Indicators of piglet survival in an outdoor farrowing system
11
Indicators of piglet survival in an outdoor farrowing system Emma M. Baxter a, ⁎, Susan Jarvis a , Lorna Sherwood a , Sheena K. Robson a , Elisabeth Ormandy b , Marianne Farish a , Kathleen M. Smurthwaite c , Rainer Roehe a , Alistair B. Lawrence a , Sandra A. Edwards d a Animal Behaviour and Welfare, Sustainable Livestock Systems, Scottish Agricultural College, West Mains Road, Edinburgh, EH9 3JG, UK b Animal Welfare Program, Room 190E, Macmillan Building, Main Mall, Vancouver, BC, Canada V6T 1Z2 c Grampian Country Food Group, Muirden, Turriff, Grampian, AB53 4NH, UK d Newcastle University, School of Agriculture, Food and Rural Development, Agriculture Building, Newcastle upon Tyne NE1 7RU, UK article info abstract Article history: Received 1 July 2008 Received in revised form 6 February 2009 Accepted 11 February 2009 Pre-weaning piglet mortality continues to be a major welfare and economic concern. In outdoor farrowing systems, there is a particular need to broaden breeding goals by incorporating selection for piglet survival to improve both productivity and welfare. This study aimed to identify behavioural and physiological survival indicators that are influential in outdoor systems and that could provide additional information for use when selecting for piglet survival. Data were collected from 511 piglets from Large White×Landrace×Duroc sows and Generalised Linear Mixed Models determined which indicators were most important for piglet survival in an outdoor system. With respect to prenatal mortality (surviving vs. stillborn piglets) high ponderal index (P b 0.001) or body mass index (P b 0.001) in conjunction with being born earlier in the farrowing birth order (P b 0.001) were the most important survival indicators. Birth weight (P b 0.001) and rectal temperature 1 h after birth (P = 0.032) were the most significant postnatal survival indicators. However survival indicators identified as important in indoor, conventional farrowing crates, such as landmark behaviours (latency to reach the udder, a teat and to suckle colostrum), were not influential in this system. These results highlight the importance of studying potential indicators of survival in alternative farrowing systems to the farrowing crate. © 2009 Elsevier B.V. All rights reserved. Keywords: Piglet survival Outdoor systems Welfare Survival indicators 1. Introduction With recent estimates of total pre-weaning piglet mortal- ity ranging between 15–20% (Leenhouwers et al., 2002; Meat and Livestock Commission, 2006), and environmental mod- ifications, such as the farrowing crate, no longer having an impact on reducing this level (Edwards, 2002), piglet survival continues to be a major welfare and economic concern. Environmental solutions to piglet mortality have included the widespread use of the farrowing crate. However this system imposes many restrictions, both behavioural and physical, on the sow and raises serious welfare concerns (Jarvis et al., 1997; Lawrence et al., 1994). With increasing pressure to abolish the farrowing crate, investigations into improving survival in alternative farrowing systems are essential. In general, outdoor systems are perceived by consumers as more humane, environmentally friendly, traditional and sustain- able (Edwards, 2005). Although challenges exist for animals kept outdoors, particularly with respect to climatic extremes (Algers and Jensen, 1990; Edwards and Zanella, 1996) and potential bio-security risks (e.g. Callaway et al., 2005), there is little doubt that this environment gives the opportunity for behavioural freedom. Current estimates of live-born mortality in the UK (11.8% indoor and 10.5% outdoor; Meat Livestock and Commission, 2006) suggest that the outdoor environ- ment offers a competitive level of survival. In order to further improve survival in these extensive farrowing systems, genetic selection programmes with broader breeding goals that incorporate piglet survival traits could be implemented. Livestock Science 124 (2009) 266–276 ⁎ Corresponding author. Tel.: +44 131 5353214; fax: +44 131 5353121. E-mail address: [email protected] (E.M. Baxter). 1871-1413/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.livsci.2009.02.008 Contents lists available at ScienceDirect Livestock Science journal homepage: www.elsevier.com/locate/livsci
Transcript of Indicators of piglet survival in an outdoor farrowing system

Livestock Science 124 (2009) 266–276
Contents lists available at ScienceDirect
Livestock Science
j ourna l homepage: www.e lsev ie r.com/ locate / l ivsc i
Indicators of piglet survival in an outdoor farrowing system
Emma M. Baxter a,⁎, Susan Jarvis a, Lorna Sherwood a, Sheena K. Robson a, Elisabeth Ormandy b,Marianne Farish a, Kathleen M. Smurthwaite c, Rainer Roehe a, Alistair B. Lawrence a,Sandra A. Edwards d
a Animal Behaviour and Welfare, Sustainable Livestock Systems, Scottish Agricultural College, West Mains Road, Edinburgh, EH9 3JG, UKb Animal Welfare Program, Room 190E, Macmillan Building, Main Mall, Vancouver, BC, Canada V6T 1Z2c Grampian Country Food Group, Muirden, Turriff, Grampian, AB53 4NH, UKd Newcastle University, School of Agriculture, Food and Rural Development, Agriculture Building, Newcastle upon Tyne NE1 7RU, UK
a r t i c l e i n f o
⁎ Corresponding author. Tel.: +44 131 5353214; faxE-mail address: [email protected] (E.M. Bax
1871-1413/$ – see front matter © 2009 Elsevier B.V.doi:10.1016/j.livsci.2009.02.008
a b s t r a c t
Article history:Received 1 July 2008Received in revised form 6 February 2009Accepted 11 February 2009
Pre-weaning piglet mortality continues to be amajor welfare and economic concern. In outdoorfarrowing systems, there is a particular need to broaden breeding goals by incorporatingselection for piglet survival to improve both productivity and welfare. This study aimed toidentify behavioural and physiological survival indicators that are influential in outdoorsystems and that could provide additional information for use when selecting for pigletsurvival. Data were collected from 511 piglets from Large White×Landrace×Duroc sows andGeneralised Linear Mixed Models determined which indicators were most important for pigletsurvival in an outdoor system. With respect to prenatal mortality (surviving vs. stillbornpiglets) high ponderal index (Pb0.001) or body mass index (Pb0.001) in conjunction withbeing born earlier in the farrowing birth order (Pb0.001) were the most important survivalindicators. Birth weight (Pb0.001) and rectal temperature 1 h after birth (P=0.032) were themost significant postnatal survival indicators. However survival indicators identified asimportant in indoor, conventional farrowing crates, such as landmark behaviours (latency toreach the udder, a teat and to suckle colostrum), were not influential in this system. Theseresults highlight the importance of studying potential indicators of survival in alternativefarrowing systems to the farrowing crate.
© 2009 Elsevier B.V. All rights reserved.
Keywords:Piglet survivalOutdoor systemsWelfareSurvival indicators
1. Introduction
With recent estimates of total pre-weaning piglet mortal-ity ranging between 15–20% (Leenhouwers et al., 2002; Meatand Livestock Commission, 2006), and environmental mod-ifications, such as the farrowing crate, no longer having animpact on reducing this level (Edwards, 2002), piglet survivalcontinues to be a major welfare and economic concern.Environmental solutions to piglet mortality have included thewidespread use of the farrowing crate. However this systemimposes many restrictions, both behavioural and physical, onthe sow and raises serious welfare concerns (Jarvis et al.,1997; Lawrence et al., 1994). With increasing pressure to
: +44 131 5353121.ter).
All rights reserved.
abolish the farrowing crate, investigations into improvingsurvival in alternative farrowing systems are essential. Ingeneral, outdoor systems are perceived by consumers as morehumane, environmentally friendly, traditional and sustain-able (Edwards, 2005). Although challenges exist for animalskept outdoors, particularly with respect to climatic extremes(Algers and Jensen, 1990; Edwards and Zanella, 1996) andpotential bio-security risks (e.g. Callaway et al., 2005), there islittle doubt that this environment gives the opportunity forbehavioural freedom. Current estimates of live-bornmortalityin the UK (11.8% indoor and 10.5% outdoor; Meat Livestockand Commission, 2006) suggest that the outdoor environ-ment offers a competitive level of survival. In order to furtherimprove survival in these extensive farrowing systems,genetic selection programmes with broader breeding goalsthat incorporate piglet survival traits could be implemented.

267E.M. Baxter et al. / Livestock Science 124 (2009) 266–276
However before such breeding programmes can be put intopractice, information on piglet survival indicators that areinfluential in outdoor systems needs to be collected todetermine traits for selection.
The focus of most studies in outdoor farrowing systemshas been directed towards gathering piglet mortality andperformance information (Edwards et al., 1994; Johnson et al.,2001; O'Reilley et al., 2006) or investigating elements of theenvironment that influence mortality (e.g. temperaturedifferentials, hut and fender design- (McGlone and Hicks,2000; Johnson and McGlone, 2003)), rather than detaileddata collection on individual animals. Behavioural studieshave been limited to the maternal ability of sows (Jensen,1986, 1988; Wülbers-Mindermann et al., 2002; Vieuille et al.,2003; Held et al., 2006), a factor thought to be paramount indetermining the number of piglets raised in outdoor systems(Arey, 1997). Given the challenges of detailed data collectionin these environments, few studies exist which focus onindividual piglet characteristics. Those that do, concentrate onbehavioural aspects, such as suckling frequency and mother–young interactions (Newberry and Woodgush, 1985; Hotzelet al., 2004), although a notable exception is the study of Riartet al. (2000) whichmeasured rectal temperatures and time tofirst colostrum intake in outdoor and indoor piglets. However,the majority of studies identifying characteristics associatedwith improved piglet survival have been conducted in indoor,relatively controlled conditions (Herpin et al., 1996;Tuchscherer et al., 2000; Herpin et al., 2001; Baxter et al.,2008) with indicators of viability being fairly consistentacross studies (e.g. birth weight, thermoregulatory ability,landmark behaviours). However, it should not be assumedthat the relative importance of survival indicators will be
Fig. 1. Diagrammatic representation (not to scale) of the farrowing paddock, farrowinand the farrowing and safe huts had 3.09 m2
floor space (2.22 m×1.39 m), roof d1.00 m×1.07 m and a height of 0.46 m.
generic across environments. Therefore it is important tostudy piglet survival indicators in different farrowing envir-onments and, in this study, we investigated behavioural andphysiological indicators of piglet survival in a population ofpiglets on an outdoor farm.
2. Materials and methods
2.1. Animals and husbandry
Thirty-eight, fifth parity Large White×Landrace×Durocsows were randomly selected from a herd of 384. The sowswere artificially inseminatedwith semen from Landrace boars(PIC Ltd, Abingdon, UK). They were part of a larger geneticstudy (Roehe et al., 2009) involving two selection lineswhich were balanced within our experimental sample.
The experiment took place on a commercial outdoor pigfarm in Aberdeenshire in the UK, with sows and pigletsmanaged under typical commercial conditions. All sows wereartificially inseminated in a nearby indoor facility. After beingserved, sows were grouped together (approximately 20 sowsper group) in outdoor gestation paddocks. Four weeks afterservice, sows were pregnancy tested with an ultrasoundscanner. The 38 experimental sows were divided betweenfour farrowing batches over time. The batches farrowed atthree week intervals between May and August 2005. Sowswere moved from gestation paddocks to individual farrowingpaddocks approximately 10 days before the farrowing duedate. The individual farrowing paddocks were approximately20 m×20 m, separated by electric fences (Fig. 1). Farrowingoccurred in double-skinned insulated huts, with slopedwooden walls and a floor area of 3.09 m2 (2.22 m×1.39 m),
g hut and experimental set-up. The paddock was approximately 20 m×20 mimension of 2.22 m×1.22 m and a height of 1.09 m. The fender measured

268 E.M. Baxter et al. / Livestock Science 124 (2009) 266–276
roof dimension of 2.22 m×1.22 m and a height of 1.09 m. Atthe entrance of each hut was a fender that measured1.00 m×1.07 m and a height of 0.46 m to keep the neonatalpiglets within the hut vicinity. Huts were initially beddedwith long-stemmed barley straw to a depth of approximately10–12 cm. This was replenished when needed. After weaning,huts were moved to a new location and the bedding wasremoved from the paddocks. Pregnant sows were offered threekgperdayof a diet containing12.74%CP,13.32MJDEkg−1. Afterfarrowing, lactation diet (17%CP,13.75MJ DE kg−1)was offeredat a rate of three kg per day, followed by 0.5 kg increments eachday until seven kg, and then followed by one kg incrementseach day up to amaximum of 12 kg until weaning. Sows had adlibitumaccess towater froma trough. Farrowingwas allowed tooccur naturally, although staff were present for data collection.The experimental protocol limited intervention by trained staffwith a farrowing sow to circumstances where a piglet had notappeared within three hours of fluids being seen or where athree-hour interval had passed between piglet births. Nointerventionswere necessary in this study. Temperaturewithinthe huts was recorded remotely during farrowing using athermal imager (IRISYS™ PCI Imager, Integrated InfraredSystems Limited). Hut wall temperature averaged 14.7 °C(±S.E.M. 0.53) (range 2.9–25.7 °C). Temperature at theudder of the sow averaged 31.5 °C (±0.62) (range 22.5–37.1 °C). Approximately 24 h after piglets were born theyunderwent standard husbandry procedures, including tail-docking, teeth-clipping and ear tagging. Average age atweaning for the piglets was 28 days (±0.33).
2.2. Data collection
Back-fat measurements (taken ultrasonically at the P2position; 6.5 cm from the midline at the level of the last rib)and condition scores (1–5 scale of increasing condition;Muirhead and Alexander, 1997) were taken just prior to thesows moving into their farrowing accommodation. Before thesows were moved for farrowing, the huts were modified forsafe data collection and filming: Infra-red (IR) video cameras(Low-lux B/W waterproof cameras: SK-2020XC/SO, RF Con-cepts Ltd, Belfast, Northern Ireland) were positioned in a safe,discreet place (Fig. 1) and additional IR lighting was fitted tothe farrowing huts to ensure that the full hut area could beseen on camera and that 24 h filming caused no disturbanceto the sow and her piglets. Video recording started approxi-mately three days before the farrowing due date andcontinued for two days post-farrowing. The farrowingpaddocks and huts were remotely situated, and thereforepower was supplied via a “Super-Silenced” generator (MOSAGE6000 SXGS Yanmar diesel). The generator was running 24
Table 1Definitions of the different causes of piglet death after post-mortem examination.
Type of death Description
Stillborn Fully formed piglet, observed by experimenters not to brlung tissue is removed and placed in water; lung tissue d
Starvation Piglet very thin, dehydrated skin with pin bones and indicolostrum in the stomach.
Crushed Piglet body has signs of trauma; squashed or bruised in aOther Scour, blind anus, splayed legs, deformations, disease and
h a day and because it was turned on from three days pre-farrowing, sows had a chance to habituate to any noise. Anadditional hut was placed back-to-back with the sow's ownhut (Fig. 1) to facilitate safe data collection. This “safe” hutallowed two experimenters to be present during farrowing tocollect data and manipulate camera angles if needed, withminimum disturbance to the sow. Data collection wasconcentrated on the piglets and when sows gave birth pigletscould be removed from the birth site simply by reaching infrom the safe hut. When sows farrowed in a less accessiblearea (e.g. with piglets born in the centre of the farrowing hut),piglets were gathered carefully, using a shepherd's crook.Behavioural and physiological data were collected during thefarrowing period and for 24 h post-farrowing. Piglets weremonitored until weaning, when they were weighed (WW).Monitoring the piglets involved checking the huts daily tocount litter size, check health and to see if there were anydead piglets. Any piglets dying between birth and weaningwere recorded and cause of death was ascertained by post-mortem dissection and examination of internal organs (seeTable 1 for descriptions). All farrowings were attended andtherefore stillborn piglets could be recorded accurately. Ifconfirmation of stillbirth was needed, post-mortem examina-tion of the lungs was carried out. Mummified piglets wererecorded but not included in any analyses. Physiological datacollection began at the birth of the first piglet for each litter.
2.3. Farrowing measurements and behavioural observations
Information collected within litter consisted of birth littersize (LS), birth order (BO), birth interval (BI) (the intervalbetween each piglet being born) and cumulative farrowingduration (CFD) (the elapsed time between the birth of thefirst piglet and each subsequent piglet). Immediately afterbirth, the vitality (V) of the piglet was visually assessed andscored using a categorical scale:
1. No movement, no breathing after 15 s;2. No body or leg movement after 15 s, piglet is breathing or
attempting to breathe (coughing, spluttering, clearing itslungs);
3. Piglet shows some movement, breathing or attempting tobreathe and rights itself onto its sternum within 15 s;
4. Good movement, good breathing, piglet attempts to standwithin 15 s.
A blood glucose sample was taken at birth (BG) from theumbilical cord as a mixture of venous and arterial blood andwas analysed using a blood glucose monitor (Accu-chekAdvantage Glucose Meter™, Roche Diagnostics, West Sussex,UK). Piglets were weighed at birth (BW) and a crown to rump
eathe after birth, periople still present on hooves. If confirmation is needed,oes not float.vidual ribs visible, vertebrae prominent. Dissection shows piglet has no
ppearance or with broken limbs. Dissection discounts other causes of death.any unidentified causes.

269E.M. Baxter et al. / Livestock Science 124 (2009) 266–276
length (CRL) was measured (the supine length of the pigletfrom the crown of its head to the base of its tail). From thesemeasurements, ponderal index (PI; birth weight (kg)/crown-rump length (m)3) and body mass index (BMI; birth weight/crown-rump length2) were calculated for each piglet. Pigletgender was noted and birth rectal temperature was recorded(BT) using a digital thermometer (BF-169 Flexible tip digitalthermometer, Farlin Infant Products Corporation, Taiwan).The piglet then underwent a righting response test (R): thepiglet was placed gently on its back onto a concave beanbag(dimensions: 25 cm width×15 cm length), then it wasreleased and its latency to right itself onto its feet was timed.The piglets were given 15 s to right themselves, if they failedto do this within 15 s a latency of 16 s was noted. Once a piglethad undergone these measurements, a birth order numberwas written on its back and it was returned to the sow at itsbirth site. Processing each piglet in this way took approxi-mately four minutes. Piglets were processed with care bytrained staff tominimise handling stress and any confoundingeffects in the data. Piglet behaviour data were collected fromvideo play-back and latencies to key landmark behaviourswere recorded from birth, with processing time subtractedfrom these latencies:
• Latency to reach the udder (U): time between birth andwhen contact between the piglet's snout and the sow'sudder was made.
• Latency to reach a functional teat (T): time between birthand when firm contact between the piglet's open mouthand a functional teat (not a non-functional or inverted teat)was made.
• Latency to suckle (S): time between birth and firm andprolonged (over two seconds) contact between the piglet'smouth and a functional teat was made. The piglet typicallyadopted a brace position, rapid mouth and muzzle motionsoccurred and frantic teat-seeking stopped.
Rectal temperatures were recorded at 1 h (T1) and 24 h(T24) after birth. Approximately 24 h after birth, a secondblood glucose (G24) sample was taken when the piglet wastail docked; after the tip of the tail was docked this discardedtissue was used to provide a blood sample. Piglets wereweighed again at 24 h old (W24) and the percentage weightchange over this 24 h period was calculated.
2.4. Statistical analysis
Generalised LinearMixedModels (GLMMs)werefitted to thebinary data (piglets were either dead or alive) to identifyindicators of piglet survival. Some potential indicator variables(BI, CFD, landmark behaviours and T24) were not normallydistributed and so were natural logarithm transformed prior toGLMManalysis. Analyseswere split according to the two distinctmortality types: firstly, stillborn piglets and surviving piglets (i.e.those that were weaned) were compared and all models wereclassified as prenatal. Secondly, live-born piglets that subse-quently died before weaning (dying) and surviving piglets werecompared and all models were classified as postnatal. All modelstook the following format: mortality was fitted as the responsevariable, with sow nested within farrowing batch number fittedas random factors to take into account litter differences and anyvariation in batch number respectively. Potential indicator
variables were fitted into the models as fixed effects. GLMMswere fittedwith a Binomial distribution and a Logit link function.
GLMMs were initially carried out in a univariate basiswhere each potential indicator variable was fitted as the soleexplanatory variable in the model to identify its importancewith respect to survival and both prenatal and postnatalresults are presented from these univariate analyses. Anyvariables from these univariate analyses that were significantat 5% were carried forward for more detailed multivariateanalysis where GLMMs comprising more than one explana-tory variable were considered using a forward selectionprocess. This determined which combinations of variablesbest explained variation in survival and the final prenatal andpostnatal multivariate models are presented in the results.Models were built up by considering pairs of variables,followed by adding a third variable where appropriate. Thiswas subject to the constraint that certain variables did notappear together in the same model because they were ratiosinvolving birth weight or crown-rump length or both (i.e. PI,BMI) and so were highly correlated with each other.
The statistical significance of each term in the GLMM wasassessed using the χ2 approximation to the Wald test; thisassesses the contributions of individual terms in the fixed effectscomponent of the models (Genstat, 11th edition, Lawes Agricul-tural Trust, VSN International Ltd, Oxford, UK). Sensitivity andspecificity were used to assess the adequacy of eachmodel fittedand to compare fitted models. Sensitivity is the proportion ofdead piglets which are correctly predicted to be dead using thefittedmodel, and specificity is the proportion of piglets survivingwhich are correctly predicted as not dying using the fittedmodel(Baxter et al., 2008). Spearman's rank correlations were used toidentify relationships between variables. Genstat software (Gen-stat,11th edition, Lawes Agricultural Trust, VSN International Ltd,Oxford, UK) was used for all statistical analyses.
3. Results
Thirty-eight sows produced 511 piglets. Formortality data, all38 litters were used however, because of simultaneous farrow-ings, two of the litters had very little behavioural andphysiological data collected, so were removed from additionalanalysis.
3.1. Mortality
The 38 sows produced 511 piglets, of which 99 died pre-weaning. Total pre-weaning mortality (prenatal mortalities +postnatal mortalities) was 19.38% and total live-born mortalitywas 14.29%. Of the piglets that died 26 were stillborn, 42 werecrushed, 10 died from starvation and 21 died from other causes(see Table 1). Selection line was found not to have a significanteffect on either total pre-weaning mortality (W1=1.77P=0.183) or live-born mortality (W1=0.11 P=0.744).
3.2. Prenatal survival indicators
3.2.1. Farrowing traitsTable 2 shows the relationships between traits found to be
significant with respect to mortality in the GLMM analysis and itllustrates that the majority of farrowing traits (litter size,birth order, birth interval and cumulative farrowing
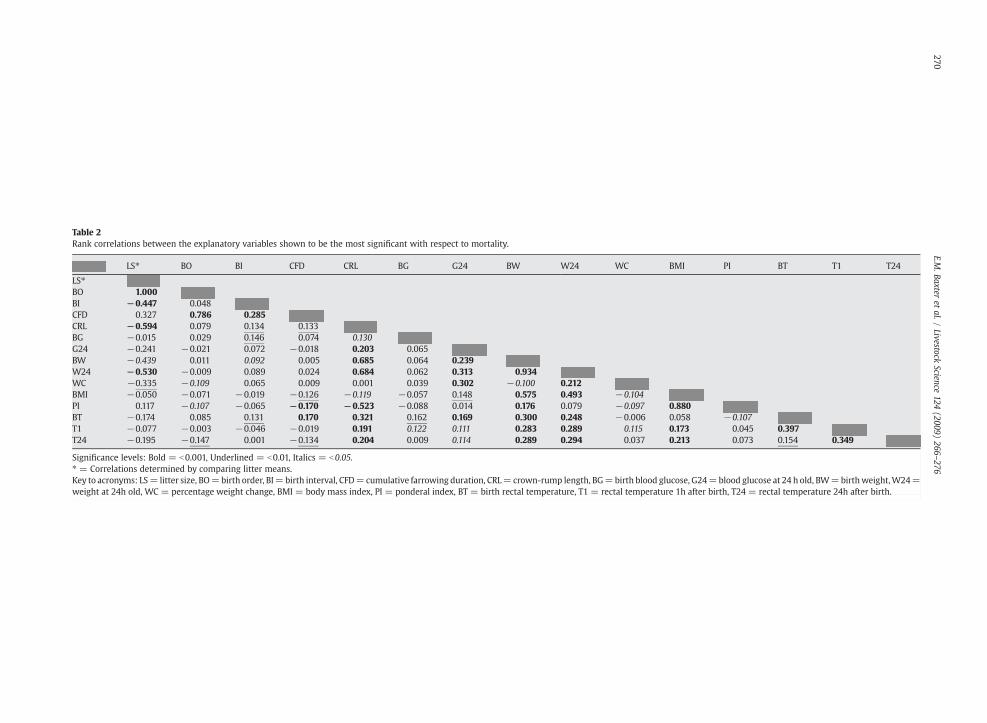
Table 2Rank correlations between the explanatory variables shown to be the most significant with respect to mortality.
LS⁎ BO BI CFD CRL BG G24 BW W24 WC B I PI BT T1 T24
LS⁎BO 1.000BI −0.447 0.048CFD 0.327 0.786 0.285CRL −0.594 0.079 0.134 0.133BG −0.015 0.029 0.146 0.074 0.130G24 −0.241 −0.021 0.072 −0.018 0.203 0.065BW −0.439 0.011 0.092 0.005 0.685 0.064 0.239W24 −0.530 −0.009 0.089 0.024 0.684 0.062 0.313 0.934WC −0.335 −0.109 0.065 0.009 0.001 0.039 0.302 −0.100 0.212BMI −0.050 −0.071 −0.019 −0.126 −0.119 −0.057 0.148 0.575 0.493 −0.104PI 0.117 −0.107 −0.065 −0.170 −0.523 −0.088 0.014 0.176 0.079 −0.097 0 80BT −0.174 0.085 0.131 0.170 0.321 0.162 0.169 0.300 0.248 −0.006 0 58 −0.107T1 −0.077 −0.003 −0.046 −0.019 0.191 0.122 0.111 0.283 0.289 0.115 0 73 0.045 0.397T24 −0.195 −0.147 0.001 −0.134 0.204 0.009 0.114 0.289 0.294 0.037 0 13 0.073 0.154 0.349
Significance levels: Bold = b0.001, Underlined = b0.01, Italics = b0.05.⁎ = Correlations determined by comparing litter means.Key to acronyms: LS= litter size, BO=birth order, BI= birth interval, CFD=cumulative farrowing duration, CRL= crown-rump length, BG=birth blood glu se, G24=blood glucose at 24 h old, BW=birthweight,W24=weight at 24h old, WC = percentage weight change, BMI = body mass index, PI = ponderal index, BT = birth rectal temperature, T1 = rectal temperature h after birth, T24 = rectal temperature 24h after birth.
270E.M
.Baxteret
al./Livestock
Science124
(2009)266
–276
M
.8
.0
.1
.2
co1
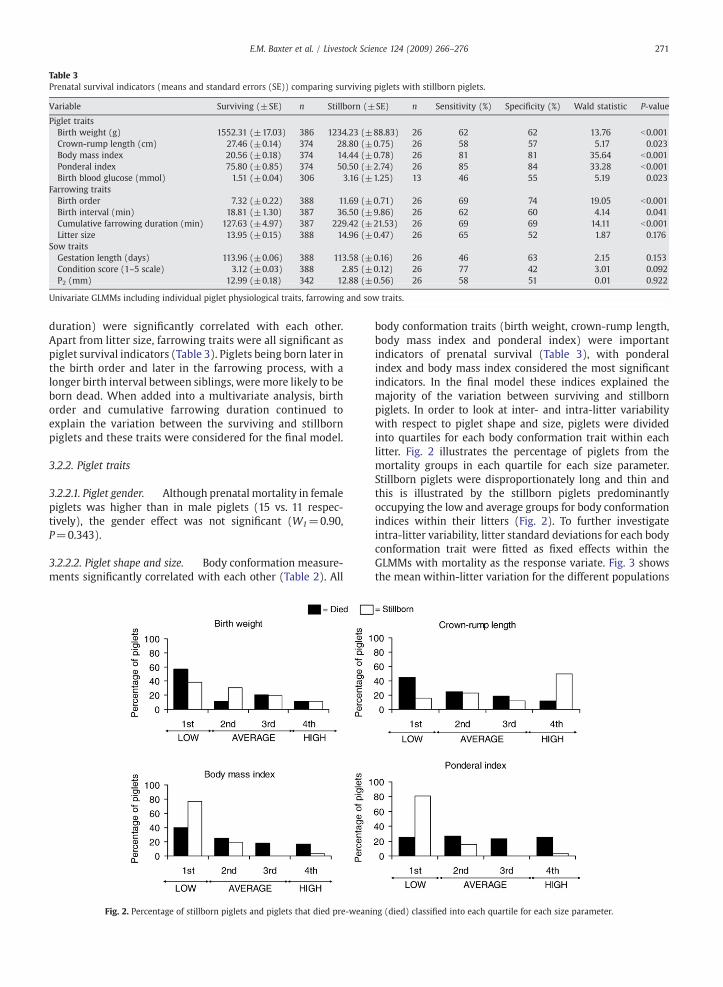
Table 3Prenatal survival indicators (means and standard errors (SE)) comparing surviving piglets with stillborn piglets.
Variable Surviving (±SE) n Stillborn (±SE) n Sensitivity (%) Specificity (%) Wald statistic P-value
Piglet traitsBirth weight (g) 1552.31 (±17.03) 386 1234.23 (±88.83) 26 62 62 13.76 b0.001Crown-rump length (cm) 27.46 (±0.14) 374 28.80 (±0.75) 26 58 57 5.17 0.023Body mass index 20.56 (±0.18) 374 14.44 (±0.78) 26 81 81 35.64 b0.001Ponderal index 75.80 (±0.85) 374 50.50 (±2.74) 26 85 84 33.28 b0.001Birth blood glucose (mmol) 1.51 (±0.04) 306 3.16 (±1.25) 13 46 55 5.19 0.023
Farrowing traitsBirth order 7.32 (±0.22) 388 11.69 (±0.71) 26 69 74 19.05 b0.001Birth interval (min) 18.81 (±1.30) 387 36.50 (±9.86) 26 62 60 4.14 0.041Cumulative farrowing duration (min) 127.63 (±4.97) 387 229.42 (±21.53) 26 69 69 14.11 b0.001Litter size 13.95 (±0.15) 388 14.96 (±0.47) 26 65 52 1.87 0.176
Sow traitsGestation length (days) 113.96 (±0.06) 388 113.58 (±0.16) 26 46 63 2.15 0.153Condition score (1–5 scale) 3.12 (±0.03) 388 2.85 (±0.12) 26 77 42 3.01 0.092P2 (mm) 12.99 (±0.18) 342 12.88 (±0.56) 26 58 51 0.01 0.922
Univariate GLMMs including individual piglet physiological traits, farrowing and sow traits.
271E.M. Baxter et al. / Livestock Science 124 (2009) 266–276
duration) were significantly correlated with each other.Apart from litter size, farrowing traits were all significant aspiglet survival indicators (Table 3). Piglets being born later inthe birth order and later in the farrowing process, with alonger birth interval between siblings, were more likely to beborn dead. When added into a multivariate analysis, birthorder and cumulative farrowing duration continued toexplain the variation between the surviving and stillbornpiglets and these traits were considered for the final model.
3.2.2. Piglet traits
3.2.2.1. Piglet gender. Although prenatal mortality in femalepiglets was higher than in male piglets (15 vs. 11 respec-tively), the gender effect was not significant (W1=0.90,P=0.343).
3.2.2.2. Piglet shape and size. Body conformation measure-ments significantly correlated with each other (Table 2). All
Fig. 2. Percentage of stillborn piglets and piglets that died pre-weani
body conformation traits (birth weight, crown-rump length,body mass index and ponderal index) were importantindicators of prenatal survival (Table 3), with ponderalindex and body mass index considered the most significantindicators. In the final model these indices explained themajority of the variation between surviving and stillbornpiglets. In order to look at inter- and intra-litter variabilitywith respect to piglet shape and size, piglets were dividedinto quartiles for each body conformation trait within eachlitter. Fig. 2 illustrates the percentage of piglets from themortality groups in each quartile for each size parameter.Stillborn piglets were disproportionately long and thin andthis is illustrated by the stillborn piglets predominantlyoccupying the low and average groups for body conformationindices within their litters (Fig. 2). To further investigateintra-litter variability, litter standard deviations for each bodyconformation trait were fitted as fixed effects within theGLMMs with mortality as the response variate. Fig. 3 showsthe mean within-litter variation for the different populations
ng (died) classified into each quartile for each size parameter.
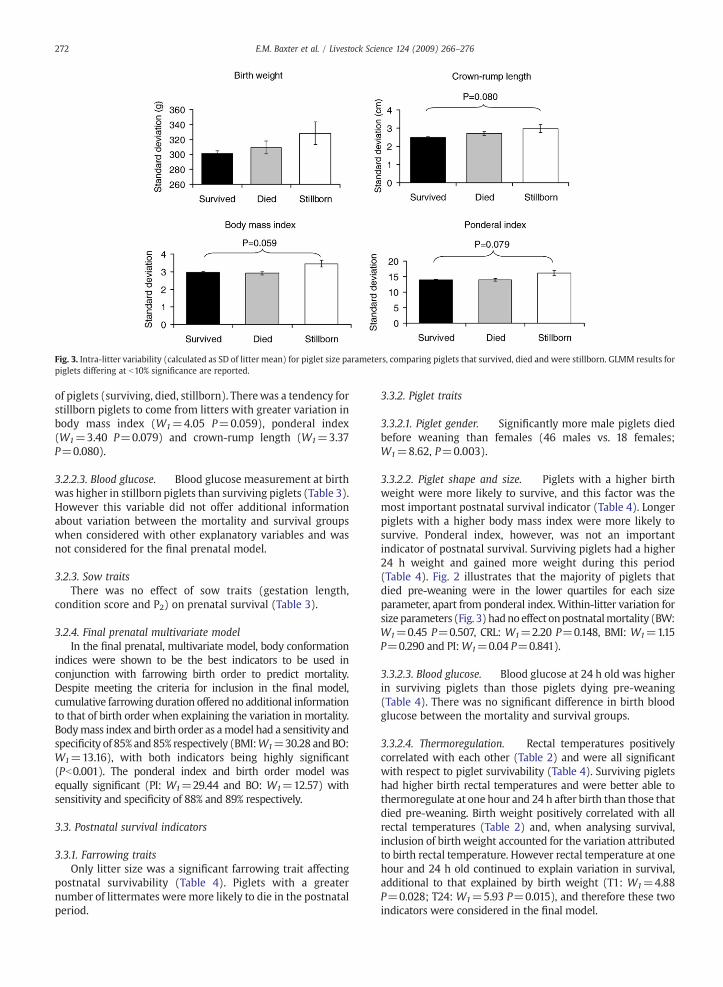
Fig. 3. Intra-litter variability (calculated as SD of litter mean) for piglet size parameters, comparing piglets that survived, died and were stillborn. GLMM results forpiglets differing at b10% significance are reported.
272 E.M. Baxter et al. / Livestock Science 124 (2009) 266–276
of piglets (surviving, died, stillborn). Therewas a tendency forstillborn piglets to come from litters with greater variation inbody mass index (W1=4.05 P=0.059), ponderal index(W1=3.40 P=0.079) and crown-rump length (W1=3.37P=0.080).
3.2.2.3. Blood glucose. Blood glucose measurement at birthwas higher in stillborn piglets than surviving piglets (Table 3).However this variable did not offer additional informationabout variation between the mortality and survival groupswhen considered with other explanatory variables and wasnot considered for the final prenatal model.
3.2.3. Sow traitsThere was no effect of sow traits (gestation length,
condition score and P2) on prenatal survival (Table 3).
3.2.4. Final prenatal multivariate modelIn the final prenatal, multivariate model, body conformation
indices were shown to be the best indicators to be used inconjunction with farrowing birth order to predict mortality.Despite meeting the criteria for inclusion in the final model,cumulative farrowing duration offered no additional informationto that of birth order when explaining the variation in mortality.Bodymass index and birth order as amodel had a sensitivity andspecificity of 85% and 85% respectively (BMI:W1=30.28 and BO:W1=13.16), with both indicators being highly significant(Pb0.001). The ponderal index and birth order model wasequally significant (PI: W1=29.44 and BO: W1=12.57) withsensitivity and specificity of 88% and 89% respectively.
3.3. Postnatal survival indicators
3.3.1. Farrowing traitsOnly litter size was a significant farrowing trait affecting
postnatal survivability (Table 4). Piglets with a greaternumber of littermates were more likely to die in the postnatalperiod.
3.3.2. Piglet traits
3.3.2.1. Piglet gender. Significantly more male piglets diedbefore weaning than females (46 males vs. 18 females;W1=8.62, P=0.003).
3.3.2.2. Piglet shape and size. Piglets with a higher birthweight were more likely to survive, and this factor was themost important postnatal survival indicator (Table 4). Longerpiglets with a higher body mass index were more likely tosurvive. Ponderal index, however, was not an importantindicator of postnatal survival. Surviving piglets had a higher24 h weight and gained more weight during this period(Table 4). Fig. 2 illustrates that the majority of piglets thatdied pre-weaning were in the lower quartiles for each sizeparameter, apart from ponderal index. Within-litter variation forsize parameters (Fig. 3) hadnoeffect onpostnatalmortality (BW:W1=0.45 P=0.507, CRL: W1=2.20 P=0.148, BMI: W1=1.15P=0.290 and PI:W1=0.04 P=0.841).
3.3.2.3. Blood glucose. Blood glucose at 24 h old was higherin surviving piglets than those piglets dying pre-weaning(Table 4). There was no significant difference in birth bloodglucose between the mortality and survival groups.
3.3.2.4. Thermoregulation. Rectal temperatures positivelycorrelated with each other (Table 2) and were all significantwith respect to piglet survivability (Table 4). Surviving pigletshad higher birth rectal temperatures and were better able tothermoregulate at one hour and 24 h after birth than those thatdied pre-weaning. Birth weight positively correlated with allrectal temperatures (Table 2) and, when analysing survival,inclusion of birth weight accounted for the variation attributedto birth rectal temperature. However rectal temperature at onehour and 24 h old continued to explain variation in survival,additional to that explained by birth weight (T1: W1=4.88P=0.028; T24: W1=5.93 P=0.015), and therefore these twoindicators were considered in the final model.
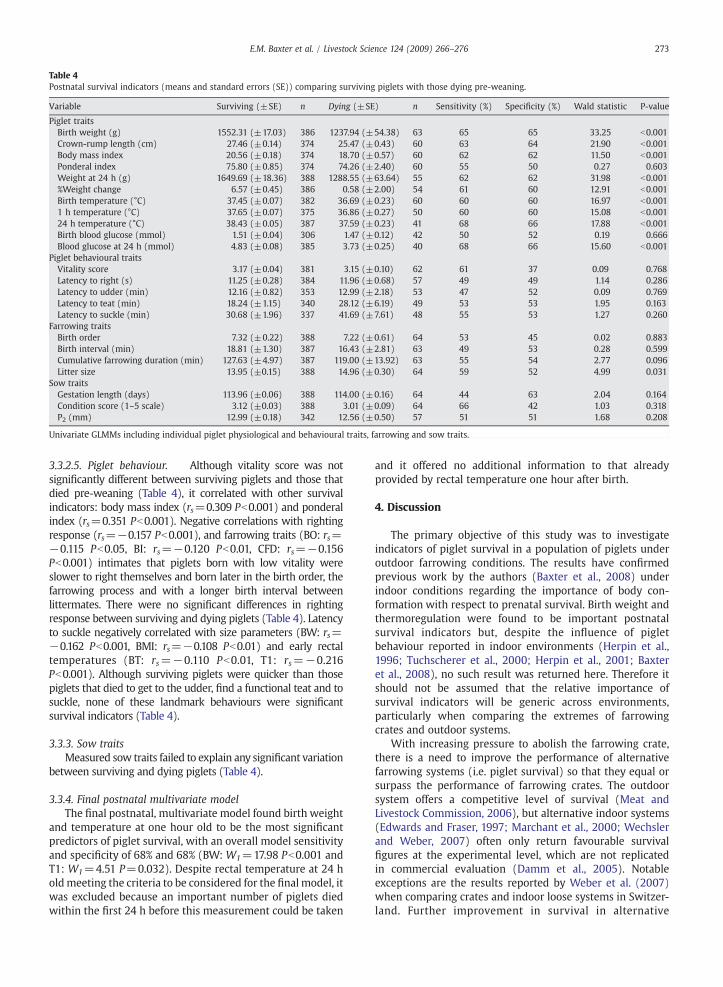
Table 4Postnatal survival indicators (means and standard errors (SE)) comparing surviving piglets with those dying pre-weaning.
Variable Surviving (±SE) n Dying (±SE) n Sensitivity (%) Specificity (%) Wald statistic P-value
Piglet traitsBirth weight (g) 1552.31 (±17.03) 386 1237.94 (±54.38) 63 65 65 33.25 b0.001Crown-rump length (cm) 27.46 (±0.14) 374 25.47 (±0.43) 60 63 64 21.90 b0.001Body mass index 20.56 (±0.18) 374 18.70 (±0.57) 60 62 62 11.50 b0.001Ponderal index 75.80 (±0.85) 374 74.26 (±2.40) 60 55 50 0.27 0.603Weight at 24 h (g) 1649.69 (±18.36) 388 1288.55 (±63.64) 55 62 62 31.98 b0.001%Weight change 6.57 (±0.45) 386 0.58 (±2.00) 54 61 60 12.91 b0.001Birth temperature (°C) 37.45 (±0.07) 382 36.69 (±0.23) 60 60 60 16.97 b0.0011 h temperature (°C) 37.65 (±0.07) 375 36.86 (±0.27) 50 60 60 15.08 b0.00124 h temperature (°C) 38.43 (±0.05) 387 37.59 (±0.23) 41 68 66 17.88 b0.001Birth blood glucose (mmol) 1.51 (±0.04) 306 1.47 (±0.12) 42 50 52 0.19 0.666Blood glucose at 24 h (mmol) 4.83 (±0.08) 385 3.73 (±0.25) 40 68 66 15.60 b0.001
Piglet behavioural traitsVitality score 3.17 (±0.04) 381 3.15 (±0.10) 62 61 37 0.09 0.768Latency to right (s) 11.25 (±0.28) 384 11.96 (±0.68) 57 49 49 1.14 0.286Latency to udder (min) 12.16 (±0.82) 353 12.99 (±2.18) 53 47 52 0.09 0.769Latency to teat (min) 18.24 (±1.15) 340 28.12 (±6.19) 49 53 53 1.95 0.163Latency to suckle (min) 30.68 (±1.96) 337 41.69 (±7.61) 48 55 53 1.27 0.260
Farrowing traitsBirth order 7.32 (±0.22) 388 7.22 (±0.61) 64 53 45 0.02 0.883Birth interval (min) 18.81 (±1.30) 387 16.43 (±2.81) 63 49 53 0.28 0.599Cumulative farrowing duration (min) 127.63 (±4.97) 387 119.00 (±13.92) 63 55 54 2.77 0.096Litter size 13.95 (±0.15) 388 14.96 (±0.30) 64 59 52 4.99 0.031
Sow traitsGestation length (days) 113.96 (±0.06) 388 114.00 (±0.16) 64 44 63 2.04 0.164Condition score (1–5 scale) 3.12 (±0.03) 388 3.01 (±0.09) 64 66 42 1.03 0.318P2 (mm) 12.99 (±0.18) 342 12.56 (±0.50) 57 51 51 1.68 0.208
Univariate GLMMs including individual piglet physiological and behavioural traits, farrowing and sow traits.
273E.M. Baxter et al. / Livestock Science 124 (2009) 266–276
3.3.2.5. Piglet behaviour. Although vitality score was notsignificantly different between surviving piglets and those thatdied pre-weaning (Table 4), it correlated with other survivalindicators: body mass index (rs=0.309 Pb0.001) and ponderalindex (rs=0.351 Pb0.001). Negative correlations with rightingresponse (rs=−0.157 Pb0.001), and farrowing traits (BO: rs=−0.115 Pb0.05, BI: rs=−0.120 Pb0.01, CFD: rs=−0.156Pb0.001) intimates that piglets born with low vitality wereslower to right themselves and born later in the birth order, thefarrowing process and with a longer birth interval betweenlittermates. There were no significant differences in rightingresponse between surviving and dying piglets (Table 4). Latencyto suckle negatively correlated with size parameters (BW: rs=−0.162 Pb0.001, BMI: rs=−0.108 Pb0.01) and early rectaltemperatures (BT: rs=−0.110 Pb0.01, T1: rs=−0.216Pb0.001). Although surviving piglets were quicker than thosepiglets that died to get to the udder, find a functional teat and tosuckle, none of these landmark behaviours were significantsurvival indicators (Table 4).
3.3.3. Sow traitsMeasured sow traits failed to explain any significant variation
between surviving and dying piglets (Table 4).
3.3.4. Final postnatal multivariate modelThe final postnatal, multivariate model found birth weight
and temperature at one hour old to be the most significantpredictors of piglet survival, with an overall model sensitivityand specificity of 68% and 68% (BW: W1=17.98 Pb0.001 andT1: W1=4.51 P=0.032). Despite rectal temperature at 24 holdmeeting the criteria to be considered for the final model, itwas excluded because an important number of piglets diedwithin the first 24 h before this measurement could be taken
and it offered no additional information to that alreadyprovided by rectal temperature one hour after birth.
4. Discussion
The primary objective of this study was to investigateindicators of piglet survival in a population of piglets underoutdoor farrowing conditions. The results have confirmedprevious work by the authors (Baxter et al., 2008) underindoor conditions regarding the importance of body con-formation with respect to prenatal survival. Birth weight andthermoregulation were found to be important postnatalsurvival indicators but, despite the influence of pigletbehaviour reported in indoor environments (Herpin et al.,1996; Tuchscherer et al., 2000; Herpin et al., 2001; Baxteret al., 2008), no such result was returned here. Therefore itshould not be assumed that the relative importance ofsurvival indicators will be generic across environments,particularly when comparing the extremes of farrowingcrates and outdoor systems.
With increasing pressure to abolish the farrowing crate,there is a need to improve the performance of alternativefarrowing systems (i.e. piglet survival) so that they equal orsurpass the performance of farrowing crates. The outdoorsystem offers a competitive level of survival (Meat andLivestock Commission, 2006), but alternative indoor systems(Edwards and Fraser, 1997; Marchant et al., 2000; Wechslerand Weber, 2007) often only return favourable survivalfigures at the experimental level, which are not replicatedin commercial evaluation (Damm et al., 2005). Notableexceptions are the results reported by Weber et al. (2007)when comparing crates and indoor loose systems in Switzer-land. Further improvement in survival in alternative

274 E.M. Baxter et al. / Livestock Science 124 (2009) 266–276
farrowing systems could utilise genetic selection strategiestargeting specific survival traits. However, as evidenced by theresults from this study, detailed investigations into how thesow and her piglets interact with the specific farrowingenvironment to promote survival are needed in order toimplement these programmes.
4.1. Mortality
Overall, the total mortality figures for this study are similarto those reported from commercial units (approximately20%). The live-born mortality figures are higher than thosereported by UK recording schemes (Meat and LivestockCommission, 2006). However, this could be a result of theaccurate post-mortem analysis performed on every pigletdying pre-weaning. In the outdoor industry, practicalities ofproduction limit the accuracy of recording the actual numberof deaths and the cause. Subjective assessment of pre-weaning deaths can result in a constrained choice of causes,leading to a discrepancy in the reported figures with inflatednumbers of deaths being attributed to crushing or stillbirths(Edwards et al., 1994).
4.2. Prenatal survival indicators
This study confirmed previous results (Baxter et al., 2008)that piglet shape and size are vital components of prenatalsurvival. Piglets that were disproportionately long and thinwere more likely to be born dead and piglets that had a lowerbody mass index were more likely to die postnatally. Althoughnot as important as body conformation indices with respect toprenatal survival, birth weight was a significant survivalindicator and many authors have highlighted this trait as themost important survival parameter (Kerr and Cameron, 1995;Roehe, 1999; Roehe and Kalm, 2000; Knol et al., 2002a,b;Arango et al., 2005). It could be the key trait in selectionprogrammes for decreasing mortality, as it has a greaterheritability (0.05 for direct effects and 0.22 for maternaleffects—Roehe, 1999) than piglet survival per se. However, therelationship between birth weight and survival is a quadraticone (Roehe and Kalm, 2000),with the extremes in birthweightat equal risk ofmortality, and selection solely for increased birthweight could therefore compromise survival and litter size,with which it has a negative genetic correlation. Within-litteruniformity is as important as high average birth weight forpiglet survival (Rydhmer, 2000; Knol et al., 2002a). This is not anew concept (English and Smith, 1975), yet it is still an area inneed of progress. Selecting for an optimum, rather than highbirth weight and for within-litter homogeneity of birth weightis a suggested strategy to improve survival inpolytocous species(Damgaard et al., 2003; Bolet et al., 2007). The results from thepresent study suggest that optimum shape as well as weightshould be considered within breeding programmes to improvepiglet survival.
Farrowing traits are additional factors influencing still-births. Longer farrowing durations incur greater risks forthose piglets born later in the birth order, with the greaterincidence of premature umbilical cord rupture and subse-quent hypoxia (Herpin et al., 2001). In this study, theseaspects of farrowing were significant prenatal survivalindicators.
4.3. Postnatal survival indicators
Birth weight and thermoregulation were the most impor-tant postnatal survival indicators. Typically, birth weightexplains the variation in thermoregulation, with a closecorrelation between these factors. Smaller piglets have highersurface area to volume ratios, resulting in greater suscept-ibility to hypothermia (Herpin et al., 2002a). In this studyrectal temperature immediately after birth had an effect onsurvival, with piglets that died pre-weaning having lowerbirth rectal temperatures. This supports our previous findings(Baxter et al., 2008), and suggests that susceptible pigletsmight have lower homeothermic capabilities. Piglets havebeen shown to differ in such capability (Herpin et al., 2002b),with influential factors including birth weight, thematurationof skeletal muscle metabolism and function, as well asgenotype and vigour at birth. Temperature one hour afterbirth explained additional variance in mortality to thatalready explained by birth weight. Within the first hourafter birth, thermoregulation will be compromised in allpiglets as a result of evaporation of the placental fluids andconsequent cooling. Susceptible piglets fail to recover fromthis initial temperature drop, and hypothermia then affectslatency to suckle leading to starvation, subsequent lethargyand ultimately crushing by the sow (Weary et al., 1996). Inthis study the negative correlation between latency to suckleand rectal temperature one hour after birth supports thisscenario.
Piglet behaviour is a key aspect in adaptation toextrauterine life. Piglets that are quicker to perform land-mark behaviours, particularly to suckle colostrum, increasetheir chances of survival (Tuchscherer et al., 2000; Herpinet al., 2001; Baxter et al., 2008). In this study, latency tosuckle was not a significant survival indicator, with highvariability within survival categories. This conflicts withwork conducted in indoor environments (Tuchscherer et al.,2000; Herpin et al., 2001; Baxter et al., 2008) and a studyobserving hypothermia in outdoor piglets (Riart et al.,2000). The latter authors found positive relationshipsbetween temperature and suckling in both environmentsbut, in contrast to our study, they concluded that latency tosuckle was more vital for outdoor piglets than indoorpiglets to preserve homeothermy. Despite the conclusionsof these authors (Riart et al., 2000), they did not subse-quently track piglets in the outdoor environment, withmortality not recorded, and the impact this landmarkbehaviour had on survival was not determined but assumedby the authors. The failure for latency to suckle to be acritical survival factor in the current study could actually bean influence of the environment. In farrowing huts, deep-straw bedding has multiple functions: it acts as suitablenesting material for the pre-parturient sow, it is a malle-able, non-uniform mattress cushioning sow posturechanges and offering a buffer to piglets from fatal crushingand it creates a microclimate for the vulnerable newborns.This latter function might diminish the influence of timingon landmark behaviours. In the indoor crated environment,suckling quickly is vital, primarily to preserve home-othermy in the piglets. The absence of substantial, if any,nesting material means the newborn piglet, wet fromplacental fluids, is exposed and experiencing rapid cooling

275E.M. Baxter et al. / Livestock Science 124 (2009) 266–276
even in temperature-controlled environments. Recentresearch in indoor loose-housed systems (Malmkvist etal., 2006) has recognised that even in a temperature-con-trolled environment the newborn piglet is highly suscep-tible to chilling. This group have investigated the influenceof under-floor heating on piglet viability and showed agreater proportion of piglets survived in the treatment withunder-floor heating. They go on to suggest that this could bea result of potentially vulnerable piglets (i.e. low birthweight, no colostrum ingestion), at risk from hypothermia,being kept warmer by the under-floor heating, thus givingthem a longer opportunity to achieve their first sucklingevent.
In detailed experiments on environment–temperatureinteractions (Mount, 1967), the insulating properties ofstraw (2.5 cm deep) were measured and found to reduceheat loss to the floor by 40% when compared to uninsulatedconcrete. Deep bedding slows the rate of heat loss and has athermal resistance 11 times greater than that of concrete slatsand 22 times greater than that of solid, wet concrete flooring(Wathes and Whittemore, 2006). Detailed experiments byAlgers and Jensen (1990) on the nest-sites of free-rangingpigs concluded that deep-straw nests provide an optimummicroclimate for the neonate. The deep-straw bed in thecurrent study simulates a more natural nesting site, and thestraw facilitates physical removal of membranes and absorp-tion of birth fluids, reducing the impact of placental fluidevaporation and buffering the piglet from immediate suscept-ibility to the extrauterine environment. Domestication of thepig over centuries has failed to alter the innate need for a sowto nest-build (Jensen, 2002), probably because of theextremely high survival value of this function under naturalconditions. Potentially, it has also failed to alter the impor-tance of the nest as a microclimate balancing the thermo-regulatory demands of both the piglet and the sow.
5. Conclusions and implications
Survival indicators identified as important in outdoorsystems were piglet body shape and size, as well as farrowingtraits and thermoregulation. With respect to prenatal survival,this study has confirmed previous work by the authors (Baxteret al., 2008) that piglet body shape ismore important thanbirthweight and this has implications for genetic selection strategiestargeting birth weight as the most important survival trait.These results suggest that optimum body shape as well asoptimum weight should be considered as a survival trait,regardless of farrowing system. With respect to postnatalsurvival, there is a suggestion that behavioural parametersknown to be important in indoor systems, might not be asinfluential in outdoor systems. This illustrates the need forfurther detailed investigations of individual behavioural andphysiological characteristics under alternative environments tothe farrowing crate, in order to identify appropriate traits foruse in breeding programmes incorporating welfare as well asproduction goals.
This study concentrated on piglet survival traits; howeverthe behaviour of the mother is likely to be a critical factor forpiglet survival and further investigations taking into accountthese important sow factors in outdoor systems are beingconducted.
Acknowledgements
The authors would like to thank owners and staff of theparticipating farm and the technical staff who worked on theproject. The study was part of the Welfare Quality® researchproject which has been co-financed by the EuropeanCommission, within the 6th Framework Programme, contractNo. FOOD-CT-2004-506508. The text represents the authors'views and does not necessarily represent a position of theCommission who will not be liable for the use made of suchinformation. The work was also funded by the ScottishGovernment. We are grateful to the contribution of geneticinformation and animals from the Genomum project(LS3103), which is funded by the Department for Environ-ment, Food and Rural Affairs (Defra) and the ScottishGovernment, with substantial support from partners: Gram-pian Country Food Group (GCFG), PIC International, JSRGenetics Ltd, and SSPCA.
References
Algers, B., Jensen, P., 1990. Thermal microclimate in winter farrowing nests offree-ranging domestic pigs. Livest. Prod. Sci. 25, 177–181.
Arango, J., Misztal, I., Tsuruta, S., Culbertson, M., Herring, W., 2005. Threshold-linear estimation of genetic parameters for farrowing mortality, littersize, and test performance of LargeWhite sows. J. Anim Sci. 83, 499–506.
Arey, D.S., 1997. Behavioural observations of peri-parturient sows and thedevelopment of alternative farrowing accommodation: a review. Anim.Welf. 6, 217–229.
Baxter, E.M., Jarvis, S., D'Eath, R.B., Ross, D.W., Robson, S.K., Farish, M.,Nevison, I.M., Lawrence, A.B., Edwards, S.A., 2008. Investigating thebehavioural and physiological indicators of neonatal survival in pigs.Theriogenology 69, 773–783.
Bolet, G., Garreau, H., Joly, T., Theau-Clement, M., Falieres, J., Hurtaud, J.,Bodin, L., 2007. Genetic homogenisation of birth weight in rabbits:indirect selection response for uterine horn characteristics. Livest. Sci.111, 28–32.
Callaway, T.R., Morrow, J.L., Johnson, A.K., Dailey, J.W., Wallace, F.M.,Wagstrom, E.A., McGlone, J.J., Lewis, A.R., Dowd, S.E., Poole, T.L.,Edrington, T.S., Anderson, R.C., Genovese, K.J., Byrd, J.A., Harvey, R.B.,Nisbet, D.J., 2005. Environmental prevalence and persistence of Salmo-nella spp. in outdoor swine wallows. Foodborne Pathogens Dis. 2,263–273.
Damm, B.I., Pedersen, L.J., Heiskanen, T., Nielsen, N.P., 2005. Long-stemmedstraw as an additional nesting material in modified Schmid pens in acommercial breeding unit: effects on sow behaviour, and on pigletmortality and growth. Appl. Anim. Behav. Sci. 92, 45–60.
Damgaard, L.H., Rydhmer, L., Lovendahl, P., Grandinson, K., 2003. Geneticparameters for within-litter variation in piglet birth weight and changein within-litter variation during suckling. J. Anim Sci. 81, 604–610.
Edwards, S.A., 2002. Perinatal mortality in the pig: environmental orphysiological solutions? Livest. Prod. Sci. 78, 3–12.
Edwards, S.A., 2005. Product quality attributes associated with outdoor pigproduction. Livest. Prod. Sci. 94, 5–14.
Edwards, S.A., Zanella, A., 1996. Producao de suinos ao ar livre na Europa:bem-estar e consideracoes ambientais. [Pig production in outdoorsystems in Europe: produciton, welfare and environmental considera-tions]. Hora Veterinara 92, 86–93.
Edwards, S.A., Fraser, D., 1997. Housing systems for farrowing and lactation.Pig J. 39, 77–89.
Edwards, S.A., Smith, W.J., Fordyce, C., Macmenemy, F., 1994. An analysis ofthe causes of piglet mortality in a breeding herd kept outdoors. Vet. Rec.135, 324–327.
English, P.R., Smith, W.J., 1975. Some causes of death in neonatal piglets. Vet.Ann. 15, 95–104.
Held, S., Mason, G., Mendl, M., 2006. Maternal responsiveness of outdoorsows from first to fourth parities. Appl. Anim. Behav. Sci. 98, 216–233.
Herpin, P., LeDividich, J., Hulin, J.C., Fillaut, M., DeMarco, F., Bertin, R., 1996.Effect of the level of asphyxia during delivery on viability at birth andearly postnatal vitality of newborn pigs. J. Anim Sci. 74, 2067–2075.
Herpin, P., Hulin, J.C., Le Dividich, J., Fillaut, M., 2001. Effect of oxygeninhalation at birth on the reduction of early postnatal mortality in pigs.J. Anim Sci. 79, 5–10.

276 E.M. Baxter et al. / Livestock Science 124 (2009) 266–276
Herpin, P., Damon, M., Le Dividich, J., 2002a. Development of thermoregula-tion and neonatal survival in pigs. Livest. Prod. Sci. 78, 25–45.
Herpin, P., Lossec, G., Schmidt, I., Cohen-Adad, F., Duchamp, C., Lefaucheur, L.,Goglia, F., Lanni, A., 2002b. Effect of age and cold exposure onmorphofunctional characteristics of skeletal muscle in neonatal pigs.Pflugers Archiv-Euro. J. Phys. 444, 610–618.
Hotzel, M.J., Pinheiro, M.F., Wolf, F.M., Costa, O.A.D., 2004. Behaviour of sowsand piglets reared in intensive outdoor or indoor systems. Appl. Anim.Behav. Sci. 86, 27–39.
Jarvis, S., Lawrence, A.B., Mclean, K.A., Deans, L.A., Chirnside, J., Calvert, S.K.,1997. The effect of environment on behavioural activity, ACTH, beta-endorphin and cortisol in pre-farrowing gilts. Anim Sci. 65, 465–472.
Jensen, P., 1986. Observations on the maternal behaviour of free-rangingdomestic pigs. Appl. Anim. Behav. Sci. 16, 131–142.
Jensen, P., 1988. Maternal behaviour and mother–young interactions duringlactation in free-ranging domestic pigs. Appl. Anim. Behav. Sci. 20,297–308.
Jensen, P., 2002. Behaviour of pigs. In: Jensen, P. (Ed.), The Ethology ofDomestic Animals. CABI Publishing, Wallingford, UK, pp. 159–172.
Johnson, A.K., McGlone, J.J., 2003. Fender design and insulation of farrowinghuts: effects on performance of outdoor sows and piglets. J. Anim Sci. 81,955–964.
Johnson, A.K., Morrow-Tesch, J.L., McGlone, J.J., 2001. Behavior and perfor-mance of lactating sows and piglets reared indoors or outdoors. J. AnimSci. 79, 2571–2579.
Kerr, J.C., Cameron, N.D., 1995. Reproductive-performance of pigs selected forcomponents of efficient lean growth. Anim Sci. 60, 281–290.
Knol, E.F., Ducro, B.J., van Arendonk, J.A.M., van der Lende, T., 2002a. Direct,maternal and nurse sow genetic effects on farrowing-, pre-weaning- andtotal piglet survival. Livest. Prod. Sci. 73, 153–164.
Knol, E.F., Leenhouwers, J.I., van der Lende, T., 2002b. Genetic aspects of pigletsurvival. Livest. Prod. Sci. 78, 47–55.
Lawrence, A.B., Petherick, J.C., Mclean, K.A., Deans, L.A., Chirnside, J., Vaughan,A., Clutton, E., Terlouw, E.M.C., 1994. The effect of environment onbehavior, plasma-cortisol and prolactin in parturient sows. Appl. Anim.Behav. Sci. 39, 313–330.
Leenhouwers, J.I., Knol, E.F., van der Lende, T., 2002. Differences in lateprenatal development as an explanation for genetic differences in pigletsurvival. Livest. Prod. Sci. 78, 57–62.
Malmkvist, J., Pedersen, L.J., Damgaard, B.M., Thodberg, K., Jørgensen, E.,Labouriau, R., 2006. Does floor heating around parturition affect thevitality of piglets born to loose housed sows? App. Anim. Beh. Sci. 99,88–105.
Marchant, J.N., Rudd, A.R., Mendl, M.T., Broom, D.M., Meredith, M.J., Corning,S., Simmins, P.H., 2000. Timing and causes of piglet mortality inalternative and conventional farrowing systems. Vet. Rec. 147, 209–214.
McGlone, J.J., Hicks, T.A., 2000. Farrowing hut design and sow genotype(Camborough-15 vs 25% Meishan) effects on outdoor sow and litterproductivity. J. Anim Sci. 78, 2832–2835.
Meat and Livestock Commission, 2006. The Pig Yearbook 2006.Mount, L.E., 1967. The heat loss from new-born pigs to the floor. Res. Vet. Sci.
8, 175–186.Muirhead, M.R., Alexander, T.J.L., 1997. Managing pig health and the
treatment of disease: a reference for the farm. 5 M Enterprises. Sheffield,UK, p. 459.
Newberry, R.C., Woodgush, D.G.M., 1985. The suckling behavior of domesticpigs in a semi-natural environment. Behaviour 95, 11–25.
O'Reilly, K.M., Harris, M.J., Mendl, M., Held, S., Moinard, C., Statham, P.,Marchant-Forde, J., Green, L.E., 2006. Factors associated with preweaningmortality on commercial pig farms in England and Wales. Vet. Rec. 159,193–196.
Riart, G.R., Edwards, S.A., English, P.R., 2000. Assessment of hypothermia inoutdoor newborn piglets and comparison with an indoor system.Proceedings of the British Society of Animal Science, p. 139.
Roehe, R., 1999. Genetic determination of individual birth weight and itsassociation with sow productivity using Bayesian analyses. J. Anim. Sci.77, 330–343.
Roehe, R., Kalm, E., 2000. Estimation of genetic and environmental riskfactors associated with pre-weaning mortality in piglets using general-ized linear mixed models. Anim. Sci. 70, 227–240.
Roehe, R., Shrestha, N.P., Mekkawy, W., Baxter, E.M., Knap, P.W., Smurthwaite,K.M., Jarvis, S., Lawrence, A.B., Edwards, S.A., 2009. Genetic analyses ofpiglet survival and individual birth weight on first generation dataof a selection experiment for piglet survival under outdoor conditions.Livestock Science, Corrected Proof 121, 173–181.
Rydhmer, L., 2000. Genetics of sow reproduction, including puberty, oestrus,pregnancy, farrowing and lactation. Livest. Prod. Sci. 66, 1–12.
Tuchscherer, M., Puppe, B., Tuchscherer, A., Tiemann, U., 2000. Earlyidentification of neonates at risk: traits of newborn piglets with respectto survival. Theriogenology 54, 371–388.
Vieuille, C., Berger, F., Le Pape, G., Bellanger, D., 2003. Sow behaviour involvedin the crushing of piglets in outdoor farrowing huts—a brief report. Appl.Anim. Behav. Sci. 80, 109–115.
Wathes, C., Whittemore, C.T., 2006. Environmental management of pigs. In:Kyriazakis, I., Whittemore, C.T. (Eds.), Whittemore's Science and Practiceof Pig Production. Blackwell Publishing, Oxford, pp. 533–592.
Weary, D.M., Pajor, E.A., Thompson, B.K., Fraser, D., 1996. Risky behaviour bypiglets: a trade off between feeding and risk of mortality by maternalcrushing? Anim. Beh. 51, 619–624.
Weber, R., Keli, N., Fehr, M., Horat, R., 2007. Piglet mortality on farms usingfarrowing systems with or without crates. Anim. Welf. 16, 277–279.
Wechsler, B., Weber, R., 2007. Loose farrowing systems: challenges andsolutions. Anim. Welf. 16, 295–307.
Wulbers-Mindermann, M., Algers, B., Berg, C., Lundeheim, N., Sigvardsson, J.,2002. Primiparous and multiparous maternal ability in sows in relationto indoor and outdoor farrowing systems. Livest. Prod. Sci. 73, 285–297.