Induction of CTL response by a minimal epitope vaccine in HLA A∗0201/DR1 transgenic mice:...
16
Induction of CTL Response by a Minimal Epitope Vaccine in HLA A*0201/DR1 Transgenic Mice: Dependence on HLA Class II Restricted T H Response Lbachir BenMohamed, Radhika Krishnan, Jeffrey Longmate, Catherine Auge, Ldito Low, James Primus, and Don J. Diamond ABSTRACT: CTL play a pivotal role in the immune response during viral infections. In this study, the HLA class II restricted T H requirement for optimal in vivo induction of HLA class I restricted CTL responses has been investigated. Towards this goal, transgenic mice expressing both HLA class I (A*0201 or A2.1) and class II (DRB1*0101 or DR1) molecules have been derived. Immunization of these mice with an HLA A*0201-re- stricted and CMV-specific CTL epitope (pp65 495–503 ), and either of three different tetanus toxin-derived MHC class II-binding T H epitopes, resulted in a vigorous CTL response. CTL specific for the pp65 495–503 epitope were dramatically enhanced in mice expressing both the HLA- DR1 and HLA-A*0201 transgenes. Notably, preinjec- tion of three TT peptides (TT 639 – 652 , TT 830 – 843 , and TT 947–967 ) increased the capability of HLA A*0201/DR1 Tg mice to respond to subsequent immunization with the T H 1 CTL peptide mixture. These results indicate that the use of HLA A*0201/DR1 Tg mice constitute a ver- satile model system (in lieu of immunizing humans) for the study of both HLA class I and class II restricted T-cell responses. These studies provide a rational model for the design and assessment of new minimal-epitope vaccines based on their in vivo induction of a pathogen-specific CTL response. Human Immunology 61, 764 –779 (2000). © American Society for Histocompatibility and Immu- nogenetics, 2000. Published by Elsevier Science Inc. KEYWORDS: T help; cytotoxic T lymphocyte; HLA- DR1; HLA-A*0201; transgenic mice ABBREVIATIONS PBMC peripheral blood mononuclear cells CRA chromium release assay IVS in vitro stimulation LN lymph nodes Tg transgenic TT tetanus toxin INTRODUCTION During the past decade, considerable interest has been focused on specifically inducing CTL to prevent and/or control chronic viral diseases. Among the numerous ap- proaches investigated, are epitope based studies using synthetic peptides derived from the processed form of viral proteins which bind to MHC class I alleles [1– 4]. Protective antiviral immunity using CTL epitope pep- tides has been achieved in several experimental models of infection [5–9]. The epitope-based approach offers sev- eral potential advantages. Selection of naturally processed epitopes may allow the immune system to focus on highly conserved and immunodominant epitopes of a pathogen [10]. The epitope-based approach may also augment CD8 1 CTL responses with the inclusion of high potency CD4 1 T H (helper T lymphocyte) epitopes into the vaccine construct to sustain antiviral CD8 1 T-cell activity [11, 12]. Several investigators have examined the potential of From the Laboratory of Vaccine Research (L.BM., R.K., J.L., C.A., L.L.), the Department of Biostatistics, Division of Information Sciences (J.L.), City of Hope National Medical Center, Duarte, California, USA, the Beckman Research Institute of the City of Hope (D.J.D.), Duarte, California, USA, and the Department of Pathology, Vanderbilt University Medical Center (J.P.), Nashville, Tennessee, USA. Address reprint requests to: Don J. Diamond, Ph.D., Laboratory of Vaccine Research, Beckman Research Institute of the City of Hope, 1500 E. Duarte Road, Duarte, CA 91010, USA; Fax: (626) 301-8981; E-Mail: [email protected]. Received February 10, 2000; revised April 13, 2000; accepted April 17, 2000. This work is supported by grants from the Leukemia Society of America to D.J.D. (LSA #98-6116), and grants from the NCI (CA77544 and CA30206-Project III) and CA33572 as a Cancer Center core grant to the City of Hope. Human Immunology 61, 764 –779 (2000) 0198-8859/00/$–see front matter © American Society for Histocompatibility and Immunogenetics, 2000 Published by Elsevier Science Inc. PII S0198-8859(00)00139-7
Transcript of Induction of CTL response by a minimal epitope vaccine in HLA A∗0201/DR1 transgenic mice:...

Induction of CTL Response by a MinimalEpitope Vaccine in HLA A*0201/DR1Transgenic Mice: Dependence on HLAClass II Restricted TH Response
Lbachir BenMohamed, Radhika Krishnan,Jeffrey Longmate, Catherine Auge, Ldito Low,James Primus, and Don J. Diamond
ABSTRACT: CTL play a pivotal role in the immuneresponse during viral infections. In this study, the HLAclass II restricted TH requirement for optimal in vivoinduction of HLA class I restricted CTL responses hasbeen investigated. Towards this goal, transgenic miceexpressing both HLA class I (A*0201 or A2.1) and classII (DRB1*0101 or DR1) molecules have been derived.Immunization of these mice with an HLA A*0201-re-stricted and CMV-specific CTL epitope (pp65495–503),and either of three different tetanus toxin-derived MHCclass II-binding TH epitopes, resulted in a vigorous CTLresponse. CTL specific for the pp65495–503 epitope weredramatically enhanced in mice expressing both the HLA-DR1 and HLA-A*0201 transgenes. Notably, preinjec-tion of three TT peptides (TT639 – 652, TT830 – 843, and
TT947–967) increased the capability of HLA A*0201/DR1Tg mice to respond to subsequent immunization with theTH 1 CTL peptide mixture. These results indicate thatthe use of HLA A*0201/DR1 Tg mice constitute a ver-satile model system (in lieu of immunizing humans) forthe study of both HLA class I and class II restricted T-cellresponses. These studies provide a rational model for thedesign and assessment of new minimal-epitope vaccinesbased on their in vivo induction of a pathogen-specificCTL response. Human Immunology 61, 764–779 (2000).© American Society for Histocompatibility and Immu-nogenetics, 2000. Published by Elsevier Science Inc.
KEYWORDS: T help; cytotoxic T lymphocyte; HLA-DR1; HLA-A*0201; transgenic mice
ABBREVIATIONSPBMC peripheral blood mononuclear cellsCRA chromium release assayIVS in vitro stimulation
LN lymph nodesTg transgenicTT tetanus toxin
INTRODUCTIONDuring the past decade, considerable interest has beenfocused on specifically inducing CTL to prevent and/or
control chronic viral diseases. Among the numerous ap-proaches investigated, are epitope based studies usingsynthetic peptides derived from the processed form ofviral proteins which bind to MHC class I alleles [1–4].Protective antiviral immunity using CTL epitope pep-tides has been achieved in several experimental models ofinfection [5–9]. The epitope-based approach offers sev-eral potential advantages. Selection of naturally processedepitopes may allow the immune system to focus onhighly conserved and immunodominant epitopes of apathogen [10]. The epitope-based approach may alsoaugment CD81 CTL responses with the inclusion ofhigh potency CD41 TH (helper T lymphocyte) epitopesinto the vaccine construct to sustain antiviral CD81
T-cell activity [11, 12].Several investigators have examined the potential of
From the Laboratory of Vaccine Research (L.BM., R.K., J.L., C.A.,L.L.), the Department of Biostatistics, Division of Information Sciences(J.L.), City of Hope National Medical Center, Duarte, California, USA,the Beckman Research Institute of the City of Hope (D.J.D.), Duarte,California, USA, and the Department of Pathology, Vanderbilt UniversityMedical Center (J.P.), Nashville, Tennessee, USA.
Address reprint requests to: Don J. Diamond, Ph.D., Laboratory ofVaccine Research, Beckman Research Institute of the City of Hope, 1500 E.Duarte Road, Duarte, CA 91010, USA; Fax: (626) 301-8981; E-Mail:[email protected].
Received February 10, 2000; revised April 13, 2000; accepted April 17,2000.
This work is supported by grants from the Leukemia Society of America toD.J.D. (LSA #98-6116), and grants from the NCI (CA77544 andCA30206-Project III) and CA33572 as a Cancer Center core grant to theCity of Hope.
Human Immunology 61, 764–779 (2000)0198-8859/00/$–see front matter© American Society for Histocompatibility and Immunogenetics, 2000
Published by Elsevier Science Inc. PII S0198-8859(00)00139-7

using TT (tetanus toxin) derived peptides to inducestrong TH responses, to strengthen the potency of vaccinepreparations [8, 13, 14]. A large proportion of the pop-ulation has previously been vaccinated with tetanus tox-oid vaccine preparations, therefore, most subjects haveTT responsive precursor cells. Because a need for potentTH responses to maximize the duration of activity ofadoptively transferred CTL or peptide vaccines has beenshown, it would be of great value to harness the immune-stimulating capacity of tetanus vaccinations [15, 16].With the aid of the murine transgenic model discussedbelow, immunogenic determinants from TT protein,which bind to multiple HLA DR alleles [17–21] havebeen evaluated for optimal induction of HLA-A*0201-restricted and CMV-specific CTL responses in vivo.
As a preclinical model, a transgenic (Tg) mouse thatexpresses both HLA-A*0201 class I and HLA-DR1 classII molecules was established. The HLA A*0201 trans-gene used in these studies encodes a chimeric molecule inwhich the a3 domain of the HLA-A*0201 heavy chainwas replaced with the corresponding domain from themurine H-2 Kb heavy chain [22]. The HLA-DR1 trans-gene is a chimeric (human/mouse) molecule(DRB1*0101, DRA1*0101) in which the a2 domain ofthe HLA-DR1 a chain was replaced with the corre-sponding domain from the murine I-Ea chain, therebyenhancing interaction with murine CD4 [23]. This wasdone since, at least for some HLA class II molecules,there is a species barrier to the interaction with murineCD4 [24]. The model has been used to test the role ofHLA class II in the induction and potency of HLA classI restricted CTL responses against a CMV-specific CTLepitope.
We report that both HLA-A*0201 and HLA-DR1transgenes are functional in vivo as MHC class I and classII restriction elements, respectively, and that the productof the HLA-DR1 transgene enhances HLA-A*0201-re-stricted CMV-specific CTL responses. Of particular in-terest is the fact that a single dose of the HLA-A*0201restricted pp65495–503 CTL epitope, in combinationwith three different TT derived TH epitopes; TT639–652,TT830–843, and TT947–967, resulted in the induction of avigorous CMV-specific CTL response in HLA-A*0201/DR1 Tg mice. These results have both theoretical andpractical importance for the development of prophylacticand therapeutic peptide vaccines against pathogens.
MATERIALS AND METHODSGeneration of Tg Mice Expressing HLA-DR1 andHLA-A*0201 MoleculesThe HLA-A*0201 transgene is a chimeric molecule con-sisting of the a1 and a2 domains of the HLA-A*0201molecule and the a3 domain of murine H-2Kb [22, 25].
The substitution of the a3 domain of the murine Kb
molecule is required to strengthen the interactions withendogenous murine CD8, producing a more vigorousimmune response [26, 27]. The Tg mice expressing theHLA class I gene were derived from homozygousC57BL/6 (H-2Kb) mice [22]. The HLA-DR1 transgenewas inserted into B10.M/Sn (H-2f) mice, which lackmurine I-E molecules that might form heterodimerswith human Tg MHC class II chains [28–31]. To gen-erate Tg mice carrying both human HLA class I and classII alleles, we crossed HLA-A*0201 Tg [22, 25] withHLA-DR1 Tg founders [28, 32] (kind gift of D. Zaller,Merck, Sharpe & Dohme, Rahway, NJ, USA). Progeny(F1) were first screened for the presence of both theHLA-A*0201 and HLA-DR1 transgenes by PCR (seebelow for PCR conditions). The expression of HLA-A*0201 and HLA-DR1 molecules in the positive prog-eny was confirmed by flow cytometric analyses of spleno-cytes from individual mice using antibodies directedagainst the individual transgene products (see below). Tgparental mice derived from inbred colonies of HLA-A*0201 and HLA-DR1 mice and the progeny were bredand maintained in the animal facility of the City of Hope(Duarte, CA, USA).
Polymerase Chain Reaction
Expression of DRB1*0101 and HLA A*0201 trans-genes, was detected by PCR using sequence-specific PCRprimers as described previously [33]. Briefly, DNA wasisolated from 4-mm ear/tail punches from Tg animals.Skin punches were treated for 3 to 4 hours in 50 mMTris-HCl, pH 8.0, 20 mM NaCl, 1 mM EDTA, 1% SDSand proteinase K (1 mg/ml), followed by an extractionwith phenol-chloroform. PCR was performed in tubescontaining mouse DNA (50 ml of genomic DNA forDRB1*0101 and 20 ml for HLA A*0201), 50 mM KCl,20 mM Tris-HCl, 2.5 mM MgCl2, 2.5 % DMSO, 100mM of each deoxynucleotide triphosphate, 0.1 mM ofeach primer, and 2.5 U of Taq polymerase. Ten cycles ofa PCR program consisting of 1-min denaturation at94°C, 1-min annealing at 65°C, and 2-min extension at72°C followed with twenty additional cycles consistingof 1-min denaturation at 94°C, 30 sec annealing at 55°C,and 1-min extension at 72°C were performed for detec-tion of the HLA-A*0201 transgene. Thirty cycles of aprogram consisting of 30 sec denaturation at 94°C, 45sec annealing at 57°C, and 1-min extension at 72°C wereperformed for HLA-DR1 transgene. For detection of theHLA-A*0201 transgene, the sense primer used was 59-CCT CGT CCC CAG GCT CT-39 and the antisenseprimer was 59-TGG CCC CTG GTA CCC GT-39 [34].For the DRB1*0101 a chain transgene, the sense primerwas 59-CAT GCC CTT CAC CCA GAC CTT GTC-39
765Response to Minimal Epitope CTL Vaccine

and the antisense primer was 59-ATC GTC CAA ATTCTT CAA GCC GCC-39 [28].
Flow Cytometric AnalysisTo assess the expression of HLA-DR1 and HLA-A*0201molecules, single-cell splenocyte suspensions were pre-pared by homogenizing tissue in 3 ml PBS containing5% FCS, 5 mM EDTA, and 0.02% NaN3 (PBS-5%buffer). Cells were washed in cold PBS-5% buffer andincubated with BB7.2 (anti-HLA-A*0201, ATCC) orL-243 (anti-HLA-DR, PharMingen, San Diego, CA,USA) mAbs for 20–30 min at 4°C, followed by FITC-labeled goat anti-mouse IgG2a/2b (PharMingen) for an-other 20–30 min at 4°C as described previously [25,28]. The BB7.2 mAb was purified from supernatant ofthe HB82 hybridoma (ATCC) (gift of S. Kovats, City ofHope, Duarte, CA, USA). Propidium iodide was used toexclude dead cells. For each sample, 5000–10,000 cellswere analyzed by flow cytometry with a FACSCalibur(Becton Dickinson, San Jose, CA, USA) and analyzedwith CellQuest software (Becton Dickinson) on an inte-grated power Macintosh G3 (Apple Computers, Cuper-tino, CA, USA).
Viruses and Cell LinesThe AD169 strain of human CMV was used in allexperiments and was propagated and titered as previ-ously described [3]. Briefly, the virus was propagated inMRC-5, a human fibroblast cell line by three serialpassages. Infectious supernatant was harvested to prepareviral stocks when MRC-5 cells showed 100% cytopathiceffect. Dermal HLA matched (A2, B51, B55) and HLAmismatched (A1, A30, B13, B18) fibroblasts lines weregenerated by expanded culture of skin biopsies. Fibro-blasts were maintained in DMEM supplemented with10% heat-inactivated FCS, 2 mM glutamine, 100 U/mlpenicillin, 100 mg/ml streptomycin and were used as
target cells between the 4th and 12th passages. T2 cells[35], HLA-matched EBV-LCL (Epstein-Barr virus trans-formed lymphocyte cell lines), expressing A*0201, B51,and B55, as well as mismatched EBV-LCL expressingA1, A30, B13, and B18 were used to assay the MHCrestriction of effector cells. EBV-LCL and T2 cell lineswere maintained in RPMI-1640 with 10% FCS and weresplit 1:2 in fresh medium 1 d before use.
Synthetic PeptidesThe HLA-A*0201-restricted CTL epitope derived fromthe CMV tegument protein (pp65), referred to aspp65495–503 peptide (NH2-NLVPMVATV-COOH),was used in this study [3]. The five TT peptides used inthis study as TH epitopes have been described previously:TT590–603, TT615-629, and TT639–652 [19]; andTT830–843 and TT947–967 [17, 18, 20, 21]. The peptidesequences of these TT TH epitopes are shown in Table 1.All synthetic peptides were synthesized on an ABI 432instrument using standard F-Moc technology (PE Ap-plied Biosystems, Foster City, CA, USA). Peptides werecleaved from the Wang resin using TFA: Ethanedithiol:Thioanisole: Anisole: (90:5:3:2) followed by ether (meth-yl-t-butyl ether) extraction and lyophilization. Stock so-lutions of all synthetic peptides were made at 5 mM ina solution of 0.1% acetic acid and 10% DMSO, andpurity evaluated on a C18 HPLC column (Vydac Separa-tions, Hesperia, CA, USA) using a Shimadzu SCL-10AVP (Shimadzu, Columbia, MD, USA) chromatographicsystem.
Immunization ProceduresFor all immunization experiments, HLA-DR1 and/orHLA-A*0201 mice were 6–8 weeks of age. Mice wereimmunized with pp65495–503 and/or TT peptides in PBSemulsified with IFA (Gibco BRL, Gaithersburg, MD,USA), as previously described [2]. Mice were immunized
TABLE 1 Amino acid sequence of TT peptides used in this study
Tetanus peptide AA sequence
HLA class IIrestriction
(DR b alleles)DR allelic frequencies(caucasian population) References
TT590–603 NH2-TKIYSYFPSVISKV-COOH DR1, 2, 3, 4, 7 81.2% (93.7%) [19][44]
TT615–629 NH2-VRDIIDDFTNESSQK-COOH DR2, 4, 7, 13 74.0% (91.4%) [19][44]
TT639–652 NH2-VSTIVPYIGPALNI-COOH DR1, 2, 3, 4 67.2% (79.7%) [19][71]
TT830–843 NH2-QYIKANSKFIGITE-COOH DR1, 2, 3, 4, 5 7,8, 9, 11, 13, 51
85.5% (94.2%) [48][44]
TT947–967 NH2-FNNFTVSFWLRVPKVSASHLE-COOH DR1, 7, 9, 11 65.3% (57.2%) [19][71, 18]
Known HLA-DR restriction for each TT peptide and the cumulative phenotypic frequencies of the indicated HLA-DR antigens as an average of the frequenciesfor all major ethnicities are shown. Frequencies for each HLA-DR phenotype are also shown in the Caucasian population in parentheses.
766 L. BenMohamed et al.

subcutaneously at the base of the tail with 100 nmol ofeach peptide. For some experiments, mice were boosted2–3 weeks later with 100 nmol of the same peptide inIFA. Control mice were injected subcutaneously with100 ml of sterile PBS, emulsified with IFA. Twelve daysafter peptide immunization, spleens were harvested andcell suspensions were restimulated using peptide-sensi-tized stimulator cells (see below). All procedures wereconducted with approval of the City of Hope Institu-tional Animal Care and Use Committee and according tothe National Institutes of Health Guide for the Care andUse of Laboratory Animals.
In Vitro Proliferation Assays of Human PBMCVenous blood (40 ml) was collected from normal donorsinto heparinized tubes at the City of Hope Blood DonorCenter. Peripheral blood mononuclear cells (PBMC) wereseparated from whole blood by centrifugation on a Ficoll-Hypaque density gradient (Amersham Pharmacia, Upp-sala, Sweden) and were washed and cultured as previouslydescribed [36]. PBMC were cultured with or without TTpeptides in RPMI-1640 medium supplemented with10% human AB1 serum at 23105 cells/well in flat-bottomed 96-well plates in a total volume of 200 ml.Triplicate wells were set up for each individual TTpeptide (90, 30, 10, 3, 1, and 0.3 mM) or condition,among which included wells for tetanus toxoid (10, 3, 1,0.3, and 0.1 Lyme factor/ml). PPD (10 mg/ml), a puri-fied protein derivative of Mycobacterium tuberculosis, wasused as a positive control Ag (kindly provided by PasteurMerieux Connaught, Swiftwater, PA, USA). Cell suspen-sions were incubated for 5 days at 37°C in a 5% CO2incubator. Cultures were pulsed with 1 mCi of [3H]-thymidine per well for 16 h before harvesting, andcounted in a Trilux Microbeta 1450 plate liquid scintil-lation counter (Wallac, Turku, Finland). Results areexpressed as D cpm (D cpm 5 cpm in the presence ofantigen-cpm without antigen) and as stimulation indices(SI 5 cpm in the presence of antigen/cpm withoutantigen). A proliferative response was deemed as positivewhen the D cpm . 1000 cpm and SI . 2.5 [37].
Cellular Immune Responses in MiceLymphocyte proliferation assays.
Twelve days after immunization the spleen, inguinal,and para-aortic lymph nodes (LN) were removed fromanimals, and placed into ice-cold DMEM medium.Spleen and LN single-cell suspensions were washed twicewith DMEM supplemented with 15 mM HEPES,5.5031025 M-mercaptoethanol (Gibco BRL), 2 mMglutamine, 50 U of penicillin/ml and 50 mg of strepto-mycin/ml. Spleen and LN single-cell suspensions wereadjusted to 43106 cells/ml in complete medium supple-mented with 10% heat-inactivated fetal calf serum
(FCS), and used as previously described [37]. Equal vol-umes of cells and complete medium or complete mediumwith recall TT peptides were mixed to give a finalconcentration of 23106 cells/ml in medium alone or inmedium with peptide at 90, 30, 10, 3, or 1 mM con-centration. The cell suspensions were incubated for 72 hat 37°C in 5 % CO2. One mCi of [3H]-thymidine(Dupont NEN, Boston, MA, USA) was added to eachwell, 16 h before the cells were harvested onto glass fiberfilters, and then counted on a Matrix 96 direct ioniza-tion-counter (Packard Instruments, Meriden, CT, USA)and the incorporated radioactivity determined as de-scribed previously. All cultures were set up in quadru-plicate. Results are expressed as the mean cpm of cell-associated [3H]-thymidine recovered from wellscontaining Ag subtracted from the mean cpm of cell-associated [3H]-thymidine recovered from wells withoutAg (D cpm; average of quadruplicate). The T-cell mito-gen Concanavalin A (Sigma, St. Louis, MO, USA), wasused as a positive control. Proliferation results wereconfirmed by repeating each experiment twice.
DTH AssayThe DTH reaction was elicited by injection of 5 mg ofTT639–652, T830–843, or TT947–967 TH peptide in PBSin a 25 ml volume into the right hind footpad of eachmouse, using a Hamilton microliter 702 syringe (Ham-ilton Company, Reno, NV, USA). The left hind footpadwas injected with 25 ml of PBS as a control. Using avernier caliper, footpad swelling was determined 24, 48,and 72 h later as measure of DTH reaction as previouslydescribed [38].
Footpad HistologySeventy-two hours after boosting with recall peptides,footpad tissues were excised immediately after sacrifice ofmice, washed in PBS, fixed in 4% paraformaldehyde inPBS for 7 to 10 days, and footpad histology performed aspreviously described [39]. For light microscopy, footpadtissues were fixed in 10% neutral buffered formalin andprocessed for paraffin embedding. Serial 4-mm sectionswere stained with hematoxylin/eosin and examined usinga Leitz DM-RB light microscope (Leica MicrosystemsInc., Deerfield, IL, USA). Preparations were photo-graphed using digitizing SPOT 1.1.0 camera (DiagnosticInstruments, Inc., Sterling Heights, MI, USA) and ana-lyzed with the Image Pro 3.0 software (Media Cybernet-ics, Silver Spring, MD, USA) on a personal computer(Gateway P5-100).
Generation of CTL Cultures and ChromiumRelease AssaysThe method used for generation of CTL lines has beenpreviously described [3]. Immune spleen and LN cells
767Response to Minimal Epitope CTL Vaccine

were re-stimulated in vitro for 5–6 days on autologousspleen feeder cells that were first lipopolysaccharide(LPS) stimulated for 72 h then acid treated and loadedwith pp65495–503 peptide according to a modification ofWentworth et al. [40]. An additional in vitro restimula-tion was performed on days 5–6 following the firststimulation, using peptide-pulsed and irradiated T2 cellsas APC and irradiated spleen cells as feeders. Growthfactors in the form of 5% (vol/vol) Rat T-Stim (Collab-orative Biomedical Products, Bedford, MA, USA) wereadded to the culture 2 days after each in vitro restimu-lation. Six days after the second in vitro restimulation,cytotoxic activity was assessed against peptide-sensitizedT2 or HLA-A*0201 positive B-LCL target cells. Targetsincubated with relevant and irrelevant peptide were pel-leted, resuspended in 100 mCi of Na-51CrO4 per 106
cells (Amersham Corp., Arlington Heights, IL, USA),and incubated for 1 h at 37°C in a humidified 5% CO2incubator. The cells were rinsed three times and resus-pended at 105/ml, and 0.1 ml of the cell preparation wasadded to round-bottomed microtiter wells. Varyingnumbers of effector splenocytes were added in triplicatein 0.1 ml of medium to achieve the desired E/T ratio.Autologous and mismatched human fibroblast (0.53106
cells) were split into 25 cm2 flasks overnight followed byincubation with 400 IU of human IFN-g for 48 h asdescribed [3]. Cells were then infected with CMV (AD-169 strain) at a multiplicity of infection of 5 for approx-imately 12 h. Cells were labeled with Na-51CrO4 for 45min and washed twice prior to use as targets (2000/well)in a standard 4 h chromium release assays (CRA) in a96-well U-bottom plate. For spontaneous 51Cr-release,0.1 ml of media was added to the labeled target cells inplace of effector cells. For maximum 51Cr release, 0.1 mlof 2% SDS (United States Biochemical Co., Cleveland,OH, USA) was added to the labeled target cells. After4 h at 37°C in a 5% CO2 incubator, the microtiter plateswere centrifuged at 200 RPM for 2 min (RT-2000D,Sorvall Instruments, Newtown, CT, USA). Supernatantswere removed (0.1 ml) from each well and counted on aCobra II (Packard Instrument, Meriden, CT, USA)gamma counter. Data are expressed as: percentage of 51Crrelease 5 100 3 (experimental cpm 2 spontaneouscpm)/(maximum release cpm 2 spontaneous releasecpm). Specific lysis, presented, is a result of subtractingcpm from untreated cells from treated cells. Experimen-tal data had calculated spontaneous release , 15% forB-LCL and T2 cells and , 25% for fibroblasts.
mAb Blocking of CTL ActivityHLA-A*0201/DR1 Tg mice were immunized with TTand/or pp65495–503 peptides, and splenocytes were iso-lated and stimulated as described above. Stimulatedsplenocytes were then incubated with target HLA
A*0201-matched B-LCL with or without the anti-CD8mAb 53-6.7 (PharMingen). Identically prepared cultureswere incubated with anti-CD4 mAb GK1.5 (PharMin-gen) or an isotype-matched control mAb, at 10 mg/mlduring the lysis portion of the assay.
Statistical Analysis
All data shown are representative of at least two inde-pendent experiments. The data are expressed as themean 6 SEM, and analysis of variance methods withspecific constants were used to compare mean specificlysis under different experimental conditions. All exper-iments analyzed were balanced, and F-tests were used topredict false positive rates.
RESULTS
Selection and Characterization of TTImmunodominant TH Epitopes
In vitro proliferative response of human PBMC to TT andTT-derived peptides.
Previous studies have defined TT-derived CD41 TH
epitopes that promiscuously bound to DR alleles asdetected by in vitro primary immunization of humanPBMC [17, 19, 41] (Table 1). However, it was unclearwhich promiscuous TT epitope caused superior immuneresponses in the context of the broadest number of DRalleles. TT protein and five of the most promising TH
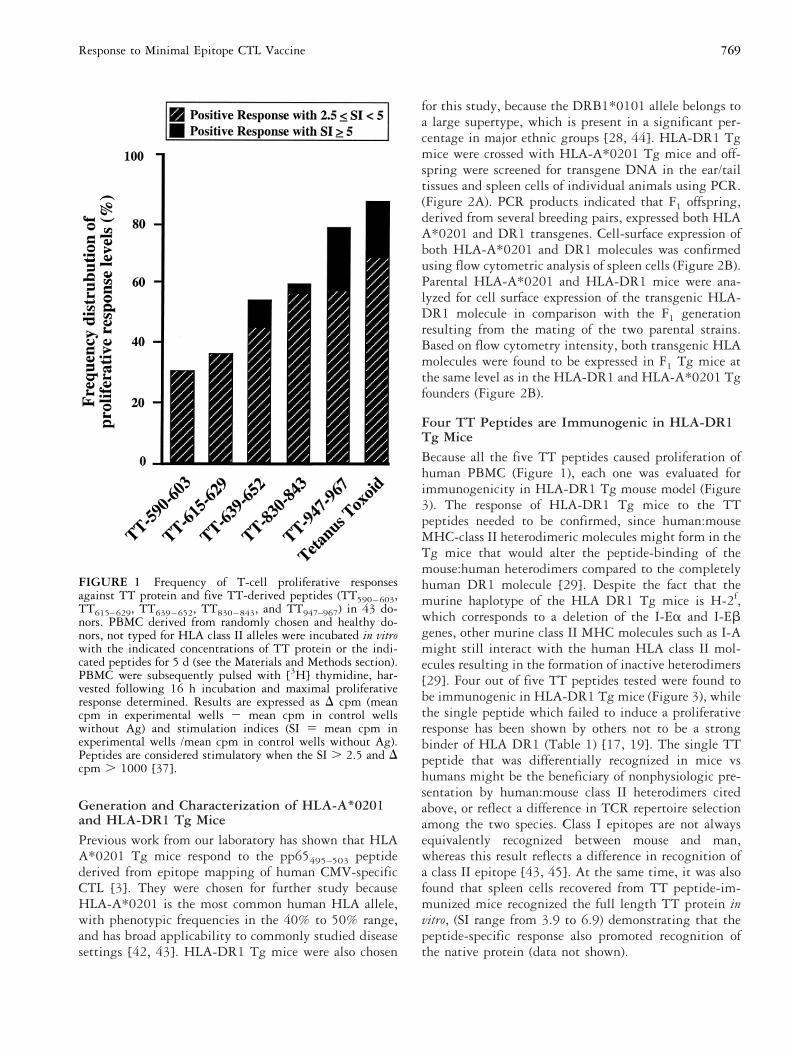
epitopes were used to stimulate human T-cell prolifera-tive responses in 43 randomly chosen donors (MHChaplotypes unknown). As a control, PBMC were testedfor their proliferative response to the Ag PPD. Specificcell proliferative responses were observed for TT proteinas well as for every TT peptide, indicating that each ofthe five chosen sequences contained T cell determinants(Figure 1). T-cell proliferative responses directed primar-ily, although not exclusively, to three peptides(TT639–652, TT830–843 and TT947–967), were detected inPBMC of the 43 donors tested (Figure 1). The frequencyof responders (SI Ä 2.5 and . cpm . 1000) was 53%for TT639–652, 58% for TT830–843 and up to 79% forTT947–967 peptide. However, the frequency of respondersto TT590–603 and TT615–629 peptides were relatively low(38% and 30%, respectively) (Figure 1). TT639–652,TT830–843 and TT947–967 peptides were found to stim-ulate elevated T proliferative responses, with SI 5 5,whereas none of the TT590–603 and TT615–629 peptideswas able to stimulate a high level of response. Theseproliferative responses were dependent on the CD41 Tcell subset, as they were blocked by a mAb against CD4(data not shown). In contrast, a mAb against CD8 didnot block proliferative responses (data not shown).
768 L. BenMohamed et al.
Generation and Characterization of HLA-A*0201and HLA-DR1 Tg MicePrevious work from our laboratory has shown that HLAA*0201 Tg mice respond to the pp65495–503 peptidederived from epitope mapping of human CMV-specificCTL [3]. They were chosen for further study becauseHLA-A*0201 is the most common human HLA allele,with phenotypic frequencies in the 40% to 50% range,and has broad applicability to commonly studied diseasesettings [42, 43]. HLA-DR1 Tg mice were also chosen
for this study, because the DRB1*0101 allele belongs toa large supertype, which is present in a significant per-centage in major ethnic groups [28, 44]. HLA-DR1 Tgmice were crossed with HLA-A*0201 Tg mice and off-spring were screened for transgene DNA in the ear/tailtissues and spleen cells of individual animals using PCR.(Figure 2A). PCR products indicated that F1 offspring,derived from several breeding pairs, expressed both HLAA*0201 and DR1 transgenes. Cell-surface expression ofboth HLA-A*0201 and DR1 molecules was confirmedusing flow cytometric analysis of spleen cells (Figure 2B).Parental HLA-A*0201 and HLA-DR1 mice were ana-lyzed for cell surface expression of the transgenic HLA-DR1 molecule in comparison with the F1 generationresulting from the mating of the two parental strains.Based on flow cytometry intensity, both transgenic HLAmolecules were found to be expressed in F1 Tg mice atthe same level as in the HLA-DR1 and HLA-A*0201 Tgfounders (Figure 2B).
Four TT Peptides are Immunogenic in HLA-DR1Tg MiceBecause all the five TT peptides caused proliferation ofhuman PBMC (Figure 1), each one was evaluated forimmunogenicity in HLA-DR1 Tg mouse model (Figure3). The response of HLA-DR1 Tg mice to the TTpeptides needed to be confirmed, since human:mouseMHC-class II heterodimeric molecules might form in theTg mice that would alter the peptide-binding of themouse:human heterodimers compared to the completelyhuman DR1 molecule [29]. Despite the fact that themurine haplotype of the HLA DR1 Tg mice is H-2f,which corresponds to a deletion of the I-Ea and I-Ebgenes, other murine class II MHC molecules such as I-Amight still interact with the human HLA class II mol-ecules resulting in the formation of inactive heterodimers[29]. Four out of five TT peptides tested were found tobe immunogenic in HLA-DR1 Tg mice (Figure 3), whilethe single peptide which failed to induce a proliferativeresponse has been shown by others not to be a strongbinder of HLA DR1 (Table 1) [17, 19]. The single TTpeptide that was differentially recognized in mice vshumans might be the beneficiary of nonphysiologic pre-sentation by human:mouse class II heterodimers citedabove, or reflect a difference in TCR repertoire selectionamong the two species. Class I epitopes are not alwaysequivalently recognized between mouse and man,whereas this result reflects a difference in recognition ofa class II epitope [43, 45]. At the same time, it was alsofound that spleen cells recovered from TT peptide-im-munized mice recognized the full length TT protein invitro, (SI range from 3.9 to 6.9) demonstrating that thepeptide-specific response also promoted recognition ofthe native protein (data not shown).
FIGURE 1 Frequency of T-cell proliferative responsesagainst TT protein and five TT-derived peptides (TT590–603,TT615–629, TT639–652, TT830–843, and TT947–967) in 43 do-nors. PBMC derived from randomly chosen and healthy do-nors, not typed for HLA class II alleles were incubated in vitrowith the indicated concentrations of TT protein or the indi-cated peptides for 5 d (see the Materials and Methods section).PBMC were subsequently pulsed with [3H] thymidine, har-vested following 16 h incubation and maximal proliferativeresponse determined. Results are expressed as D cpm (meancpm in experimental wells 2 mean cpm in control wellswithout Ag) and stimulation indices (SI 5 mean cpm inexperimental wells /mean cpm in control wells without Ag).Peptides are considered stimulatory when the SI . 2.5 and Dcpm . 1000 [37].
769Response to Minimal Epitope CTL Vaccine
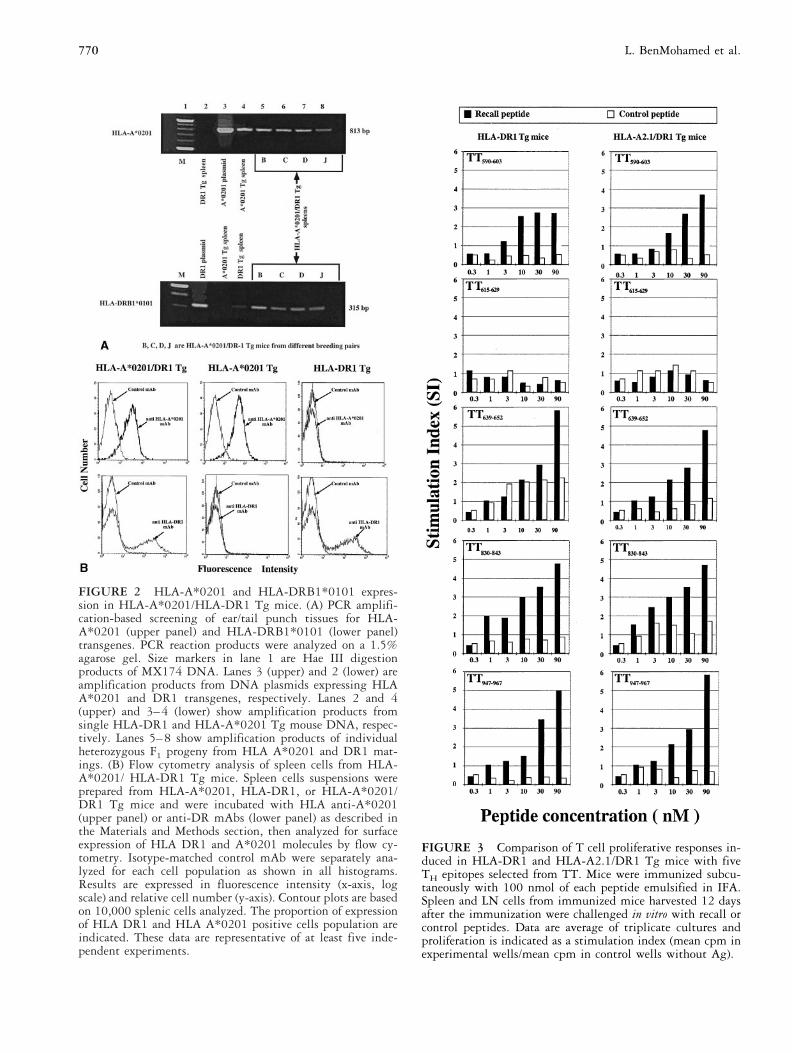
FIGURE 2 HLA-A*0201 and HLA-DRB1*0101 expres-sion in HLA-A*0201/HLA-DR1 Tg mice. (A) PCR amplifi-cation-based screening of ear/tail punch tissues for HLA-A*0201 (upper panel) and HLA-DRB1*0101 (lower panel)transgenes. PCR reaction products were analyzed on a 1.5%agarose gel. Size markers in lane 1 are Hae III digestionproducts of MX174 DNA. Lanes 3 (upper) and 2 (lower) areamplification products from DNA plasmids expressing HLAA*0201 and DR1 transgenes, respectively. Lanes 2 and 4(upper) and 3–4 (lower) show amplification products fromsingle HLA-DR1 and HLA-A*0201 Tg mouse DNA, respec-tively. Lanes 5–8 show amplification products of individualheterozygous F1 progeny from HLA A*0201 and DR1 mat-ings. (B) Flow cytometry analysis of spleen cells from HLA-A*0201/ HLA-DR1 Tg mice. Spleen cells suspensions wereprepared from HLA-A*0201, HLA-DR1, or HLA-A*0201/DR1 Tg mice and were incubated with HLA anti-A*0201(upper panel) or anti-DR mAbs (lower panel) as described inthe Materials and Methods section, then analyzed for surfaceexpression of HLA DR1 and A*0201 molecules by flow cy-tometry. Isotype-matched control mAb were separately ana-lyzed for each cell population as shown in all histograms.Results are expressed in fluorescence intensity (x-axis, logscale) and relative cell number (y-axis). Contour plots are basedon 10,000 splenic cells analyzed. The proportion of expressionof HLA DR1 and HLA A*0201 positive cells population areindicated. These data are representative of at least five inde-pendent experiments.
FIGURE 3 Comparison of T cell proliferative responses in-duced in HLA-DR1 and HLA-A2.1/DR1 Tg mice with fiveTH epitopes selected from TT. Mice were immunized subcu-taneously with 100 nmol of each peptide emulsified in IFA.Spleen and LN cells from immunized mice harvested 12 daysafter the immunization were challenged in vitro with recall orcontrol peptides. Data are average of triplicate cultures andproliferation is indicated as a stimulation index (mean cpm inexperimental wells/mean cpm in control wells without Ag).
770 L. BenMohamed et al.

Evaluation of TT Peptides in HLA-DR1/A*0201Tg MiceA further question was whether any of the five TTpeptides would be immunogenic in a mouse that ex-pressed both I-E and I-A MHC class II genes, such as inthe case of the HLA-A*0201/DR1 Tg mice (H-2f/H-2b).In two separate experiments, a single injection ofTT590–603, TT639–652, TT830–843, or TT947–967 pep-tides resulted in a substantial TH proliferative responsein HLA-A*0201/DR1 Tg mice (Figure 3). However,TT615–629, which is antigenic in humans (this study and[17–19, 46]), did not result in a substantial proliferativeresponse in HLA-DR1 or -A*0201/DR1 Tg mice (Fig-ure 3). Inhibition studies with both anti-CD4 and CD8mAbs were conducted by adding them to separate ali-quots of the same spleen culture for the length of theproliferation assay (72 h). Only the anti-CD4 and not theCD8 or an irrelevant isotype-matched mAb showed adose-dependent inhibition of TH proliferative responses(data not shown). To exclude murine MHC class IIheterodimers as the presenting molecules for these TTpeptides as opposed to HLA-DR1, non-Tg C57BL/6were immunized under the same conditions as the Tgmice. Consistent with their inability to bind I-Ab,TT590–603, TT639–652, TT830–843, and TT947–967 wereunable to generate a specific T cell proliferative responsein nontransgenic C57BL/6 control mice [47, 48] (datanot shown). Collectively, these results indicate that fourTT TH peptides TT590–603, TT639–652, TT830–843, andTT947–967, which induced human PBMC proliferation invitro, are immunogenic in HLA-DR1 Tg as well as inHLA-A*0201/DR1 Tg mice. Among the four TT pep-tides, TT639–652, TT830–843, and TT947–967 were foundto induce strong proliferation of human PBMC in vitro aswell in DR1 Tg mice in vivo. The data also demonstratethat HLA-DR1 molecules are capable of presenting pro-cessed TT peptides in either HLA-DR1 or -A*0201/DR1 Tg mice, and in the latter case endogenous murineMHC molecules did not inhibit presentation of the pep-tides.
Delayed-type Hypersensitivity Immune Responsesto TT639–652, TT830–843, and TT947–967 Peptides inHLA-A*0201/DR1 Tg MiceTo determine the ability of TT peptides to promote invivo cell-mediated immunity in the HLA-A*0201/DR1Tg model, delayed-type hypersensitivity (DTH) immuneresponses was assessed after immunization with TT pep-tides. TT639–652, TT830–843, or TT947–967 peptidesalone, or in combination with pp65495–503 peptide wereadministrated s.c. in separate groups of HLA-A*0201/DR1 Tg mice. Twelve days later, DTH response waselicited by injecting 5 mg/mouse of the individual recallpeptides (i.e., TT639–652, TT830–0843, TT947–967, or
pp65495–503 peptide) into the right footpad. Footpadswelling was measured 24, 48, and 72 h after the injec-tion, and compared with the left footpad injected withsaline as a control. As shown in Figure 4A, there was anequivalent DTH response generated in HLA-A*0201/DR1 Tg mice by TT639–652, TT830–843, and TT947–967peptides. However, no measurable DTH response wasfound when the pp65495–503 CTL epitope was tested.The development of the DTH response after challengewith recall peptides TT639–652, TT830–843, orTT947–967, but not with the pp65495–503 peptide, wasaccompanied by a strong mononuclear cellular infiltrateas determined by footpad histology (Figure 4B). On theother hand, no DTH response was induced in the footpadreceiving PBS alone or in non-immunized control mice(Figures 4A and B). Therefore, the three TT peptidesTT639–652, TT830–843, and TT947–967, stimulate a DTHresponse, which is of equivalent magnitude for each TTpeptide and substantially greater than the DTH responseinduced by pp65495–503 CTL epitope. These results showthat the TT peptides can also generate an in vivo response(DTH), and extend the usefulness of the HLA-A *0201/DR1 Tg mouse model to address in vivo cell mediatedimmune responses.
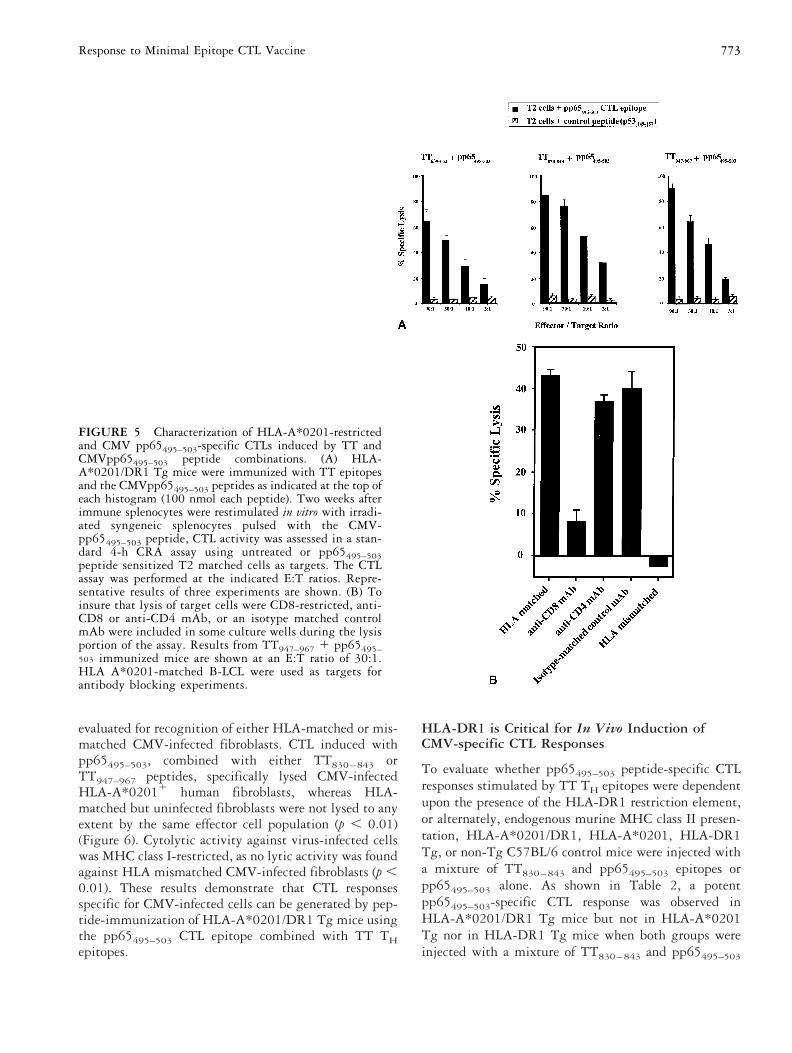
Generation of pp65-specific andHLA-A*0201-restricted CTL in HLA-A*0201/DR1Tg MiceHLA-dependent recognition of class I and II restrictedantigens in the same mouse model would allow thetesting and optimization of vaccines being developed forhuman use, without the complication of partial depen-dence on murine MHC recognition systems. CTL re-sponses in HLA-A*0201/DR1 Tg mice were evaluatedafter injection with the HLA-A*0201-restricted CMVpp65495–503 CTL peptide combined with three HLA-DR1 restricted TT peptides (Figure 3). The pp65495–503CTL peptide was previously shown to induce strong invitro CTL responses in HLA-A*0201 and CMV-seropos-itive humans or in HLA-A*0201 Tg mice in combina-tion with the PADRE TH epitope [3, 49]. As shown inFigure 5A, pp65495–503 combined with TT639–652,TT830–843, or TT947–967 peptides was found to behighly immunogenic with a single dose eliciting a strongpeptide-specific CTL response. The CTL responses inmice that received pp65495–503 combined withTT830–843 or TT947–967 peptides were of similar mag-nitude (p 5 0.06) but greater than the CTL responseinduced when the pp65495–503 epitope was combinedwith the TT639–652 peptide (p , 0.01). Thus, CTLresponses could be generated in vivo by these peptideformulations in a hierarchical fashion in which some TTTH epitopes were more potent than others in stimulatingCTL immunity (Figure 5A). CTL responses in the pres-
771Response to Minimal Epitope CTL Vaccine

ence of the other two TT peptides (TT590–603 andTT615–629) were minimal, and may reflect there de-creased ability to stimulate TH responses in mice andhumans (Figures 1 and 3) compared with the three TTpeptides shown in Figure 5 (data not shown). The pep-tide-specific CTL induced in HLA-A*0201/DR1 Tgmice using the TT TH epitopes were of the CD81
phenotype inasmuch as specific lysis was significantlyinhibited by mAbs against CD8 (p , 0.01), but onlyslightly inhibited by mAbs against CD4 (p 5 0.015).The CTL were also found to be HLA-restricted as therewas cytolytic activity against HLA-matched B-LCL butnot HLA-mismatched B-LCL (p , 0.01) (Figure 5B).
Immunization with pp65495–503 and TT EpitopeMixtures Results in Anti-CMV CTL Activity
The immune response elicited by CTL1TH epitope rec-ognition must promote lysis of virally-infected APC, inorder for this Tg model to be useful in anti-viral research.In the CMV model, it was previously shown that spleno-cytes from peptide immunized mice could recognize andkill CMV-infected human fibroblasts in an HLA-re-stricted manner [3]. The recognition of CMV-infectedtargets was evaluated in HLA-A*0201/DR1 Tg mice,immunized once with CTL 1 TH peptides emulsified inIFA. Following two IVS, spleen cell suspensions were
FIGURE 4 Ag-specific DTH responses inHLA/A*0201/DR1 Tg mice primed with CMVpp65495–503 and TT peptides. (A) Footpadswelling of HLA-A*0201/DR1 Tg mice primedwith CMV pp65495–503 1 TT639–652, CMVpp65495–503 1 TT830–843, or CMV pp65495–503 1 TT947–967 TH peptides and boosted, 2weeks after priming, with individual recall pep-tide (i.e., pp65495–503, TT639–652, TT830–843, orTT947–967) or with saline. Footpad swelling wasmeasured 48 h after boost. (B) Histochemicalanalysis of footpads from HLA-A*0201/DR1 Tgmice primed with CMV pp65495–503 in combi-nation with TT639–652, TT830–843, or TT947–967TH peptides and boosted 2 weeks after primingwith TT639–652 (a), or TT830–843 (b), orTT947–967 (c), or PBS (d). Mice were killed 48 hafter boosting, and tissues from the distal foot-pad were stained with hematoxylin and eosin.Original magnification, 3400 for all sections. Arepresentative result of three experiments isshown.
772 L. BenMohamed et al.
evaluated for recognition of either HLA-matched or mis-matched CMV-infected fibroblasts. CTL induced withpp65495–503, combined with either TT830–843 orTT947–967 peptides, specifically lysed CMV-infectedHLA-A*02011 human fibroblasts, whereas HLA-matched but uninfected fibroblasts were not lysed to anyextent by the same effector cell population (p , 0.01)(Figure 6). Cytolytic activity against virus-infected cellswas MHC class I-restricted, as no lytic activity was foundagainst HLA mismatched CMV-infected fibroblasts (p ,0.01). These results demonstrate that CTL responsesspecific for CMV-infected cells can be generated by pep-tide-immunization of HLA-A*0201/DR1 Tg mice usingthe pp65495–503 CTL epitope combined with TT THepitopes.
HLA-DR1 is Critical for In Vivo Induction ofCMV-specific CTL Responses
To evaluate whether pp65495–503 peptide-specific CTLresponses stimulated by TT TH epitopes were dependentupon the presence of the HLA-DR1 restriction element,or alternately, endogenous murine MHC class II presen-tation, HLA-A*0201/DR1, HLA-A*0201, HLA-DR1Tg, or non-Tg C57BL/6 control mice were injected witha mixture of TT830–843 and pp65495–503 epitopes orpp65495–503 alone. As shown in Table 2, a potentpp65495–503-specific CTL response was observed inHLA-A*0201/DR1 Tg mice but not in HLA-A*0201Tg nor in HLA-DR1 Tg mice when both groups wereinjected with a mixture of TT830–843 and pp65495–503
FIGURE 5 Characterization of HLA-A*0201-restrictedand CMV pp65495–503-specific CTLs induced by TT andCMVpp65495–503 peptide combinations. (A) HLA-A*0201/DR1 Tg mice were immunized with TT epitopesand the CMVpp65495–503 peptides as indicated at the top ofeach histogram (100 nmol each peptide). Two weeks afterimmune splenocytes were restimulated in vitro with irradi-ated syngeneic splenocytes pulsed with the CMV-pp65495–503 peptide, CTL activity was assessed in a stan-dard 4-h CRA assay using untreated or pp65495–503peptide sensitized T2 matched cells as targets. The CTLassay was performed at the indicated E:T ratios. Repre-sentative results of three experiments are shown. (B) Toinsure that lysis of target cells were CD8-restricted, anti-CD8 or anti-CD4 mAb, or an isotype matched controlmAb were included in some culture wells during the lysisportion of the assay. Results from TT947–967 1 pp65495–503 immunized mice are shown at an E:T ratio of 30:1.HLA A*0201-matched B-LCL were used as targets forantibody blocking experiments.
773Response to Minimal Epitope CTL Vaccine
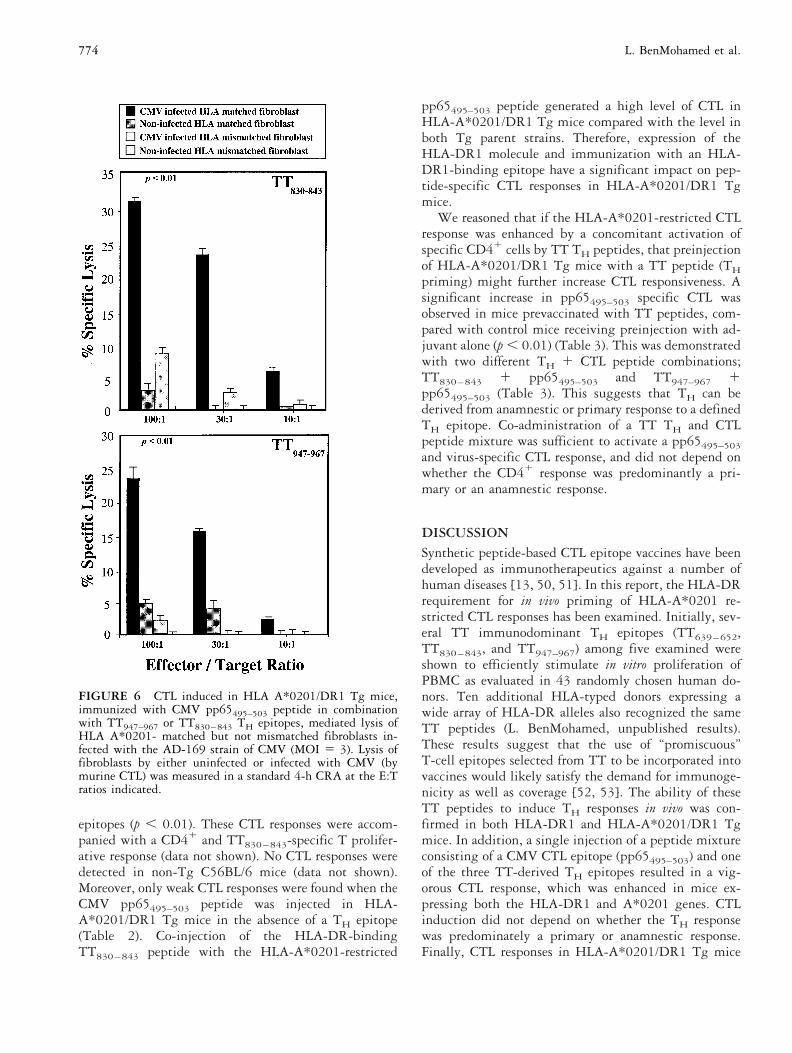
epitopes (p , 0.01). These CTL responses were accom-panied with a CD41 and TT830–843-specific T prolifer-ative response (data not shown). No CTL responses weredetected in non-Tg C56BL/6 mice (data not shown).Moreover, only weak CTL responses were found when theCMV pp65495–503 peptide was injected in HLA-A*0201/DR1 Tg mice in the absence of a TH epitope(Table 2). Co-injection of the HLA-DR-bindingTT830–843 peptide with the HLA-A*0201-restricted
pp65495–503 peptide generated a high level of CTL inHLA-A*0201/DR1 Tg mice compared with the level inboth Tg parent strains. Therefore, expression of theHLA-DR1 molecule and immunization with an HLA-DR1-binding epitope have a significant impact on pep-tide-specific CTL responses in HLA-A*0201/DR1 Tgmice.
We reasoned that if the HLA-A*0201-restricted CTLresponse was enhanced by a concomitant activation ofspecific CD41 cells by TT TH peptides, that preinjectionof HLA-A*0201/DR1 Tg mice with a TT peptide (THpriming) might further increase CTL responsiveness. Asignificant increase in pp65495–503 specific CTL wasobserved in mice prevaccinated with TT peptides, com-pared with control mice receiving preinjection with ad-juvant alone (p , 0.01) (Table 3). This was demonstratedwith two different TH 1 CTL peptide combinations;TT830–843 1 pp65495–503 and TT947–967 1pp65495–503 (Table 3). This suggests that TH can bederived from anamnestic or primary response to a definedTH epitope. Co-administration of a TT TH and CTLpeptide mixture was sufficient to activate a pp65495–503and virus-specific CTL response, and did not depend onwhether the CD41 response was predominantly a pri-mary or an anamnestic response.
DISCUSSIONSynthetic peptide-based CTL epitope vaccines have beendeveloped as immunotherapeutics against a number ofhuman diseases [13, 50, 51]. In this report, the HLA-DRrequirement for in vivo priming of HLA-A*0201 re-stricted CTL responses has been examined. Initially, sev-eral TT immunodominant TH epitopes (TT639–652,TT830–843, and TT947–967) among five examined wereshown to efficiently stimulate in vitro proliferation ofPBMC as evaluated in 43 randomly chosen human do-nors. Ten additional HLA-typed donors expressing awide array of HLA-DR alleles also recognized the sameTT peptides (L. BenMohamed, unpublished results).These results suggest that the use of “promiscuous”T-cell epitopes selected from TT to be incorporated intovaccines would likely satisfy the demand for immunoge-nicity as well as coverage [52, 53]. The ability of theseTT peptides to induce TH responses in vivo was con-firmed in both HLA-DR1 and HLA-A*0201/DR1 Tgmice. In addition, a single injection of a peptide mixtureconsisting of a CMV CTL epitope (pp65495–503) and oneof the three TT-derived TH epitopes resulted in a vig-orous CTL response, which was enhanced in mice ex-pressing both the HLA-DR1 and A*0201 genes. CTLinduction did not depend on whether the TH responsewas predominately a primary or anamnestic response.Finally, CTL responses in HLA-A*0201/DR1 Tg mice
FIGURE 6 CTL induced in HLA A*0201/DR1 Tg mice,immunized with CMV pp65495–503 peptide in combinationwith TT947–967 or TT830–843 TH epitopes, mediated lysis ofHLA A*0201- matched but not mismatched fibroblasts in-fected with the AD-169 strain of CMV (MOI 5 3). Lysis offibroblasts by either uninfected or infected with CMV (bymurine CTL) was measured in a standard 4-h CRA at the E:Tratios indicated.
774 L. BenMohamed et al.
were of CD81 CD42 phenotype, and resulted in specificlysis of human HLA-A*0201 positive CMV-infected fi-broblasts. Overall, the data show that use of HLA-A*0201/DR1 Tg mice offers a new animal model systemto study the role of HLA class II-restricted TH epitopesin the development of HLA class I restricted CTL re-sponses.
The peptide-based approach was pursued based onprevious studies showing that immunization with syn-thetic peptides can induce protective immunity in ex-perimental models of infection [8]. The capacity of CTLand TH antigenic peptides to efficiently induce CTLresponses has been demonstrated both in experimentalmodels [54, 55] and in humans [13, 14, 56]. In thisreport, it was shown that CTL which lyse CMV-infectedfibroblasts can be induced in HLA-A*0201/DR1 Tgmice after immunization with a mixture of pp65495–503CTL and TT TH synthetic peptides (Figure 6). It was alsofound that injection of a minimal CMV-specific CTLepitope (pp65495–503) into HLA-A*0201/DR1 Tg micefailed to induce peptide and virus-specific CTL responsesunless it was combined with a TH peptide (Table 2).These results support the concept that in vivo induction
of CD81 CTL responses using minimal antigenic pep-tides requires the stimulation of both CD41 and CD81
T cell subsets. Whether cognate CD41 T cell help willfurther enhance the induction and the magnitude ofCMV specific CTL responses as recently shown for severalmodels, remains to be determined [57, 58].
CTL are critical components of protective immunityagainst viral infections, but the requirements for in vivopriming of CTL are not completely understood. Al-though it is generally accepted that TH cells are essentialfor CTL priming with synthetic peptides, there has beenconflicting data from some experimental virus infections[59, 60]. With respect to synthetic peptides encodingCTL epitopes, several studies point to a mandatory needfor TH lymphocyte stimulation to induce optimal CTLresponses [54, 55, 57, 61]. In contrast, some 8- to12-mer synthetic peptides corresponding to the naturallyprocessed minimal CTL epitope, could exhibit in vivoCTL immunogenicity independent of TH cells [62–65].
In the present study, HLA-A*0201/DR1 Tg micewere used to address the question if a CMV minimal CTLepitope requires TH cell stimulation to induce CTLresponses. The pp65495–503 CTL epitope was coinjected
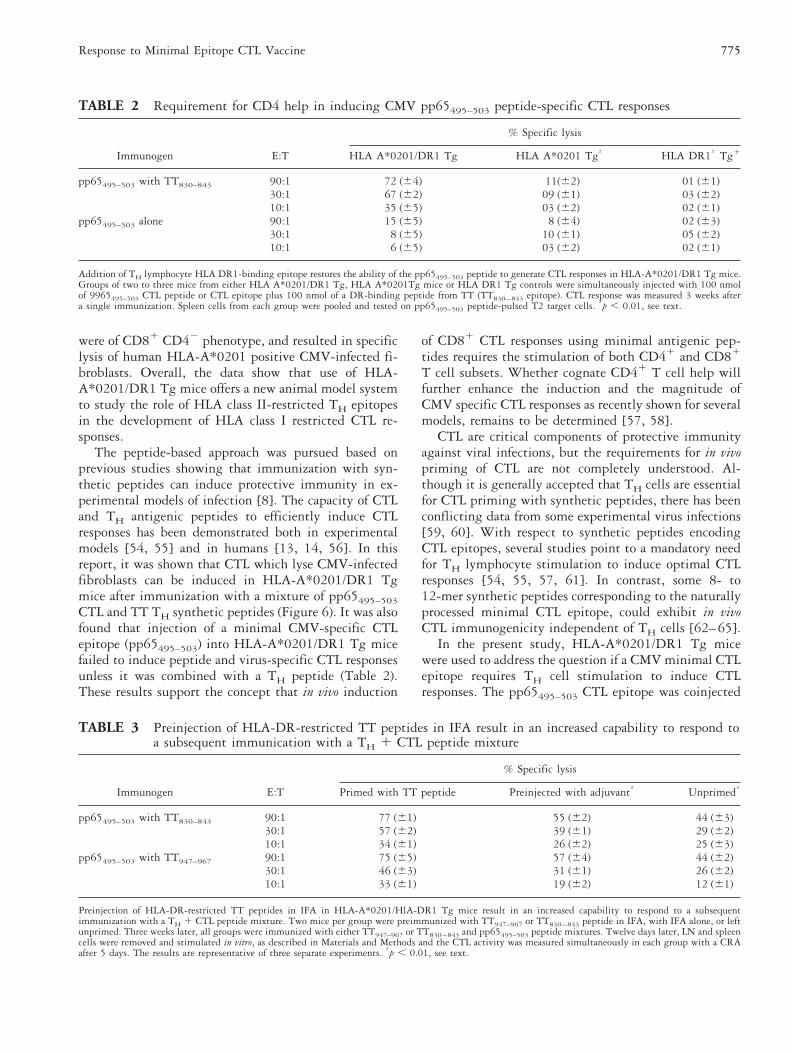
TABLE 2 Requirement for CD4 help in inducing CMV pp65495–503 peptide-specific CTL responses
Immunogen E:T
% Specific lysis
HLA A*0201/DR1 Tg HLA A*0201 Tg† HLA DR1† Tg1
pp65495–503 with TT830–843 90:1 72 (64) 11(62) 01 (61)30:1 67 (62) 09 (61) 03 (62)10:1 35 (65) 03 (62) 02 (61)
pp65495–503 alone 90:1 15 (65) 8 (64) 02 (63)30:1 8 (65) 10 (61) 05 (62)10:1 6 (65) 03 (62) 02 (61)
Addition of TH lymphocyte HLA DR1-binding epitope restores the ability of the pp65495–503 peptide to generate CTL responses in HLA-A*0201/DR1 Tg mice.Groups of two to three mice from either HLA A*0201/DR1 Tg, HLA A*0201Tg mice or HLA DR1 Tg controls were simultaneously injected with 100 nmolof 9965495–503 CTL peptide or CTL epitope plus 100 nmol of a DR-binding peptide from TT (TT830–843 epitope). CTL response was measured 3 weeks aftera single immunization. Spleen cells from each group were pooled and tested on pp65495–503 peptide-pulsed T2 target cells. †p , 0.01, see text.
TABLE 3 Preinjection of HLA-DR-restricted TT peptides in IFA result in an increased capability to respond toa subsequent immunication with a TH 1 CTL peptide mixture
Immunogen E:T
% Specific lysis
Primed with TT peptide Preinjected with adjuvant† Unprimed†
pp65495–503 with TT830–843 90:1 77 (61) 55 (62) 44 (63)30:1 57 (62) 39 (61) 29 (62)10:1 34 (61) 26 (62) 25 (63)
pp65495–503 with TT947–967 90:1 75 (65) 57 (64) 44 (62)30:1 46 (63) 31 (61) 26 (62)10:1 33 (61) 19 (62) 12 (61)
Preinjection of HLA-DR-restricted TT peptides in IFA in HLA-A*0201/HlA-DR1 Tg mice result in an increased capability to respond to a subsequentimmunization with a TH 1 CTL peptide mixture. Two mice per group were preimmunized with TT947–967 or TT830–843 peptide in IFA, with IFA alone, or leftunprimed. Three weeks later, all groups were immunized with either TT947–967 or TT830–843 and pp65495–503 peptide mixtures. Twelve days later, LN and spleencells were removed and stimulated in vitro, as described in Materials and Methods and the CTL activity was measured simultaneously in each group with a CRAafter 5 days. The results are representative of three separate experiments. †p , 0.01, see text.
775Response to Minimal Epitope CTL Vaccine

with or without CD41 TH epitopes derived from TTprotein into Tg mice expressing HLA-A*0201 and/orHLA-DR1 molecules. The results established that acti-vation of CMV specific CTL responses by the optimalpp65495–503 CTL peptide required DR-restricted T-cellhelp (Table 2). Significant enhancement of CTL re-sponses were observed in HLA-A*0201/DR1 Tg micecompared with HLA-A*0201 Tg mice, when bothgroups were immunized with the HLA-A*0201-re-stricted CTL epitope in combination with three differentCD41 TH epitopes from TT protein; TT639–652,TT830–843, and TT947–967 (Figure 5A). Preinjection ofTT639–652, TT830–843, or TT947–967 T helper peptidesincreased the capability of the Tg mice to respond to asubsequent immunization with a TH 1 CTL peptidemixture (Table 3). There was no stimulatory effect ofpreinjection with adjuvant alone on subsequent injectionwith the TH 1 CTL peptide mixture (Table 3). Thissuggests that, in the presence of a pre-existing responseto a defined TH epitope, administration of TH 1 CTLpeptide mixture may be sufficient to activate a specificanti-peptide CTL response. This could, in principle, beapplied to human vaccination, due to the fact that a largeproportion of the population is preimmunized with TT[21, 66, 67].
The notion that CD41 TH lymphocytes are critical formaintaining CMV-specific CTL function is not new. THepitopes might play an important role not only foroptimal induction of CTL responses, as reported in thisarticle, but also for maintenance of CTL memory asreported by others [68, 69]. In studies on the CTLresponse to CMV in BMT recipients receiving adoptivetransfer of autologous CD81 T cells, patients who main-tained strong CTL responses over time had recoveredendogenous CD41 TH lymphocytes, whereas those whodid not recover sufficient CD41 TH failed to maintaintheir CTL responses [68, 69]. Using in vivo depletion ofmurine CD41 T cells or MHC class II knockout mice,previous studies have demonstrated a major role formurine CD41 T cells in the induction and maintenanceof CD81 CTL responses [54, 57, 64]. Therefore, thera-peutic vaccination should be targeted towards enhancingboth CD41 and CD81 T cell virus-specific responses[11]. The present studies describe, for the first time, a Tgmouse model expressing both HLA class I (A*0201) andHLA class II (DRB1*0101) molecules in which the micewere fully capable of mounting both HLA class I and IIrestricted T-cell responses after immunization with CTLand TH peptides. Results of the present study suggestthat the HLA-A*0201/DR1 Tg mouse model may pro-vide insights in the role of the human MHC class II inthe induction, duration, and strength of class I restrictedCTL responses to prevent and/or control chronic viral andparasitic diseases.
Conventional strategies of peptide immunizationbased upon HLA-allele specific peptide motifs have lim-ited ability to elicit immune responses in the outbredhuman population of MHC haplotypes. This geneticallyrestricted recognition of CTL or TH epitopes is a seriousobstacle to vaccine design [70]. However, TH peptidesare usually more “promiscuous” in binding to MHC classII molecules than CTL epitopes are for MHC class I. Thisopens the possibility that the TH epitopes are less alleledependent and thereby require less variations for univer-sal recognition by humans. The use of “promiscuous”T-cell epitopes selected from TT would likely satisfy thedemand for immunogenicity as well as coverage [18, 20,52, 71]. To further reduce the number of individualvaccines tailored to individual MHC alleles, investigatorshave characterized HLA class I and II supertypes, whichare subsets of overlapping peptide-binding specificitiescomprising a high percentage of outbred populations[44, 52]. Therefore, a large proportion of the .200different known HLA class I and II alleles could begrouped into a few broad HLA supertypes (HLA super-motifs). Sette and collaborators [53] have suggested thatHLA-A*0201 is part of a group of HLA alleles, desig-nated the A3 supertype, which is comprised of over 40%of known HLA class I alleles. Investigators have alsorecently identified several HLA-DR supertypes charac-terized by overlapping peptide-binding specificities[44]. The HLA-DR1 allele used in this study belong tothe largest DR family, designated the DR1-4-7 super-type, which would encompass 50% to 80% of mostpolymorphic populations [44]. Because of the high rep-resentation of the A3 and DR supertypes in most majorethnic groups, development of the HLA-A*0201/DR1Tg mice would be of potentially broad significance.
In summary, we describe a model system using HLA-A*0201/DR1 Tg mice, which offers a system to addressthe role of HLA class II-restricted TH epitopes in thedevelopment of HLA class I restricted T-cell responses.In addition, we show experimental data highlighting theimportance of CD41 TH lymphocytes in the optimal invivo induction of CTLs against a representative viralpathogen, indicating that TH epitopes have to be in-cluded in the design of epitope-based vaccines againstCMV or other pathogens.
ACKNOWLEDGMENTS
The authors would like to thank the following people forassistance in conducting the studies described in this report:Dr. Susan Kovats (Division of Immunology, COH) for helpwith PCR analysis and flow cytometry; Alice Huang for experthistochemical analysis of mouse tissues (Department of Ana-tomic Pathology, COH); Dr. Corinna La Rosa and Julie Papp,M.S. (Department of Hematology Research, COH) for provid-ing fibroblasts, B-LCL, and titered CMV stocks; and Heather
776 L. BenMohamed et al.

Adams, Donna Isbell, and Dr. Herod Howard (Director) of theCOH Animal Care Facility for care and husbandry of thetransgenic mice used in these studies. Dr. Joshua Ellenhornand Dr. Michel Denis of COH are acknowledged for assistancein interpretation of DTH studies and a critique of several earlydrafts of the manuscript. Dr. Dennis Zaller (Merck, Sharp andDohme, Rahway, NJ, USA) is gratefully acknowledged forproviding animals to create a breeding colony of HLA-DR1mice for this study. The expert organization and typing of thismanuscript was coordinated by Rose-Marie Imstepf and CaroleSmith, who are gratefully acknowledged.
REFERENCES
1. Aichele P, Hengartner H, Zinkernagel RM, Schulz M:Antiviral cytotoxic T cell response induced by in vivopriming with a free synthetic peptide. J Exp Med 171:1815, 1990.
2. BenMohamed L, Gras-Masse H, Tartar A, Daubersies P,Brahimi K, Bossus M, Thomas A, Druhile P: Lipopeptideimmunization without adjuvant induces potent and long-lasting B, Thelper, and CTL responses against a malarialiver stage antigen in mice and chimpanzees. Eur J Im-munol 27:1242, 1997.
3. Diamond DJ, York J, Sun J, Wright CL, Forman SJ:Development of a candidate HLA A*0201 restricted pep-tide-based vaccine against human cytomegalovirus infec-tion. Blood 90:1751, 1997.
4. Kast WM, Roux L, Curren J, Blom HJ, Voordouw AC,Meloen RH, Kolakofsky D, Melief CJ: Protection againstlethal Sendai virus infection by in vivo priming of virus-specific cytotoxic T lymphocytes with a free syntheticpeptide. Proc Natl Acad Sci USA 88:2283, 1991.
5. Scalzo AA, Elliott SL, Cox J, Gardner J, Moss DJ, Suhr-bier A: Induction of protective cytotoxic T cells to murinecytomegalovirus by using a nonapeptide and a human-compatible adjuvant (Montanide ISA 720). J Virol 69:1306, 1995.
6. Feltkamp MC, Smits HL, Vierboom MP, Minnaar RP, deJongh BM, Drijfhout JW, ter Schegget J, Melief CJ, KastWM: Vaccination with cytotoxic T lymphocyte epitope-containing peptide protects against a tumor induced byhuman papillomavirus type 16-transformed cells. EurJ Immunol 23:2242, 1993.
7. Sarin PS, Mora CA, Naylor PH, Markham R, Schwartz D,Kahn J, Heseltine P, Gazzard B, Youle M, Rios A: HIV-1p17 synthetic peptide vaccine HGP-30: induction of im-mune response in human subjects and preliminary evi-dence of protection against HIV challenge in SCID mice.Cell Mol Biol (Noisy-le-grand) 41:401, 1995.
8. Franke ED, Corradin G, Hoffman SL: Induction of pro-tective CTL responses against the Plasmodium yoelii cir-cumsporozoite protein by immunization with peptides.J Immunol 159:3424, 1997.
9. Blaney JEJ, Nobusawa E, Brehm MA, Bonneau RH,Mylin LM, Fu TM, Kawaoka Y, Tevethia SS: Immuniza-
tion with a single major histocompatibility complex classI- restricted cytotoxic T-lymphocyte recognition epitopeof herpes simplex virus type 2 confers protective immu-nity. J Virol 72:9567, 1998.
10. van der Most RG, Sette A, Oseroff C, Alexander J, Murali-Krishna K, Lau LL, Southwood S, Sidney J, Chesnut RW,Matloubian M, Ahmed R: Analysis of cytotoxic T cellresponses to dominant and subdominant epitopes duringacute and chronic lymphocytic choriomeningitis virus in-fection. J Immunol 157:5543, 1996.
11. Zajac AJ, Murali-Krishna K, Blattman JN, Ahmed R:Therapeutic vaccination against chronic viral infection:the importance of cooperation between CD41 and CD81T cells. Curr Opin Immunol 10:444, 1998.
12. Firat H, Garcia-Pons F, Tourdot S, Pascolo S, Scardino A,Garcia Z, Michel ML, Jack RW, Jung G, KosmatopoulosK, Mateo L, Suhrbier A, Lemonnier FA, Langlade-De-moyen P: H-2 class I knockout, HLA-A2.1-transgenicmice: a versatile animal model for preclinical evaluation ofantitumor immunotherapeutic strategies [In Process Ci-tation]. Eur J Immunol 29:3112, 1999.
13. Vitiello A, Ishioka G, Grey HM, Rose R, Farness P,LaFond R, Yuan L, Chisari FV, Furze J, Bartholomeuz R:Development of a lipopeptide-based therapeutic vaccineto treat chronic HBV infection. I. Induction of a primarycytotoxic T lymphocyte response in humans. J Clin Invest95:341, 1995.
14. Livingston B, Crimi C, Grey H, Ishioka G, Chisari FV,Fikes J, Grey HM, Chesnut R, Sette A: The hepatitis Bvirus-specific CTL responses induced in humans by li-popeptide vaccination are comparable to those elicited byacute viral infection. J Immunol 159:1383, 1997.
15. Livingston B, Alexander J, Crimi C, Oseroff C, Celis E,Daly K, Guidotti LG, Chisari FV, Fikes J, Chesnut RW,Sette A: Altered helper T lymphocyte function associatedwith chronic hepatitis B virus infection and its role inresponse to therapeutic vaccination in humans. J Immunol162:3088, 1999.
16. Reece JC, McGregor D, Geysen HM, Rodda SJ: Scanningfor T helper epitopes with human PBMC using pools ofshort synthetic peptides. EMBO J 172:241, 1994.
17. Demotz S, Grey HM, Appella E, Sette A: Characterizationof a naturally processed MHC class II- restricted T- celldeterminant of hen egg lysozyme. Nature 342:682, 1989.
18. Panina-Bordignon P, Tan A, Termijtelen A, Demotz S,Corradin G, Lanzavecchia A: Universally immunogenic Tcell epitopes: promiscuous binding to human MHC classII and promiscuous recognition by T cells. Eur J Immunol19:2237, 1989.
19. Reece JC, Geysen HM, Rodda SJ: Mapping the majorhuman T helper epitopes of tetanus toxin. The emergingpicture. J Immunol 151:6175, 1993.
20. Valmori D, Sabbatini A, Lanzavecchia A, Corradin G,Matricardi PM: Functional analysis of two tetanus toxin
777Response to Minimal Epitope CTL Vaccine

universal T cell epitopes in their interaction with DR1101and DR1104 alleles. J Immunol 152:2921, 1994.
21. Valmori D, Pessi A, Bianchi E, Corradin G: Use of humanuniversally antigenic tetanus toxin T cell epitopes as car-riers for human vaccination. J Immunol 149:717, 1992.
22. Mizobata S, Clarke P, Mann J, Horton L, Primus J:Cytotoxic T cell responses for human carcinoembryonicantigen (CEA) epitopes in mice transgenic for HLAA*0201/kb and human CEA. Submitted.
23. Yamamoto K, Fukui Y, Esaki Y, Inamitsu T, Sudo T,Yamane K, Kamikawaji N, Kimura A, Sasazuki T: Func-tional interaction between human histocompatibility leu-kocyte antigen (HLA) class II and mouse CD4 molecule inantigen recognition by T cells in HLA-DR and DQ trans-genic mice [see comments]. J Exp Med 180:165, 1994.
24. Vignali DA, Moreno J, Schiller D, Hammerling GJ: Spe-cies-specific binding of CD4 to the b2 domain of majorhistocompatibility complex class II molecules. J Exp Med175:925, 1992.
25. Vitiello A, Marchesini D, Furze J, Sherman LA, ChesnutRW: Analysis of the HLA-restricted influenza-specificcytotoxic T lymphocyte response in transgenic mice car-rying a chimeric human- mouse class I major histocom-patibility complex. J Exp Med 173:1007, 1991.
26. Irwin MJ, Heath WR, Sherman LA: Species-restrictedinteractions between CD8 and the alpha 3 domain of classI influence the magnitude of the xenogeneic response. JExp Med 170:1091, 1989.
27. Theobald M, Biggs J, Hernandez J, Lustgarten J, LabadieC, Sherman LA: Tolerance to p53 by A2.1-restrictedcytotoxic T lymphocytes. J Exp Med 185:833, 1997.
28. Rosloniec EF, Brand DD, Myers LK, Whittington KB,Gumanovskaya M, Zaller DM, Woods A, Altmann DM,Stuart JM, Kang AH: An HLA-DR1 transgene conferssusceptibility to collagen-induced arthritis elicited withhuman type II collagen. J Exp Med 185:1113, 1997.
29. Woods A, Chen HY, Trumbauer ME, Sirotina A, Cum-mings R, Zaller DM: Human major histocompatibilitycomplex class II-restricted T cell responses in transgenicmice [see comments]. J Exp Med 180:173, 1994.
30. Vu TH, Begovich AB, Tacchini-Cottier FM, Jones PP:Molecular defects in the non-expressed H-2 E alpha genesof the f and q haplotypes. J Immunol 142:2936, 1989.
31. Begovich AB, Vu TH, Jones PP: Characterization of themolecular defects in the mouse E b f and E b q genes.Implications for the origin of MHC polymorphism. J Im-munol 144:1957, 1990.
32. Altmann DM, Douek DC, Frater AJ, Hetherington CM,Inoko H, Elliott JI: The T cell response of HLA-DRtransgenic mice to human myelin basic protein and otherantigens in the presence and absence of human CD4. JExp Med 181:867, 1995.
33. Hurley CK, Schreuder GM, Marsh SG, Lau M, MiddletonD, Noreen H: The search for HLA-matched donors: asummary of HLA-A*, -B*, - DRB1/3/4/5* alleles and
their association with serologically defined HLA-A, -B,-DR antigens. Tissue Antigens 50:401, 1997.
34. Krausa P, Brywka M3, Savage D, Hui KM, Bunce M,Ngai JL, Teo DL, Ong YW, Barouch D, Allsop CE, et al.:Genetic polymorphism within HLA-A*02: significant al-lelic variation revealed in different populations. TissueAntigens 45:223, 1995.
35. Wei ML, Cresswell P: HLA-A2 molecules in an antigen-processing mutant cell contain signal sequence-derivedpeptides [see comments]. Nature 356:443, 1992.
36. Fidock DA, Gras-Masse H, Lepers JP, Brahimi K, Ben-Mohamed L, Mellouk S, Guerin-Marchand C, Londono A,Raharimalala L, Meis JF: Plasmodium falciparum liverstage antigen-1 is well conserved and contains potent Band T cell determinants. [Published erratum appears inJ Immunol 1994;153(11):5347] J Immunol 153:190,1994.
37. Bottius E, BenMohamed L, Brahimi K, Gras H, LepersJP, Raharimalala L, Aikawa M, Meis J, Slierendregt B,Tartar A, Thomas A, Druilhe P: A novel Plasmodiumfalciparum sporozoite and liver stage antigen (SALSA)defines major B, T helper, and CTL epitopes. J Immunol156:2874, 1996.
38. Kikuta K, Kurata H, Nagamine T, Aizawa C, Ueno Y,Kurata T, Tamura S: Enhancement of DTH response bycholera toxin B subunit inoculated intranasally togetherwith influenza HA vaccine. Microbiol Immunol 34:337,1990.
39. Moutouh L, Corbeil J, Richman DD: Recombinationleads to the rapid emergence of HIV-1 dually resistantmutants under selective drug pressure. Proc Natl Acad SciUSA 93:6106, 1996.
40. Wentworth PA, Celis E, Crimi C, Stitely S, Hale L, TsaiV, Serra HM, Del Guercio MF, Livingston B, Alazard D,et al.: In vitro induction of primary, antigen-specific CTLfrom human peripheral blood mononuclear cells stimu-lated with synthetic peptides. Mol Immunol 32:603,1995.
41. Ho RJ, Burke RL, Merigan TC: Physical and biologicalcharacterization of antigen presenting liposome formula-tions: relative efficacy for the treatment of recurrent gen-ital HSV-2 in guinea pigs. Antiviral Res 13:187, 1990.
42. Lee TD, Vemuri S: MacProMass: a computer program tocorrelate mass spectral data to peptide and protein struc-tures. Biomed Environ Mass Spectrom 19:639, 1990.
43. Wentworth PA, Vitiello A, Sidney J, Keogh E, ChesnutRW, Grey H, Sette A: Differences and similarities in theA2.1-restricted cytotoxic T cell repertoire in humans andhuman leukocyte antigen-transgenic mice. Eur J Immu-nol 26:97, 1996.
44. Southwood S, Sidney J, Kondo A, Del Guercio MF, Ap-pella E, Hoffman S, Kubo RT, Chesnut RW, Grey HM,Sette A: Several common HLA-DR types share largelyoverlapping peptide binding repertoires. J Immunol 160:3363, 1998.
778 L. BenMohamed et al.

45. Geluk A, Taneja V, van Meijgaarden KE, Zanelli E,Abou-Zeid C, Thole JE, de Vries RR, David CS, Otten-hoff TH: Identification of HLA class II-restricted deter-minants of Mycobacterium tuberculosis-derived proteinsby using HLA-transgenic, class II-deficient mice. ProcNatl Acad Sci USA 95:10797, 1998.
46. Ho PC, Mutch DA, Winkel KD, Saul AJ, Jones GL,Doran TJ, Rzepczyk CM: Identification of two promiscu-ous T cell epitopes from tetanus toxin. Eur J Immunol20:477, 1990.
47. Men Y, Thomasin C, Merkle HP, Gander B, Corradin G:A single administration of tetanus toxoid in biodegrad-able microspheres elicits T cell and antibody responsessimilar or superior to those obtained with aluminumhydroxide. Vaccine 13:683, 1995.
48. Alexander J, Sidney J, Southwood S, Ruppert J, Oseroff C,Maewal A, Snoke K, Serra HM, Kubo RT, Sette A:Development of high potency universal DR-restrictedhelper epitopes by modification of high affinity DR-blocking peptides. Immunity 1:751, 1994.
49. La Rosa C, Forman SJ, Diamond DJ: Development of anin vitro stimulation protocol to measure CMV specificCTL activity using HLA-restricted CTL epitopes. Blood92:518a, 1998.
50. Wentworth P, Sette A, Southwood S, del Guercio M,Wong N, Kubo RT, Volenec J, Walker BD, Kalams S,Cotton D, Chesnut RW: Development of a CTL epitope-based vaccine for the treatment of HIV infection. ConfAdv AIDS Vaccine Dev XX:60, 1997.
51. Elliott SL, Pye S, Le T, Mateo L, Cox J, Macdonald L,Scalzo AA, Forbes CA, Suhrbier A: Peptide based cyto-toxic T-cell vaccines: delivery of multiple epitopes, help,memory and problems. Vaccine 17:2009, 1999.
52. Sidney J, Del Guercio MF, Southwood S, Engelhard VH,Appella E, Rammensee HG, Falk K, Rotzschke O,Takiguchi M, Kubo RT: Several HLA alleles share over-lapping peptide specificities. J Immunol 154:247, 1995.
53. Sidney J, Southwood S, Del Guercio MF, Grey HM,Chesnut RW, Kubo RT, Sette A: Specificity and degen-eracy in peptide binding to HLA-B7-like class I mole-cules. J Immunol 157:3480, 1996.
54. Fayolle C, Deriaud E, Leclerc C: In vivo induction ofcytotoxic T cell response by a free synthetic peptide re-quires CD41 T cell help. J Immunol 147:4069, 1991.
55. Widmann C, Romero P, Maryanski JL, Corradin G, Val-mori D: T helper epitopes enhance the cytotoxic responseof mice immunized with MHC class I-restricted malariapeptides. J Immunol Meth 155:95, 1992.
56. Brander C, Corradin G, Hasler T, Pichler WJ: Peptideimmunization in humans: a combined CD81/CD41 Tcell-targeted vaccine restimulates the memory CD4 T cellresponse but fails to induce cytotoxic T lymphocytes(CTL). Clin Exp Immunol 105:18, 1996.
57. Shirai M, Pendleton CD, Ahlers J, Takeshita T, NewmanM, Berzofsky JA: Helper-cytotoxic T lymphocyte (CTL)
determinant linkage required for priming of anti-HIVCD81 CTL in vivo with peptide vaccine constructs.J Immunol 152:549, 1994.
58. Bennett SR, Carbone FR, Karamalis F, Miller JF, HeathWR: Induction of a CD81 cytotoxic T lymphocyte re-sponse by cross-priming requires cognate CD41 T cellhelp. J Exp Med 186:65, 1997.
59. Keene JA, Forman J: Helper activity is required for the invivo generation of cytotoxic T lymphocytes. J Exp Med155:768, 1982.
60. Zhou X, Berglund P, Zhao H, Liljestrom P, Jondal M:Generation of cytotoxic and humoral immune responsesby nonreplicative recombinant Semliki Forest virus. ProcNatl Acad Sci USA 92:3009, 1995.
61. Sauzet JP, Gras-Masse H, Guillet JG, Gomard E: Influ-ence of strong CD4 epitope on long-term virus-specificcytotoxic T cell responses induced in vivo with peptides.Int Immunol 8:457, 1996.
62. Zhou X, Berg L, Motal UM, Jondal M: In vivo primaryinduction of virus-specific CTL by immunization with9-mer synthetic peptides. J Immunol Meth 153:193,1992.
63. Fayolle C, Abdel-Motal UM, Berg L, Deriaud E, JondalM, Leclerc C: Induction of cytotoxic T-cell response byoptimal-length peptides does not require CD41 T-cellhelp. Immunology 89:41, 1996.
64. Gao XM, Zheng B, Liew FY, Brett S, Tite J: Priming ofinfluenza virus-specific cytotoxic T lymphocytes vivo byshort synthetic peptides. J Immunol 147:3268, 1991.
65. Dyall R, Vasovic LV, Molano A, Nikolic-Zugic J: CD4-independent in vivo priming of murine CTL by optimalMHC class I- restricted peptides derived from intracellu-lar pathogens. Int Immunol 7:1205, 1995.
66. Etlinger HM, Gillessen D, Lahm HW, Matile H, Schon-feld HJ, Trzeciak A: Use of prior vaccinations for thedevelopment of new vaccines. Science 249:423, 1990.
67. Reece JC, McGregor DL, Geysen HM, Rodda SJ: Scan-ning for T helper epitopes with human PBMC using poolsof short synthetic peptides. J Immunol Meth 172:241,1994.
68. Walter EA, Greenberg PD, Gilbert MJ, Finch RJ, Wa-tanabe KS, Thomas ED, Riddell SR: Reconstitution ofcellular immunity against cytomegalovirus in recipientsof allogeneic bone marrow by transfer of T-cell clonesfrom the donor. N Engl J Med 333:1038, 1995.
69. Riddell SR, Greenberg PD: Adoptive immunotherapywith antigen-specific T cells. In Thomas ED, Blume KG,Forman SJ (eds): Hematopoietic Cell Transplantation,2nd edn. Malden, MA: Blackwell Science Inc., 1999.
70. Letvin NL: Progress in the development of an HIV-1vaccine [see comments]. Science 280:1875, 1998.
71. Demotz S, Lanzavecchia A, Eisel U, Niemann H, Wid-mann C, Corradin G: Delineation of several DR-re-stricted tetanus toxin T cell epitopes. J Immunol 142:394, 1989.
779Response to Minimal Epitope CTL Vaccine