
Integrating cortisol and isotopic analyses of archaeological hair: Elucidating juvenile ante-mortem...
18
Integrating Cortisol and Isotopic Analyses of Archeological Hair: Reconstructing Individual Experiences of Health and Stress Emily C. Webb, 1 * Christine D. White, 1 Stan Van Uum, 2 and Fred J. Longstaffe 3 1 Department of Anthropology, The University of Western Ontario, London, ON, Canada 2 Department of Medicine, The University of Western Ontario, London, ON, Canada 3 Department of Earth Sciences, The University of Western Ontario, London, ON, Canada KEY WORDS stress; cortisol; stable isotopes; mummies; hair ABSTRACT Archeological hair from 14 adults from the Nasca Region, Peru (c. AD1–1000) was analyzed for carbon and nitrogen isotopic compositions and cortisol levels. We investigated the relationship between isotopic compositions, which reflect diet, and cortisol, which reflects biogenic cortisol production and chronic stress. Using a case study approach, we determined that there are consistent changes in cortisol production associated with the rapid dietary change characteristic of local mobility. Moreover, changes in nitrogen- and carbon- isotope compositions, when integrated with cortisol lev- els, enabled inferences to be made about nitrogen metab- olism and carbon routing, and elucidated the nature of potential stressors in the months before death. The iso- topic and cortisol data suggested a relatively high rate of exposure to stress that is consistent with what is known about the Nasca Region social and physical envi- ronments. Of the 14 adults included in this study, six likely suffered from illness/trauma before death, and a further three experienced stress without an observable associated change in isotopic composition. Five individu- als also experienced increased stress related to local mobility, inferred from co-occurring changes in cortisol production and dietary shifting. The integration of corti- sol and isotopic data revealed individual characteristics of hidden frailty and risk that would not be apparent using more traditional methods of evaluating health sta- tus. This approach will provide a powerful enhancement to the understanding of stress, morbidity, and well-being developed through skeletal analysis. Am J Phys Anthro- pol 000:000–000, 2014. V C 2014 Wiley Periodicals, Inc. Hair retains a sequential archive of physical, environ- mental, and dietary information, and is a valuable resource for both archeological and clinical research investigating behavior, health, and stress. Carbon- and nitrogen-isotope analysis of hair in archeological contexts has been used to reconstruct seasonal dietary change, res- idential mobility, and geographic relocation, ante-mortem diet and seasonality of death, and to investigate the met- abolic impact of trauma and illness (White, 1993; Schwarcz and White, 2004; Knudson et al., 2007; White et al., 2009; Williams et al., 2011; Knudson et al., 2012; Williams and Katzenberg, 2012; Webb et al., 2013). Because hair can be noninvasively sampled, there is also a sizable body of clinical isotopic research focusing on starvation, dietary habits and nutrition, geographic relo- cation, pregnancy and specific diseases (O’Connell and Hedges, 1999; Bol and Pflieger, 2002; Fuller et al., 2004, 2005; McCullagh et al., 2005; Petzke et al., 2005, 2006; Mekota et al., 2006; Huelsemann et al., 2009; Mekota et al., 2009; Valenzuela et al., 2011). Cortisol, a hormone produced in response to stress, can also be measured in hair. Clinical studies have measured systemic cortisol lev- els during pregnancy, and explored the impact of chronic stress, perceived stress, and specific diseases on cortisol production (Kalra et al., 2007; Van Uum et al., 2008; Kirschbaum et al., 2009; Thomson et al., 2009; Gow et al., 2010; Dettenborn et al., 2012; Russell et al., 2012). Cortisol has also been successfully extracted from archeo- logical hair samples and has been used to document indi- vidual experiences of stress in the months preceding death (Webb et al., 2010). For this study, hair samples from 14 adults excavated at Cahuachi and near Huaca del Loro in the Southern Nasca Region of Peru (AD1-1000) were analyzed to determine carbon- and nitrogen-isotope compositions (Webb et al., 2013) and cortisol levels. Good biomolecular preservation has been established through a pilot study of Peruvian mummies from several sites, including the Nasca Region (Webb et al., 2010). The Nasca Region remains in particular were chosen for additional study because of the generally low frequency of stress markers on bones and teeth for previously analysed skeletal material from this context (Kellner, 2002; Cagigao, 2009), suggesting that understanding ante-mortem expe- riences may be critical to elucidating stress and death in Grant sponsor: Social Sciences and Humanities Research Council of Canada, the Natural Sciences and Engineering Research Council of Canada, the Canada Research Chairs Program, the Physicians’ Services Incorporated Foundation, Canada Foundation for Innova- tion and Ontario Research Fund. *Correspondence to: Emily Webb, Organic Geochemistry Unit, School of Chemistry, Cantock’s Close, University of Bristol, Bristol BS8 1TS, United Kingdom. E-mail: [email protected] Received 4 August 2014; revised 16 October 2014; accepted 12 November 2014 DOI: 10.1002/ajpa.22673 Published online 00 Month 2014 in Wiley Online Library (wileyonlinelibrary.com). Ó 2014 WILEY PERIODICALS, INC. AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 00:00–00 (2014)
Transcript of Integrating cortisol and isotopic analyses of archaeological hair: Elucidating juvenile ante-mortem...

Integrating Cortisol and Isotopic Analyses ofArcheological Hair: Reconstructing IndividualExperiences of Health and Stress
Emily C. Webb,1* Christine D. White,1 Stan Van Uum,2 and Fred J. Longstaffe3
1Department of Anthropology, The University of Western Ontario, London, ON, Canada2Department of Medicine, The University of Western Ontario, London, ON, Canada3Department of Earth Sciences, The University of Western Ontario, London, ON, Canada
KEY WORDS stress; cortisol; stable isotopes; mummies; hair
ABSTRACT Archeological hair from 14 adults fromthe Nasca Region, Peru (c. AD1–1000) was analyzed forcarbon and nitrogen isotopic compositions and cortisollevels. We investigated the relationship between isotopiccompositions, which reflect diet, and cortisol, whichreflects biogenic cortisol production and chronic stress.Using a case study approach, we determined that thereare consistent changes in cortisol production associatedwith the rapid dietary change characteristic of localmobility. Moreover, changes in nitrogen- and carbon-isotope compositions, when integrated with cortisol lev-els, enabled inferences to be made about nitrogen metab-olism and carbon routing, and elucidated the nature ofpotential stressors in the months before death. The iso-topic and cortisol data suggested a relatively high rateof exposure to stress that is consistent with what is
known about the Nasca Region social and physical envi-ronments. Of the 14 adults included in this study, sixlikely suffered from illness/trauma before death, and afurther three experienced stress without an observableassociated change in isotopic composition. Five individu-als also experienced increased stress related to localmobility, inferred from co-occurring changes in cortisolproduction and dietary shifting. The integration of corti-sol and isotopic data revealed individual characteristicsof hidden frailty and risk that would not be apparentusing more traditional methods of evaluating health sta-tus. This approach will provide a powerful enhancementto the understanding of stress, morbidity, and well-beingdeveloped through skeletal analysis. Am J Phys Anthro-pol 000:000–000, 2014. VC 2014 Wiley Periodicals, Inc.
Hair retains a sequential archive of physical, environ-mental, and dietary information, and is a valuableresource for both archeological and clinical researchinvestigating behavior, health, and stress. Carbon- andnitrogen-isotope analysis of hair in archeological contextshas been used to reconstruct seasonal dietary change, res-idential mobility, and geographic relocation, ante-mortemdiet and seasonality of death, and to investigate the met-abolic impact of trauma and illness (White, 1993;Schwarcz and White, 2004; Knudson et al., 2007; Whiteet al., 2009; Williams et al., 2011; Knudson et al., 2012;Williams and Katzenberg, 2012; Webb et al., 2013).Because hair can be noninvasively sampled, there is alsoa sizable body of clinical isotopic research focusing onstarvation, dietary habits and nutrition, geographic relo-cation, pregnancy and specific diseases (O’Connell andHedges, 1999; Bol and Pflieger, 2002; Fuller et al., 2004,2005; McCullagh et al., 2005; Petzke et al., 2005, 2006;Mekota et al., 2006; Huelsemann et al., 2009; Mekotaet al., 2009; Valenzuela et al., 2011). Cortisol, a hormoneproduced in response to stress, can also be measured inhair. Clinical studies have measured systemic cortisol lev-els during pregnancy, and explored the impact of chronicstress, perceived stress, and specific diseases on cortisolproduction (Kalra et al., 2007; Van Uum et al., 2008;Kirschbaum et al., 2009; Thomson et al., 2009; Gowet al., 2010; Dettenborn et al., 2012; Russell et al., 2012).Cortisol has also been successfully extracted from archeo-logical hair samples and has been used to document indi-vidual experiences of stress in the months precedingdeath (Webb et al., 2010).
For this study, hair samples from 14 adults excavatedat Cahuachi and near Huaca del Loro in the SouthernNasca Region of Peru (AD1-1000) were analyzed todetermine carbon- and nitrogen-isotope compositions(Webb et al., 2013) and cortisol levels. Good biomolecularpreservation has been established through a pilot studyof Peruvian mummies from several sites, including theNasca Region (Webb et al., 2010). The Nasca Regionremains in particular were chosen for additional studybecause of the generally low frequency of stress markerson bones and teeth for previously analysed skeletalmaterial from this context (Kellner, 2002; Cagigao,2009), suggesting that understanding ante-mortem expe-riences may be critical to elucidating stress and death in
Grant sponsor: Social Sciences and Humanities Research Councilof Canada, the Natural Sciences and Engineering Research Councilof Canada, the Canada Research Chairs Program, the Physicians’Services Incorporated Foundation, Canada Foundation for Innova-tion and Ontario Research Fund.
*Correspondence to: Emily Webb, Organic Geochemistry Unit,School of Chemistry, Cantock’s Close, University of Bristol, BristolBS8 1TS, United Kingdom. E-mail: [email protected]
Received 4 August 2014; revised 16 October 2014; accepted 12November 2014
DOI: 10.1002/ajpa.22673Published online 00 Month 2014 in Wiley Online Library
(wileyonlinelibrary.com).
� 2014 WILEY PERIODICALS, INC.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 00:00–00 (2014)

the Nasca Region. Both isotopic and cortisol data areincorporated sequentially into hair as it grows, andreflect approximately the same period of time for eachindividual. Combining these datasets has the potentialto provide unique insight into the often hidden issues offrailty, risk and ante-mortem stress, which are central toour understanding of health, life, and death in ancientsocieties. Here, we use a case study approach to demon-strate the interpretive potential of integrating isotopicand cortisol data to explore diet, stress, and health, andto describe individual experiences in the months or yearspreceding death.
BIOCHEMICAL ANALYSIS OF ARCHEOLOGICALHAIR
Hair growth considerations
Hair grows at an average rate of �0.35 mm/day, or1 cm/month, and is metabolically inactive, retaining asequential archive of biochemical information (O’Connelland Hedges, 1999). Hair growth rate cannot be deter-mined after death, and has been observed to range from0.6 to 2 cm per month among modern humans (LeBeauet al., 2011). There is a 14-day lag between hair forma-tion and emergence of the mature hair from the scalp,and this creates a blind spot covering the two weeksimmediately preceding death (Nakamura et al., 1982;O’Connell and Hedges, 1999). Hairs that have completedgrowth are periodically shed, and subsequently replacedby new hairs. Hair grows during the anagen phase (last-ing �two to three years) and it is during this phase thatnew information is incorporated. The anagen phase isfollowed by the catagen, or quiescent, phase, which lastsfrom one to four weeks, and the telogen phase, whichlasts from one to three months. The hair is finally shedwhen a new anagen phase begins in the follicle, and theold hair is pushed out by the new hair. Typically, 80–85% of scalp hair is in anagen phase, 1–2% in catagenphase, and 10–20% in telogen phase, but these propor-tions may change seasonally or with illness (Hardingand Rogers, 1999; Schwertl et al., 2003). In a mixed-phase hair sample, temporal trends may be obscuredand intersegment variation in isotopic composition willbe attenuated (Williams et al., 2011); thus, using mixed-phase samples risks under-representing the true extentof variability in hair biomolecular data. The proportionof actively growing hairs in a random sample of hair canalso vary among individuals (Williams et al., 2011). Dif-ferences in cortisol levels and isotopic compositionsacross bundles of hair from different parts of the scalpwould hypothetically occur if the proportions of anagen:catagen: telogen hair differed substantially within eachbundle. Finally, Sauv�e et al. (2007) determined thatthere were no statistically significant differences amongcortisol levels from five different parts of the scalp. Thecortisol measurements ranged from 50 to 86.5 pg/mg,which is small relative to the scale of variation weobserved for cortisol levels in archeological hair. There isno evidence to suggest intrinsic biological variation inisotopic composition of hair from different parts of thescalp.
Biomolecules, including cortisol and protein, are incor-porated into the hair fiber as it grows. The cortisol levelmeasured in a 1 cm segment of hair reflects the amountof free cortisol circulating in the body during the periodof growth (i.e., roughly one month). Similarly, keratind13C and d15N values reflect the isotopic composition of
food consumed during the period of growth. Becausehair in anagen phase grows continuously, a change indiet can be detected in hair keratin isotopic compositionswithin a few days. Variation is dampened to someextent, however, by the buffering effect of incorporatingamino acids into keratin from the body protein pool,which will still contain carbon and nitrogen from the‘old’ diet. On the basis of controlled research withhuman scalp hair, complete isotopic equilibration of thebody protein pool with a new diet may take up to 12months for carbon, and approximately five months fornitrogen (O’Connell and Hedges, 1999). Because hairgrows quickly, shifts in dietary isotopic composition arereflected in changing keratin d13C and d15N values incre-mentally along the hair shaft. A correlative relationshipbetween carbon- and nitrogen-isotope compositionsunder conditions of rapidly changing diet has beenobserved (Webb et al., 2013). Further, the relationshipbetween keratin d13C and d15N values becomes weakduring equilibration, since the nitrogen- and carbon-isotope compositions of the body pool are not adjustingto the new constant diet at the same rate (Webb et al.,2013). It is possible that there is an offset between corti-sol and isotopic data as it is archived in hair, whereinmeasured cortisol levels in hair change more rapidlythan isotopic compositions, which may lag behind by afew days. We contend that the temporal offset betweencortisol and dietary information is likely mitigated tosome extent by the sampling interval of 1 cm used here,but, in the absence of clinical research, we caution thatthe temporal match may not be exact.
Carbon- and nitrogen-isotope analysis
The hair fiber is composed of dead keratinized cells.Keratin d13C and d15N values reflect the isotopic compo-sition of dietary protein when diet is protein sufficient(Ambrose, 1993; Petzke et al., 2005). Keratin protein iscomposed of essential amino acids, which must be routedfrom dietary protein, and nonessential amino acids,which may be routed from dietary protein or synthesizedfrom stored carbohydrates, lipids, or proteins (Schoeller,1999; McCullagh et al., 2005). The d13C values reflectrelative proportions of C3 and C4 foods directly con-sumed or in the diet of consumed animals. If, however,carbohydrate intake is inadequate, lipolysis (breakdownof stored fats) may be accelerated. Although the relativeproportions of C3 and C4 foods consumed would notchange, the d13C values could decrease in response tothe change in carbon source (i.e., from dietary carbohy-drates to body lipids) because body lipids are 13C-depleted relative to diet (Lee-Thorp et al., 1989; Mekotaet al., 2006, 2009). Keratin d15N values are used toassess trophic level of consumed protein, and consumertissues are always 15N-enriched by 3–4& per trophiclevel relative to diet (Ambrose, 1993, 2000). Recent bio-archeological and clinical research has demonstratedthat d15N values can be significantly influenced by theenvironment, by metabolic and physiological stress (e.g.,pregnancy, nutritional inadequacy), and by pathologicalconditions (e.g., disease, illness, and trauma) (Heatonet al., 1986; Hobson et al., 1993; Katzenberg and Lovell,1999; Fuller et al., 2004, 2005; Petzke et al., 2006;Mekota et al., 2006, 2009; Williams et al., 2011; Olsenet al., 2013).
In healthy individuals, the rate of protein synthesisequals the rate of protein breakdown and loss, and this
2 E.C. WEBB ET AL.
American Journal of Physical Anthropology

homeostatic state is reflected in body nitrogen balance.This balance responds to changes in amino acid avail-ability (Waterlow, 1999). The metabolic amino acid poolis composed of amino acids derived from dietary proteinand catabolized body protein. The body is in positivenitrogen balance when synthesis exceeds breakdown(e.g., during tissue repair after trauma, pregnancy), andnewly synthesized proteins will be 15N-depleted relativeto diet.
In negative nitrogen balance, catabolized body proteinamino acids make up a greater proportion of the aminoacids available for synthesis, leading to progressive 15Nenrichment of the metabolic amino acid pool. Negativenitrogen balance is induced by stress (e.g., infection,fever, anxiety, inadequate diet; Hobson et al., 1993; Scho-eller, 1999). The progressive enrichment of the metabolicamino acid pool during negative nitrogen balance occursbecause nitrogen from catabolized body proteins is 15N-enriched relative to diet. Isotopic fractionation (differen-tial partitioning of 15N vs. 14N) is thought to occur duringdeamination and transamination reactions as nitrogen istransported to the liver for elimination. Isotopically-lightnitrogen (14N) is favored for excretion in the form of urea,resulting in an overall enrichment of the metabolic aminoacid pool (Hobson et al., 1993; Schoeller, 1999). This isthe basis for both the trophic level effect previouslydescribed, and the ability to detect metabolic changesassociated with stress via nitrogen-isotope analysis.
Cortisol
The glucocorticoid hormone cortisol is produced inresponse to real and perceived stress, and it plays a keyrole in mediating the body’s physiological stress response(O’Connor et al., 2000; Davenport et al., 2006; Miller et al.2007; Van Uum et al., 2008; Novak et al., 2013). Clinicalresearch has demonstrated that hair cortisol levels reflectstress-induced changes in cortisol production by thehypothalamic-pituitary-adrenal axis (Davenport et al.2006; Kalra et al., 2007; Van Uum et al., 2008; Kirsch-baum et al., 2009). In clinical contexts, cortisol in bloodplasma, saliva, or urine is typically measured at specificpoints in time over short experimental time periods,whereas hair, as a continuously growing tissue, is well-suited to longer term retrospective studies of stress. Theprimary metabolic effect of cortisol is to promote gluconeo-genesis, which mobilises stored energy from adipose andbody proteins when dietary sources and glucose reservesare depleted. Cortisol also accelerates both proteolysis andlipolysis, and modulates the immune system by directingthe duration and magnitude of the inflammatory responseand lymphocyte maturation (Sapolsky et al., 2000). Corti-sol can therefore be seen as an important mechanismthrough which experienced physiological and psychosocialstresses influence the physical body (Miller et al., 2007).
Cortisol levels in hair reflects systemic cortisol levels inthe body, and have been widely used in clinical researchto retrospectively examine exposure to stress and hor-mone production (e.g., Kirschbaum et al., 2009; Detten-born et al., 2012). The preservation of biogenic patternsof cortisol production in archeological hair has also beendemonstrated (Webb et al., 2010). Free cortisol is incorpo-rated into hair follicles via the bloodstream, as are othersteroid hormones, drugs, and biomolecules. Althoughthere is no evidence to suggest that cortisol, once incorpo-rated, migrates through the hair shaft (Novak et al.,2013), there are several factors that can impact the detec-
tion of biogenic cortisol levels, such as leaching, contami-nation or modification of biogenic cortisol. Externalcontamination from the burial environment is unlikely,since cortisol is only produced by mammals. A possiblesource of exogenous cortisol may be animal fats used inthe manufacture of soaps or shampoos but, if this wereoccurring, a systematic distribution of contaminant corti-sol along the hair shaft or obliteration of intersegmentaldifferences would be expected. The most likely form ofdiagenetic alteration in an archeological context is leach-ing. Cortisol is most effectively removed from hair byrepeated or prolonged exposure to a solvent (e.g., wateror alcohol, Davenport et al., 2006; Novak et al., 2013;Thomson, personal communication), particularly at ele-vated temperatures. In modern studies, for example, theamount of cortisol retained in hair further than 3 cmfrom the scalp decreases sharply, which was attributed toloss of cortisol through washing with alcohol-containingshampoos (Kirschbaum et al., 2009). In archeological hairsamples, changing levels of cortisol along individual hairbundles therefore most likely reflect biogenic patterns ofcortisol production, even if the absolute amount of cortisolmay be somewhat lower due to post-mortem loss.
INTEGRATING ISOTOPIC AND CORTISOL DATA
Dietary shifting and psychosocial stress
For an essentially healthy individual (i.e., not suffer-ing significant physiological and/or metabolic perturba-tion) consuming a nutritionally adequate diet, carbon-,and nitrogen-isotope compositions may vary minimallyor seasonally. Among ancient Peruvians, this variation istypically less than 2.7& in d13C over the length of thehair sample, or is in a fashion consistent with acquiringfoods from multiple production zones, either by move-ment of food or local mobility (Williams et al., 2012;Webb et al., 2013). Diurnal variations in cortisol produc-tion, including age- and sex-related differences (VanCauter et al., 1996; Kudielka et al., 2004), are subsumedwithin the sampling resolution for this study. Clinicalresearch suggests that there may be seasonal variationin cortisol production (Hansen et al., 2001; Matchocket al., 2007), but the extent to which such patterningmay be detectable in hair is unknown.
Psychosocial and physical stress could cause cortisolproduction to increase without an associated change inisotopic composition from one month to the next. Socialstress, particularly threats to physical integrity, as wellas low controllability of stressors and individual psycho-logical responses to stress can all influence cortisol pro-duction (Miller et al., 2007). Exploiting multipleproduction zones through local mobility involves signifi-cant movement across the landscape. In the Andes, thismovement would have involved the extra effort requiredfor travel up and down altitudinal gradients. Psychoso-cial and physical stressors related to travel are alsolikely to contribute to experienced stress as reflected incortisol levels. Exposure to new pathogens, adaptation toa different social or physical environment, or leaving astressful environment to settle in a less stressful locale(or vice versa) are all potential stressors that may bedirectly associated with residential mobility.
Pregnancy
Clinical and bioarcheological research have demon-strated that pregnancy can be inferred with considerable
INTEGRATING CORTISOL AND ISOTOPIC DATA FROM HAIR 3
American Journal of Physical Anthropology

certainty using the isotopic composition of hair keratin.During pregnancy, dietary intake, particularly proteinintake, should increase in order to meet the nutritionalrequirements associated with maintaining maternalhealth and foetal growth and development (Mojtahediet al., 2002). Under nutritionally adequate conditions,nitrogen balance is positive, and increasing nutritionaldemands are met by normal routing of dietary aminoacids for tissue synthesis. The d13C values of tissues syn-thesized during this time are expected to reflect diet,and the d15N values are expected to decrease (by 1–1.5&; Fuller et al., 2005). If there are periods of nutri-tional stress during pregnancy (e.g., morning sickness)or if diet is at times nutritionally inadequate, this willinduce negative nitrogen balance, and body stores willbe mobilized to support tissue synthesis. Under theseconditions, the d15N values are expected to increase (by0.5–1&; Fuller et al., 2005; Williams et al., 2011). Dur-ing pregnancy, systemic cortisol levels increase toapproximately two to three times above individual base-line values in the third trimester, and return to thebaseline within days after birth (Kirschbaum et al.,2009).
Malnutrition and starvation
Isotopic studies of malnutrition and starvation haveused anorexia nervosa as a clinical analogy for under-standing changes in metabolism associated with calorie-restricted and/or nutritionally inadequate diets. Whenthe diet is inadequate, fat stores are progressivelydepleted and body proteins are mobilized. As malnutri-tion progresses to starvation, either because of decreasedfood intake or decreased intestinal absorption, fat storesare exhausted and body protein catabolism continues(Mekota et al., 2006, 2009). Since catabolized body pro-teins will make up an increasingly greater proportion ofnitrogen-containing compounds in the metabolic aminoacid pool, additional 15N-enrichment relative to diet (by1–2&) will be observed in synthesized tissues (Hobsonet al., 1993; Mekota et al., 2006, 2009). Mobilization offat stores may also cause a decrease in d13C values (Lee-Thorp et al., 1989; Mekota et al., 2006, 2009). In caseswhere increased nutritional intake leads to recoveryfrom malnutrition and the return of a healthy metabo-lism, it is expected that d15N values will initiallydecrease (transition from 15N-enriched stored protein todietary protein), and then equilibrate to the nitrogen-isotope composition of diet (Mekota et al., 2006, 2009).
Clinically, weight loss via calorie restriction leads tominimal increase in plasma cortisol levels (i.e., consum-ing a low calorie, but nutritionally adequate, diet).Severe malnutrition and starvation may; however,induce marked increase in plasma cortisol, likely causedby both increased secretion of cortisol and hypometabo-lism (Fichter et al., 1986; Støving et al., 1999; Johnstoneet al., 2004; Manary et al., 2006). Malnutrition and star-vation are often accompanied by infection or illness,which cause increased cortisol production that may notbe directly associated with restricted diet.
Trauma and illness
During tissue repair after fracture or trauma, nutri-tional state influences nitrogen balance. The body willbe in positive nitrogen balance if increased nutritionaldemands are met by diet or in negative nitrogen balanceif diet is inadequate or pain, infection, or fever co-occurs.
The d13C values of hair keratin synthesized during theperiod of interest do not change uniformly in response tothe injury and will reflect diet (Williams et al., 2011).Changes in d15N values of 61–1.5& associated withtrauma, pain and reinjury have also been observed (seealso Katzenberg and Lovell, 1999; Williams et al., 2011;Olsen et al., 2013). Systemic infection, fever, diarrhea,and/or viral illness all induce negative nitrogen balance(i.e., a catabolic response), and could thus potentiallycause an increase in d15N values (Beisel, 1972; Scrim-shaw and San Giovanni, 1997). This catabolic responsebegins at the onset of infection or illness, and persistsinto convalescence (Beisel, 1972). If malnutritionbecomes a complication, the d13C values may alsodecrease (section “Malnutrition and Starvation”). Com-bined physiological and psychological stress associatedwith trauma and/or infection typically lead to higheraverage cortisol levels (Sapse, 1997; Beishuizen et al.,2001; Ball, 2008). In clinical contexts, patients under-going treatment for multiple traumas or systemic infec-tion had high cortisol levels that gradually decreasedduring recovery (Beishuizen et al., 2001). Although long-term high levels of cortisol in the body suppress immunefunction, cortisol does play an important role in immuneresponse to injury and illness over the short-term.
Limitations
Although both clinical and bioarcheological researchhave examined cortisol production and variability in ker-atin isotopic composition under various physiological,metabolic, and psychosocial conditions, there has notbeen a clinical study where both isotopic and cortisoldata were collected from the hair of the same individu-als. Moreover, modern clinical data cannot be applieduncritically to individuals from archeological contexts.Clinical data addressing, for example, metabolic changesassociated with disease processes, will involve individu-als undergoing treatment and consuming a balanceddiet who, for the most part, were likely to have beenfairly healthy before becoming ill. By comparison,although ancient people in the Nasca Region most likelyresponded to and cared for illness or injury, treatmentwas ultimately unsuccessful for the individuals exam-ined in this study. Finally, the overall state of healthprior to illness or trauma (i.e., stress history extendingfurther back than the length of the hair sample), is oftennot accessible for individuals from archeological con-texts, although recent stress experiences and evidence ofchildhood and long-term stress can sometimes beobserved. The proposed expectations for cortisol and iso-topic data and their relationship to various stressors aresummarized in Table 1.
ARCHEOLOGICAL CONTEXT
Nasca society
The Nasca polity inhabited the Rio Grande de NascaDrainage, Peru, during the Early Intermediate Period(AD1–750; Fig. 1). During the Early Nasca period (AD1–450), settlements emerged in the upper valleys of thedrainage, and construction and use of the ceremonialcenter, Cahuachi, reached its apogee (Silverman, 1993;Vaughn, 2009). Cahuachi is thought to have functionedprimarily as a pilgrimage centre with a small elite popu-lation described as a “centralized theocratic authority”(Orefici, 2006, p. 184). Feasting and rituals focused on
4 E.C. WEBB ET AL.
American Journal of Physical Anthropology

cyclical agricultural events are suggested to have drawnparticipants from the surrounding valleys (Silverman,1993). The transitional Middle Nasca period (AD450–550) is characterized by a dramatic reworking of iconog-raphy and the cessation of major construction at Cahua-chi (Silverman, 1993; Schreiber and Lancho Rojas,2003). During this period, the typically unstable Andeanclimate became even more unpredictable as the result ofsevere droughts (Thompson et al., 1985). From c.AD250until the end of the Middle Horizon (c.AD1000), climatein the Nasca Region became increasingly arid (Eitelet al., 2005; Eitel and Machtle, 2009; Bird et al., 2011).Because of the low river volume and inconsistent avail-ability of surface water, the Nasca people developed asystem of wells and irrigation canals (puquios) to accessmore reliable subterranean water and thereby increaseagricultural production in the dry middle valleys(Schreiber and Lancho Rojas, 2003). Kantner andVaughn (2012) hypothesize that the theocratic elite atCahuachi may have lost social and ideological powerbecause of their inability to mitigate the effects of thedrought.
The populace coalesced into a few large settlementsduring the Late Nasca period (AD550–750; Schreiberand Lancho Rojas, 2003; Schreiber, 2005). This popula-tion aggregation was concomitant with increased socialcomplexity and greater representation of conflict-relatedthemes on pottery. Towards the end of the Early Inter-mediate Period, evidence of damaging El Ni~no effectshas been observed, including flooding at Cahuachi (Ore-fici and Drusini, 2003; Beresford-Jones et al., 2009). TheWari polity expanded into Nasca c. AD750, marking thebeginning of the Middle Horizon (locally the Loro Period,AD750–1000). Wari settlements were establishedthroughout the Southern Nasca Region, and Wariceramics are found in Nasca cemeteries and at habita-tion sites along with the local Loro style (Strong, 1957;Schreiber, 2001, 2005). The local population of the north-ern river valleys decreased as Nasca people moved southto the Las Trancas river valley where a large local cen-ter, Huaca del Loro, was established (Schreiber, 2001;Conlee and Schreiber, 2006). A recent archeological sur-vey (Edwards, 2010) further indicates that the Warimade important changes to the landscape of the upperand mid-valleys of the SNR, establishing several settle-ments and constructing roads that linked the head-
waters of the Nasca river tributaries with the coastalplains.
Bioarcheological perspective on Nasca stressand health status
Elevated rates of accidental trauma and trauma asso-ciated with violence have been observed in skeletal sam-ples from the Las Trancas and Palpa river valleys(Kellner, 2002; Cagigao, 2009). The incidence of traumaindicative of interpersonal conflict (e.g., cranial andcranio-facial trauma, parry fractures) is relatively highduring all periods compared to samples from otherregions globally (Kellner, 2002). Interpersonal violence isoften motivated by changing social and physical environ-mental conditions, such as uncertain resource availabil-ity or conflict with expanding polities (Tung, 2007). Tung(2007) observed that Wari imperial rule was associatedwith considerable presence of trauma consistent withinterpersonal violence in peripheral Wari settlements
Fig. 1. Map of Rio Grande de Nasca Drainage indicatinglocation of sampled burials.
TABLE 1. Summary of expectations for cortisol and isotopic data
Isotopic data Cortisol levels
Seasonal diet �62.7& change in d13CSome change in d15N values
Low stress: minimal variabilityPsychosocial stress: increased cortisol
Travel �62.7& change in d13C.Possible increase in d15N values with infection
Physical stress: increased cortisol.Exposure to new pathogens: increased cortisol
Malnutrition andstarvation
�1–2& increase in d15N values.Probable decrease in d13C values.Recovery indicated by decrease in d15Nvalues followed by equilibration with diet
If severe, cortisol production will increase
Trauma and illness Good nutrition: some decrease in d15N values.Poor nutrition: some increase in d15N values(�61–1.5&).Recovery indicated by equilibration of d15Nvalues with diet.Minimal change in d13C values.
High cortisol levels that decrease duringrecovery
INTEGRATING CORTISOL AND ISOTOPIC DATA FROM HAIR 5
American Journal of Physical Anthropology
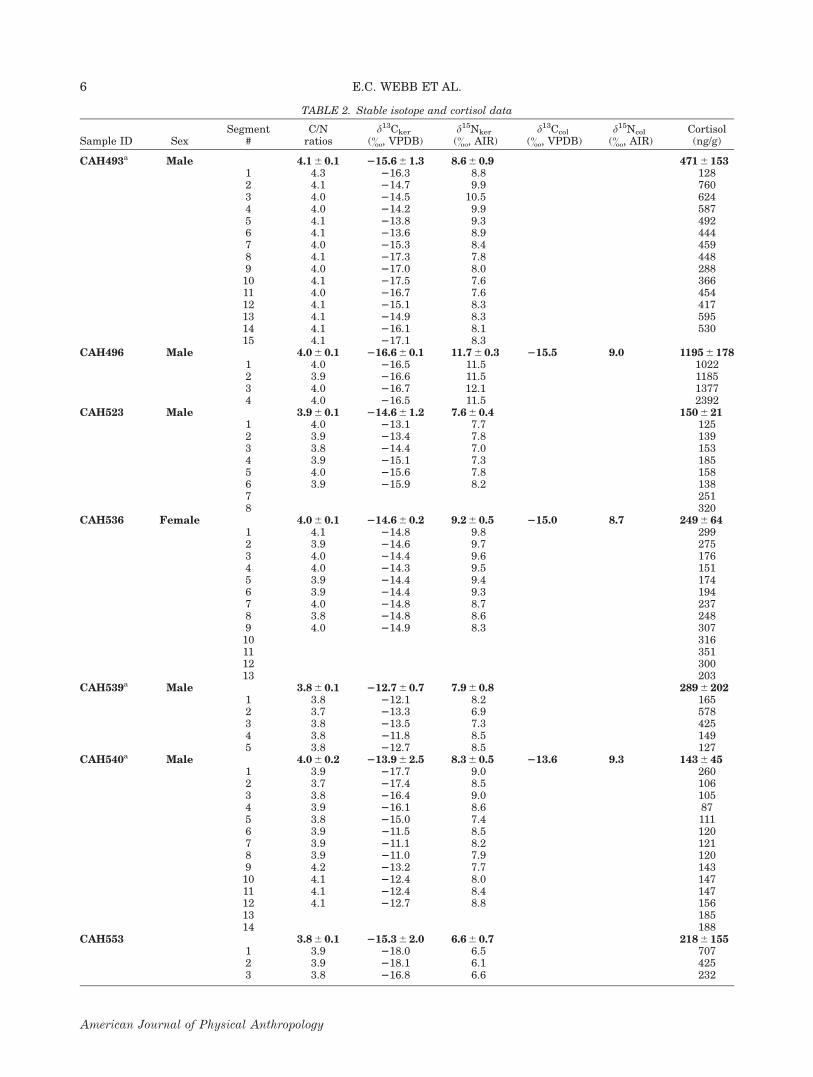
TABLE 2. Stable isotope and cortisol data
Sample ID SexSegment
#C/N
ratiosd13Cker
(&, VPDB)d15Nker
(&, AIR)d13Ccol
(&, VPDB)d15Ncol
(&, AIR)Cortisol(ng/g)
CAH493a Male 4.1 6 0.1 215.6 6 1.3 8.6 6 0.9 471 6 1531 4.3 216.3 8.8 1282 4.1 214.7 9.9 7603 4.0 214.5 10.5 6244 4.0 214.2 9.9 5875 4.1 213.8 9.3 4926 4.1 213.6 8.9 4447 4.0 215.3 8.4 4598 4.1 217.3 7.8 4489 4.0 217.0 8.0 28810 4.1 217.5 7.6 36611 4.0 216.7 7.6 45412 4.1 215.1 8.3 41713 4.1 214.9 8.3 59514 4.1 216.1 8.1 53015 4.1 217.1 8.3
CAH496 Male 4.0 6 0.1 216.6 6 0.1 11.7 6 0.3 215.5 9.0 1195 6 1781 4.0 216.5 11.5 10222 3.9 216.6 11.5 11853 4.0 216.7 12.1 13774 4.0 216.5 11.5 2392
CAH523 Male 3.9 6 0.1 214.6 6 1.2 7.6 6 0.4 150 6 211 4.0 213.1 7.7 1252 3.9 213.4 7.8 1393 3.8 214.4 7.0 1534 3.9 215.1 7.3 1855 4.0 215.6 7.8 1586 3.9 215.9 8.2 1387 2518 320
CAH536 Female 4.0 6 0.1 214.6 6 0.2 9.2 6 0.5 215.0 8.7 249 6 641 4.1 214.8 9.8 2992 3.9 214.6 9.7 2753 4.0 214.4 9.6 1764 4.0 214.3 9.5 1515 3.9 214.4 9.4 1746 3.9 214.4 9.3 1947 4.0 214.8 8.7 2378 3.8 214.8 8.6 2489 4.0 214.9 8.3 30710 31611 35112 30013 203
CAH539a Male 3.8 6 0.1 212.7 6 0.7 7.9 6 0.8 289 6 2021 3.8 212.1 8.2 1652 3.7 213.3 6.9 5783 3.8 213.5 7.3 4254 3.8 211.8 8.5 1495 3.8 212.7 8.5 127
CAH540a Male 4.0 6 0.2 213.9 6 2.5 8.3 6 0.5 213.6 9.3 143 6 451 3.9 217.7 9.0 2602 3.7 217.4 8.5 1063 3.8 216.4 9.0 1054 3.9 216.1 8.6 875 3.8 215.0 7.4 1116 3.9 211.5 8.5 1207 3.9 211.1 8.2 1218 3.9 211.0 7.9 1209 4.2 213.2 7.7 14310 4.1 212.4 8.0 14711 4.1 212.4 8.4 14712 4.1 212.7 8.8 15613 18514 188
CAH553 3.8 6 0.1 215.3 6 2.0 6.6 6 0.7 218 6 1551 3.9 218.0 6.5 7072 3.9 218.1 6.1 4253 3.8 216.8 6.6 232
6 E.C. WEBB ET AL.
American Journal of Physical Anthropology

TABLE 2. Continued
Sample ID SexSegment
#C/N
ratiosd13Cker
(&, VPDB)d15Nker
(&, AIR)d13Ccol
(&, VPDB)d15Ncol
(&, AIR)Cortisol(ng/g)
4 3.8 213.3 8.0 2115 3.8 211.3 8.2 1636 3.7 212.1 7.0 1127 3.8 214.1 5.9 1548 3.8 215.8 5.7 1539 3.8 216.0 6.3 20610 3.7 215.7 6.5 14711 3.7 215.9 6.4 17112 3.7 215.9 6.4 12313 3.8 215.5 6.5 18614 3.8 215.0 6.3 133b
15 3.9 216.8 5.9 133b
CAH558a 4.0 6 0.0 216.2 6 2.9 6.9 6 1.2 955 6 1371 3.9 218.1 6.2 10382 3.9 218.3 5.9 8503 4.0 217.7 6.3 11184 4.0 215.6 7.6 9885 3.9 211.4 8.7 782
LTR590 Female 3.8 6 0.1 213.8 6 0.9 9.4 6 1.4 805 6 3161 3.7 214.7 8.4 9652 3.7 215.3 8.2 12613 3.7 214.6 8.5 12284 3.7 212.6 10.6 10735 3.7 212.0 11.1 7426 3.7 213.2 9.2 5527 3.7 214.8 7.9 4628 3.8 214.7 8.3 5329 3.8 214.6 8.5 51510 3.7 214.0 9.2 52411 3.7 213.7 9.8 56912 3.8 213.3 10.2 74313 3.8 213.4 10.0 137514 3.8 213.7 10.4 72815 3.8 212.9 11.516 3.8 213.0 11.717 3.8 212.8 11.218 3.8 212.1 11.819 3.9 213.4 9.220 3.8 214.4 7.821 3.8 214.6 7.822 3.8 214.4 8.223 3.9 214.5 8.124 3.9 214.5 8.0
LTR593a Male 3.7 6 0.1 213.9 6 1.4 7.6 6 0.3 636 6 1271 3.9 215.9 7.3 8112 3.7 215.6 8.0 6853 3.6 215.7 8.4 4934 3.6 215.6 7.9 5755 3.6 214.8 7.4 5366 3.8 214.7 7.0 5927 3.6 215.0 7.1 5758 3.6 214.9 7.1 5259 3.7 214.0 7.5 62410 3.7 213.5 7.5 53211 3.7 212.9 7.8 613b
12 3.7 212.6 7.6 613b
13 3.7 212.6 7.7 613b
14 3.7 212.2 7.7 897b
15 3.8 212.4 7.6 897b
16 3.8 212.5 7.8 897b
17 3.7 212.6 7.7 807b
18 3.7 212.6 7.6 807b
LTR616 Male 3.7 6 0.0 213.5 6 0.5 9.2 6 0.2 596 6 3221 3.7 213.8 9.2 16532 3.7 214.5 9.2 7753 3.7 214.0 9.1 6264 3.6 213.5 9.3 6705 3.6 212.9 9.1 4836 3.7 212.6 9.2 457
INTEGRATING CORTISOL AND ISOTOPIC DATA FROM HAIR 7
American Journal of Physical Anthropology
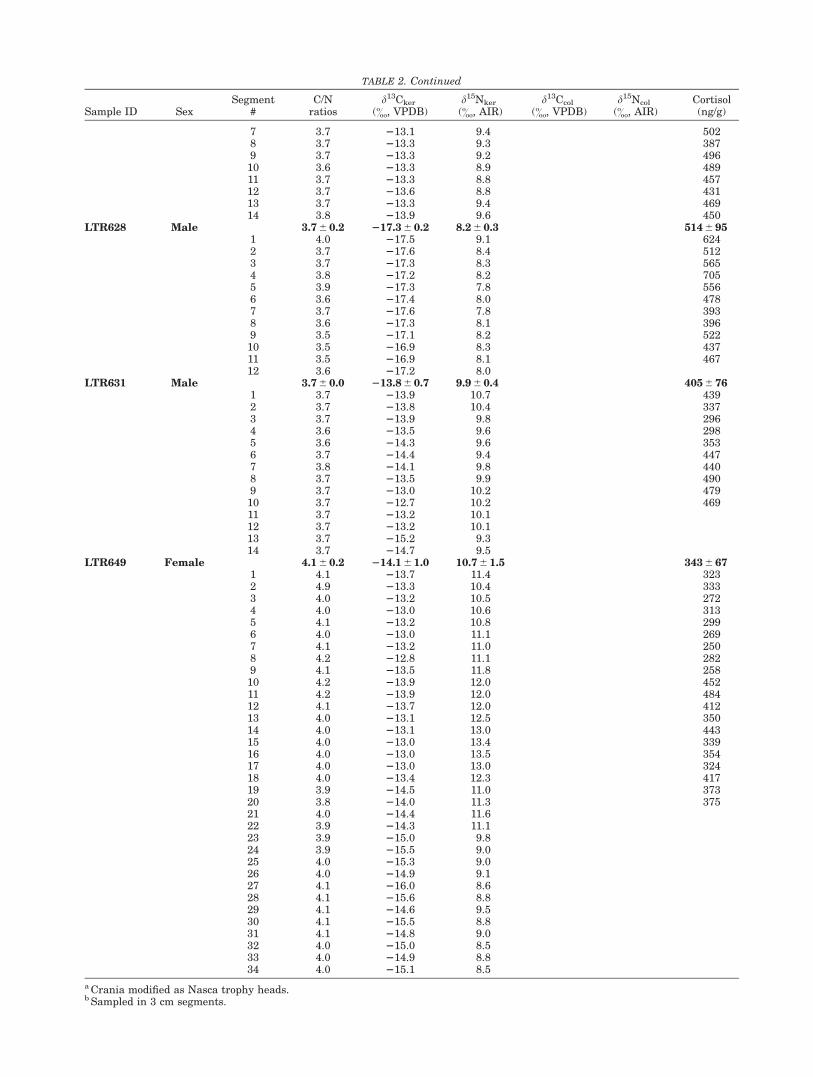
TABLE 2. Continued
Sample ID SexSegment
#C/N
ratiosd13Cker
(&, VPDB)d15Nker
(&, AIR)d13Ccol
(&, VPDB)d15Ncol
(&, AIR)Cortisol(ng/g)
7 3.7 213.1 9.4 5028 3.7 213.3 9.3 3879 3.7 213.3 9.2 49610 3.6 213.3 8.9 48911 3.7 213.3 8.8 45712 3.7 213.6 8.8 43113 3.7 213.3 9.4 46914 3.8 213.9 9.6 450
LTR628 Male 3.7 6 0.2 217.3 6 0.2 8.2 6 0.3 514 6 951 4.0 217.5 9.1 6242 3.7 217.6 8.4 5123 3.7 217.3 8.3 5654 3.8 217.2 8.2 7055 3.9 217.3 7.8 5566 3.6 217.4 8.0 4787 3.7 217.6 7.8 3938 3.6 217.3 8.1 3969 3.5 217.1 8.2 52210 3.5 216.9 8.3 43711 3.5 216.9 8.1 46712 3.6 217.2 8.0
LTR631 Male 3.7 6 0.0 213.8 6 0.7 9.9 6 0.4 405 6 761 3.7 213.9 10.7 4392 3.7 213.8 10.4 3373 3.7 213.9 9.8 2964 3.6 213.5 9.6 2985 3.6 214.3 9.6 3536 3.7 214.4 9.4 4477 3.8 214.1 9.8 4408 3.7 213.5 9.9 4909 3.7 213.0 10.2 47910 3.7 212.7 10.2 46911 3.7 213.2 10.112 3.7 213.2 10.113 3.7 215.2 9.314 3.7 214.7 9.5
LTR649 Female 4.1 6 0.2 214.1 6 1.0 10.7 6 1.5 343 6 671 4.1 213.7 11.4 3232 4.9 213.3 10.4 3333 4.0 213.2 10.5 2724 4.0 213.0 10.6 3135 4.1 213.2 10.8 2996 4.0 213.0 11.1 2697 4.1 213.2 11.0 2508 4.2 212.8 11.1 2829 4.1 213.5 11.8 25810 4.2 213.9 12.0 45211 4.2 213.9 12.0 48412 4.1 213.7 12.0 41213 4.0 213.1 12.5 35014 4.0 213.1 13.0 44315 4.0 213.0 13.4 33916 4.0 213.0 13.5 35417 4.0 213.0 13.0 32418 4.0 213.4 12.3 41719 3.9 214.5 11.0 37320 3.8 214.0 11.3 37521 4.0 214.4 11.622 3.9 214.3 11.123 3.9 215.0 9.824 3.9 215.5 9.025 4.0 215.3 9.026 4.0 214.9 9.127 4.1 216.0 8.628 4.1 215.6 8.829 4.1 214.6 9.530 4.1 215.5 8.831 4.1 214.8 9.032 4.0 215.0 8.533 4.0 214.9 8.834 4.0 215.1 8.5
a Crania modified as Nasca trophy heads.b Sampled in 3 cm segments.

outside the Nasca drainage, and Wari-affiliated individu-als in the Las Trancas skeletal sample had a similarlyhigh incidence of cranial trauma (Kellner, 2002). Finally,Cagigao (2009) noted that, among Early IntermediatePeriod Nasca individuals from sites in the Palpa rivervalley, the frequencies of accidental trauma (e.g., limbfractures) and trauma associated with violence were50% and 38%, respectively. Combined data from thesestudies indicate that both physical and psychosocialstress caused by interpersonal violence and accidentaltrauma were likely significant during the Early Interme-diate Period and Middle Horizon.
Bioarcheological analysis of a skeletal sample exca-vated from the Las Trancas river valley cemeteriesrevealed an overall decline in health status from EarlyNasca to the Loro period, but, compared to other archeo-logical skeletal samples, the Nasca were not underexcessive stress during any period (Kellner, 2002). Nascaindividuals experienced physiological stress during child-hood (indicated by the presence of linear enamel hypo-plasias), localized and systemic infections(osteoperiostitis) and anaemia (potentially caused byparasitic infection, infectious disease and/or chronic diar-rhea) (Kellner, 2002). Increased population density oftenleads to a decline in health because of increased patho-gen transmission and waste disposal problems. Stressmay be alleviated by improving socioeconomic condi-tions, but social development may aggravate and/or gen-erate stress as well (Goodman et al., 1988). Exposure tovarious stressors does not mean that there will be aharmful impact on an individual’s health. Impact is afunction of duration and severity as well as an individu-al’s ability to cope with a given stressor (Sapolsky, 2004).Typically, prolonged stress episodes, and many infectiousagents and soft tissue diseases/injuries, will not leaveclear skeletal evidence. Moreover, individuals withlesions may be “paradoxically” healthier that those with-out, since, for skeletal traces to become evident, the indi-vidual must survive the stress episode and/or live for asignificant amount of time with a disease (Wood et al.,1992).
METHODS
Sampling
Hair samples were collected at the State Collection forAnthropology and Palaeoanatomy, Munich, Germany inFebruary and September 2008. The remains are fromHeinrich Ubbelohde-Doering’s 1932 field season, duringwhich he excavated �50 graves in the Rio Grande deNasca drainage (Ubbelohde-Doering, 1958, 1966). Hairsampled from fourteen adults from Cahuachi (CAH) andfrom cemeteries south of Huaca del Loro in the Las Tran-cas river valley (LTR) was analyzed for cortisol levels andcarbon- and nitrogen-isotope compositions. This group ofsamples includes five crania that have been modified asNasca trophy heads (Tello, 1918; Proulx, 1989; Browneet al., 1993; Silverman, 1993; Verano, 1995; Proulx, 2001;Verano, 2003; Knudson et al., 2009). Hair samples werecollected with care taken to maintain orientation andalignment of individual hair strands within each samplebundle. Each bundle of hair was sectioned into 1 cm seg-ments, with each segment representing approximatelyone month of growth based on the average growth rate of0.35 mm/day for scalp hair. Hair lengths in this studyranged from three to 34 cm, resulting in individual tem-
poral records ranging from approximately three monthsto nearly three years.
Laboratory procedures
For carbon- and nitrogen-isotope analysis, each hairsample bundle was cleaned of loose particulate matter bywiping gently with ethanol. Hair bundles were then sec-tioned and each segment sample was placed in a glassvial. Lipids and decomposition fluid residues wereremoved by soaking hair segments in a 2:1 chloroform:methanol solution for 24 h. Each segment sample wasthen rinsed with fresh solution and, when necessary, thisprocess was repeated. Segment samples were air-dried for12 to 24 h, and then finely minced and weighed (510 6 10lg) into tin capsules for isotopic analysis.
Isotopic analysis of keratin was performed using aCostech Elemental Analyzer interfaced with a ThermoFinnigan DeltaPlus XL mass spectrometer. The d13C val-ues were calibrated to VPDB (Coplen, 1994, 1996) usingIAEA-CH-6 and IAEA NBS-22, and the d15N values werecalibrated to AIR (Mariotti, 1983; Hoefs, 2004) usingIAEA-N1 and IAEA-N2. Analytical precision was deter-mined through duplicate analyses of samples, and was60.2& for d13Cker, and 60.1& for d15Nker. Accuracy wasassessed using a laboratory keratin standard, which gavean average d13C value of –24.06 6 0.05&, and an averaged15N value of 16.26 6 0.09&, which compare well withits accepted values (–24.04& and 16.36&, respectively).Isotopic analyses were performed at The University ofWestern Ontario, London, Canada, in the Laboratory forStable Isotope Science.
The methodology used to measure cortisol abundance inhair is described in detail in Van Uum et al. (2008). At least10 mg of hair per segment is required for cortisol analysis.Hair segments were placed in glass vials and the hair wasvery finely minced with surgical scissors. One millilitre ofmethanol (>98%) was added to extract the steroid. Thevials were then sealed and incubated for 16 h, whileshaking at 100 rpm and heating at 50�C. The methanolwas then removed from the vial, transferred to test tubesand evaporated by heating to 40�C under a nitrogenstream. The residue was reconstituted in 250 ml ofphosphate-buffered saline (PBS) at pH 8.0. The PBS mix-ture was then analyzed using a commercially availablesalivary enzyme immunoassay kit (ELISA, ALPCO Diag-nostics). All measurements were done in duplicate, andthe average difference between duplicate analyses was643 ng/g. For this procedure, the typical intraday coeffi-cient of variation is 7.2% in a hair sample with 64 ng/g ofcortisol and 6.0% at 629 ng/g. The interday variation is10.6% and 7.6% for 49 and 548 ng/ml, respectively.Cross-reactivities of steroids in the salivary ELISA kitare as follows: cortisol 100%, corticosterone 31%, proges-terone <2%, deoxycortisol <2%, dexamethasone <2%,estriol <0.001%, estrone <0.001%, and testosterone<0.001%. The lower limit of detection for cortisol is 1.14ng/ml (information from manufacturer), and the absolutecortisol extraction recovery is 88% in a 100 ng/ml stand-ard, and 87% for a 2 ng/ml standard (Van Uum et al.,2008). Cortisol analyses were performed at The Univer-sity of Western Ontario, London, Canada.
RESULTS
The carbon- and nitrogen-isotope results for hair kera-tin and collagen (when available; Webb et al., 2013) are
INTEGRATING CORTISOL AND ISOTOPIC DATA FROM HAIR 9
American Journal of Physical Anthropology

presented in Table 2 and Figure 2. All averages arereported as mean 6 1r unless otherwise stated. Themean C/N ratio for hair keratin was 3.9 [3.7–4.1]. Therewas no significant correlation between carbon- andnitrogen-isotope compositions and C/N ratios (mean P-value 5 0.44, C/N vs. d13Cker; mean P-value 5 0.51, C/Nvs. d15Nker). The isotopic composition of keratin is there-fore unlikely to have been extensively altered postmor-tem (O’Connell and Hedges, 1999). The cortisol resultsare also presented in Table 2. Webb et al. (2010) estab-lished that, with good tissue preservation and intermentin a dry burial environment, biogenic patterns of cortisolproduction can be observed along an archeological hairsample. The sample means are calculated as a mean ofaverage cortisol levels for each individual. The adultmean cortisol level is 522 6 368 ng/g for the 14 individu-als included in this study (Fig. 3). The mean male corti-sol level is 526 6 402 ng/g, and the mean female cortisollevel is 466 6 298 ng/g. This difference is not statisticallysignificant (Mann–Whitney, P 5 0.926). There is also nostatistically significant difference between Cahuachi(500 6 480 ng/g) and Las Trancas adults (550 6 167 ng/g;MW, P 5 0.197). Individuals modified into trophy headsdid not differ from the rest of the sample in any way.
Because all individuals have distinct experiences ofhealth, well-being and stress, each individual includedin this paper has been considered as a separate casestudy (see Webb et al., 2013 for a more detailed discus-sion of the isotopic data). The case studies are organ-ised by most likely cause(s) of stress, inferred from theinformation provided in section “Integrating Isotopic
and Cortisol Data.” For some individuals, however,there is overlap among potential causes of stress. Forsix adults (CAH 523, CAH 553, CAH 558, LTR 590,LTR 593, and LTR 649), stress experiences wereclosely associated with dietary change consistent withresidential mobility. In three adults (CAH 496, LTR593, and LTR 628), isotopic and cortisol results sug-gest illness and/or trauma. For two adults (CAH 493and CAH 540), nutritional stress and illness/traumaco-occurred. Neither the isotopic nor cortisol resultsindicate that any of the three females in this samplewere pregnant during the periods of time representedby their hair samples. Nonspecific stress along thehair sample and in the month(s) preceding death wasobserved for four adults (CAH 536, CAH 539, LTR 616,and LTR 631).
DISCUSSION
Mobility
Here, residential mobility refers to the relocation of anindividual or group among places of residence, whereaslocal mobility refers to cyclical or periodic rapid move-ment. The topography of the Andean Region is such thatisotopically and ecologically distinct production zonesare comparatively close together (Murra, 1972; Tieszenet al., 1979; Tomczak, 2003). As a result, isotopic recon-struction of short-term dietary shifting can be used toinfer residential and local mobility (White et al., 2009;Webb et al., 2013). Local mobility has been inferred from
Fig. 2. Carbon- and nitrogen-isotope compositions for individuals (A) engaged in local mobility, (B) who relocated, (C) with sig-nificant nondietary change, and (D) with seasonally-shifting diets.
10 E.C. WEBB ET AL.
American Journal of Physical Anthropology
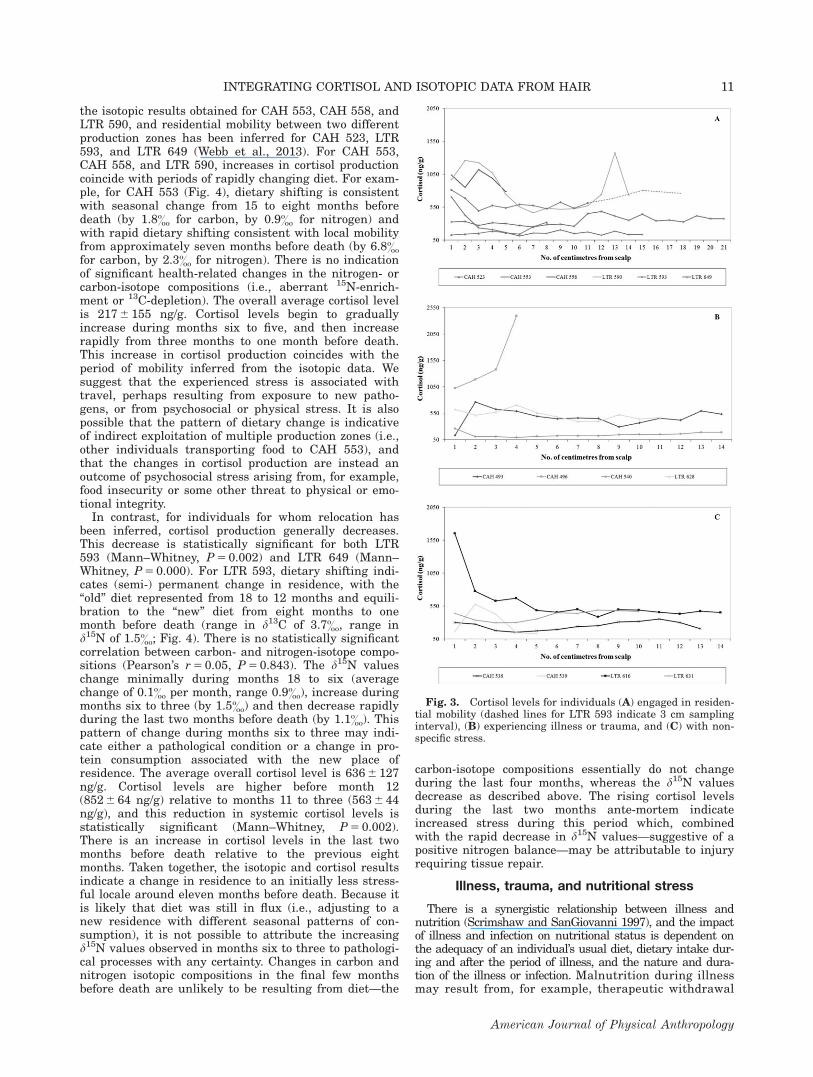
the isotopic results obtained for CAH 553, CAH 558, andLTR 590, and residential mobility between two differentproduction zones has been inferred for CAH 523, LTR593, and LTR 649 (Webb et al., 2013). For CAH 553,CAH 558, and LTR 590, increases in cortisol productioncoincide with periods of rapidly changing diet. For exam-ple, for CAH 553 (Fig. 4), dietary shifting is consistentwith seasonal change from 15 to eight months beforedeath (by 1.8& for carbon, by 0.9& for nitrogen) andwith rapid dietary shifting consistent with local mobilityfrom approximately seven months before death (by 6.8&
for carbon, by 2.3& for nitrogen). There is no indicationof significant health-related changes in the nitrogen- orcarbon-isotope compositions (i.e., aberrant 15N-enrich-ment or 13C-depletion). The overall average cortisol levelis 217 6 155 ng/g. Cortisol levels begin to graduallyincrease during months six to five, and then increaserapidly from three months to one month before death.This increase in cortisol production coincides with theperiod of mobility inferred from the isotopic data. Wesuggest that the experienced stress is associated withtravel, perhaps resulting from exposure to new patho-gens, or from psychosocial or physical stress. It is alsopossible that the pattern of dietary change is indicativeof indirect exploitation of multiple production zones (i.e.,other individuals transporting food to CAH 553), andthat the changes in cortisol production are instead anoutcome of psychosocial stress arising from, for example,food insecurity or some other threat to physical or emo-tional integrity.
In contrast, for individuals for whom relocation hasbeen inferred, cortisol production generally decreases.This decrease is statistically significant for both LTR593 (Mann–Whitney, P 5 0.002) and LTR 649 (Mann–Whitney, P 5 0.000). For LTR 593, dietary shifting indi-cates (semi-) permanent change in residence, with the“old” diet represented from 18 to 12 months and equili-bration to the “new” diet from eight months to onemonth before death (range in d13C of 3.7&, range ind15N of 1.5&; Fig. 4). There is no statistically significantcorrelation between carbon- and nitrogen-isotope compo-sitions (Pearson’s r 5 0.05, P 5 0.843). The d15N valueschange minimally during months 18 to six (averagechange of 0.1& per month, range 0.9&), increase duringmonths six to three (by 1.5&) and then decrease rapidlyduring the last two months before death (by 1.1&). Thispattern of change during months six to three may indi-cate either a pathological condition or a change in pro-tein consumption associated with the new place ofresidence. The average overall cortisol level is 636 6 127ng/g. Cortisol levels are higher before month 12(852 6 64 ng/g) relative to months 11 to three (563 6 44ng/g), and this reduction in systemic cortisol levels isstatistically significant (Mann–Whitney, P 5 0.002).There is an increase in cortisol levels in the last twomonths before death relative to the previous eightmonths. Taken together, the isotopic and cortisol resultsindicate a change in residence to an initially less stress-ful locale around eleven months before death. Because itis likely that diet was still in flux (i.e., adjusting to anew residence with different seasonal patterns of con-sumption), it is not possible to attribute the increasingd15N values observed in months six to three to pathologi-cal processes with any certainty. Changes in carbon andnitrogen isotopic compositions in the final few monthsbefore death are unlikely to be resulting from diet—the
carbon-isotope compositions essentially do not changeduring the last four months, whereas the d15N valuesdecrease as described above. The rising cortisol levelsduring the last two months ante-mortem indicateincreased stress during this period which, combinedwith the rapid decrease in d15N values—suggestive of apositive nitrogen balance—may be attributable to injuryrequiring tissue repair.
Illness, trauma, and nutritional stress
There is a synergistic relationship between illness andnutrition (Scrimshaw and SanGiovanni 1997), and the impactof illness and infection on nutritional status is dependent onthe adequacy of an individual’s usual diet, dietary intake dur-ing and after the period of illness, and the nature and dura-tion of the illness or infection. Malnutrition during illnessmay result from, for example, therapeutic withdrawal
Fig. 3. Cortisol levels for individuals (A) engaged in residen-tial mobility (dashed lines for LTR 593 indicate 3 cm samplinginterval), (B) experiencing illness or trauma, and (C) with non-specific stress.
INTEGRATING CORTISOL AND ISOTOPIC DATA FROM HAIR 11
American Journal of Physical Anthropology
of food, decreased intestinal absorption of differentnutrients and vitamins, or an array of metabolicchanges associated with the catabolic response to ill-ness. Moreover, poor diet can increase susceptibility toinfection or illness, and if severe, can impair resistanceto infection. Nutritional status during tissue synthesisand repair post-trauma impacts both the isotopic com-position of newly formed tissues, and the likelihood ofco-occurring fever or infection (Beisel, 1972; Scrim-
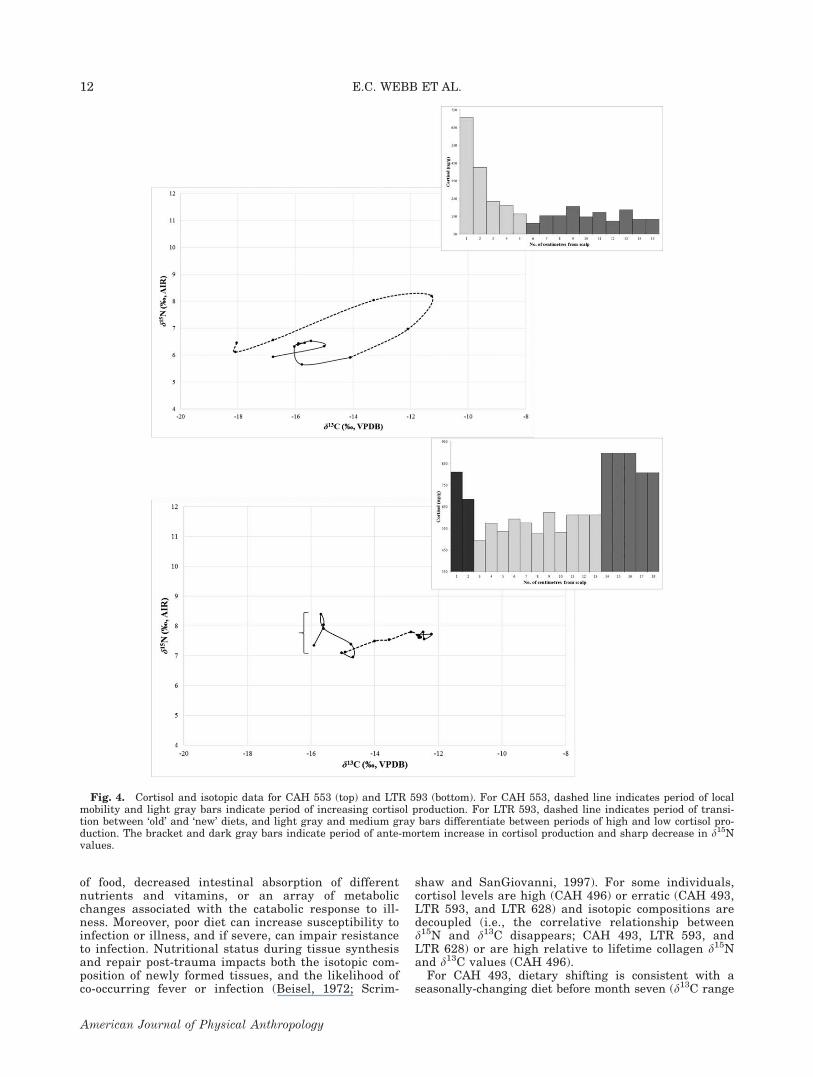
shaw and SanGiovanni, 1997). For some individuals,cortisol levels are high (CAH 496) or erratic (CAH 493,LTR 593, and LTR 628) and isotopic compositions aredecoupled (i.e., the correlative relationship betweend15N and d13C disappears; CAH 493, LTR 593, andLTR 628) or are high relative to lifetime collagen d15Nand d13C values (CAH 496).
For CAH 493, dietary shifting is consistent with aseasonally-changing diet before month seven (d13C range
Fig. 4. Cortisol and isotopic data for CAH 553 (top) and LTR 593 (bottom). For CAH 553, dashed line indicates period of localmobility and light gray bars indicate period of increasing cortisol production. For LTR 593, dashed line indicates period of transi-tion between ‘old’ and ‘new’ diets, and light gray and medium gray bars differentiate between periods of high and low cortisol pro-duction. The bracket and dark gray bars indicate period of ante-mortem increase in cortisol production and sharp decrease in d15Nvalues.
12 E.C. WEBB ET AL.
American Journal of Physical Anthropology
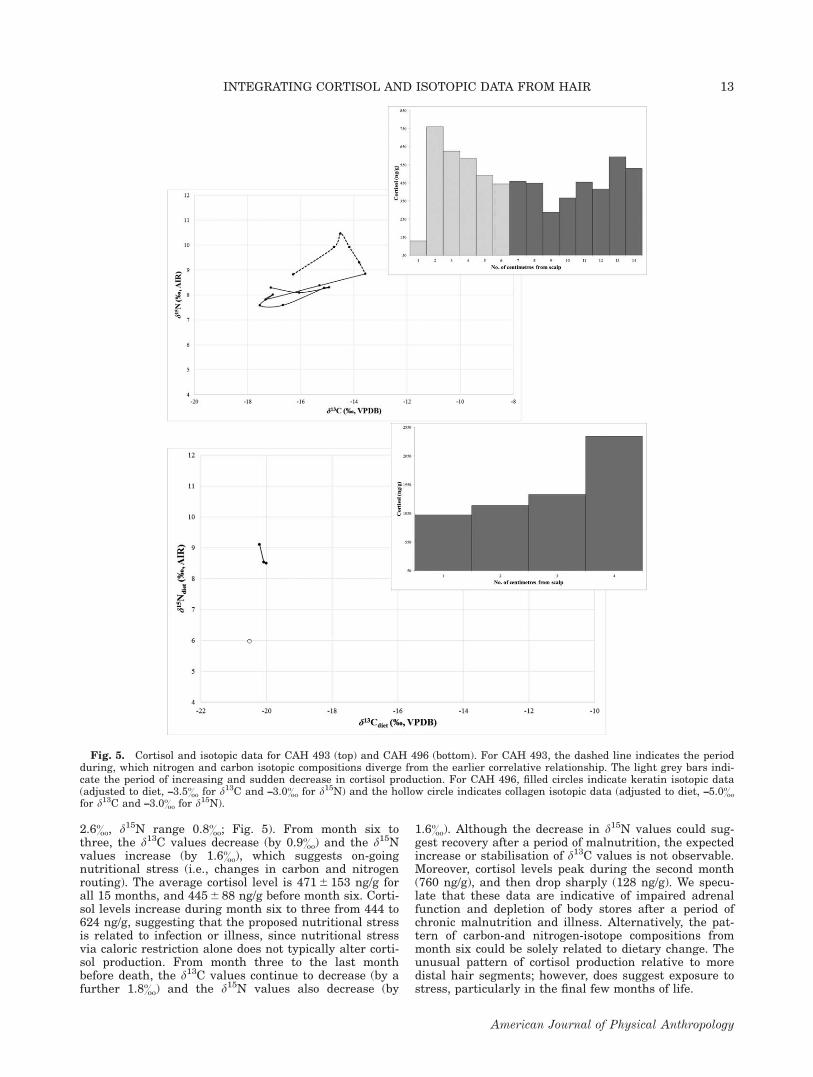
2.6&, d15N range 0.8&; Fig. 5). From month six tothree, the d13C values decrease (by 0.9&) and the d15Nvalues increase (by 1.6&), which suggests on-goingnutritional stress (i.e., changes in carbon and nitrogenrouting). The average cortisol level is 471 6 153 ng/g forall 15 months, and 445 6 88 ng/g before month six. Corti-sol levels increase during month six to three from 444 to624 ng/g, suggesting that the proposed nutritional stressis related to infection or illness, since nutritional stressvia caloric restriction alone does not typically alter corti-sol production. From month three to the last monthbefore death, the d13C values continue to decrease (by afurther 1.8&) and the d15N values also decrease (by
1.6&). Although the decrease in d15N values could sug-gest recovery after a period of malnutrition, the expectedincrease or stabilisation of d13C values is not observable.Moreover, cortisol levels peak during the second month(760 ng/g), and then drop sharply (128 ng/g). We specu-late that these data are indicative of impaired adrenalfunction and depletion of body stores after a period ofchronic malnutrition and illness. Alternatively, the pat-tern of carbon-and nitrogen-isotope compositions frommonth six could be solely related to dietary change. Theunusual pattern of cortisol production relative to moredistal hair segments; however, does suggest exposure tostress, particularly in the final few months of life.
Fig. 5. Cortisol and isotopic data for CAH 493 (top) and CAH 496 (bottom). For CAH 493, the dashed line indicates the periodduring, which nitrogen and carbon isotopic compositions diverge from the earlier correlative relationship. The light grey bars indi-cate the period of increasing and sudden decrease in cortisol production. For CAH 496, filled circles indicate keratin isotopic data(adjusted to diet, –3.5& for d13C and –3.0& for d15N) and the hollow circle indicates collagen isotopic data (adjusted to diet, –5.0&
for d13C and –3.0& for d15N).
INTEGRATING CORTISOL AND ISOTOPIC DATA FROM HAIR 13
American Journal of Physical Anthropology
For CAH 496, the isotopic compositions are essentiallystable (d13C range 0.2&, d15N range 0.6&) during thethree-month period represented (Fig. 5). The d15Ncol
value is 19.0& and the d13Ccol is –15.5&. Adjusting ker-atin and collagen isotopic compositions to compensatefor different tissue-diet isotopic offsets reveals an ante-mortem increase of 12.7& for d15N values and essen-tially no change in d13C values relative to long-termdiet. The average cortisol level is 1494 6 616 ng/g andcortisol production decreases from month four to themonth before death. Although the change in d15Nker val-ues relative to long-term diet (d15Ncol) may result fromconsumption of higher trophic level protein, the absenceof a co-occurring change in the d13C values and veryhigh cortisol levels suggest otherwise. It is instead likelythat the high d15Nker values indicate a change in nitro-gen metabolism. The profound negative nitrogen balanceand persistent hypercortisolemia may thus be indicativeof critical illness.
Nonspecific stress
Cortisol is produced in response to both physiologicaland psychosocial stressors (Miller et al., 2007). Eachindividual’s response to a given stressful situation isdependent on his or her ability to cope, mediated by per-ception of the situation and by overall state of health(McEwen, 1998). Metabolic changes could also bemasked by dietary shifting or provision of a nutritionallyadequate diet during the period of illness. Moreover, aspreviously mentioned, there is a two-week blind spotimmediately preceding death (Nakamura et al., 1982).Hence, for individuals for whom death was compara-tively sudden, it may not be possible to make inferencesabout stress experiences and morbidity using archeologi-cal hair. The lack of a significant change in hair cortisollevels may thus make a comparatively sudden death themore likely explanation for these individuals. For CAH
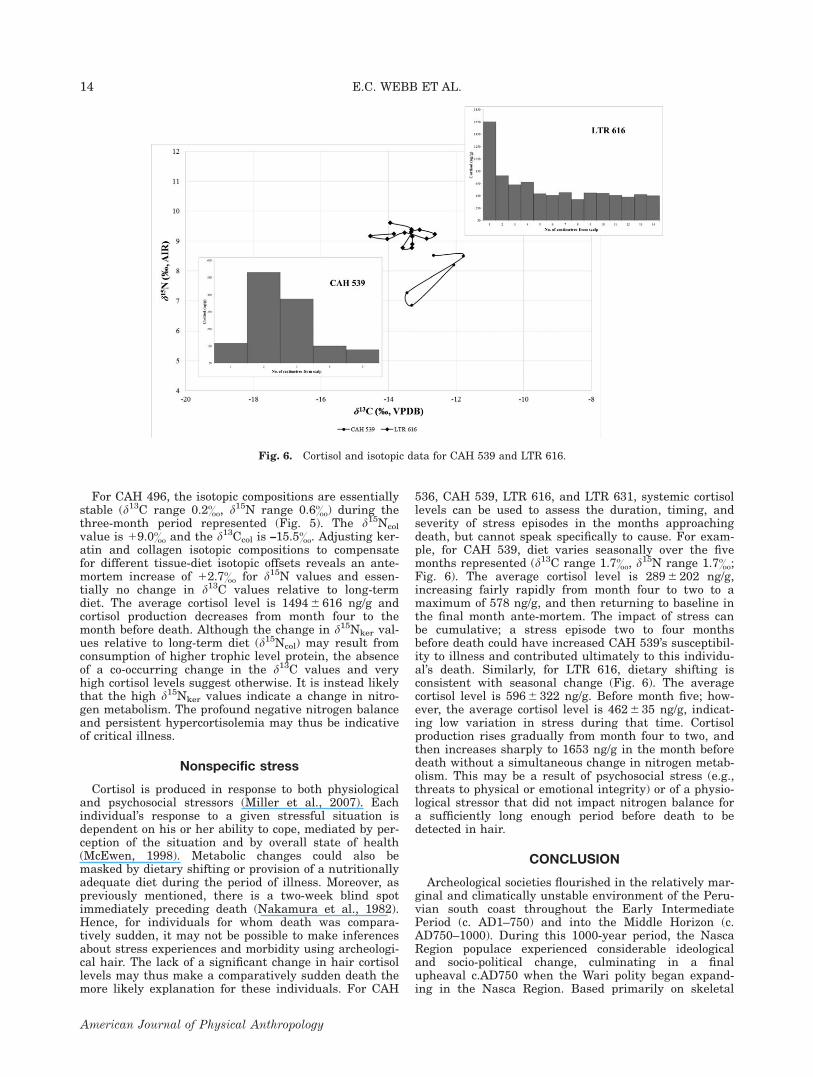
536, CAH 539, LTR 616, and LTR 631, systemic cortisollevels can be used to assess the duration, timing, andseverity of stress episodes in the months approachingdeath, but cannot speak specifically to cause. For exam-ple, for CAH 539, diet varies seasonally over the fivemonths represented (d13C range 1.7&, d15N range 1.7&;Fig. 6). The average cortisol level is 289 6 202 ng/g,increasing fairly rapidly from month four to two to amaximum of 578 ng/g, and then returning to baseline inthe final month ante-mortem. The impact of stress canbe cumulative; a stress episode two to four monthsbefore death could have increased CAH 539’s susceptibil-ity to illness and contributed ultimately to this individu-al’s death. Similarly, for LTR 616, dietary shifting isconsistent with seasonal change (Fig. 6). The averagecortisol level is 596 6 322 ng/g. Before month five; how-ever, the average cortisol level is 462 6 35 ng/g, indicat-ing low variation in stress during that time. Cortisolproduction rises gradually from month four to two, andthen increases sharply to 1653 ng/g in the month beforedeath without a simultaneous change in nitrogen metab-olism. This may be a result of psychosocial stress (e.g.,threats to physical or emotional integrity) or of a physio-logical stressor that did not impact nitrogen balance fora sufficiently long enough period before death to bedetected in hair.
CONCLUSION
Archeological societies flourished in the relatively mar-ginal and climatically unstable environment of the Peru-vian south coast throughout the Early IntermediatePeriod (c. AD1–750) and into the Middle Horizon (c.AD750–1000). During this 1000-year period, the NascaRegion populace experienced considerable ideologicaland socio-political change, culminating in a finalupheaval c.AD750 when the Wari polity began expand-ing in the Nasca Region. Based primarily on skeletal
Fig. 6. Cortisol and isotopic data for CAH 539 and LTR 616.
14 E.C. WEBB ET AL.
American Journal of Physical Anthropology

remains, Kellner (2002) concluded that, as a group, thepeople inhabiting the Nasca Region were not underexcessive physiological stress as compared to otherarcheological skeletal samples, but that there was a com-paratively high incidence of trauma indicative of inter-personal violence. The data presented here enhance thisunderstanding of the impact of stress, as well as thepotential causes of that stress.
Two distinct types of movement have been inferredfrom the isotopic data for several individuals: local mobil-ity and relocation. Cortisol production and, by inference,experienced stress (i.e., physical and/or psychosocial)were consistently different between those who wereactively moving and those who relocated. For those indi-viduals engaged in local mobility, increases in cortisol lev-els were coincident with periods of rapid change inisotopic composition over comparatively large ranges (e.g.,CAH 553). Although metabolic changes in isotopic compo-sition would likely be masked by rapid dietary shifting(and would enable differentiation between pathologicaland psychological stress), these data allow unique insightinto the physical and psychosocial stress associated withmovement among production zones. In contrast, compar-ing cortisol levels before and after (semi-) permanent relo-cation reveals a significant reduction in cortisolproduction, and therefore in stress, associated with reset-tling in a new locale. This finding raises the possibilitythat the motivation for relocation may have been relatedto quality of life, and that these individuals chose to moveto a place where they would be exposed to fewer/lesssevere stressors. The combined datasets enabled thereconstruction of individual experiences of residentialmobility and shed light on the affective experiences ofthese ancient inhabitants of the Nasca Region.
Considered together, the isotopic and cortisol data sug-gest a relatively high rate of exposure to stress that is con-sistent with what is known about the Nasca social andphysical environments. Of the 14 adults included in thisstudy, six likely suffered from illness/trauma before death,and a further three experienced stress without an observ-able associated change in isotopic composition. For someindividuals, repeated stress episodes during the year pre-ceding death are apparent, and indicate cumulative stress.For the most part, however, there is a period of stress andillness extending between two and six months ante-mortem, which in agreement with Kellner’s (2002)appraisal of overall Nasca health status, suggests thatthese individuals were moderately healthy and able tocope for some time before eventually succumbing. Thesedata thus reveal individual characteristics of frailty andrisk that would not be apparent using more traditionalmethods of evaluating health status, and also supportpopulation-level inferences. Thus, this analytical approachprovides a powerful enhancement to the understanding ofstress, morbidity and well-being developed through skele-tal analysis. Finally, it should be noted that neither cortisolnor isotopic data would reflect evidence for stress or meta-bolic changes associated with illness occurring within twoweeks of death. Barring much older traumas, it is alsolikely that all such individuals would appear essentiallyhealthy based on a skeletal assessment.
The combination of nitrogen- and carbon-isotope andsystemic cortisol level analyses has enabled detaileddescriptions of individual experiences during the periodpreceding death represented by each hair sample. These14 case studies demonstrate both inter- and intraindivid-ual differences in diet, health, stress, and residential
mobility, and emphasize the interaction of these factorsin producing the biochemical record archived in hair.The patterns of cortisol production associated with dif-ferent forms of residential mobility (as inferred from die-tary shifting) were surprisingly consistent amongindividuals. Beyond a series of case studies, this articlefurther provides a useful set of working guidelines forintegrating isotopic and cortisol data to make increas-ingly specific inferences about the causes and impact ofexperienced stress. By using cortisol levels to identifystressful periods, the physiological, metabolic, or behav-ioral (i.e., diet and/or mobility) causes for change innitrogen- and carbon-isotope compositions were clarifiedconsiderably. Future clinical research exploring the rela-tionships among nitrogen- and carbon-isotope composi-tion, nitrogen metabolism, and cortisol production inhealthy and ill adults would add greater certainty to theinferences made.
ACKNOWLEDGMENTS
The authors thank Dr. Gisela Grupe, Dr. GeorgeMcGlynn, and the State Collection for Anthropology andPalaeoanatomy, Munich, Germany for providing archeo-logical samples for analysis, and Dr. Karyn Olsen for col-lecting samples. They also thank Rachel Gow foranalytical support. This is Laboratory for Stable IsotopeScience Contribution #316.
LITERATURE CITED
Ambrose SK. 1993. Isotopic analysis of paleodiets: methodologi-cal and interpretive considerations. In: Sandford MK, editor.Investigations of ancient human tissue: chemical analyses inanthropology. Langhorne, Pennsylvania: Gordon and BreachScience. p 59–130.
Ambrose SK. 2000. Controlled diet and climate experiments onnitrogen isotope ratios of rats. In: Ambrose SH, KatzenbergMA, editors. Biogeochemical approaches to paleodietary anal-ysis. New York: Kluwer Academic/Plenum Publishers.
Ball SG. 2008. Metabolic and endocrine changes in sepsis andthe catabolic state. In: Baudouin S, editor. Sepsis. London:Springer-Verlag. p 11–16.
Beisel WR. 1972. Interrelated changes in host metabolism dur-ing generalized infectious illness. Am J Clin Nutr 25:1254–1260.
Beishuizen A, Thijs LG, Vermes I. 2001. Patterns ofcorticosteroid-binding globulin and the free cortisol index dur-ing septic shock and multitrauma. Intens Care Med 27:1584–1591.
Beresford-Jones DG, Torres SA, Whaley OQ, Chepstow-LustyAJ. 2009. The role of prosopis in ecological and landscapechange in the Samaca Basin, Lower Ica Valley, South CoastPeru from the Early Horizon to the Late Intermediate Period.Lat Am Antiq 20:303–332.
Bird BW, Abbott MB, Vuille M, Rodbell DT, Stansell ND,Rosenmeier MF. 2011. A 2300-year-long annually resolvedrecord of the South American summer monsoon from thePeruvian Andes. Proc Natl Acad Sci U S A 108:8583–8588.
Bol R, Pflieger C. 2002. Stable isotope (13C, 15N and 34S) anal-ysis of the hair of modern humans and their domestic ani-mals. Rapid Commun Mass Spectrom 16:2195–2200.
Browne DM, Silverman H, Garcia R. 1993. A cache of Nascatrophy heads from Cerro Carapo, Peru. Lat Am Antiq 4:274–294.
Cagigao ET. 2009. Talking bones: bioarchaeological analysis ofindividuals from Palpa. In: Reindel M, Wagner G, editors.New technologies for archaeology. Berlin: Springer-Verlag. p141–158.
Conlee CA, Schreiber KJ. 2006. The role of intermediate elitesin the Balkanization and reformation of post-Wari society in
INTEGRATING CORTISOL AND ISOTOPIC DATA FROM HAIR 15
American Journal of Physical Anthropology

Nasca, Peru. In: Elson CM, Covey RA, editors. Intermediateelites in Pre-Columbian states and empires. Tucson: Univer-sity of Arizona Press. p 94–111.
Coplen TB. 1994. Reporting stable hydrogen, carbon and oxygenisotope abundances. Pure Appl Chem 66:271–276.
Coplen TB. 1996. New guidelines for reporting stable hydrogen,carbon, and oxygen isotope-ratio data. Geochim CosmochimActa 60:3359–3360.
Davenport M, Tiefenbacher S, Lutz CK, Novak MA, Meyer JS.2006. Analysis of endogenous cortisol concentrations in thehair of rhesus macaques. Gen Comp Endocr 147:255–261.
Dettenborn L, Muhtz C, Skoluda N, Stalder T, Steudte S,Hinkelmann K, Otte C. 2012. Introducing a novel method toassess cumulative steroid concentrations: increased hair corti-sol concentrations over 6 months in medicated patients withdepression. Stress 15:348–353.
Edwards MJ. 2010. Archaeological investigations at Pataraya: aWari outpost in the Nasca valley of Southern Peru. Disserta-tion, University of California.
Eitel B, Hecht S, M€achtle B, Schukraft G, Kadereit A, WagnerGA, Kromer B, Unkel I, Reindel M. 2005. Geoarchaeologicalevidence from desert loess in the Nazca-Palpa region, south-ern Peru: palaeoenvironmental changes and their impact onpre-Columbian cultures. Archaeometry 47:137–158.
Eitel B, M€achtle B. 2009. Man and environment in the easternAtacama Desert (Southern Peru): Holocene climate changesand their impact on Pre-Columbian cultures. In: Reindel M,Wagner G, editors. New technologies for archaeology.Springer-Verlag: Berlin. p 17–37.
Fichter MM, Pirke K-M, Holsboer F. 1986. Weight loss causesneuroendocrine disturbances: experimental study in healthystarving subjects. Psychiatr Res 17:61–72.
Fuller BT, Fuller JT, Sage NE, Harris DA, O’Connell TC,Hedges REM. 2004. Nitrogen balance and d15N: why you’renot what you eat during pregnancy. Rapid Commun MassSpectrom 18:2889–2896.
Fuller BT, Fuller JT, Sage NE, Harris DA, O’Connell TC,Hedges REM. 2005. Nitrogen balance and d15N: why you’renot what you eat during nutritional stress. Rapid CommunMass Spectrom 19:2497–2506.
Goodman A, Thomas R, Swedlund A, Armelagos G. 1988. Bio-cultural perspectives on stress in prehistoric, historical andcontemporary population research. Yearb Phys Anthropol 31:169–202.
Gow R, Thomson S, Rieder M, Van Uum S, Koren G. 2010. Anassessment of cortisol analysis in hair and its clinical applica-tions. Forensic Sci Int 196:32–37.
Hansen AM, Garde AH, Skovgaard LT, Christensen JM. 2001.Seasonal and biological variation of urinary epinephrine, nor-epinephrine, and cortisol in healthy women. Clin Chim Acta309:25–35.
Harding H, Rogers G. 1999. Physiology and growth of humanhair. In: Robertson J, editor. Forensic examination of hair.Philadelphia: Taylor and Francis. p 1–77.
Heaton THE, Vogel JC, von la Chevallerie G, Collet G. 1986.Cli-matic influence on the isotopic composition of bone nitrogen.Nature 322:822–823.
Hobson K, Alisauskas R, Clark R, 1993. Stable-nitrogen isotopeenrichment in avian tissues due to fasting and nutritionalstress: implications for isotopic analysis of diet. Condor 95:388–394.
Hoefs J. 2004. Stable isotope geochemistry. New York: Springer.Huelsemann F, Flenker U, Koehler K, Schaenzer W. 2009.
Effect of a controlled dietary change on carbon and nitrogenstable isotope ratios of human hair. Rapid Commun MassSpectrom 23:2448–2454.
Johnstone AM, Faber P, Andrew R, Gibney ER, Elia M, LobleyGM, Stubbs RJ, Walker BJ. 2004. Influence of short-term die-tary weight loss on cortisol secretion and metabolism in obesemen. Eur J Endocrinol 150:185–194.
Kalra S, Einarson A, Karaskov T, Van Uum S, Koren G. 2007.The relationship between stress and hair cortisol in healthypregnant women. Clin Invest Med 30:E103–E107.
Kantner J, Vaughn KJ. 2012. Pilgrimage as costly signal: reli-giously motivated cooperation in Chaco and Nasca.J Anthropol Archaeol 31:66–82.
Katzenberg MA, Lovell NM. 1999. Stable isotope variation inpathological bone. Int J Osteoarchaeol 9:316–324.
Kellner CM. 2002. Coping with environmental and social chal-lenges in prehistoric Peru: bioarchaeological analyses ofNasca populations. Santa Barbara, California: University ofCalifornia.
Kirschbaum C, Tietze A, Skoluda N, Dettenborn L. 2009. Hairas a retrospective calendar of cortisol production—Increasedcortisol incorporation into hair in the third trimester of preg-nancy. Psychoneuroendocrino 34:32–37.
Knudson KJ, Aufderheide AE, Buikstra JE. 2007. Seasonalityand paleodiet in the Chiribaya polity of southern Peru.J Archaeol Sci 34:452–462.
Knudson KJ, Pestle WJ, Torres-Rouff C, Pimentel G. 2012.Assessing the life history of an Andean traveller through bio-geochemistry: stable and radiogenic isotope analyses ofarchaeological human remains from Northern Chile. Int JOsteoarchaeol 22:435–451.
Knudson KJ, Williams SR, Osborn R, Forgey K, Williams PR.2009. The geographic origins of Nasca trophy heads usingstrontium, oxygen, and carbon isotope data. J AnthropolArchaeol 28:244–252.
Kudielka BM, Buske-Kirschbaum A, Hellhammer DH,Kirschbaum C. 2004. HPA axis responses to laboratory psy-chosocial stress in healthy elderly adults, younger adults, andchildren: impact of age and gender. Psychoneuroendocrino 29:83–98.
LeBeau MA, Montgomery MA, Brewer JD. 2011. The role ofvariations in growth rate and sample collection on interpret-ing results of segmental analyses of hair. Forensic Sci Int210:110–116.
Lee-Thorp JA, Sealy JC, van der Merwe NJ. 1989. Stable car-bon isotope ratio differences between bone collagen and boneapatite, and their relationship to diet. J Archaeol Sci 16:585–599.
Manary MJ, Muglia LJ, Vogt SK, Yarasheski KE. 2006. Cortisoland its action on the glucocorticoid receptor in malnutritionand acute infection. Metabolism 55:550–554.
Mariotti A. 1983. Atmospheric nitrogen is a reliable standardfor natural 15N abundance measurements. Nature 303:685–687.
Matchock RL, Dorn LD, Susman EJ. 2007. Diurnal and sea-sonal cortisol, testosterone, and DHEA rhythms in boys andgirls during puberty. Chronobiol Int 24:969–990.
McCullagh JSO, Tripp JA, Hedges REM. 2005. Carbon isotopeanalysis of bulk keratin and single amino acids from Britishand North American hair. Rapid Commun Mass Spectrom 19:3227–3231.
McEwen B.S. 1998. Protective and damaging effects of stressmediators. New Engl J Med 338:171–179.
Mekota A, Grupe G, Ufer S, Cuntz U. 2006. Serial analysis ofstable nitrogen and carbon isotopes in hair: monitoringstarvation and recovery phases of patients suffering fromanorexia nervosa. Rapid Commun Mass Spectrom 20:1604–1610.
Mekota A, Grupe G, Ufer S, Cuntz U. 2009. Identifying starva-tion episodes using stable isotopes in hair. Rechtsmedizin 19:431–440.
Miller G, Chen E, Zhou ES. 2007. If it goes up, must it comedown? Chronic stress and the hypothalamic-pituitary-adrenocortical axis in humans. Psychol Bull 133:25–45.
Mojtahedi, M., de Groot, L., Boekholt, H.A. van Raaij, J.M.A.2002. Nitrogen balance of healthy Dutch women before andduring pregnancy. Am J Clin Nutr. 75:1078–1083.
Murra JV. 1972. El control vertical de un maximo de pisos eco-logicos en las economia de las sociedades Andinas. Visita dela Provincia de Leon de Huanuco en 1562. Documentos por laHistoria y Etnologia de Huanuco y la Selva Central 2:427–476.
Nakamura K, Schoeller D, Winkler FJ, Schmidt HL. 1982. Geo-graphical variation in the carbon isotope composition of the
16 E.C. WEBB ET AL.
American Journal of Physical Anthropology

diet and hair in contemporary man. Biomed Mass Spectrom9:390–394.
Novak MA, Hamel AF, Kelly BJ, Dettmer AM, Meyer JS. 2013.Stress, the HPA axis, and nonhuman primate well-being: areview. Appl Anim Behav Sci 143:135–149.
O’Connell TC, Hedges REM. 1999. Investigations into the effectof diet on modern human hair isotopic values. Am J PhysAnthropol 108:409–425.
O’Connor TM, O’Halloran DJ, Shanahan F. 2000. The stressresponse and the hypothalamic–pituitary–adrenal axis: frommolecule to melancholia. Q J Med 93:323–333.
Olsen KC, White CD, Longstaffe FJ, von Heyking K, McGlynnG, Grupe G, R€uhli FJ. 2013. Intraskeletal isotopic composi-tions (d13C, d15N) of bone collagen: nonpathological andpathological variation. Am J Phys Anthropol 153:598–604.
Orefici G. 2006 Reconstructing Nasca domestic and ceremoniallife: a view from Cahuachi and Pueblo Viejo. In: Shimada I,Baba H, Shinoda K, Mashiro O, editors. Nasca, wonder of theworld, messages etched on the desert floor. Tokyo: TBS. p176–184.
Orefici G, Drusini A. 2003. Nasca: Hipotesis y Evidencias de suDesarrollo Cultural. BresciaLima: Centro Italiano Studi eRicerche Archeologiche Precolombiane, Lima, Peru.
Petzke KJ, Boeing H, Klaus S, Metges CC. 2005. Carbon andnitrogen stable isotopic composition of hair protein and aminoacids can be used as biomarkers for animal-derived dietaryprotein intake in humans. J Nutr 135:1515–1520.
Petzke KJ, Feist T, Fleig WE, Metges CC. 2006. Nitrogen iso-topic composition in hair protein is different in liver cirrhoticpatients. Rapid Commun Mass Spectrom 20:2973–2978.
Proulx DA. 1989. Nasca trophy heads: victims of warfare or rit-ual sacrifice? In: Tkaczuk DC, Vivian BC, editors. Cultures inconflict: current archaeological perspectives. Calgary: Univer-sity of Calgary Archaeological Association. p 73–85.
Proulx DA. 2001. Ritual uses of trophy heads in ancient Nascasociety. In: Benson EP, Cook AG, editors. Ritual sacrifice inancient Peru. Austin: University of Texas Press. p 119–136.
Russell E, Koren G, Rieder M, Van Uum S. 2012. Hair cortisolas a biological marker of chronic stress: current studies,future directions and unanswered questions. Psychoneuroen-docrino 37:589–601.
Sapolsky RM. 2004. Social status and health in humans andother animals. Annu Rev Anthropol 33:393–418.
Sapolsky RM, Romero LM, Munck AU. 2000. How do glucocorti-coids influence stress responses? Integrating permissive, sup-pressive, stimulatory and preparative actions. Endocr Rev 21:55–89.
Sapse AT. 1997. Cortisol, high cortisol diseases and anti-cortisoltherapy. Psychoneuroendocrino 22(S1):S3–S10.
Sauv�e B, Koren G, Walsh G, Tokmakejian S, Van Uum SH.2007. Measurement of cortisol in human hair as a biomarkerof systemic exposure. Clin Invest Med 30:E183–E191.
Schreiber K. 2005. Imperial agendas and local agency: Waricolonial strategies. In: Stein GJ, editor. The archaeology ofcolonial encounters: comparative perspectives. Santa Fe:School of American Research Press. p 237–262.
Schreiber K, Lancho Rojas J. 2003. Irrigation and society in thePeruvian desert: the puquios of Nasca. New York: LexingtonBooks.
Schreiber KJ. 2001. The Wari Empire of Middle Horizon Peru:the epistemological challenge of documenting an empire with-out documentary evidence. In: Alcock SE, D’Altroy TN, Morri-son KD, editors. Empires. Cambridge: University ofCambridge Press. p 70–92.
Schwarcz H, White C. 2004. The grasshopper or the ant?Cultigen-use strategies in ancient Nubia from C-13 analysisof human hair. J Archaeol Sci 31:753–762.
Schoeller DA. 1999. Isotope fractionation: why aren’t we whatwe eat. J Archaeol Sci 26:667–673.
Schwertl M, Auerswald K, Schnyder H. 2003. Reconstruc-tion of the isotopic history of animal diets by hair seg-mental analysis. Rapid Commun Mass Spectrom 17:1312–1318.
Scrimshaw NS, SanGiovanni JP. 1997. Synergism of nutrition,infection and immunity: an overview. Am J Clin Nutr 66:464S–477S.
Silverman H. 1993. Cahuachi in the ancient Nasca world. IowaCity: University of Iowa Press.
Støving RK, Hangaard J, Hansen-Nord M, Hagen C. 1999. Areview of endocrine changes in anorexia nervosa. J PsychiatRes 33:139–152.
Strong WD. 1957. Paracas, Nazca and Tiahuanacoid Culturalrelationships in south coastal Peru. Mem Soc Am Archaeol13:1–48.
Tello JC. 1918. El uso de las cabezas humanas artificialmentemomificadas y su representacion en el antiguo arte peruano.Revista Universitaria 2:477–533.
Thompson LG, Mosley-Thompson E, Bolzan JF, Koci BR. 1985.A 1500-year record of tropical precipitation in ice cores fromthe Quelccaya Ice Cap, Peru. Science 229:971–973.
Thomson S, Koren G, Fraser L-A, Rieder M, Friedman TC, VanUum SHM. 2009. Hair analysis provides a historical record ofcortisol levels in Cushing’s syndrome. Exp Clin Endocr Diab117:1–6.
Tieszen LL, Senyimba MM, Imbamba SK, Troughton JH. 1979.The distribution of C3 and C4 grasses and carbon isotope dis-crimination along an altitudinal and moisture gradient inKenya. Oecologia 37:337–350.
Tomczak PD. 2003. Prehistoric diet and socioeconomic relation-ships within the Osmore Valley of southern Peru. J AnthropolArchaeol 22:262–278.
Tung TA. 2007. Trauma and violence in the Wari Empire of thePeruvian Andes: Warfare, raids, and ritual fights. Am J PhysAnthropol 133:941–956.
Ubbelohde Doering H. 1958. Bericht €uber arch€aologische Fel-darbeitein in Per�u. Ethnos 2–4:67–99.
Ubbelohde Doering H. 1966. On the royal highways of the Inca.New York: Frederick A. Praeger Publishers.
Valenzuela LO, Chesson LA, O’Grady SP, Cerling TE,Ehleringer JR. 2011. Spatial distributions of carbon,nitrogen and sulfur isotope ratios in human hair across thecentral United States. Rapid Commun Mass Spectrom 25:861–868.
Van Cauter E, Leproult R, Kupfer DJ. 1996. Effects of genderand age on the levels and circadian rhythmicity of plasmacortisol. J Clin Endocr Metab 81:2468–2473.
Van Uum SHM, Sauv�e B, Fraser LA, Morley-Forster P, PaulTL, Koren G. 2008. Elevated content of cortisol in hair ofpatients with severe chronic pain: a novel biomarker forstress. Stress 11:483–488.
Vaughn KJ. 2009. The ancient Andean village: Marcaya in pre-hispanic Nasca. Tucson: University of Arizona Press.
Verano JW. 1995. Where do they rest? The treatment of humanofferings and trophies in ancient Peru. In: Dillehay T, editor.Tombs for the living: Andean mortuary practices. Washington,DC: Dumbarton Oaks Research Library and Collection. p189–228.
Verano JW. 2003. Mummified trophy heads from Peru: diagnos-tic features and medicolegal significance. J Forensic Sci 48:525–530.
Waterlow JC. 1999. The mysteries of nitrogen balance. NutrRes Rev 12:25–54.
Webb E, Thomson S, Nelson A, White C, Rieder M, Koren G,Van Uum S. 2010. Assessing individual systemic stressthrough cortisol analysis of archaeological hair. J ArchaeolSci 37:807–812.
Webb E, White C, Longstaffe F. 2013. Dietary shifting in theNasca Region as inferred from the carbon-and nitrogen-iso-tope compositions of archaeological hair and bone. J ArchaeolSci 40:129–139.
White CD. 1993. Isotopic determination of seasonality in dietand death from Nubian mummy hair. J Archaeol Sci 20:657–666.
White CD, Nelson AJ, Longstaffe FJ, Grupe G, Jung A. 2009. Land-scape bioarchaeology at Pacatnamu, Peru: inferring mobilityfrom d13C and d15N values of hair. J Archaeol Sci 36:1527–1537.
INTEGRATING CORTISOL AND ISOTOPIC DATA FROM HAIR 17
American Journal of Physical Anthropology

Williams JS, Katzenberg AM. 2012. Seasonal fluctuations in diet anddeath during the late horizon: a stable isotopic analysis of hair andnail from the central coast of Peru. J Archaeol Sci 39:41–57.
Williams LJ, White CD, Longstaffe FJ. 2011. Improving stableisotopic interpretations made from human hair through
reduction of growth cycle error. Am J Phys Anthropol 145:125–136.
Wood JW, Milner GR, Harpending HC, Weiss KM. 1992. Theosteological paradox: problems of inferring prehistoric healthfrom skeletal samples. Curr Anthropol 33:343–370.
18 E.C. WEBB ET AL.
American Journal of Physical Anthropology




















