result that is directlyassociated with variations in the seawater temperature ofintertidal rock pools and with the patterns of occupationof S. viridis and G. laevifrons of the intertidal habitat ofthe three studied localities.
Acknowledgements We thank M. Aldana and A. Munoz for valu-able comments on the manuscript. This study was funded byFondecyt grant 2010218 to J. P. and FONDAP-FONDECYTgrant 1501-0001 to the Centre for Advanced Studies in Ecology andBiodiversity, Programs 1 (FB) and 5 (FPO).
References
Aldana M, Pulgar J, Ogalde, Ojeda FP (2002) Morphometric andparasitological evidence for ontogenetic and geographic dietaryshifts in fish. Bull Mar Sci 70:55–74
Angilletta MJ Jr, Niewiarowski PH, Navas CA (2002) The evolu-tion of thermal physiology in ectotherms. J Thermal Biol27:249–268
Bennett B, Griffiths CL (1984) Factors affecting the distribution,abundance and diversity of rock-pool fishes on the Cape Pen-insula, South Africa. S Afr J Zool 19:97
Bennett B, Griffiths CL , Penrith M (1983) The diets of littoral fishfrom the Cape Peninsula. S Afr Tydskr Dierk 18:343–352
Bennett BA (1987) The rock-pool community of Koppie Allen andassessment of the importance of Cape rock-pools as nurseriesfor juvenile fish. S Afr J Zool 22:25–32
Bozinovic F, Rosenmann M (1988) Energetics and food require-ments of the female snake Phillodryas chamissons during thebreeding season. Oecologia 75:282–284
Bozinovic F, Rosenmann M (1989) Maximum metabolic rate ofrodents: physiological and ecological consequences on distri-butional limits. Funct Ecol 3:173–181
Bozinovic F, Rosenmann M, Novoa FF, Medel RG (1995) Med-iterranean-type of climatic adaptation in the physiologicalecology of rodent species. In: Kalin-Arroyo MT, Zedler PH,Fox MD (eds) Ecology and biogeography of mediterraneanecosystems in Chile, California and Australia). Springer, BerlinHeidelberg New York, pp 347–362
Bridges CR (1993) Ecophysiology of intertidal fish. In: Rankin JC,Jensen FB (eds) Fish ecophysiology. Chapman and Hall, Lon-don, pp 375–400
Chown SL, Gaston KL (2000) Areas, cradles and museums: thelatitudinal gradient in species richness. Trends Ecol Evol 8:310–315
Collette BB (1986) Resilence of the fish assemblages in New Eng-land tidepools. US Fish Bull 84:200–204
Conover DO (1998) Local adaptation in marine fishes: evidenceand implications for stock enhancement. Bull Mar Sci 62:477–493
Conover DO, Present TMC (1990) Countergradient variation ingrowth rate: compensation for length of the growing seasonamong Atlantic silversides from different latitudes. Oecologia83:316–324
Cossins AR, Bowler K (1987) Temperature biology of animals.Chapman and Hall, New York
Davis BJ (1977) Distribution and temperature adaptation in theteleost fish Genus Gibbonsia. Mar Biol 42:320–345
Davis BJ (2000) Spatial and seasonal patterns of habitat parti-tioning in a guild of Southern California tidepool fish. Mar EcolProg Ser 196:256–268
Gaston KJ, Spicer JL (1998) Do upper thermal tolerances differ ingeographically separated populations of the beachflea Orchestiagammarellus (Crustacea: Amphipoda)? J Exp Mar Biol Ecol229:265–276
Gibson RR (1972) The vertical distribution and feeding relation-ship of intertidal fish on the Atlantic coast of France. J AnimEcol 41:189–207
Gibson RN (1982) Recent studies on the biology of intertidal fish.Oceanogr Mar Biol Annu Rev 20:263–414
Gibson RN, Yoshiyama RM (1999) Intertidal fish communities. In:Horn MH, Martin KL, Chotkowski MA (eds) Intertidal Fish.Life in two worlds. Academic Press, San Diego, Calif., pp 95–125
Gosline WA (1965) Vertical distribution of inshore fishes in theupper water layers of the Hawaiians islands. Ecology 46:823–881
Graham JB (1970) Temperature sensivity of two species of inter-tidal fish. Copeia 1970:49–56
Granterner A, Taborsky M (1998) The metabolic rates associatedwith resting, and with the performance of agonist, submissiveand digging behaviours in the cichlid fish Neolmprolonus pulcher(Pisces: Cichlidae). J Comp Physiol B Biochem Syst EnvironPhysiol 168:427–433
Grossman GD (1982) Dynamics and organization of a rockyintertidal fish assemblage: the persistence and resilence oftaxocene structure. Am Nat 119: 611–637
Grossman GD (1986) Food resource partitioning in a rockyintertidal fish assemblage. J Zool 1:317–335
Hernandez CE, Neill PE, Pulgar JM, Ojeda FP, Bozinovic F (2002)Water temperature fluctuations and territoriality in the inter-tidal zone: two possible explanations for the vertical distribu-tion of body size in Graus nigra (Kyphosidae). J Fish Biol61:472–488
Horn MH, Gibson RN (1988) Intertidal fish. Sci Am 258:54–60Horn MH, Martin KL, Chotkowski MA (1999) Intertidal fiskes:
life in two worlds. Academic Press, San Diego, Calif.Huey RB, Bennett AF (1987) Phlyogenetic studies of coadaptation:
preferred temperatures versus optimal performance tempera-tures of lizards. Evolution 41:1098–1115
Lenski RE, Bennett AF (1993) Evolutionary responses of Escher-ichia coli to thermal stress. Am Nat 142:S47–S64
Lonsdale DJ, Levinton JS (1985) Latitudinal differentiation incopepod growth: an adaptation to temperature. Ecology66:1397–1407
Metaxas A, Scheibling RE (1993) Community structure andorganization of tidepools. Mar Ecol Prog Ser 98:187–198
Morritt D, Ingolfsson A (2000) Upper thermal tolerances of thebeach flea Orchestia gammarellus Pallas (Crustacea: Amphi-poda: Talitridae) associated with hot springs in Iceland. J ExpMar Biol Ecol 255:215–227
Moyle PB, Cech JJ (1982) Fish: an introduction to Ichthyology.Prentice Hall, Englewood Cliffs, N.J.
519

Munoz AA, Ojeda FP (1997) Feeding guild structure of a rockyintertidal fish assemblage in central Chile. Environ Biol Fish49:471–479
Munoz AA, Ojeda FP (1998) Guild structure of carnivorousintertidal fish of the Chilean coast: implications of ontogeneticdietary shifts. Oecologia 114:563–573
Nelson JS (1994) Fish of the world. Wiley, New YorkNewell RC (1970) Biology of intertidal animals. Marine Ecological
Survey, UKNorris KS (1963) The functions of temperature in the ecology of
the percoid fish Girella nigricans (Aires). Ecol Monogr 33:23–62Ojeda FP, Labra FA, Munoz AA (2000) Biogeographic patterns of
Chilean littoral fish. Rev Chil Hist Nat 73:625–641Parsons PA (1993) The importance and consequences of stress in
living and fossil populations: from life-history variations toevolutionary change. Am Nat 142:S6–S20
Piersma T, Drent J (2003) Phenotypic flexibility and the evolutionof organismal design. Trends Ecol Evol 18:228–233
Prochazka K, Griffiths CL (1992) The intertidal fish fauna of thewet coast of South Africa species, community and biogeo-graphic patterns. S Afr J Zool 27:115–121
Pulgar J, Bozinovic F, Ojeda FP (1999) Behavioral thermoregula-tion in the intertidal fish Girella laevifrons (Kyphosidae): theeffect of starvation. Mar Freshwater Behav Physiol 32:27–38
Ralston SL, Horn MH (1986) High tide movements of the tem-perate-zone herbivorous fish Cebidichthys violaceus (Girardi) asdetermined by ultrasonic telemetry. J Exp Mar Biol Ecol 98:35–50
Ricklefs RR (1990) Ecology. Freeman, New YorkRogers SC, Weatherley AH (1993) The use of opercular muscle
myograms as an indicator of the metabolic costs of fish activityin rainbow trout, S. gairdneri Richardson, as determined byradiotelemetry. J Fish Biol 23:535–547
Spicer JL, Gaston KG (1999) Physiological diversity and its eco-logical implications. Blackwell, Oxford
Stearns SC (1982) The evolutionary significance of phenotypicplasticity. Bioscience 7:436–445
Stephen CA, Porter WP (1993) Temperate, activity, and lizard lifehistories. Am Nat 42:273–293
Stephenson TA, Stephenson A (1972) Life between tidemarks onrocky shore. Freeman, San Francisco, Calif.
Stepien CA (1990) Population structure, diets and biogeographicrelationships of a rocky intertidal fish assemblage in centralChile: high levels of herbivory in a temperate system. Bull MarSci 47(3):598–612
Thompson DA, Lehner CE (1976) Resilence of a rocky intertidalfish community in a physically unstable environment. J ExpMar Biol Ecol 22:1–29
Tracy CR, Christian KA (1986) Ecological relations among space,time, and thermal niche axes. Ecology 67:609–615
Truchot JP, Duhanel-Jouve A (1980) Oxygen and carbon dioxidein the marine intertidal environments: diurnal and tide changesin rockpools. Respir Physiol 39:241–254
Van Rooij JM, Videler JJ (1996) Estimating oxygen uptake ratefrom ventilation frequency in the reef fish Sparisoma viride. MarEcol Prog Ser 132:21–41
Varas E, Ojeda FP (1990) Intertidal fish assemblage of the centralChilean coast: diversity, abundance and trophic patterns. RevBiol Mar 25:59–70
Weise W (1991) Limitation of energy acquisition and energy use insmall poikilotherms: evolutionary implications. Funct Ecol5:234–240
Yoshiyama RM (1980) Food habits of three species of rockyintertidal sculpins (Cottidae) in central California. Copeia 1980:515–525
Yoshiyama RM (1981) Distribution and abundance patterns ofrocky intertidal fishes in central California. Environ Biol Fish6:315–332
Zander CD, Nieder J, Martin K (1999) Vertical distribution pat-tern. In: Horn MH, Martin KL, Chotkowski MA (eds) Inter-tidal fish: life in two worlds. Academic Press, San Diego, Calif.,pp 264–296
Zar J (1996) Biostatistical analysis. Prentice Hall, Englewood Cliffs,N.J.
520




















