
Individual differences in local gray and white matter volumes reflect differences in temperament and...
12
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of Individual differences in local gray and white matter volumes reflect differences in temperament and...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Research Report
Individual differences in local gray and white matter volumesreflect differences in temperament and character: A voxel-basedmorphometry study in healthy young females
Peter Van Schuerbeeka,⁎, Chris Baekenb, Rudi De Raedtc, Johan De Meya, Rob Luypaerta
aDepartment of Radiology, UZ Brussel, Vrije Universiteit Brussel (VUB), Laarbeeklaan 101, 1090 Brussels, BelgiumbDepartment of Psychiatry, UZ Brussel, Vrije Universiteit Brussel (VUB), Laarbeeklaan 101, 1090 Brussels, BelgiumcDepartment of Experimental Clinical and Health Psychology, Ghent University, Henri Dunantlaan 2, 9000 Ghent, Belgium
A R T I C L E I N F O A B S T R A C T
Article history:Accepted 19 November 2010Available online 29 November 2010
The psychobiological personality model of Cloninger distinguishes four heritabletemperament traits (harm avoidance (HA), novelty seeking (NS), reward dependence (RD)and persistence (P)) and three character traits (self-directedness (SD), cooperativeness (CO)and self-transcendence (ST)) which develop during lifetime. Prior research already showedthat individual differences in temperament are reflected in structural variances in specificbrain areas. In this study, we used voxel-basedmorphometry (VBM) to correlate the differenttemperament and character traits with local gray and white matter volumes (GMV andWMV) in young healthy female volunteers. We found correlations between thetemperament traits and GMV and WMV in the frontal, temporal and limbic regionsinvolved in controlling and generating the corresponding behavior as proposed inCloninger's theory: anxious for HA, impulsive for NS, reward-directed for RD and goal-directed for P. The character traits correlated with GMV and WMV in the frontal, temporaland limbic regions involved in the corresponding cognitive tasks: self-reflection for SD,mentalizing and empathizing with others for CO and religious belief for ST. This studyshows that individual variations in brain morphology can be related to the temperamentand character dimensions, and lends support to the hypothesis of a neurobiological basis ofpersonality traits.
© 2010 Elsevier B.V. All rights reserved.
Keywords:PersonalityTemperamentCharacterVBMBrain
B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2
⁎ Corresponding author. Fax: +32 2 477 53 90.E-mail addresses: [email protected] (P. Van Schuerbeek), [email protected] (C. Baeken),
[email protected] (R. De Raedt), [email protected] (J. De Mey), [email protected] (R. Luypaert).Abbreviations: WMV, white matter volume; GMV, gray matter volume; HA, harm avoidance; NS, novelty seeking; RD, reward
dependence; P, persistence; SD, self-directedness; CO, cooperativeness; ST, self-transcendence; PD, personality disorder; MD, mooddisorder; ACC, anterior cingulate cortex; PCC, posterior cingulate cortex; MFC, medial frontal cortex; PFC, prefrontal cortex; OFC,orbitofrontal cortex; DTI, Diffusion tensor imaging; MTR, Magnetic transfer ratio
0006-8993/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.brainres.2010.11.073
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res

Author's personal copy
1. Introduction
Biopsychosocial theories propose that personality traits arerelated to genetic and neurobiological markers. In the late1980s and early 1990s, Robert Cloninger presented hispsychobiological model of personality to account for normaland abnormal variations in temperament and characterbetween individuals (Cloninger, 1986, 1987; Cloninger et al.,1993). This model focused on the structure of learning abilitieswithin the person, based on the neurobiology of functionalbrain systems to account for the evolution and function of thebrain. He distinguished four temperament dimensions (harmavoidance (HA), novelty seeking (NS), reward dependence (RD)and persistence (P)) and three character dimensions (self-directedness (SD), cooperativeness (CO) and self-transcendence(ST)). HA, NS, RD and P respectively reflect one's tendency toinhibit behavior to avoid punishment, to initiate a behaviortowards novelty, to generate a reward related behavior and tomaintainanongoingbehavior despite anabsenceof reward. SD,CO and ST respectively describes one's view on the self as anautonomous individual, as an integral part of human societyand as a part of a broader universe including its tendencytowards spiritualism.
Both major components of personality, temperament andcharacter, are differentiated by their biologic and socialdeterminants. The temperament traits are found to begenetically determined, heritable and relatively stable duringlifetime. It is postulated that they determine one's dispositionto the early emotions of fear, anger and attachment and one'sautomatic behavioral responses to the environmental stimuliof danger, novelty and reward. The character traits are thecomplex result of learning, maturation and socio-culturalfactors and evolve during the entire life. They were added tothe model to account for individual differences in themodulation of automatic behavioral responses, regulated bytemperament, by changes in the significance and salience ofstimuli due to the individual concepts of identity. It ispostulated that the character traits are involved in highercognitive processes.
In this two component model, character adapts one'sheritable personality traits and behavioral patterns to one'sown socio-cultural environment and experiences which mayprevent the maladaptive impact of the temperament traits.
To measure all personality dimensions for clinical andresearch purpose, the Temperament and Character Inventory(TCI) questionnaire was developed (Cloninger et al., 1993).
The Cloninger personality model has been introduced todistinguish normal from abnormal personality. In general,patients suffering froma personality disorder (PD) are found tobe characterized by immature character traits and extremetemperament traits. In clinical practice, the temperament aswell as the character dimensions were found to be of interestin the assessment and the differential diagnosis of PDs(Cloninger et al., 1993; Richter and Brändström, 2009; Svrakicet al., 2002). Furthermore, all seven personality dimensionshave proven to be useful in understanding the epidemiology,co-morbidity and treatment of mental illnesses such as mooddisorders (MD) (Cloninger et al., 1998, 2006; Farmer and Seeley,2009).
In addition to changes in mood and personality, local brainmorphology and functional alterations in PD and MD patientswere revealed in several brain imaging studies (Drevets et al.,2008; Goodman et al., 2007; McCloskey et al., 2005). Toinvestigate a possible link between structural abnormalitiesand personality traits, Iidaka et al. (2006) and Gardini et al.(2009) conducted a voxel-based morphometry (VBM) study onhealthy young adults, not suffering from a major mentalillness. Both studies showed evidence for a correlation betweenindividual structural variances in specific brain regions and thetemperament dimensions. They hypothesized that the struc-tural variances of interest, related to personality, are geneticallydetermined so the character dimensions were not taken intoaccount.
Kaasinen et al. (2005) conducted a similar VBM study inolder adults to investigate a possible correlation betweenmorphologic alterations in the brain and personality changesdue to aging. In this study all personality traits were taken intoaccount but only a correlation between ST and local graymatter volume (GMV) in the temporal cortex was found.
To correlate brain function to personality, Sugiura et al.(2000) used the SPECT imaging technique to voxel-wisecorrelate the temperament traits to cerebral blood flow. Ina replication of this study, but with all seven personalitydimensions included, Turner et al. (2003) found that individualdifferences in brain function in some regions may not onlyreflect differences in temperament but also differences incharacter.
These studies suggest that meaningful associations existbetween neurobiology and personality variance. However,the findings of Kaasinen et al. (2005) and Turner et al. (2003)suggest that the expression of personality traits may be relatedto neurodevelopmental variability, and the work of Iidaka et al.(2006) and Gardini et al. (2009) took only into account thetemperament traits that are genetic determined and believed tobe rather stable. Therefore, our aim was to go beyond thesestudies, extendingouranalysis to the characterdimensions thatdevelop over the first decades of life. We hypothesized that acorrelate of one's temperament and character, as described byCloninger's model, can be found in local brain morphology. Toexclude confounding variations of age and gender in brainmorphology andpersonality scores (Goodet al., 2001;Miettunenet al., 2007; Yamasue et al., 2008a,b), only females in a narrowage rangewere included. Due to the fact that the character traitsdevelop during maturation and maturation in the brain isshown to influence local GMV as well as white matter volumes(WMV) (Lenroot and Giedd, 2006), we performed our analysis onboth GMV and WMVmaps.
2. Results
In our final cohort of 68 subjects, themeanTCI scoreswere 13±6 for HA, 22±6 for NS, 19±3 for RD, 5±2 for P, 35±5 for SD, 36±4for CO, and 8±6 for ST.
In Tables 1 and 2 the correlations found for the tempera-ment and character traits respectively were summarized. Theresults from the GMV and WMV analyses are indicated byGMV and WMV, respectively. The coordinates reported in thetables are Talairach coordinates.
33B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2
Author's personal copy
3. Discussion
In the present study we investigated individual regional GMVand WMV differences related to all seven personality dimen-sions of Cloninger's model in young female adults. This studyshows that individual variations in brain morphology can berelated to the temperament dimensions as well as to thecharacter dimensions. These results lend support to thehypothesis of a neurobiological basis of personality traits as
already suggested by other authors (Iidaka et al., 2006; Gardiniet al., 2009; Sugiura et al., 2000; Turner et al., 2003).
We note that our results suggest that both temperament andcharacter are reflected inneuroanatomy, unless their differencesin nature. These results suggest that brain maturation isinfluenced by both the genetic as well as the environmentaldetermined component of personality. In twin and familystudies, the influence of genetic and environmental factors onbrain development, maturation and aging in a regional specificmanner has been revealed. It has been shown that these
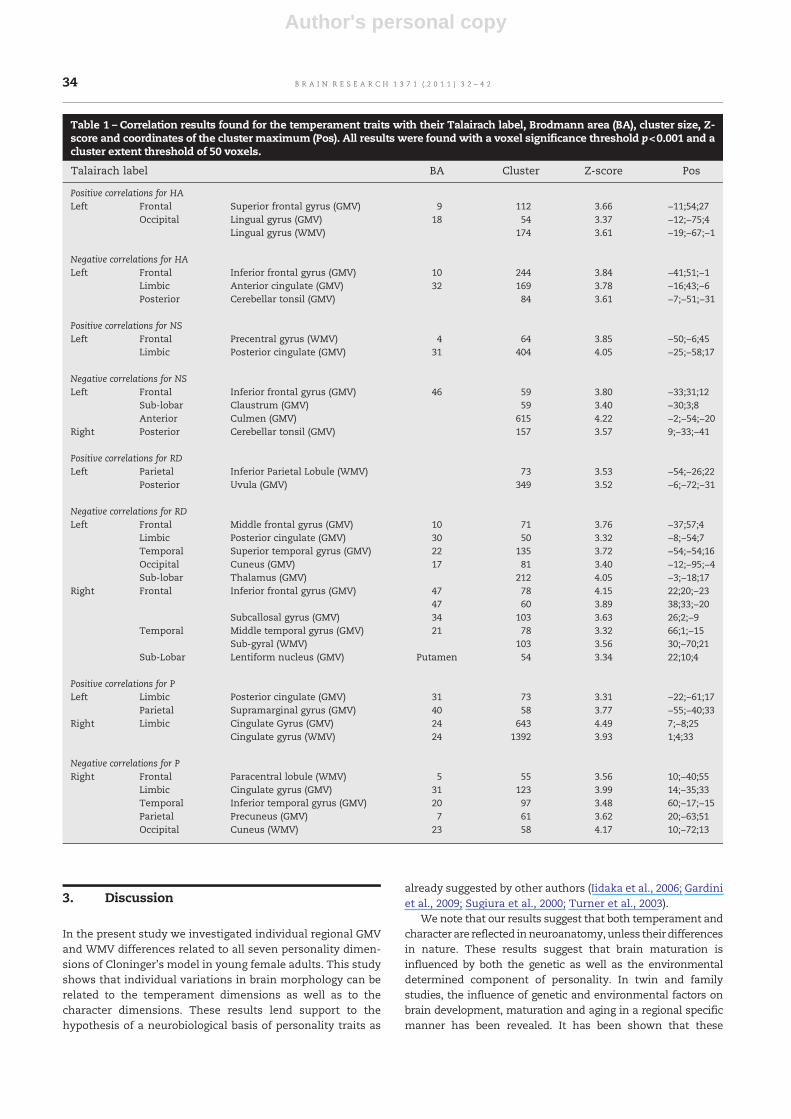
Table 1 – Correlation results found for the temperament traits with their Talairach label, Brodmann area (BA), cluster size, Z-score and coordinates of the cluster maximum (Pos). All results were found with a voxel significance threshold p<0.001 and acluster extent threshold of 50 voxels.
Talairach label BA Cluster Z-score Pos
Positive correlations for HALeft Frontal Superior frontal gyrus (GMV) 9 112 3.66 −11;54;27
Occipital Lingual gyrus (GMV) 18 54 3.37 −12;−75;4Lingual gyrus (WMV) 174 3.61 −19;−67;−1
Negative correlations for HALeft Frontal Inferior frontal gyrus (GMV) 10 244 3.84 −41;51;−1
Limbic Anterior cingulate (GMV) 32 169 3.78 −16;43;−6Posterior Cerebellar tonsil (GMV) 84 3.61 −7;−51;−31
Positive correlations for NSLeft Frontal Precentral gyrus (WMV) 4 64 3.85 −50;−6;45
Limbic Posterior cingulate (GMV) 31 404 4.05 −25;−58;17
Negative correlations for NSLeft Frontal Inferior frontal gyrus (GMV) 46 59 3.80 −33;31;12
Sub-lobar Claustrum (GMV) 59 3.40 −30;3;8Anterior Culmen (GMV) 615 4.22 −2;−54;−20
Right Posterior Cerebellar tonsil (GMV) 157 3.57 9;−33;−41
Positive correlations for RDLeft Parietal Inferior Parietal Lobule (WMV) 73 3.53 −54;−26;22
Posterior Uvula (GMV) 349 3.52 −6;−72;−31
Negative correlations for RDLeft Frontal Middle frontal gyrus (GMV) 10 71 3.76 −37;57;4
Limbic Posterior cingulate (GMV) 30 50 3.32 −8;−54;7Temporal Superior temporal gyrus (GMV) 22 135 3.72 −54;−54;16Occipital Cuneus (GMV) 17 81 3.40 −12;−95;−4Sub-lobar Thalamus (GMV) 212 4.05 −3;−18;17
Right Frontal Inferior frontal gyrus (GMV) 47 78 4.15 22;20;−2347 60 3.89 38;33;−20
Subcallosal gyrus (GMV) 34 103 3.63 26;2;−9Temporal Middle temporal gyrus (GMV) 21 78 3.32 66;1;−15
Sub-gyral (WMV) 103 3.56 30;−70;21Sub-Lobar Lentiform nucleus (GMV) Putamen 54 3.34 22;10;4
Positive correlations for PLeft Limbic Posterior cingulate (GMV) 31 73 3.31 −22;−61;17
Parietal Supramarginal gyrus (GMV) 40 58 3.77 −55;−40;33Right Limbic Cingulate Gyrus (GMV) 24 643 4.49 7;−8;25
Cingulate gyrus (WMV) 24 1392 3.93 1;4;33
Negative correlations for PRight Frontal Paracentral lobule (WMV) 5 55 3.56 10;−40;55
Limbic Cingulate gyrus (GMV) 31 123 3.99 14;−35;33Temporal Inferior temporal gyrus (GMV) 20 97 3.48 60;−17;−15Parietal Precuneus (GMV) 7 61 3.62 20;−63;51Occipital Cuneus (WMV) 23 58 4.17 10;−72;13
34 B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2

Author's personal copy
influences can be found throughout the whole life (Lenroot andGiedd, 2008; Schmitt et al., 2007). In current theories, the genesdonot directly code for behavior and cognition but rather define theprocesses that promote neuronal cell growth and their mutualinteractions that give rise to the observed behaviors. In otherwords, it has beenhypothesized that the brain is an intermediatefactor between behavior, cognition and genes and that brainmorphology and function reflect individual differences inbehavior and cognition (Plomin and Kosslyn, 2001).
In the personality theory of Cloninger, the temperamenttraits define one's automatic responses to emotional stimuliwhile character modulates these responses through cognitiveprocesses. The processing of these stimuli and generatingthese responses take place by neuronal processes in the brain.As hypothesized, our results suggest that one's personalityhas an influence on how his brain develop and mature. Thestudies of Gardini et al. (2009) and Iidaka et al. (2006) alreadyshowed the likeliness of this hypothesis.
In the following paragraphs we will give a brief discussionfor each personality trait separately.
3.1. Harm avoidance
Compared to the results of Gardini et al. (2009) and Yamasueet al. (2008a), a negative correlation between HA and GMV inthe left prefrontal cortex (PFC) was found. Consistent with theresults of Iidaka et al. (2006) a positive correlationwas found inthe left orbitofrontal cortex (OFC). The possible relation
between the amygdala GMV and HA, as found by Iidaka et al.(2006), or the hippocampus, as found by Yamasue et al.(2008a), was not found. Moreover, none of these authors didestablish a correlation between GMV and HA in the anteriorcingulate cortex (ACC) as we did.
In clinical practice, HA is used to probe an anxiety-relatedpersonality trait (Jylha and Isometsa, 2006). A neural networkincluding the ACC and the PFC is involved in the perceptionand regulation of fear and anxiety (LeDoux, 2000). A key playerin this network is theamygdalawhich is involved in generatingautonomic responses to threatening stimuli. The amygdalaresponse is controlled and modulated through connectionsfrom theACCand frontal cortex. In a study of Spampinato et al.(2009), a negative correlation between GMV in the ACC and thefrontal cortex and self-reported anxiety measures have beenrevealed. Interesting to note is that studies on patients with ananxiety disorder reported morphological abnormalities invarious brain regions including a reduction of GMV in theACC and frontal cortex (Ferrari et al., 2008).
We conclude that our results provide further evidence for apossible relation between HA and individual variations inbrain structure in the ACC and frontal cortex (see Fig. 1), bothinvolved in anxiety-related behaviors.
3.2. Novelty seeking
In line with the study of Gardini et al. (2009) we found acorrelation between NS and GMV in the motor (BA4) and
Table 2 – Correlation results found for the character traits with their Talairach label, Brodmann area (BA), cluster size, Z-scoreand coordinates of the cluster maximum (Pos). All results were found with a voxel significance threshold p<0.001 and acluster extent threshold of 50 voxels.
Talairach label BA Cluster Z-score Pos
Positive correlations for SDRight Temporal Superior temporal gyrus (GMV) 38 57 3.24 32;6;−28
Occipital Middle occipital gyrus (GMV) 37 201 3.57 37;−67;−1
Negative correlations for SDLeft Frontal Middle frontal gyrus (GMV) 8 53 3.46 −46;21;38
Sub-gyral (WMV) 521 3.74 −24;34;6Temporal Sub-gyral (WMV) 75 3.60 −39;−38;2Sub-lobar Extra-nuclear (WMV) 887 3.95 −21;−26;25Anterior Culmen (GMV) 164 3.39 −5;−48;−7
Right Limbic Cingulate gyrus (WMV) 690 4.21 19;−15;30
Positive correlations for CORight Temporal Superior temporal gyrus (GMV) 22 71 3.65 59;1;4
Negative correlations for COLeft Frontal Precentral gyrus (WMV) 4 258 4.22 −19;−29;72
Posterior Inferior semi-lunar lobule (GMV) 76 3.27 −3;−63;−45Right Frontal Medial frontal gyrus (WMV) 54 3.37 15;−1;59
Positive correlations for STRight Temporal Middle temporal gyrus (GMV) 21 64 3.41 52;2;−14
Parietal Inferior parietal gyrus (GMV) 40 469 4.31 58;2;−14
Negative correlations for STLeft Frontal Superior frontal gyrus (GMV) 10 496 4.28 −15;64;18
Temporal Inferior temporal gyrus (GMV) 20 106 3.82 −65;−35;−19Parietal Sub-gyral (WMV) 55 3.67 −12;−35;46
Right Frontal Superior frontal gyrus (GMV) 10 111 3.63 3;61;24
35B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2

Author's personal copy
posterior cingulate cortex (PCC). In linewith the results of Iidakaet al. (2006) we found a correlation between NS and GMV inthe left PFC. These findings are of interest as that the theoryof Cloninger (1987) defined NS as one's tendency to initiatebehaviors such as impulsivity and exploratory activity inresponse to novelty, while the PFC and PCC have been shownto be part of a neuronal network for taking impulsive decisions,response inhibition and impulsive control (Kalenscher et al.,2006; Crews and Boettiger, 2009; Sripada et al., 2010; Kim andLee, 2010). Our results were not in line with the results ofMatsuo et al. (2008) who correlated GMV to the subjects' traitimpulsivity and found strong correlations in the orbitofrontalcortex (OFC).
Previous studies linking personality to brain structuresremoved the cerebellum prior to analysis. In this study wetook the cerebellum into account and found a negativecorrelation with NS in the GMV of the left vermis. This is inline with studies on patients suffering an attention/hyperac-tivity disorder (ADHD) who were found to score high on NS
(Anckarsäter et al., 2006) and showed reduced GMV in thevermis and the frontal cortex.
We conclude that our results give support to a possiblerelation between individual variations in brain structure andNS in frontal and cingulate regions (see Fig. 2) known to beinvolved in generating an impulsive behavior. Future researchshould take the cerebellum into account as well to confirm ourresults for this region and to reveal its role in attentional andimpulsive control.
3.3. Reward dependence
In contrast to the results of Gardini et al. (2009) and Iidaka et al.(2006) we did not found any correlation between local GMVand RD in the right caudate nucleus and we found a negativecorrelation while Iidaka et al. (2006) reported a positive cor-relation in the right temporal cortex . On the other hand, ourresults showed correlations between RD and GMV in the OFC,temporal cortex and in the putamen as reported by Lebretonet al. (2009).
In the theory of Cloninger, RD is associated with one'stendency to respond intensely to social rewards (e.g. gettingapproval) (Cloninger, 1987). In the brain, social rewards areprocessed using the same fronto-limbic neural circuit asprimary (e.g. getting food when hungry) and secondary (e.g.winning money) rewards. Key regions in this reward systemare the dopaminergic neurons in the basal ganglia for rewardprediction and adaptation, the thalamus for reward consump-tion and the PFC and OFC for guiding social and goal-directedbehavior (Behrens et al., 2009; O'Doherty, 2004; Rademacheret al., 2010).
Outside the reward circuit, we found correlations betweenRD and GMV in temporal and occipital regions. In the study ofMobbs et al. (2003) the temporal cortex has been found to beinvolved in processing humor which is considered as a socialreward. Other examples of social rewards are the others smile,glance and laugh. For the perception of the others facialexpressions, eye gaze and speech, a functional networkstarting in the occipital and temporal cortex towards limbicand frontal areas have been revealed (Belin et al., 2004; Haxbyet al., 2002).
Fig. 1 – Correlation results found for the temperament traitHA overlaid on a T1 MNI template image in the ACC and PFC.
Fig. 2 – Correlation results found for the temperament trait NS overlaid on a T1 MNI template image in the PFC (A), the PCC(B) and the vermis (C).
36 B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2

Author's personal copy
Weconclude that our results support evidence for a relationbetween RD and GMV in the frontal, limbic and temporal areas(see Fig. 3) involved in social interactions, reward processingand generating a social behavior.
3.4. Persistence
In the first version of Cloninger's model, P was considered tobe a subscale of RD (Cloninger, 1987). In a later version P wasadded as an independent temperament trait (Cloninger et al.,1993).
Iidaka et al. (2006) and Gardini et al. (2009) included P as anindependent trait, but no positive correlations with GMV werefound by Iidaka et al. (2006) while Gardini et al. (2009) foundsignificant positive correlations with GMV in the precuneus,the paracentral lobule and the parahippocampal gyrus. Theselatter results could not be confirmed by our study, althoughin our study a negative correlation with GMV was found in
the precuneus and with WMV in the paracentral lobule. Inaddition P showed a correlation with GMV and WMV in thecingulate gyrus (see Fig. 4).
To generate persistent, goal-directed actions it is importantto be able tomake predictions of future rewards and to balancethese rewards to the costs and risks to reach them. Thesepredictions and actions should be updated constantly due tochanging situations and errors. In the brain, the cingulategyrus has been shown to be a key player in decisionmaking asit weights costs and benefits of actions, monitors errors and isinvolved in adapting the action value to changing environ-ments by evaluating the new outcome to the expected andprevious rewards (Rushworth and Behrens, 2008).
We conclude that our results suggest a possible link betweenGMV and P in the neural network involved in generating apersistent, goal-directed behavior.
3.5. Self-directedness
In Cloninger's theory SD is associated with one's view onthe self (Cloninger et al., 1993). This self-judgment results inreflective self appraisals about our actions, abilities, attitudesand emotions and our beliefs about how we are perceived byothers. Imaging studies investigating brain activity duringself-reflecting tasks have consistently revealed a neuralnetwork including the ACC, the posterior cingulate cortex(PCC) and the medial frontal cortex (MFC) and less consistent-ly, the temporal cortex for the appraisal of the self (Ochsneret al., 2005; van der Meer et al., 2010). In line with thesefindings we found a correlation between SD and GMV in theMFC and the temporal cortex and between SD andWMV in theACC and PCC (see Fig. 5).
We conclude that our study supports the evidence that SDcan be associated with local GMV and WMV in the brain inthose regions involved in self-reflection.
3.6. Cooperativeness
In this study we found a correlation between CO and WMV inthe medial frontal and precentral gyrus (see Fig. 6) andbetween CO and GMV in the superior temporal gyrus. This
Fig. 3 – Correlation results found for the temperament trait RD overlaid on a T1MNI template image in the OFC (A) and temporalcortex (B).
Fig. 4 – Correlation results found for the temperament trait Poverlaid on a T1 MNI template image in the cingulate cortex.
37B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2

Author's personal copy
result is of interest as that in the theory of Cloninger, COhas been introduced to account for individual differences inidentification with and acceptance of other people (Cloningeret al., 1993). Identification with someone else means that oneis able to understand the others' mental and emotional stateby mentalizing his actions and by empathizing with hisfeelings. Both processes are hypothesized to be the result ofimitating or mirroring the others' feelings and intentions bya mirror neuronal system in the brain starting in the pre-motor andmotor cortex towards the limbic (empathizing) andprefrontal (mentalizing) cortex during the perception ofemotional expressions and motor actions of others (Hookeret al., 2010; Singer, 2006). The superior temporal cortex hasbeen shown to be involved in mentalizing processes (Hookeret al., 2010). A possible link between brain morphology in thementalizing and empathizing network (the social-brain) haspreviously been revealed in the study of Yamasue et al.(2008b).
We conclude that our results suggest a possible relationbetween CO and brainmorphology in those regions involved inmentalizing and empathizing processes, although we cannotstate that the found correlations between WMV and CO in themotor related cortex are really related with the mirror neurons.
3.7. Self-transcendence
In line with the results of Kaasinen et al. (2005) we found acorrelation between ST and GMV in themiddle temporal gyrus(see Fig. 7). In addition to this we also found correlations in theparietal and frontal cortex.
In the theory of Cloninger et al. (1993), ST has beenintroduced to account for one's tendency towards identifyingthe self as part of a broader universe, spiritualism and religion.In the brain, religiosity has been correlated with structural andfunctional variability in the temporal, parietal, limbic andfrontal cortex (Harris et al., 2009; Kapogiannis et al., 2009a,b).Moreover, in a recent study of Urgesi et al. (2010), damage totemporoparietal structures has been found to cause differ-ences in individual ST scores.
In conclusion, our results support a possible relationbetween local GMV and ST in those regions involved intranscendent behaviors.
As discussed in the previous paragraphs, there was partialagreement between our results and those of Iidaka et al. (2006)and Gardini et al. (2009). However, some methodologicaldifferences between these studies must be pointed out, thatmay provide an explanation for the lack of agreement. Inour study we restricted our subject group to young females toexclude effects of age and gender on personality and braindevelopment. Therefore, we cannot generalize our results toolder or younger females or men. A second methodologicaldifference is the use of the DARTEL algorithm which has beenintroduced as an improvement of the optimized VBM approach(Ashburner, 2007). Finally, we used a multiple linear regressionmodel with all personality scales included as independentvariables instead of performing a separate regression analysisfor each personality trait. In this way, our correlations foreach personality dimension are not only controlled for age,total GMV and WMV, but also for the other dimensions.
Fig. 6 – Correlation results found for the character trait CO overlaid on a T1 MNI template image in the motor (A) and pre-motor(B) cortex.
Fig. 5 – Correlation results found for the character trait SDoverlaid on a T1 MNI template image in the ACC and PCC.
38 B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2

Author's personal copy
4. Limitations
In this study, we did not use diffusion tensor imaging (DTI) ormagnetic transfer ratio (MTR) techniques to gain more infor-mation about the micro-structural processes underlying ourWMV results. AsWMV is not only dependent on the numbers ofaxons butalsoon their diameter and the thickness of themyelinsheath surrounding the axons (Paus, 2010), differences inWMVcan hardly be interpreted as differences in inter-regionalconnectivity. Although aswhitemattermaturation is the resultof axon-related andmyelin-related processes,WMV is used as acommon parameter in brain maturation and aging studies(Lenroot and Giedd, 2006). Despite a correlation between WMVand GMV and functional performances have been reported(Brickman et al., 2006, 2007), we do not state that our resultssuggest any difference in behavioral or cognitive neuronalprocesses in the brain between individuals due to differences intheir personality traits.
Other factors such as learning and training (Draganski etal., 2004) have been shown to change brain morphology aswell. In our study we did not any special preparation orselection of our volunteers to control for these environmentalfactors. Moreover, as long as these environmental factors donot interact with personality in any sense, we considered theircontribution to GMV and WMV as random variations that areincluded in the noise term in our regression model. On theother hand, we cannot exclude that personality has aninfluence on training and learning. Even if some or all of ourresults found were due to learning and training, it is still ofinterest to find that personality has an influence on the GMVand WMV changes induced by environmental factors. Ascorrelation results do not show any causality between twofactors, a third interacting factor cannot be excluded.
5. Conclusions
We conclude that the results of our exploratory study supportthe findings of other groups (Gardini et al., 2009; Iidaka et al.,2006), who showed the temperament traits to correlate withindividual structural variations in the brain and we extended
these results to the character dimensions. Moreover, weshowed that structural variations occur in those regionsinvolved in generating the corresponding behavior as statedin the psychobiological theory of Cloninger. However, theexistence of a direct relation between local GMV and WMVvariations and variations in the corresponding functionalperformances is still a matter of debate.
6. Experimental procedures
6.1. Participants
We recruited 80 native Belgian female volunteers (mean age:23±3 years) from staff members and students at our hospitalor the participating universities: the Vrije Universiteit Brussel(VUB) and the Ghent University, by local advertising. Eachparticipant had to be medication free (except for birth-controlmedication), right-handed as assessed with the Van Strienquestionnaire (Van Strien, 2001), free of any psychiatricdisorder as assessed with the Dutch version of the Mini-International Neuropsychiatric Interview (Mini, Sheehanet al., 1998), without any personal psychiatric disorder historyand being non depressed (defined as having a score lower than9 on the 21 item Beck Depression Inventory (BDI, Beck andSteer, 1984)). All volunteers gave their written informedconsent and were financially compensated. This study wasapproved by the Institutional Ethical Board of the UniversityHospital of the Vrije Universiteit Brussel (UZ Brussel) and inaccordance with the guidelines laid down in the declaration ofHelsinki (2004).
6.2. TCI questionnaire
All participants completed the Dutch version of the TCIquestionnaire (de la Rie et al., 1998) by answering “True” or“False” to 240 statements. To measure the personality dimen-sions, these statements were categorized in 24 lower orderpersonality dimensions (4 for HA and NS, 3 for RD and ST, 5 forSD and CO) and P (Cloninger et al., 1993). According to thenumber of lower order dimensions, a maximum score of 40was given to HA and NS, 30 to RD and ST, 10 to P and 50 to SD
Fig. 7 – Correlation results found for the character trait ST overlaid on a T1 MNI template image in the temporal (A) and frontal(B) cortex.
39B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2

Author's personal copy
and CO. Six participants did not complete the questionnaireand were excluded from further analyses.
6.3. Imaging protocol
This study was integrated in fMRI studies running at ourinstitution between 2004 and 2008. In those years one upgradeof our scanner system was performed and a new scanner wastaken in service. This resulted in a division of our study group inthree subgroups with a slightly different scanning protocol. Afirst group of 31 participants was scanned on a 1.5 T PhilipsIntera (release 8) MRI scanner (Philips, Best, The Netherlands)with a quadrature transmit-receive head coil and a 3D T1-TFEsequence characterized by TI/TR/TE=1501/16/4.6 ms, flipangle=30°, FOV=240×240×200 mm, resolution=1×1×2 mmand 100 axial slices. The second group of 19 participants wasscanned on the same scanner upgraded to release 11 with amulti-channelheadcoil andthesamesequence. The third groupof 30 participants was scanned on a 3 T Philips Achieva (release2.5) system (Philips, Best, The Netherlands) with a quadraturetransmit-receiveheadcoil anda3DT1-TFE sequencewithTI/TR/TE=940.4/7.6/3.7 ms, flip angle=8°, FOV=240×240×200 mm,resolution=1×1×2mm and 100 axial slices.
6.4. VBM analysis and statistics
All data preprocessing and analysis was done in SPM5(Statistical Parametric Mapping, Wellcome Department ofImagingNeuroscience, London, UK) following theVBM-DARTELanalysis procedure as introduced by Ashburner and Friston(2000) but with the modification proposed by McLaren et al.(2010) andBergouignan et al. (2009). In a first step,weperformeda segmentation of the individual brains into GMV and WMVmaps. Thesemaps contain theproportionofGMVandWMV inavoxel respectively. The segmentation algorithm included anaffine, linear transformation step to align the individual brainsto the GMV andWMV templates and a resampling step to bringthe data to an isotropic resolution of 1.5 mm.
In a second step, we performed a normalization of theindividual GMV andWMVmaps tomap them inMNI space. Toimprove the accuracy of the normalization step, the “Diffeo-morphic Anatomical Registration using Exponentiated LieAlgebra” (DARTEL) algorithm (Ashburner, 2007) as implemen-ted in the SPM5 DARTEL toolbox was used to create a grouptemplate and to calculate the individual non-linear transfor-mation to this template. After the DARTEL transformation, thesegmented images were modulated by multiplying the voxelvalues with the Jacobian determinants of the deformationfield, to correct for local expansion and contraction and to endup with tissue volume proportion maps. The final normaliza-tion to MNI space was done using the script of McLaren et al.(2010). To avoid smoothing, this script performs an affinetransformation of the coordinate axes defined in the image fileheader rather than performing an affine transformation andresampling of the image. Finally, an isotropic Gaussiansmoothing step with 8 mm FWHM was done.
Prior to the analysis, the sample homogeneity was checkedwith the sample homogeneity check option in the VBM5.1toolbox (http://dbm.neuro.uni-jena.de/vbm/check-sample-homogeneity). This check was done by calculating the square
distance of each image to the sample mean based on a voxel-by-voxel analysis. A systematic error between the 1.5 and 3 Tdata was not found (results not shown). After this check, sixsubjects, one of group 2 and five of group 3,were excluded fromfurther analysis due to differences in segmentation accuracy.
The final analysis was done with a multiple linearregression algorithm in SPM5 once for the GMV maps andonce for the WMV maps with an absolute mask threshold setat 0.1. This absolute mask threshold means that a proportionof at least 10% of the voxel should be white matter or graymatter respectively in all brains to be included in the analysis.
In our model we introduced all personality traits asindependent variables (Miettunen et al., 2008). To correct forunwanted morphological variations, age and total GMV andWMV were included as covariates. As suggested by Kruggelet al. (2010), we added scanner group as covariate to take intoaccount possible local differences in segmentation accuracydue to differences in image quality between the 1.5 and 3 Tdata (Jovicich et al., 2009; Kruggel et al., 2010).
To take in account both type I and type II errors (Liebermanand Cunningham, 2009; Loring et al., 2002), we performed theanalysis without a multiple comparison correction, a voxelsignificance threshold p<0.001 and a cluster extent thresholdof 50 voxels.
The cluster position in theMNI space obtained in SPM5wastransformed to Talairach space coordinates using the Matlab(v7.5.0 R2007b) script mni2tal (http://eeg.sourceforge.net/mri-doc/mri_toolbox/mni2tal.html). These Talairach coordinateswere entered in the Talairach Client (v2.4) database (http://www.talairach.org/client.html) to find the correspondingTalairach labels.
Acknowledgments
This research was supported by a grant from the ScientificFund W. Gepts UZ Brussel.
R E F E R E N C E S
Anckarsäter, H., Larson, T., Hakansson, C., Jutblad, S.B., Niklasson,L., Nydeén, A., Wentz, E., Westergren, S., 2006. The impact ofADHD and autism spectrum disorders on temperament,character, and personality development. Am. J. Psychiatry 163,1239–1244.
Ashburner, J., 2007. A fast diffeomorphic image registrationalgorithm. Neuroimage 38, 95–113.
Ashburner, J., Friston, K.J., 2000. Voxel-based morphometry—themethods. Neuroimage 11, 805–821.
Beck, A.T., Steer, R.A., 1984. International consistencies of theoriginal and revised Beck Depression Inventory. J. Clin. Psychol.40, 1365–1367.
Behrens, T.E.J., Hunt, L.T., Rushworth, F.S., 2009. The computationof social behavior. Science 324, 1160–1164.
Belin, P., Fecteau, S., Bédard, C., 2004. Thinking the voice:neural correlates of voice perception. Trends Cogn. Sci. 8,129–135.
Bergouignan, L., Chupin, M., Czechowska, Y., Kinkingnéhun, S.,Lemogne, C., Le Bastard, G., Lepage, M., Garnero, L., Colliot, O.,Fossati, P., 2009. Can voxel based morphometry, manualsegmentation and automated segmentation equally detect
40 B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2

Author's personal copy
hippocampal volume differences in acute depression?Neuroimage 45, 29–37.
Brickman, A.M., Zimmerman, M.E., Paul, R.H., Grieve, S.M., Tate,D.F., Cohen, R.A., Williams, L.M., Clark, C.R., Gordon, E., 2006.Regional white matter and neuropsychological functioningacross the adult lifespan. Biol. Psychiatry 60, 444–453.
Brickman, A.M., Habeck, C., Zarahn, E., Flynn, J., Stern, Y., 2007.Structural MRI covariance patterns associated with normalaging and neuropsychological functioning. Neurobiol. Aging28, 284–295.
Cloninger, C.R., 1986. A unified biosocial theory of personality andits role in the development of anxiety states. Psychiatr. Dev. 4,167–226.
Cloninger, C.R., 1987. A systematic method for clinical descriptionand classification of personality variants. Arch. Gen. Psychiatry44, 573–588.
Cloninger,C.R., Svrakic, D.M., Przybeck,T.R., 1993.Apsychobiologicalmodel of temperament and character. Arch. Gen. Psychiatry 50,975–990.
Cloninger, C.R., Bayon, C., Svrakic, D.M., 1998. Measures oftemperament and character in mood disorders: a model offundamental states as personality types. J. Affect. Disord. 51,21–32.
Cloninger, C.R., Svrakic, D.M., Przybeck, T.R., 2006. Can personalityassessment predict future depression? A twelve-monthfollow-up of 631 subjects. J. Affect. Disord. 92, 35–44.
Crews, F.T., Boettiger, C.A., 2009. Impulsivity, frontal lobes and riskfor addiction. Pharmacol. Biochem. Behav. 93, 237–247.
de la Rie, S.M., Duijsens, I.J., Cloninger, C.R., 1998. Temperament,character and personality disorders. J. Pers. Disord. 12, 362–372.
Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn, U., May,A., 2004. Neuroplasticity: changes in gray matter induced bytraining. Nature 427, 311–312.
Drevets, W.C., Price, J.L., Furrey, M.L., 2008. Brain structural andfunctional abnormalities in mood disorders: implications forneurocircuitry models of depression. Brain Struct. Funct. 213,93–118.
Farmer, R.F., Seeley, J.R., 2009. Temperament and characterpredictors of depressed mood over a 4-year interval. Depress.Anxiety 26, 371–381.
Ferrari, M.C., Busatto, G.F., McGuire, P.K., Crippa, J.A., 2008.Structural magnetic resonance imaging in anxiety disorders:an update of research findings. Rev. Bras. Psiquiatr. 30, 251–264.
Gardini, S., Cloninger, C.R., Venneri, A., 2009. Individualdifferences in personality traits reflect structural variance inspecific brain regions. Brain Res. Bull. 79, 265–270.
Good, C.D., Johnsrude, I.S., Ashburner, J., Henson, R.N., Friston, K.J.,Frackowiak, R.S., 2001. A voxel-based morphometric study ofageing in 465 normal adult human brains. Neuroimage 14,21–36.
Goodman,M., Triebwasser, J., Shah,S.,New,A.S., 2007.Neuroimagingin personality disorders: current concepts, findings, andimplications. Psychiatr. Ann. 37, 100–108.
Harris, S., Kaplan, J.T., Curiel, A., Bookheimer, S.Y., Iacoboni, M.,Cohen, M.S., 2009. The neural correlates of religious andnonreligious belief. PloS ONE 4, e7272 doi:10.1371/journal.pone.0007272.
Haxby, J.V., Hoffman, E.A., Gobbini, M.I., 2002. Human neuralsystems for face recognition and social communication. Biol.Psychiatry 51, 59–67.
Hooker, C.I., Verosky, S.C., Germine, L.T., Knight, R.T., D'Esposito,M., 2010. Neural activity during social signal perceptioncorrelates with self-reported empathy. Brain Res. 1308,100–113.
Iidaka, T., Matsumoto, A., Ozaki, N., Suzuki, T., Iwata, N.,Yamamoto, Y., Okada, T., Sadato, N., 2006. Volume of leftamygdala subregion predicted temperamental trait of harmavoidance in female young subjects. A voxel-basedmorphometry study. Brain Res. 1125, 85–93.
Jovicich, J., Czanner, S., Han, X., Salat, D., van der Kouwe, A., Quin,B., Pacheco, J., Albert, M., Killiany, R., Blacker, D., Maguire, P.,Rosas, D., 2009. MRI-derived measurements of humansubcortical, ventricular and intracranial brain volumes:reliability effects of scan sessions, acquisition sequences,data analyses, scanner upgrade, scanner vendors and fieldstrengths. Neuroimage 46, 177–192.
Jylha, P., Isometsa, E., 2006. Temperament, character andsymptoms of anxiety and depression in the generalpopulation. Eur. Psychiatry 21, 389–395.
Kaasinen, V., Maguire, R.P., Kurki, T., Brück, A., Rinne, J.O., 2005.Mapping brain structure and personality in late adulthood.Neuroimage 24, 315–322.
Kalenscher, T., Ohmann, T., Güntürkün, O., 2006. Theneuroscience of impulsive and self-controlled decisions.Int. J. Psychophysiol. 62, 203–211.
Kapogiannis, D., Barbey, A.K., Su, M., Krueger, F., Grafman, J.,2009a. Neuroanatomical variability of religiosity. PloS ONE 4,e7180 doi:10.1371/journal.pone.0007180.
Kapogiannis, D., Barbey, A.K., SU, M., Zamboni, G., Krueger, F.,Grafman, J., 2009b. Cognitive and neural foundations ofreligious belief. Proc. Natl. Acad. Sci. U. S. A. 106, 4879–4881.
Kim, S., Lee, D., 2010, Prefrontal cortex and impulsive decisionmaking. Biol. Psychiatry. doi:10.1016/j.biopsych.2010.07.005.
Kruggel, F., Turner, J., Muftuler, L.T., The Alzheimer's DiseaseNeuroimaging Initiative, 2010. Impact of scanner hardware andimaging protocol on image quality and compartment volumeprecision in the ADNI cohort. Neuroimage 49, 2123–2133.
Lebreton, M., Barnes, A., Miettunen, J., Peltonen, L., Ridler, K.,Veijola, J., Suckling, J., Jarvelin, M.R., Jones, P.B., Bullmore, E.T.,Murray, G.K., 2009. The brain structural disposition to socialinteraction. Eur. J. Neurosci. 29, 2247–2252.
LeDoux, J.E., 2000. Emotion circuits in the brain. Annu. Rev.Neurosci. 23, 155–184.
Lenroot, R.K., Giedd, J.N., 2006. Brain development in children andadolescents: insights from anatomical magnetic resonanceimaging. Neurosci. Biobehav. Rev. 30, 718–729.
Lenroot, R.K., Giedd, J.N., 2008. The changing impact of genes andenvironment on brain development during childhood andadolescence: initial findings from a neuroimaging study ofpediatric twins. Dev. Psychopathol. 20, 1161–1175.
Lieberman, M.D., Cunningham, W.A., 2009. Type I and type II errorconcerns in fMRI research: re-balancing the scale. Soc. Cogn.Affect. Neurosci. 4, 423–428.
Loring, D.W., Meador, K.J., Allison, J.D., Pillai, J.J., Lavi, T., Lee, G.P.,Balan, A., Dave, V., 2002. Now you see it, now you don't:statistical andmethodological considerations in fMRI. EpilepsyBehav. 3, 539–547.
Matsuo, K., Nicoletti, M., Nemoto, K., Hatch, J.P., Marco, A.M.P.,Nery, F.G., Soares, J.C., 2008. A voxel-basedmorphometry studyof frontal gray matter correlates of impulsivity. Hum. BrainMapp. 30, 1188–1195.
McCloskey, M.S., Phan, K.L., Caccaro, E.F., 2005. Neuroimaging andpersonality disorders. Curr. Psychiatry Rep. 7, 65–72.
McLaren, D.G., Kostmatka, K.J., Kastman, E.K., Bendlin, B.B.,Johnson, S.C., 2010. Rhesus macaque brain morphometry:a methodological comparison of voxel-wise approaches.Methods 50, 157–165.
Miettunen, J., Veijola, J., Lauronen, E., Kantojärvi, L., Joukamaa, M.,2007. Sexdifferences inCloninger's temperamentdimensions—ameta-analysis. Compr. Psychiatry 48, 161–169.
Miettunen, J., Lauronen, E., Kantojärvi, L., Veijola, J., Joukamaa, M.,2008. Inter-correlations between Cloninger's temperamentdimensions—a meta-analysis. Psychiatry Res. 160, 106–114.
Mobbs, D., Greicius, M.D., Abdel-Azim, E., Menon, V., Reiss, A.L.,2003. Humor modulates the mesolimbic reward centers.Neuron 40, 1041–1048.
Ochsner, K.N., Beer, J.S., Robertson, E.R., Cooper, J.C., Gabrieli,J.D.E., Kihlstrom, J.F., D'Esposito, M., 2005. The neural correlates
41B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2

Author's personal copy
of direct and reflected self-knowledge. Neuroimage 28,797–814.
O'Doherty, J.P., 2004. Reward representations and reward-relatedlearning in the human brain: insights from neuroimaging.Curr. Opin. Neurobiol. 14, 769–776.
Paus, T., 2010. Growth of white matter in the adolescent brain:myelin or axon? Brain Cogn. 72, 26–35.
Plomin, R., Kosslyn, S.M., 2001. Genes, brain and cognition. Nat.Neurosci. 4, 1153–1154.
Rademacher, L., Krach, S., Kohls, G., Irmak, A., Gründer, G.,Spreckelmeyer, K.N., 2010. Dissociation of neural networks foranticipation and consumption ofmonetary and social rewards.Neuroimage 49, 3276–3285.
Richter, J., Brändström, S., 2009. Personality disorder diagnosis bymeans of the Temperament and Character Inventory. Compr.Psychiatry 50, 347–352.
Rushworth, M.F., Behrens, T.E., 2008. Choice, uncertainty andvalue in prefrontal and cingulate cortex. Nat. Neurosci. 11,389–397.
Schmitt, J.E., Eyler, L.T., Giedd, J.N., Kremen, W.S., Kendler, K.S.,Neale, M.C., 2007. Review of twin and family studies onneuroanatomic phenotypes and typical neurodevelopment.Twin Res. Hum. Genet. 10, 683–694.
Sheehan, D.V., Lecrubier, Y., Sheehan, K.H., Amorim, P., Janavs, J.,Weiller, E., Hergueta, T., Baker, R., Dunbar, G.C., 1998. The MiniInternational Neuropsychiatric Interview (M.I.N.I.): thedevelopment and validation of a structured diagnosticpsychiatric interview for DSM-IV and ICD-10. J. Clin. Psychiatry.59 (Suppl. 20), 22–33 quiz 34–57. M.I.N.I. Plus (Nederlandstaligevertaling, Van Vliet, Leroy and Van Megen, 2000).
Singer, T., 2006. The neuronal basis and ontogeny of empathy andmind reading: review of literature and implications for futureresearch. Neurosci. Biobehav. Rev. 30, 855–863.
Spampinato, M.V., Wood, J.N., De Simone, V., Grafman, J., 2009.Neural coorelates of anxiety in healthy volunteers: avoxel-based morphometry study. J. Neuropsychiatry Clin.Neurosci. 21, 199–205.
Sripada, C.S., Gonzalez, R., Luan, Phan K., Liberzon, I., 2010. Theneural correlates of intertemporal decision-making:contributions of subject value, stimulus type, and traitimpulsivity. Hum. Brain Mapp. doi:10.1002/hbm.21136.
Sugiura, M., Kawashima, R., Nakagawa, M., Okada, K., Sato, T.,Goto, R., Sato, K., Ono, S., Schormann, T., Zilles, K., Fukuda, H.,2000. Correlation between human personality and neuralactivity in cerebral cortex. Neuroimage 11, 541–546.
Svrakic, D.M., Draganic, S., Hill, K., Bayon, C., Przybeck, T.R.,Cloninger, C.R., 2002. Temperament, character, and personalitydisorders: etiologic, diagnostic, treatment issues. ActaPsychiatr. Scand. 106, 189–195.
Turner, R.M., Hudson, I.L., Butler, P.H., Joyce, P.R., 2003. Brainfunction and personality in normal males: a SPECT studyusing statistical parametric mapping. Neuroimage 19,1145–1162.
Urgesi, C., Aglioti, S.M., Skrap, M., Fabbro, F., 2010. The spiritualbrain: selective cortical lesions modulate humanself-transcendence. Neuron 65, 309–319.
van der Meer, L., Costafreda, S., Aleman, A., David, A.S., 2010.Self-reflection and the brain: a theoretical review andmeta-analysis of neuroimaging studies with implications forschizophrenia. Neurosci. Biobehav. Rev. 34, 935–946.
Van Strien, J., 2001. Handvoorkeur en taaldominantie. Neuropraxis5, 10–15.
World Medical Association General Assembly, 2004. World MedicalAssociation Declaration of Helsinki: ethical principles formedical research involving human subjects. J. Int. Bioéthique 15,124–129.
Yamasue, H., Abe, O., Suga, M., Yamada, H., Inoue, H., Tochigi, M.,Rogers, M., Aoki, S., Kato, N., Kasai, K., 2008a. Gender-commonand -specific neuroanatomical basis of human anxiety-relatedpersonality traits. Cereb. Cortex 18, 46–52.
Yamasue, H., Abe, O., Suga, M., Yamada, H., Rogers, M.A., Aoki, S.,Kato, N., Kasai, K., 2008b. Sex-linked neuroanatomical basisof human altruistic cooperativeness. Cereb. Cortex 18,2331–2340.
42 B R A I N R E S E A R C H 1 3 7 1 ( 2 0 1 1 ) 3 2 – 4 2




















