
Infertility in Herero pastoralists of southern Africa
19
AMERICAN JOURNAL OF HUMAN BIOLOGY 3:135-153 (1991) Infertility in Herero Pastoralists of Southern Africa RENEE PENNINGTON AND HENRY HARPENDING Department of Anthropology, Pennsylvania State University, University Park, PA 16802 ABSTRACT Transition from low to high rates of fertility among Herero pastoralists of the northern Kalahari of Botswana is examined.Total fertility rates have increased from 2.65 in the first half of this century to 7.02 in the last decade, while postreproductive women report having had only 3.47 births. We use an indirect estimator of the fertility of mothers to show that the Herero have been afflicted with abnormally low fertility since early in this century. Although the several possible causes of subfertility in this population, including disease, mater- nal health, and child care practices are examined,it is concludedthat the effects of venereal diseases are most likely responsible for abnormally low fertility. The dramatic increase in fertility following treatment of a major cause of infertility underscores the potential impact diseases may have on human reproductive patterns. This study is the first to document subfertility in an ethnic group of southern Africa. Many models of human evolution have stressed food acquisition as an important selective variable in the past. It has been suggested that differences in the quantity of fat stores and nutritional status of women are responsible for variation in levels of fer- tility and in the survivorship of offspring (see Huss Ashmore, 1980). For exam le, fat energy reserves during pregnancy and lac- tation (Hytten, 1980; Crawford, 1980; Harpending et al., 1990). Similarly, women who space births too closely may de lete fecting fecundity and the survivorship of young. Unfortunately, only a few studies rovide strong empirical support for these y otheses. n contrast, man diseases are known to have a ronounce B effect on reproductive of disease in the shaping of human re roduc- diminution of a major cause of infertility in a southern African population corresponds to a nearly threefold increase in fertility in the last three decades. Our analysis of maternity histories collected during a survey of Herero pastoralists living in the Kalahari Desert of northwestern Botswana indicates that, rior tility marked b truncated reproductive stores in females have been consi 2 ered as energy and nutritional stores, adverse P y af- [P success. P et, few models incorporate the role tive patterns. In this paper, we show l ! ow the to about 1960, women had low overal f fer- careers and that t K e unusually low reproduc- tive rates antedate the turn of the century (Pennington and Harpending, 1990). The impact of disease on re roductive success onstrated for nutritional models, suggesting that the influence of disease on fertility in human evolution merits further inquiry. This study is the first to present evidence of subfertility in southern Africa, a region that reviously has not been included in the well-known “African infertility belt” (Coale, 1968; Caldwell and Caldwell, 1983; Frank, 1983).Botswana, like the rest of the African continent, is characterized b population the world. We describe an ethnically distinct minorit PO ulation in Botswana with a pat- tern of Z r t i h y very different from the aver- age. Although the potential effects of nutrition and nonvenereal diseases on fertil- ity are considered, the pattern of fertility among Herero is most consistent with one burdened by the sequelae of venereal infec- tion. METHODS Study population and sampling area The Herero are Bantu-speaking pastoral- ists who live in scattered villages and cattle- may thus be far greater t K an has been dem- growth rates that are among t TI e highest in Received September 12,1990; accepted January 7, 1991. Renee Pennington’s current address is Center for Demography and Ecology, University of Wisconsin, Madison, WI 53706. 01991 Wiley-Liss, Inc
Transcript of Infertility in Herero pastoralists of southern Africa

AMERICAN JOURNAL OF HUMAN BIOLOGY 3:135-153 (1991)
Infertility in Herero Pastoralists of Southern Africa RENEE PENNINGTON AND HENRY HARPENDING Department of Anthropology, Pennsylvania State University, University Park, PA 16802
ABSTRACT Transition from low to high rates of fertility among Herero pastoralists of the northern Kalahari of Botswana is examined. Total fertility rates have increased from 2.65 in the first half of this century to 7.02 in the last decade, while postreproductive women report having had only 3.47 births. We use an indirect estimator of the fertility of mothers to show that the Herero have been afflicted with abnormally low fertility since early in this century. Although the several possible causes of subfertility in this population, including disease, mater- nal health, and child care practices are examined, it is concluded that the effects of venereal diseases are most likely responsible for abnormally low fertility. The dramatic increase in fertility following treatment of a major cause of infertility underscores the potential impact diseases may have on human reproductive patterns. This study is the first to document subfertility in an ethnic group of southern Africa.
Many models of human evolution have stressed food acquisition as an important selective variable in the past. It has been suggested that differences in the quantity of fat stores and nutritional status of women are responsible for variation in levels of fer- tility and in the survivorship of offspring (see Huss Ashmore, 1980). For exam le, fat
energy reserves during pregnancy and lac- tation (Hytten, 1980; Crawford, 1980; Harpending et al., 1990). Similarly, women who space births too closely may de lete
fecting fecundity and the survivorship of young. Unfortunately, only a few studies
rovide strong empirical support for these y otheses.
n contrast, man diseases are known to have a ronounce B effect on reproductive
of disease in the shaping of human re roduc-
diminution of a major cause of infertility in a southern African population corresponds to a nearly threefold increase in fertility in the last three decades. Our analysis of maternity histories collected during a survey of Herero pastoralists living in the Kalahari Desert of northwestern Botswana indicates that, rior
tility marked b truncated reproductive
stores in females have been consi 2 ered as
energy and nutritional stores, adverse P y af-
[ P success. P et, few models incorporate the role
tive patterns. In this paper, we show l! ow the
to about 1960, women had low overal f fer-
careers and that t K e unusually low reproduc-
tive rates antedate the turn of the century (Pennington and Harpending, 1990). The impact of disease on re roductive success
onstrated for nutritional models, suggesting that the influence of disease on fertility in human evolution merits further inquiry.
This study is the first to present evidence of subfertility in southern Africa, a region that reviously has not been included in the well-known “African infertility belt” (Coale, 1968; Caldwell and Caldwell, 1983; Frank, 1983). Botswana, like the rest of the African continent, is characterized b population
the world. We describe an ethnically distinct minorit PO ulation in Botswana with a pat- tern of Z r t i h y very different from the aver- age. Although the potential effects of nutrition and nonvenereal diseases on fertil- ity are considered, the pattern of fertility among Herero is most consistent with one burdened by the sequelae of venereal infec- tion.
METHODS Study population and sampling area
The Herero are Bantu-speaking pastoral- ists who live in scattered villages and cattle-
may thus be far greater t K an has been dem-
growth rates that are among t TI e highest in
Received September 12,1990; accepted January 7, 1991. Renee Pennington’s current address is Center for Demography
and Ecology, University of Wisconsin, Madison, WI 53706.
01991 Wiley-Liss, Inc

136 R. PENNINGTON AND H. HARPENDING
posts in the remote Kalahari Desert of northwestern Botswana. We interviewed Herero living in western Ngamiland in the area west of the Okavango Delta in the Qangwa, NxauNxau, Xaudum, and XaiXai valleys; in Sehitwa and along the ridge southeast of Lake Ngami between Bodibeng and Tsoku; and between Tsau and Shakawe along the western edge of the Okavango Delta (see Fig. 11. Herero speakers who iden- tified themselves as ethnically Herero or Mbanderu (the two Herero-speaking tribes in Botswana) were classified as Herero. Most are descended from either two Herero or two Mbanderu parents. Most live in grass-roofed mud houses without electricity of running water and make their living from cattle. Many also tend gardens and small stock. A few Herero living in the larger villages are engaged in wage employment, but it is un- like1 that their income contributes signifi-
living in the remote areas included in this study.
The size of the Herero population in Botswana is unknown but probably numbers on the order of 10,000-15,000 individuals (Pennington and Harpending, 1990) out of a national opulation of about one million (Central gatistics Office, 1981). Most of the Herero in Botswana originally arrived as refugees durin the Herero-German war of 1904-1907. Un a py with German colonial
critical defeat in the Battle of Waterberg in August of 1904, they dispersed, many of them crossing the waterless Omaheke Desert to Botswana. Although many Herero died of thirst or starvation during the flight, tens of thousands of those remaining in Namibia (then Southwest Africa) became the victims of a German genocide campaign (Bley, 1971; Bridgeman, 1981). At least 1,700-1,800 Herero must have settled in Botswana as a result of the war (Alnaes, 19891, although the true number of refugees may have been closer to 5,800 individuals (Pennington and Harpendin 1990). In
declining population until fertility began to increase in the 1960s, and their intrinsic growth rate is now about 3.5% per year (Pennington and Harpending, 1990). The majority live in Ngamiland, where we ascer- tained about 3,500 living individuals. Since the turn of the century, the Herero have transformed from destitute refugees to pros- perous cattle farmers. They are regarded as
cant P y to the subsistence base of Herero
rule, they revolte III in 1904. Following their
Botswana, they appear to have f een a slowly
staunch traditionalists who have remained an ethnically distinct minority population in Botswana.
Because of its harsh climate and scarcity of permanent sources of water, much of Ngamiland is uninhabited. In the occupied northern valleys included in this study, pop- ulation densities are less than 0.1 persons/ km2 compared to 1 to 10 personskm’ in the Lake N ami area (Central Statistics Office,
35°C in the warm months of the year, but in the winter months it becomes cold enough to freeze standing water. The rainy season typ- ically begins in November or December and begins tapering off in March. Levels of pre- cipitation vary widely across narrow geo- graphic ranges and from year to year such that areas separated by only a few kilome- ters may receive significantly different amounts of rain in any year. Although drought years are not uncommon, the driest part of Ngamiland receives an average of 400 mdyear (Central Statistics Office, 1988). Throughout the Kalahari rainfall exceeds that of “true” deserts, but the tendency for evaporation to exceed precipitation through- out the year produces an arid, desert-like environment. Conse uently, Botswana is
prevalent in the moist tropics.
1981). i mbient temperatures may excede
not overwrought wit Il the many diseases
Data collection Maternity histories of 611 Herero were
collected from July 1987 to March 1989 in two person-years of field work by the au- thors. These histories were obtained by
Herero settlements scattered out the study area. All women of
age and older, living or visiting at each homestead, were interviewed to as- certain their birth year and the years of birth and death of children. Men were interviewed about the births of their wives, both livin and dead. Both men and women were aske to provide re roductive histories of deceased
tive histories were obtained (see Penning- ton, 1990a; Harpending and Pennington, 1991, for more details).
All Herero are familiar with the European calendar, but few know the Gregorian year of a vital event if it occurred before about 1960. However, most know the Herero name of the year in which the event occurred. Tradition- ally Herero have assigned names to years that reflect some important event occuring in that year. These year names can be se-
8 daughters. A P together, 611 female reproduc-

INFERTILITY IN HERERO PASTORALISTS 137
Dobe
Fig. 1. Detailed map of study area in northwestern Botswana.
quenced and aligned with Gregorian years so that they can be used to date the years of birth and death of individuals. We used the list published in Vedder (1966b) to date vital events occurring prior to this century. The list of Mbanderu year names in Almagor (1980) was used to date events occurring in the 20th century. Officially, Mbanderu and Herero maintain separate year lists, but there is a great deal of overlap between them. Reuse of some year names occurs, such as ojozombahu (the year of locusts), which was used in both 1917 and 1924. Careful questionin of informants was usually suffi-
names occurring in this century, but some- times there was confusion concerning year names in earlier centuries. The year list compiled from published sources was sup- plemented in the field by knowledgeable in- dividuals and is given in Pennington (1990a).
The birth ear of a Herero is, in a sense,
name several others who were born the same year. We used references to agemates to obtain the birth years of several individuals whose birth year was otherwise unknown. In addition, Herero who have been to school frequently know the exact dates of birth and death of their children. Since discrepancies about month or season of birth occasionally occurred between reports about individuals
cient to dif B erentiate between duplicate year
part of his i B entity so that most are able to
that were multiply ascertained, years of vital events are all reported in whole years only.
The method of dating vital events using year names is advantageous because it is not susceptible to common problems of age esti- mation found in many populations, such as age-hea ing on multiples of five (Brass,
variation among sources in the correct se- quence of Herero and Mbanderu year names so that the assi ed year of a vital event is
error results from traditional years begin- ning and ending at the start of each rainy season, usually November or December, rather than in Januar . Consequently, someone born in Novem l er 1919 may be recorded in our database as being born in 1920 if they reported their birth year using a Herero year name.
RESULTS Desire for large families is universal in
Africa such that childlessness is a heartfelt tragedy (Caldwell and Caldwell, 1987). In this section we describe abnormally low lev- els of fertility among Herero, including child- lessness, using several different measures of fertility. First, age-specific fertility rates are presented to illustrate the unusually low rates of fertility characterizing Herero re- roductive erformance rior to the 1960s.
1968). 8 n the other hand, there is some
occasionally off r y a year or two. Additional
b e show t K at fertility \as gradually in-

138 R. PENNINGTON AND H. HARPENDING
creased and is presently at its highest level in at least a century. Next, the completed family size distributions of post-reproduc- tive women indicate temporal variation in fertility levels among birth cohorts of women as well as an unusually high frequency of small family sizes. These family size distri- butions are used to compute parity progres- sion ratios to illustrate the onset of sterility in Herero. Finally, we compare the parity distributions and arity progression ratios with those impute c f from reports of maternal family size using a technique described by Har ending and Draper (1990).
Akhough contrace tive use is advocated
cently, few women voluntarily limited their fertility (Frank, 1987). In Botswana, the Family Health Survey (which included few, if any, Herero) re orted that 15% of rural
tives (Manyeneng et al., 1985). However, since these reports are based on self-reports of current use, they cannot be used to esti- mate numbers of births averted. A few Her- ero told us that they used contraceptives, but these tended to be older women whose chances of having another birth were low anyhow. In addition, contraceptives are fre- quently used for spacing, rather than stop- ping, births (Wray et al., 1987) or for preventing conceptions with the “wrong” man. Given the currently hi h birth rate
argue that a significant number of births are averted by contraceptives and, more impor- tantly, that any contraceptive use was prac- ticed in the ast that would account for the
Age -sp ecific ferlility Age-specific fertility rates (ASFRs) are
computed by summing the number of births to women at each age and dividing this sum by the number of women at risk at that age. The total fertility rate (TFR) is the number of live births a woman would have at the end of childbearing for a set of ASFRs. If the fertil- ity rates of a o ulation are constant
independent of parity, the TFR will be equal to the cohort fertility rate, the expected num- ber of births occurring to women of a birth cohort at the end of childbearing, and the completed family size (CFS), the number of live births retrospectively reported b
bearing (Newell, 1988).
throughout Africa ( 6 age, 19881, until re-
women used some f orm of modern contracep-
among Herero women, it wou 7 d be hard to
low rates of P ertility reported below.
through time an B P i maternal mortality is
women who have reached the end of chi1 B -
Parts A and B of Table 1 are the sum of the number of births and person-years of expo- sure occurring to married and unmarried Herero women combined for five-year age classes in four periods. Part C of the table shows the yearly ASFR for seven age classes and the TFRs (the sum of five times each ASFR) computed from Parts A and B for each period. The number of births and person- years of exposure by single year are reported in Pennington (1990a). Altogether, there were 12,774 erson-years and 1,921 births in 1909 througRl986.
Since this analysis is primarily based on fertility histories re orted by living women, it has been assumec! that the fertility histo- ries are representative of all women, i.e., women who have died do not have higher or lower fertility than women alive toda . It has also been assumed that all births an B deaths occur on the last day of a year so that any woman known to be alive in any year contrib- utes a full year of risk for her age in that year. These tabulations begin in 1909 be- cause that is the year the oldest woman in the sample was 15 and end in 1986, the year prior to the start of this study. The fertility rates of women reproducing in 1909-1956 have been combined in this analysis due to data sparseness during the first two de- cades, especially at the oldest a es. How- ever, no variability in rates are evi i ent when age-specific fertility rates are computed for narrower time periods.
The plot of the ASFRs is shown in Figure 2. The curves for the different time periods show that fertility was lowest in the period 1909-1956 in which the TFR was only 2.65. Fertility began increasing among women of all ages in the period 1957-1966, resulting in a TFR of 7.02 for the most recent period, 1977-1986. Each eriod shows a pattern of
tions in which it has been shown that mari- tal fertility rates tend to peak in the mid- twenties and decline thereafter (Henry, 1961; Wood, 1989). The high recent fertility rates compare with TFRs of 6.2 in the 1981 census population of Botswana as a whole (Central Statistics Office, 1981) and with 6.5 in the 1984 Botswana Family Health Survey (Manyeneng et al., 1985). The lower fertility of the Botswana population ma be attribut- able to ethnic differences, to B ifferences in rural and urban fertility, or to the timing of the surveys. Urban dwellers in the family health survey, for example, had TFRs of one less than the rural population (Manyeneng
fertility typical o P natural fertility popula-

139 INFERTILITY IN HERERO PASTORALISTS
TABLE 1 . Age-specific fertility rates for four periods
Age 1909-56 1957-66 1967-76 1977-86 Total Part A: Number of person-years by age in four periods
15-19 20-24 25-29 30-34 35-39 40-44 45-49 Total
15-19 20-24 25-29 30-34 35-39 40-44 45-49 Total
15-19 20-24
1146 427 932 412 752 386 590 335 373 370 190 395 73 297
4056 2622
447 399 422 394 373 323 367
2725
151 161 83 27 16 5 0
443
0.1318 0.1727
Part B: Number of births to women by age in four periods
75 86 83 71 36 27 16 4
312
114 96 71 43 22 5
437
Part C: Age-specific fertility rates in four periods
0.1756 0.1924 0.2015 0.2857
25-29 0.1104 0.1839 0.2275 30-34 0.0458 0.1075 0.1802 35-39 0.0429 0.0730 0.1153 40-44 0.0263 0.0405 0.0681
713 644 441 397 422 391 363
3371
156 224 132 99 81 31 6
729
0.2188 0.3478 0.2993 0.2494 0.1919 0.0793
2733 2387 2001 1716 1538 1299 1100
12774
468 582 382 233 167 74 15
1921
45-49 0.0000 0.0135 0.0136 0.0165 TFR 2.6493 3.9772 5.4141 7.0153
0.4
0.351
0.3- 2 2 $ 0.25- E g 0.2- k V) 0.15-
0.1 ~
0.05-
Q
c r ._ m
0-
/ hl
TFRs: 1909: 2.65
1967: 5.41 1977: 7.02
1957: 3.98
Age class
Fig. 2. Age-specific fertility rates in four periods
et al., 1985). On the other hand, postrepro- ductive women in both the 1981 census and family health survey had completed family sizes of more than six compared to less than
four for the Herero. Based on a survey of 106 Mbanderu women living in the Lake Ngami area in the late 1970s, Almagor (1982) esti- mates their TFR at 6.45, a value intermedi-

140 R. PENNINGTON AI
ate to the rates we report for 1967-1976 and
The increase in fertility in the last half of this century is unusually dramatic. When estimates of fertility are based on retrospec- tive reports, the possibility that older women are underreporting births must be consid- ered. Several lines of evidence indicate that estimates of fertility are not biased by older women “forgetting” births or by underreport- ing deceased offspring. First, women with low fertility report their births as occurring while youn . Women with low fertility
than having their few births dispersed throughout their reproductive spans. The truncated reproductive careers resulted in an unusually low mean age at childbearing of about 22 years during the infertile periods (Pennington and Harpending, 1990). If women underreport births due to memory la se, they should be more likely to forget
those born later in repro uction. Second, older women report higher mortalit amon their young during infancy and ciildhoo8 than younger women (Harpending and Pen- nington, 1991) so that underreporting of de- ceased offspring cannot account for the increase in fertility. Moreover, offspring born to women early in re roduction are
ceased than offspring born later in reproduc- tion, yet, the majority of births are reported as occurring earlier in reproduction. The secondary sex ratio is also near unity, indi- cating that Herero are not preferentially re orting births of one sex more than the
fertility in the first half of this century is also supported by historical accounts in which the possibility that the Herero were actually committing “tribal suicide” by refusing to bear children was considered (Steenkamp, 1944; Gibson, 1959).
Family size distribution Direct. The total fertility rates of periods
are useful to illustrate temporal trends in fertilit or to make comparisons in the level of fertifity between populations at a particu- lar time. However, unless fertility is con- stant, TFRs are not good summaries of the experiences of cohorts of women. For exam- ple, women aged 4 0 4 4 in eriod 1977-1986
15-19 in 1947-1965. Thus, none of the peri- ods represents the experience of a single
1977-1986.
ceased chi1 I f bearing at young ages rather
a of P spring born earlier in re roduction than
older and therefore more li E ely to be de-
ot r: er. That the Herero had abnormally low
were born in 1932-1946; t K ese women were
\ID H. HARPENDING
cohort of women. The TFR of each period reflects the fertility experience of women born in four different decades, during which time fertility has undergone a dramatic tran- sition. In contrast, completed family size (CFS), which is the mean number of live births born to a group of women at the end of childbearing, reflects the experience of a seemingly more homogeneous, or better de- fined group of women, although it reveals nothin about the timing of reproduction.
fertility of small populations in which there are too few individuals to calculate reliable estimates of age-specific rates. The CFS also can be accurately calculated in opulations
can report if they have reached menopause. Thus, it is a good statistic for making com- parisons with other anthropological popula- tions.
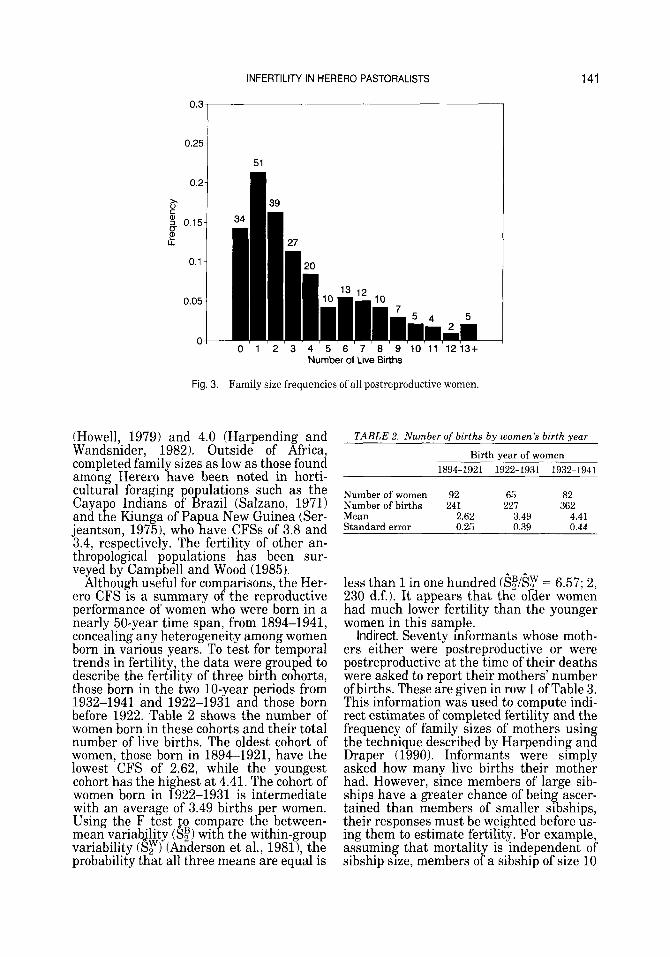
Famil size is the number of live births a
family sizes amon 239 postreproductive
1986) is shown in Figure 3. It is apparent that most women had small families-about 15% had no births and more than half had two or fewer. Famil sizes of only one are
occurring at parity six. Altogether the CFS of these women is 3.47, which is quite low. A CFS less than about 5 in natural fertilit
unusual circumstances such as sterilizing diseases or subfecundity. Almagor (1982) found that 11 Mbanderu women aged 4 5 4 9 in his study in the 1970s had an average of 5.18 births, but this sample is small and does not represent a defined birth cohort of women. However, few data on fertility of other African pastoralists are available for comparison. Among nomadic and sedentary Baggara cattle pastoralists in the Sudan, CFSs were 4.6 and 5.5, respectively (Henin, 1968). In the Sahel, postreproductive women among nomadic Tamasheq and Fu- lani pastoralists had mean family sizes ranging from 4.6 to 7.1 (Hill et al., 1983). Another group of Turkana women who had spent their reproductive years as nomadic pastoralists had 7.32 births (Brainard, 1986). One of the lowest CFSs in Africa was observed among the !Kung Bushmen, Khoisan hunter-gatherers residing in the same area as many of the Herero in this study. !Kung CFS has been estimated at 4.7
The C f S may also be the best summary of
in which women cannot report t g eir age but
woman K as had. The frequency of various
women (those age li 45 years or more in
the most fre uent. T K e distribution appears to be bimoda 4 , however, with a smaller peak
population tends to be associated wit i
INFERTILITY IN HERERO PASTORALISTS
0.25 0’3.
0.2-
x C
U
U
% 0.15- P
0.1 -
0.05-
07
39 34
141
Number of Live Births
Fig. 3. Family size frequencies of all postreproductive women.
(Howell, 1979) and 4.0 (Harpending and Wandsnider, 1982). Outside of Africa, completed family sizes as low as those found among Herero have been noted in horti- cultural foraging o ulations such as the
jeantson, 19751, who ave CFSs of 3.8 and 3.4, respectively. The fertility of other an- thropological populations has been sur- veyed by Campbell and Wood (1985).
Although useful for com arisons, the Her-
performance of women who were born in a nearly 50-year time span, from 1894-1941, concealing any heterogeneity among women born in various years. To test for temporal trends in fertility, the data were ou ed to
those born in the two 10-year eriods from 1932-1941 and 1922-1931 an8 those born before 1922. Table 2 shows the number of women born in these cohorts and their total number of live births. The oldest cohort of women, those born in 1894-1921, have the lowest CFS of 2.62, while the youngest cohort has the highest at 4.41. The cohort of women born in 1922-1931 is intermediate with an average of 3.49 births per women. Using the F test t o com are the between-
variability CST) (Anderson et al., 19811, the probability that all three means are equal is
Caya o Indians o h! razil (Salzano, 1971)
Yl and t R e Kiunga of Pa ua New Guinea (Ser-
era CFS is a summary o I p the reproductive
describe the fertility of three birt T R co orts,
mean variability ($1 wit K the within-group
TABLE 2. Number of births by women’s birth year
Birth year of women 1894-1921 1922-1931 1932-1941
Number of women 92 65 82 Number of births 241 227 362 Mean 2.62 3.49 4.41 Standard error 0.25 0.39 0.44
less than 1 in one hundred C S ~ l S ~ = 6.57; 2, 230 d.f.1. It appears that the older women had much lower fertility than the younger women in this sample.
Indirect. Seventy informants whose moth- ers either were postreproductive or were postreproductive at the time of their deaths were asked to report their mothers’ number of births. These are given in row 1 of Table 3. This information was used to compute indi- rect estimates of completed fertility and the frequency of family sizes of mothers using the technique described by Harpending and Draper (1990). Informants were sim ly
had. However, since members of large sib- ships have a greater chance of being ascer- tained than members of smaller sibships, their responses must be weighted before us- ing them to estimate fertility. For example, assuming that mortality is independent of sibship size, members of a sibship of size 10
asked how many live births their mot I: er

142 R. PENNINGTON AND H. HARPENDING
TABLE 3. Weighted number of mothers with various family sizes'
Familv size 1 2 3 4 5 6 7 8 9 10 11 12 13
Observed number 4 7 6 8 8 12 9 6 2 4 2 0 2 Weighted number 4 3.50 2 2 1.60 2 1.29 0.75 0.22 0.40 0.18 0 0.15
'Family sizes are reported by offspring. Family sizeis the number of live births. Reported number is the number of individualsreporting their mother having each family size. Weighted number is the reported number divided by family size.
have a ten times greater chance of being ascertained than individuals from sibshi s
Harpending and Dra er (1990) is a forma !Y - of size one. The technique described
ized method for weig R ting each observation such that, if ni is the number of informants with sibships of size i, i = 1,2,3 ,.., k, where k is the maximum family size, then the im- puted number of mothers mi can be esti- mated by
n i
i mi = -
The estimated frequency of each sibship size (i > 0) is thus
m i f . - - ' - C m ,
1
The estimated CFS of women having at least one live birth is then
C n i
i "=s i
The resulting imputed number of mothers havin various family sizes is given in the
used to generate the imputed (indirect) fam- ily size distribution in Figure 4, which also shows for comparison the directly obtained family size distribution already given in Fig- ure 3. The indirect estimate closely approxi- mates that of the direct estimate. The CFSs of women having at least one live birth are also similar, = 3.9 for the indirect esti-
The advantage of computing estimates of average family sizes indirectly is that it al- lows for inferences to be made about events further in the past. The average birth year of the mothers of informants was 1907 com- pared to 1925 for the fertility histories of the postreproductive women, making them 18
secon ! row of Table 3. These data have been
mate and 4.0 ? or the direct estimate.
years older on average. Fertility is of consid- erable interest among Herero, and infor- mants res onded with confidence when
That the inferences made from these indirect estimates are based on data that are not gross1 in error is su ported by the fact that
women who had been independently inter- viewed. Both answers matched exactly. Her- ero also tend to be knowledgeable about their mothers' pregnancy wastage.
Because it is of interest to know the antiq- uity of low fertility among the Herero, these data were examined to identify temporal changes in fertility by dividing the data into two maternal birth cohorts. Information about parit of mothers who were born prior
after 1907 was partitioned so as to divide the number of informants reporting on their mothers' fertility exactly in half. The num- ber of informants re orting coming from each sibship size and t K e resulting weighted number of mothers by cohort is given in Table 4. The estimate of the average famil size of mothers who had at least one birt and the standard error (using the formula in Harpending and Draper, 1990) in the older cohort is 4.7 -C 0.70 compared to only 3.3 2 0.35 in the ounger cohort. The un- weighted average 8 irth years of mothers in each cohort were 1891 (ranging from 1870 to 1906) and 1922 (ranging from 1907 to 1941) so that the older set of mothers were, on the average, born 29 years earlier. The indirect estimates of CFSs compare with means of 3.0, 4.0, and 5.3 for the direct estimates of women born in the periods 1894-1921, 1922-1931, and 1932-1941, respectively. It ap ears that fertility was higher among the
creased a ain. If the trend in the data is
greatest impact among women born in the two decades after the turn of the century. This implies that the cause of low fertility may have its roots early in this century.
reporting t R eir mothers' number of births.
14 in F ormants supp[ed information about
to 1907 an dy of mothers who were born in or
E
01 (P er mothers, bottomed out, and then in-
genuine, t a en the cause of infertility had its

INFERTILITY IN HERERO PASTORALISTS 143
Fromwomen "'- I
0.05
Number of Live Births
Fig. 4. Imputed family size frequencies. Frequency of directly obtained family sizes are shown for comparison.
TABLE 4. Weighted number of mothers by birth cohort'
Family size 1 2 3 4 5 6 7 8 9 10 11 12 13
Mothers born in 1870-1906
Observed number 0 4 2 3 6 8 5 3 2 2 0 0 0 Weighted number 0 2 0.67 0.75 1.20 1.33 0.71 0.38 0.22 0.20 0 0 0
Mothers born in 1907-1941
Reported number 4 3 4 5 2 4 4 3 0 2 2 0 2 Weighted number 4 1.50 1.33 1.25 0.40 0.67 0.57 0.38 0 0.20 0.18 0 1.15
'Family sizes are reported by offspring. Family size is the number of live births. Reported number is the number of individuals reporting their mother having each family size. Weighted number is the reported number divided by family size.
It is ossible that the difference in fertility
an artifact of disruption from the Herero- German war or to differences in the quality of life in Namibia and Botswana. All but a few women in both cohorts, however, either entered or spent the majority of their child- bearing years in Botswana so that the poten- tial effects of these factors are probably small in this sample of women.
Parity progression ratios The family size distributions obtained
both directly and indirectly were used to com Ute parity progression ratios (PPRs). Eacg PPR i, i = 0, 1 , 2 ,.., k-1, where k is the maximum family size, is the pro ortion of
levels 1 etween the two cohorts of mothers is
women who, having had an i th birt R , have an
i + 1 th birth. The two sets of parity ro res- sion ratios are shown in Figure 5. $he%PR for i = 0 is about .85, indicating that about 85% of all women have a first birth, or, conversely, that 15% never have at least one birth. The rest of the PPRs are around .75, indicating that about 25% of women become sterile after each birth until it is zero at the maximum family size. Since the number of women who have zero births cannot be esti- mated indirectly, the indirect parity rogres-
indirect estimates are close at lower parities, further indicating that the attern of fertil-
about the reproductive behavior of Herero women. The overall shape of the curves is
sion ratios starts at i = 1. The PPRs f! rom the
it is similar for both sets o P data. The shape o P these curves also indicates something

144
0.7- 0.8-
0.6-
R. PENNINGTON AND H. HARPENDING
~
0.9 ‘1
7 tii 0.5 Indirect a: L 0.3 “‘1 0.21
0 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 Progression
Fi,y 5. ,Parity progression ratios of Herero women. Direct is the PPR computed from 239 ferti ity histories. Indirect is the PPR of the im uted parity distribution estimated from 70 informants reporting on their mothers’ number orbirths.
concave downwards, a sha e characteristic
1979). The shape of the PPR curve in contra- cepting populations tends to be concave up- wards due to most women terminating re- production at lower parities so that PPRs decline more rapidly. It is not surprising to find the concave downwards shape of the parity progression ratios since few deliber- ately control their fertility, and in fact there would be some concern with the quality of the data were this pattern absent. Charac- terizing the Herero as a natural fertility
opulation is useful, however, because it can
roductive behavior are
of natural fertility popu r ations (Howell,
ge assumed that factors
rated with the number already have.
Discussion Demographers have been aware of sub-
stantial variation in levels of fertility in African census and survey data for several decades. Coale (1968) observed a “high fertil- ity rid e” and a low fertility region in popu-
which total fertility rates ranged from 3.5 to more than 8. Overall, TFRs for the sub- Saharan countries he included (which did not include southern Africa) were 6.1-6.2. However, the distribution was patchy,
lation % ata from the 1950s and early 1960s in
having regions with very high fertilit neighboring those with much lower ferti P - ity. Romaniuk (1968) examined underlyin regional variation that is lost in nationa level opulation data. Both Romaniuk (1968)
of low fertility, such as marriage patterns, induced abortion, contraception, malnutri- tion, and disease, and found venereal disease to be an outstanding correlate of subfertility. Many of these data have been reworked in more recent surveys on infertility by Frank (19831, Caldwell and Caldwell (1983), and Doenges and Newman (19891, and all impli- cate venereal disease as a causal agent.
Interestingly, Adadevoh (1974) dismissed low fertility in Botswana as being due to emigration of males seeking employment in other areas. Unlike the Tswana, however, few Herero have worked in the South African mines. Demographers have also attributed low fertility rates re orted by older women to
some of their children, especially if the childred died young (Brass, 1968). The pat- tern of low fertility among Herero, however, is better described by disease rather than misreporting. There is no evidence in these data indicating underreporting of deceased children or to suggest memory lapse among older women.
The low rates of fertility and the high
1 and R dadevoh (19741 considered correlates
their “forgetting” t R at they gave birth to

INFERTILITY IN HERERO PASTORALISTS 145
proportion of childless postreproductive women suggests that Herero are afflicted with the same infectious infertility charac- terizing other African populations. In the absence of contraceptive use and pathologi- cal causes, childlessness rates of about 3% are considered normal (Bongaarts and Pot- ter, 1983). Although about 15% of postrepro- ductive Herero women never had a live birth, this is a low childlessness rate compared to many of the low fertility populations de- scribed by Frank (19831, in which rates were as high as 30-40%. However, many of the postreproductive Herero women terminated childbearin early in reproduction (Pen- nington an Har ending, 1990) so that over
Herero total fertility rates are among the lowest reported. The increase in the rates began in the period 1957-1966 at about the same time mobile dispensaries began visit- ing northwestern Botswana (Dam and Mo- losiwa, 1987). Toda there are permanent
workers in the few large villages of Ngami- land.
Accepting venereal disease as the major cause of low fertility rates among Herero, fertility began increasing in the 1960s as a result of fewer women becoming sterile from infection, rather than as a result of women becoming more fertile. Especially before the appearance of penicillin-resistant strains of venereal disease, administration of antibiot- ics at dispensaries would drastically reduce the incidence of pathological sterility result- ing from the se uelae of pelvic infections among women. Zonsequently, women who became sexually active after about 1960 (those born after about 19451, when treat- ment became available, would be at lower risk of becoming infertile than women born before then, since they are the first cohort to have access to treatment. Those who became menopausal before 1960 (those born before about 19151, assuming that the effect the sterilizing disease is long-standing, would be the most severely affected since the have been at risk throughout their repro B uctive span. Women born after about 1915 would have some advantage since they were able to seek treatment and, if they have not already become sterile, have less of a chance of be- coming so before menopause.
The continued increase in age-specific fer- tility at younger ages after 1960 can be at- tributed to increasingly more women seeking treatment for infection and in-
50% of them t d p ha two or fewer births. Thus,
medical clinics staf P ed with trained health
creases in the availability of health services in rural Botswana (Ministry of Health, 1982). The higher fertility of the youngest cohort of postreproductive women is consis- tent with the suggestion that women repro- ducin in the 1960s would benefit from the intro B uction of antibiotics. Few women born before 1921 would benefit from access to the dispensaries introduced in the 1960s, and these women have si ificantly lower com-
cohorts of postreproductive women. The an- tiquit of infectious infertilit among Herero
fertility obtained indirectly from offspring. However, fertility among the oldest mothers of informants appears to be higher than the fertility of younger women, suggesting that the cause of sterility may have been intro- duced around the turn of the century.
DISEASE AND FERTILITY Disease can affect fertility by causing
sterility or by lengthening birth intervals. Birth intervals can be lengthened by in- creases in pregnancy wastage or by lowered fecundability. The effect of infections such as tuberculosis (TB), malaria, filariasis, schistosomiasis, and sexually transmitted diseases (STDs) on fertility are described in McFalls and McFalls (1984), and their epide- miology in sub-Saharan Africa has been re- viewed by Belse (1976). Among these, pelvic
coccal, chlamydial, and other lower genital tract infections, and syphilis stand out as major correlates of low fertility in sub-Sa- haran Africa (Belsey, 1976; Muir and Belse ,
1983; Mabey et al., 1985; Cates et al., 1988). Other infections appear to have either smaller effects on fertility, have a low known incidence in sub-Saharan Africa (McFalls and McFalls, 1984; Belse , 19761, or have
Falls and McFalls, 1984). The potential im- pact on fertility of those diseases occurring in Botswana are considered below. The correla- tion between the diseases and the attern of
intervals) is discussed.
pleted family sizes t r an the two younger
is evi B enced by the similarly P ow estimates of
inflammatory B isease resulting from gono-
1980; Frank, 1983; Caldwell and Caldwe Y 1,
been ignored in studies o P subfertility (Mc-
Herero fertility (early sterility and P ong birth
Tuberculosis Genital TB occurs when the bacillus Myco-
bacterium tuberculosis (though sometimes Mycobacterium bovis or other bacteria are involved) s reads from its site of primary infection (t K e lung in the case of M. tubercu-

146 R. PENNINGTON AND H. HARPENDING
Zosis) through the lymph and vascular sys- tems into the reproductive organs (McFalls and McFalls, 1984). Sterility in females re- sults from the bilateral formation of tuber- cles in the Fallopian tubes. These tubercles may impede movement of germ cells through the tubes making conception and implanta- tion improbable. In severe cases, genital TB results in total tuba1 closure. Since genital TB is frequent1 symptomless, the pro or-
unknown. Most estimates are based on the number of women reporting to infertility clinics who are diagnosed as having genital TB so that the prevalence in the eneral
time scale necessar for TB to rogress to
1 een established. However, some research- ers have argued that genital TB may have a significant im act on reproductive perfor-
ample, are convinced that genital TB is largely responsible for low fertility among Black Americans earlier in this century rather than venereal disease as su gested
1984). In any case, only a fraction of TB infections
affects women of reproductive age or younger, and then only a fraction of these cases affects the genitals so that the inci- dence of TB must be much higher for the enital form to effect fertility at a PO ulation
Botswana for many years, its antiquity is unknown. The prevalence of the disease amon Herero is also unknown. In Ngami- land, $ B is the major cause of institutional deaths (those in hospitals and other health clinics), although a significant pro ortion of
BCG vaccine (Ministry of Health, 1982). Twenty ears a 0, TB was quite prevalent among ! h n g (8ruswell and Hansen, 1976, p. 169). Howell (1979, p. 63)found that about 56% of !Kung tested positive for TB infection and that TB was a major cause of death among them. However, during field work among the same !Kung population in Ngam- iland, one of us (H.H.) found a lower fre- quency of !Kung testing positive for TB, although TB was common among the !Kung living in Ghanzi. Due to their higher stan- dard of living, infection rates of Herero are probably much less than those of the impov- erished !Kung. During the field work for this study, both investigators heard of only a few Herero who had been afflicted with TB in the
tion of cases of J B that affect the genita P s is
population is unknown. Additional 9 y, the
enital infection an B cause steri P ity has not
mance. McFal P s and McFalls (19841, for ex-
by others (Farley, 1970; Wright an i Pirie,
fevel. Although TB has been en c f emic in
children are now being inoculate LQ with the
past. In Namibia, O’Keefe and colleagues (1988) found that just under 70% of !Kung patients had TB infections, com ared to less
live among Herero, who employ many of them as cattle hands.
In sum, it is unlikely that the frequency of TB among Herero is high enough to explain much of their low levels of fertility. More- over, hi h rates of TB infection reported
rather than decreases, in fertility among Herero, and TB is still endemic. TB is also not generally responsive to treatments avail- able at health posts, so there appears to be no mechanism by which changes in fertility would correspond to changes in the preva- lence of TB.
Malaria Pregnancy wastage may be induced by
malaria due to high fevers occurring durin
destruction. Malaria has been prevalent in northwestern Botswana for centuries and was remarked upon by Andersson (1987) during his 19th-century travels. However, malaria probably has had negligible measur- able effects on fertility among Herero for several reasons. First, most are only sporad- ically exposed to infection. The Anopheles mosquito that transmits infection to humans reproduces in pools of standing water. Since the semidesert environment west of the Oka- vango Delta occupied by Herero rovides a
rain season only, mosquito concentrations are 3: igh for only a few months per year. Given the wide variability in annual rainful in this area, risk of malarial infection varies considerably year to year. The mosquito breeding season is also truncated due to the coldest part of the year following the rains, cold bein nonconducive to mosquito repro- duction. 8 n the other hand, mosquitos are more like1 to persist in areas along the
and around Lake N ami, which is a d lake
the Okavango by way of the Thamalakane River. The Mbanderu heartland is Lake Ngami, so the risk of malaria infection among Herero may var geographically.
lengthen birth intervals among Herero ex- ists, malaria has ersisted throughout the decades in which P ertility has been increas- ing. Also, gven its seasonal pccurrence in Botswana, only a small fraction of women
that 10% among Herero. Most o P these !Kung
among t a e !Kung coincide with increases,
infection and to anemia subsequent to ce a
favorable mosquito environment : uring the
Okavango 6 elta, which is flooded annually,
bed that occasional 7 y fills from overf T ow of
While the potential P or pregnancy loss to

INFERTILITY IN HERERO PASTORALISTS 147
would be at risk during a given outbreak. The Herero also appear to have some resis- tance to malaria, once again, in comparison to the !Kung who frequently occup the same
malaria was epidemic among Herero and !Kung in the Dobe and Magopa areas of our study. Many more !Kung sought treatment from us at our camp for malaria symptoms than Herero living in the same area, and communication with the local dispensary in- dicates that Herero were not preferentially seeking treatment from the area medical clinic instead. It may be that the Herero style of dress affords them better protection against mosquito bites. Most women of reproductive age wear full-length 19th century-style dresses, underlayered with at least three petticoats. On the other hand, clothing may not explain variation in risk of malaria since men and children in the two groups do not contrast as sharply in their dress.
Sc his tosomiasis The impact of schistosomiasis on fertility
is not clear. Worm infection results from skin penetration of water-borne Schistosoma lar- vae, which undergo a portion of their life c cle in certain freshwater snails. It is t H ought that schistosomiasis may affect fer- tility by producing lesions in genital tissues. One of the best known studies of the involve- ment of the genital tract by Schistosoma comes from an autopsy study in Zimbabwe in which 50% of women were found to have lesions in elvic organs, most commonly the
McFalls (1984) believe that schistosomiasis probably has little effect on fecundity on the population level because most lesions found on reproductive organs do not appear severe enough to impair reproductive function. However, a study b Bullou h (1976) found
who were childless or suffering from second- ary sterility to be about twice the rate found in a group of control women. While the actual lesions may have a minimal effect on repro- ductive performance, these findings sug est
infection of the tubes, although schistosomal salpingitis is not unknown either.
In any case, Schistosoma infection ap- ears to be rare or unknown among Herero.
Howell (1979, . 65) reported that this dis-
neighbors in the Dobe area, where there are no permanent fresh water sources other
homesteads as Herero. In Marc E of 1988,
tubes (Ge P fand et al., 1971). McFalls and
the incidence of S. 1 % aemato ium in women
that schistosomiasis may promote secon c f ary
ease is unusua P among the Herero’s !Kung
than wells. Schistosoma have infested the Okavan o Delta, es ecially in Maun. Since
infection from this source is low. Further- more, Geldenhuys and colleagues (1967) found the prevalence of schistosomiasis to be relatively rare among all peoples of the Kala- hari. Consequently, it cannot account for the low levels of fertility observed among Herero.
Sexually transmitted diseases
Herero Yive away P rom the Delta, risk of
Pelvic inflammato disease. Pelvic inflam-
the occurrence of salpingitis and is often diagnosed in women reporting lower ab- dominal pain in association with cervical, uterine, and adnexal tenderness (Cates et al., 1988). Women suffering from PID have er thema, edema, and us or exudate in the
1969). Chlamydia trachomatis and Neisseria gonorrhea are two sexually transmitted bac- teria to which the majorit of cases of PID
(Belsey, 1976; Mabey, et al. 1985; Cates et al., 1987). Other bacteria have been impli- cated as well, but the possibility that these are secondary infections of C. trachomatis or N. gonorrhea has not been ruled out.
C. trachomatis and N . gonorrhea cause PID by ascending from the lower genital to upper genital tract through the cervix. The disease is self-limiting, but damage to the reproductive tract results from scarring of one or both Fallopian tubes during postinfec- tion healing. These scars can impede tubal motility or damage tubal mucosa and cilia, thereby interferin with the movement of
reducing the likelihood of fertilization. These same factors impede the transfer of zygotes to the uterus and in fact increase the probability of ectopic regnancy in women
occlusion can result from a single PID epi- sode with the risk of occlusion increasing with repeated infection. Although infection can occur in only one Fallopian tube, bilat- eral infection is normal so that bilateral occlusion and infertility appear to be the ultimate outcome of repeated infection. Some evidence indicates that PID may cause ovarian failure such as through thickening of the ovarian ca sule, thereby reventing re-
Although the fertility prognosis of PID epi- sodes is good for early treatment, once scar- ring or occlusion have occurred phar-
matory disease (P Y D) generally refers to
fa i lopian tubes (Jaco I& son and Westrom,
have been attributed in su i -Saharan Africa
gametes through t a e reproductive tract and
with PID (Muir and f; elsey, 1980). Tuba1
lease of ova ( K cFalls and d cFalls, 1984).

148 R. PENNINGTON AND H. HARPENDING
macological treatment of PID a parently
The frequency of C. trachomatis- and N . gonorrhea-associated PID episodes are high- est at the beginning of the menstrual cycle (Cates et al., 1988). The biotic environment of the lower genital tract may favor prolifer- ation of unfavorable organisms whose chances of entering the upper re roductive
cervical mucus plug, which is thought to act as a barrier against disease, during menses. The opportunity for infection may also be greater during menses due to the flux of menstrual blood through the Fallopian tubes and to greater vulnerability of tissue ex- posed by the sloughing of the endometrium. Consequently, variation in the length of menses and the frequenc of cycles may be
vi uals and opulations in which these fac-
by later sexual maturity and prolonged post- partum amenorrhea may also be character- ized by a lower incidence of PID than where menses and sexual activity follow parturi- tion more quickly. Since women are amenor- rheic during pregnancy, pregnancy would seemingly provide an additional protective component against PID. Several studies show, however, that a high percentage of women who have chlamydia1 or gonorrheal infection present in the endocervical canal experience PID episodes following parturi- tion.
Syphilis. The effects of syphilis on fertility in sub-Saharan Africa have received less attention. The fact that sy hilis increases
rility and because it is largely a self-limiting disease with respect to fertility may be why it has received less attention.
Syphilis is caused by Treponema dum. Fetuses of women afflicted wit pri- mary or secondary s philis have a hi h
tus may be either aborted, still born, or live born having congenital syphilis. The out- come of pregnancy has been related to the stage of maternal infection. Infected moth- ers who conceive in the earl sta e of syphilis
chances of bearing a live-born child, either with or without con enital syphilis, increase
Syphilis becomes latent in most women after about two years, after which the chances of bearing healthy children are then quite
does not improve reproductive per P ormance.
tract may be greater due to the P oss of the
im ortant correlates of PI 6 risk among indi-
tors B E vary. T us, populations characterized
pregnancy wastage rather t K an causing ste-
tallC-
chance of becoming in P ected. An infected B e-
are most likely to abort t i i f eir etuses. The
with the duration o B infection in the mother.
good. Untreated primary or secondary syph- ilis tends to produce immunity to reinfection. Reinfection is most likely in individuals who receive early treatment, but the chances of reinfection diminish the later treatment oc- curs. In sum, syphilis can be an important cause of pregnancy loss, but its effect is limited to only one or two pregnancy losses in most women.
Prevalence of sexual1 transmitted diseases
s phiyis are widespread throughout Africa. gnce these infections are transmitted in the same way, they tend to be revalent in the
effects of one from another. Moreover, in Africa, both aws and endemic syphilis are
nonvenereally but are caused by Tre onema
tinguishable from T. pal f idum. Measuring the prevalence of venereal s philis where
difficult. Yaws was noted by early 16th- century explorers in Africa, about the same time venereal syphilis became imported from its probable origin in the New World, but the relationships of these diseases are unclear. All three (yaws, endemic syph- ilis, and venereal syphilis) cause cross- immunity, and since yaws and endemic syphilis are childhood diseases, venereal syphilis does not become prevalent in areas where the other Treponema prevail. How- ever, since the yaws eradication campaigns in the 1950s (Guthe et al., 1972), the inci- dence of syphilis has increased. At the same time, infectious infertility has decreased in these areas as a secondary effect of mass panicillin in'ections.
(Manson-Bahr, 1950; Hunter et al., 19761, while endemic syphilis is most common in arid regions (Hunter et al., 1976). Neither Manson-Bahr (1950) nor Hunter et al. (1976) include Botswana in the laces they mention
syphilis was seen there in children in the 1930s and 1940s (Kuczynski, 1949) suggest- ing nonvenereal transmission of the disease there. Guthe and colleagues (1972) reported that nonvenereal syphilis was endemic along the eastern edge of Botswana and in south- eastern Namibia along the southwestern border of Botswana. The Kgalagadi in Botswana are said to have a high prevalence of endemic syphilis, but no figures were available (Murray et al., 1952, Murray, 1957,
amon Herero. Gonorr l ea, chlamydia, and
same places, making it dif P icult to sort out
prevalent. T K ese diseases are transmitted
either serologically or mor hological P y indis-
there is yaws or endemic syphi P is is therefore
Yaws ten d s to be found in the moist tropics
as having yaws or en dp emic syphilis, but

INFERTILITY IN HEAERO PASTORALISTS 149
cited in Nurse and Jenkins, 1977). Endemic sy hilis was also observed among G/wi and Gfana Bushmen (Nurse et al., 1973) but apparently not among !Kung. Based on a response from the Hi h Commissioner in Pretoria, Hacket (19537 reported that aws
rica, although it was known to occur among miners.
Of the ethnic groups in which either of these diseases are known to be prevalent, none live in close proximity to Herero. The revalence of both yaws and endemic syphi- F is have been correlated with environmental
factors, such as poor hygiene, which may explain why it is found in some ethnic groups but not others. Today, venereal syphilis is widespread in Botswana, but the antiquity of syphilis and other venereal diseases among Herero is unknown. Both Vedder (1966a) and Irle (1906, cited in Poewe, 1985) suggest that the Nama may have introduced venereal sy hilis to the Herero tribes in
ties (see Pennington, 1990a). In 1928, Ved- der (1966a, p. 204) noted the opularit of
b sufferers of venereal diseases and by
voted an essay on the causes of low fertility among Herero in which venereal diseases were considered an important correlate. Gib- son (1959) also noted a shortage of children in the population yramid constructed from
of the small Sam le size, Gibson was reluc-
lation pyramid was a result of low ferti ity. Government documents cited in Kuczyn-
ski (1949) indicate that venereal diseases were clearly widespread in Botswana in 1940. During field working in Botswana 1968-1969, one of us (H.H.) noted that Her- ero women frequently complained of pelvic pain and while men frequently asked for treatment of symptoms of gonorrhea. During our field work in 1987-1989, we heard com-
laints from only a few women who typically Rad already sought treatment for their symptoms from medical clinics. Conse- quently, it appears that most Herero are seeking treatment for venereal infections before they cause severe damage. Today, PID is included in the Medical Statistics report and thousands of cases of gonorrhea and syphilis were reported (Ministry of Health, 1982). In sum, there is evidence of a long history of both syphilis and gonorrhea
was not endemic in Botswana or Sout i Af-
about 1850 a wing the Herero-Nama hostili-
“witchdoctors” who were most P y “consu 9 ted
c 3; ildless women.” Steenkamp (1944) de-
a survey of Ngami P and Herero, but, because
tant to conclude t K at his narrow based opu- P
among Herero. Most certainly other sexually transmitted genital tract infections that could lead to PID are prevalent among them as well.
Discussion Of the diseases considered, PID roduces a
pattern of fertility consistent wit K the pat- tern observed arnon Herero. Although we
the major cause of infertility among women seeking medical treatment for infertility (Belsey, 1976; Mabey et al., 1985; Cates et al., 1987). Some researchers are skeptical of accepting that venereal diseases are res on-
the African continent and point out that high frequencies of venereal infections do not al- ways correspond to low fertility (Belsey, 1976; Caldwell and Caldwell, 1983; Doenges and Newman, 1989). However, current rates of venereal infection do not necessarily re- flect the risk of women who became sterile in the past. Also, areas reporting high preva- lences of venereal diseases tend to be areas where women are being treated for them. STDs do their dama e when they are not
expected to be greatest where medical treat- ment is lacking.
ECOLOGICAL FACTORS In addition to disease, ecological variables
have also been proposed as constraints on fertility. The critical fat hypothesis (Frisch and McArthur, 1974; Frisch, 1978) sug- gested that a minimum fat store was neces- sary for the onset of menarche and for maintaining menstrual cyclin . Under this
turition and during lactation among women in mar ‘nally nourished populations would cause f l elayed ovulatory cycling and longer birth intervals. Longer birth intervals result in lower fertility since fewer births can occur in the re roductive span. However, this hy-
Rterature (c.f. Johnston et al., 1975; Huff- man et al., 1978; Huss-Ashmore, 1980; Bon- gaarts, 1982). In any case, undernutrition is not likely to be a factor affecting the Herero, who frequently suffer from obesity (O’Keefe et al., 1988).
Breastfeeding patterns are also known to account for variation in birth intervals and, therefore, variation in levels of fertility be- tween populations. Frequent and intense
lack su porting me C F ical evidence, in other areas o F Africa PID has been implicated as
sible for the low levels of fertility throug : out
treated, and their e B fects on fertility are
hypothesis, depleted fat stores !! ollowing par-
othesis R as not found much support in the

150 R. PENNINGTON AND H. HARPENDING
suckling is thought to reduce secretion of gonadotropins, inhibiting follicular develop- ment (McNeilly et al., 1988). Previously, lac- tation was thought to su press cycling by causin circulation of high f evels of prolactin
al., 1986; McNeilly et al., 1988). The end result is that women who breastfeed their babies frequently and longer postpartum have prolonged birth intervals and, there- fore, have lower fertility than women whose infants suckle less intensively. The contra- ceptive effect of lactation varies but lasts several months. One of the longest delays associated with breastfeeding patterns on cycling has been found among the Gainj of New Guinea, where the median duration of anovulatory lactation was 20 months (Wood et al., 1985).
A population-wide reduction in the fre- uency and intensity of breastfeeding has
(Knodel, 1977; Bongaarts, 1981). Careful ob- servations on Herero breastfeeding were not made, but there are no obvious behavioral indications that would suggest that Herero are carin for infants differently than they
were ever observed bottle feeding their in- fants; one claimed her breasts did not pro- duce enough milk to satisfy her baby and the other was feeding her newborn triplets. Mothers carry their infants tied to their backs throughout the day; they remove the infants and offer them their breast whenever they become restless. Herero did not express strong beliefs about weaning children, but both youn and old women remarked that it
after about one year of age. These women said that a child should be weaned if his or her mother becomes pregnant because they consider breastmilk harmful to the nursing child. A large proportion of children are weaned in the second year of life when they are fostered out to relatives (Pennington, 1990b). We noted that supplementary foods, such as corn porridge, were commonly of- fered and accepted by infants as young as three months of age. While the on-demand style of feeding is consistent with the type thought to maximize the contraceptive effect of breastfeeding, this effect would be less- ened by the early introduction of supplemen- tary foods. Nevertheless, most women in this study, regardless of age, did not have subse- quent births until at least two years postpar- tum (Pennington, 1990a).
The !Kung are interesting to contrast with
(e.g., 2 onner and Worthman, 1980; Stern et
t a e potential to cause an increase in fertility
have in t a e past. Only two Herero women
was consi f ered correct to wean children only
the Herero because they live in the same ecological zone but are ethnically and demo-
aphically very different. The !Kung are Eown for their long birth intervals (having a mean interval between live births of 35.4 months) and may breastfeed their children for as long as five years (Howell, 1979). Because of the association between pro- lon ed breastfeeding and delayed ovulatory
unusual !Kung reproductive pattern have been proposed. It has been hypothesized that !Kung women breastfeed longer due to short- ages of suitable bush food for weanin or
Lee, 19797. &hers have suggested that !Kung who space their births widely have increased reproductive success because they have reduced mortalit among their oung
Jones, 1986). Pennington and Harpending (1988), however, found no association be- tween the fertility of !Kung women and the survivorship of their offs ring.
edly contributes to wide birth s acing among
ship between lactation and long birth inter- vals is obscured in longer intervals. For example, a !Kung child is usually weaned once his or her mother recognizes her next pregnancy (Howell, 1979; Konner and Worthman, 1980). Thus, women with low fecundability wean their children at older ages so that rolonged breastfeeding ma be
Harpending and Dra er (1990) have argued
were afflicted with athological infertility. Interestingly, Howel P (1979) considered but dismissed venereal diseases as a complete explanation for the low levels of fertility she observed among !Kung.
Maternal de letion models have also been
strain female fertility. For example, women who have closely spaced births are thought to become nutritionally depleted because they are unable to recover maternal re- sources between regnancies (e.g. Winikoff, 1983; Pebley a n t DaVanzo, 1988; Winikoff and Castle, 1988). Women with the highest parity at the youngest ages living in the most marginal environments are most affected. The significance of these models in a human evolutionary framework is that depletion is thought to lead to lower survivorship of mothers and their offspring. But support for
cyc F ing, a variety of explanations for the
because women could carry on1 one in f ant on foragin tri s into the bush ( i5 leek, 1928;
(Blurton Jones and Jibly, 1978; B r" urton
Although prolonged P actation undoubt-
!Kung, the causal direction o P the relation-
an e€fect rat i er than a cause of low fertigty.
that the !Kung, like t K eir Herero neighbors,
proposed in w K ich ecologcal variables con-

INFERTILITY IN HERERO PASTORALISTS 151
these models in this context is contradictory, partly due to methodological problems. Most studies measure nutritional status usin an-
it is assumed that women with less fat have lower nutritional status. Some studies have found that women with higher parity are more depleted than lower parity women of the same age, but others have not. Interpre- tation of these results also hinges on the correlation between amounts of subcutane- ous fat and health, although more recise
theless, results are often inconclusive be- cause heterogeneity in offsprin mortality
guishable from the effects of maternal de le-
deaths may also shorten birth intervals be- cause they remove the contraceptive effect of lactation; consequently, separatin the ef-
vival is com lex. In fact, few Herero are probably un ! ernourished (OKeefe et al., 1988) and experience heterogeneity in the survivorship of their offspring (Harpending and Pennington, 1991).
CONCLUSION This paper has documented unusually low
rates of fertility among Herero. Throughout the first half of this century, many women apparently ex erienced truncated re roduc-
rates have increased from a low of 2.65 to 7.02. The increases in fertility coincide with the appearance of rural health posts in northwestern Botswana. After considering the effects of various diseases, it appears that the low fertility rates of Herero are most likely the result of pelvic infections in women.
The importance of ecolo ical and behavior factors, such as breastfee fing practices and nutrition, on Herero fertility are discussed. While these factors undoubtedly account for some of the variation in fertility among Her- ero women, their importance is overshad- owed by the apparent effect of sterilizing disease. It has been argued that nutritional stress has played an important role in the evolution of human reproductive patterns. This study suggests, however, that the ef- fects of disease alone may have a far greater impact on the fertility of women than the effects of poor nutritional status. The possi- bility that correlates of maternal nutritional status are spurious relationships due to het-
thropometric measures of fat stores in w a 1ch
measurements are coming into use. h ever-
among high parity mothers may fJ e indistin-
tion (Winikoff and Castle, 1988). In P ant
fect of short birth intervals on in f ant sur-
tive careers. 2 ince about 1960, total P ertility
erogeneity among women in poor health merits further consideration. This is es e-
correlations between parity and fertility run in the same direction, whether the rimary
words, a woman's chances of becoming sub- fertile due to disease increase with parity while her maternal energy stores decrease with parity.
Finally, analyses of Herero fertility and mortality suggest that increases in health care may have a more substantial effect on fertility than mortality. Among Herero, mi- nor improvements in health care have re- sulted in a nearly threefold increase in the birth rate, while analysis of mortality among Herero indicate that survivorship has in- creased by only a few ears (Harpending and Pennington, 1990). %'hese trends suggest that decreases in sterility rather than in- creases of survivorship are responsible for transforming the African continent into one of the fastest growing regions in the world.
ACKNOWLEDGMENTS We thank Kenneth Weiss, James Wood,
Clifford Clogg, Jeffrey Kurland, Alan Rog- ers, and Albert0 Palloni for commenting on the manuscript. Project field work was sup- ported by the Wenner-Gren Foundation for Anthro ological Research (RP), the Hill
(RP), and National Institute on Agin grant AGO3110 (HH and RP) to Christine f r and Jennie Keith. We are grateful to the Ofice of the President of the Republic of Botswana for ermission to carr out this research and to
Museum for support and encouragement.
cially important in light of the fact t K at
cause is disease or undernutrition. P n other
Fund o k? The Pennsylvania State University
;F . Mpulubusi an d others at the National
REFERENCES Adadevoh BK (ed.) (1974) Sub-fertility and Infertility in
Africa. Ibadan, Nigeria: Claxton Press. Almagor U (1980) Some notes on the Mbanderu calen-
dar. Botswana Notes and Records 12:67-82. Almagor U (1982) A note on the fertility of the Mbanderu
women. Botswana Notes and Records 14:23-25. Alnaes K (1989) Living with the past: The songs of the
Herero in Botswana. Africa 59:267-299. Anderson DR, Sweeney DJ, and Williams TA (1981)
Introduction to Statistics: An Applications Approach. St. Paul: West.
Anderson CJ (1987) Lake Ngami, 2nd ed. Capetown, South Africa: C. Struik.
Belsey MA (1976) The epidemiology of infertility: A review with articular reference t o sub-Saharan Af- rica. Bull W 8 0 54:319-341.
Bleek D (1928) The Naron: A Bushman Tribe of the Central Kalahari. Cambridge: Cambridge University Press.

152 R. PENNINGTON AND H. HARPENDING
Bley H (1971) South-west Africa Under German Rule, 1894-1914. Evanston: Northwestern University Press.
Blurton Jones N (1986) Bushman birth s acin A test for optimal interbirth intervals. EtKol gociobiol
Blurton Jones N and Sibly RM (1978) Testing ada tive ness of culturally determined behaviour: Do Buslman women maximize their reproductive success by spac- ing births widely and foraging seldom? In Blurton Jones N and Reynolds V (eds): Human Behaviour and Adaptation. London: Taylor and Francis, pp. 135-155.
Bongaarts J (1981) The impact on fertility of traditional and chan ‘n child spacing ractices In Page H J and Lesthaegfe 8 (eds.;: Child-&acing in Tropical Africa. London: Academic Press, pp. 111-129.
Bongaarts J (1982) Does malnutrition affect fecundity? A summary of the evidence. Science 208:564-569.
Bongaarts J and Potter RG (1983) Fertility, Biology, and Behavior: An Analysis of the Proximate Determi- nants. New York: Academic Press.
Brainard J (1986) Differential mortality in Turkana agriculturalists and pastoralists. Am J Phys Anthro-
Brass WI (1968) The use of existing data. In Caldwell JC and Okonjo C (eds.): The Population ofTropical Africa. New York: Columbia University Press, pp. 172-178.
Bridgeman JM (1981) The Revolt of the Hereros. Berke- ley: University of California Press.
Bullou h CHW (1976) Infertilit and bilharziasis of the femaye genital tract. South d Med J 83:819-822.
Caldwell JC and Caldwell P (1983) The demographic evidence for the incidence and cause of abnormally low fertility in tropical Africa. World Health Stat Q 36: 2-34.
Caldwell JC and Caldwell P (1987) The cultural context of high fertility in sub-Saharan Africa. Popul Dev Rev 13:409-437.
Campbell KL and Wood JW (1985) Fertility in tradi- tional societies. In Diggory P, Potts M, and Teper S (eds.): Natural Human Fertility. Houndmills, En- gland: MacMillan, pp. 39-69.
Cates W, Aral SO, and Rolfs RT (1988) Patho hysiology and epidemiology of sexually transmitted {iseases in relation to pelvic inflammatory disease and infertility. International Union for the Scientific Study of Popu- lation, Johns Hopkins University.
Cates W, Farley TMM, and Rowe PJ (1987) Infections, pregnancies, and infertility: Perspectives on preven- tion. Fertil Steril47:964-968.
Central Statistics Office (1981) Population and Hous- ing Census, Republic of Botswana. Census Admin- istrativemechnical Report and National Statistical Tables. Gaborone, Botswana: Central Statistics Of- fice.
Central Statistics Office (1988) Statistical Bulletin. Vol. 13, no. 1. Gaborone, Botswana: Central Statistics Of- fice.
Coale AJ (1968) Estimates of fertility and mortality in tropical Africa. In Caldwell JC and Okonjo C (eds.): The Population of Tropical Africa. New York: Colum- bia University Press, pp. 179-186.
Crawford MA (1980) Lipid requirements during preg- nancy and lactation. In Santos W, Lopes N, Barbos JJ, and Chaves D (eds): Nutrition and Food Science: Present Knowledge and Utilization (Vol. 3): Nutri- tional Biochemistry and Pathology. New York: Ple- num Press, pp. 37-42.
Dam CJ van and Molosiwa K (1987) A control pro-
7:91-105.
pol 70:525-536.
gramme for sexually transmitted diseases in Botswana. Proposal to the Ministry of Health, Botswana, July.
Doenges CE and Newman J L (1989) Impaired fertility in tropical Africa. Geog Rev 79:99-111.
Farley R (1970) The Growth of the Black Population. Chicago: Markham.
Frank 0 (1983) Infertility in Sub-Saharan Africa: Esti- mates and implications. Popul Dev Rev 9:137-144.
Frank 0 (1987) The demand for fertility control in sub-Saharan Africa. Stud Fam Plan 18:181-201.
Frisch R (1978) Population, food intake, and fertility. Science 199:22-30.
Frisch RE and McArthur JW (1974) Menstrual cycles: Fatness as a determinant of minimum wei ht for height necessary for the maintenance or onset. gcience 185:949-951.
Geldenhuys PJ, Hallett AF, Visser PF, and Malcolm AC (1967) Bilharzia survey in the eastern Ca rivi, north- ern Bechuanaland and northern South 6 e s t Africa. South Afr Med J 41:767-771.
Gelfand M, Ross MD, Blair DM, and Weber MC (1971) Distribution and extent of schistosomiasis in female pelvic organs, with special reference to the genital tract, as determined at autopsy. Am J Trop Med Hyg 20:846-849.
Gibson GD (1959) Herero marriage. Rhodes Livingstone J 24:l-37.
Guthe T, Ridet J , Vorst F, D’Costa J, and Grab B (1972) Methods for the surveillance of endemic treponema- toses and sero-immunolo ‘cal investigations of “disap- pearing” disease. Bull W E 0 46:l-14.
Hackett CJ (1953) Extent and nature of the yaws prob- lem in Africa. In First International Symposium on Yaws Control, World Health Organization (Mono- graph No. 15). Geneva: World Health Organization,
Harpending H and Draper P (1990) Estimating parity of parents: An application to the history of infertility among the !Kung of southern Africa. Hum Biol
Harpending H, Draper P, and Pennington R (1990) Cultural evolution, parental care, and mortality. In Swedlund A and Armelagos G (eds.): Health and Dis- ease in Transitional Societies. South Hadley, MA: Bergin and Garvey, pp. 241-255.
Harpending H and Pennin on R (1991) Age-structure and sex-biased mortafty amon Herero and Mbanderu peoples of the northern Kafahari. Hum Biol (in press).
Harpending H and Pennington R (1990) Product-limit estimates of adult survivorship among southern Afri- can pastoralists. Unpublished.
Harpending H and Wandsnider L (1982) Population structures of Ghanzi and Ngamiland !Kung. In Craw- ford MH and Mielke JH (eds.): Current Developments inAnthropologica1 Genetics. New York: Plenum Press,
Henin RA (1968) Fertility differentials in the Sudan (with reference to the nomadic and settled popula- tions). Popul Stud 22:147-168.
Henry L (1961) Some data on natural fertility. Eugen Q 8 3 - 9 1 .
Hill AJ, Randall SC, and van den Eerenbeemt ML (1983) Infant and child mortality in rural Mali (Research Paper No. 83-51 London: Centre for Population Stud- ies.
Howell N C1979) Demography of the Dobe !Kung. New York: Academic Press.
Huffman SL, Alauddin Chowdhury AKM, and Mosley
pp. 129-182.
62:195-203.
pp. 29-50.

INFERTILITY IN HE RERO PASTORALISTS 153
WH (1978) Postpartum amenorrhea: How is it affected by maternal nutritional status? Science 200:1155- 1157.
Hunter GW, Swartzwelder JC, and Clyde DF (1976) Tropical Medicine, 5th ed. Philadelphia: Saunders.
Huss-Ashmore R (1980) Fat and fertility: Demographic implications of differential fat storage. Am J Phys Anthropol23:65-91.
Hytten FE (1980) Nutritional aspects of human preg- nancy. In Aebi H and Whitehead R (eds.): Maternal Nutrition During Pregnancy and Lactation. Bern: Hans Guber, pp. 27-38.
Irle J (1906) Die Herero. Gutersloh, Germany: Bertels- mann.
Jacobson L and Westrom L (1967) Objectivized dia of acute pelvic inflammatory disease. Am J 8;;; Gynecol105:1088-1098.
Johnston FE, Roche AF, Schell LM, and Wettenhall HNB (1975) Critical weight at menarche: Critique of a hypothesis. Am J Dis Child 129:19-23.
Knodel J (1977) Breast-feeding and population growth. Science 198:llll-1115.
Konner M and Worthman C (1980) Nursing frequency, gonadal function, and birth spacing among !Kung hunter-gatherers. Science 207:788-791.
Kuczynski RR (1949) Demographic Survey of the British Colonial Empire. Vol. 11. London: Oxford University Press.
Lee R (19791 The !Kung San. Cambridge: Cambridge University Press.
Mabe DCW, Ogbaselassie G, Robertson JN, Heckels JE , andrWard ME (1985) Tuba1 infertility in the Gambia: Chlamydia1 and gonoccal serology in women with tuba1 occlusion compared with pregnant controls. Bull WHO 63: 1107-1 113.
Manson-Bahr PH (ed.) (1950) Manson’s Tropical Dis- eases. A Manual of the Diseases of Warm Climates. 13th ed. Baltimore: Williams & Wilkins Company.
Manyeneng WG, Khulumani P, Larson MK and Way AA (1985) Botswana Family Health Survey. Gaborone, Botswana: Ministry of Health.
McFalls JA and McFalls JH (19841 Disease and Fertility. Orlando: Academic Press.
McNeilly AS, Howie PW, and Glasier A (1988) Lactation and the return of ovulation. In Diggory P, Potts M, and Teper S (eds.): Natural Human Fertility. London: MacMillan, pp. 102-117.
MinistryofHealth(1982) Medical Statistics, 1982. Gabo- rone, Botswana: Central Statistics Office.
Muir DG and Belsey MA (19801 Pelvic inflammator disease and its consequences in the developing world: Am J Obstet Gynecol 7:913-928.
Newel1 C (1988) Methods and Models in Demography. New York: Guilford Press.
Nurse GT and Jenkins T (1977) Health and the Hunter- Gatherer. Monographs in Human Genetics, Vol. 8. New York: S. Krager.
Nurse GT, Tanaka N, MacNab G, and Jenkins T (1973) Non-venereal syphilis and Austrailian antigen among the Giwi and Giiana San of the Central Kalahari Reserve. Central Afr J Med 19:207-213.
O’Keefe SJD, Rund JE, Marot NR, Symmonds KL, and Berger GMB (1988) Nutritional status, dietary intake and disease patterns in rural Herero, Kavangos and Bushmen in South West Afr icaamibia . South Afr Med J 73:643-648.
Pa e H (1988) Fertility and famil planning in sub- ,faharan Africa. In van de Walle 8, Ohadike PO, and Sala-Diakanda MD (eds.): The State ofAfrican Demog-
raphy. International Union for the Scientific Study of Population, pp. 29-45.
Pebley AR and DaVanzo J (1988) Maternal depletion and child survival in Guatemala and Malaysia. Paper pre- sented at the 1988 Annual Meetings of the Population Association of America, New Orleans.
Pennington R (1990a) Fertility of the Herero and Mbanderu pastoralists ofBotswana. PhD dissertation, The Pennsylvania State University, University Park.
Pennington R (1990b) Fostering as a reproductive strat- egy among southern African pastoralists. Ethol Socio- biol (in press).
Pennington R and Harpending H (1988) Fitness and fertility in Kalahari !Kung. Am J Phys Anthropol
Pennington R and Harpending H (1990) The effect of infertility on the population structure of the Herero and Mbanderu of southern Africa. Social Biology (sub- mitted).
Poewe Ki1985) The Namibian Herero, a History ofTheir Psychosocial Disintegration and Survival. New York: Edwin Mellen.
Romaniuk A (1968) Infertility in tropical Africa. In Caldwell JC and Okonjo C (eds): The Population of Tropical Africa. New York: Columbia University Press, pp. 214-224.
Salzano FM (1971) Demographic and genetic interrela- tionships among the Cayapo Indians of Brazil. Social Biol18:148-157.
Serjeantson S (1975) Marriage patterns and fertility in three Papua New Guinean populations. Hum Biol 47:399%413.
Steenkamp WP (1944) Is the South-West African Herero Committing Race Suicide? Capetown: Unie- Volkspers.
Stern JM, Konner M, Herman TN, and Reichlin S (1986) Nursing behaviour, prolactin and postpartum amen- orrhoea during prolonged lactation in American and !Kung mothers. Clin Endo 25:247-258.
Truswell AS and Hansen JDL (1976) Medical research among the !Kung. In Lee RB and I DeVore (eds.): Kalahari Hunter-Gatherers. Cambridge: Harvard University Press, pp. 166-194.
Vedder H (1966a) The Herero. In Hahn CHL, Vedder H, and Fourie L ieds.): Native Tribes ofSouthwest Africa. New York: Barnes & Noble, pp. 153-211 (reprint of 1928 edition).
Vedder H (1966b) South West Africa in Early Times (translated and edited by CG Hall). London: Frank Cass & Co. (reprint of 1938 edition).
Winikoff B (1983) The effects of birth spacing on child and maternal health. Stud Fam Plan 14:231-245.
Winikoff B and Castle MA(1988) The maternal depletion syndrome: Clinical diagnosis or eco-demographic con- dition. Biol SOC 5:163-170.
Wood JW (1989) Fecundity and natural fertility in hu- mans. In Milligan S (ed.): Oxford Reviews in Biology (Vol. 11). Oxford: Oxford University Press, pp. 61-109.
Wood JW, Lai D, Johnson PL, Campbell KL, and Maslar IA(1985) Lactation and birth spacing in highland New Guinea. J Biosoc Science (Supplement) 9:159-173.
Way AA, Cross A, and Kumar S (1987) Family planning in Botswana, Kenya and Zimbabwe. Int Fam Perspect
Wright P and Pirie P (1984) A false fertility transition: The case of American Blacks. Pa ers of the East-West Population Institute, No. 90. Ronolulu: East-West Center.
77:303-319.
13:7-11.




















