
Influence of extracellular osmolarity and mechanical stimulation on gene expression of...
10
Influence of Extracellular Osmolarity and Mechanical Stimulation on Gene Expression of Intervertebral Disc Cells K. Wuertz, 1 J.P.G. Urban, 2 J. Klasen, 3 A. Ignatius, 1 H.-J. Wilke, 1 L. Claes, 1 C. Neidlinger-Wilke 1 1 Institute of Orthopaedic Research and Biomechanics, University of Ulm, Helmholtzstrasse 14, 89081 Ulm, Germany 2 Physiology Laboratory, University of Oxford, Oxford, England 3 Neurosurgical Department, University of Ulm/Guenzburg, Ulm, Germany Received 25 July 2006; accepted 30 March 2007 Published online 13 June 2007 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/jor.20436 ABSTRACT: Intervertebral discs (IVD) have a higher extracellular osmolarity than most other tissues; moreover their osmolarity changes by around 25% during each diurnal cycle. In this study, changes in aggrecan, collagen I and collagen II expression of IVD cells were examined after exposure to osmotic environment alterations or mechanical stimulation under different osmotic conditions. Human and bovine IVD cells seeded in three-dimensional (3D) collagen type I matrices were cultured under hypo-osmotic (300 mOsm), iso-osmotic (400 mOsm), or hyperosmotic (500 mOsm) conditions. Osmolarity-induced changes in gene expression of IVD cells were measured after 5 days. Load- induced changes in gene expression under the different osmotic conditions were measured after application of hydrostatic pressure (0.25 MPa, 0.1 Hz, 30 min) or cyclic strain (4%, 1 Hz, 24 h). The results showed that IVD cells respond strongly to changes in the osmotic environment by altering mRNA expression. Human cells cultured over 5 days increased expression of aggrecan and collagen II in both nucleus and annulus cells under increasing osmolarity. In contrast, collagen I expression was inhibited at high osmolarity in both cell types. Mechanically induced alterations in gene expression appear to have only modest effects on matrix protein expression, but the same stimulus partly resulted in an inhibition or stimulation of gene expression, depending on the osmotic conditions. This study showed that the osmotic environment does not only have an appreciable effect on gene expression but also affects responses to mechanical stimuli. This suggests that the osmotic conditions cannot be ignored when examining physiological and pathological behavior of IVD cells. ß 2007 Orthopaedic Research Society. Published by Wiley Periodicals, Inc. J Orthop Res 25:1513– 1522, 2007 Keywords: intervertebral disc cells; osmolarity; mechanical stimulation; gene expression INTRODUCTION The extracellular matrix of the intervertebral disc (IVD), like that of other load-bearing cartilage, is rich in aggrecan and hence in negatively charged glycosaminoglycans (GAGs). These regulate the ionic composition of the IVDs extracellular matrix through the Gibbs-Donnan equilibrium condi- tions. The IVDs matrix has a high concentration of positive ions such as Na þ and K þ compared to serum and indeed most tissues 1 with ion concen- trations varying directly with fixed charge density; Na þ concentrations are thus greatest in the nucleus pulposus (NP) and lowest in the outer annulus fibrosus (AF). The mobile ions play a vital role in the biomechanical functioning of the disc as they impart a high extracellular osmolarity to the tissue 2 and enable it to retain fluid even under high external loads. 3 As the IVD is an osmotic system, its hydration is not constant but varies with applied load. In vivo loading is applied mainly by body weight and muscle activity. Loads are lowest at night during rest (typically 0.1–0.2 MPa). 4 During the day’s activities, intradiscal pressures can rise to 3 MPa. 4 As a result, fluid is expressed from the IVD during day (up to 25% of fluid loss) and re-imbibed at night when the loads are reduced. 3 Consequently, as a result of this diurnal change of fluid, the extra- cellular osmolarity changes proportionately. Cells of the IVD thus have to adapt to these changes in extracellular osmolarity. Although surprisingly little is known about the adaptation process in the IVD, changes in extracellular osmolarity as in most cell types 5 regulate cell volumes of NP, AF, and notochordal disc cells. 6,7 Changes in the osmotic conditions result in changes in [Ca 2þ ] in the IVD 7 JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007 1513 Correspondence to: C. Neidlinger-Wilke (Telephone: 49- (0)731-5002-3482; Fax: 49-(0)731-5002-3498; E-mail: [email protected]) ß 2007 Orthopaedic Research Society. Published by Wiley Periodicals, Inc.
Transcript of Influence of extracellular osmolarity and mechanical stimulation on gene expression of...

Influence of Extracellular Osmolarity and MechanicalStimulation on Gene Expression of Intervertebral Disc Cells
K. Wuertz,1 J.P.G. Urban,2 J. Klasen,3 A. Ignatius,1 H.-J. Wilke,1 L. Claes,1 C. Neidlinger-Wilke1
1Institute of Orthopaedic Research and Biomechanics, University of Ulm, Helmholtzstrasse 14, 89081 Ulm, Germany
2Physiology Laboratory, University of Oxford, Oxford, England
3Neurosurgical Department, University of Ulm/Guenzburg, Ulm, Germany
Received 25 July 2006; accepted 30 March 2007
Published online 13 June 2007 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/jor.20436
ABSTRACT: Intervertebral discs (IVD) have a higher extracellular osmolarity than most othertissues; moreover their osmolarity changes by around 25% during each diurnal cycle. In this study,changes in aggrecan, collagen I and collagen II expression of IVD cells were examined after exposureto osmotic environment alterations or mechanical stimulation under different osmotic conditions.Human and bovine IVD cells seeded in three-dimensional (3D) collagen type I matrices were culturedunder hypo-osmotic (300 mOsm), iso-osmotic (400 mOsm), or hyperosmotic (500 mOsm) conditions.Osmolarity-induced changes in gene expression of IVD cells were measured after 5 days. Load-induced changes in gene expression under the different osmotic conditions were measured afterapplication of hydrostatic pressure (0.25 MPa, 0.1 Hz, 30 min) or cyclic strain (4%, 1 Hz, 24 h). Theresults showed that IVD cells respond strongly to changes in the osmotic environment by alteringmRNA expression. Human cells cultured over 5 days increased expression of aggrecan and collagen IIin both nucleus and annulus cells under increasing osmolarity. In contrast, collagen I expressionwas inhibited at high osmolarity in both cell types. Mechanically induced alterations in geneexpression appear to have only modest effects on matrix protein expression, but the same stimuluspartly resulted in an inhibition or stimulation of gene expression, depending on the osmoticconditions. This study showed that the osmotic environment does not only have an appreciable effecton gene expression but also affects responses to mechanical stimuli. This suggests that the osmoticconditions cannot be ignored when examining physiological and pathological behavior of IVD cells.
� 2007 Orthopaedic Research Society. Published by Wiley Periodicals, Inc. J Orthop Res 25:1513–
1522, 2007
Keywords: intervertebral disc cells; osmolarity; mechanical stimulation; geneexpression
INTRODUCTION
The extracellular matrix of the intervertebral disc(IVD), like that of other load-bearing cartilage, isrich in aggrecan and hence in negatively chargedglycosaminoglycans (GAGs). These regulate theionic composition of the IVDs extracellular matrixthrough the Gibbs-Donnan equilibrium condi-tions. The IVDs matrix has a high concentrationof positive ions such as Naþ and Kþ compared toserum and indeed most tissues1 with ion concen-trations varying directly with fixed charge density;Naþ concentrations are thus greatest in thenucleus pulposus (NP) and lowest in the outerannulus fibrosus (AF). The mobile ions play a vitalrole in the biomechanical functioning of the disc as
they impart a high extracellular osmolarity to thetissue2 and enable it to retain fluid even underhigh external loads.3
As the IVD is an osmotic system, its hydration isnot constant but varies with applied load. In vivoloading is applied mainly by body weight andmuscle activity. Loads are lowest at night duringrest (typically 0.1–0.2 MPa).4 During the day’sactivities, intradiscal pressures can rise to 3 MPa.4
As a result, fluid is expressed from the IVD duringday (up to 25% of fluid loss) and re-imbibed at nightwhen the loads are reduced.3 Consequently, as aresult of this diurnal change of fluid, the extra-cellular osmolarity changes proportionately. Cellsof the IVD thus have to adapt to these changes inextracellular osmolarity. Although surprisinglylittle is known about the adaptation process in theIVD, changes in extracellular osmolarity as in mostcell types5 regulate cell volumes of NP, AF, andnotochordal disc cells.6,7 Changes in the osmoticconditions result in changes in [Ca2þ] in the IVD7
JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007 1513
Correspondence to: C. Neidlinger-Wilke (Telephone: 49-(0)731-5002-3482; Fax: 49-(0)731-5002-3498;E-mail: [email protected])
� 2007 Orthopaedic Research Society. Published by Wiley Periodicals,Inc.

and in addition, volume changes correlate withrapid changes in sulphated GAG production.2
Apart from the effects of applied load on fluidexpression, various loading regimes in vivo subjectthe disc routinely to flexion, bending, or torsionresulting in complex load-induced signals to thecells. Changes in hydrostatic pressure,4,8,9 cyclicstrain,10,11 or shear stress12 have all been found toinfluence cellular behavior directly. Althoughmechanical stress in vivo in the IVD is associatedwith changes in hydration and therefore also withchanges in osmolarity, most in vitro studies onmechanobiology in the IVD (especially when usingcell culture models) have ignored the role of theosmotic environment. Therefore, little is knownabout the interactions between the extracellularosmotic and mechanical environment on IVD cellbehavior. In a previous study,13 we were able toshow that human NP and AF cells cultured in athree-dimensional (3D) collagen type I matrixresponded to the application of hydrostatic pres-sure and cyclic strain by altering mRNA expressionof several genes known to be relevant in IVD matrixturnover; however, this former study was carriedout under iso-osmotic conditions. Thus, this studyaddressed the following questions:
How is gene expression of IVD cells related to theextracellular osmolarity in 3D culture?
How is gene expression of IVD cells influenced bymechanical stimulation (hydrostatic pressure, cyc-lic strain) under different osmotic conditions?
MATERIALS AND METHODS
Isolation and Culture of Bovine Cells
Caudal IVDs were dissected under sterile conditionsfrom the tails of five 18- to 24-month old bovines within1 h of slaughter. For each animal nucleus pulposus (NP),transition zone (TZ), and annulus fibrosus (AF) weredissected from the upper eight levels and pooled by zone.The tissue was chopped finely and digested over 4 h inDulbeccos’s modified Eagle’s medium (DMEM Hepes;Gibco, Karlsruhe, Germany) with collagenase-1 (0.8 mg/mlNP, 2 mg/mlTZ, 3 mg/mlAF), DNAse (1.3 U/mlNP/TZ/AF)and protease type 1 (0.0 mg/mlNP, 0.6 mg/mlTZ/AF)(all Sigma Chemical Co, Deisenhofen, Germany). Thedigestion medium was supplemented with 500 U/mlpenicillin (Biochrom, Berlin, Germany), 500 mg/mlstreptomycin sulphate (Biochrom, Berlin, Germany),and 12.5 mg/ml fungizone (Gibco, Karlsruhe, Germany).Culture medium was adjusted to iso-osmotic conditions(400 mOsm6) by salt solution addition: 5M NaCl (Riedel-de-Haen, Seelze, Germany) and 0.4M KCl (Merck,Darmstadt, Germany) at 1.5% v/v. Cells were subcultur-ed in monolayer up to passage 1 or 2, using iso-osmoticDMEM Hepes with 100 U/ml penicillin, 100 mg/mlstreptomycin sulphate, 1.25 mg/ml fungizone, 5% v/v
fetal bovine serum (Biochrom), 1% v/v nonessential aminoacids (Biochrom) and 1% v/v L-glutamine (Biochrom).13,14
Isolation and Culture of Human Cells
Human IVD cells were isolated from biopsy samples(n¼ 9) of patients undergoing surgery for disc hernia-tion (Table 1). Tissue was separated into NP and AF bythe surgeon and digested within 2 h of excision asdescribed above except that enzyme concentrationsdiffered: collagenase 1 (0.8 mg/mlNP, 2 mg/mlAF), DNAse(2.6 units/mlNP/AF), and protease type 1 (0.0 mg/mlNP,0.6 mg/mlAF). Not all donors expressed collagen II afterexpansion in monolayer (up to passage 2 or 3), resultingin a smaller group size for collagen II.
Cell Culture (General)
Cell culture experiments with or without mechanicalstimulation are described in the following paragraphsand diagrammed in Figure 1.
Cell Culture under Controlled Osmolarity(without Mechanical Stimulation)
Human NP and AF cells were cultured under differentosmotic conditions for 5 days. Cells were seeded incollagen type I gels at a final concentration of 3 mg/ml�1
and an in vivo like cell density of 0.3� 106 cells/ml(Ars Arthro AG, Esslingen, Germany) as previouslydescribed.13 Gels of 3 ml were poured into the respectiveculture dishes, allowed to polymerize and covered with2 ml of serum-free culture medium. Osmolarity of theculture medium was adjusted to 300, 400, or 500 mOsmin order to simulate the physiological osmotic range thatoccurs in vivo. A mean value of 400 mOsm was chosen torepresent the average iso-osmotic in vivo situation;300 mOsm represents a lower and 500 mOsm a higherphysiological range After 5 days, gels were dissolved(collagenase type I, 6000 U/ml, Sigma) and cells werecentrifugated and used for RNA preparation.
Application of Hydrostatic Pressure underControlled Osmolarity
For the application of hydrostatic pressure, human(n¼ 4) and bovine (n¼ 6) NP cells were seeded in the3D collagen type I gels and covered with 2 ml serum-freeculture medium at the defined osmolarities describedabove. After 3 days of preculture, hydrostatic pressurewas applied as described previously.13 Briefly, cell-seeded gels were stimulated inside a costum-madepressure chamber by air compression. Intermittenthydrostatic pressure was applied at a magnitude of0.25 MPa during a stimulation period of 30 min with afrequency of 0.1 Hz (simulating a moderate in vivoloading case). Control dishes, cultured at the sameosmolarities, were placed in an identical apparatusbut no pressure was applied. Following mechanical
1514 WUERTZ ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007 DOI 10.1002/jor

Table 1. Patient Details
No. Age SexDiscLevel
SurgeryDue to
DiscRegion
Data Used in Figure
2A 2B 3A 4A 4C
1 49 F L4/5 Herniation NP* x x2 37 M L4/5 Herniation NP x x2 37 M L4/5 Herniation AF* x x3 71 M L4/5 Herniation NP x x3 71 M L4/5 Herniation AF x x4 37 F L5/S1 Herniation NP* x x x4 37 F L5/S1 Herniation AF* x x5 42 M L# Herniation NP x x6 44 M L4/5 Herniation NP x x x7 51 M L4/5 Herniation NP x x x8 49 F L5/S1 Herniation AF x x9 29 M L1/2 Herniation NP x x
Abbreviations: M, male; F, female; NP, nucleus pulposus; AF, annulus fibrosus.*Collagen II expression detectable.#Lumbar disc, level unknown.
Figure 1. Flowchart of all cell culture experiments (with or without mechanical stimulation).
DISC CELLS: OSMOLARITY AND MECHANICAL LOAD 1515
DOI 10.1002/jor JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007

stimulation, both controls and pressurized samples werecultured for further 24 h in an incubator beforedissolving gels (as described above) and examiningcellular responses. Loading time and analysis time werechosen accordingly to results in our pre-experimentsand data from the literature.15
Application of Cyclic Strain underControlled Osmolarity
Human (n¼ 4) and bovine (n¼ 4) AF cells were used forcyclic strain experiments. Additionally, human NP cells(n¼ 4) were stretched in order to check whether cellsfrom the central region of the IVD also respond to cyclicstrain. Cells were seeded in 3D collagen gels at a densityof 0.3� 106 cells/ml and poured into rectangular costum-made silicon dishes as previously described.13 After2 days of pre-culture, cyclic strain was applied at 4%(equivalent to 40,000 mstrain) and 1 Hz with a stimula-tion period of 24 h. Knobs on the surface of the dishesrelayed the strain to the gels. Control gels were insertedin similar silicon dishes but unattached to the surface toprevent any mechanical stimulus; control gels existedfor each of the defined osmotic conditions. Followingmechanical stimulation, both control and loaded gelswere cultured for further 24 h in an incubator beforedissolving the gels (as described above) and examiningcellular responses.
RNA Isolation and Real-Time Quantitative RT-PCR
Cellular responses were examined as previouslydescribed.13 Briefly, total RNA was prepared, measured,and transcribed into cDNA. Based on published genesequences (NCBI Entrez search system), specific bovineand human primers were designed for the matrix
proteins aggrecan, collagen I, and collagen II (seeTable 2). Gene expression was determined by real-timequantitative RT-PCR on the iCycler analysis system(Biorad, Munich, Germany) with use of standards forquantification (obtained by cloning as described13).GAPDH was chosen as the housekeeping gene afterverifying that its expression in stimulated samples andunstimulated control samples differed by only 0.1concerning the ct value (Dct). The average ct value ofeach duplicate measurement of each sample wasnormalized to the GAPDH in each sample (e.g.,Dct¼ ct(aggrecan)� ct(GAPDH)). DDct values were calculat-ed by referring Dct values to the respective controland relative mRNA was then calculated by theequation 2DDct.
Data Collection and Statistical Analysis
Gene expression of hypo-osmotic, iso-osmotic, andhyperosmotic cultures was measured. For human cellcultures without mechanical stimulation, gene expres-sion after 5 days of culture at 400 mOsm and 500 mOsmwas compared to 300 mOsm. Ratio of gene expression ineach subgroup was statistically tested by use of the t-test(p< 0.05). In addition the nonparametric Wilcoxon2 sample test (p< 0.05) was performed in order todetermine differences between gene expression atdifferent osmotic conditions.
Effects of mechanical stimulation were statisticallytested by use of the t-test (stimulated group vs. associatedcontrol group, p< 0.05). Stimulation-induced effects forthe three different osmotic conditions were compared ingroups by the nonparametric Wilcoxon 2 sample test(p< 0.05) for all osmotic conditions. Results are shown asan increase or a decrease in gene expression in the
Table 2. Primer Sequences
Gene Species Sequence (Forward and Reverse Primer)
SequenceAccessionNumber
Aggrecan Bovine 50-ACA GCG CCT ACC AAG ACA AG-30 U7661550-ACG ATG CCT TTT ACC ACG AC-30
Collagen I Bovine 50-TGA CCT CAA GAT GTG CCA CT-30 AF01717850-ACC AGA CAT CCC TCT TGT CC-30
Collagen II Bovine 50-CCT GTA GGA CCT TTG GGT CA-30 X0242050-ATA GCG CCG TTG TGT AGG AC-30
GAPDH Bovine 50-ACC CAG AAG ACT GTG GAT GG- 30 U8504250-CAA CAG ACA CGT TGG GAG TG-30
Aggrecan Human 50-TCT GTA ACC CAG GCT CCA AC-30 XM_00770150-CTG GCA AAA TCC CCA CTA AA-30
Collagen I Human 50-TGACCTCAAGATGTGCCACT-30 AF01717850-ACC AGA CAT CCC TCT TGT CC-30
Collagen II Human 50-CGC ACCTGCAGAGACCTGAA-30 XM_05648150-TCT TCT TGG GAA CGT TTG CTG G-30
GAPDH Human 50-GAAGGTGAAGGTCGGAGTC-30 J0403850-GAA GAT GGT GATGGG ATT TC-30
1516 WUERTZ ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007 DOI 10.1002/jor
stimulated samples compared to the control samples (inpercentage).
RESULTS
Results show that gene expression of IVD cells isstrongly influenced by changes in the extracellularosmolarity. After 5 days of culture, large differ-ences in gene expression between the chosenosmotic conditions could be noted. The effects ofload application on matrix protein expression weremodest when compared to those of osmolarity;nevertheless, the osmotic environment influencedthe changes in mRNA expression due to mechan-ical stress.
Cell Culture at Controlled Osmolarity (withoutMechanical Stimulation)
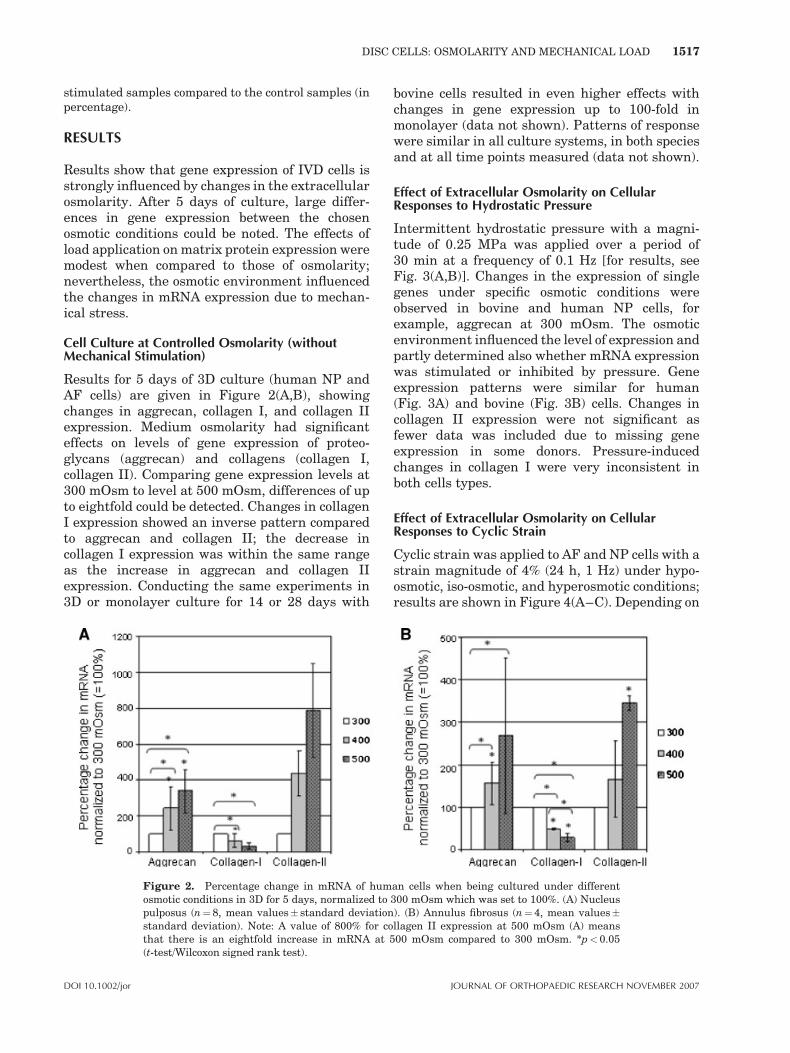
Results for 5 days of 3D culture (human NP andAF cells) are given in Figure 2(A,B), showingchanges in aggrecan, collagen I, and collagen IIexpression. Medium osmolarity had significanteffects on levels of gene expression of proteo-glycans (aggrecan) and collagens (collagen I,collagen II). Comparing gene expression levels at300 mOsm to level at 500 mOsm, differences of upto eightfold could be detected. Changes in collagenI expression showed an inverse pattern comparedto aggrecan and collagen II; the decrease incollagen I expression was within the same rangeas the increase in aggrecan and collagen IIexpression. Conducting the same experiments in3D or monolayer culture for 14 or 28 days with
bovine cells resulted in even higher effects withchanges in gene expression up to 100-fold inmonolayer (data not shown). Patterns of responsewere similar in all culture systems, in both speciesand at all time points measured (data not shown).
Effect of Extracellular Osmolarity on CellularResponses to Hydrostatic Pressure
Intermittent hydrostatic pressure with a magni-tude of 0.25 MPa was applied over a period of30 min at a frequency of 0.1 Hz [for results, seeFig. 3(A,B)]. Changes in the expression of singlegenes under specific osmotic conditions wereobserved in bovine and human NP cells, forexample, aggrecan at 300 mOsm. The osmoticenvironment influenced the level of expression andpartly determined also whether mRNA expressionwas stimulated or inhibited by pressure. Geneexpression patterns were similar for human(Fig. 3A) and bovine (Fig. 3B) cells. Changes incollagen II expression were not significant asfewer data was included due to missing geneexpression in some donors. Pressure-inducedchanges in collagen I were very inconsistent inboth cells types.
Effect of Extracellular Osmolarity on CellularResponses to Cyclic Strain
Cyclic strain was applied to AF and NP cells with astrain magnitude of 4% (24 h, 1 Hz) under hypo-osmotic, iso-osmotic, and hyperosmotic conditions;results are shown in Figure 4(A–C). Depending on
Figure 2. Percentage change in mRNA of human cells when being cultured under differentosmotic conditions in 3D for 5 days, normalized to 300 mOsm which was set to 100%. (A) Nucleuspulposus (n¼8, mean values� standard deviation). (B) Annulus fibrosus (n¼ 4, mean values�standard deviation). Note: A value of 800% for collagen II expression at 500 mOsm (A) meansthat there is an eightfold increase in mRNA at 500 mOsm compared to 300 mOsm. *p<0.05(t-test/Wilcoxon signed rank test).
DISC CELLS: OSMOLARITY AND MECHANICAL LOAD 1517
DOI 10.1002/jor JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007
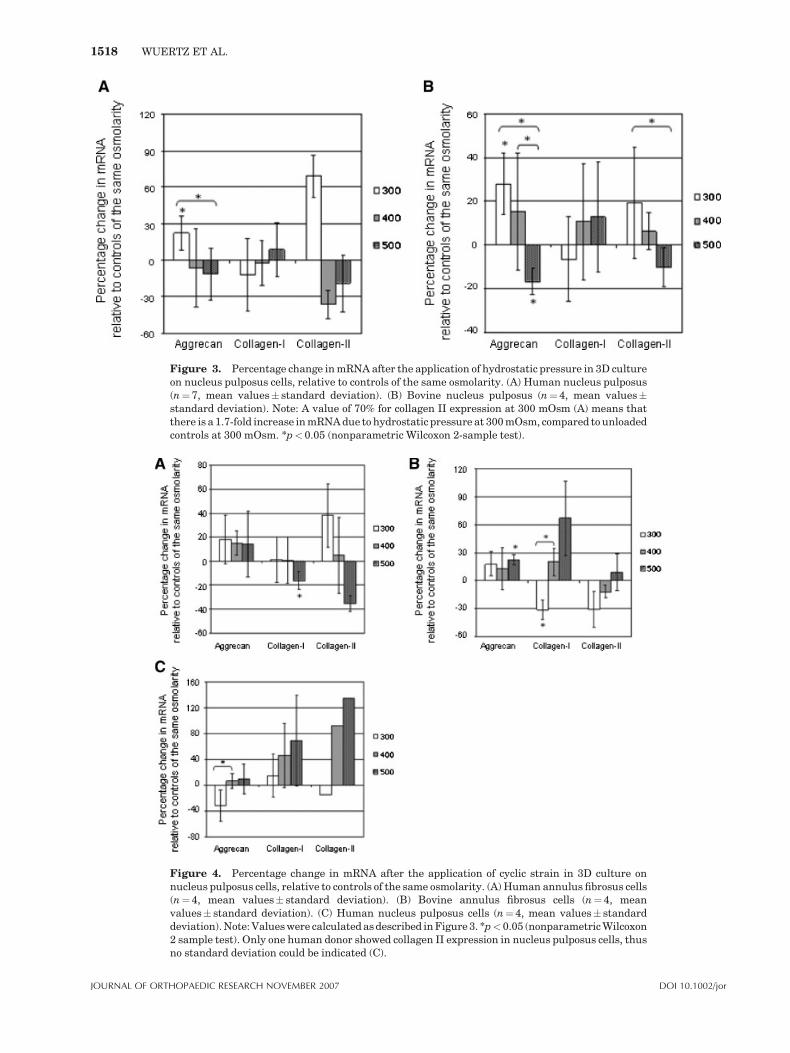
Figure 3. Percentage change in mRNA after the application of hydrostatic pressure in 3D cultureon nucleus pulposus cells, relative to controls of the same osmolarity. (A) Human nucleus pulposus(n¼7, mean values� standard deviation). (B) Bovine nucleus pulposus (n¼4, mean values�standard deviation). Note: A value of 70% for collagen II expression at 300 mOsm (A) means thatthere is a 1.7-fold increase in mRNA due to hydrostatic pressure at 300 mOsm, compared to unloadedcontrols at 300 mOsm. *p<0.05 (nonparametric Wilcoxon 2-sample test).
Figure 4. Percentage change in mRNA after the application of cyclic strain in 3D culture onnucleus pulposus cells, relative to controls of the same osmolarity. (A) Human annulus fibrosus cells(n¼4, mean values� standard deviation). (B) Bovine annulus fibrosus cells (n¼ 4, meanvalues� standard deviation). (C) Human nucleus pulposus cells (n¼4, mean values� standarddeviation). Note: Values were calculated as described in Figure 3. *p<0.05 (nonparametric Wilcoxon2 sample test). Only one human donor showed collagen II expression in nucleus pulposus cells, thusno standard deviation could be indicated (C).
1518 WUERTZ ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007 DOI 10.1002/jor

species and IVD zone, changes in mRNA levelscould be observed for some single genes underspecific osmotic conditions, for example, collagen Iin bovine AF cells at 300 mOsm (Fig. 4B).Aggrecan expression in bovine and human AFcells seemed to be independent from changes inthe extracellular osmolarity and tended to belightly stimulated by the application of cyclicstrain at all osmotic values with significant resultsonly in bovine AF cells at 500 mOsm. Effects oncollagen I and collagen II expression differed forbovine and human AF cells. Changes in collagen IIexpression were not significant as fewer data wasincluded due to missing gene expression in somedonors. Human NP cells didn’t show significantchanges in gene expression in response to cyclicstrain (Fig. 4C); trends were not similar toresponses of human AF cells (Fig. 4A).
DISCUSSION
In this study, we showed that gene expression ofIVD cells is influenced by the osmotic environment(Fig. 2). Although only results of human IVD cellsafter 5 days of osmotic culture were illustrated,similar patterns of response with even highereffects were seen for longer culture periods (14 or28 days) in bovine cells, especially in monolayer(data not illustrated). In comparison to the osmoticenvironment, mechanical stimulation had less ofan effect on cellular responses; however, this studyshowed that loading effects, though being minor,partly depend on the osmotic situation (Figs. 3, 4).
Application of hydrostatic pressure increasedaggrecan and collagen II expression in human andbovine NP cells under hypo-osmotic conditions butthe identical pressure application inhibited expres-sion of aggrecan and collagen II in a hyperosmoticenvironment (Fig. 3). Expression of collagen I,however, was not influenced by hydrostatic pres-sure, regardless of the osmotic environment(Fig. 3). Gene expression in response to theapplication of cyclic strain was also sensitive tothe osmotic environment but the effects variedwith cell type (Fig. 4); collagen I and collagen IIexpression were inhibited in a hyperosmoticenvironment in human AF cells, but tended to bestimulated in human NP cells. It has to be notedthat the osmotic environment differs in the zonesof the IVD with higher values in NP. Osmoticconditions named as hypo-, iso-, or hyperosmoticwere chosen identically for cells of each zone,representing a mean value for the disc with
consequent light discrepancies from the real valueof the respective zone.
Despite the large diurnal changes in extracel-lular osmolarity in the disc in vivo because of load-induced water expression and re-imbibition, only afew studies have investigated the role of osmolarityon matrix synthesis or gene expression in IVD orother cartilages to date. In agreement with ourfindings (data not illustrated), Ishihara and collea-gues observed an increase of sulphated GAGsynthesis in bovine NP cells with increase inosmolarity.6 In an organ culture system, Hascht-mann and colleagues found that an increase inosmolarity induced a (tendential) upregulation ofaggrecan and downregulation of collagen I expres-sion,16 similar to our findings. Studies on articularchondrocytes showed that extracellular osmolarityinfluenced matrix synthesis and regulated aggre-can promoter activity as well as aggrecan expres-sion and sulphated GAG synthesis.17–20 Bycontrast, pig notochordal NP cells in alginateculture showed little response to changes inextracellular osmolarity in regard to expression ofaggrecan, collagen I and collagen II;21 only cells ofthe TZ responded and showed a higher expressionof collagen I under hypo-osmotic conditions. It hasto be noted that our study as well as most otherrecent studies were based on isolated cells. Cellculture studies can only serve as a model with aconsequent lack of transferability to the in vivosituation. It is not certain up to now if the observedeffects would appear in the same way in tissueexplants or in the tissue in vivo as it is wellknown that cell–matrix interactions may play animportant role in signal transduction. Never-theless, IVD cells seem to be sensitive to changesin their osmotic environment, occurring in vivobecause of load application or during degenerativeprocesses.
Loading of the IVD results in circadian changesin its hydration and thus osmolarity. Changes inloading and activity result in expression of around20% to 25% of the disc’s fluid.22 The extracellularosmolarity thus varies widely over the space of afew hours; in the NP where extracellular osmolar-ity is greatest, it is estimated that osmolarity variesfrom 450 mOsm to 550 mOsm, depending on thetime of day.23 Not only does the change inosmolarity influence cell function strongly17–20
but it also affects responses to mechanical signalsas shown here (Figs. 3, 4); cellular responses topressure or deformation could thus vary dependingon time of day with pressure application stimulat-ing aggrecan expression in the morning day(400 mOsm) but inhibiting it in the evening
DISC CELLS: OSMOLARITY AND MECHANICAL LOAD 1519
DOI 10.1002/jor JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007

(500 mOsm; Fig. 3). Mechanical stimulation underdifferent osmotic conditions may help simulatingcircadian changes in the physicochemical environ-ment in the disc matrix under loading. Themechanisms involved in the osmotic response andtheir interactions are unknown but changes in cellvolume could be partly involved. The volume of disccells, like those of other mammalian cells,24 alterswith change in extracellular osmolarity; volumefalls under hyperosmotic conditions through fluidloss and cells swell under hypo-osmotic conditionsbecause of water imbibition;6,7,25 these findingwere confirmed in our study histologically and bySEM (not shown).
The effect of changes in volume on cell physiol-ogy has hardly been examined in disc cells thoughosmolarity has been shown to regulate (Ca2þ)itransients in notochordal NP cells; in chondrocytes,volume changes are known to have powerfuleffects on cells, altering transporter activity,activating Ca2þ transients and initiating signalingcascades and gene expression.24,26–28 It has beenshown that swollen or shrunken articular chon-drocytes respond differentially to mechanical sti-mulation; thus sensitivity of cells to mechanicalloading as well as the consequent responses to thestimuli appear influenced by the osmotic environ-ment.29 In summary, application of mechanicalload is not only a stimulus for cells on its own, butalso results in changes of the osmotic environmentbecause of fluid expression, hence affecting furtherresponses to mechanical signals.
It should be noted that cellular responses toosmolarity are time dependent in other cell typesincluding articular cartilage cells. In chondrocytes,exposure to a hyperosmotic environment initiallyleads to a fall in sulphate incorporation and proteinsynthesis; later, cells adapt and synthesis isstimulated.17 Here, the disc cells were exposed tolong-term incubation at different osmolarities andby analogy with cartilage cells, it appears that theyhad adapted to changing conditions. Modeling thediurnal osmotic changes will claim to analyzechanges in gene expression occurring after hoursrather than days. Thus, cellular responses seen inour experiments might not be directly transferableto diurnal osmotic changes in vivo. Our resultsare however likely to simulate long-term changesas seen, for example, during disc degeneration.Hydration of the disc and extracellular osmolarityof disc cells is altered during disc degeneration. Oneof the major changes in disc degeneration is a fall inaggrecan content30 with consequent falls in fixedcharge density and extracellular osmolarity. Cellsin degenerated discs thus exist under hypo-osmotic
conditions compared to those in normal discs.According to the results in this study, the fall inosmolarity could reduce aggrecan and collagen IIexpression substantially (Fig. 2). Thus the fall inextracellular osmolarity that is present duringdisc degeneration might reduce matrix synthesis,further promoting degenerative processes. Never-theless, it still remains unclear if the observedchanges in gene expression would affect proteinlevels as well; experiments analyzing the influenceof the extracellular osmolarity on protein synthesiswill be conducted in future studies.
It has to be noted that this study has some mainlimitations: As many factors (such as loadingsituation, age, degeneration) influence the extra-cellular osmolarity, our simplified cell culturemodel cannot be a perfect simulation of the complexin vivo situation. Though the chosen range from300 to 500 mOsm is physiological, it does notperfectly represent the situation in both NP and AFtissue. Due to differences in the biochemicalcomposition, these tissue structures have a differ-ent osmotic environment with higher values forNP. It is important to note that this experimentserves only as a model and, as such, is characterizedby a limited transferability to the in vivo situation.In addition, differences to the in vivo situationoccur due to the fact that expansion in monolayer isnecessary to obtain a sufficient cell number. Thismay induce changes in phenotype and thuschanges in gene expression profiles (mainly forcollagen type II) as described in the literature.31,32
Due to limited starting cell numbers and lowerproliferation rates, human IVD cells had to beexpanded in monolayer for up to three passagesalthough it is known that de-differentiation (atleast of AF cells) occurs mostly after passage 231;this explains why collagen type II expression insome human donors could not be detected anymore.Variation in our data might thus be due todifferences in monolayer passaging; in addition,human disc biopsies were obtained from donorswith different age, degeneration degree, and fromdifferent disc levels. All these factors as well as thefact that it is hard to distinguish between NP andAF tissue in a herniated disc might be responsiblefor the lack of uniformity in our results. Anotherlimitation can be found in the choice of a collagentype I gel as a scaffold for both, NP and AF cells.In vivo, AF cells are embedded in a matrix thatpredominantly consists of collagen type I, whereasNP cells are mostly surrounded by collagen type IIand aggrecan; thus, a collagen type I gels is moresuitable for AF cells. However, culturing IVD cellsin a 3D environment such as a collagen type I gel is
1520 WUERTZ ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007 DOI 10.1002/jor

more appropriate than monolayer culture which isoften performed in cell culture experiments.
IVD cells respond to changes in the extracellularosmolarity by altering cell volume and geneexpression. Increasing the osmolarity resulted inan increase of aggrecan and collagen II expression.A fall in extracellular osmolarity as seen indegenerated discs resulted in altered mRNA levelsthat might further promote degenerative processesif transferred to protein levels. The effects ofmechanical stimulation were less profound, butpartly depended on the osmotic environment;shifting from hypo-osmotic to hyperosmotic condi-tions could invert responses to the same mechan-ical stimulus. Pressurization of human nucleuscells as well as stretching of human annulus cellstended to result in an increased expression ofaggrecan and collagen II under hypo-osmoticconditions.
ACKNOWLEDGMENTS
The authors gratefully acknowledge Mrs. DanielaWasiak for excellent technical assistance andMr. Herbert Schmitt for expert construction of thestimulation apparatus. Special thanks go to ArsArthroAG (Esslingen, Germany) for providing the collagen gelas well as to Mr. Schwarz (Ulmer Fleisch GmbH, Ulm,Germany) for placing the bovine tails at our disposal.This work was supported by the European Unionresearch project EURODISC QLK6-CT-2002-02582 aswell as by the German Research Council DFG. Jill Urbanis an Arthritis Research Campaign Senior ResearchFellow.
REFERENCES
1. Urban JP, Maroudas A. 1981. Swelling of the interverteb-ral disc in vitro. Connect Tissue Res 9:1–10.
2. Urban JP, Maroudas A, Bayliss MT, et al. 1979. Swellingpressures of proteoglycans at the concentrations found incartilaginous tissues. Biorheology 16:447–464.
3. Urban JP, McMullin JF. 1985. Swelling pressure of theintervertebral disc: influence of proteoglycan and collagencontents. Biorheology 22:145–157.
4. Wilke HJ, Neef P, Caimi M, et al. 1999. New in vivomeasurements of pressures in the intervertebral disc indaily life. Spine 24:755–762.
5. Lang F, Busch GL, Ritter M, et al. 1998. Functionalsignificance of cell volume regulatory mechanisms. PhysiolRev 78:247–306.
6. Ishihara H, Warensjo K, Roberts S, et al. 1997. Proteogly-can synthesis in the intervertebral disk nucleus: the roleof extracellular osmolality. Am J Physiol 272:C1499–C1506.
7. Pritchard S, Erickson GR, Guilak F. 2002. Hyperosmoti-cally induced volume change and calcium signaling in
intervertebral disk cells: the role of the actin cytoskeleton.Biophys J 83:2502–2510.
8. Adams MA, McNally DS, Dolan P. 1996. ‘‘Stress’’ distribu-tions inside intervertebral discs. The effects of age anddegeneration. J Bone Joint Surg [Br] 78:965–972.
9. Nachemson A. 1995. Lumbar intradiscal pressure—experimental studies on post-mortem materials. Essen.
10. Shirazi-Adl A, Ahmed AM, Shrivastava SC. 1986. Afinite element study of a lumbar motion segmentsubjected to pure sagittal plane moments. J Biomech 19:331–350.
11. Stokes IA. 1987. Surface strain on human intervertebraldiscs. J Orthop Res 5:348–355.
12. Iatridis JC, ap Gwynn I. 2004. Mechanisms for mechanicaldamage in the intervertebral disc annulus fibrosus. JBiomech 37:1165–1175.
13. Neidlinger-Wilke C, Wurtz K, Liedert A, et al. 2005. Athree-dimensional collagen matrix as a suitable culturesystem for the comparison of cyclic strain and hydrostaticpressure effects on intervertebral disc cells. J NeurosurgSpine 2:457–465.
14. Bibby SR, Urban JP. 2004. Effect of nutrient deprivationon the viability of intervertebral disc cells. Eur Spine J13:695–701.
15. Maclean JJ, Roughley P, Alini M, et al. 2006. In vivo discremodeling, kinetics of mRNA expression in response to asingle loading event. Paper presented at: 33rd AnnualMeeting of the ISSLS. Bergen, Norway.
16. Haschtmann D, Stoyanov JV, Ferguson SJ. 2006. Influ-ence of diurnal hyperosmotic loading on the metabolismand matrix gene expression of a whole-organ interverteb-ral disc model. J Orthop Res 24:1957–1966.
17. Hopewell B, Urban JP. 2003. Adaptation of articular chon-drocytes to changes in osmolality. Biorheology 40: 73–77.
18. Palmer GD, Chao PH, Raia F, et al. 2001. Time-dependentaggrecan gene expression of articular chondrocytes inresponse to hyperosmotic loading. Osteoarthritis Cartilage9:761–770.
19. Schneiderman R, Keret D, Maroudas A. 1986. Effects ofmechanical and osmotic pressure on the rate of glycosa-minoglycan synthesis in the human adult femoral headcartilage: an in vitro study. J Orthop Res 4:393–408.
20. Urban JP, Hall AC, Gehl KA. 1993. Regulation of matrixsynthesis rates by the ionic and osmotic environment ofarticular chondrocytes. J Cell Physiol 154:262–270.
21. Chen J, Baer AE, Paik PY, et al. 2002. Matrix proteingene expression in intervertebral disc cells subjected toaltered osmolarity. Biochem Biophys Res Commun 293:932–938.
22. Paajanen H, Lehto I, Alanen A, et al. 1994. Diurnal fluidchanges of lumbar discs measured indirectly by magneticresonance imaging. J Orthop Res 12:509–514.
23. Urban JP. 2002. The role of the physicochemical environ-ment in determining disc cell behaviour. Biochem SocTrans 30:858–864.
24. Lang F, Busch GL, Volkl H. 1998. The diversity of volumeregulatory mechanisms. Cell Physiol Biochem 8:1–45.
25. Bayliss MT, Urban JP, Johnstone B, et al. 1986. In vitromethod for measuring synthesis rates in the intervertebraldisc. J Orthop Res 4:10–17.
26. Bush PG, Hall AC. 2001. The osmotic sensitivity of isolatedand in situ bovine articular chondrocytes. J Orthop Res19:768–778.
27. Hung CT, LeRoux MA, Palmer GD, et al. 2003. Disparateaggrecan gene expression in chondrocytes subjected to
DISC CELLS: OSMOLARITY AND MECHANICAL LOAD 1521
DOI 10.1002/jor JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007

hypotonic and hypertonic loading in 2D and 3D culture.Biorheology 40:61–72.
28. Hall AC, Starks I, Shoults CL, et al. 1996. Pathways for Kþtransport across the bovine articular chondrocyte mem-brane and their sensitivity to cell volume. Am J Physiol270:C1300–C1310.
29. Bush PG, Hodkinson PD, Hamilton GL, et al. 2005.Viability and volume of in situ bovine articular chondro-cytes-changes following a single impact and effectsof medium osmolarity. Osteoarthritis Cartilage 13:54–65.
30. Lyons G, Eisenstein SM, Sweet MB. 1981. Biochemicalchanges in intervertebral disc degeneration. BiochimBiophys Acta 673:443–453.
31. Chou AI, Bansal A, Miller GJ, et al. 2006. The effect ofserial monolayer passaging on the collagen expressionprofile of outer and inner anulus fibrosus cells. Spine 31:1875–1881.
32. Marlovits S, Hombauer M, Truppe M, et al. 2004. Changesin the ratio of type-I and type-II collagen expression duringmonolayer culture of human chondrocytes. J Bone JointSurg [Br] 86:286–295.
1522 WUERTZ ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH NOVEMBER 2007 DOI 10.1002/jor




















