
Inhibitory effect of state independent ponto-geniculo-occipital waves on seizure occurrence induced...
10
Inhibitory effect of state independent ponto-geniculo-occipital waves on seizure occurrence induced by local application of penicillin into the temporal lobe amygdala Ignacio Ramírez Salado ⁎ , Ana Paula Rivera García, Manuel Alejandro Cruz Aguilar, José María Calvo Departamento de Cronobiología, División de Investigaciones en Neurociencias, Instituto Nacional de Psiquiatría “Ramón de la Fuente Muñiz”, Calz. México-Xochimilco 101, Col. Sn Lorenzo Huipulco, 14370. México, D.F. México abstract article info Article history: Received 23 May 2008 Received in revised form 19 June 2008 Accepted 10 July 2008 Available online 15 July 2008 Keywords: Amygdala Carbachol Epilepsy Peribrachial area (PBL) Ponto-geniculo-occipital waves (PGO) Sleep Purpose: In order to explore the possible inhibitory role of the phasic phenomena of REM sleep ponto- geniculo-occipital (PGO) waves over epilepsy, seizure activity produced by topic administration of Na- penicillin (PCN) has been analyzed during sustained PGO waves irrespective of current state. PGO waves were induced by the injection of carbachol in the peribrachial area. Methods: The development of acute experimental epilepsy was compared among nine chronically implanted, adult, male cats, by means of polygraphic 23h recordings. Our protocol consisted of sets of 4 trials: carbachol; PCN; carbachol followed by PCN and finally PCN followed by carbachol. Each cat received one single set and all trials were carried out with a seven days interval, in order to compare the epileptic activity both in the presence of PGOs and without them. Results: PGO waves 1) exert an inhibitory influence over number and duration of the Generalized Convulsive Seizures (GCSs) and 2) spike frequency; 3) increase the latency of GCSs; and 4) restores sleep alterations produced by experimental epilepsy. Conclusions: PGO waves exhibit an inhibitory influence over seizures induced by PCN. These data support the hypothesis that this phasic phenomenon of REM sleep have a depressant effect on epilepsy, inhibit seizures and normalize sleep architecture changes induced by seizures. We suggest that one possible function of PGO activity is to protect the brain from intense changes in neuronal excitability; namely convulsive activity. © 2008 Elsevier Inc. All rights reserved. 1. Introduction Clinical studies on sleep and epilepsy have shown that the rapid eye movement sleep (REM) (Aserinsky and Kleitman, 1953) has an inhibitory influence on epileptic seizures. The onset of generalized convulsive seizures (GCSs) never occurs during episodes of REM sleep (Gloor et al., 1977). Moreover, during this stage, the number of focal epileptic discharges is lower than the number found during wakeful- ness (W) or during slow wave sleep (SWS) (Delange et al., 1962; Cadilhac, 1982). When focal discharges occur during REM sleep, their amplitude is lowered and their propagation to other brain regions diminishes, becoming confined to the primary epileptic focus site (Montplaisir et al., 1982). Experimental studies in the cat also support the premise that REM sleep has an inhibitory effect on epilepsy. The threshold for electro- convulsive induced seizures reaches its highest values during REM sleep, as compared to those recorded during W and SWS stages (Cohen et al., 1970). In a pioneer experimental study in cats in 1971, Fernández- Guardiola and Ayala (1971) found that towards the end and during the extinction of experimental GCS in cats, both onset of rapid eye movements, sustained hippocampal theta activity and muscle atonia were similar to those occurring during REM sleep. These findings lead the authors to propose for the first time that the mechanisms responsible for REM sleep are similar to those responsible for seizure extinction. In previous studies we found that REM sleep also has an inhibitory effect on epileptogenesis. We compared the amygdaloid kindling development during REM sleep to the one delivered during W. We found that kindling development is significantly delayed when electrical stimuli are delivered during a burst of ponto-geniculo- occipital waves (PGO) of spontaneous REM sleep (Calvo, 1991), demonstrating that these potentials play an important role in the inhibition over seizures development. 1.1. PGO-activity PGO waves appear in REM sleep as well as in the immediately preceding slow wave sleep episode, the stage known as “sommeil phasique a ondes lentes” (SPHOL) (Thomas and Benoit, 1967), which Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697 Abbreviations: AM, temporal lobe amygdala; EE, experimental epilepsy; EEG, electroencephalogram; GCSs, generalized convulsive seizures; LDT, laterodorsal tegmentum; NPO, nucleus pontis oralis; PBL, peribrachial area; PCN, Na-penicillin; PGO, ponto-geniculo-occipital waves; PPT, pedunculopontine tegmentum; REM, rapid eye movements sleep; SPHOL, sommeil phasique à ondes lentes; SWS, slow wave sleep; SWS1, slow wave sleep 1; SWS2, slow wave sleep 2; W, wakefulness. ⁎ Corresponding author. E-mail address: [email protected] (I.R. Salado). 0278-5846/$ – see front matter © 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.pnpbp.2008.07.004 Contents lists available at ScienceDirect Progress in Neuro-Psychopharmacology & Biological Psychiatry journal homepage: www.elsevier.com/locate/pnpbp
-
Upload
unadmexico -
Category
Documents
-
view
0 -
download
0
Transcript of Inhibitory effect of state independent ponto-geniculo-occipital waves on seizure occurrence induced...

Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
Contents lists available at ScienceDirect
Progress in Neuro-Psychopharmacology &Biological Psychiatry
j ourna l homepage: www.e lsev ie r.com/ locate /pnpbp
Inhibitory effect of state independent ponto-geniculo-occipital waves on seizureoccurrence induced by local application of penicillin into the temporal lobe amygdala
Ignacio Ramírez Salado ⁎, Ana Paula Rivera García, Manuel Alejandro Cruz Aguilar, José María CalvoDepartamento de Cronobiología, División de Investigaciones en Neurociencias, Instituto Nacional de Psiquiatría “Ramón de la Fuente Muñiz”, Calz. México-Xochimilco 101, Col. Sn LorenzoHuipulco, 14370. México, D.F. México
Abbreviations: AM, temporal lobe amygdala; EE,electroencephalogram; GCSs, generalized convulsivetegmentum; NPO, nucleus pontis oralis; PBL, peribracPGO, ponto-geniculo-occipital waves; PPT, pedunculopoeye movements sleep; SPHOL, sommeil phasique à ondesSWS1, slow wave sleep 1; SWS2, slow wave sleep 2; W,⁎ Corresponding author.
E-mail address: [email protected] (I.R. Salado).
0278-5846/$ – see front matter © 2008 Elsevier Inc. Aldoi:10.1016/j.pnpbp.2008.07.004
a b s t r a c t
a r t i c l e i n f oArticle history:
Purpose: In order to explor Received 23 May 2008Received in revised form 19 June 2008Accepted 10 July 2008Available online 15 July 2008Keywords:AmygdalaCarbacholEpilepsyPeribrachial area (PBL)Ponto-geniculo-occipital waves (PGO)Sleep
e the possible inhibitory role of the phasic phenomena of REM sleep ponto-geniculo-occipital (PGO) waves over epilepsy, seizure activity produced by topic administration of Na-penicillin (PCN) has been analyzed during sustained PGO waves irrespective of current state. PGO waves wereinduced by the injection of carbachol in the peribrachial area.Methods: The development of acute experimental epilepsy was compared among nine chronically implanted,adult, male cats, by means of polygraphic 23h recordings. Our protocol consisted of sets of 4 trials: carbachol;PCN; carbachol followed by PCN and finally PCN followed by carbachol. Each cat received one single set andall trials were carried out with a seven days interval, in order to compare the epileptic activity both in thepresence of PGOs and without them.Results: PGO waves 1) exert an inhibitory influence over number and duration of the Generalized ConvulsiveSeizures (GCSs) and 2) spike frequency; 3) increase the latency of GCSs; and 4) restores sleep alterationsproduced by experimental epilepsy.Conclusions: PGO waves exhibit an inhibitory influence over seizures induced by PCN. These data support thehypothesis that this phasic phenomenon of REM sleep have a depressant effect on epilepsy, inhibit seizuresand normalize sleep architecture changes induced by seizures. We suggest that one possible function of PGOactivity is to protect the brain from intense changes in neuronal excitability; namely convulsive activity.
© 2008 Elsevier Inc. All rights reserved.
1. Introduction
Clinical studies on sleep and epilepsy have shown that the rapideye movement sleep (REM) (Aserinsky and Kleitman, 1953) has aninhibitory influence on epileptic seizures. The onset of generalizedconvulsive seizures (GCSs) never occurs during episodes of REM sleep(Gloor et al., 1977). Moreover, during this stage, the number of focalepileptic discharges is lower than the number found during wakeful-ness (W) or during slow wave sleep (SWS) (Delange et al., 1962;Cadilhac, 1982). When focal discharges occur during REM sleep, theiramplitude is lowered and their propagation to other brain regionsdiminishes, becoming confined to the primary epileptic focus site(Montplaisir et al., 1982).
Experimental studies in the cat also support the premise that REMsleep has an inhibitory effect on epilepsy. The threshold for electro-
experimental epilepsy; EEG,seizures; LDT, laterodorsal
hial area; PCN, Na-penicillin;ntine tegmentum; REM, rapidlentes; SWS, slow wave sleep;wakefulness.
l rights reserved.
convulsive induced seizures reaches its highest valuesduringREMsleep,as compared to those recorded during W and SWS stages (Cohen et al.,1970). In a pioneer experimental study in cats in 1971, Fernández-Guardiola and Ayala (1971) found that towards the end and during theextinction of experimental GCS in cats, both onset of rapid eyemovements, sustained hippocampal theta activity and muscle atoniawere similar to those occurring during REM sleep. These findings leadthe authors to propose for the first time that the mechanismsresponsible for REM sleep are similar to those responsible for seizureextinction. In previous studies we found that REM sleep also has aninhibitory effect on epileptogenesis. We compared the amygdaloidkindling development during REM sleep to the one delivered duringW.We found that kindling development is significantly delayed whenelectrical stimuli are delivered during a burst of ponto-geniculo-occipital waves (PGO) of spontaneous REM sleep (Calvo, 1991),demonstrating that these potentials play an important role in theinhibition over seizures development.
1.1. PGO-activity
PGO waves appear in REM sleep as well as in the immediatelypreceding slow wave sleep episode, the stage known as “sommeilphasique a ondes lentes” (SPHOL) (Thomas and Benoit, 1967), which

1689I.R. Salado et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
occurs just seconds before REM sleep onset. PGO waves of SPHOLoccur always in presence of sleep spindles, have a lesser frequencyrate, and occur mainly in a simple pattern, unlike to those of REMsleep.Whenever the PGOwaves increase their incidence, among othersigns (muscular atonia, EEG desynchronization and rapid eye move-ments) a REM sleep episode is initiated.
The occurrence of REM sleep has been shown to be regularlyassociated with the deactivation of a neuronal group (REM-off cells)located in the raphe nuclei (serotonergic population) and the nor-adrenergic group in the locus coeruleus, as well as with the reciprocalactivation of one neuronal population (REM-on cells), localized in thePeribrachial area (PBL) (Datta and Hobson, 1994; Datta and MacLean,2007). Among the many REM-on cell groups, one selectively activesubpopulation is the PGO burst cell complex of the PBL region. Spe-cifically, the pedunculopontine tegmentum (PPT), is involved in theREM generation through cholinergic cell activation, which in turn isregulated by glutamate (Datta, 2006). Since REM sleep is physiologi-cally mediated by cholinergic mechanisms (Datta et al., 1991), thecaudo-lateral part of PBL region is involved in the homeostaticregulation of REM sleep, and is the main structure responsible for PGOgeneration. A prolonged increase in REM sleep and other PGO relatedstates, following PBL stimulation by carbachol, has been demonstrated(Calvo et al., 1992; Datta et al., 1992). Taken together, these obser-vations indicate a physiological coupling between the PBL cholinergic/PGO trigger zone and the REM generator network that is both strongand enduring.
On the other hand some forebrain areas such as the limbic system,promote the generation of REM sleep, as well as the phasic elements ofthis phase (Calvo et al., 1987). Anatomical studies have shownreciprocal projections between the PBL and the amygdaloid centralnucleus (Nomura et al., 1979; Amaral et al., 1992). Electrophysiologicalstudies have demonstrated that REM PGO waves generated in thepons, but propagated and recorded in the LGB, also propagate with ahigher latency to the amygdala of the temporal lobe (40 ms), dorsalhippocampus (25 ms) and to the anterior and posterior regions of thecyngulus (30 ms) (Calvo and Fernández-Guardiola, 1984).
We suggest for the first time that the phasic phenomena of REMsleep, the PGOwaves, participate in seizures inhibition. In order to testthis, we induced PGO potentials, unrelated to those of REM sleep, bymeans of the microinjection of carbachol in the PBL (Vivaldi et al.,1980; Datta et al., 1992), and to induce experimental epilepsy, wemicroninjected Na-penicillin (PCN) into the temporal lobe amygdala(AM) (Fernández-Guardiola et al., 1991), with the aim of under-standing how PGO phenomena and epileptic activity interact, wepredict that experimentally induced PGO activity will have aninhibitory effect over seizures. This possible inhibitory effect will beassessed in two different trials: PGO effect over established epilepticactivity and PGO effect followed by epileptic activity. Both trials willallow us to analyze if PGO can only abolish previously establishedepileptic activity or if PGO waves can also avoid the generation of anepileptic focus. If so, PGO waves will probably have a restoring effectover sleep architecture changes induced by seizures, effect measuredthroughout the whole 23h recordings.
2. Methods
2.1. Surgical preparation
The experiments were carried out on nine freely moving,chronically implanted, adult, male cats (Felis domesticus), weighingbetween 3.2 and 3.8 kg. Animals were anesthetized with Propofol(200 mg/150 ml). The cats were prepared with stereotaxically (SniderandNiemer,1961) implanted electrodes in order tomake conventionalsleep recordings (stainless steel electrodes placed epidurally to recordthe electrocorticogram of the sensoriomotor and visual cortices, EEG;Nail-shaped electrodes placed in the supraorbital cavity to register the
electrooculogram, EOG; electromyogram of the nuchal muscles, EMG),and PGO wave recordings of both lateral geniculate bodies (A=6.5;L=9.5; V=+3.0). For the purpose ofmicroinjection of carbachol into thepontine brain stem and Na-penicillin (PCN) into the AM, stainless steelguide tubes were placed in all cats. Also all cats (N=9) had bilateralguide tubes (26 gauge) aimed stereotaxically at the PBL area(Angle=38°; P=4.0, L=4.5, V=−2.5). For PCN microinjection, bilateralguide tubes were placed above both temporal lobe amygdala (AM)(A=11.5; L=10; V=−0.5). Two stainless-steel wires were also adhered,exceeding the guide tubes by 1 mm, in order to record the elec-troenchephalographic activity of both AMs. All guide tubes were fixedin place using dental acrylic. A 31 gauge stainless steel sty1et was keptinserted in the guide tubes to ensure patency. The tips of the guidetubes were positioned 3 mm above the target sites. All experimentswere carried out following the technical guidelines for the production,care and use of laboratory animals, issued by the SAGARPA (NOM-062-ZOO-2002) and approved by the ethics committee of the InstitutoNacional de Psiquiatría “Ramón de la Fuente Muñiz”.
2.2. Recording and microinjection
Immediately after surgery, the animals were housed in a sound-attenuated recording chamber (101×71×64 cm) and allowed torecover for at least 15 days. During this period, the animals werehabituated to the same light-dark cycle as that maintained duringexperiments (light 20:00–8:00 h). The cats remained in the recordingchamber throughout the recovery and habituation periods, as well asduring the experiment sessions. Animals were fed ad libitum andwerekept at room temperature.
After three consecutive polygraphic recordings of 23 h (controlrecordings), the PBL area was stimulated cholinergically (carbamyl-choline chloride, Sigma). A Hamilton syringe, mounted in a manualmicrodrive, was used to deliver 4.0 µg carbachol with a volume of250 nl, over a 1 min period, after which, the cannula was left in placefor 1 min to prevent backflow. The dose of carbachol was defined onthe basis of previous dose–response studies (Baghdoyan et al., 1989).Seven days after PBL stimulation, we injected the AM of the temporallobe by delivering 250 IU of PCN dissolved in 1 µl of saline solution(0.9%) for the generation of the experimental epilepsy (baselinerecordings). For carbachol or PCN administration, a stainless steelcannula (31-gauge) 3 mm longer than the guide tube was used. Allrecordings were performed between 9:00 am and 8:00 am of thefollowing day. Pharmacological effects were assessed in all trials byscoring polygraphic recordings over a 23h period, evaluating sleep-waking states (Sterman et al., 1965; Ursin and Sterman, 1981) as wellas epileptic activity.
Our protocol consisted of carbachol injection (N=9) to induce PGOwave activity (PGO), unrelated to current state; and PCN injection(n=9) to induce experimental epilepsy (EE); carbachol followed by atopic PCN injection (n=9) (PGO+EE); and finally PCN followed bycarbachol injection (n=9) (EE+PGO). Drug trials were performed inthe order described above, with an interval of one week between eachone, and were always unilateral and ipsilateral to the initialmicroinjection site (e.g. whenever the left AMG was microinjected,the left PBL was stimulated). Contralateral injections were avoideddue to the lack of direct connections between AM and PBL. Each catwas submitted to a single series of these trials.
In the PGO+EE trial, we allowed a window of 30 min for theestablishment of PGO waves, after which the PCNwas administered forthe EE. For the EE+PGO trial, wewaited 30min for the establishment ofepileptiform activity and then the carbachol was injected. At theconclusion of the drug trials, one 23-hr control saline recording wasperformed in the PGO generator area. In order to localize effectiveinjection sites, where carbachol had induced PGO waves and wherePCN had evoked epileptic activity, we verified the location of the steelguide tubes, using the rapid procedure (Guzmán-Flores et al., 1958). All

Fig.1.Histological localization of Na–penicillin and carbachol injection sites. A. Schematic coronal representation at level A:11.5 of the injection sites into the AM. Black dots representinjection sites fromwhich Na–penicillin induced seizure activity. B. Schematic coronal section of the cat's brain stem illustrated at level P:4.0. Dots represent carbachol injection sitesfromwhich state independent PGO waves were elicited. Abbreviations of anatomical terms: 5 M, motor trigeminal nucleus; 5MT, motor trigeminal tract; 5ST, spinal trigeminal tract;BC, brachium conjunctivum; BN, amygdaloid basal nucleus; CN, amygdaloid central nucleus; FTG, gigantocellular tegmental field; FTL, lateral tegmental field; LC, nucleus of the locuscoeruleus; PBL, peribrachial area; R, raphe.
1690 I.R. Salado et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
animals included in this work had effective microinjection sites of thePBL region and AM (Fig. 1).
2.3. Scoring and analysis of behavioral states and PGO
Wakefulness, slow wave sleep 1 (SWS1), slow wave sleep 2 (SWS2),SPHOL episodes, and REM-sleep stages were visually determined fromsixty three 23 h polygraphic recordings: 27 from control recordings, and9 for eachPGO, EE, PGO+EEandEE+PGO trial (N=36). Statistical analysisof percentage, number,mean duration and latencywas calculatedwith asoftwaredesigned in our laboratory to evaluate sleepon the cat. Speciallythe latency to the first SWS1 and REM sleep onset was analyzed aftereach of the different experimental conditions. In EE, PGO + EE and EE +PGO latency was evaluated after PCN administration. These variables
Fig. 2. Representative polygraphic recording that demonstrate PGO waves triggered after 10recorded in both LGBs. Under this condition PGO waves appear in all behavioral states scoreContinuous trace (2 min) represents a transition fromW to SWS1 accompanied by PGO waveVC, visual cortex; MC, motor cortex; R-AM and L-AM right and left amygdala; R-LGB and L-LG
were averaged for each cat in each experimental condition and thenaveraged for all the cats in each trial, after which this data was plotted.Statistical analysis was performed and differences betweenmean valuesof control and experimental conditions were evaluated by one wayanalysis of variance (ANOVA) for repeated measures. The main effectswere analyzed with the pairwise comparisons tests of Bonferroni whichmet the criterion for significance of P=0.05.
Bilateral PGO waves were quantified visually by counting all LGBwaveforms characterized by an initial negative peak followed by asmaller, slower positive peak with an amplitude at least twice that ofbackgroundactivity (Brooks andGershon,1971). Analysis of PGOdensity(number of PGOwaves/min), wasmade on the first five SPHOL episodesand in the five REM sleep stages that have occurred after the beginningof the recordings. The differences between mean density values of PGO
min of carbachol injection into the PBL region. Note the enhancement of PGO activityd, while in normal conditions PGO waves appear only in SPHOL and REM sleep stages.s. Note the eye movements, persistent muscle atonia, and activated EEG. Abbreviations:B right and left lateral geniculate body; EOG, electrooculogram; EMG, electromyogram.

Fig. 3. Epileptic spikes' density, for every 30 min from Na-penicillin, during 9 h. Left panel; average values in Na–penicillin (EE, squares) (n=9) and during Carbachol+Na–penicillintrial (PGO+EE, circles) (n=9). Right panel; average values in Na–penicillin (squares) and Na–penicillin+carbachol trial (EE+PGO, circles) (n=9); same values for Na–penicillin in bothpanels (squares). Note that in experimental situations with sustained PGO activity, a progressive decline in spike frequency was observed. This reduction was accompanied by adiminishment (clinically observed) of epileptic behavioral stages IV, V and VI, whereas a lesser but still evident reduction of stages I, II and III was also observed. Data were obtainedfrom 27 trials, and values are expressed as mean±SEM; asterisks indicate the levels of statistical significance, T test; ⁎Pb0.05, ⁎Pb0.005, ⁎⁎⁎Pb0.0005, as compared to Na–penicillin.
1691I.R. Salado et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
wave bursts were statistically evaluated in each experimental situationby T tests (P=0.05).
2.4. Statistical analysis of epilepsy
Three major dependent variables were assessed: (1) generalizedseizures number, (2) spike wave density and (3) duration of theelectrophysiological epileptic seizures. The behavioral effects pro-duced by EE were also clinically observed. Spike wave density wasmeasured as the total number of spikes that occurred during a 3 minperiod (spikes/min). This data was obtained each 30 min in order toobtain 18 data samples (9 h), regardless of the sleep-waking stage. Weperformed one T test for each one of the 18 samples, in order tocompare the ones from EE with both PGO + EE and EE + PGO samples.The duration (time in seconds of each seizure) and total number ofseizures were scored, finally T tests were performed.
3. Results
3.1. Activation of the pontine PGO wave generator
When carbachol was injected into the PBL, PGO waves wereobserved in the LGB within 10 min. These PGO waves were observedover a sustained 18 to 20h period, and showed a constant pattern intheir occurrence, consisting of 4–6 wave bursts, followed by aninterval of 4–6 s. This activity occurred irrespective of the behavioralstate as illustrated in Fig. 2.
Fig. 4. Average number and duration of generalized convulsive seizures (GCSs) that have occerror bars indicate the SEM of each population. Note the smaller number of seizures during(PGO + EE). Asterisks indicate the levels of statistical significance of the difference between
3.2. Experimental epilepsy before PGO wave activation
PCN induces focal epileptiform activity, when injected into thetemporal lobe AM (250 IU/1 µl). This model of epilepsy resembles akindling-like state, provoked only with a single microinjection of PCNin freely moving cats (Fernandez-Guardiola et al., 1991), in which thesix behavioral stages appearwithin 6 h, as described byWada and Sato(Wada and Sato, 1974). All of these behavioral stages were observedafter EE, but its development was time dependent, progressive andincreasingly severe.
EE provoked high voltage biphasic spikes that appear with a shortlatency (1min post injection), which at first showed a simple pattern, butafter 7min became complex (with post-discharge) and began to spread toother structures (premotor Cortex 7 min; both LGB 10 min; visual andmotor cortex 11 min; and finally the contralateral AM 28 min). Epilepticspikeanalysis showed itshighestvaluesduring thefirst 30min,decreasingafter 60min and remaining constant during the following 8h, afterwhich,spike density began to decrease (Fig. 3). In addition to the epileptic spikes,all animals showed focal seizures (AM epileptic activity withoutpropagation), as well as EEG seizures without motor displays (with adurationbetween40 to60 s)whichweclassifiedas secondarygeneralizedseizures, and finally GCSs which lasted between 90 and 120 s. EEprogressively provoked all the behavioral stages described by Wada andSato. These convulsive andbehavioral displayshad an average timespanof9 h, yet the epileptic spike activity lasted for 24 h (decreasing their rateacross timeandfinallyoccurringonly inSWS-2).GCSs latency ranged from35 to 55 min and were observed in all animals. Status epilepticus
urred throughout the 23 h of polygraphic sleep recordings. Bars represent mean value;the PGO activated trials, and the time reduction of seizures in carbachol+Na-penicillinmeans; T test, ⁎ Pb0.01; ⁎⁎ Pb0.001 as compared with EE.

1692 I.R. Salado et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
(recurrent seizures without complete recovery between attacks orvirtually persistent seizure activity) occurred in 3 cats (one single time)in which were necessary to administer diazepam i.p. 1.2 mg/kg. The laterintervention excluded these recordings from further analysis and this trialwas performed once again one week later.
3.3. Inhibition of experimental epilepsy after PGO wave activation
In the PGO+EE trial, sustained PGO waves delayed (up to 10 min)spike installation as comparedwith EE. Also, spike propagation occurredwith a higher latency, whereas in premotor cortex is 25 min instead of7 min. LGB ipsilateral to the cholinergically activated pontine generatornever manifested epileptic activity propagation. Spike density analysis,showed top frequency in EE (20.7±SD 10.4) in the first 30 min, but inPGO + EE (4.6±SD 3.8) and EE+PGO (11.8±SD 6.2) spike frequency wassignificantly diminished during the first 30 min (T test, Pb0.0005 andPb0.05 respectively as compared to EE). From the 3rd hour ofregistration onwards, spike frequency reduced progressively, reachingsignificantly lesser values in PGO+EE (4.8±SD4.3) and inEE+ PGO (6.7±SD 3.7) in opposition to EE (15.0±SD 10.3) (Fig. 3).
During EE the total number of GCSs reached 8.0±SD 4.1, carbacholmicroinfusion, on the other hand, caused a significant decrease in thenumber of seizures to more than a half in PGO+EE (3.1±SD 5.0)(Pb0.001 and EE + PGO) (3.4±SD 2.9) (Pb0.01) (Fig. 4). Also, theaverage duration of seizures was significantly shortened, only in thePGO + EE condition (34.9±SD 11.0) (Pb0.01), as compared to EE (45.7±
Fig. 5. Representative polygraphic recordings which show electrographic differences betwee(EE+PGO, intermediate trace) and carbachol+Na-penicillin seizures (PGO+EE, bottom tracePGO activity (PGO+EE and EE+PGO). Traces obtained from one cat.
SD 17.3) Fig. 4. Notably, the latency for the first seizure in PGO+EE andEE+PGO was drastically increased (over double) to 128±SD 52.0 min(average for both conditions), instead of the 45±SD 10 min latency forthe first GCS in EE. Unlike EE condition, the seizures during PGO+EEand EE+PGO never became GCSs, therefore, we classified them assecondary generalized convulsive seizures.
Electrographic differences between EE seizures and EE+PGO andPGO + EE seizures were observed clinically. Mainly, in EE+PGO adiminishment in spike voltage, as well as in seizure duration wasobserved, also the spikes occurred without post-discharge, unlikethose occurred in EE. On the other hand, during PGO+EE trial, spikeswere depressed drastically, even though seizures still appeared butwith a significant decrease in duration (Fig. 5). While during EE, theepileptiform activity lasts for up to 24 h, whenever PGO activity wasinduced (PGO + EE and EE + PGO) the duration of the epileptic activitywas shortened, lasting for only 5 h (average).
3.4. Sleep organization after experimental epilepsy (EE)
TheANOVA showed significant differences inWpercentage betweentrials (F=6.602, df=4, P=0.031). Bonferroni's Test within subjectscontrasts showed differences for: WCON (28.7±SD 6.3) vs. WEE (43.9±13.3) (F=15.301; df=1; P=0.004). The ANOVA showed significantdifferences in REM sleep percentage between trials (F=50.874, df=4,P=0.000). Bonferroni's Testwithin subjects contrasts showeddifferencesfor: REMCON (13.04±SD 2.5) vs. REMPGO (25.09±SD 3.5) (F=56.584, df=1,
n seizures induced by Na–penicillin (EE, upper trace), Na–penicillin+Carbachol seizures). Note, a reduction of spike frequency and a decline in seizure duration during induced
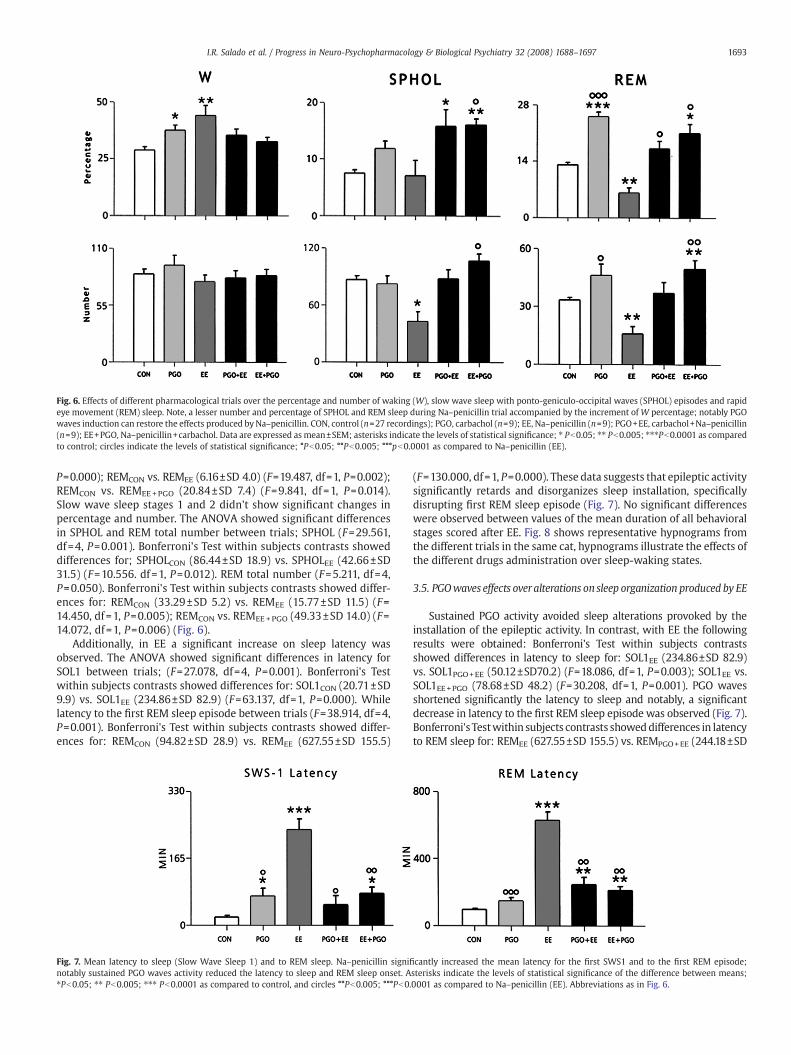
Fig. 6. Effects of different pharmacological trials over the percentage and number of waking (W), slow wave sleep with ponto-geniculo-occipital waves (SPHOL) episodes and rapideye movement (REM) sleep. Note, a lesser number and percentage of SPHOL and REM sleep during Na–penicillin trial accompanied by the increment of W percentage; notably PGOwaves induction can restore the effects produced by Na–penicillin. CON, control (n=27 recordings); PGO, carbachol (n=9); EE, Na–penicillin (n=9); PGO+EE, carbachol+Na–penicillin(n=9); EE+PGO, Na–penicillin+carbachol. Data are expressed as mean±SEM; asterisks indicate the levels of statistical significance; ⁎ Pb0.05; ⁎⁎ Pb0.005; ⁎⁎⁎Pb0.0001 as comparedto control; circles indicate the levels of statistical significance; °Pb0.05; °°Pb0.005; °°°pb0.0001 as compared to Na–penicillin (EE).
1693I.R. Salado et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
P=0.000); REMCON vs. REMEE (6.16±SD 4.0) (F=19.487, df=1, P=0.002);REMCON vs. REMEE+PGO (20.84±SD 7.4) (F=9.841, df =1, P=0.014).Slow wave sleep stages 1 and 2 didn't show significant changes inpercentage and number. The ANOVA showed significant differencesin SPHOL and REM total number between trials; SPHOL (F=29.561,df=4, P=0.001). Bonferroni's Test within subjects contrasts showeddifferences for; SPHOLCON (86.44±SD 18.9) vs. SPHOLEE (42.66±SD31.5) (F=10.556. df =1, P=0.012). REM total number (F=5.211, df=4,P=0.050). Bonferroni's Test within subjects contrasts showed differ-ences for: REMCON (33.29±SD 5.2) vs. REMEE (15.77±SD 11.5) (F=14.450, df=1, P=0.005); REMCON vs. REMEE +PGO (49.33±SD 14.0) (F=14.072, df=1, P=0.006) (Fig. 6).
Additionally, in EE a significant increase on sleep latency wasobserved. The ANOVA showed significant differences in latency forSOL1 between trials; (F=27.078, df=4, P=0.001). Bonferroni's Testwithin subjects contrasts showed differences for: SOL1CON (20.71±SD9.9) vs. SOL1EE (234.86±SD 82.9) (F=63.137, df=1, P=0.000). Whilelatency to the first REM sleep episode between trials (F=38.914, df=4,P=0.001). Bonferroni's Test within subjects contrasts showed differ-ences for: REMCON (94.82±SD 28.9) vs. REMEE (627.55±SD 155.5)
Fig. 7. Mean latency to sleep (Slow Wave Sleep 1) and to REM sleep. Na–penicillin signifinotably sustained PGO waves activity reduced the latency to sleep and REM sleep onset. A⁎Pb0.05; ⁎⁎ Pb0.005; ⁎⁎⁎ Pb0.0001 as compared to control, and circles °°Pb0.005; °°°Pb0.
(F=130.000, df=1, P=0.000). These data suggests that epileptic activitysignificantly retards and disorganizes sleep installation, specificallydisrupting first REM sleep episode (Fig. 7). No significant differenceswere observed between values of the mean duration of all behavioralstages scored after EE. Fig. 8 shows representative hypnograms fromthe different trials in the same cat, hypnograms illustrate the effects ofthe different drugs administration over sleep-waking states.
3.5. PGOwaves effects over alterations on sleep organization produced by EE
Sustained PGO activity avoided sleep alterations provoked by theinstallation of the epileptic activity. In contrast, with EE the followingresults were obtained: Bonferroni's Test within subjects contrastsshowed differences in latency to sleep for: SOL1EE (234.86±SD 82.9)vs. SOL1PGO+EE (50.12±SD70.2) (F=18.086, df=1, P=0.003); SOL1EE vs.SOL1EE+PGO (78.68±SD 48.2) (F=30.208, df=1, P=0.001). PGO wavesshortened significantly the latency to sleep and notably, a significantdecrease in latency to the first REM sleep episode was observed (Fig. 7).Bonferroni's Testwithin subjects contrasts showeddifferences in latencyto REM sleep for: REMEE (627.55±SD 155.5) vs. REMPGO+EE (244.18±SD
cantly increased the mean latency for the first SWS1 and to the first REM episode;sterisks indicate the levels of statistical significance of the difference between means;0001 as compared to Na–penicillin (EE). Abbreviations as in Fig. 6.
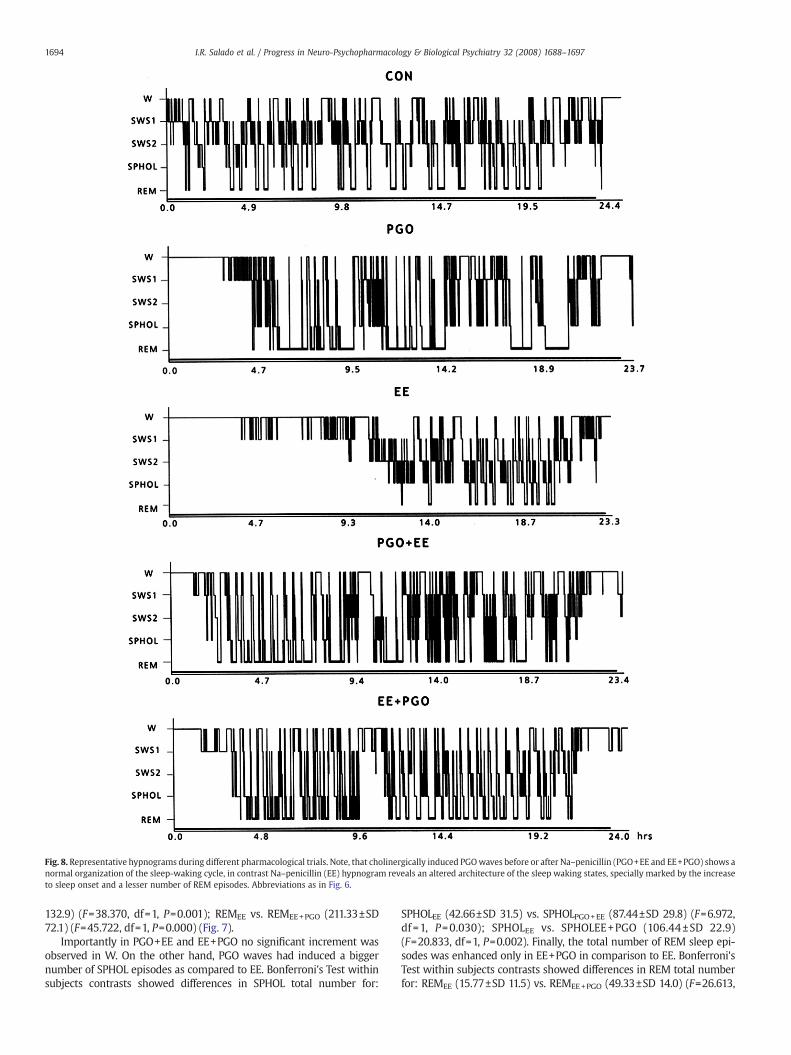
Fig. 8. Representative hypnograms during different pharmacological trials. Note, that cholinergically induced PGOwaves before or after Na–penicillin (PGO+EE and EE+PGO) shows anormal organization of the sleep-waking cycle, in contrast Na–penicillin (EE) hypnogram reveals an altered architecture of the sleep waking states, specially marked by the increaseto sleep onset and a lesser number of REM episodes. Abbreviations as in Fig. 6.
1694 I.R. Salado et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
132.9) (F=38.370, df=1, P=0.001); REMEE vs. REMEE+PGO (211.33±SD72.1) (F=45.722, df=1, P=0.000) (Fig. 7).
Importantly in PGO+EE and EE+PGO no significant increment wasobserved in W. On the other hand, PGO waves had induced a biggernumber of SPHOL episodes as compared to EE. Bonferroni's Test withinsubjects contrasts showed differences in SPHOL total number for:
SPHOLEE (42.66±SD 31.5) vs. SPHOLPGO+EE (87.44±SD 29.8) (F=6.972,df =1, P=0.030); SPHOLEE vs. SPHOLEE+PGO (106.44±SD 22.9)(F=20.833, df=1, P=0.002). Finally, the total number of REM sleep epi-sodes was enhanced only in EE+PGO in comparison to EE. Bonferroni'sTest within subjects contrasts showed differences in REM total numberfor: REMEE (15.77±SD 11.5) vs. REMEE+PGO (49.33±SD 14.0) (F=26.613,

Fig. 9. Effects of the different pharmacological trials over PGO waves density (during SPHOL episodes and during rapid eye movement REM sleep episodes). Note, an importantreduction of PGO density during Na–penicillin (REM sleep episodes), and the enhancement produced by carbachol injection (SPHOL and REM episodes). Carbachol+Na–penicillin(PGO+EE) and Na–penicillin+carbachol (EE+PGO) trials, showed an unexpected enhancement of PGO waves density in both SPHOL and REM stages. Data are expressed as mean±SEM; asterisks indicate the levels of statistical significance; ⁎ pb0.05; ⁎⁎ pb0.005; ⁎⁎⁎pb0.0001 as compared to control; and circles °pb0.05; as compared to EE.
1695I.R. Salado et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
df=1, P=0.001). The effects of PGO + EE and EE + PGO over SPHOL andREM percentages are presented in Fig. 6.
After the analysis of PGO waves density during REM sleep andSPHOL episodes, we found that density showed important changesthroughout different trials; we performed T tests between trials andPGO condition produced a long-term enhancement of PGO wavedensity during SPHOL (86.7±SD 13.3) (Pb0.0001) and during REM(94.5±SD 19.6) (Pb0.005), as compared with control (SPHOL 25.4±SD10.8; REM 70.1±SD 18.0). Notably, in EE, PGO density diminishedduring REM episodes (48.4±SD 8.1) Pb0.0001 as compared withcontrol (70.1±SD 17.8). Major density changes were observed as wellin PGO + EE (86.3±SD 15.6) and in EE + PGO (86.9±SD 8.4), as com-pared with EE density (48.4±SD 8.2) and to control condition. Also,PGO density during SPHOL episodes significantly increased in PGO+EE(57.0±SD 24.6) and in EE+PGO (66.6±SD 14.0) as compared withcontrol density (Pb0.0001). Finally, a significant augment wasobserved as well in both PGO+EE and EE+PGO trials in comparisonto EE (24.8±SD 6.6) (Pb0.0001). The data are presented in Fig. 9.
4. Discussion
The results obtained in the present study show that PGO wavesunrelated to REM sleep exert an inhibitory influence over seizuresinduced by EE. Due to this inhibition, PGO activity restores and evenpromotes sleep onset and maintenance antagonizing sleep alterationsinduced by epilepsy. Moreover, these results also confirm our previousfindings in which the inhibitory influence of REM sleep over kindlingdevelopment was demonstrated (Calvo, 1991). PGO potentials,responsible for the rapid eye movements of REM sleep, whichpropagate from the pons to several forebrain regions, have beencorrelated with increased neuronal firing throughout the brain andwith inhibited transmission in the relay of sensory pathways, as aconsequence of neuronal depolarization (Jouvet, 1972; Callaway et al.,1987). Therefore, PGO-related phasic potentials resulting from PGOactivity propagation (Calvo and Fernández-Guardiola, 1984), mayaffect the neuronal excitability of the amygdala and this change maythus prevent the hyperexcitation of the same structure, as occurring inthe epileptiform activity.
The present study corroborates as well, that carbachol microinfu-sion in the PBL region, induces PGO wave activity, irrespective of thesleep stage (Datta et al., 1991, 1992), and also establishes precisely thatthis phenomenon lasts from 18 to 20 h.
PGOs have an inhibitory effect on seizure occurrence. This becomesevident by the fact that during the first 4 h of PGO potentials, that wereboth sustained andunrelated to REMsleep,we observed epileptic spikeswith lower voltage, without propagation to other brain structures and
with a significantly lesser frequency, when compared to those whichoccurred in the absence of PGOs. Notably, even though spikes werepresent, only the behavioral stages I and IIwere reached,whereas stagesIII, IV, V were abolished. Furthermore, although seizures indeedoccurred, these were only restricted to an electrographic discharge(EEG seizure), never reaching themotor displays of Stage VI (GCSs). Thisabolishment may be explained by the typical muscular atonia whichaccompanies carbachol application in the PBL region. Nevertheless, sucheffect lasts for only 20–40 min post stimulation, after this period suchatonia cannot inhibit any of the motor components of seizures. Thus,PGO waves might participate as an electrophysiological factor thatdiminishes the motor displays of seizures through the isolation of theepileptic activity solely to the primary focus. Notably, during this period,REM sleep episodes have not occurred yet, as there is a slight increase inlatency for this stage. Therefore, the inhibitory effect over seizures anditsmotor components in the first hours can only be due to the sustainedoccurrence of PGO activity.
Another result also upholding the premise of an inhibitory effect ofPGO potentials over seizures is the latency to the first seizure. In thecase of EE, the first GCS occurred between 35 and 55 min aftermicroinjection. In contrast, whenever PGO waves were present, thefirst seizure was significantly delayed, with twice the time to occur(averaging 128 min). Also, both the average number and length ofseizures were significantly shortened with PGO activity compared tothose during EE. The increased time lapse in seizure latency and thelack of any behavioral components inherent to seizures, can probablybe explained by the delay in the spike propagation to other brainregions and by the decline in amplitude and frequency duringsustained PGO activity. Notably, such effect was obtained duringPGO waves, even after a second (EE+PGO) and third (PGO+EE) Na–penicillin application respectively; in both trials a kindling-like effectby repeated chemical stimulation could be expected. Nevertheless,such phenomenon did not occur, suggesting that PGO waves couldalso be avoiding a potential kindling-like effect.
On the other hand, it is important to consider that PGO waves,unrelated to REM sleep, are brief electrophysiological phenomena(150 to 200 ms) which may provoke an intermittent inhibition at thesite of the origin of the epileptic activity, and that sustained epilepticabolition during the REM stage not only involves PGO potentials, butalso includes the participation of other electrophysiological andneurochemical components, such as atonia and EEG desynchroniza-tion, which are inherent to this sleep phase. Immediate effects ofcarbachol in the PBL region may be explained in terms of muscarinicreceptor activation. The long-term effects suggests that carbachol mayalso induce a long-lasting change in PBL neuronal responsiveness viasecond messengers that may account for an increased PBL cholinergic

1696 I.R. Salado et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
output reflected in PGO activity enhancement. It has been frequentlysuggested that the extinction of a convulsive seizure may be due to anactive depressor mechanism, in this case PGO sustained activity,which might be playing a critical role in the protection of theamygdala from epileptic spikes, acting as a “gating”mechanism in theAM which in turn promotes PGO waves maintenance with itsconsequent REM sleep installation.
Nevertheless it is important to notice, that while spontaneous REMsleep episodes, abolish epileptic activity solely during the phase time,during state independent PGO activity, epileptic spikes and seizuresinhibition is sustained and long-lasted. This sustained inhibition mightbe explained by PGO potentials that propagate to the AM, therefore,impeding the synchronization of the same structure. We suggest thatalthough the AM is already stimulated by Na–penicillin, it may beenhancing PGO activity through the anatomical pathway (i.e. amygda-lofugal bundle) with the pontine regions that generate PGO waves andREM sleep, specifically the PBL region. In this context, the AM may befacilitating sleep mechanisms involved in seizure inhibition, thusdepressing seizure mechanisms and acting as a protective forebraincomponent rather than as an epileptic promoter site. Simple andsustained PGOactivityoriginated in thePBLmaybe restricting abnormalelectric activity to the AM. This restriction avoids the thalamicstimulation from the AM as well. Also, PBLmay be suppressing seizuresthrough thedirect inhibition of thalamicneuronalpopulationsbymeansof the anatomical pathways between the PBL and the thalamus, namely,PPT and LDT nuclei, structures involved in the EEG desynchronizationduring REM sleep. Finally, inhibition of themotor component of seizuresmay be due to the deactivation of cortical pathways from the thalamusas well. Thus, the suppression of epileptic activity during PGO wavesdepends directly on PBL activationwith the consequent thalamocorticalphasic inhibition.
Another major feature of the present work is the restoring role ofsustained PGOwaves over both architecture and temporal organizationof the sleep waking cycle alterations following EE, indicating that thesepotentials have a facilitating role on the installation and maintenancemechanisms of REM sleep, even in the presence of epileptic activity.
Under the EE condition a significant increase in W percentage wasobserved, also sleep latency (SWS1) was robustly augmented. On theother hand, both PGO+EE and EE+PGO recordings showed asignificant decrease in W percentage, and therefore a reduction insleep latency, evidenced by a restoration over the whole sleeporganization. Also, a significant increase in REM sleep percentagewas observed in both PGO+EE and EE+PGO compared with EE.
Theoutcomesof thepresent study ledus to propose that PGOactivityisworking through a two-waypath: in a short term, it abolishes epilepticactivity by decreasing spike density, frequency and propagation, and,once thismonophasic activity is controlled, PGOactivitynormalizes (in along term) the whole sleep waking cycle by activating the neuralmechanisms responsible for REM sleep. However the S–W cycle duringPGO+EE and EE+PGO resemblesmore to carbachol trials, than to controlones, since PGO promotes REM sleep generation. Thus, it may bepossible that PGO activity enhances REM sleep onset (after Na–penicillin) as a preventing mechanism against seizure.
Altogether, these results support the inhibitor role of PGOwaves overseizures, and corroborate the notion that the physiological mechanismsof REM sleep are similar to those of epileptic inhibition. This may occurbymeans of desynchronization of anterior structures, in such away thatPGO waves are impeding the hypersynchronized firing activity of AMneurons, which is necessary for epileptic propagation and therefore, forthe occurrence of GCSs. The PCN epileptogenic effect is due to a partialblocking of the GABA-mediated inhibition (Loeb et al., 1985; Chatt andEbersole, 1988). However the paroxistic activity can be modified bystimulating regions distant from the primary focus by means of severalprocedures (Gasteiger et al., 1985; La Grutta and Sabatino, 1988;McLachlan and Bihari, 1990; Fernández-Guardiola et al., 1999; Kumarand Raju 2001; Magdaleno-Madrigal et al., 2002). Our protocol allowed
us todemonstrate that the epileptogenic effect canbe revertedbymeansof cholinergic stimulation of the PBL region, since the reciprocalconnections between PBL and AM may be activating each other, inorder to facilitate the generation and maintenance of the phasicphenomena of REM sleep (PGOs).
These findings provide evidence for future studies designed toidentify the neuronal mechanisms responsible for the interactionsbetween sleep and epilepsy. Specifically we will analyze whetherepileptic inhibition can also be obtained throughout the whole REMsleep stage induction, by topic application of carbachol in the nucleuspontis oralis (NPO). Also, through the administration of GABAergicagonists into the NPO, and muscarinic receptor antagonists in PBLregion, we will assess whether REM and PGO depressed mechanismsexert a promoting and enhancing effect over epilepsy.
Acknowledgements
The authors thank Mr. Carlos Jímenez, Mr. Carlos Camacho andBiologist Pilar Chiappa for their excellent technical assistance. We arealso grateful to PhD. Isabel Pérez Monfort for revising the Englishversion of this manuscript. This work was supported by Project 3340from the Instituto Nacional de Psiquiatría “Ramónde la FuenteMuñiz”.
References
Amaral DG, Price JL, Pitkänen A, Carmichael ST. Anatomical organization of the primateamygdaloid complex. In: Aggleton JP, editor. The Amygdala: Neurobiological Aspectsof Emotion,Memory, andMental Dysfunction. New York:Willey-Liss; 1992. p.1–66.
Aserinsky E, Kleitman N. Regularly occurring periods of eye motility and concomitantphenomena during sleep. Science 1953;118:273–4.
Baghdoyan HA, Lydic R, Callaway CW, Hobson JA. The carbachol induced enhancementof desynchronized sleep signs is dose dependent and antagonized by centrallyadministered atropine. Neuropsychopharmacology 1989;2:67–9.
Brooks DC, Gershon MD. Eye movement potentials in the oculomotor and visual systemof the cat: a comparison of reserpine induced waves with those present duringwakefulness and rapid eye movement sleep. Brain Res. 1971;27:223–39.
Cadilhac J. Complex partial seizures and REM sleep. In: Sterman MB, Shouse MN,Passouant P, editors. Sleep and Epilepsy. NewYork: Academic Press; 1982. p. 315–24.
Callaway CW, Lydic R, Baghdoyan HA, Hobson JA. Pontogeniculooccipital waves:spontaneous visual system activity during rapid eye movement sleep. Cell. Mol.Neurobiol. 1987;7:105–49.
Calvo JM. Amygdaloid kindling during wakefulness and paradoxical sleep in the cat. 1.Inhibitory influence of paradoxical sleep on kindling development. Epilepsy Res.1991;9:113–20.
Calvo JM, Fernandez-Guardiola A. Phasic activity of basolateral amygdala, cingulategyrus, and hippocampus during REM sleep in the cat. Sleep 1984;7:202.
Calvo JM, Badillo S, Morales-Ramirez M, Palacios-Salas P. The role of the temporal lobeamygdala in ponto-geniculo-occipital activity and sleep organization in cats. BrainRes. 1987;403:22–30.
Calvo JM, Datta S, Quattrochi J, Hobson JA. Cholinergic microestimulation of theperibraquial nucleus in the cat. II. Delayed and prolonged increases in REM sleep.Arch. Ital. Biol. 1992;130:285–301.
Cohen H, Thomas J, Dement WC. Sleep stages, REM deprivation and electroconvulsivethreshold in the cat. Brain Res. 1970;19:317–21.
Chatt AB, Ebersole JS. Comparision between strychnine and penicillin epileptogenesissuggest that propagating epileptiform abnormalities require the potentiation ofthalamocortical circuitry in neocortical layer 4. Exp. Neurol. 1988;100:365–80.
Datta S. Activation of phasic pontine-wave generation: a mechanism for sleep-dependent memory processing. Sleep Biol. Rhythms 2006;4:16–26.
Datta S, Hobson JA. Neuronal activity in the caudolateral peribrachial pons: relationshipto PGO waves and rapid eye movements. J. Neurophysiol. 1994;71(1):95–109.
Datta S, Calvo JM, Quattrochi J, Hobson JA. Long-term enhancement of REM sleepfollowing cholinergic stimulation. NeuroReport 1991;2:619–22.
Datta S, Calvo JM, Quattrochi J, Hobson JA. Cholinergic microstimulation of theperibrachial nucleus in the cat. 1. Immediate and prolonged increase in ponto-geniculo-occipital waves. Arch. Ital. Biol. 1992;130:263–84.
Datta S, Maclean RR. Neurobiological mechanisms for the regulation of mammaliansleep-wake behavior: reinterpretation of historical evidence and inclusion ofcontemporary cellular and molecular evidence. Neurosci Biobehav Rev 2007;30(5):775–824.
Delange M, Caston P, Cadilhac J, Passouant P. Etude du sommeil de nuit au coursd'épilepsies centrencéphaliques et temporales. Rev. Neurol. (Paris) 1962;106:106–13.
Fernandez-Guardiola A, Ayala F. Red nucleus fast activity and signs of paradoxical sleepappearing during the extinction of experimental seizures. Electroencephalogr. Clin.Neurophysiol 1971;30:547–55.
Fernandez-Guardiola A, Martinez A, Gutierrez R, Fernandez-Mas R. Amygdalinepenicillin focus replicates and modulates electrical amygdaloid Kindling in thecat. Proc. West. Pharmacol. Soc. 1991;34:219–22.

1697I.R. Salado et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 1688–1697
Fernandez-Guardiola A, Martinez A, Valdéz-Cruz A, Magdaleno-Madrigal VM, MartínezD, Fernández-Mas R. Vagus nerve prolonged stimulation in cats: effects onepileptogenesis (Amygdala Electrical Kindling): behavioral and electrographicchanges. Epilepsia 1999;40(7):822–9.
Gasteiger LE, Albowitz B, Barken MF. Interictal after discharge in focal penicillinepilepsy: block by thalamic cooling. Exp. Neurol. 1985;88:349–59.
Gloor P, Quesney LF, Zumstein H. Pathophysiology of generalized epilepsy in the cat: therole of cortical and subcortical structures. II Topical application of penicillin to thecerebral cortex and to subcortical structures. Electroencephalogr. Clin. Neurophy-siol. 1977;43:79–94.
Guzmán-Flores C, Alcaráz M, Fernandez-Guardiola A. Rapid procedure to localizeelectrodes in experimental neurophysiology. Bol. Estud. Med. Biol. (México)1958;16:29–31.
Jouvet M. The role of monoamines and acetylcholine containing neurons in theregulation of the sleep-waking cycle. In: Jouvet M, Moruzzi G, editors. Neuropy-siology and Neurochemistry of Sleep and Wakefulness. Reviews of Physiology.Heidelberg: New York; 1972. p. 166–307.
Kumar P, Raju TR. Seizure susceptibility decreases with enhancement of rapid eyemovement sleep. Brain Res. 2001;922:299–304.
La Grutta V, Sabatino M. Focal hippocampal epilepsy: effect of caudate stimulation. Exp.Neurol. 1988;99:38–49.
Loeb C, Bo GP, Scotto A, Benassi B, Besio G, Minardi P, Faverio A. GABA and phospholipidsin penicillin-induced seizures. Exp. Neurol. 1985;9:278–80.
Magdaleno-Madrigal VM, Valdéz-Cruz A, Martinez-Vargas D, Martinez A, Almazán S,Fernández-Mas R, Fernandez-Guardiola A. Effect of electrical stimulation of the
nucleus of the solitary tract on the development of electrical amygdaloid kindling inthe cat. Epilepsia 2002;43(9):964–9.
McLachlan RS, Bihari F. Secondary generalization of seizures from a cortical penicillinfocus following stimulation of the basal forebrain. Exp. Neurol. 1990;109:237–42.
Montplaisir J, Lavardiere M, Saint Hilaire JM. Sleep and focal epilepsy: contribution ofdepth recording. In: Sterman MB, Shouse MN, Passouant P, editors. Sleep andEpilepsy. New York: Academic Press; 1982. p. 301–14.
Nomura S, Mizuno N, Itoh K, Matsuda K, Sugimoto T, Nakamura Y. Localization ofparabrachial nucleus neurons projecting to the thalamus or the amygdala in the catusing horseradish peroxidase. Exp. Neurol. 1979;64(2):375–85.
Snider RS, Niemer WT, editors. A Stereotaxic Atlas of the Cat Brain. Chicago Il: ChicagoPress; 1961.
StermanMB, Knauss T, Lehman D, Clemente CD. Circadian sleep and waking patterns inthe laboratory cat. Electroencephalogr. Clin. Neurophysiol. 1965;19:509–17.
Thomas J, Benoit O. Individualisation d'un sommeil a ondes lentes et activité phasique.Brain Res. 1967;5:221–35.
Ursin R, Sterman MB. A manual for standarized scoring of sleep and waking in the adultcat. Brain Information Service / Brain Research Institute. Los Angeles: University ofCalifornia; 1981.
Vivaldi E, McCarley RW, Hobson JA. Evocation of desynchronized sleep signs bychemical microestimulation of the pontine brainstem. In: Hobson JA, Brazier MAB,editors. The Reticular Formation Revisited. New York: Raven Press; 1980. p. 513–29.
Wada JA, Sato M. Generalized convulsive seizures induced by daily electricalstimulation of the amygdala. Correlative electrographic and behavioral features.Neurology 1974;24:565–74.




















