
Diversity in hydrogen evolution from bidirectional hydrogenases in cyanobacteria from terrestrial,...
10
Journal of Biotechnology 162 (2012) 105–114 Contents lists available at SciVerse ScienceDirect Journal of Biotechnology jou rn al hom epage: www.elsevier.com/locate/jbiotec Diversity in hydrogen evolution from bidirectional hydrogenases in cyanobacteria from terrestrial, freshwater and marine intertidal environments Ankita Kothari, Ruth Potrafka, Ferran Garcia-Pichel ∗ School of Life Sciences, Arizona State University, Tempe, AZ 85287-4501, USA a r t i c l e i n f o Article history: Received 11 January 2012 Received in revised form 20 March 2012 Accepted 17 April 2012 Available online 4 July 2012 Keywords: Oxygenic phototrophs Photohydrogen Hydrogenases Microbial mats Soils microbes Plankton a b s t r a c t We characterized a set of 36 strains of cyanobacteria isolated from terrestrial, freshwater and marine intertidal settings to probe their potential to produce hydrogen from excess reductant, in the hope of finding novel strains with improved traits for biohydrogen production. The set was diverse with respect to origin, morphology, taxonomy and phylogeny. We found that about one half of the strains could produce hydrogen from hydrogenases in standard assays, a trait that corresponded invariably with the presence of homologues of the gene hoxH, coding for subunit H in the bidirectional Ni–Fe hydroge- nase. Strains from freshwater and intertidal settings had a high incidence of hydrogen producing, hoxH containing strains, but all terrestrial isolates were negative for both. While specific rates of hydrogen production varied among strains, some novel strains displayed rates several fold higher than those pre- viously reported. We detected two different patterns in hydrogen production. Pattern 1, corresponding to that previously known in Synechocystis PCC 6803, encompassed strains whose hydrogenase system produced hydrogen only temporarily to revert to hydrogen consumption within a short time and after reaching moderate hydrogen concentrations. Cyanobacteria displaying pattern 2, in the genera Lyngbya and Microcoleus, tended to have higher rates, did not reverse the direction of the reaction and reached much higher concentrations of hydrogen at steady state, making them of interest as potential platforms for biohydrogen production. Published by Elsevier B.V. 1. Introduction Interest in the generation of renewable fuels has gained momen- tum in the last decades in the face of global warming associated with the continued use of fossil fuels, and because of the finite nature of their reserves. Biohydrogen production from photosyn- thetic organisms constitutes a conceptually promising avenue in renewable bioenergy, because it would couple directly solar radi- ant energy, essentially inexhaustible, to the generation of clean, carbon-neutral biofuels, particularly if water-splitting (oxygenic) phototrophs were used (Weaver et al., 1980; Akkerman et al., 2002; Prince and Kheshgi, 2005). In spite of the efforts geared toward developing biological platforms for photohydrogen production, none has yet reached large-scale production stages. Cyanobacteria, the only group of oxygenic phototrophs among the bacteria, have been regarded as good models for research and eventual applica- tion in this area for several reasons: they are capable of growth with minimal nutritional requirements, they are demonstrable Abbreviation: PCC, Pasteur culture collection. ∗ Corresponding author. Tel.: +1 480 727 7534. E-mail addresses: [email protected] (A. Kothari), [email protected] (R. Potrafka), [email protected] (F. Garcia-Pichel). producers of hydrogen gas under certain physiological conditions, and some can be genetically modified with ease. Among cyanobac- teria, three different enzymes participate in hydrogen metabolism (Ghirardi et al., 2007; Tamagnini et al., 2007): nitrogenase, and two types of Ni–Fe hydrogenases (uptake and bidirectional). The nitrogenase produces H 2 at the cost 2 ATP per mol, and it does so as a byproduct of its main reaction, the assimilatory reduction of N 2 (Phelps and Wilson, 1941; Hyndman et al., 1953; D’Eustachio and Hardy, 1964) In principle, production of biohydrogen based on nitrogenase systems requires significant modifications of the enzyme or cumbersome growth conditions in order to promote proton reduction and prevent N 2 reduction. In spite of this, sig- nificant improvements have recently been achieved by extending the panoply of cyanobacterial species investigated, in order to take advantage of pre-existing physiological diversity (Bandyopadhyay et al., 2010). The hydrogen produced by nitrogenase is often recy- cled back into metabolic reducing equivalents by means of the uptake hydrogenase (Peterson and Burris, 1978; Eisbrenner and Evans, 1983). Under physiological conditions, and as the name sug- gests, the latter enzyme can only consume, rather than produce H 2 , and so does not constitute a viable platform for biohydrogen production; in fact, it needs to be inactivated to improve yields of nitrogenase-based hydrogen production (Happe et al., 2000; Lindblad et al., 2002; Yoshino et al., 2007). 0168-1656/$ – see front matter. Published by Elsevier B.V. http://dx.doi.org/10.1016/j.jbiotec.2012.04.017
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Diversity in hydrogen evolution from bidirectional hydrogenases in cyanobacteria from terrestrial,...

Dc
AS
a
ARRAA
KOPHMSP
1
twntracpPdntbtw
(
0h
Journal of Biotechnology 162 (2012) 105– 114
Contents lists available at SciVerse ScienceDirect
Journal of Biotechnology
jou rn al hom epage: www.elsev ier .com/ locate / jb io tec
iversity in hydrogen evolution from bidirectional hydrogenases inyanobacteria from terrestrial, freshwater and marine intertidal environments
nkita Kothari, Ruth Potrafka, Ferran Garcia-Pichel ∗
chool of Life Sciences, Arizona State University, Tempe, AZ 85287-4501, USA
r t i c l e i n f o
rticle history:eceived 11 January 2012eceived in revised form 20 March 2012ccepted 17 April 2012vailable online 4 July 2012
eywords:xygenic phototrophshotohydrogenydrogenasesicrobial mats
a b s t r a c t
We characterized a set of 36 strains of cyanobacteria isolated from terrestrial, freshwater and marineintertidal settings to probe their potential to produce hydrogen from excess reductant, in the hope offinding novel strains with improved traits for biohydrogen production. The set was diverse with respectto origin, morphology, taxonomy and phylogeny. We found that about one half of the strains couldproduce hydrogen from hydrogenases in standard assays, a trait that corresponded invariably with thepresence of homologues of the gene hoxH, coding for subunit H in the bidirectional Ni–Fe hydroge-nase. Strains from freshwater and intertidal settings had a high incidence of hydrogen producing, hoxHcontaining strains, but all terrestrial isolates were negative for both. While specific rates of hydrogenproduction varied among strains, some novel strains displayed rates several fold higher than those pre-viously reported. We detected two different patterns in hydrogen production. Pattern 1, corresponding
oils microbeslankton
to that previously known in Synechocystis PCC 6803, encompassed strains whose hydrogenase systemproduced hydrogen only temporarily to revert to hydrogen consumption within a short time and afterreaching moderate hydrogen concentrations. Cyanobacteria displaying pattern 2, in the genera Lyngbyaand Microcoleus, tended to have higher rates, did not reverse the direction of the reaction and reachedmuch higher concentrations of hydrogen at steady state, making them of interest as potential platformsfor biohydrogen production.
. Introduction
Interest in the generation of renewable fuels has gained momen-um in the last decades in the face of global warming associatedith the continued use of fossil fuels, and because of the finiteature of their reserves. Biohydrogen production from photosyn-hetic organisms constitutes a conceptually promising avenue inenewable bioenergy, because it would couple directly solar radi-nt energy, essentially inexhaustible, to the generation of clean,arbon-neutral biofuels, particularly if water-splitting (oxygenic)hototrophs were used (Weaver et al., 1980; Akkerman et al., 2002;rince and Kheshgi, 2005). In spite of the efforts geared towardeveloping biological platforms for photohydrogen production,one has yet reached large-scale production stages. Cyanobacteria,he only group of oxygenic phototrophs among the bacteria, have
een regarded as good models for research and eventual applica-ion in this area for several reasons: they are capable of growthith minimal nutritional requirements, they are demonstrableAbbreviation: PCC, Pasteur culture collection.∗ Corresponding author. Tel.: +1 480 727 7534.
E-mail addresses: [email protected] (A. Kothari), [email protected]. Potrafka), [email protected] (F. Garcia-Pichel).
168-1656/$ – see front matter. Published by Elsevier B.V.ttp://dx.doi.org/10.1016/j.jbiotec.2012.04.017
Published by Elsevier B.V.
producers of hydrogen gas under certain physiological conditions,and some can be genetically modified with ease. Among cyanobac-teria, three different enzymes participate in hydrogen metabolism(Ghirardi et al., 2007; Tamagnini et al., 2007): nitrogenase, andtwo types of Ni–Fe hydrogenases (uptake and bidirectional). Thenitrogenase produces H2 at the cost 2 ATP per mol, and it does soas a byproduct of its main reaction, the assimilatory reduction ofN2 (Phelps and Wilson, 1941; Hyndman et al., 1953; D’Eustachioand Hardy, 1964) In principle, production of biohydrogen basedon nitrogenase systems requires significant modifications of theenzyme or cumbersome growth conditions in order to promoteproton reduction and prevent N2 reduction. In spite of this, sig-nificant improvements have recently been achieved by extendingthe panoply of cyanobacterial species investigated, in order to takeadvantage of pre-existing physiological diversity (Bandyopadhyayet al., 2010). The hydrogen produced by nitrogenase is often recy-cled back into metabolic reducing equivalents by means of theuptake hydrogenase (Peterson and Burris, 1978; Eisbrenner andEvans, 1983). Under physiological conditions, and as the name sug-gests, the latter enzyme can only consume, rather than produce
H2, and so does not constitute a viable platform for biohydrogenproduction; in fact, it needs to be inactivated to improve yieldsof nitrogenase-based hydrogen production (Happe et al., 2000;Lindblad et al., 2002; Yoshino et al., 2007).
1 iotech
nhTtebrotcTepsbpfbAftS6toat2
pabgagncefBiodahdsm
aibpcmtimfl(scdet
06 A. Kothari et al. / Journal of B
Certain cyanobacteria, however, host a bidirectional hydroge-ase that can catalyze both the production and the uptake ofydrogen under physiological conditions (Fujita and Myers, 1965).his enzyme is attractive because it is naturally much less sensitiveo long-term inactivation by O2 than other hydrogenases (Vignaist al., 2001; Cournac et al., 2004; Vignais and Colbeau, 2004) andecause it does not depend on nitrogen metabolism. Much of theesearch on biohydrogen production in cyanobacteria has focusedn this enzyme as platform. Physiological studies of the bidirec-ional hydrogenase revealed that it can play multiple roles in theell: it can be a part of fermentative metabolism (Oost et al., 1989;roshina et al., 2002), act as an emergency electron valve to releasexcess electrons during photosynthesis (Appel et al., 2000), anderhaps also as a general regulator for maintaining optimal redoxtate in the cell (Carrieri et al., 2011). Direct production of hydrogeny the bidirectional hydrogenase with electrons coming from thehotolysis of water has been demonstrated during the transitionrom (anaerobic) dark to light conditions in Synechocystis PCC 6803,ut this production is slow and very transient (Houchins, 1984;ppel and Schulz, 1998). In fact, one of the major disadvantages
or sustained H2 production via the bidirectional hydrogenase ishe easy reversal of the reaction direction (Tamagnini et al., 2007).imilarly, in our experience, when we subjected Synechocystis PCC803 to the specific hydrogenase activity assay we observed thathe production was transient and the enzyme soon reverted to H2xidation. Genetic modification of standard strains has been anpproach used in trying to address the shortcomings of the bidirec-ional hydrogenase platform (Cournac et al., 2004; Gutthann et al.,007; Marques et al., 2011; Masukawa et al., 2002).
Alternatively, one can also try a bio-prospecting approach, torobe existing biological diversity in the search for natural vari-nts of the enzyme or its regulatory networks that constituteetter production platforms. Perhaps with this in mind, extensiveenomic surveys have been conducted on aquatic environments inn attempt to understand the distribution and diversity of the hoxHene, coding for the large H subunit of the bidirectional hydroge-ase. Some authors suggested that bidirectional hydrogenases areommon where anaerobic or micro-aerobic conditions may be rel-vant ecologically, and hence hox genes are highly unlikely to beound at high frequency in open ocean waters (Ludwig et al., 2006).arz et al. (2010) could show a clear increase in abundance of hoxH
n freshwater and marine coastal environments, with respect to thepen oceans. Using a more physiological approach, focused on pro-uction by N-fixing strains, Allahverdiyeva et al. (2010) surveyed
large number of existing isolates from culture collections forydrogen evolution. They found that about 50% of strains producedetectable amounts of H2, and about 2% displayed production ratesimilar to those obtained in standard strains optimized by geneticodification.Our study is novel in carrying out a blend of both of the
forementioned bio-prospecting approaches, by surveying newlysolated cultures from diverse environments for the presence of theidirectional hydrogenase gene and a concurrent quantitative com-arison of their hydrogenase activities under non nitrogen-fixingonditions. We targeted cyanobacteria from terrestrial environ-ents, since no bidirectional hydrogenase genes originating in
hese environments were known from public databases, suggest-ng that they may have been differentially under-sampled. Marine
icrobial intertidal mats were also of special interest since a highux of hydrogen had been reported from these cyanobacterial matsSkyring et al., 1989; Hoehler et al., 2001). To this we added aurvey of freshwater plankton, a habitat well known to harbor
yanobacteria with bidirectional hydrogenases and hydrogen pro-ucing capabilities (Schmitz et al., 1995; Appel et al., 2000; Schutzt al., 2004; Allahverdiyeva et al., 2010). We deemed it importanto carry out these determinations in newly isolated strains, givennology 162 (2012) 105– 114
the tendency for phenotype loss that may ensue during continuedcultivation in the laboratory.
2. Materials and methods
2.1. Sampling and isolation of strains.
Cyanobacterial strains were isolated from terrestrial, freshwater plankton and marine benthos. The terrestrial cyanobacte-rial strains were isolated from different layers of a gypsum outcrop(34◦19′12′′N; 106◦42′39.1′′S) protruding from the biological soilcrust in the Chihuahuan Desert at the Sevilleta LTER, Palo DuroCanyon, NM, U.S.A. Pulverized sample from each layer was usedto inoculate liquid BG11 media, with 0.1 g/L cyclohexamide to pre-vent fungal growth. Strains were isolated from these enrichmentsby repeated streaking on petri dishes (Rippka, 1988). The fresh-water cyanobacterial strains were isolated from the epilimneticwaters of Saguaro Lake (33◦33′56′′N, 111◦32′10′′W), Lake Roo-sevelt (33◦40′18′′N, 111◦09′40′′W) and from the benthos of a coastalhot spring in Puertecitos, Baja California, Mexico (30◦20′46.6′′N;114◦38′11.2′′S). Water was filtered onto GF/F filters, and thebiomass on the filter used to inoculate liquid BG11 media. Repeatedstreaking as above led to isolation of clonal strains of cyanobacte-ria. Marine microbial intertidal mats were sampled from GuerreroNegro, Baja California Sur, Mexico, with the low- and mid-tide matsbeing collected at 27◦45′32.2′′N; 113◦59′51.3′′W and the high-tidemat at 27◦44′34.6′′N; 113◦57′2.5′′W (Rothrock and Garcia-Pichel,2005). Small pieces of low, mid and high tide mats were inoculateddirectly on plates of IMR media (Eppley et al., 1968) solidified with1% noble agar. Filaments of cyanobacteria moved or grew out of theinoculum and into the agar from where they could be sequentiallyplated. As observed previously (Garcia-Pichel et al., 1996), placingthe field sample on the media, selected for fast growing oppor-tunistic cyanobacterial species instead of the major mat-formingcyanobacteria. Therefore pieces of these mats were also viewedunder a dissecting microscope and filaments which morpholog-ically resembled Microcoleus (Garcia-Pichel et al., 1996), Lyngbya(Castenholz, 2001) and Calothrix (Rippka et al., 1979) were pulledout by micromanipulation using watchmaker’s forceps, draggedthough the agar for cleaning (Prufert-Bebout and Garcia-Pichel,1994) and allowed to grow on fresh media. Thereafter, a singlemotile filament (hormogonium) was picked and allowed to growinto a culture, three consecutive times, to establish a clonal culture.In addition, other strains available in the laboratory or obtainedfrom public culture collections were used to complement the sur-vey or to serve as positive or negative controls. All strains used inthis study were clonal and mono-cyanobacterial, but many con-tained contaminant heterotrophic bacteria. For experimentation,the level of contamination was monitored by microscopic observa-tion under phase contrast optics, and did not exceed 0.01% of thecyanobacterial biomass (assessed as bio-volume).
2.2. Cultivation and maintenance
All non-marine isolates were grown on BG11 medium (Rippkaet al., 1979) except for those that could fix nitrogen, which weregrown on a nitrate and ammonium free version, BG11◦ medium.A 1:1 mixture of medium IMR and ASN III medium (Eppleyet al., 1968) set at 3% salinity worked best for all Microcoleusstrains. For the rest of marine strains IMR media (3%) was used.For strains Chlorogloeopsis O-89-Cgs., Scytonema hyalinum FGP-
7A, Microcoleus vaginatus 9802, Chroococcidiopsis sp. CCMEE 029,Calothrix BECD30, Mastigocoleus BC008, Euhalothece MPI N303 andMicrocoleus chthonoplastes NDN-I media was used as described foreach (Garcia-Pichel and Castenholz, 1991; Yeager et al., 2007; Billi
iotech
e1cNenpp
2
PetTnnp(cs(atiAqQgIicsk
sMwaloRnlacmwsn
2
rwacTfwbots
A. Kothari et al. / Journal of B
t al., 2000; Chacon et al., 2006; Garcia-Pichel et al., 1998, 2010,996). For PCC strains, media used were as recommended by theulture collection. All cultures were supplemented with 0.5 �MiSO4, not always present in all formulations, and necessary tonsure a supply of Ni for appropriate function of Ni–Fe hydroge-ases. Cultures were maintained in appropriate standard medialates with 1% agar at 28 ◦C and at a light intensity of 20 �molhoton m−2 s−1.
.3. Molecular analyses
Genomic DNA preparations were obtained from all strains afterCI (phenol; chloroform; isoamyl alcohol) extraction (Countwayt al., 2005). DNA in the extracts was quantified using fluorome-ry of ethidium bromide-stained 1% agarose electrophoresis gels.o verify genetic homogeneity, and to have an idea of the phyloge-etic placement, we amplified 16S rRNA genes fragments from eachewly isolated strains. For this we used the cyanobacteria-specificrimers (CYA106F and CYA781R) and amplification conditions ofNübel et al., 1997). The presence of bidirectional hydrogenase-oding genes was assessed by PCR amplification using the hoxHpecific primers and amplification protocols described by BarzBarz et al., 2010). A Bio-Rad iCycler Thermal Cycler was used forll PCR reactions. All reactions included a positive and a nega-ive control. Any reactions yielding no amplification were repeatedndependently three times, before reporting results as negative.mplification products were documented on 1% agarose gels foruantity and expected size. Excised bands were purified using theIAquick Gel Extraction Kit (Qiagen Sample and Assay Technolo-ies) and used as template for commercial Sanger sequencing.n most cases both forward and reverse primers were used forndependent sequencing reactions of the same template, and theonsensus sequence used for further analysis. NCBI nucleotide blastearches were performed to assign the phylogenetically closestnown/named strains to those newly isolated and sequenced here.
In addition, this tentative placement based on 16S rRNA geneequence was confirmed by phylogenetic reconstructions usingEGA 5 (Tamura et al., 2011). About 600 bp of 16S rRNA sequencesere aligned using ClustalW. The alignment was manually curated,
nd Kimura 2 parameter model was used to construct maximumikelihood trees with 1000 bootstrap replicates. Since the sequencesbtained from Chroococcidiopsis RP110, 114, 116, 118, TolypothrixP 102 and Leptolyngbya RP108 were less than 400 bp, they wereot used in the analysis. For hoxH, the DNA sequences were trans-
ated into amino acid sequence and aligned by ClustalW. Thelignment was manually checked and all the hoxH sequences wereropped to a common length of 300 amino acids. MEGA 5 deter-ined WAG to be the best model, and a maximum likelihood treeith 1000 bootstrap replicates was constructed using it. Since the
equence obtained from Anabaena 901 was less than 100 AA, it wasot used in the phylogenetic analysis.
.4. Standard assay for hydrogen production
Most cultures did not grow as homogenous suspensions, butather as biofilms or clumps. Batch cultures of 100 ml volumeere grown in cotton-plugged 250 ml Erlenmeyer flasks at 28 ◦C
nd 20 �mol photon m−2 s−1, without additional bubbling. Smalllumps of biomass from log phase cultures were used for the assay.he clumps were loosened by means of forceps, suspended inresh medium, and placed in a custom made, air tight chamberith continuous stirring, so as to achieve biomass concentrations
etween 3 and 10 �g (Chl a) ml−1. Chl a was measured a posteri-ri. The media used for specific hydrogenase activity assay washe same as that used for their growth, except for nitrogen fixingtrains. The latter were measured in N-replete BG11 medium. The
nology 162 (2012) 105– 114 107
chamber was endowed with a miniature Clark-type electrode tomonitor hydrogen partial pressure. The electrode was connected toa pico-ammeter set at a voltage of 0.8 V. An A/D converter allowedthe current signal data to be read on a computer using Sensor traceBasic software. All electrodes and peripherals were from Unisense,Aarhus, Denmark. Before each measurement, the electrode wassubject to a 2-point calibration in culture medium bubbled witheither air (0% H2) or bubbled with a custom gas mixture of 10% H2in N2. Media sans cyanobacteria constituted a negative control. Allmeasurements were made in airtight glass chambers provided byUnisense. Controls were routinely run and there was always neg-ligible leakage. The specific hydrogenase activity assay was run indark using whole cells (in vivo) by the addition of methyl viologen(5 mM, final concentration) and freshly prepared sodium dithionite(10 mM, final concentration), (Appel et al., 2000). Sodium dithionitemakes the assay solution anaerobic (Jhaveri and Sharma, 1968) andreduces methyl viologen (Mayhew, 1978). The reduced methyl vio-logen acts as an electron carrier, supplying the reducing equivalentsto the bidirectional hydrogenase eliciting hydrogen production(Houchins and Burris, 1981; Houchins, 1984). The assay solutionswere always bluish at the end of the 24 h assays, indicating the con-tinued presence of reduced methyl viologen. Chlorophyll contentwas measured sacrificially in the spent suspension after extractionwith 100% methanol (MacKinney, 1941).
3. Results
3.1. Diversity of the set of strains surveyed
The survey involved a total of 36 strains from diverse envi-ronments. Table 1 gathers information on origin and taxonomicassignments. Strains are arranged there by ecological origin. In all,11 strains were of terrestrial origin, 7 from freshwater systemsand 18 originated in the marine intertidal benthos. In certain casesmultiple isolates from a single genus/species were obtained andstudied, which allowed us to probe internal variability. In termsof taxonomic diversity, all major groups were represented, butnot equally. Most strains (19) belong to Subsection III (i.e., orderOscillatoriales), 8 are in Section IV (order Nostocales), and 5 arein Section II (Pleurocapsales). Sections I (Chroococcales) and VI(Stigonematales) are only represented each by 2 strains. The phy-logenetic placement of the strains according to 16S rRNA genetrees indicate that the survey encompasses also a wide represen-tation of cyanobacterial biodiversity (Supplementary Information.1), but that it is not exhaustive in its coverage as several impor-tant phylogenetic clusters are not encompassed. Light microscopicimages depicting the morphology of strains surveyed can be seenin Fig. 1. Morphological diversity in this set of strains spans mostof that known for all cyanobacteria, from simple unicells (such asSynechocystis sp.) to large, complex, multicellular, branching fila-mentous strains (such as Mastigocoleus sp.).
3.2. Identity of the isolates and phylogenetic placements.
In most cases, PCR amplification of 16S rRNA genes yieldedclean products of about 600 bp in length that resulted in good-quality Sanger sequences with no background and few hard tocall bases (less than 1% undefined bases), indicating the absenceof mixed or multiple alleles in the amplificate and corroboratingthat each strain was likely monocyanobacterial, as also indicatedby microscopic observation. In a few cases, however, we obtained
multiple sequences. Such strains were submitted to further roundsof purification until a complete separation was achieved; thosethat did not result in a complete separation were not used inthis study. BLAST analyses against public databases of 16S rRNA
108A
. K
othari et
al. /
Journal of
Biotechnology 162 (2012) 105– 114
Table 1Cyanobacterial strains used in this survey, with their origin, morphological and phylogenetic assignments and the presence of hoxH in their genome as determined by PCR amplification.
Origin Strain Morphogenusa Closest 16S rRNA-based cultivated relativeb Similarity(%)c
hoxHpresence
Closest hoxH alleleb Similarity(%)c
Reference
Terrestrial desertRP110 Chroococcidiopsis Chroococcidiopsis sp. CC1 95 – – – This studyRP114 Chroococcidiopsis Pleurocapsa concharum 89 – – – This studyRP116 Chroococcidiopsis Chroococcidiopsis sp. CC1 93 – – – This study
This studyRP118 Chroococcidiopsis Chroococcidiopsis sp. CC1 95 – – – This studyRP102 Tolypothrix Tolypothrix sp. LQ-10 98 – – – This studyRP108 Leptolyngbya Leptolyngbya sp. CCALA 094 98 – – – This studyFGP-7A Scytonema NAd NA – – – (Yeager et al., 2007)PCC 9802 Microcoleus vaginatus NA NA – – – (Garcia-Pichel et al., 1996)CCMEE 029 Chroococcidiopsis NA NA – – – (Billi et al., 2000)
Concrete sidewalk O-89-Cgs Chlorogloeopsis NA NA – – – (Garcia-Pichel andCastenholz, 1991)
Soil/Cycad symbiont PCC 73102 Nostoc NA NA – – – Rippka et al., 1979
FreshwaterPond PCC 6803 Synechocystis sp. NA NA + Synechocystis PCC6803 100 –Lake plankton 901 Anabaena/Nostoc Anabaena oscillarioides BO HINDAK 1984/43 97 + Lyngbya majuscula CCAP 1446/4 83 This study
081 Oscillatoria Leptolyngbya sp. LEGE 92 + Lyngbya majuscula CCAP 1446/4 69 This study005 Oscillatoria Limnothrix redekei 2LT25S01 100 + Spirulina subsalsa FACHB351 77 This study
Hot Spring NH Anabaena/Nostoc Nostoc sp. ‘Mollenhauer 1:1–067’ 98 – – – This study011 Oscillatoria Limnothrix redekei 2LT25S01 100 + Spirulina subsalsa FACHB351 77 This studyHS Oscillatoria Limnothrix redekei 2LT25S01 99 + Lyngbya majuscula CCAP1446/4 70 This study
Marine benthosMicrobial mat BL G Lyngbya Lyngbya aestuarii PCC7419 99 + Lyngbya majuscula CCAP1446/4 89 This study
BL E Lyngbya Lyngbya aestuarii PCC7419 99 + Lyngbya majuscula CCAP1446/4 89 This studyBL J Lyngbya Lyngbya aestuarii PCC7419 99 + Lyngbya majuscula CCAP1446/4 89 This studyBL AA Lyngbya Lyngbya aestuarii PCC7419 99 + Lyngbya majuscula CCAP1446/4 89 This studyBM 001 Microcoleus Microcoleus chthonoplastes CCY9602 100 + Spirulina subsalsa FACHB351 72 This studyBM 002 Microcoleus Microcoleus chthonoplastes CCY9602 100 + Spirulina subsulsa FACHB351 72 This studyBM 003 Microcoleus Microcoleus chthonoplastes CCY9602 100 + Spirulina subsalsa FACHB351 72 This studyBC 2 Calothrix Calothrix sp. BC001 97 – – – This studyBC 4 Calothrix Calothrix sp. BC001 97 – – – This study7Y Oscillatoria Spirulina sp. SCyano57 93 – – – This study7C Oscillatoria Spirulina sp. SCyano57 93 – – – This studyG066 Oscillatoria Geitlerinema sp. kopara-FE 99 + ND This studyPCC 7419 Lyngbya NA NA + Lyngbya majuscula CAP1446/4 92 –NDN-I Microcoleus NA NA + Spirulina subsalsa FACHB351 76 (Garcia-Pichel et al., 1996)PCC 7420 Microcoleus NA NA + Microcoleus chthonoplastes PCC7420 100 (Garcia-Pichel et al., 1996)MPI N303 Euhalothece NA NA + Euhalothece halophytica ATCC43922 82 (Garcia-Pichel et al., 1998)
Carbonate surfaces BECD30 Calothrix sp. NA NA – – – (Chacon et al., 2006)BC008 Mastigocoleus testarum NA NA – – – (Garcia-Pichel et al., 2010)
ND for not determined.a Assigned according to Bergey’s manual (Castenholz, 2001).b According to NCBI BLAST search.c Of the isolate’s sequence to that of the closest cultivated strain in database.d NA stands for not applicable.

A. Kothari et al. / Journal of Biotechnology 162 (2012) 105– 114 109
F . (A) CR rocoleL aloth
smbBIcnlOlb
3
fsSphtaln
ig. 1. Morphological diversity of the strains surveyed. Scale bars represent 10 �mP108, a thin filamentous oscillatorian. (D) Mastigocoleus testarum BC003. (E) Micimnothrix sp. 081. (I) Oscillatoria sp. 7C. (J) Microcoleus chthonoplastes BM 001. (K) C
equences were used to confirm the validity of the generic assign-ents based on morphology. In general the assignments obtained
y BLAST corresponded to the morphogenus assignment based onergey’s Manual, Phylum BX Cyanobacteria section (see Table 1).
n a few cases, when the closest match had low similarity to anyultivated strains (7Y, 7C and RP114), morphogenus and phyloge-etic assignment differed. This is due to the fact that the isolates
ikely represent novel cyanobacterial taxa in need of description.ther mismatches were between thin filamentous strains (Oscil-
atoria/Leptolyngbya/Geitlerinema) where the microbiological andotanical nomenclatural traditions may differ.
.3. Patterns of hoxH detectability
We sought to detect the presence the hoxH gene that codesor the H subunit of the bidirectional NiFe hydrogenase, usingpecific PCR amplification. Four strains were included as controls.ynechocystis PCC 6803 (Yang and Mcfadden, 1994) and M. chthono-lastes PCC 7420 were known to posses hox operons containing aoxH gene in their genome, and were used as gene positive con-
rol, whereas Nostoc punctiforme PCC 73102 (Meeks et al., 2001)nd M. vaginatus PCC 9802 (Starkenburg et al., 2011) lacked homo-ogues in their completely sequenced genomes and were used asegative controls. All controls behaved as predicted (Table 1). Ofhroococcidiopsis RP114. (B) Tolypothrix RP102, detail of filament. (C) Leptolyngbyaus vaginatus PCC 9802. (F) Euhalothece sp. MPI N303. (G) Limnothrix sp. 005. (H)rix sp. BC2, with terminal heterocysts. (L) Lyngbya sp. BL J.
the strains tested, approximately half were positive for hoxH. Cer-tain clear patterns of distribution of hoxH emerged based on thisanalysis. While negatives for hoxH were seen in all environments(Table 1), none of the cyanobacterial strains from terrestrial envi-ronments possessed amplifiable homologues. In contrast, 87% ofthe strains isolated from fresh water contained the gene. Marinebenthic strains displayed 67% hoxH gene positives. The patterns ofdistribution of hoxH sequence obtained from multiple strain iso-lates of a single genus/species displayed a high level of consistency(i.e., all M. chthonoplastes were positive, while all Chroococcidiop-sis sp. were negative). In addition all strains in the same genusthat were isolated from the same environment had 100% sequencesimilarity. At higher taxonomic level there were no discernible pat-terns. For example Group IV (Nostocales) cyanobacteria containedboth positive strains and negative strains, as did Group III. Fig. 2depicts a maximum likelihood hoxH tree based on 300 amino acidlong sequences from 36 strains, of which 14 were novel. Well-supported clades of Arthospira, Limnothrix, Lyngbya and Microcoleushox sequences could be resolved.
3.4. Physiology of hydrogen production
A total of 32 strains were tested for hydrogen production viathe hydrogenase assay. Approximately half of the strains tested

110 A. Kothari et al. / Journal of Biotechnology 162 (2012) 105– 114
Fig. 2. Maximum likelihood tree of cyanobacterial sequences of the hoxH subunit in the bidirectional Ni–Fe hydrogenase, based on 300 amino acids. Bootstrap values (%) forw trees.c
pbhNFmu1v8GcBs
ell supported nodes (>50) are indicated, obtained from analyses of 1000 replicate
lades are indicated to the right.
roduced detectable hydrogen (Table 2). A direct correlationetween the presence of hoxH (Table 1) and the ability to evolveydrogen in the assay was evident from all the strains tested.o evolution was detected in any hoxH-negative strains, either.or quantitative comparisons, we determined RH – the maxi-al (typically also the initial) rate of hydrogen production per
nit biomass. The range in RH (gathered in Table 2; between4 and 156 nmol �g [Chl a]−1 h−1) was wider than the range ofalues already reported in the literature, typically from 30 to0 nmol (�g Chl a)−1 h−1 (Appel et al., 2006; Gutekunst et al., 2006;
utthann et al., 2007; Schutz et al., 2004). The highest values wereonsistently seen in marine intertidal Lyngbya isolates, with strainL J displaying average rates as high as five fold faster than thoseeen in Synechocystis PCC 6803 under identical assay conditions,The sequence from Chloroflexus aurantiacus J-10-fl was used as outgroup. Coherent
but also with considerable variability among Lyngbya strains. In all,however, this is only a moderate improvement with respect to RH.
We also quantified certain other aspects of the dynamics of H2production that have not usually been considered or reported indetail in the literature. As advanced in the introduction, the pro-duction of hydrogen is reported to be only transient, and, afterH2 reaches a certain maximum hydrogen concentration, [H2]M attime TR, H2 production stops and reverts to consumption, bringingconcentrations to very low levels or even consuming all availablehydrogen. These dynamics are exemplified in Synechocystis PCC
6803, shown in Fig. 3a. We found, however, that these parametersvaried widely among the different strains (Table 2). Amongst theH2 producing strains, 2 distinct dynamic patterns of H2 productioncould be distinguished, which we call Patterns 1 and 2. Pattern 1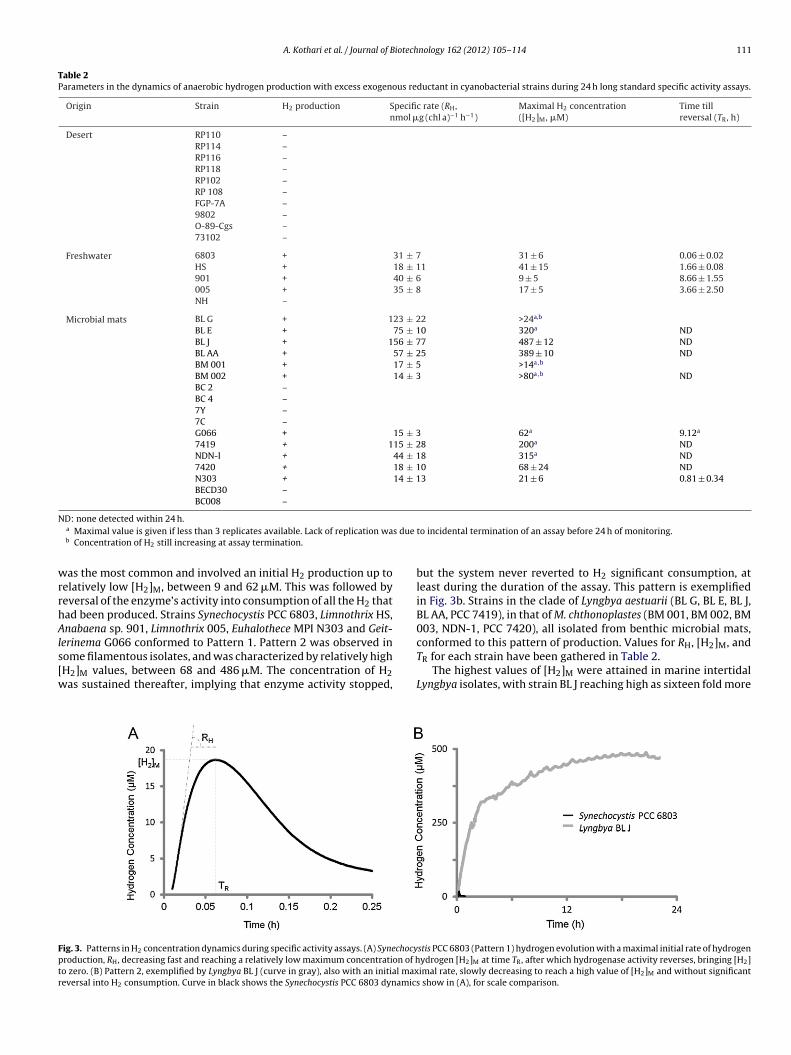
A. Kothari et al. / Journal of Biotechnology 162 (2012) 105– 114 111
Table 2Parameters in the dynamics of anaerobic hydrogen production with excess exogenous reductant in cyanobacterial strains during 24 h long standard specific activity assays.
Origin Strain H2 production Specific rate (RH,nmol �g (chl a)−1 h−1)
Maximal H2 concentration([H2]M, �M)
Time tillreversal (TR, h)
Desert RP110 –RP114 –RP116 –RP118 –RP102 –RP 108 –FGP-7A –9802 –O-89-Cgs –73102 –
Freshwater 6803 + 31 ± 7 31 ± 6 0.06 ± 0.02HS + 18 ± 11 41 ± 15 1.66 ± 0.08901 + 40 ± 6 9 ± 5 8.66 ± 1.55005 + 35 ± 8 17 ± 5 3.66 ± 2.50NH –
Microbial mats BL G + 123 ± 22 >24a,b
BL E + 75 ± 10 320a NDBL J + 156 ± 77 487 ± 12 NDBL AA + 57 ± 25 389 ± 10 NDBM 001 + 17 ± 5 >14a,b
BM 002 + 14 ± 3 >80a,b NDBC 2 –BC 4 –7Y –7C –G066 + 15 ± 3 62a 9.12a
7419 + 115 ± 28 200a NDNDN-I + 44 ± 18 315a ND7420 + 18 ± 10 68 ± 24 NDN303 + 14 ± 13 21 ± 6 0.81 ± 0.34BECD30 –BC008 –
Ns due
wrrhAls[w
Fptr
D: none detected within 24 h.a Maximal value is given if less than 3 replicates available. Lack of replication wab Concentration of H2 still increasing at assay termination.
as the most common and involved an initial H2 production up toelatively low [H2]M, between 9 and 62 �M. This was followed byeversal of the enzyme’s activity into consumption of all the H2 thatad been produced. Strains Synechocystis PCC 6803, Limnothrix HS,nabaena sp. 901, Limnothrix 005, Euhalothece MPI N303 and Geit-
erinema G066 conformed to Pattern 1. Pattern 2 was observed in
ome filamentous isolates, and was characterized by relatively highH2]M values, between 68 and 486 �M. The concentration of H2as sustained thereafter, implying that enzyme activity stopped,ig. 3. Patterns in H2 concentration dynamics during specific activity assays. (A) Synechocyroduction, RH, decreasing fast and reaching a relatively low maximum concentration of ho zero. (B) Pattern 2, exemplified by Lyngbya BL J (curve in gray), also with an initial maxeversal into H2 consumption. Curve in black shows the Synechocystis PCC 6803 dynamics
to incidental termination of an assay before 24 h of monitoring.
but the system never reverted to H2 significant consumption, atleast during the duration of the assay. This pattern is exemplifiedin Fig. 3b. Strains in the clade of Lyngbya aestuarii (BL G, BL E, BL J,BL AA, PCC 7419), in that of M. chthonoplastes (BM 001, BM 002, BM003, NDN-1, PCC 7420), all isolated from benthic microbial mats,conformed to this pattern of production. Values for RH, [H2]M, and
TR for each strain have been gathered in Table 2.The highest values of [H2]M were attained in marine intertidalLyngbya isolates, with strain BL J reaching high as sixteen fold more
stis PCC 6803 (Pattern 1) hydrogen evolution with a maximal initial rate of hydrogenydrogen [H2]M at time TR, after which hydrogenase activity reverses, bringing [H2]imal rate, slowly decreasing to reach a high value of [H2]M and without significant
show in (A), for scale comparison.

1 iotech
tcsai
4
orsanfoiInvrt
cltEowoodotfsca(sarar
actfhhcfhPthresih
nO
12 A. Kothari et al. / Journal of B
han that seen in Synechocystis PCC 6803 under identical assayonditions, but also with considerable variability among Lyngbyatrains. This coupled with the lack of reversal of enzyme under thessay conditions in Microcoleus and Lyngbya strains is a significantmprovement with respect to [H2]M and TR.
. Discussion
Ours is the first survey to encompass concurrent assessmentf the hoxH gene presence and H2 evolution capacity from excesseductant amongst newly isolated cyanobacterial strains. It isignificant that the correspondence between gene presence andctivity was absolute (Table 2). Since whenever hoxH was absent,o H2 could be detected, it is likely that this enzyme is responsible
or the activity in a vast majority of cyanobacteria. Nitrogenase, thenly other hydrogen-producing enzyme known in cyanobacteria,s not expressed in nitrogen replete conditions used in our assays.n addition, it has been established in literature that the nitroge-ase is not capable of evolving hydrogen in the presence of methyliologen and sodium dithionite (Daday et al., 1979). Thus, the bidi-ectional hydrogense is the most probable source of hydrogen inhis assay.
Phylogenetic analyses of hoxH confirmed the degree of overallonservation, and allowed us to increase the resolution of the evo-utionary history of the enzyme within oxygenic phototrophs, fromhose that have been presented elsewhere (see Vignais et al., 2001).ven though the tree was not well-resolved with respect to higher-rder relationships among clades, all sequences fit clearly andithin the “cyanobacterial” lineage, and well-populated clusters
f strains seemed to contain consistently closely related versionsf the gene (Fig. 3). This seems to speak for an evolutionary pathevoid of events of lateral transfer. This is possibly a consequencef the rather complex composition of the hox operon itself, and tohe presence of necessary “maturation’ systems that help obtainunctional enzymes (Hoffmann et al., 2006). Among the databaseequences in Fig. 2, there does seem to be a couple of cases oflear “misplacement”: a Mastigocladus sequence, which should bemong the heterocystous clade, falls among a group of unicellularsSynechocystis, Microcystis, Cyanothece etc.), and a single Arthrospiratrain falls away from its main clade. It is hard to interpret this datas being due to lateral transfer events, rather than just being theesult of database entry or other analytical errors, and it woulddvisable to re-sequence those two strains before attempting toead too much into it.
Interestingly, this Ni–Fe hydrogenase is clearly not universalmong cyanobacteria. Our results and those of others tend toonverge in a figure around 50% incidence. Clearly the bidirec-ional hydrogenase system is subject to facile evolutionary lossrom cyanobacterial genomes. We could show, as other have alsointed (Barz et al., 2010) that the prevalence of these bidirectionalydrogenase genes was clearly not independent from habitat. Weurrently have no satisfactory explanation as to why cyanobacteriarom terrestrial environments are virtually devoid of bidirectionalydrogenases [we hesitate to write completely, given that NostocCC 7422, a symbiont with cycads and probably having a terres-rial part of its life cycle is known to possess a known bidirectionalydrogenase gene (Yoshino et al., 2007)]. It is possible that ter-estrial environments, which tend to be oxic, may select againstnzymes that are involved in anaerobic pathways or are very sen-itive to oxygen, but then again, the epilimnion of freshwater lakess also a rather oxygenated environment, and yet the incidence of
oxH-positive strains there was quite large.With respect to the discovery of novel strains of high biotech-ological potential our findings had two clearly different aspects.n the one hand, we did not detect maximal specific activities
nology 162 (2012) 105– 114
that were much faster than those previously reported in the lit-erature. Lyngbya BL J was 5 fold more potent than SynechocystisPCC 6803. However, in the literature, a range of specific hydro-genase activities have been reported for Synechocystis PCC 6803itself (Appel et al., 2006; Baebprasert et al., 2010; Gutekunst et al.,2006; Gutthann et al., 2007; Schutz et al., 2004) While most arein the range of several tens of nmol (H2) per �g Chl a and hour,Schutz et al. reported as much as 235. It should be noted that it isdifficult to compare the absolute values of hydrogenase activitiesobtained from different labs due to difference in growth condi-tions, experimental set ups and methods of measuring hydrogenproduction. If one takes the typical variability into account, theimprovement may be just a mere two fold; including the high-level outlier value, no improvement at all. However, the fact thatwe discovered the presence of “Pattern 2” cyanobacteria, havingbidirectional hydrogenases that run unidirectionally under excessreductant, may be quite relevant for potential biotechnologicalapplications. Cyanobacteria in this class can accumulate high con-centrations of hydrogen in the medium, thus facilitating eventualhydrogen harvesting in large-scale processes. Unfortunately, allstrains conforming to Pattern 2, belong to the Oscillatoriales, (fila-mentous, non-heterocystous cyanobacteria), which are notoriouslydifficult to grow in large scale, and are not yet amenable to even thesimplest forms of genetic modification. Therefore future attemptsto use these cyanobacteria as platform for hydrogen productionwill have to rely on optimization through variations of basic culti-vation and physiological parameters. As an immediate goal, it willbe imperative to carry out a detailed study of the enzymology, phys-iology and the molecular basis of hydrogen generation in modelsstrain of Pattern 2. Differences in genomic sequence and architec-ture, enzyme maturation, and regulation of the enzyme must beunderstood. We note that a simple comparison of the 300 aminoacids in our hoxH alignment did not reveal any obvious differencein sequence between the Pattern 1 and 2 cyanobacteria. Heterolo-gous expression of Pattern 2 hydrogen generating systems undercontrolled conditions in model strains such as Synechocystis PCC6803 may eventually constitute the best platform.
Contribution
Ferran Garcia-Pichel and Ankita Kothari provided the concept.Isolation of strains from desert and their 16s gene amplificationwas carried out by Ruth Potrafka. Isolation of strains from freshwa-ter and marine microbial mats was by AK, who also carried out allother molecular, physiological and phylogenetic work. AK and FGPwrote the manuscript. All authors have read and approved the finalmanuscript.
Acknowledgements
This research and A.K were supported by an endowment fromB. Swette through the ASU President’s Fusion Fund. The fundingsource had no involvement in experimental design, data collection,analysis, report writing or decision to submit article.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.jbiotec.2012.04.017.
References
Akkerman, I., Janssen, M., Rocha, J., Wijffels, R.H., 2002. Photobiological hydrogenproduction: photochemical efficiency and bioreactor design. International Jour-nal of Hydrogen Energy 27, 1195–1208.

iotech
A
A
A
A
B
B
B
B
C
C
C
C
C
D
D
E
E
F
G
G
G
G
G
G
G
H
H
H
A. Kothari et al. / Journal of B
llahverdiyeva, Y., Leino, H., Saari, L., Fewer, D.P., Shunmugam, S., Sivonen, K., Aro,E.M., 2010. Screening for biohydrogen production by cyanobacteria isolatedfrom the Baltic Sea and Finnish lakes. International Journal of Hydrogen Energy35, 1117–1127.
ppel, J., Gutekunst, K., Hoffmann, D., Lommer, M., Egert, M., Suzuki, I., Schulz-Friedrich, R., 2006. Metal dependence and intracellular regulation of thebidirectional NiFe-hydrogenase in Synechocystis sp. PCC 6803. InternationalJournal of Hydrogen Energy 31, 1452–1459.
ppel, J., Phunpruch, S., Steinmuller, K., Schulz, R., 2000. The bidirectional hydro-genase of Synechocystis sp. PCC 6803 works as an electron valve duringphotosynthesis. Archives of Microbiology 173, 333–338.
ppel, J., Schulz, R., 1998. Hydrogen metabolism in organisms with oxygenic pho-tosynthesis: hydrogenases as important regulatory devices for a proper redoxpoising? Journal of Photochemistry and Photobiology B: Biology 47, 1–11.
aebprasert, W., Lindblad, P., Incharoensakdi, A., 2010. Response of H2 productionand hox-hydrogenase activity to external factors in the unicellular cyanobac-terium Synechocystis sp. strain PCC 6803. International Journal of HydrogenEnergy 35, 6611–6616.
andyopadhyay, A., Stockel, J., Min, H.T., Sherman, L.A., Pakrasi, H.B., 2010. High ratesof photobiological H2 production by a cyanobacterium under aerobic conditions.Nature Communications 1, 139.
arz, M., Beimgraben, C., Staller, T., Germer, F., Opitz, F., Marquardt, C., Schwarz,C., Gutekunst, K., Vanselow, K.H., Schmitz, R., LaRoche, J., Schulz, R., Appel, J.,2010. Distribution analysis of hydrogenases in surface waters of marine andfreshwater environments. Plos One 5, e13846.
illi, D., Friedmann, E.I., Hofer, K.G., Caiola, M.G., Ocampo-Friedmann, R.,2000. Ionizing-radiation resistance in the desiccation-tolerant cyanobacteriumChroococcidiopsis. Applied and Environmental Microbiology 66, 1489–1492.
arrieri, D.W.K., Eckert, C., Yu, J., Maness, P.C., 2011. The role of the bidirectionalhydrogenase in cyanobacteria. Bioresource Technology 102, 8368–8377.
astenholz, R.W., 2001. Phylum BX. Cyanobacteria. In: Bergey’s Manual of System-atic Bacteriology, 2nd edn, pp. 473–487.
hacon, E., Berrendero, E., Pichel, F.G., 2006. Biogeological signatures of microboringcyanobacterial communities in marine carbonates from Cabo Rojo, Puerto Rico.Sedimentary Geology 185, 215–228.
ountway, P.D., Gast, R.J., Savai, P., Caron, D.A., 2005. Protistan diversity estimatesbased on 18S rDNA from seawater incubations in the western North Atlantic.Journal of Eukaryotic Microbiology 52, 95–106.
ournac, L., Guedeney, G., Peltier, G., Vignais, P.M., 2004. Sustained photoevolutionof molecular hydrogen in a mutant of Synechocystis sp. strain PCC 6803 defi-cient in the type I NADPH-dehydrogenase complex. Journal of Bacteriology 186,1737–1746.
aday, A., Lambert, G.R., Smith, G.D., 1979. Measurement in vivo of hydrogenase-catalysed hydrogen evolution in the presence of nitrogenase enzyme incyanobacteria. Biochemical Journal 177, 139–144.
’Eustachio, A.J., Hardy, R.W.F., 1964. Reductants and electron transport in nitrogenfixation. Biochemical and Biophysical Research Communications 15, 319–323.
isbrenner, G., Evans, H.J., 1983. Aspects of hydrogen metabolism in nitrogen-fixinglegumes and other plant-microbe associations. Annual Review of Plant Physiol-ogy 34, 105–136.
ppley, R.W., Holmes, R.W., Strickland, J.D.H., 1968. Sinking rates of marine phyto-plankton measured with a fluorometer. Journal of Experimental Marine Biologyand Ecology 1, 191–208.
ujita, Y., Myers, J., 1965. Hydrogenase and NADP-reduction reactions by a cell-freepreparation of Anabaena cylindrica. Archives of Biochemistry and Biophysics 111,619–625.
arcia-Pichel, F., Castenholz, R.W., 1991. Characterization and biological implica-tions of scytonemin, a cyanobacterial sheath pigment. Journal of Phycology 27,395–409.
arcia-Pichel, F., Nübel, U., Muyzer, G., 1998. The phylogeny of unicellular, extremelyhalotolerant cyanobacteria. Archives of Microbiology 169, 469–482.
arcia-Pichel, F., Ramirez-Reinat, E., Gao, Q.J., 2010. Microbial excavation of solidcarbonates powered by P-type ATPase-mediated transcellular Ca2+ transport.Proceedings of the National Academy of Science of the United States of America107, 21749–21754.
arcia-Pichel, F., PrufertBebout, L., Muyzer, G., 1996. Phenotypic and phylogeneticanalyses show Microcoleus chthonoplastes to be a cosmopolitan cyanobacterium.Applied and Environmental Microbiology 62, 3284–3291.
hirardi, M.L., Posewitz, M.C., Maness, P.C., Dubini, A., Yu, J.P., Seibert, M., 2007.Hydrogenases and hydrogen photoproduction in oxygenic photosyntheticorganisms. Annual Review of Plant Biology 58, 71–91.
utekunst, K., Hoffmann, D., Lommer, M., Egert, M., Schulz–Friedrich, R., Appel,J., 2006. Metal dependence and intracellular regulation of the bidirectionalNiFe–hydrogenase in Synechocystis sp. PCC 6803. International Journal of Hydro-gen Energy 31, 1452–1459.
utthann, F., Egert, M., Marques, A., Appel, J., 2007. Inhibition of respiration andnitrate assimilation enhances photohydrogen evolution under low oxygenconcentrations in Synechocystis sp. PCC 6803. Biochimica et Biophysica Acta-Bioenergetics 1767, 161–169.
appe, T., Schutz, K., Bohme, H., 2000. Transcriptional and mutational analysis ofthe uptake hydrogenase of the filamentous cyanobacterium Anabaena variabilis
ATCC 29413. Journal of Bacteriology 182, 1624–1631.oehler, T.M., Bebout, B.M., Des Marais, D.J., 2001. The role of microbial mats in theproduction of reduced gases on the early Earth. Nature 412, 324–327.
offmann, D., Gutekunst, K., Klissenbauer, M., Schulz-Friedrich, R., Appel, J., 2006.Mutagenesis of hydrogenase accessory genes of Synechocystis sp. PCC 6803.
nology 162 (2012) 105– 114 113
Additional homologues of hypA and hypB are not active in hydrogenase matu-ration. The FEBS Journal 273, 4516–4527.
Houchins, J.P., 1984. The physiology and biochemistry of hydrogen metabolism incyanobacteria. Biochimica et Biophysica Acta 768, 227–255.
Houchins, J.P., Burris, R.H., 1981. Comparative characterization of 2 distinct hydro-genases from Anabaena sp. strain-7120. Journal of Bacteriology 146, 215–221.
Hyndman, L.A., Burris, R.H., Wilson, P.W., 1953. Properties of hydrogenase fromAzotobacter vinelandii. Journal of Bacteriology 65, 522–531.
Jhaveri, A.S., Sharma, M.M., 1968. Absorption of oxygen in aqueous alkaline solutionsof sodium dithionite. Chemical Engineering Science 23, 1–8.
Lindblad, P., Christensson, K., Lindberg, P., Fedorov, A., Pinto, F., Tsygankov, A., 2002.Photoproduction of H-2 by wildtype Anabaena PCC 7120 and a hydrogen uptakedeficient mutant: from laboratory experiments to outdoor culture. InternationalJournal of Hydrogen Energy 27, 1271–1281.
Ludwig, M., Schulz-Friedrich, R., Appel, J., 2006. Occurrence of hydrogenases incyanobacteria and anoxygenic photosynthetic bacteria: implications for thephylogenetic origin of cyanobacterial and algal hydrogenases. Journal of Molec-ular Evolution 63, 758–768.
MacKinney, G., 1941. Absorption of light by chlorophyll solutions. Journal of Biolog-ical Chemistry 140, 315–322.
Marques, A.E., Barbosa, A.T., Jotta, J., Coelho, M.C., Tamagnini, P., Gouveia, L., 2011.Biohydrogen production by Anabaena sp. PCC 7120 wild-type and mutantsunder different conditions: light, nickel, propane, carbon dioxide and nitrogen.Biomass and Bioenergy 35, 4426–4434.
Masukawa, H., Mochimaru, M., Sakurai, H., 2002. Hydrogenases and photobiologicalhydrogen production utilizing nitrogenase system in cyanobacteria. Interna-tional Journal of Hydrogen Energy 27, 1471–1474.
Mayhew, S.G., 1978. Redox potential of dithionite and So2- from equilibriumreactions with flavodoxins, methyl viologen and hydrogen plus hydrogenase.European Journal of Biochemistry 85, 535–547.
Meeks, J.C., Elhai, J., Thiel, T., Potts, M., Larimer, F., Lamerdin, J., Predki, P., Atlas,R., 2001. An overview of the genome of Nostoc punctiforme, a multicellular,symbiotic cyanobacterium. Photosynthesis Research 70, 85–106.
Nübel, U., Garcia-Pichel, F., Muyzer, G., 1997. PCR primers to amplify 16S rRNA genesfrom cyanobacteria. Applied and Environmental Microbiology 63, 3327–3332.
Oost, J., Bulthuis, B.A., Feitz, S., Krab, K., Kraayenhof, R., 1989. Fermentationmetabolism of the unicellular cyanobacterium Cyanothece PCC 7822. Archivesof Microbiology 152, 415–419.
Peterson, R.B., Burris, R.H., 1978. Hydrogen metabolism in isolated heterocysts ofAnabaena-7120. Archives of Microbiology 116, 125–132.
Phelps, A.S., Wilson, P.W., 1941. Occurrence of hydrogenase in nitrogen-fixing organ-isms. Proceedings of the Society for Experimental Biology and Medicine 47,473–476.
Prince, R.C., Kheshgi, H.S., 2005. The photobiological production of hydrogen:potential efficiency and effectiveness as a renewable fuel. Critical Reviews inMicrobiology 31, 19–31.
Prufert-Bebout, L., Garcia-Pichel, F., 1994. Field and cultivated Microcoleus chthono-plastes: the search for clues to its prevalence in marine microbial mats. In: NATOASI SERIES, G 35, pp. 111–116.
Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M., Stanier, R.Y., 1979. Genericassignments, strain histories and properties of pure cultures of cyanobacteria.Journal of General Microbiology 111, 1–61.
Rippka, R., 1988. Isolation and purification of cyanobacteria. Methods in Enzymology167, 3–27.
Rothrock, M.J., Garcia-Pichel, F., 2005. Microbial diversity of benthic mats along atidal desiccation gradient. Environmental Microbiology 7, 593–601.
Schmitz, O., Boison, G., Hilscher, R., Hundeshagen, B., Zimmer, W., Lottspeich, F.,Bothe, H., 1995. Molecular biological analysis of a bidirectional hydrogenasefrom Cyanobacteria. European Journal of Biochemistry 233, 266–276.
Schutz, K., Happe, T., Troshina, O., Lindblad, P., Leitao, E., Oliveira, P., Tamagnini,P., 2004. Cyanobacterial H-2 production—a comparative analysis. Planta 218,350–359.
Skyring, G.W., Lynch, R.M., Smith, G.D., 1989. Quantitative relationship betweencarbon, hydrogen and sulfur metabolism in cyanobacterial mats. In: Cohen, Y.,Rosenberg, E. (Eds.), Microbial Mats: Physiological Ecology of Benthic MicrobialCommunities. ASM, Washington, DC, pp. 170–179.
Starkenburg, S.R., Reitenga, K.G., Freitas, T., Johnson, S., Chain, P.S.G., Garcia-Pichel,F., Kuske, C.R., 2011. Genome of the cyanobacterium Microcoleus vaginatus FGP-2, a photosynthetic ecosystem engineer of arid land soil biocrusts worldwide.Journal of Bacteriology 193, 4569–4570.
Tamagnini, P., Leitao, E., Oliveira, P., Ferreira, D., Pinto, F., Harris, D.J., Heidorn, T.,Lindblad, P., 2007. Cyanobacterial hydrogenases: diversity, regulation and appli-cations. FEMS Microbiology Review 31, 692–720.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S., 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolu-tionary distance, and maximum parsimony methods. Molecular Biology andEvolution 28, 2731–2739.
Troshina, O., Serebryakova, L., Sheremetieva, M., Lindblad, P., 2002. Production ofH2 by the unicellular cyanobacterium Gloeocapsa alpicola CALU 743 during fer-mentation. International Journal of Hydrogen Energy 27, 1283–1289.
Vignais, P.M., Billoud, B., Meyer, J., 2001. Classification and phylogeny of hydroge-
nases. FEMS Microbiology Review 25, 455–501.Vignais, P.M., Colbeau, A., 2004. Molecular biology of microbial hydrogenases. Cur-rent Issues in Molecular Biology 6, 159–188.
Weaver, P.F., Lien, S., Seibert, M., 1980. Photobiological production of hydrogen. SolarEnergy 24, 3–45.

1 iotech
Y
Y
14 A. Kothari et al. / Journal of B
ang, X.Y., Mcfadden, B.A., 1994. The Complete DNA-sequence and replication anal-
ysis of the plasmid-Pcb2.4 from the cyanobacterium Synechocystis PCC 6803.Plasmid 31, 131–137.eager, C.M., Kornosky, J.L., Morgan, R.E., Cain, E.C., Garcia-Pichel, F., Housman,D.C., Belnap, J., Kuske, C.R., 2007. Three distinct clades of cultured heterocys-tous cyanobacteria constitute the dominant N-2-fixing members of biological
nology 162 (2012) 105– 114
soil crusts of the Colorado Plateau, USA. FEMS Microbiology Ecology 60,
85–97.Yoshino, F., Ikeda, H., Masukawa, H., Sakurai, H., 2007. High photobiologicalhydrogen production activity of a Nostoc sp. PCC 7422 uptake hydrogenase-deficient mutant with high nitrogenase activity. Marine Biotechnology 9,101–112.




















